 Tatiana Begun
Tatiana Begun Adrian Teacă
Adrian Teacă Mihaela Mureşan
Mihaela Mureşan Pedro A. Quijón
Pedro A. Quijón Selma Menabit
Selma Menabit Victor Surugiu
Victor Surugiu- 1National Institute for Research and Development on Marine Geology and Geoecology–GeoEcoMar, Bucharest, Romania
- 2Department of Biology, University of Prince Edward Island, Charlottetown, PE, Canada
- 3Department of Microbiology, Institute of Biology Bucharest, Bucharest, Romania
- 4Faculty of Biology, “Alexandru Ioan Cuza” University of Iaşi, Iaşi, Romania
Exposure to a variety of anthropogenic stressors has prompted the creation of a growing number of marine protected areas (MPAs). Unfortunately, long-term changes in both the sources of stress and the extent or operation of the MPAs have also occurred, at a pace that is often faster than the research trying to understand the consequences of those changes. This study addresses this challenge by focusing on two MPAs located in the southern Romanian Black Sea, namely the Cape Tuzla and the Mangalia Sulfur springs. Using 95 seafloor samples collected over a 7-year period, we explored distinct benthic habitats and their associated communities in a depth gradient common to both MPAs. We described benthic communities, their density, biomass, and diversity and subsequently used them as indicators of the health of the seafloor. Specifically, we used the M-AMBI*(n) index to assess the ecological status of the benthic habitats found in both MPAs. Our results identified seven distinct habitats across infralittoral and circalittoral areas and described species composition and diversity levels, reaching up to a grand total of 119 benthic taxa. These communities were often dominated by polychaetes and crustaceans in terms of density and by mollusks in terms of biomass. Several mollusks represent key ecosystem engineer species creating or modifying their local habitat conditions. Our results revealed that two thirds of the sampling stations could be characterized as having a good ecological status (GES), and were therefore dominated by species normally considered as either sensitive or indifferent to organic pollution. Meanwhile, the other third of the stations was labeled as non-GES, was dominated by opportunistic species and represent a concern for the coastline system. This study calls for further research to carefully describe the valuable coastal habitats under the frame of MPAs, and to assess the effects of specific pollutants and the response of organisms that seem most affected by these stressors.
Introduction
Biodiversity and ecological integrity are essential to the function and services provided by marine ecosystems (Rees et al., 2020). The description of the structure and dynamics of these ecosystems relies on basic information about species occurrence, composition, and distribution (Danovaro et al., 2008; Hillebrand et al., 2017), but integrative measures such as biodiversity is a priority in a way to ascertain the ecological status and to evaluate the targets for protection measures (Teixeira et al., 2016). Nowadays, the biodiversity and integrity of many marine ecosystems are threatened by habitat degradation and species loss, two issues often associated with overfishing, pollution, ocean acidification, and climate change (Airoldi and Beck, 2007; Halpern et al., 2008, 2015; Claudet and Fraschetti, 2010). To counteract such impacts, a growing number of Marine Protected Areas (MPAs) has been established around the globe (Cabral et al., 2020). MPAs are popular management tools to safeguard biodiversity and ecosystem services. However, some MPAs fail to meet their conservation goals and are quickly tagged as “paper parks” that provide limited protection to their biota (Rife et al., 2013; Plumeridge and Roberts, 2017).
Much like other coastlines, Romanian MPAs have faced scrutiny over their effectivity achieving conservation goals. This has been in part due to missteps during their establishment, which failed to involve relevant stakeholders and reach public consensus, leading to hard-to-solve conflicts (Begun et al., 2018). Meanwhile, global impacts on biodiversity, ecosystem services and society continue to accelerate (IPBES, 2019), and the assessment of MPAs’ benefits becomes even more necessary. Information on macrozoobenthic communities’ features offers an opportunity for such assessment, given the integrative nature of seafloor habitats and biota. However, historical changes on management and data gaps represent in some cases, as in the Romanian Black Sea region, a challenge that must be carefully overcome.
The Romanian Regional Context
Biodiversity research on the Romanian shelf grew vigorously after WWII from the seminal work of Professor M. Bacescu and collaborators. Between the 1950s and 1970s, numerous species were described, including micro- and macrophytobenthos, Nemertini, Nematoda, Polychaeta, Mollusca, and Crustacea (Bacescu et al., 1971). The taxonomic and ecological contributions of these researchers remained underrecognized in a period of major environmental changes in the Black Sea (Gomoiu, 1992; Mee, 1992; Zaitsev, 1993; Bologa et al., 1995; Leppäkoski and Mihnea, 1996): growing coastal euthrophication (Mee and Topping, 1999; Cociaşu et al., 2008) and associated events of phytoplankton and macroalgal blooms (Bologa et al., 1995; Moncheva et al., 1995; Bodeanu, 2002), resulted in population declines or extinctions (Gomoiu, 1983, 1992), the introduction of new species (Gomoiu et al., 2002), and the modification of sediment regimes due to hydrographic disruption in tributary rivers (Panin et al., 1996; Panin and Jipa, 1998; Duţu et al., 2018). The combined impact of these stressors led to the restructuring of benthic assemblages with 60% of benthic fauna to be lost in three decades and local declines in population numbers and diversity (Petranu, 1997; Gomoiu, 1998).
The lag in marine benthic research in a period of such striking changes has been linked to the small number of specialists and limitations in resources due to the broader economic decline in the region (Gomoiu et al., 2003/2004). After 1990, the socio-economic recession in the riparian Black Sea countries and the introduction of national and international programs aiming at reducing river inputs and eutrophication, gave the Romanian coastline an opportunity to recover. However, despite the slow recovery observed in various marine ecosystems in the region (Mee et al., 2005; Friedrich et al., 2014; Mureşan et al., 2020; Teacă et al., 2020), it remained fragile and vulnerable to anthropogenic impacts, ecological accidents, and climate change.
Marine Protected Areas’ Benthic Communities and Objectives
Research on marine ecosystems, in general, and benthic communities, in particular, intensified in the 2000s with the application of the European Directives and the growth of research funding. Some of that research was directed toward macrozoobenthos in MPAs (Abaza and Godeanu, 2008; Abaza, 2010; Niţă et al., 2012), mapping of benthic habitats according to the Natura 2000 classification (Zaharia et al., 2012; Ungureanu et al., 2015), the study of benthic communities associated with rocky bottom substrates and macrophytic algae (Teacă et al., 2006a,b, 2010; Filimon et al., 2016), and the assessment of circalittoral habitats (Begun et al., 2010, 2018; Abaza et al., 2018a,2019; Mureşan and Teacă, 2019; Mureşan et al., 2019; Teacă et al., 2019, 2020). Today, the Romanian MPA network encompasses 7,457 km2 of the Romanian shelf distributed among ten MPAs. Two MPAs in particular, the Cape Tuzla (ROSCI0273) and the Mangalia submerged Sulfur springs (ROSCI0094), are examined in this study and together represent 1.8% (107.3 km2) of the surface of all Romanian protected areas. Such area was reached only recently, when the area of both MPAs increased by 65 and 93%, respectively, following the approval of the MMAP Order no. 46/2016.
The growth of these protected areas also entailed a challenge now faced by many other MPAs: the knowledge on large portions of protected seafloor remains extremely limited, despite the need to apply European Union’s policies, such as the Marine Strategy Framework Directive, the Birds and Habitats Directives (Nature Directives hereafter) and the most recent European Green Deal (EC, 2019) and Biodiversity Strategy for 2030 (EC, 2020). As in other MPAs, the description, mapping and status of macrozoobenthic communities in the Cape Tuzla and the Mangalia Sulfur springs MPAs are critical and are examined in this study. We aimed to assess biodiversity, map benthic habitats according to the EUNIS classification, and to achieve a synoptic overview of the state of these communities. We attempted this by embracing the modern approach to management of marine habitats of European policies, such as the 2008/56/EC Directive, which requires European Member States to achieve a Good Environmental Status (GES) by 2020.
Materials and Methods
The Study Area
The Black Sea is an inland sea with a maximum depth of 2,245 m (Sorokin, 2002), connected in the northeast with the Sea of Azov and in the southwest with the Sea of Marmara and further south with the Aegean and Mediterranean Seas (Figure 1). The Romanian part of the Black Sea coastline spans 244 km and can be divided in a northern part, between Musura Bay and the Cape Midia Harbor jetties, and a southern part between the Cape Midia and the Romanian-Bulgarian border. The southern part includes the most diverse geomorphological features of the coastline (Panin and Jipa, 1998; Panin, 2005), and is characterized by the presence of almost continuous loess cliffs (12–20 m high). This area is also more stable in terms of geodynamic processes than the northern deltaic coast (Panin and Jipa, 1998), but is more exposed to anthropogenic stressors. The shoreline south of Constanta does receive a scarce input of sediments from the Danube system, so the sediments in this area are mainly of organic origin, derived from mollusk shells, and affected by continuous erosion caused by inappropriate hydrotechnical protective works.
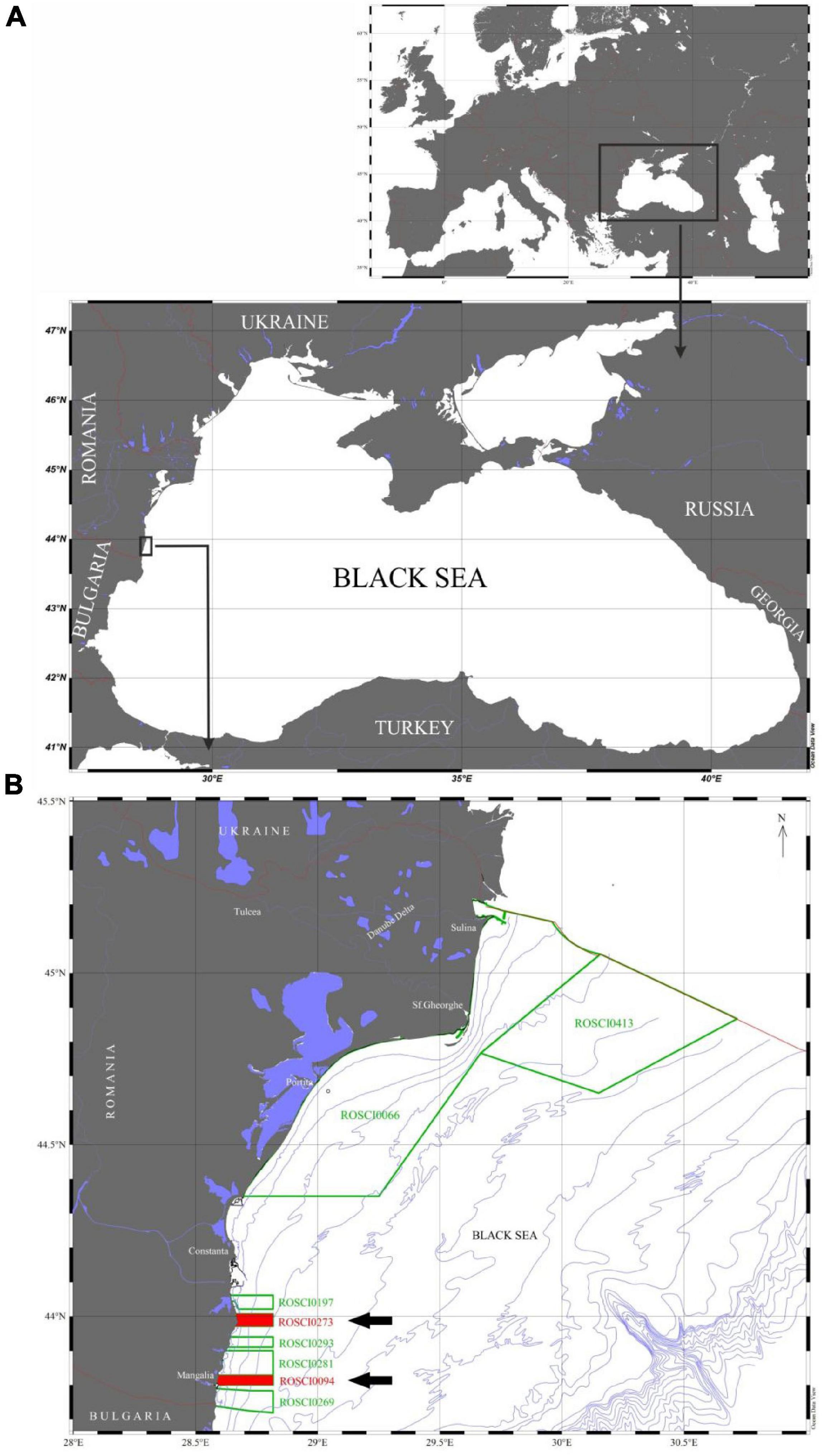
Figure 1. Maps outlining (A) the approximate location of the study area in the southern Romanian Black Sea, and (B) the location of eight Natura 2000 Marine Protected Areas (MPAs) highlighting (red polygons pointed by arrows) the Cape Tuzla MPA (ROSCI0273) and the Mangalia submarine Sulfur springs MPA (ROSCI0094).
The two MPAs assessed in this study, the Cape Tuzla and the Mangalia Sulfur springs are located in the Romanian southern part extending down to 50 m depth (Figure 1). They occupy 107.3 km2 and comprise loess cliffs which are constituted by discontinuous Quaternary red clays and Sarmatian limestone. Their erosion has created a heterogeneous subtidal bottom with extensive blocks of rock and boulder fields going down to 25 m in depth (Panin, 2005). These MPAs were officially designated under the EC Habitats Directive in 2009 (Decision 2009/92/EEC) to protect these unique and rare marine ecosystems, the submerged sulfur springs, extensive canopies of Cystoseira and Zostera meadows, and their associated natural resources from multiple socio-economic impacts. The MPAs support a rich biodiversity and have been recognized as biodiversity hotspots and important nursery grounds for several protected fish species through Habitat Directives and have been also listed in the Annex II of the Directive 92/43/EEC. They have been also included in the national list of the Natura 2000 sites, hosting four distinct habitats (Table 1).

Table 1. Habitats listed in the “Habitats Directive” of the Cape Tuzla and the Mangalia Sulfur springs MPAs (ROSCI0273 and ROSCI0094, respectively), according to the Natura 2000 classification.
Data Collection and Analyses
During the seven field campaigns conducted between 2012 and 2018, 75 stations were sampled in total—47 stations in the Cape Tuzla and 28 stations in the Mangalia Sulfur springs MPAs (Figure 2). Other 20 samples were also collected for visual inspection and habitat identification on board of the research vessel R/V “Mare Nigrum.” The W-SE bathymetry gradient of the study areas ranged between 11.1 and 50.4 m deep. Folk 5/7 was used for sediment classification of the marine habitat delineation according to the EMODnet Seabed Habitats substrate and National classification schemes (Populus et al., 2017; Vasquez et al., 2020). Habitat distribution maps were then developed using the combination of sediment type, depth and point (station) data. Such habitat distribution was subsequently confirmed (in 2021) using remote sensing techniques such as multibeam and Side-Scan Sonar (Popa, Pers. Comm.). A single sample was collected at each station using van Veen grabs with grasping areas of 0.135 m2 and 0.125 m2. Samples were sieved through a 0.5 mm mesh (Bacescu et al., 1971; Todorova and Konsulova, 2005) and the retained organisms were preserved in 4% neutralized formalin-seawater solution. Density and biomass (wet weight) were estimated as ind. m––2 and g m––2. Species nomenclature was checked using the World Register of Marine Species1 and all the specimens collected were deposited at the National Research-Development Institute for Marine Geology and Geoecology—GeoEcoMar.
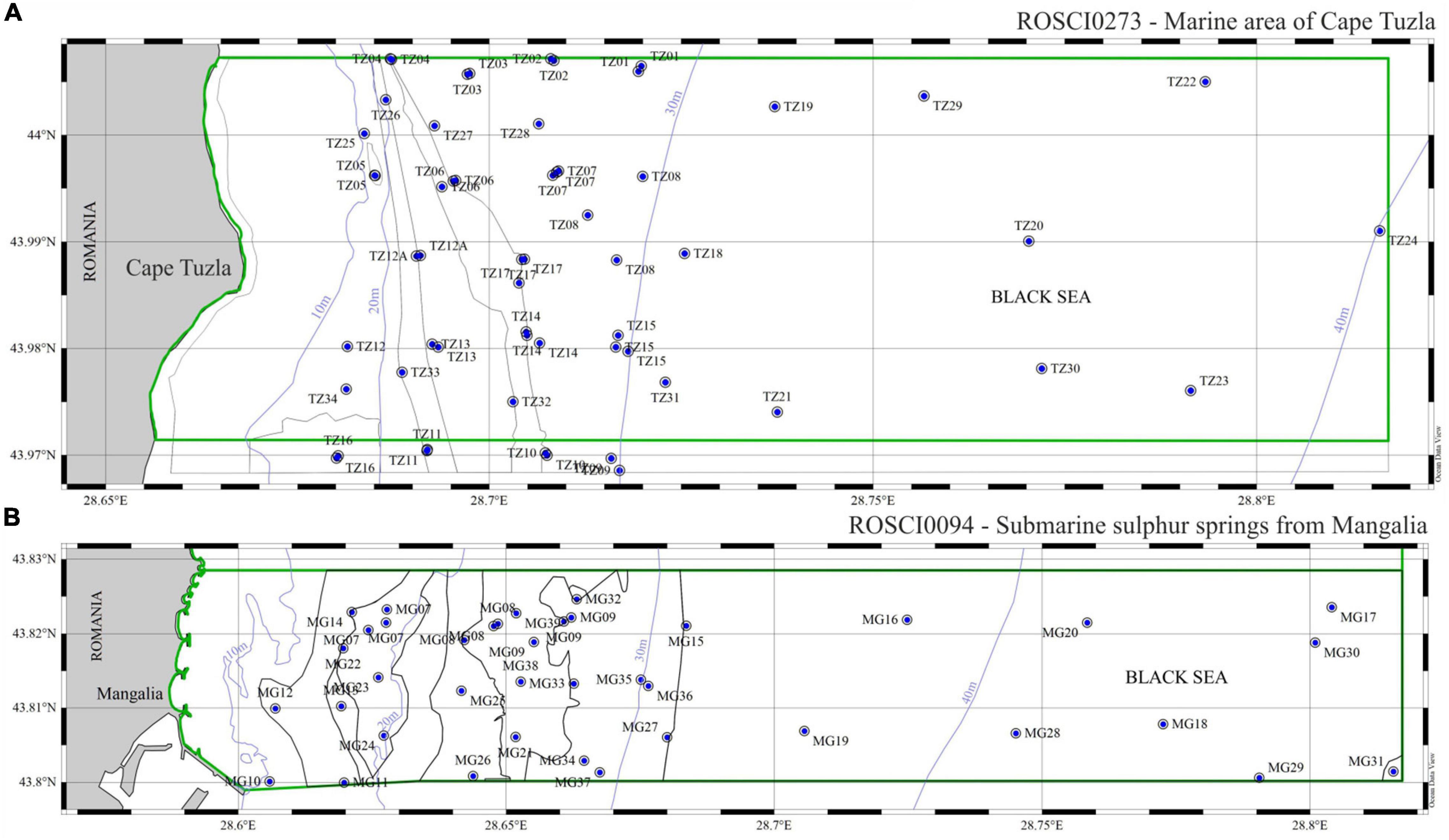
Figure 2. Maps outlining the position of the 47 samples collected in the Cape Tuzla MPA (A) and 28 in the Mangalia Sulfur spring MPA (B), between 2012 and 2018.
Macrozoobenthic community structure was analyzed using species composition (S), their density (A), dominance (D), frequency (F), diversity (H’), and biomass (B). Diversity was calculated using the Shannon-Wiener index (H’) on a log2 base (Shannon and Weaver, 1963). Spatial analyses of that data were conducted with Ocean Data View, version 5.4.0 (Schlitzer, 2018). Univariate and multivariate analyses were performed using the PRIMER 7 and PERMANOVA + software package, version 7.0.17. Multidimensional Scaling (MDS) and cluster analyses, in particular, were conducted with density and biomass, but the biomass offered a better visualization of the dissimilarity between stations and is the one presented here. The AZTI Marine Biotic Index, AMBI (Borja et al., 2000), and the multivariate AMBI, also called M-AMBI (Muxika et al., 2007), were calculated using the software (version 5.0),2 and the assignment of the identified species to the five Ecological Groups (EG) was based on the species list (June 2017) in the AMBI index software (Borja and Muxika, 2005). Subsequently, M-AMBI*(n) was calculated using the arithmetic mean of the minimum and maximum normalized AMBI, H’ and S (Sigovini et al., 2013). The use of AMBI and related indexes follows the recommendations given by National Authority Reports (Abaza et al., 2018b) and the intercalibration initiative between Romania and Bulgaria (Todorova et al., 2018). For the infralittoral and circalittoral sedimentary habitats, a threshold value of 0.68 for M-AMBI*(n) was used to discriminate between “Good Environmental Status” (GES) and non-GES (Abaza et al., 2018b).
Results
Species Composition
A total of 119 taxa were identified from both MPAs along the 7-year study period (Supplementary Material 1). Diversity was dominated by polychaetes (28.6%), followed by mollusks (25.2%), crustaceans (25.2%), and to a lesser extent by nemerteans (5%), cnidarians (5%), and “others,” encompassing a combined 11% of all other taxa identified. 95 taxa were found in the Cape Tuzla MPA, dominated by polychaetes, molluscs, and crustaceans (26, 25, and 24%, respectively). Fairly similar results were gathered in the Mangalia Sulfur springs MPA, where 98 taxa belonged to twelve groups and were dominated by the same taxa (28.6, 26.5, and 23.5%, respectively). 72 taxa (60.5%) were found in both MPAs, while 23 and 26 taxa were collected exclusively in the Cape Tuzla and the Mangalia Sulfur springs MPAs, respectively (Supplementary Material 1). Out of the species collected, five are currently listed in the Red Book of the Invertebrates from Romania (Murariu and Maican, 2021): Upogebia pusilla, Carinina heterosoma, Liocarcinus navigator, Gastrosaccus sanctus, and Brachynotus sexdentatus.
Benthic Habitat Distributions (Size and Extent) and Their Communities
Our point-sample study covered an overall area of 92% of the Cape Tuzla MPA and 90% of the Mangalia Sulfur springs MPA. Seven large benthic habitats were delineated (see below) based on the broad approach of manually drawing on a sampling point grid, following community structure in terms of presence of ecosystem engineers, their coverage, and the type of substrate. Cluster and multidimensional scaling plot (MDS) using biomass data (see section “Materials and Methods”) showed that the dissimilarity between benthic assemblages in different stations ranged from 83 to 96% (Figure 3), whereas similarity values ranged between 34 and 56% (Figure 4) (ANOSIM global R = 0.552, p = 0.1%, permutations = 999). Besides the clear spatial ordination based on bootstrap biomass, the results showed resemblance between habitats with similar type of substrate (as in the case of Infralittoral and Circalittoral coarse sediments). Moreover, according to the SIMPER analysis (see Supplementary Table 2), the species which contributed the most to the habitat differentiation were ecosystem engineers, including Mytilus galloprovincialis, Chamelea gallina, and Upogebia pusilla.
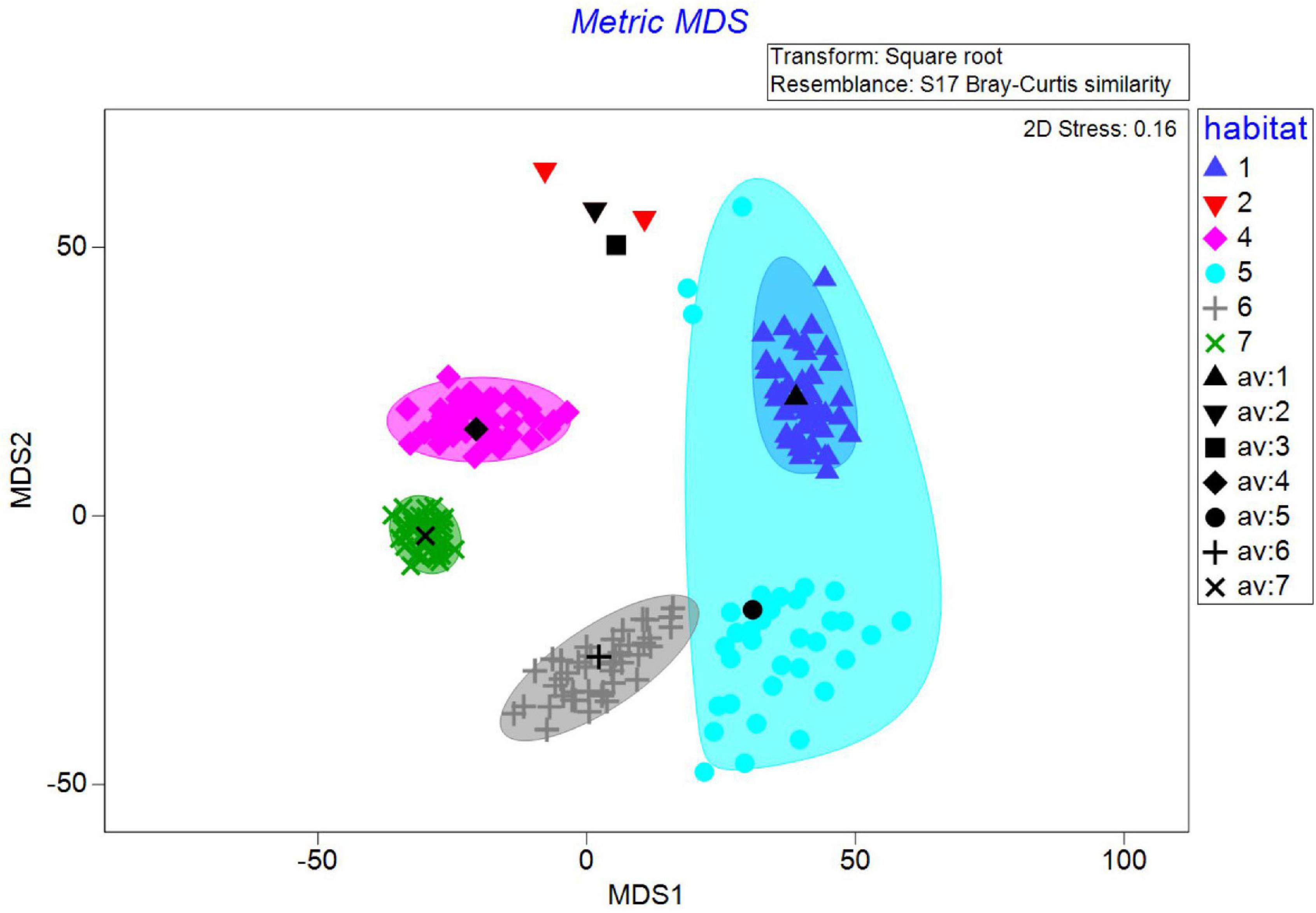
Figure 3. Two-dimensional, metric multidimensional scaling plot showing ordination of benthic habitats based on bootstrapped values calculated for 43 replications per group. Bootstrap averages (colored symbols), group averages (black symbols) and 95% confidence intervals (colored ellipses) based on bootstrap sampling with replacement are also shown. 1, Infralittoral shelly coarse sand and shellbed with various infauna; 2, Infralittoral fine and medium sand dominated by Chamelea gallina; 3, Infralittoral muddy sand with Upogebia pusilla; 4, Circalittoral mud and sandy mud with Upogebia pusilla; 5, Circalittoral shelly coarse sediment with various infauna; 6, Circalittoral mixed sediments with various infauna and mussel beds; 7, Circalittoral mud with Spisula subtruncata.
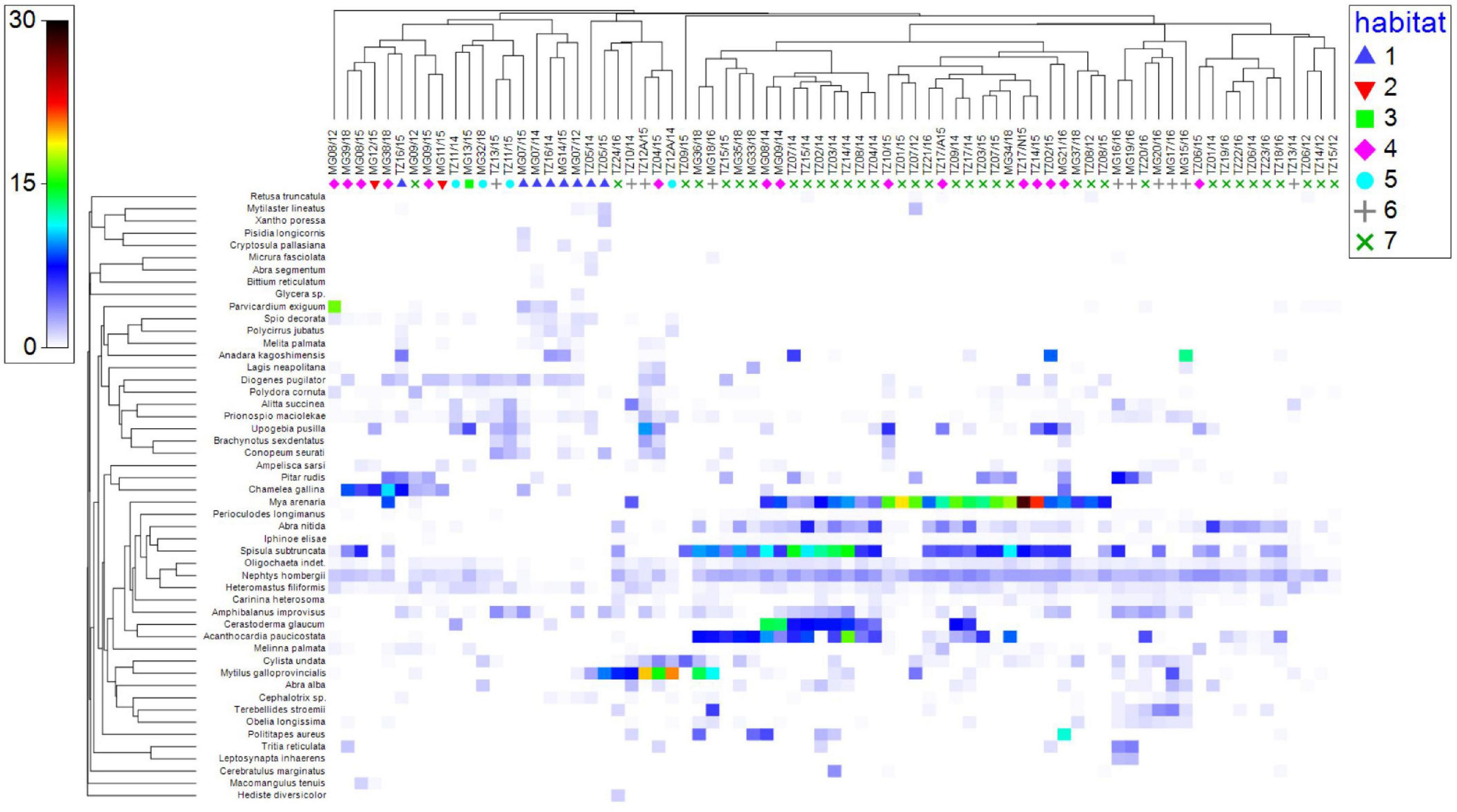
Figure 4. Shade plot of square root transformed abundances for the 50 most abundant species. Species clustering based on index of association. Habitat codes as in Figure 3.
Following the depth gradient, three infralittoral and four circalittoral habitats were distinguished (Table 2 and Figure 5). For both MPAs, the total number of species in the infralittoral and circalittoral added up to 62 and 103, respectively. Benthic communities were dominated by polychaetes (23 and 28 species in infra- and circalittoral, respectively) and crustaceans (18 and 24 species, respectively). Among polychaetes, 17 species were common to habitats present in both biozones, whereas 85% of the abundance and 75% of the biomass were accounted by the detritivorous species Heteromastus filiformis and Prionospio maciolekae and the predatory species Nephtys hombergii and Micronephthys longicornis (Supplementary Material 1). Mollusks reach the highest diversity in circalittoral habitats (27 in comparison to 11 species in the infralittoral), due mainly to soft-bottom engineering species such as Acanthocardia paucicostata, Papilicardium papillosum, Spisula subtruncata, Abra alba, Abra nitida, Pitar rudis, and Modiolula phaseolina.
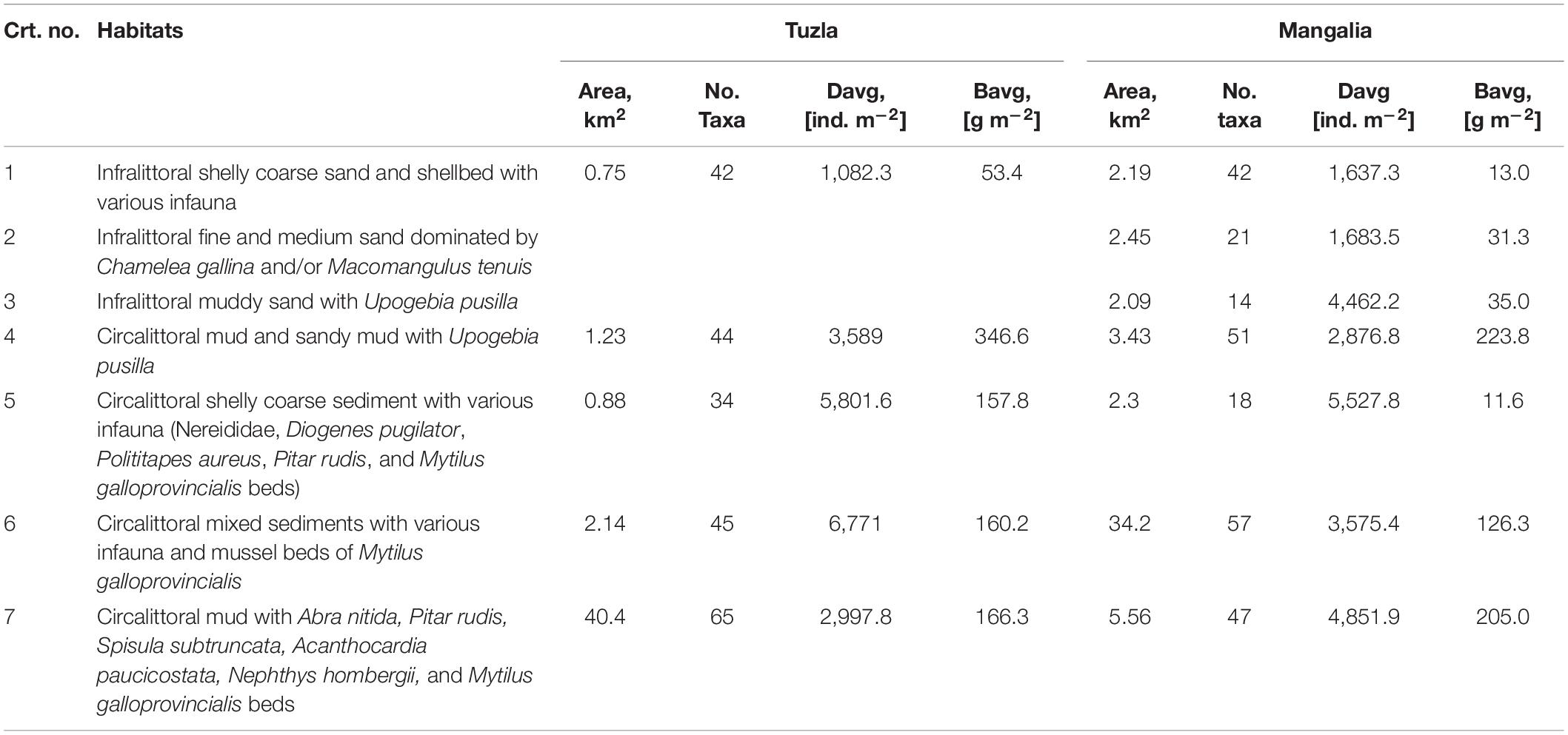
Table 2. The main habitats described in the Cape Tuzla and the Mangalia Sulfur springs MPAs, according to the EUNIS classification (Davg, average density; Bavg, average biomass).
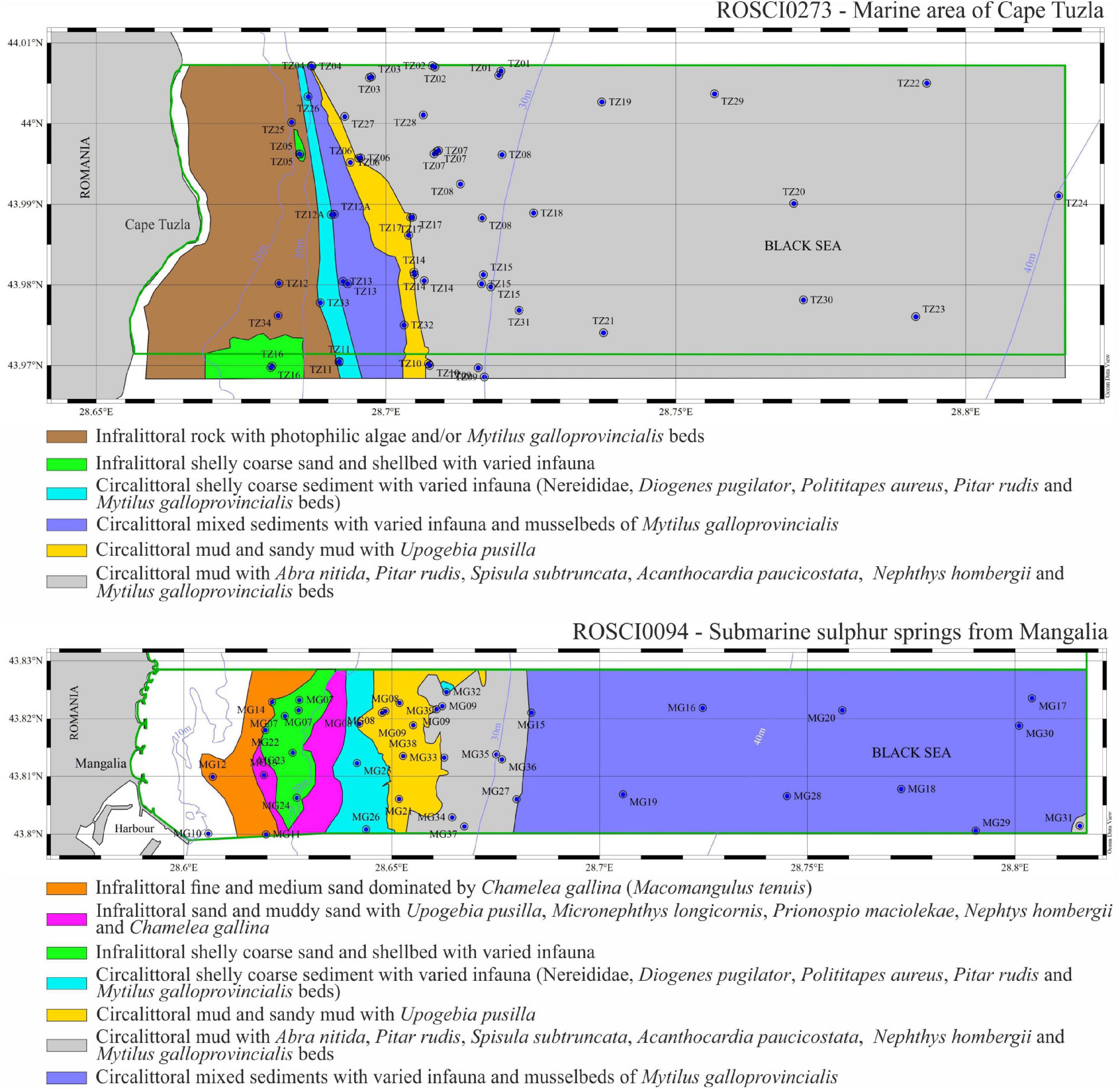
Figure 5. Maps of distribution of six benthic habitats in the ROSCI273 Cape Tuzla and seven in the ROSCI0094 Mangalia submarine sulfur springs, classified according to their sedimentological and faunistic features, based on the Romanian Broad Habitat Types (see section “Materials and Methods” for details). Purple lines are depth contours whereas green lines identify the limits of the MPA.
The infralittoral rocks with photophilic algae and/or associated Mytilus galloprovincialis reefs were treated as a major geological unit and ranged between 1 and 20 m deep (based on Bacescu et al., 1971; Teacă et al., 2010; Teacă et al., pers. obs.), with a larger breadth in the Cape Tuzla MPA than in the Mangalia Sulfur springs MPA. The seabed in the former MPA was a rugged landscape marked by boulders of different sizes, whereas the seabed in the latter was rather flat in its upper part and more irregular toward deeper seafloor. Nearby and as a continuation of these reefs, the following benthic habitats were described (Table 2):
Habitat (1): Infralittoral shelly coarse sand. A habitat built from biogenic material (bivalve shells) from nearby rocky communities, supporting two prominent associations: M. galloprovincialis and its epibenthos (Amphibalanus improvisus, Dysidea fragilis, Obelia sp.) and interstitial tube-dwelling polychaetes and vagile crustaceans (P. maciolekae, H. filiformis, and Melita palmata), forming patchy habitats covering < 5% of the MPAs areas. Besides, these patchy habitats sustained high diversities (42 species), dominated by detritivorous polychaetes (80% of the abundance) and suspension feeding mollusks (80% of the biomass) (Figures 6, 7 and Supplementary Material 1).
(2): Infralittoral fine and medium sand dominated by Chamelea gallina. The rocky infralittoral facies favors the accumulation of sandy sediments and the formation of this shallow habitat (5–18 m deep), which is dominated by a thriving specialized community formed of Chamelea gallina and Ampelisca sarsi along with the polychaetes M. longicornis and H. filiformis. Species richness amounted to ∼20 species (Table 2), dominated numerically by annelids and by mollusks in terms of biomass. This habitat covered 4% of the Mangalia Sulfur springs MPA.
(3): Infralittoral muddy sand. Infra- and circalittoral coarser habitats border this small habitat (17–23 m deep; 3.62% of the area of the Mangalia Sulfur springs MPA; Figures 6, 7), characterized mainly by Upogebia pusilla which reached up to 118 ind. m–2 (mean: 14.8 ind. m–2). Detritivorous annelids (H. filiformis and oligochaetes) numerically dominated this habitat (3,641 ind. m–2).
(4): Circalittoral mud and sandy mud with U. pusilla. An extension into deeper waters of the prior habitat, where muddy sediment species such as Mya arenaria, Abra nitida, and Spisula subtruncata prevailed (> 40% of biomass; Figures 6, 7) at the expense of U. pusilla (∼14% only). Annelid composition was similar to the previous habitat with the exception of N. hombergii, which here accounted for 70% of the average density. This habitat holds a higher species diversity (61 taxa) than the infralittoral (14).
(5): Circalittoral shelly coarse sediment with various infauna. This habitat followed the configuration of the circalittoral hard bottoms and was formed by clean shell debris deposited at the edge of rocky bottoms (21–23 m deep). Infauna was numerically dominated (78%) by the polychaetes P. maciolekae, M. longicornis, and H. filiformis, while the biomass was dominated (> 35%) by M. galloprovincialis. P. maciolekae was the first ranked species in terms of density (60% in circalittoral and 30% in infralittoral, respectively).
(6): Circalittoral mixed sediments with various infauna. The largest habitat in the Mangalia Sulfur springs MPA (34.2 km2) but considerably smaller in the Cape Tuzla MPA, due to steep bathymetry and deflection of fine sediments and nutrients further to the south-east. The heterogeneity of this habitat supports various trophic groups including filter feeders, grazers, detritivores and predatory species (72 species and 4,800 ind. m–2) (Figures 6, 7). Mussel beds constitute the most important habitats, and account for the highest biomass among all mollusk fauna (∼80%).
(7): Circalittoral mud. As a result of hydrodynamics, this habitat covered the largest area in the Cape Tuzla MPA (40.4 vs. 5.56 km2 in the Mangalia Sulfur springs MPA). Such conditions were reflected by the change of dominant mollusk communities, formed primarily by infaunal species such as Abra nitida, Pitar rudis, Spisula subtruncata, and Acanthocardia paucicostata, which dominated the density and biomass (366 ind. m–2 and 90.44 g m–2, respectively) and supported a species richness of 74 taxa in both MPAs.
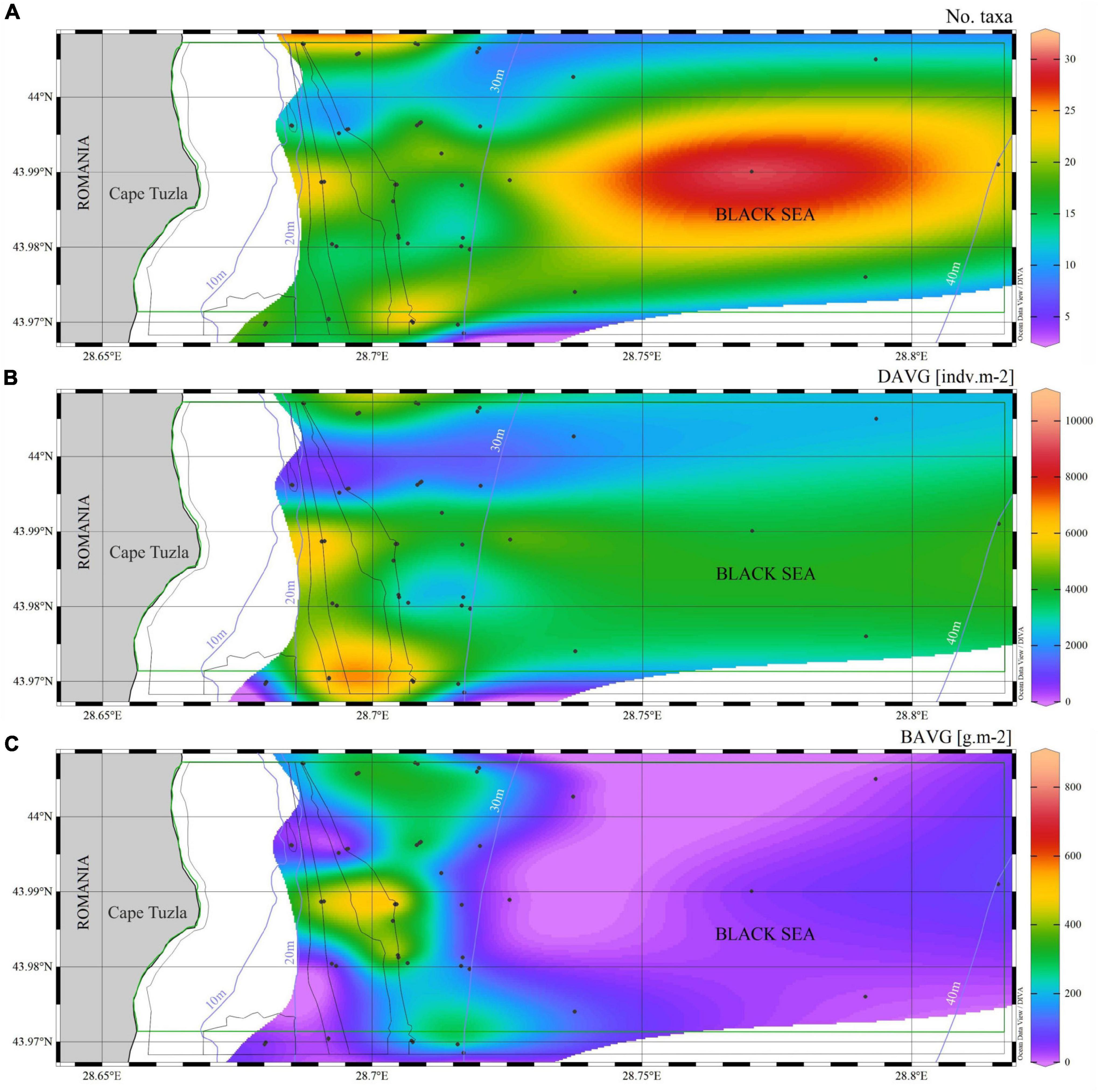
Figure 6. Contour maps showing the variation in (A) the number of macrozoobenthic taxa, (B) their mean density and (C) mean biomass in the Cape Tuzla MPA, based on samples (round symbols) collected between 2012 and 2018.
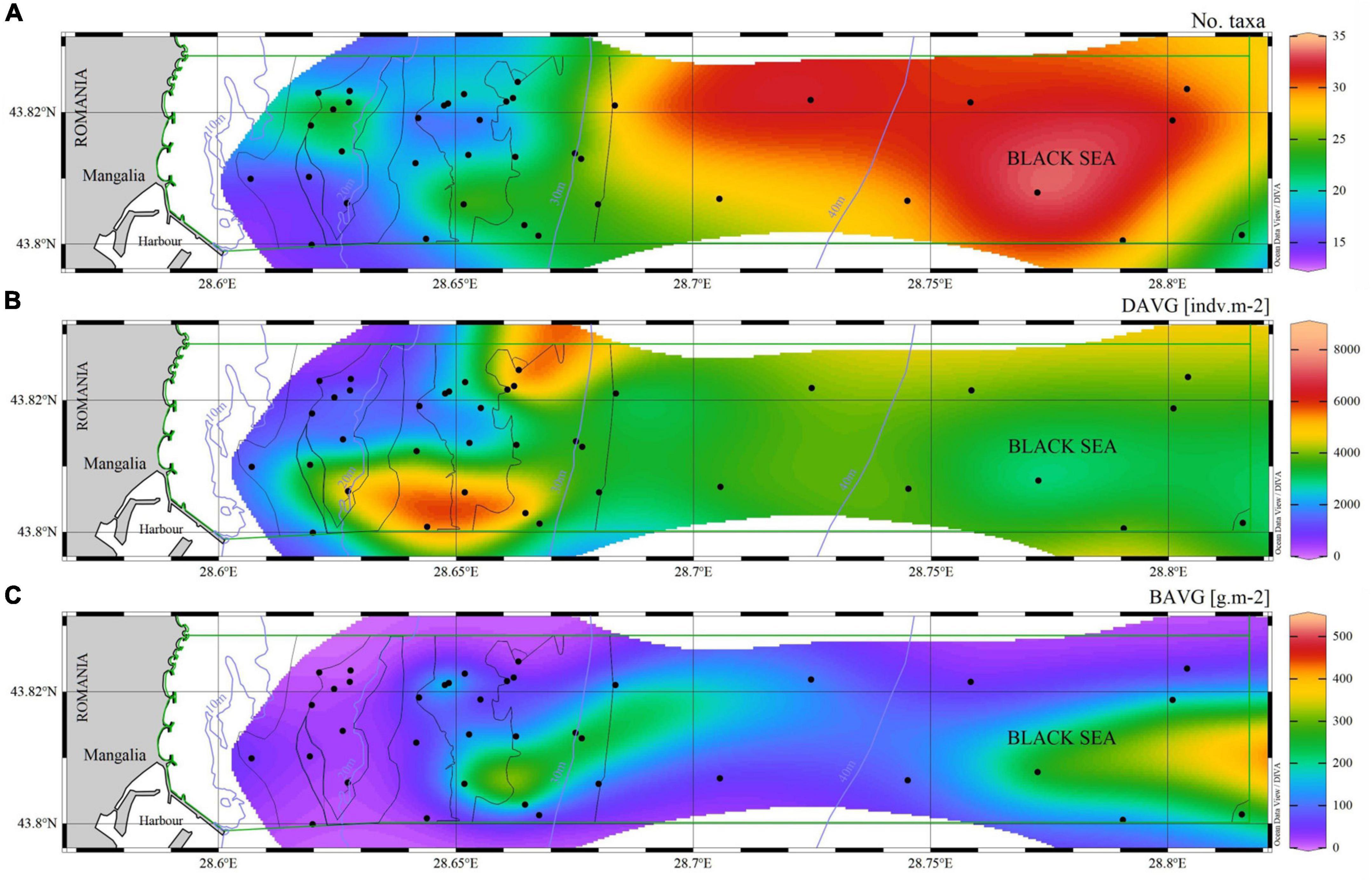
Figure 7. Contour maps showing the variation in (A) the number of macrozoobenthic taxa, (B) their mean density and (C) mean biomass in the Mangalia submarine Sulfur springs MPA, based on samples (round symbols) collected between 2012 and 2018.
Environmental Assessment
The AMBI values for the 75 stations ranged between 0.72 and 5.14, indicating that macrobenthic communities from 25 stations (33.3%) achieved GES and that from 50 stations (66.6%) did not achieved GES. M-AMBI*(n) values ranged between 0.32 and 1.00. Based on this index, GES was assigned to a total of 50 stations and non-GES to 25 stations (Figure 8). M-AMBI*(n) values were generally lower in offshore (circalittoral) than in inshore (infralittoral) bottoms. The circalittoral mud habitat achieved a 54% GES, while other circalittoral habitats (sandy mud, shelly coarse sediment, or mixed sediment) reached between 70 and 100% of GES. Infralittoral shelly coarse sand and shellbed with various infauna and infralittoral fine and medium sand dominated by Chamelea gallina habitats achieved 88 and 60% of GES, respectively. In general, relatively high local diversity (20–35 species in average per habitat) translated into high values of H’ and M-AMBI*(n) index. Only the infralittoral muddy sand habitat accounted for a 100% non-GES due to the small number of samples available in this habitat (Figure 8).
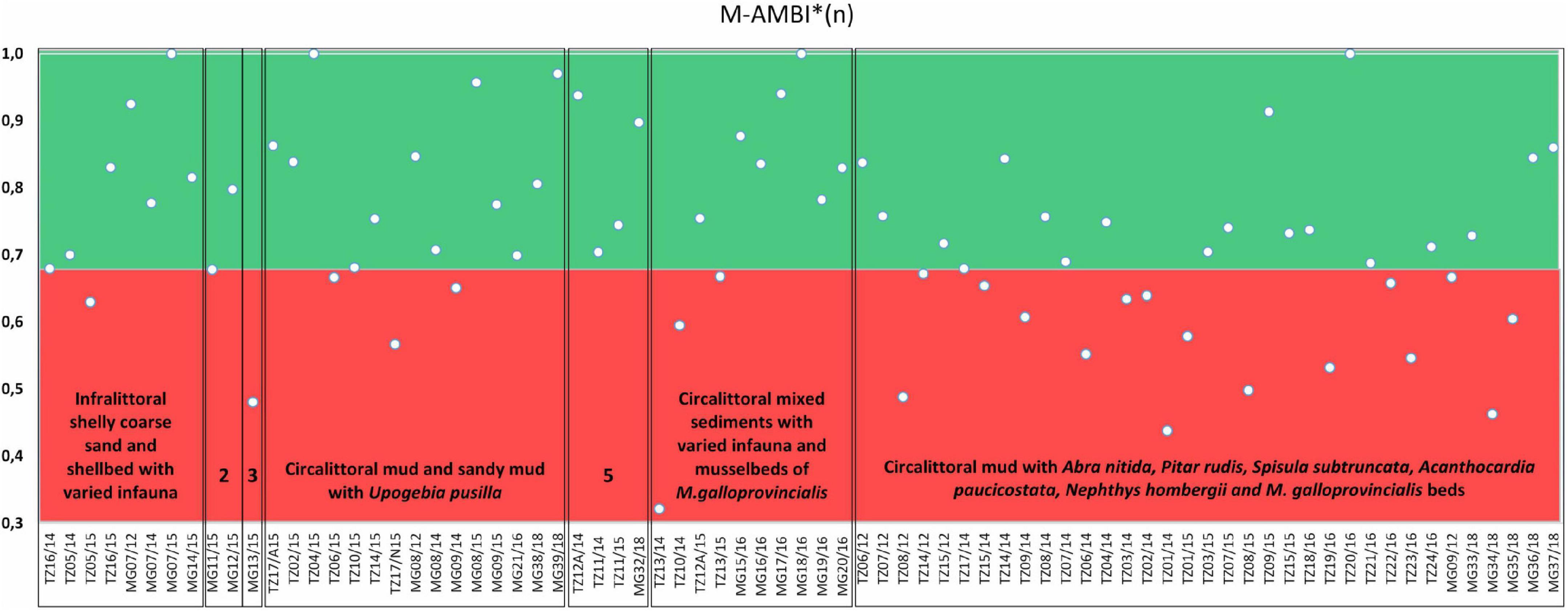
Figure 8. Diagram illustrating the variation in ecological state in the seven habitats analyzed based on M-AMBI*(n) values (Sigovini et al., 2013). The background colors distinguish samples from both MPAs in good ecological status (GES; green) and non-GES (red).
The proportion of ecological groups (EG) to which benthic species were allocated, showed the dominance of species “sensitive” and “indifferent” to organic pollution in infralittoral habitats (Chamelea gallina, Mytilaster lineatus, Parvicardium exiguum, Apherusa bispinosa, Melita palmata, Microdeutopus versiculatus, Iphinoe tenella, Diogenes pugilator, Upogebia pusilla, and Liocarcinus vernalis). In these habitats, 7–57% of the species were considered sensitive and 13–75% of the species were considered “indifferent.” In contrast, first-order “opportunistic” species (Heteromastus filiformis, Oligochaeta) were better represented in the circalittoral mud habitat (15–69%).
Discussion
Matching the description (mapping) of basic ecological patterns with the management of coastal areas is not an aspiration but rather a pressing need for society (Carr et al., 2019). The spatial analyses performed in this study can be used to elaborate and implement management measures focused on achieving habitat conservation and, for this particular region, to comply with the European directives focused on the management of marine resources and habitat protection. Habitat typology with detailed data on extent and areal cover, are expected to contribute to the assessment and design of future programs that monitor the status of the seabed (Sokolowski et al., 2021).
Benthic Habitats and Associated Diversity
Mapping provides basic information about the distribution, pattern, extent and area of habitats of conservation importance, and through that supports spatial planning and ecosystem-based management of MPAs (Todorova et al., 2015). Our study encompassed habitats associated with the first two (out of five) biological benthic zones that have been identified in the Black Sea: infralittoral, circalittoral, offshore circalittoral, bathyal, and abyssal. The two first zones, infralittoral, and circalittoral bottoms, represent almost 14% of the entire Black Sea seafloor, and ∼2.3% of the Romanian coastline. The soft-bottoms associated with infralittoral and circalittoral habitats are affected by wave action (2.9 N/m2), whilst the subsequent zones, offshore circalittoral, bathyal and abyssal bottoms, only start at the sea-bottom 9.7°C isotherm (at about 50 m depth) (Populus et al., 2017; Vasquez et al., 2020).
The seven habitats described in this study span only 107.3 km2 of the infralittoral and circalittoral zones, a relatively small surface when compared with the extent of lifeless areas like the anoxic bathyal and abyssal (Populus et al., 2017). Nevertheless, the ecological roles of these habitats are deemed essential for the overall health of the Black Sea. Important benthic species and communities associated with these habitats provide goods (such as food provision), cultural services (e.g., recreational diving in biogenic reefs) and have key regulatory functions including sediment bio-irrigation, reworking and mobilization, in addition to serving as carbon and nutrient sinks (see Lenihan and Peterson, 1998; Aller and Cochran, 2019).
Hard bottoms constituted a continuous geological unit stretching along both MPAs, and which probed to be associated with several types of benthic habitats. The main habitat linked to the rocky facies is the Infralittoral rock with photophilic algae and/or Mytilus galloprovincialis reefs, ranging between 1 and 20 m deep (Bacescu et al., 1971; Teacă et al., 2010). In the Mangalia Sulfur springs MPA, the rocky facies reach slightly deeper bottoms in the circalittoral, causing the partial overlap with the Infralittoral and Circalittoral coarse habitats, whose precise limits remain to be defined at a later stage using remote sensing. It is also noteworthy to mention that some species that inhabit shallow coarse sediments in the infralittoral and circalittoral bottoms (e.g., H. filiformis and U. pussila), exhibit a large biotope plasticity, with no apparent preferences for infralittoral sand or circalittoral muddy sands. However, such plasticity warrants further research attempting to understand the dynamics of species colonizing or migrating among distinct habitats.
In spite of their small surface (0.75–2.14 km2), the infralittoral as well as the circalittoral mud, coarse and mixed habitats, sustained a significant pool of benthic diversity. The infralittoral habitats (down to 20 m depth) had been already the subject of prior mapping research for the ‘‘Management Plan of the Tuzla and Mangalia Site of Community Importance,’’ as defined according to the Natura 2000 classification.3 However, no data was available before our study on macrozoobenthic diversity, their distribution, and benthic habitat assessment for these MPAs. Highlighting the contribution of this study also entails acknowledging its limitations: due to feasibility, not every habitat could be sampled with the same intensity. Still, this is an issue that should promote further studies on habitats still understudied, for which this study provides the first available information.
Over 80% of the species included in the Red Book of the Invertebrates from Romania are found in the infralittoral habitats (< 20 m) of the southern Romanian Black Sea coast (Murariu and Maican, 2021). Some of these species, including Dysidea fragilis, Halichondria panicea, Petricola lithophaga, Gastrana fragilis, Flexopecten glaber ponticus, Donax trunculus, Pholas dactylus, Palaemon adspersus, Eriphia verrucosa, had been already mentioned in various studies in the region (Begun et al., 2006; Teacă et al., 2006a,b, 2010; Filimon, 2020; Surugiu et al., 2021; Teacă, pers. communication). However, our study collected only a fraction of the species (5) included in the Red Book of the Invertebrates from Romania, likely due to the rather small surface of the investigated infralittoral bottoms.
To the best of our knowledge, our study provides the most detailed analysis of benthic diversity at Romanian littoral MPAs to date. This study also documents some of the highest diversity levels (a grand total of 119 taxa), considering that previous studies (i.e., Abaza et al., 2019) have reported slightly lower diversities (113 taxa) from an area encompassing all the Romanian benthic habitats. Our study is comparable with the studies conducted at the Bulgarian littoral, where fine-scale surveys mapped benthic habitats within the framework of the MSFD (Todorova et al., 2015, 2020; Nikolopoulou et al., 2021). Similar to our results, those studies revealed a good compliance between delineation of distinct benthic habitats and the presence of relevant engineering species contributing to biomass as well as both density and diversity. Such approach has served as a common assessment scheme across nations for the Black Sea EUSeaMap habitats mapping (Populus et al., 2017; Vasquez et al., 2020). It follows that several of the habitats identified within the two Romanian MPAs studied here, correspond with those described from the nearby Bulgarian coast (i.e., infralittoral sand dominated by Chamelea gallina, circalittoral mud with Spisula subtruncata, etc.). Such correspondence suggests the potential for upscaling the current study to assess regional interconnectivity among similar benthic habitats across the Black Sea region.
Environmental Assessment
The Cape Tuzla and the Mangalia Sulfur springs MPAs are wide open coastal areas exposed to a variety of anthropogenic activities (Begun et al., 2018). Shipping, tourism, and pelagic and bottom fishing can individually or in combination cause environmental disruptions and likely have an impact on benthic habitats (Begun et al., 2018). In fact, due to their small spatial extent and/or their vulnerability to environmental stressors, several species inhabiting both the infralittoral and circalittoral habitats have received a protection status by being included in the afore-mentioned Red Book of the Invertebrates from Romania (Murariu and Maican, 2021). Invasive species may also represent a threat, depending on the nature of their interactions with resident (native) species (see Gurevitch and Padilla, 2004; Quijón et al., 2017). In this area, in particular, several invaders have become established and do represent a serious threat (Dipolydora quadrilobata, Polydora cornuta, Streblospio gynobranchiata, Corambe obscura, Anadara kagoshimensis, Mya arenaria, and Amphibalanus improvisus) to both vulnerable species and the diversity of this coastline. Among them, the latest invader with rapid spreading behavior detected in Romanian benthic habitats is the spionid polychaete Streblospio gynobranchiata (Teacă et al., 2021).
Even though those stressors amount to a certain risk for the diversity in the Romanian Black Sea, these coastal habitats seem more severely affected by eutrophication and industrial pollution (Todorova et al., 2020). Despite that, our study indicates that 88 and 60% of the habitats and associated communities in the infralittoral of the Cape Tuzla and the Mangalia Sulfur springs MPAs, respectively, are achieving good ecological status (GES). Meanwhile, only ∼54% of the habitats and communities associated with circalittoral bottoms are achieving GES. The latter result contrasts with the GES of other habitats from the circalittoral (namely from mud and sandy mud, shelly coarse sediment and mixed sediment) which are achieving between 70 and 100% GES due to their higher biodiversity. These differences suggest that benthic habitats are for the most part resilient to the anthropogenic stressors present in this area, but also imply that there are particular habitats that do require further attention. We therefore call for further research to be conducted on specific sources of stress (first and foremost organic and industrial pollution) and the response of sensitive species to short- and long-term exposure to these stressors.
Recognizing the importance of mapping of benthic habitats in MPAs such as those studied here in the southern Romanian Black Sea, would be a key step toward better managing these areas. The unique biological value and aesthetic quality of the areas currently protected by the MPAs cannot oppose the reasonable use of these waters. Habitat mapping constructed from the combination of seafloor characteristics and main benthic community features represents a useful tool and therefore should play a critical role on the management of these systems. Seafloor structure and the species that colonize these habitats, even at relatively small spatial scales (e.g., Surugiu et al., 2021), not only maintain the function of these systems, but also act as integrative indicators of their quality and health. It follows that seafloor structure and function should be an integral part of the framework to integrate research and management programs (Sullivan et al., 2017). Such approach would alleviate some of the concerns that the larger temporal and spatial scope of our study makes evident, and that should be effectively communicated to coastal managers.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Author Contributions
TB and AT: conceptualization, methodology, data curation, software, writing original draft preparation, reviewing and editing, supervision, and funding acquisition. MM and SM: methodology, data curation, writing original draft preparation, reviewing and editing. PQ: conceptualization, writing original draft preparation, reviewing and editing. VS: writing original draft preparation, reviewing and editing. All authors contributed to the article and approved the submitted version.
Funding
This study was financially supported by the Romanian Ministry of Research, Innovation and Digitization in the framework of the national CORE Program projects: PN 19200302, PN 19200101 and the Program Development of the National R&D system—AMBI AQUA—No. 23PFE. TB, AT, and MM were supported by the Contract—EASME/EMFF/2020/3.1.ll/Lot3/SI2.843624 and the European Union’s Horizon 2020 BRIDGE-BS project under grant agreement no. 101000240. PQ acknowledges the continuous support of NSERC-Canada, during the preparation of this manuscript.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank all the colleagues who helped at sample sorting and made suggestions for the improvement of earlier versions of the manuscript. Likewise, we thank the valuable comments of two reviewers.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2022.845507/full#supplementary-material
Footnotes
- ^ www.marinespecies.org
- ^ http://ambi.azti.es
- ^ http://ananp.gov.ro/wp-content/uploads/PM-SCI-Word.pdf
References
Abaza, V. (2010). Zoobenthic communities structure in the eastern part of the marine reserve 2 Mai–Vama Veche under present ecological conditions J. Environ. Prot. Ecol. 11, 1013–1023.
Abaza, V., Dumitrache, C., Filimon, A., and Marin, O. (2019). Present ecological status of the main benthic habitats from the Romanian Black Sea Shelf. Cercet. Mar. 49, 74–91.
Abaza, V., Dumitrache, C., Spinu, A., and Filimon, A. (2018a). Ecological quality assessment of circalittoral major habitats using M-AMBI*(n) index. J. Environ. Prot. Ecol. 19, 564–572.
Abaza, V., Filimon, A., and Dumitrache, C. (2018b). IV.4. D1, 6 – Biodiversitate – Habitatele Bentale – Macrozoobentos, In Raportul National Privind Starea Ecologicã A Ecosistemului Marin Marea Neagrã Conform Cerinţelor Art. 17 Ale Directivei Cadru Strategia Pentru Mediul Marin (2008/56/EC). (Constanta: NIRD Antipa). 283–301.
Abaza, V., and Godeanu, S. (2008). Present state of shallow water macrozoobenthos from the marine reserve 2 mai – vama veche. Cercet. Mar. 38, 101–118.
Airoldi, L., and Beck, M. W. (2007). Loss, status and trends for coastal marine habitats of Europe. Oceanogr. Mar. Biol. Ann. Rev. 45, 345–405. doi: 10.1201/9781420050943.ch7
Aller, R. C., and Cochran, J. K. (2019). The critical role of bioturbation for particle dynamics, priming potential, and organic C remineralization in marine sediments: local and basin scales. Front. Earth Sci. 7:157. doi: 10.3389/feart.2019.00157
Bacescu, M., Müller, G. I., and Gomoiu, M.-T. (1971). “Cercetari de ecologie bentala in Marea Neagra (analiza cantitativa, calitativa si comparata a faunei bentale pontice),” in Ecologie Marina, Vol. ed. R. S. R. Acad (Bucuresti: Editura Academiei Republicii Socialiste România), 4, 1–357.
Begun, T., Teacă, A., and Gomoiu, M.-T. (2010). State of macrobenthos within Modiolus phaseolinus biocoenosis from Romanian Black Sea Continental shelf. Geoecomarina 16, 5–18.
Begun, T., Teacă, A., Gomoiu, M.-T., and Paraschiv, G. M. (2006). Present state of the sandy invertebrate populations in the Mamaia and Mangalia sector of the Romanian Black Sea coast. Geoecomarina 12, 67–77.
Begun, T., Teaca, A., Muresan, M., Vasiliu, D., Secrieru, D., and Pavel, B. (2018). Environmental assessment of two MPAs from the Romanian Black Sea Coast. J. Environ. Prot. Ecol. 19, 573–582.
Bodeanu, N. (2002). Algal blooms in Romanian Black Sea waters in the last two decades of the XXth century. Cercet. Mar. 34, 7–22.
Bologa, A. S., Bodeanu, N., Petran, A., Tiganus, V., and Zaitsev, Y. P. (1995). Major modifications of the Black Sea benthic and planktonic biota in the last three decades. Bull. Inst. Oceanogr. 1, 85–110.
Borja, A., Franco, J., and Perez, V. (2000). A marine biotic index to establish the ecological quality of soft-bottom benthos within European estuarine and coastal environments. Mar. Pollut. Bull. 40, 1100–1114. doi: 10.1016/S0025-326X(00)00061-8
Borja, A., and Muxika, I. (2005). Guidelines for the use of AMBI (AZTI’s Marine Biotic Index) in the assessment of the benthic ecological quality. Mar. Pollut. Bull. 50, 787–789. doi: 10.1016/j.marpolbul.2005.04.040
Cabral, R. B., Bradley, D., Mayorga, J., Goodell, W., Friedlander, A. M., Sala, E., et al. (2020). A global network of marine protected areas for food. Proc. Natl. Acad. Sci. U.S.A. 117, 28134–28139. doi: 10.1073/pnas.2000174117
Carr, M. H., White, J. W., Saarman, E., Lubchenco, J., Milligan, K., and Caselle, J. E. (2019). Marine protected areas exemplify the evolution of science and policy. Oceanography 32, 94–103. doi: 10.5670/oceanog.2019.315
Claudet, J., and Fraschetti, S. (2010). Human-driven impacts on marine habitats: a regional meta-analysis in the Mediterranean Sea. Biol. Conserv. 143, 2195–2206. doi: 10.1016/j.biocon.2010.06.004
Cociaşu, A., Lazăr, L., and Vasiliu, D. (2008). New tendency in nutrients evolution from coastal waters. Cercet. Mar. 38, 7–24.
Danovaro, R., Gambi, C., Dell’Anno, A., Corinaldesi, C., Fraschetti, S., Vanreusel, A., et al. (2008). Exponential decline of deep-sea ecosystem functioning linked to benthic biodiversity loss. Curr. Biol. 18, 1–8. doi: 10.1016/j.cub.2007.11.056
Duţu, F., Panin, N., Ion, G., and Tiron Duţu, L. (2018). Multibeam bathymetric investigations of the morphology and associated bedforms. Sulina Channel. Danube Delta. Geosciences 8:7. doi: 10.3390/geosciences8010007
EC (2020). EU Biodiversity Strategy for 2030. Bringing Nature Back Into our Lives. 20.5.2020 COM (2020) 380 Final. (Brussels: EC).
Filimon, A. (2020). First record of Flexopecten glaber (Linnaeus, 1758) (Bivalvia: Pectinidae) from the Romanian Black Sea shelf. Cercet. Mar. Res. Mar. 50, 186–191.
Filimon, A., Abaza, V., Marin, O., and Dumitrache, C. (2016). Community structure of zoobenthos associated with Cystoseira barbata facies from the Southern Romanian Black Sea coast. J. Environ. Prot. Ecol. 17, 942–949.
Friedrich, J., Janssen, F., Aleynik, D., Bange, H. W., Boltacheva, N., Çagatay, M. N., et al. (2014). Investigating hypoxia in aquatic environments: diverse approaches to addressing a complex phenomenon. Biogeosciences 11, 1215–1259. doi: 10.5194/bg-11-1215-2014
Gomoiu, M.-T. (1983). Sur la mortalité en masse des organismes benthiques du littoral roumain de la mer Noire. Rapp. Comm. Int. Mer. Médit. 28, 203–204.
Gomoiu, M.-T. (1992). Marine Eutrophication Syndrome in the North–Western Part of the Black Sea. Science of the Total Environment. Suppl. (Amsterdam: Elsevier Science PublishersB.v). 683–692.
Gomoiu, M.-T. (1998). “Some remarks concerning the evaluation of ecological diversity at the Romanian Black Sea coast,” in Conservation of the Biological Diversity as a Prerequisite for Sustainable Development in the Black Sea Region. NATO ASI Series 2 Environmental Security, eds V. Kotliakov, M. Uppenbrink, and V. Metreveli (Dordrecht: Kluwer Academic Publishers), 77–93. doi: 10.1007/978-94-011-5114-6_5
Gomoiu, M.-T., Alexandrov, B., Shadrin, N., and Zaitsev, Yu.P (2002). “The Black Sea – A recipient, donor and transit area for alien species,” in Invasive Aquatic Species of Europe. Distribution, Impacts and Management, eds E. Leppakoski, S. Gollasch, and S. Olenin (Dordrecht: Kluwer), 341–350. doi: 10.1007/978-94-015-9956-6_35
Gomoiu, M.-T., Secrieru, D., Paraschiv, G., Opreanu, P., and Puschiaza, D. (2003/2004). Some remarks on the ecological state of benthic populations recorded during the IAEA ’98 Black Sea cruise of “prof. Vodyanitskiy”. R/V. Geoecomarina 9, 1–12.
Gurevitch, J., and Padilla, D. K. (2004). Are invasive species a major cause of extinctions? Trends Ecol. Evol. 19, 470–474. doi: 10.1016/j.tree.2004.07.005
Halpern, B. S., Frazier, M., Potapenko, J., Casey, K. S., Koenig, K., Longo, C., et al. (2015). Spatial and temporal changes in cumulative human impacts on the world’s ocean. Nat. Commun. 6:7615. doi: 10.1038/ncomms8615
Halpern, B. S., Walbridge, S., Selkoe, K. A., Kappel, C. V., Micheli, F., D’Agrosa, C., et al. (2008). A global map of human impact on marine ecosystems. Science 319:948. doi: 10.1126/science.1149345
Hillebrand, H., Blasius, B., Borer, E., Chase, J., Downing, J., Eriksson, B. K., et al. (2017). Biodiversity change is uncoupled from species richness trends: consequences for conservation and monitoring. J. Appl. Ecol. 55, 169–184. doi: 10.5061/dryad.1sd59
IPBES (2019). Summary for Policymakers of the Global Assessment Report on Biodiversity and Ecosystem Services of the Intergovernmental Science–Policy Platform on Biodiversity and Ecosystem Services. (Bonn: IPBES).
Lenihan, H. S., and Peterson, C. H. (1998). How habitat degradation through fishery disturbance enhances impacts of hypoxia on oyster reefs. Ecol. Appl. 8, 128–140. doi: 10.1890/1051-0761(1998)008[0128:HHDTFD]2.0.CO;2
Leppäkoski, E., and Mihnea, P. E. (1996). Enclosed seas under man-induced change: a comparison between the Baltic and Black Seas. Ambio 25, 380–389.
Mee, L. D. (1992). The Black Sea in crisis: a need for concerted international action. Ambio 21, 278–286.
Mee, L. D., Friedrich, J., and Gomoiu, M. T. (2005). Restoring the Black Sea in times of uncertainty. Oceanography 18, 100–111. doi: 10.5670/oceanog.2005.45
Mee, L. D., and Topping, G. (1999). Black Sea Pollution Assessment. Black Sea Environmental Series 10:380.(New York, NY: United Nations Publications)
Moncheva, S., Petrova-Karadjova, V., and Palasov, A. (1995). “Harmful algal blooms along the Bulgarian Black Sea coast and possible pattern of fish and zoobenthic mortalities,” in Harmful Marine Algal Blooms, eds P. Lassus, G. Arzul, E. Denn, and P. Gentien (Lavoisier: Intercept Ltd).
Murariu, D., and Maican, S. (eds). (2021). The Red Book of the Invertebrates from Romania. (Bucharest: Romanian Academy Publishing House).
Mureşan, M., Begun, T., and Teacă, A. (2020). Contribution of spring and early autumn zooplankton populations to the Romanian Black Sea waters productivity, in 2019. Sci. Pap. Ser. D Anim. Sci. 63, 503–511.
Mureşan, M., and Teacă, A. (2019). The free-living nematode community structure within the Romanian circalittoral habitats. Geoecomarina 25, 7–13.
Mureşan, M., Teacă, A., Popa, A., and Begun, T. (2019). Free-living marine nematodes community structural changes within a post-dredging site at the Romanian shelf. J. Environ. Prot. Ecol. 20, 753–760.
Muxika, I., Borja, A., and Bald, J. (2007). Using historical data, expert judgement and multivariate analysis in assessing reference conditions and benthic ecological status, according to the European water framework directive. Mar. Pollut. Bull. 55, 16–29. doi: 10.1016/j.marpolbul.2006.05.025
Nikolopoulou, S., Berov, D., Klayn, S., Dimitrov, L. I., Velkovsky, K., Chatzinikolaou, E., et al. (2021). Benthic habitat mapping of Plazh Gradina – Zlatna ribka (Black Sea) and Karpathos and Saria Islands (Mediterranean Sea). Biodivers. Data J. 9:e71972. doi: 10.3897/BDJ.9.e71972
Niţă, V., Zaharia, T., Nenciu, M., Cristea, M., and T̨iganov, G. (2012). Current state overview of the Vama Veche - 2 Mai Marine Reserve. Black Sea, Romania. Aqua. Aquar. Conserv. Legis. 5, 44–54.
Panin, N. (2005). The Black Sea coastal zone – an overview. European seas: coastal zones and rivers – sea system. Geoecomarina 11, 21–40.
Panin, N., Gomoiu, M.-T., and Oaie, Gh, and Radan, S. (1996). Danube Delta-Black Sea system under global changes impact. Geoecomarina 1, 90–114.
Panin, N., and Jipa, D. (1998). Danube River sediment input and its interaction with the North-western Black Sea: results of EROS-2000 and EROS-21 projects. Geoecomarina 3, 23–35.
Petranu, A. (1997). Black Sea Biological Diversity – Romania, Black Sea Environmental. (Berlin: Springer) 4..
Plumeridge, A. A., and Roberts, C. M. (2017). Conservation targets in marine protected area management suffer from shifting baseline syndrome: a case study on the Dogger Bank. Mar. Pollut. Bull. 116, 395–404. doi: 10.1016/j.marpolbul.2017.01.012
Populus, J., Vasquez, M., Albrecht, J., Manca, E., Agnesi, S., Al Hamdani, Z., et al. (2017). EUSeaMap. A European Broad–Scale Seabed Habitat Map. (Brest: Ifremer). doi: 10.13155/49975
Quijón, P. A., Tummon Flynn, P., and Duarte, C. (2017). Beyond negative perceptions: the role of some marine invasive species as trophic subsidies. Mar. Pollut. Bull. 116, 538–539. doi: 10.1016/j.marpolbul.2017.01.020
Rees, E., Sheehan, E. V., Stewart, B. D., Clark, R., Appleby, T., Attrill, M. J., et al. (2020). Emerging themes to support ambitious UK marine biodiversity conservation. Mar. Policy 117:103864. doi: 10.1016/j.marpol
Rife, A. N., Erisman, B., Sanchez, A., and Aburto-Oropeza, O. (2013). When good intentions are not enough…Insights on networks of “paper park” marine protected areas. Conserv. Lett. 6, 200–212. doi: 10.1111/j.1755-263X.2012.00303.x
Schlitzer, R. (2018). Ocean Data View. Available online at: https://odv.awi.de. (accessed October 2021).
Shannon, C. E., and Weaver, W. W. (1963). The Mathematical Theory of Communications. (Urbana: University of Illinois Press).
Sigovini, M., Keppel, E., and Tagliapietra, D. (2013). M-AMBI revisited: looking inside a widely-used benthic index. Hydrobiologia 717, 41–50. doi: 10.1007/s10750-013-1565-y
Sokolowski, A., Jankowska, E., Balazy, P., and Jędruch, A. (2021). Distribution and extent of benthic habitats in Puck Bay (Gulf of Gdańsk, Southern Baltic Sea). Oceanologia 63, 301–320. doi: 10.1016/j.oceano.2021.03.001
Sullivan, J. M., Cerny-Chipman, E. B., Rosenberg, A. R., and Lubchenco, J. (2017). “Bridging the science-policy interface: adaptive solutions in the anthropocene,” in Conservation for the Anthropocene Ocean: Interdisciplinary Science in Support of Nature and People, eds P. S. Levin and M. R. Poe (Amsterdam: Elsevier). doi: 10.1016/B978-0-12-805375-1.00001-5
Surugiu, V., Teacă, A., Şvedu, I., and Quijón, P. A. (2021). A Hotspot in the Romanian Black Sea: eelgrass beds drive local biodiversity in surrounding bare sediments. Front. Mar. Sci. 8:745137. doi: 10.3389/fmars.2021.745137
Teacă, A., Begun, T., Gomoiu, M.-T., and Paraschiv, G. M. (2006a). The present state of the epibionthic populations to the biocoenoses of stone mussels in the shallow water of the Romanian Black Sea coast. Geoecomarina 12, 53–66.
Teacă, A., Begun, T., and Gomoiu, M.-T. (2006b). Recent data on benthic populations from hard bottom mussel community in the Romanian Black Sea coastal zone. Geoecomarina 12, 43–51.
Teacă, A., Begun, T., and Mureşan, M. (2021). Annelid invaders in the Black Sea region: the distribution of Streblospio gynobranchiata and first occurrence of Laonome xeprovala. Glob. Ecol. Conserv. 32:e01920. doi: 10.1016/j.gecco.2021.e01920
Teacă, A., Begun, T., Surugiu, V., and Gomoiu, M.-T. (2010). Changes in the structure of the rocky mussels littoral biocoenosis from the Romanian Black Sea Coast. Ann. St. Univ. Al. I. Cuza Biol. Animala 56, 7–22.
Teacă, A., Mureşan, M., Begun, T., Popa, A., and Ion, G. (2019). Marine benthic habitats within a physical disturbed site from the Romanian Coast of the Black Sea. J. Environ. Prot. Ecol. 20, 723–732.
Teacă, A., Mureşan, M., Menabit, S., Bucse, A., and Begun, T. (2020). Assessment of Romanian circalittoral soft bottom benthic habitats under Danube river influence. Reg. Stud. Mar. Sci. 40:101523. doi: 10.1016/j.rsma.2020.101523
Teixeira, H., Berg, T., Uusitalo, L., Fürhaupter, K., Heiskanen, A. S., Mazik, K., et al. (2016). A catalogue of marine biodiversity indicators. Front. Mar. Sci. 3:207. doi: 10.3389/fmars.2016.00207
Todorova, V., Abaza, V., Dumitrache, C., Todorov, E., Wolfram, G., and Salas Herrero, F. (2018). “Coastal and transitional waters Black Sea Geographic Intercalibration Group,” in Benthic Invertebrate Fauna Ecological Assessment Methods, (Luxembourg: Publication Office of the European Union). doi: 10.2760/31396
Todorova, V., Dimitrov, L., Doncheva, V., Trifonova, E., and Prodanov, B. (2015). Benthic habitat mapping in the Bulgarian Black Sea, in Proceedings of the Twelfth International Conference on the Mediterranean Coastal Environment MEDCOAST ’15, ed. E. Ozhan, (Dalyan: Mediterranean Coastal Foundation) 1, 251–262.
Todorova, V., Doncheva, V., and Trifonova, E. (2020). First implementation of marine strategy framework directive for benthic habitats assessment in the Bulgarian Black Sea. Ecol. Balkanica 3, 247–256.
Todorova, V., and Konsulova, T. (2005). Manual for Quantitative Sampling And Sample Treatment Of Marine Soft-Bottom Macrozoobenthos. (Sofia: IO-BAS) 38.
Ungureanu, G. V., Lazăr, A. V., Lazăr, R., Balahură, A. D., Ionescu, A. D., Micu, D., et al. (2015). Habitat mapping of Romanian Natura 2000 sites. A case study, “Underwater Sulfurous Seeps, Mangalia”. Ital. J. Geosci. 134, 69–73. doi: 10.3301/IJG.2014.57
Vasquez, M., Manca, E., Inghilesi, R., Martin, S., Agnesi, S., Al Hamdani, Z., et al. (2020). EUSeaMap 2019, A European Broad-Scale Seabed Habitat Map, Technical Report. (Brest: Ifremer). doi: 10.13155/74782
Zaharia, T., Micu, D., Nita, V., Maximov, V., Mateescu, R., Spinu, A., et al. (2012). Preliminary data on habitat mapping in the Romanian Natura 2000 marine sites. J. Environ. Prot. Ecol. 13, 1776–1782.
Keywords: biodiversity hotspot, benthic habitats, marine protected area, ecological status, Black Sea
Citation: Begun T, Teacă A, Mureşan M, Quijón PA, Menabit S and Surugiu V (2022) Habitat and Macrozoobenthic Diversity in Marine Protected Areas of the Southern Romanian Black Sea Coast. Front. Mar. Sci. 9:845507. doi: 10.3389/fmars.2022.845507
Received: 29 December 2021; Accepted: 31 January 2022;
Published: 04 March 2022.
Edited by:
Eva Chatzinikolaou, Institute of Marine Biology, GreeceReviewed by:
Panayota Koulouri, Institute of Marine Biology, GreeceGiorgos Chatzigeorgiou, Institute of Marine Biology, Greece
Copyright © 2022 Begun, Teacă, Mureşan, Quijón, Menabit and Surugiu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Adrian Teacă, YXRlYWNhQGdlb2Vjb21hci5ybw==
†These authors have contributed equally to this work and share senior authorship