 ChenYan Wu
ChenYan Wu XiaoGu Wang1*
XiaoGu Wang1*
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Mar. Sci. , 19 April 2022
Sec. Marine Fisheries, Aquaculture and Living Resources
Volume 9 - 2022 | https://doi.org/10.3389/fmars.2022.832817
The objective of this study was to investigate influence of meso-scale eddies in the western South China Sea on species composition and spatial patterns of fish larvae and juveniles based on the data collected during summer and autumn in 2012. Nintyeight species belonging to 38 families were collected, including 78 species in summer and 41 species in autumn. The dominant species in summer were Vinciguerria sp. (39.70%), Stephanolepis cirrhifer (11.21%), Caranx sp. (5.45%) and Benthosema pterotum (1.97%). The dominant species in autumn were Gobiidae sp. (15.32%), Ceratoscopelus townsendi (7.26%), Diaphus sp. (6.45%), Astronesthes sp. (5.65%) and Callionymidae sp. (4.03%). Abundance of fish larvae and juveniles in summer was higher than that found in autumn. Similarity cluster analysis results indicated that larval fish assemblages in summer can be divided into two communities: (1) larval fish in the cyclonic eddy; (2) larval fish in the anticyclonic eddies. While in autumn, the larval fish assemblages can be divided into three communities: (1) larval fish without influence of the cyclonic eddy; (2) larval fish influenced slightly by the cyclonic eddy; (3) larval fish gathered near the center of the cyclonic eddy. The variations of abundance and assemblage of larval fish might be attributed to sea surface temperature (SST), sea surface salinity (SSS) and sea level anomaly (SLA).
The South China Sea is a semi-enclosed sea located in the southern part of mainland China. Its topography is complex and affected by strong monsoon; therefore, the mesoscale vortex phenomenon can occur (Lan et al., 2006). The southwest monsoon first appears in May, then extends to the entire South China Sea in July and August, and continues until mid-September (Shaw and Chao, 1994). During summer the southwest monsoon prevailing, an upwelling current was found between 10° and 15°N in the western South China Sea (Xu et al., 1980). There is a cyclonic eddy near this upwelling (111°E, 13°N), which called the Vietnamese Cold Eddy (Yang and Liu, 1998), The occurrence and elimination of this cold eddy are closely related to the upwelling along the coast of Vietnam. Specialists also observed a northeast jet near 12°N, 110°~113°E (Levin et al., 2014), which can bring the cold coastal upwelling to the basin (Xie, 2003). In addition, there is an anticyclonic eddy in the southern part of the cold eddy. This anticyclonic eddy is called a dipole together with the structure of the offshore eastward jet and the cyclonic eddy on the north side. It generally appears in June and disappears in October (Wang et al., 2006).
The larvae and juvenile stage is a period of high mortality with significant changes in morphology, ecology and physiology during the life history of fish. In the marine ecosystem, survival rate of larval fish directly affects their early recruitment (Wan and Zhang, 2016). Physical phenomena such as upwelling affects primary productivity, which in turn will affect the survival and growth of larval fish (Deng et al., 1995). An interesting result was found that the upwelling areas along the east coast of Guangdong and of Taiwan shoal had high productivity and abundant fish bait, which can form good nursery ground (Wang et al., 2015). In addition, the distribution pattern of larval fish communities is closely related to the dynamic characteristics of current. High abundance of larval fish is usually limited to the upwelling area, which is in accordance with the abundance of phytoplankton and zooplankton (Hsieh et al., 2010).
In a famous study analyzing the composition and distribution of larvae and juveniles in two deep mesoscale eddies in the eastern Indian Ocean (Muhling et al., 2007), they concluded that compared with the cold-core eddy and the warm-core eddy, the composition and distribution of fish larvae and juveniles had a greater difference because of the physical structure. In addition, in the Santa Barbara Strait, the density of fish larvae and juveniles in the center of the eddy was higher than that of the surrounding environment, but the physical conditions changed in the second year and the density decreased (Nishimoto and Washburn, 2002). These changes were related to the changes in the current system and its impact on the life cycle of species. Therefore, the effects of eddies on the ecosystems may not be easy to predict because they depend on the evolution of physical structure and biological communities over time.
There has been many research on physical phenomena in the western South China Sea, but only a few biological surveys and studies in this region. For example, the correlation between phytoplankton biomass and the environment was investigated (Tang et al., 2004). The present survey is aimed to clarify the relationship between the distribution of fish larvae and juveniles and environmental factors in the western South China Sea. We are also committed to provide scientific basis for the protection and recruitment of fish larval and juveniles resources in this area.
A survey on 15 sampling stations in the western South China Sea was carried out onboard of the R/V Xiangyanghong 14 during summer and autumn of 2012 (Figure 1). Samples were taken by a WP2 plankton net (mouth diameter, 57 cm; mesh size, 200 μm). Plankton hauls were taken vertically from 200m depth to the sea surface at a speed of 1ms-1. A flowmeter (Hydro-Bios) was attached to the net mouth. The obtained samples were immediately fixed and preserved in 5% (v/v) buffered formalin-seawater solution. A stereomicroscope (Axio Zoom.V16) was used in the laboratory for species identification and individual counting, and samples were identified to the lowest taxonomic level as much as possible.
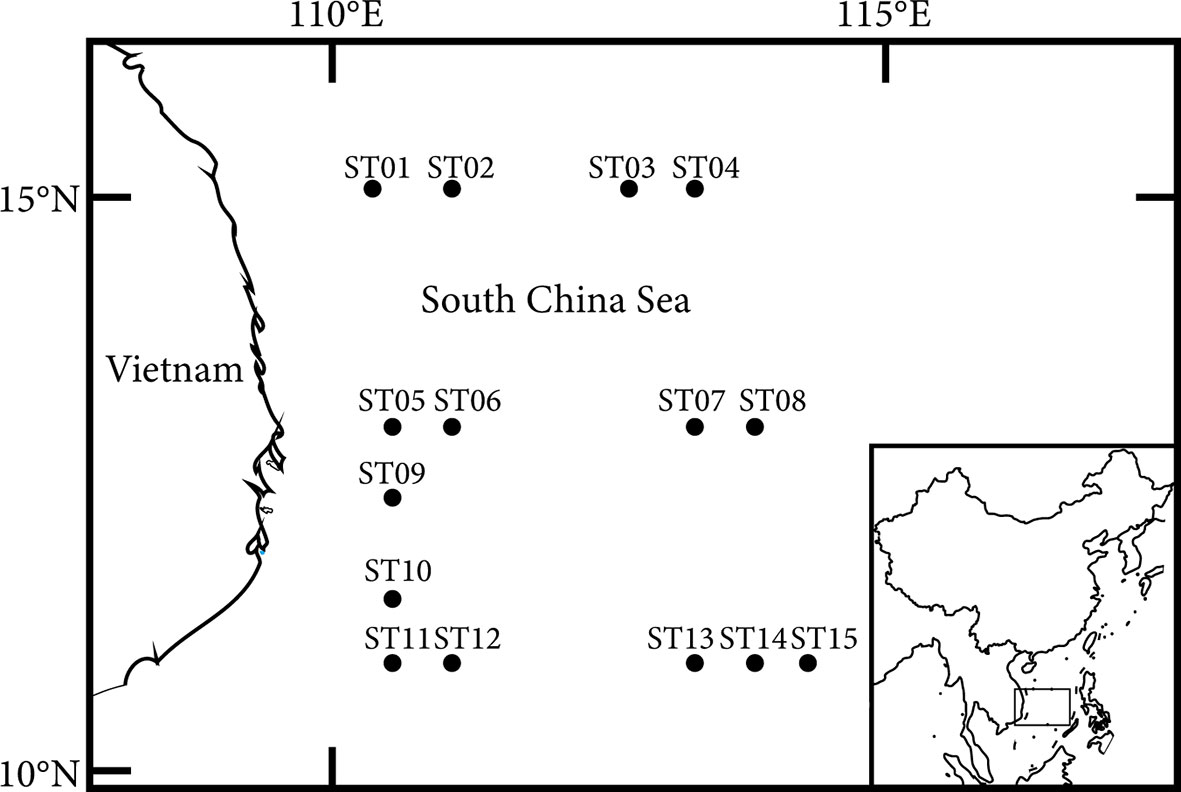
Figure 1 Sampling stations in the western South China Sea.
The sea surface temperature (SST) and chlorophyll a concentration (CHL) data with 4km resolution were downloaded from the Ocean Color Remote Sensing website of National Aeronautics and Space Administration (NASA) (https://oceancolor.gsfc.nasa.gov/, accessed by 28 Oct, 2021). The sea surface salinity (SSS) data were downloaded from the data of CATDS (Centre Aval de Traitement des Données SMOS) (http://www.salinityremotesensing.ifremer.fr/, accessed by 28 Oct, 2021). The sea level anomaly (SLA) data were downloaded from TOPEX/POSEIDON JASON satellite altimeter (http://www.aviso.oceanobs.com, accessed by 28 Oct, 2021). Ocean Data View (vers 4.0) was used for data reading, writing and editing. The specific environmental value which canonical correspondence analysis (CCA) were transformed by MATLAB.
Larval and juvenile fish abundance was standardized to numbers of individuals (ind.) per 100m3 (Li et al., 2014).
The Index of Relative Importance (IRI) (Zhu et al., 2002) was used to determine the dominant species:
N% is the relative abundance of occurrence and F% is the relative frequency of occurrence. The IRI of the dominant species should be greater than 100.
Margalef richness index (D) (Ulanowicz, 2001), Shannon-Wienerd diversity index (H’) (Wilhm, 1968) and Pielou’s index of evenness (J’) (Pielou, 1975) were used to analyse the larval community diversity.
S is the number of larval fish, N is the number of species, Pi is the proportion of individuals in taxa i.
PRIMER (vers 6.0) software package was used to perform cluster analysis and multi-dimensional scaling (MDS) in order to classify groups with distinct community structures (Bray and Curtis, 1957). The basic data used in PRIMER is fish abundance. The stress of MDS is used to test the reliability of analysis results. When 0.1<stress<0.2, it means that the result has a certain explanatory significance (Quetglas et al., 2013). The data transformed by log (x +1) before analysis (Clarke and Warwick, 2001). Canonical correspondence analysis (CCA) was performed to investigate the relationship between oceanographic conditions and larval fish compositions (Ter Braak, 1986). CANOCO (vers 6.0) software package was used to get results via the data of the fish abundance and the environmental variables. To eliminate inappropriate effect of rare species, only species with more than 1% of the total catch were chosen for the analysis.
The temperature in the northern part of the study area was relatively high in summer; however, a wide range of area with low temperature (<28°C) appeared as a large jet shape along the coastal waters of Vietnam (Figure 2A). While in autumn, the temperature in the northern part decreased significantly, and the offshore low-temperature zone disappeared (Figure 2B).
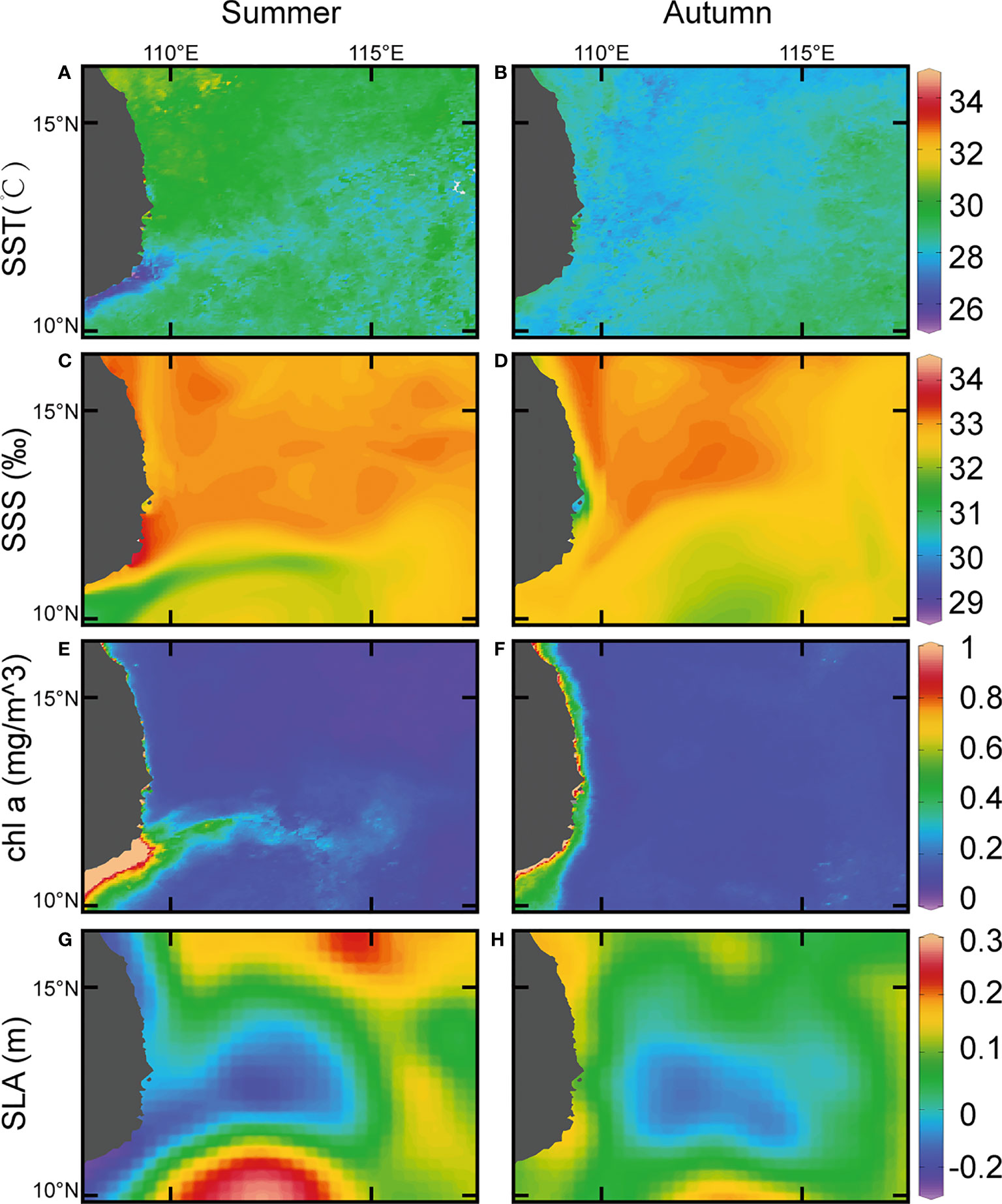
Figure 2 Sea surface temperature, salinity, chlorophyll a concentration and sea level anomaly in the western South China Sea.(A, C, E, G) in summer; (B, D, F, H) in autumn.
The salinity which influenced by the fresh water of the Mekong River gradually increased from the mouth of the Mekong River to the sea during summer and the southern part of the study area was covered by waters with low salinity (<33‰) for a long time (Figure 2C). While in autumn, the jet-shaped protrusion disappeared, and the area affected by the river’s fresh water reduced (Figure 2D).
According to the concentration of chlorophyll a from MODIS data, a large amount of phytoplankton bloomed near the Vietnam coast (10°N-12°N) during summer (Figure 2E), while in autumn, this eastward algal bloom disappeared (Figure 2F).
Sea level anomaly data disclosed that a strong cyclonic eddy happened near the area of upwelling in summer, and the anticyclonic eddies were also appeared close to the cyclonic eddy (Figure 2G). However, the intensity of the cyclonic and anticyclonic eddies weakened relatively in autumn (Figure 2H).
A total of 784 fish larvae and juveniles were collected in this survey, belonging to 98 species in 38 families (Table 1). Among them, 660 individuals belonging to 78 species were collected in summer. Stomiiformes accounted for the largest proportion, reaching 43.33% and including 11 species among 4 families; followed by Perciformes (23.48%), including 30 species among 14 families. Tetraodoniformes accounted for 12.42%, including 3 species among 2 families. As shown in Table 2, the dominant species were Vinciguerria sp. (39.70%), Stephanolepis cirrhifer (11.21%), Caranx sp. (5.45). %) and Benthosema pterotum (1.97%).
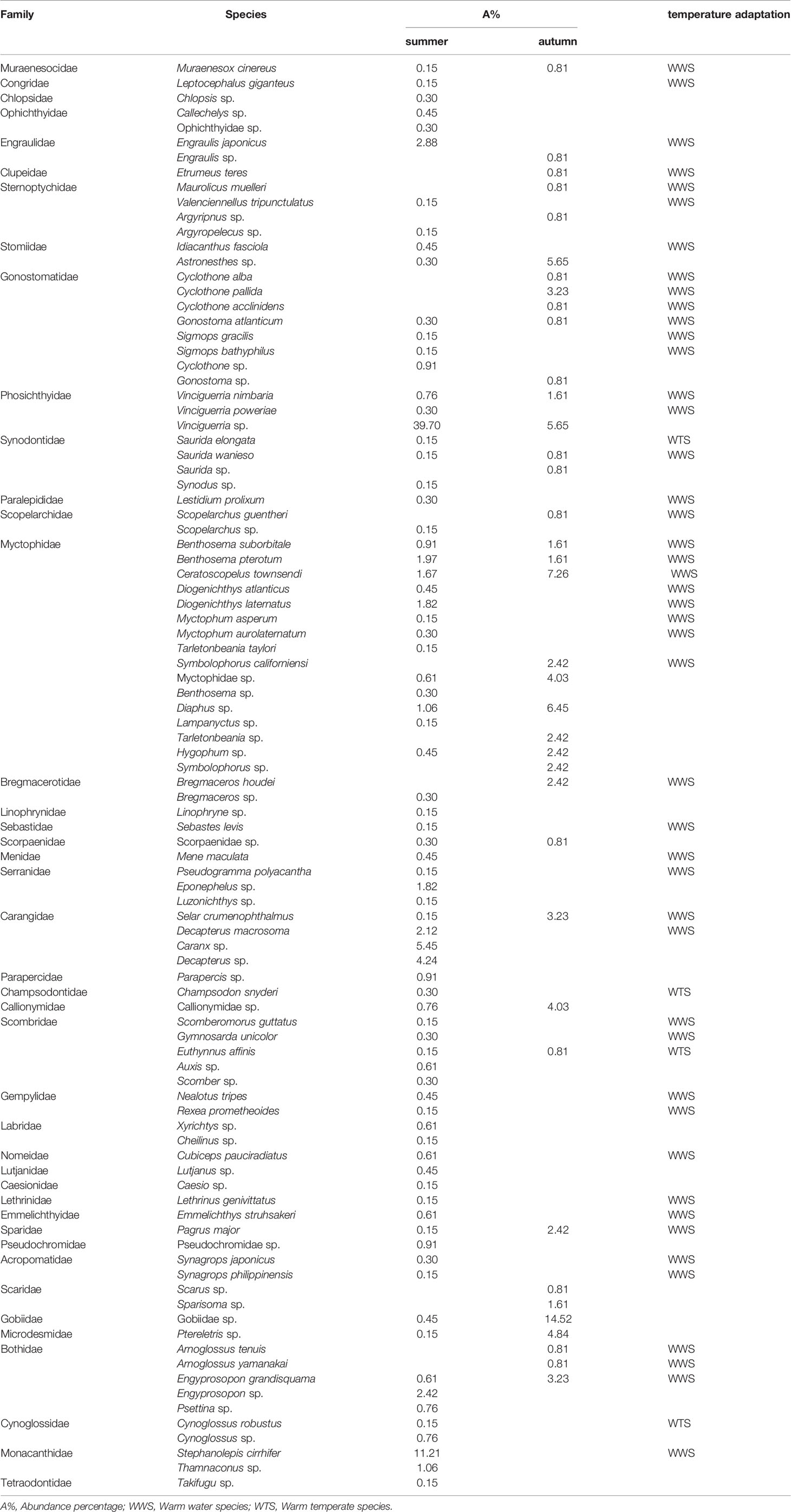
Table 1 Species composition of fish larvae and juveniles in the western South China Sea.
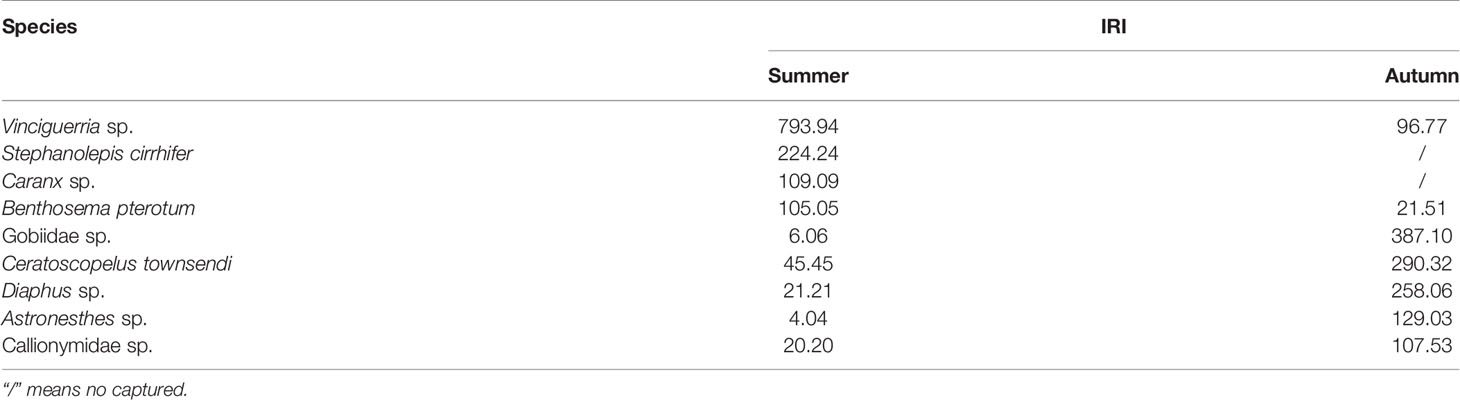
Table 2 Index of relative importance of the dominant species of larval fish in the western South China Sea.
One hundred and twenty-four individuals belonging to 41 species were collected in autumn. Perciformes accounted for the largest proportion, reaching 34.68% and including 9 species among 8 families. Myctophiformes was the second abundant group, accounting for 30.65% and including 8 species among 1 family. Stomiiformes accounted for 20.97%, including 10 species among 4 families. As shown in Table 2, the dominant species were Gobiidae sp. (15.32%), Ceratoscopelus townsendi (7.26%), Diaphus sp. (6.45%), Astronesthes sp. (5.65%) and Callionymidae sp. (4.03%).
The range of larval and juvenile fish abundance was from 3.2 to 57.1 ind/1003 in summer (Figure 3A), from 2.0 to 23.1 ind/1003 in autumn (Figure 3B). Average abundances of fish larvae and juveniles during summer and autumn are presented in Table 3. The abundance generally showed that the result in summer was higher than in autumn and differed significantly between these two seasons.
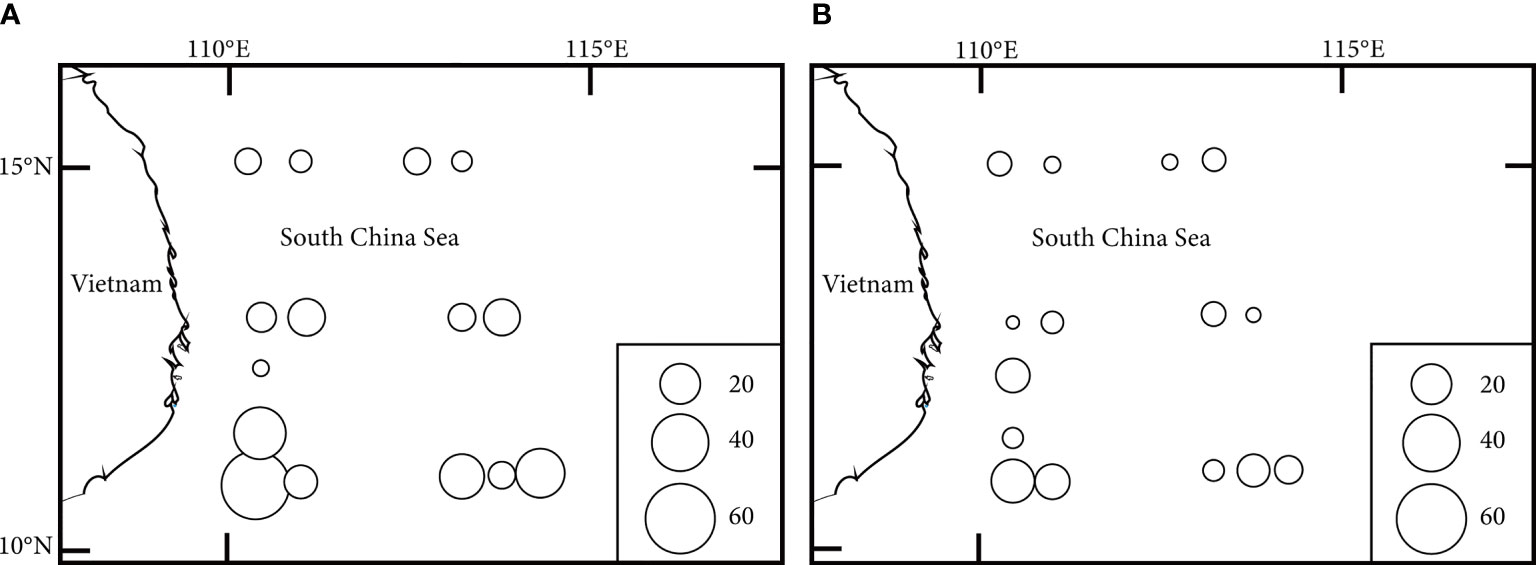
Figure 3 | The distribution of fish larvae and juveniles in the western South China Sea (circles:ind/100-3). (A) summer; (B) autumn.
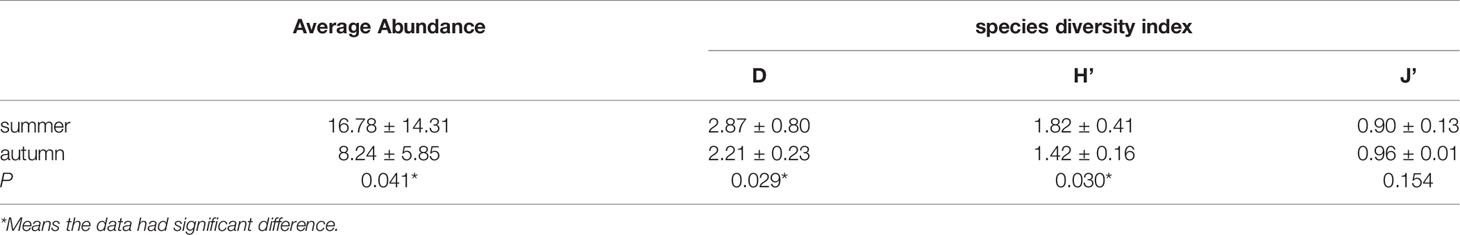
Table 3 Average abundance and diversity index of fish larvae and juveniles in the western South China Sea during summer and autumn and p values from ANOVA analysis (Significance level =0.05).
From Table 3, we can clearly see that the Margalef Richness Index (D) and the Shannon-Wiener diversity index (H’) revealed the same seasonal variation which the value was higher in summer than in autumn, particularly showed a significant difference respectively (p<0.05). However, for the Pielou’s index of evenness (J’), there was no significant difference with season(p>0.05).
The results of the cluster analysis in summer indicated (Figure 4A) that all stations can be divided into two groups at 4.38% similarity level. The assemblage of ST04, ST11, ST12, ST13 composed an anticyclonic-eddies-assemblage (AE) which came from a region of the anticyclonic eddies (combining with the above sea level anomaly image (Figure 2G), while the remaining 11 stations composed a cyclonic-eddy-assemblage (CE) which came from a region of the cyclonic eddy (combining with the above sea level anomaly image (Figure 2G). The stress (0.14<0.2) showed the reliability of the analysis result (Figure 4A). The dominant species in the AE were Vinciguerria sp., Stephanolepis cirrhifer and Caranx sp. Meanwhile, the CE was defined by Benthosema pterotum, Ceratoscopelus townsendi, Engraulis japonicus and Diogenichthys laternatus.
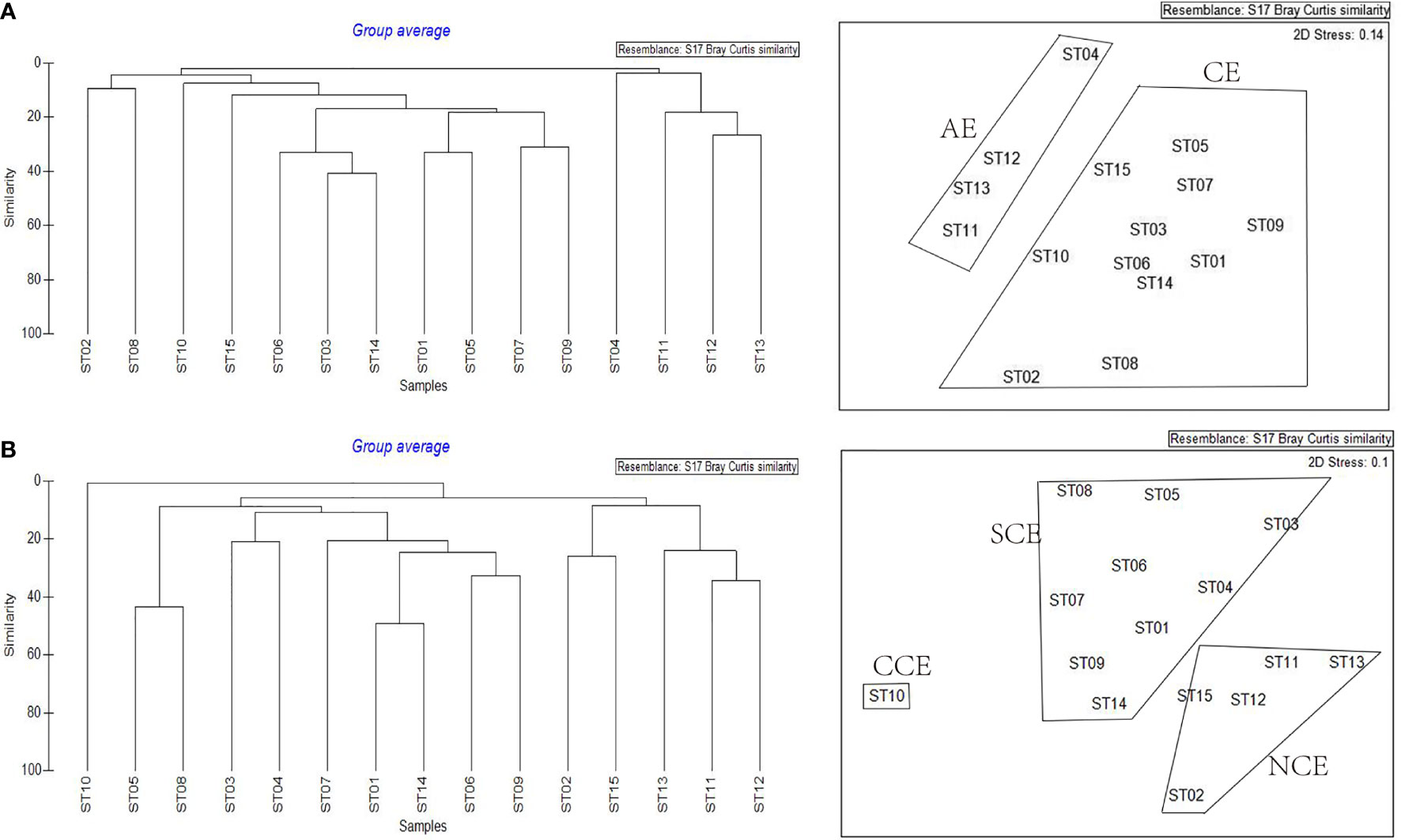
Figure 4 Cluster analysis based on the Bray-Curtis similarity of abundance at each sampling station of the western South China Sea. (A) summer; CE: larval fish in the cyclonic eddy; AE: larval fish in the anticyclonic eddies; (B) autumn; NCE: larval fish without influence of the cyclonic eddy; SCE: larval fish influenced slightly by the cyclonic eddy; CCE: larval fish gathered near the center of the cyclonic eddy.
The cluster analysis on autumn samples showed (Figure 4B) that all stations can be divided into three groups. The station of ST10 was distinguished from other groups with 0.66% similarity and grouped by itself (CCE) which near the center of the cyclonic eddy (combining with the above sea level anomaly image Figure 2H). The stations of ST02, ST11, ST12, ST13, and ST15 composed a group (NCE) which at 8.45% similarity, which were not influenced by the cyclonic eddy (combining with the above sea level anomaly image. Figure 2H). The remaining stations were classified into one group (SCE) at 8.88% similarity level, which was influenced slightly by the cyclonic eddy (combining with the above sea level anomaly image (Figure 2H). The stress (0.1<0.2) showed the reliability of the analysis result (Figure 4B). Maurolicus muelleri only occurred in the CCE group, while Diaphus sp. were found both in the NCE and SCE group. The NCE group consistently contained the highest densities of Vinciguerria sp., Astronesthes sp. and Gobiidae sp.The dominant species in the SCE group were Ceratoscopelus townsendi and Callionymidae sp.
The canonical correspondence analysis results in summer (Table 4) showed that SST、SSS and SLA had a significant effect on the fish assemblages (p<0.05). The eigenvalues were 0.667 (CCA1), 0.416 (CCA2), 0.367 (CCA3) and 0.096 (CCA4) as shown in Table 5. The correlation between axes and environmental variables were 0.924 (CCA1), 0.943 (CCA2), 0.864 (CCA3) and 0.801 (CCA4). The Monte Carlo test indicated that the CCA1 had a significant difference (F=2.5, P=0.016<0.05), while the CCA2 had no significant difference (F=1.9, P=0.096>0.05). These two ordination axes explained 32.59% variation of the composition of fish larvae and juveniles in summer. The area of the AE group was characterized with high temperature, salinity and sea level anomaly, while the environmental conditions of the CE group were on the contrary. Vinciguerria sp., Stephanolepis cirrhifer and Caranx sp. that prefer the mild environment gathered in the CE group. The environmental variables were positively correlated with assemblages (Figure 5A). On the other hand, Benthosema pterotum, Ceratoscopelus townsendi and Diogenichthys laternatus belonging to the deep-sea fish dominated the CE group. The environmental variables were negatively correlated with their spatial patterns (Figure 5A).

Table 4 Simple term effects of environmental variables (Significance level =0.05).
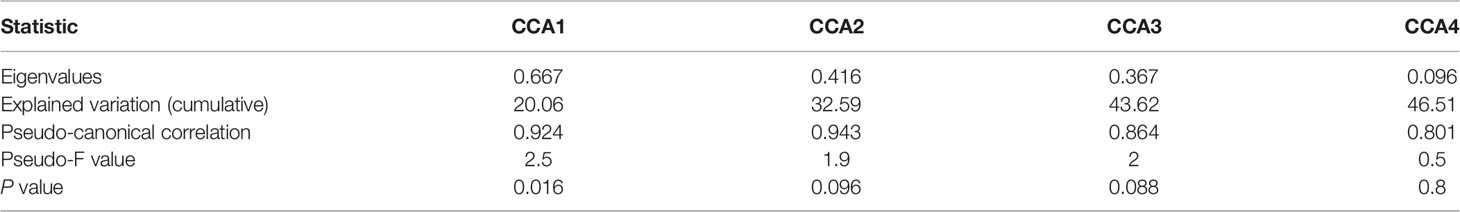
Table 5 CCA results in summer.
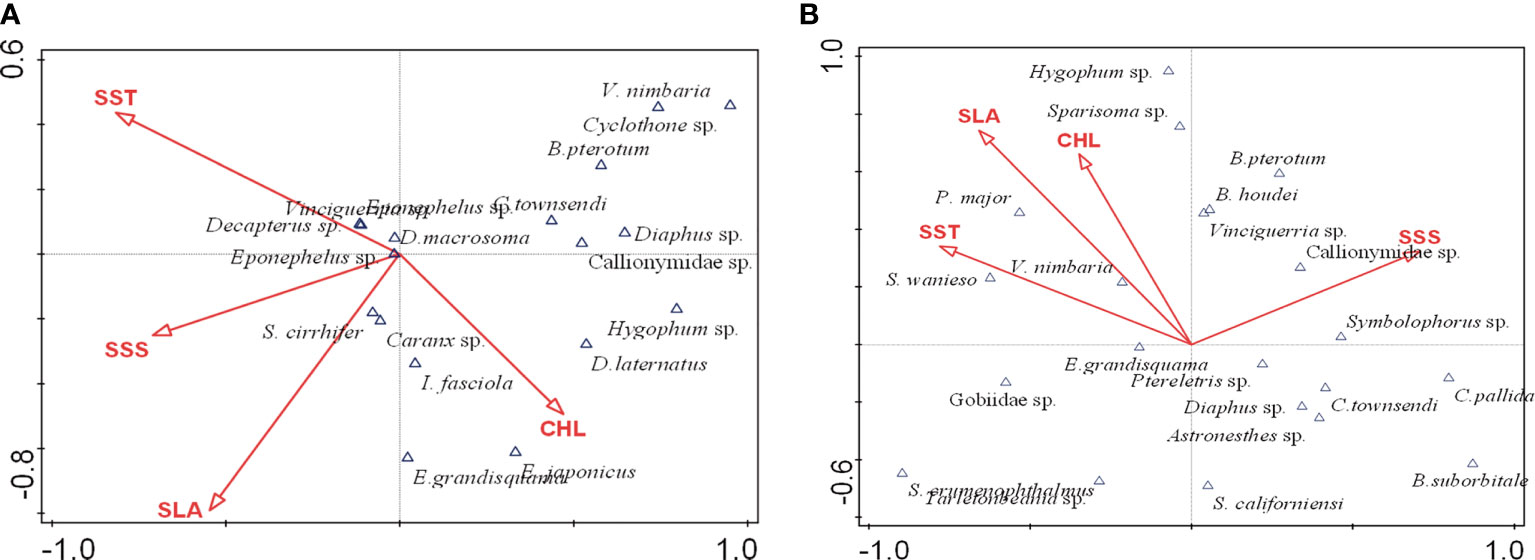
Figure 5 CCA biplot of species composition of survey stations and environmental variables in the western South China Sea. (A) summer; (B) autumn.
The canonical correspondence analysis results in autumn (Table 4) showed that SST and SLA had a significant effect on the larval and juvenile fish assemblages (p<0.05). The eigenvalues were 0.550 (CCA1), 0.312 (CCA2), 0.194 (CCA3) and 0.111 (CCA4) as shown in Table 6. The correlation between axes and environmental variables were 0.932 (CCA1), 0.915(CCA2), 0.713 (CCA3) and 0.784 (CCA4). The Monte Carlo test indicated that the CCA1 had a significant difference (F=1.7, P=0.04<0.05), while the CCA2 had no significant difference (F=1.1, P=0.718>0.05). These two ordination axes explained 24.62% variation of the composition of fish larvae and juveniles in autumn. Maurolicus muelleri was only captured in the area where the temperature and sea level anomaly were relatively low. The SCE group was dominated by Ceratoscopelus townsendi. The environmental variables were negatively correlated with their distribution (Figure 5B). However, the NCE group in the area where the temperature and sea level anomaly were relatively high which was dominantly composed by Vinciguerria sp. and Gobiidae sp. The environmental variables were positively correlated with their spatial patterns (Figure 5B).
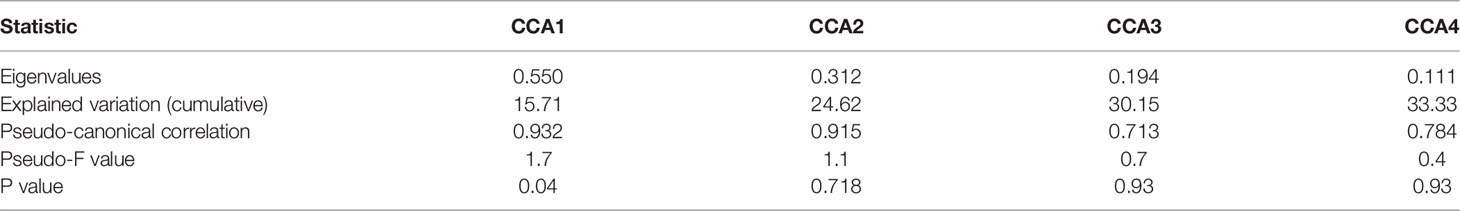
Table 6 CCA results in autumn.
There were some previous investigations of fish larvae and juveniles in the northern South China Sea. For example, in 2004-2007, 58 ichthyoplankton species belonging to 33 families were sampled, and the most significant environmental variables were water temperature, salinity and dissolved oxygen (Xiao et al., 2010). In 2006-2007, a total of 95 taxa were collected and the seasonal variation of diversity and abundance may be influenced by the South China Sea Current (Li et al., 2014). What’s more, the researchers (Zheng et al., 2016) pointed out that the coastal currents in the western South China Sea flowed to the northeast near 12°N in summer. Due to the large amount of offshore transportation of water, the alongshore water should be supplemented with sea water, so the upwelling with low-temperature and high-salinity seawater generated. The upwelling is usually accompanied by the cyclonic eddy (Yang, 2008). The present study was carried out in the western South China Sea, and the complicated hydrological conditions in this area had a significant impact on the composition and distribution of fish larvae and juveniles.
A total of 784 individuals were captured in our surveys, and the number in summer accounted for 84.18% of the total catch. The density and diversity of larval and juvenile fish in summer were higher than in autumn, which indicated that summer is the peak season for spawning in this area. The survey found that the dominant species in the CE group and SCE group were Benthosema pterotum (Myctophidae), Ceratoscopelus townsendi (Myctophidae), Engraulis japonicus (Engraulidae) and Diogenichthys laternatus (Myctophidae). The families which these species belong to were same to the results of the survey conducted in the Gulf of California where was influenced by the cyclonic eddy (Sanchez-Velasco et al., 2013). The species identified in the Gulf of California were Anchoa spp. (Engraulidae), Vinciguerria lucetia (Gonostomatidae) and Benthosema panamense (Myctophidae). Not only Sanchez’s research, but also our survey area in the eddy was dominated by the larval fish of Myctophidae and Gonostomatidae which prefer deeper layer when they grow into adult fish. How do larval and juvenile fish belong to the deep-sea fish appeared in the surface water layer? We think the upwelling accompanied by the cyclonic eddy may play an important role on the vertical transportation of larval and juvenile fish. However, this point should be verified with further study.
They Rodríguez et al. (2004) proposed that a reproductive strategy usually occurs in upwelling region. This strategy was able to cause low abundance and low diversity so that some species could prevent the offshore transport of their offspring. In addition, Diaphus sp. were found both in the NCE and SCE group. Therefore, we supposed that the larval fish may have been caught by the eddy and have been remained on the edge of the eddy, and then may be transported to the open sea. This hypothesis was almost consistent with the investigation in the Canaries CTZ during summer (Rodriguez et al., 2004). They discovered that mesoscale eddies can transport fish larvae and juveniles from shallow waters of Africa to the ocean. The stations affected by the fresh water of the Mekong River had higher abundance than other area. In Chile upwelling ecosystem, the offshore advection may partially explain the higher abundance (Landaeta et al., 2000).
This study described the impact of mesoscale eddies in the western South China Sea on fish larvae and juveniles. Contereras-Catala et al. (2012) pointed out that the abundance of fish larvae and juveniles in the center of the eddy was not as high as in the eastern upwelling region. Due to the eastern part of the Gulf of California could be affected by the upwelling. During the upwelling period, the productivity is higher and the zooplankton biomass is higher, so the abundance of fish larvae and juveniles that feed on plankton is higher.
We can see that the abundance of station 10 is higher than other stations obviously in summer. I suppose the jet had a significant influence on the abundance. The input of large amounts of nutrients and organic matter from flushing water can cause phytoplankton algal blooms to form higher primary productivity. Therefore, fish larvae and juveniles can grow comfortably. Sassa et al. (2002) pointed out that the mesopelagic fishes like Myctophidae, spend their larval stages in the upper 200m layer. Werner and Fumio (2019) found that Myctophid otoliths are the most common fossil otoliths to be found in Ncogene deep water sediments below approximately 200 m depositional depth. (Xu et al., 1980) pointed out an upwelling current was found between 10° and 15°N in the western South China Sea. The larvae and juveniles of mesopelagic fishes can be transported to the surface water easily.
This study was to explore the influence of meso-scale eddies in the western South China Sea on the species composition and distribution of fish larvae and juveniles during summer and autumn. The results indicated that the abundance and taxonomic diversity of fish larvae and juveniles in summer was higher than in autumn. The dominant species in and around the cyclonic eddy were the mixed coastal and oceanic species. We proposed that the larval fish may have been caught by the eddy and have been remained on the edge of the eddy, and then may be transported to the open sea. The variations of abundance and assemblage of larval fish might be attributed to sea surface temperature (SST), sea surface salinity (SSS) and sea level anomaly (SLA). Transportation of larval and juvenile fish of Myctophidae and Gonostomatidae from the deeper layer to the surface maybe was facilitated by the upwelling. This point should be verified with further study.
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding authors.
The animal study was reviewed and approved by Second Institute of Oceanography.
CW, XW, JZ, JJ and CL contributed to conception and design of the study. XW organized the database. JZ, JJ and CL performed the statistical analysis. CW wrote the first draft of the manuscript. All authors contributed to the article and approved the submitted version.
This work was supported by National Key Research & Development (R&D) Plan (No.2016YFC0304105).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We would like to thank Prof. Dong Sun for his valuable advices in preparing the manuscript. We also thank the editor and reviewers for their constructive comments on our manuscript. We are also grateful to the members of R/V Xiangyanghong 14 for their help during this investigation.
Bray J. R., Curtis J. T. (1957). An Ordination of the Upland Forest Communities of Southern Wisconsin. Ecol. Monogr. 27, 325–349. doi: 10.2307/1942268
Clarke K. R., Warwick R. M. (2001). Changes in Marine Communities: An Approach to Statistical Analysis and Interpretation. 2nd edition (Plymouth: Primer-e Ltd), 172.
Contereras-Catala F., Sanchez-Velasco L., Lavin M. F., Miguel F. (2012). Three-Dimensional Distribution of Larval Fish Assemblages in an Anticyclonic Eddy in a Semi-Enclosed Sea (Gulf of California). J. Plankton Res. 6, 548–562. doi: 10.1093/plankt/fbs024
Deng S., Zhong H., Wang M. (1995). On Relation Between Upwelling Off Qionghai and Fishery. J. Oceanogr. Taiwan Strait 14 (1), 51–56. (in Chinese with an English abstract).
Hsieh H. Y., Lo W. T., Liu D. C. (2010). Influence of Hydrographic Features on Larval Fish Distribution During the South-Westerly Monsoon in the Waters of Taiwan, Western North Pacific Ocean. J. Fish Biol. 76 (10), 2521–2539. doi: 10.1111/j.1095-8649.2010.02643.x
Landaeta M. F., Veas R., Letelier J., Castro L. R. (2000). Larval Fish Assemblages Off Central Chile Upwelling Ecosystem. Rev. Biol. Marina Y Oceanografia 43 (3), 569–584. doi: 10.4067/S0718-19572008000300016
Lan J., Hong J., Li P. (2006). Seasonal Variability of Cool-Core Eddy in the Western South China Sea. Adv. Earth Sci. 21 (11), 1145–1152. (in Chinese with an English abstract).
Levin J., Wilkin J. L., Castruccio F. S. (2014). Interannual Variability of the Surface Summertime Eastward Jet in the South China Sea. J. Geophys. Res. 119, 7205–7228. doi: 10.1002/2014JC010206
Li K., Yin J., Huang L. (2014). Seasonal Variations in Diversity and Abundance of Surface Ichthyoplankton in the Northern South China Sea. Acta Oceanologica Sin. 33 (12), 145–154. doi: 10.1007/s13131-014-0533-3
Muhling B. A., Beckley L. E., Olivar M. P. (2007). Icthyoplankton Assemblage Structure in Two Mesoscale Leeuwin Current Eddies, Eastern Indian Ocean. DeepSea Res. II 54, 1113–1128. doi: 10.1016/j.dsr2.2006.05.045
Nishimoto M. M., Washburn L. (2002). Patterns of Coastal Eddy Circulation and Abundance of Pelagic Juvenile Fifish in the Santa Barbara Channel, California, USA. Mar. Ecol. Prog. Ser. 241, 183–199. doi: 10.3354/meps241183
Quetglas A., Carbonell A., Snchez P. (2013). Demersal Continental Shelf and Upper Slope Eephalopod Assemblages From the Balearie Sea (North—Western Mediterranean). Biological Aspects of Some Deep-Sea Species. Estuarine, Coastal Shelf Sci. 50 (6), 739—749.
Rodriguez J. M., Barton E. D., Hernandez-Leon S. (2004). The Influence of Mesoscale Physical Processes on the Larval Fish Community in the Canaries CTZ, in Summer. Prog. Oceanogr. 62 (2/4), 171–188. doi: 10.1016/j.pocean.2004.07.006
Sanchez-Velasco L., Lavin M. F., Jimenez-Rosenberg S. P. A. (2013). Three-Dimensional Distribution of Fish Larvae in a Cyclonic Eddy in the Gulf of California During the Summer. Deep Sea Res. Part I Oceanogr. Res. Papers 75 (May), 39–51. doi: 10.1016/j.dsr.2013.01.009
Sassa C., Moser H. G., Kawaguchi K. (2002). Horizontal and Vertical Distribution Patterns of Larval Myctophid Fifishes in the Kuroshio Current Region. Fisheries Oceanogr. 11, 1–10. doi: 10.1046/j.1365-2419.2002.00182.x
Shaw P. T., Chao S. Y. (1994). Surface Circulation in the South China Sea. Deep-Sea Res. I Oceanogr. Res. Pap. 41 (11–12), 1663–1683. doi: 10.1016/0967-0637(94)90067-1
Tang D. L., Kawamura H., Dien T. V., Lee M. A. (2004). Offshore Phytoplankton Biomass Increase and Its Oceanographic Causes in the South China Sea. Marine Ecology-Progress Ser. 268 (Mar), 31–41. doi: 10.3354/meps268031
Ter Braak C. J. F. (1986). Canonical Correspondence Analysis: A New Eigenvector Technique for Multivariate Direct Gradient Analysis. Ecology 67, 1167–1179. doi: 10.2307/1938672
Ulanowicz R. E. (2001). Information Theory in Ecology. Computers&chemistry 25 (4), 393—399. doi: 10.1016/s0097-8485(01)00073-0
Wang G., Chen D., Su J. (2006). Generation and Life Cycle of the Dipole in the South China Sea Summer Circulation. J. Geophys. Res. Atmos. 111 (C6). doi: 10.1029/2005JC003314
Wang X., Yu J., Li Y. (2015). The Relationship Between Major Upwelling and the Upwelling Fishing Grounds in the South China Sea. Marine Sci. 39 (9), 131–137. doi: 10.11759/hykx20130627001
Wan R., Zhang R. (2016). Fish Eggs, Larvae and Juveniles in the Offshore Waters of China and Their Adjacent Waters (Shanghai: Shanghai Scientific & Technical Publishers).
Werner S. A., Fumio O. (2019). Lanternfish Otoliths (Teleostei,Myctophidae) From the Pliocene and Pleistocene of Japan. Rivista Italiana di Paleontol. e Stratigrafia 125 (2), 355–400. doi: 10.13130/2039-4942/11670
Wilhm J. L. (1968). Use of Biomass Units in Shannon’s Formula. Ecology 49 (1), 153–156. doi: 10.2307/1933573
Xiao Y., Wang R., Ou Q. (2010). Relationship Between Abundance Distribution of Eggs, Larvae and Juveniles and Environmental Factors in the Pearl River Estuary Waters in Spring. J. Oceanogr. Taiwan Strait 29 (4), 488–495. doi: 10.3969/J.ISSN.1000-8160.2010.04.008
Xie S. (2003). Summer Upwelling in the South China Sea and its Role in Regional Climate Variations. J. Geophys. Res. Oceans 108 (C8). doi: 10.1029/2003JC001867
Xu X., Zhang Q., Chen H. (1980). “Horizontal Circulation in South China Sea,” in Scientific Symposium of China Oceanologia et Limnologia (Beijing: SciencePress), 137–145. (in Chinese with an English abstract).
Yang S. (2008). Observations and Research on Vietnam Cold Eddies and Eddy-Induced Upwelling Current. Research (in Chinese with an English abstract).
Yang H., Liu Q. (1998). A Summary on Ocean Circulation Study of the South China Sea. Adv. Earth Sci. 13 (4), 364–368. (in Chinese with an English abstract).
Zheng S., Bao X., Yang D. (2016). The Hydrological Characteristics of Mesoscale Eddies in the South-Western Basin of the South China Sea. Trans. Oceanol. Limnol. 4, 1–10. (in Chinese with an English abstract).
Keywords: western South China Sea, fish larvae and juveniles, species composition, assemblages, meso-scale eddies
Citation: Wu CY, Wang XG, Zhong JS, Ju JL and Li CH (2022) Spatial Patterns of Larval and Juvenile Fish Assemblages in an Eddy Area in the Western South China Sea. Front. Mar. Sci. 9:832817. doi: 10.3389/fmars.2022.832817
Received: 10 December 2021; Accepted: 16 March 2022;
Published: 19 April 2022.
Edited by:
Çetin Keskin, Istanbul University, TurkeyReviewed by:
Ana Primo, University of Coimbra, PortugalCopyright © 2022 Wu, Wang, Zhong, Ju and Li. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: XiaoGu Wang, d2FuZ3hpYW9ndTE5NjhAMTI2LmNvbQ==; JunSheng Zhong, anN6aG9uZ0BzaG91LmVkdS5jbg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.