
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Mar. Sci., 16 March 2022
Sec. Marine Megafauna
Volume 9 - 2022 | https://doi.org/10.3389/fmars.2022.815684
 Justine Girardet1
Justine Girardet1 Francois Sarano2
Francois Sarano2 Gaëtan Richard3
Gaëtan Richard3 Paul Tixier4
Paul Tixier4 Christophe Guinet5
Christophe Guinet5 Alana Alexander6
Alana Alexander6 Véronique Sarano2
Véronique Sarano2 Hugues Vitry7Axel Preud’homme7
Hugues Vitry7Axel Preud’homme7 René Heuzey8
René Heuzey8 Ana M. Garcia-Cegarra9,10Olivier Adam11,12
Ana M. Garcia-Cegarra9,10Olivier Adam11,12 Bénédicte Madon13
Bénédicte Madon13 Jean-Luc Jung1*
Jean-Luc Jung1*
Adult male sperm whales (Physeter macrocephalus) are long distance runners of the marine realm, feeding in high latitudes and mating in tropical and subtropical waters where stable social groups of females and immatures live. Several areas of uncertainty still limit our understanding of their social and breeding behavior, in particular concerning the potential existence of geographical and/or social fidelities. In this study, using underwater observation and sloughed-skin sampling, we looked for male social fidelity to a specific matrilineal sperm whale group near Mauritius. In addition, we captured a wider picture of kin relationships and genetic diversity of male sperm whales in the Indian Ocean thanks to biopsies of eight individuals taken in a feeding ground near the Kerguelen and Crozet Archipelagos (Southern Indian Ocean). Twenty-six adult male sperm whales were identified when socializing with adult females and immatures off Mauritius. Sloughed-skin samples were taken from thirteen of them for genetic analysis. Long-term underwater observation recorded several noteworthy social interactions between adult males and adult females and/or immatures. We identified seven possible male recaptures over different years (three by direct observation, and four at the gametic level), which supports a certain level of male social fidelity. Two probable first- and thirty second-degree kin relationships were highlighted between members of the social unit and adult males, confirming that some of the adult males observed in Mauritian waters are reproductive. Male social philopatry to their natal group can be excluded, as none of the males sampled shared the haplotype characteristic of the matrilineal social group. Mitochondrial DNA control region haplotype and nucleotide diversities calculated over the 21 total male sperm whales sampled were similar to values found by others in the Indian Ocean. Our study strongly supports the existence of some levels of male sperm whale social fidelity, not directed to their social group of birth, in the Indian Ocean. Males sampled in breeding and feeding grounds are linked by kin relationships. Our results support a model of male mediated gene flow occurring at the level of the whole Indian Ocean, likely interconnected with large-scale geographical fidelity to ocean basin, and a small-scale social fidelity to matrilineal social groups.
Sexual dimorphism, defined as differences in external appearance or other characteristics between the two sexes of a species (Mesnick and Ralls, 2018), is widespread among animals, and especially in vertebrates (Shine, 1989). Sexual dimorphism can be behavioral and/or morphological concerning life history. Marked sexual dimorphism is present in several marine mammal species (Mesnick and Ralls, 2018). Morphological differences are obvious, for example, in elephant seals (Mirounga angustirostris and M. leonina), males being up to ten times larger than females (Le Boeuf and Laws, 1994) and in narwhals (Monodon monoceros) where males possess a tusk (Gerson and Hickie, 1985). Other species display preferential long-term association between members of the same sex, such as the Indo-Pacific bottlenose dolphins (Tursiops aduncus; Smolker et al., 1992; Galezo et al., 2017), or exhibit differences in their feeding ecology between males and females such as the resident fish-eating ecotype of killer whales of the northeastern Pacific Ocean (Orcinus orca; Beerman et al., 2016).
Sperm whales certainly display some of the most striking sexual dimorphism among cetaceans, both in terms of body size with adult males growing up to 18 m long and a weight of 45 t, while females usually remain around 11 m long for 13 t (Best, 1979; Cantor et al., 2019); but also in terms of feeding ecology, geographical distribution and social organization (Rice, 1989; Whitehead and Kahn, 1992; Teloni et al., 2008; Kobayashi et al., 2020). Male and female sperm whales live in societies that are strongly geographically segregated post-maturity (e.g., Christal, 1998; Gordon et al., 1998; Christal and Whitehead, 1999; Lyrholm et al., 1999; Whitehead et al., 2008; Labadie et al., 2018). Adult females form social units with immatures, stable over time and found all year round in warm waters at low latitudes (Whitehead and Kahn, 1992; Konrad et al., 2018; Sarano et al., 2021). In contrast, males disperse from their natal group after 6–8 years, before their sexual maturity, and move poleward to areas abundant in food (Rice, 1989). After their twenties, they make periodic forays to warmer waters for mating, with no known clear frequency, seasonal agendas nor migration routes (Best, 1979). Although we know that adult male sperm whales can travel thousands of kilometers across ocean basins (Lyrholm et al., 1999; Engelhaupt et al., 2009; Mizroch and Rice, 2012; Steiner et al., 2012), no recurrent migration routes between feeding and breeding areas have so far been identified (Cantor et al., 2019).
In cold waters, non-breeding adult males can be encountered alone or in small groups called “bachelor groups,” groups of tens of individuals of about the same age (e.g., Christal and Whitehead, 1997; Jaquet et al., 2000; Lettevall et al., 2002). They may become more and more solitary as they age (Best, 1979). In northern Norway, Nova Scotia (Canada) and Kaikōura (New Zealand) feeding grounds, no noticeable social interaction between adult males were observed when foraging (Lettevall et al., 2002; Madsen et al., 2002). Yet, some recent studies show that males can form long-term associations (Kobayashi et al., 2020) and have fluid and unstructured social interactions that allow the social transmission of depredation techniques in the Gulf of Alaska (Schakner et al., 2014) or permit coordinated anti-predator responses (Curé et al., 2013). Long-term photo-identification studies around Crozet and Kerguelen archipelagos (Crozet/Kerguelen, Southern Indian Ocean), in the Bleik Canyon (northern Norway) and in the Nemuro Strait (northern Japan) indicate that adult males exhibit site fidelity at local scales (Rødland and Bjørge, 2015; Labadie et al., 2018; Kobayashi and Amano, 2020).
In the low latitudes, the social interactions of adult male sperm whales with stable social groups of females and immatures and adult male movement patterns in breeding grounds remain poorly known. Adult males may temporarily join social units to breed and stay in the same area for periods estimated from a few hours to a few days off the Galapagos Islands (Coakes and Whitehead, 2004) to a few weeks in the West Indies (Gero et al., 2014). During this period, large males roam around, apparently avoiding one another while visiting groups of females (Cantor et al., 2019) and having limited social interactions with members of the social units (adult females and/or immatures; Gero et al., 2014). The existence of geographical and/or social fidelity is questioned in males, however, fidelity of adult males to the ocean of their birth (i.e., a large geographical scale natal philopatry) has been suggested by whaling reports (Best, 1979). Using genetic assignment, Mesnick et al. (2011) highlighted that, in the North Pacific, a higher-than randomly expected proportion of males returned to their population of origin to mate. Males sharing possible first order kinships have also been identified in the Azores and in the Chagos Archipelago (Pinela et al., 2009; Alexander et al., 2016). Photo-identification recaptures of a same male over several years in the same study area occurred in different breeding grounds of the Atlantic (in the Azores and the West Indies; Gero et al., 2014; Van der Linde and Eriksson, 2020) and of the Pacific (the Galapagos; Christal, 1998), where they may socialize with different social groups of the same vocal clan (Rendell et al., 2005). Gero et al. (2014) suggested that male fidelity to breeding sites might occur, based on the identification of the same male spanning a period of 10 years and the observation of a gathering of dozens of females and immatures around a male.
Altogether, these results suggest that some level of geographical and social fidelity could exist in male sperm whales. This hypothesis requires more evidence to be confirmed, however, long-term monitoring of adult male sperm whales is difficult. Few studies have included males in analyses when studying female social groups (e.g., Coakes and Whitehead, 2004; Rendell et al., 2005; Pinela et al., 2009; Gero et al., 2014), and this scarcity of data prevents clear conclusions concerning male sperm whale movement patterns and social fidelity being drawn.
In the Indian Ocean breeding grounds, sperm whales have been less studied than in the Pacific and the Atlantic. Several social groups have been observed (Gordon, 1987; Whitehead and Kahn, 1992; Sarano et al., 2021, 2022), and photoidentification campaigns and satellite tracks confirmed that sperm whales are common near the Mauritius and La Reunion Islands (Huijser et al., 2020; Chambault et al., 2021; Sarano et al., 2022). The predominant matrilineality of a particular social group, “Irène’s group” has been recently demonstrated near Mauritius (Sarano et al., 2021). However, except for some photo-identified individuals (Sarano et al., 2022), male sperm whales encountered within the breeding grounds of the Indian Ocean are very poorly known. More knowledge comes from the feeding grounds of the Indian Ocean, and in particular from Crozet/Kerguelen (Janc et al., 2018; Labadie et al., 2018; Richard et al., 2020), although the movement patterns between feeding and breeding grounds are not known.
In this study, we investigated the spatial and social fidelity of adult male sperm whales in the Indian Ocean. Using 9 years of monitoring based on underwater observation of sperm whale social groups off Mauritius paired with genetic information collected on individuals from both this area and the Crozet/Kerguelen region, our aims were to: (i) Assess the association patterns and genetic relatedness of adult males with the members of a resident social group with which they associate; (ii) determine the extent of genetic relatedness across adult males, and, (iii) analyze possible social and geographical fidelity of adult male sperm whales, including whether they show fidelity to their natal social group.
Field work took place off the western coast of Mauritius (Mascarenes Islands, Indian Ocean) between latitudes 20.465°S 57.334°E and 19.986°S 57.605°E, up to 15 km off the coast (Sarano et al., 2021). Sea surface and underwater observations have been carried out since 2011, during the morning (from 7.30 to 12.00 a.m.), under the auspices of a project called Maubydick led by the Marine Megafauna Conservation Organization (MMCO; Sarano et al., 2021, 2022). Since 2015, fieldwork has been conducted on a regular basis between February and May, and some sporadic observations made during the rest of the year, except in January (Supplementary Table 1).
Sperm whales were identified based on specific morphological characteristics (e.g., marks on caudal and pectoral fins and body marks, described in detail in Sarano et al., 2022). A direct sex assignation was made by underwater observation of the genital slit. An “Identity card” was established for each individual and these used to construct a catalog of individuals (Sarano et al., 2022). During underwater observation non-invasive samples from individually identified sperm whales were collected from sloughed skin fragments as described by Sarano et al. (2021).
Samples were taken only when the releasing individual could be identified, and only when a very limited number of individuals were present (e.g., no skin samples were sampled in the presence of more than three socializing individuals).
The Crozet and Kerguelen archipelagos (Crozet/Kerguelen), located in the subantarctic waters of the south Indian Ocean (respectively, 46 and 49°S), are part of the French TAAF (Terres Australes et Antarctiques Françaises). One sperm whale sample came from a stranded male found on the shore of Kerguelen in 2007. The other samples (n = 8) were collected between 2011 and 2018 from fishing vessels targeting Patagonian toothfish (Dissostichus eleginoides), a fish species that sperm whales consume both naturally and on fishing gear by removing catches (depredation behavior) (Roche and Guinet, 2007; Tixier et al., 2010; Richard et al., 2020). One sample was taken from a dead individual entangled on a longline (Richard et al., 2020) and the others were biopsies collected with a crossbow (Barnett Rhino or Barnett Wildcat), which fired a hollow-tipped biopsy dart with a floatable head (Lambersten, 1987; Tixier et al., 2019). All samples were preserved in absolute ethanol. The sampling of sperm whales at Crozet/Kerguelen was approved by the Comité de l’Environnement Polaire and the French Ministry of Research (04040.03).
All molecular analysis followed the same methodology as previously described (Alfonsi et al., 2012; Decker et al., 2017; Sarano et al., 2021). Briefly, genomic DNA was extracted from the skin and biopsy samples using the NucleoSpin DNA RapidLyse® kit (Macherey-Nagel, Düren, Germany). DNA concentrations were standardized to 10 ng/μL. Several molecular analyses were performed for each sample including molecular sexing (Richard et al., 1994), sequencing of a 638 bp fragment of the mtDNA control region (MCR: amplified with the primers DLP1.5 and DLP8G; Garrigue et al., 2004) and genotyping of 18 microsatellites loci (Supplementary Table 2).
mtDNA sequences were manually edited and aligned with Geneious Pro v.7.1 (Biomatters Ltd., Auckland, New Zealand). The 638 bp long MCR fragment used is the same region used in Sarano et al. (2021). This fragment overlapped fully with the data from Morin et al. (2018) and partially (602 bp in common) with the sequences determined by Alexander et al. (2016). It also overlapped fully with the 283 bp fragment and partially with the 563 bp fragment (514 bp in common) determined by Day et al. (2021). A new dataset that included all these sequences was constructed to allow a large-scale comparison between mitochondrial haplotypes. The numbers of haplotypes, the haplotype diversity (H) and the nucleotide diversity (π) were calculated using the program DnaSP, V.5.10.01 (Librado and Rozas, 2009). The software Arlequin, V3.5.1.2 (Excoffier et al., 2005), was used to calculate FST and ΦST, fixation index estimators for mitochondrial genomes.
Fragment sizes were determined using the “Microsatellite Plugin” of Geneious Pro v.7.1 (Biomatters Ltd., Auckland, New Zealand). All the molecular analyses were performed in at least two independent experiments, from different samples of a same individual when available, or twice from the same sample following Sarano et al. (2021). Twenty-two individuals sampled at least three times between 2017 and 2020 (Supplementary Table 3) allowed us to estimate the microsatellite-genotyping errors linked to possible poor-quality DNA extracts. We calculated an overall error rate of 2.1% per allele (52 alleles incorrect among the 2,432 scored) with this error rate then used in kinship analyses.
The procedure of anonymization of the samples described in Sarano et al. (2021) was also applied to all the samples of this new study to confirm the correspondence between field-identification of individuals (here 13 adult males and an immature female, Chesna sampled only in 2020) and genetic individuals, identified by matching genotypes in the laboratory. Briefly, when collected in the field, each skin sample was assigned to one of the individuals identified and then anonymized with an alphanumeric code. To confirm the validity of the field identifications of skin samples, all the steps of the genetic analyses were performed with anonymized skin samples: samples taken from the same individual were confirmed based on similar genotypes using the Identity Analysis function in CERVUS (Kalinowski et al., 2007) as described in Sarano et al. (2021). Genetic individuals and their corresponding samples are listed in Supplementary Table 3.
Kinship analyses were performed on the complete dataset (with duplicate samples removed), that is adult females and immatures previously analyzed (Sarano et al., 2021) with the newly sampled Chesna (sampled in 2020, Supplementary Table 3), and all the males sampled in Mauritian waters (n = 13) and in Crozet/Kerguelen (n = 8). Kinship analysis followed the same methodology as described in Sarano et al. (2021). Briefly, we first used different estimators to calculate the relatedness coefficient r between all the genotyped individuals using the R package Related (Pew et al., 2014) and the software ML relate (Kalinowski et al., 2006). Related was used to determine that the r estimators W (Wang, 2002) and L&L (Li et al., 1993) had the highest correlation between observed and expected relatedness values and were thus selected to calculate the relatedness coefficients (Supplementary Figure 1). Overall, first degree relationships, second degree relationships and unrelated individuals are distinguished by relatedness estimators, although a minority of the simulated results for different levels of relatedness overlap (Supplementary Figure 1). This means that the inferred relationships may include false positives or negative. ML relate (Kalinowski et al., 2006) was used to calculate a relatedness coefficient based on the probabilities of sharing alleles identical by descent, and to assign the most probable familial relationships [among parent–offspring (PO), full sibling (FS), half-sibling (HS), unrelated (U)] to each dyad.
The software Cervus 3.0.7 (Kalinowski et al., 2007) was also used to assign likely PO relationships. Based on the combined results of these analysis, all probable first- and second-degree kin relationships (Blouin, 2003) were listed. The consistency between familial relationships hypothesized by ML relate and r coefficient calculations was analyzed for each dyad (see also Sarano et al., 2021 for a more detailed explanation about this procedure).
A total of 26 adult males were identified by underwater observations between 2011 and 2020 off Mauritius (Supplementary Table 4). Males were observed in 2011, 2013 and yearly since 2015 when the observation effort significantly increased (Sarano et al., 2022). Since then, adult male sperm whales were sighted each year with a maximum of 10 different individuals observed in 2019. Adult males were observed during a total of 59 days over the 2015–2020 period with a maximum of 29 days in 2019 (Supplementary Table 4). Observations of adult males occurred most of the year with at least one male seen each month from February to December. Over the 2015–2020 period of observations, April was the month with the highest rate of identification (seven males). Almost half of the males were identified on at least two different days within or between years (n = 11), 15 were seen only once. When multiple sightings of the same male occurred during a given year, the longest span between the first and the last sightings was 47 days (Léonard and Jason in 2019), with a mean of 8.25 days (range = 1–47 days) (Supplementary Table 4). Three males were positively identified over multiple years: Jonas, sighted in 2018 and 2019; Navin, sighted in 2015 and in 2018; and Hugues, sighted in 2013 and again 6 years later in 2019 (Supplementary Table 4).
Different socializing behaviors were observed between adult females and/or immatures of the Irene’s group. Figure 1 shows an example of an adult male (Reza) surrounded by an adult female and seven immatures (five males and two females) of the Irène’s group. This kind of socializing behavior between an adult male and several immatures is not uncommon since it was observed and filmed 16 times in 2019, and involved 5 different adult males: Daniel, Reza, Léonard, Jason, Jonas.

Figure 1. Social interactions between an adult male (Reza, Re), an adult female (Germine, G) and different immatures of the Irène’s group: Eliot (E) 8 years-old; Arthur (A) and Romeìo (Ro) 6 years-old; Ali (Al) Daren (D) and Chesna (C) 1 year-old, and Miss Toutou (M) 3 years-old.
The arrival of Jonas and Aman in July 2018 was also a particularly interesting event: this arrival may have initiated the large gathering of females and immatures of different social units. At least 60 females and immatures were observed at this time (MMCO, Field report of the July 18, 2018). Social interactions (e.g., swimming together) were also observed between adult males present in Mauritian waters at the same time. The most striking example of these social interactions was that of Jason and Léonard. Throughout their presence, from April 23, 2019 to June 8, 2019, they were observed together at each observation (n = 11) (Supplementary Table 4).
A total of 132 sloughed skin samples were collected between 2017 and 2020 (Supplementary Table 3). They were assigned in the field to 41 different sperm whales, i.e., to 18 adult females and 10 immatures (previously analyzed in Sarano et al., 2021 except for Chesna, sampled only in 2020) and to 13 adult males (Supplementary Table 3). Mitochondrial and nuclear loci were amplified, allowing an analysis of variation over 638 bp of the MCR (Genbank references: MK907146-MK907148, MK907159, MK907163, MK907172, and MW929445-MW929452) and at 16 polymorphic microsatellite loci (Supplementary Table 2 and Supplementary Data 1). The Identity Analysis based on microsatellite polymorphisms performed in CERVUS identified thirteen genetically distinct individuals from Mauritius corresponding to the 13 adult males identified in the field (all pID < 2.45e–12). All genotypes assigned to the same individual had between 87.5 and 100% identity, and the differences were all consistent with allelic drop out. Mitochondrial haplotypes were all 100% identical between samples of the same individual. Only three skin samples had to be reassigned to another sperm whale than the one identified in the field after a posteriori careful examination of video recordings (see Supplementary Table 3; Sarano et al., 2021 for more explanation). Nine samples were taken off Crozet/Kerguelen, among which 8 genetic individuals were identified, Bio_Cro_2011_1 and Bio_Cro_2017 corresponding to the same individual (pID = 2.6e–23). Six MCR haplotypes were detected among the thirteen adult male sperm whales sampled off Mauritius (H = 0.72, π = 0.00265). Five different MCR haplotypes were identified in the eight male sperm whales sampled in Crozet/Kerguelen (H = 0.78, π = 0.00274, Genbank references MW854724-MW854731). Mitochondrial ΦST calculated between males sampled near Mauritius and those sampled in Crozet/Kerguelen was significant (ΦST = 0.136, p = 0.037), and the FST value was just above the significant value fixed to 5% (FST = 0.125, p = 0.055).
In this study, the mitochondrial haplotype names correspond to the geographical places they came from (M: Mauritius, C: Crozet, K: Kerguelen). The correspondence with the haplotypes defined by Alexander et al. (2016) is presented in Supplementary Table 5 and Figure 2. One adult male harbored the SW_M1 haplotype, corresponding to haplotype C of Alexander et al. (2016), characteristic of the Irène’s group (Sarano et al., 2021). Two others had the same haplotype (SW_MCK1) as Claire, the sole adult female of the Irene’s social group with a different MCR haplotype (Sarano et al., 2021), corresponding to the haplotype N.001.001 mainly found in the Seychelles, in the Cocos Islands and in south west Australia by Alexander et al. (2016). Another adult male had the haplotype MCK2 (differing from SW_MCK1 at position 609, Supplementary Table 5), one had the haplotype SW_M3 corresponding to the haplotype KK found almost exclusively in the Indian Ocean off Sri Lanka (Alexander et al., 2016) and off Albany in Australia (Day et al., 2021). Seven males shared the haplotype SW_MC, identical to the haplotype A.001.001, common in the Indian Ocean (Figure 2). The last male possessed a new haplotype, SW_M2, not found previously anywhere else.
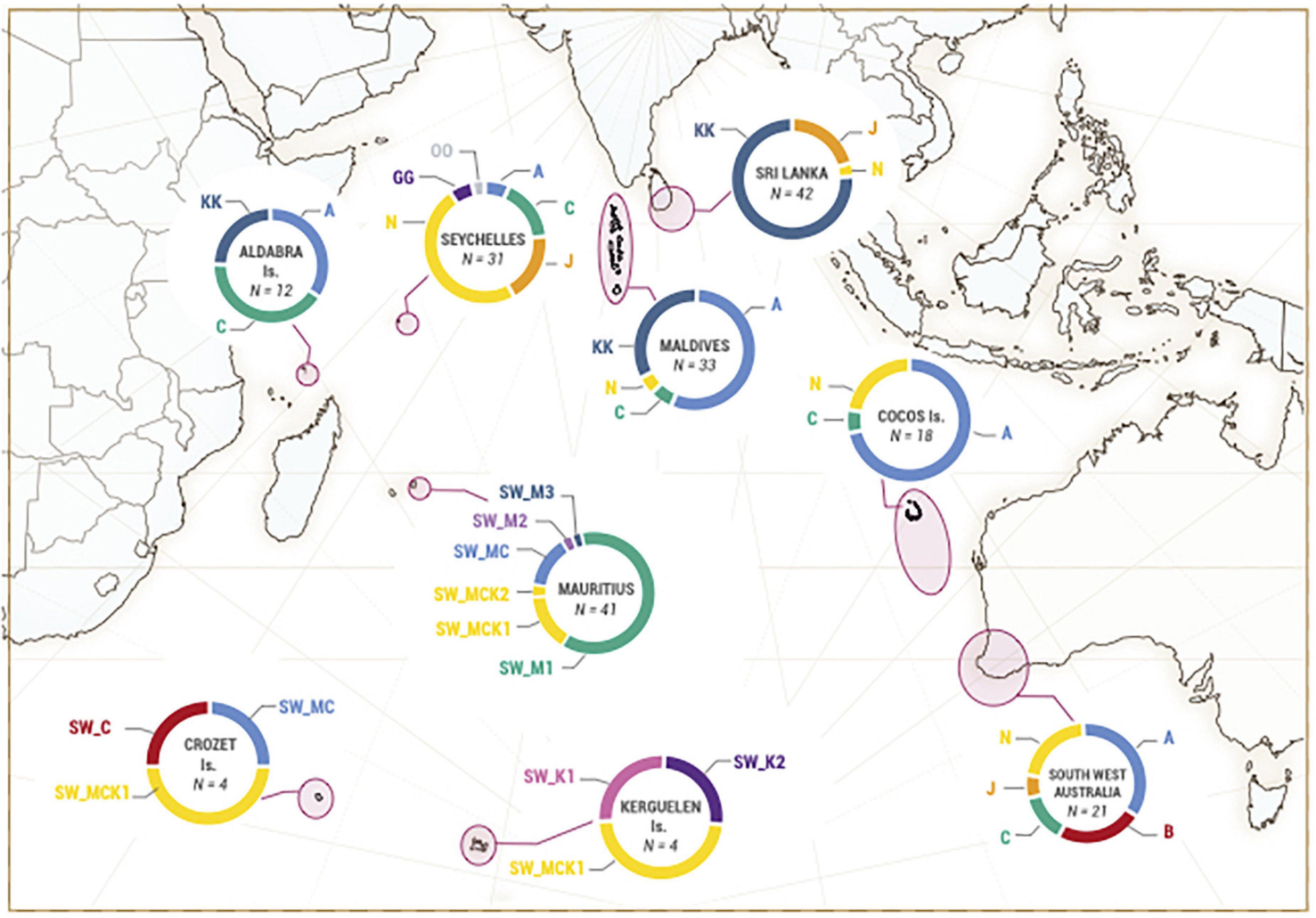
Figure 2. Geographical partitioning of mitochondrial haplotypes in the Indian Ocean determined by Alexander et al. (2016) (haplotypes named with one or two letters) and haplotypes determined during this study (haplotype names starting by SW). Identical colors indicate corresponding haplotypes (602 bp in common, see Supplementary Table 5 for correspondence of haplotype names). N: number of sperm whales for each diagram.
Kinship analysis revealed probable two first- and 21 second-degree kin relationships (12 with adult females, 9 with immatures) between the 13 adult males sampled in Mauritius and members of the Irène’s group (Figure 3, Supplementary Figure 2, and Supplementary Table 6). One adult male, Jonas, was identified as the father of Daren, a young male born in 2018; and a second adult male, Noé, was identified as the father of Lana, a young female born in 2019 (Supplementary Figure 2 and Supplementary Table 6). All but three adult males presented at least one inferred second-degree relationship with members of the Irène’s group with a maximum of four (Josuah and Léonard) (Supplementary Figure 2).
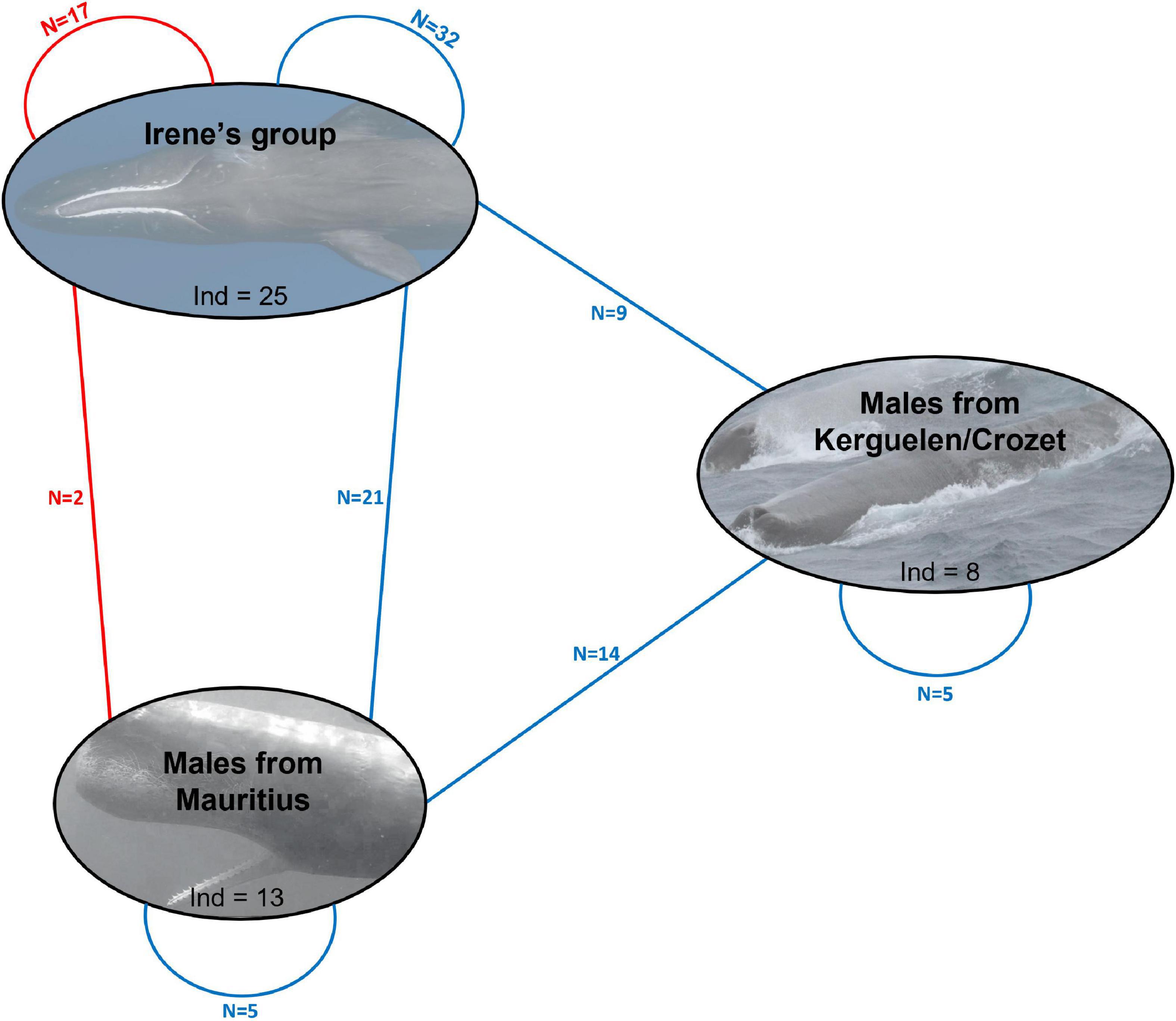
Figure 3. Schematic representation of inferred first degree (in red) and second degree (in blue) kin relationships between the members of the Irène’s group and the adult male sperm whales sampled off Mauritius and in the Sub-Antarctic waters of the South of the Indian Ocean. The number of inferred first-degree (red lines) and second-degree (blue lines) relationships between sperm whales are represented between individuals of a same group (the Irène’s social group, the group of adult males sampled in Mauritius, and the one formed by adult males sampled in Crozet-Kerguelen) and between individuals of the same groups. Detail of these relationships is presented in the Supplementary Figure 2. As stated in Sarano et al. (2021), this diagram was constructed to be consistent with the analyses conducted. Although we performed different analyses that produced similar results, uncertainty exists in the relatedness estimate calculations, which might influence some of these inferred relationships.
Four possible full sibling relationships (same mother and father) have also been discovered in the Irène’s group, two between immatures and two between adult females, three of which were highly probable: Adélie-Emy, Alexander-Zoé and Chesna-Tache Blanche (Supplementary Table 7).
Two haplotypes (SW_MCK1 and SW_MC) were found both in Crozet/Kerguelen and in Mauritian males. SW_MCK1, shared by four sub-Antarctic sperm whales (two sampled in Crozet and two in Kerguelen) was the most frequent. The haplotype SW_MC was found in one sperm whale from Crozet (Figure 2). Three other haplotypes were found in the Crozet/Kerguelen samples that were not observed among males sampled off Mauritius: SW_K1 and SW_K2, found in two sperm whales sampled in the Kerguelen and SW_C, found in one sperm whale in Crozet. SW_K1 matched the haplotype 10 defined by Day et al. (2021) found off South Australia and Victoria, and SW_K2 corresponded to the haplotype GG Alexander et al. (2016), exclusively found in the Indian Ocean in the Seychelles. SW_C corresponds to haplotype B (Alexander et al., 2016; Day et al., 2021), found in Australia (Figure 2).
Males from Kerguelen/Crozet shared no first-degree relations with the Irene’s group and had fewer probable second-degree relationships (n = 9, among which only two are found with immatures of the Irène’s group) than Mauritian males (Figure 3 and Supplementary Figure 2). However, some of these males shared high relatedness coefficient with members of the Irène’s group (for example Mystère and PM_KER_2007, r = 0.38). Among all adult males sampled off Mauritius or in the south of the Indian Ocean, 24 inferred second-degree relationships were identified (Figure 3, Supplementary Figure 2, and Supplementary Table 6).
During this study, 22 sperm whales (21 adult males and 1 immature female) were added to the 27 already analyzed in Sarano et al. (2021). The 49 sperm whales in total included in this study were the 25 members of the Irène’s social group, 2 members of another social group, “the Reshna group,” one unidentified female, 13 adult males sampled off Mauritius, and the 8 adult males sampled in Crozet/Kerguelen (the complete list is given in Supplementary Table 3). The mean relatedness of these different samples and of different combinations were calculated (Figure 4 and Supplementary Table 8). Across all the included individuals (in Mauritius and in Crozet/Kerguelen), we calculated an average r = 0.046, similar to that calculated between all adult males (r = 0.044; Supplementary Table 8). As expected, members of the mostly matrilineal Irène’s group had a higher average pairwise r (r = 0.067, Figure 4).
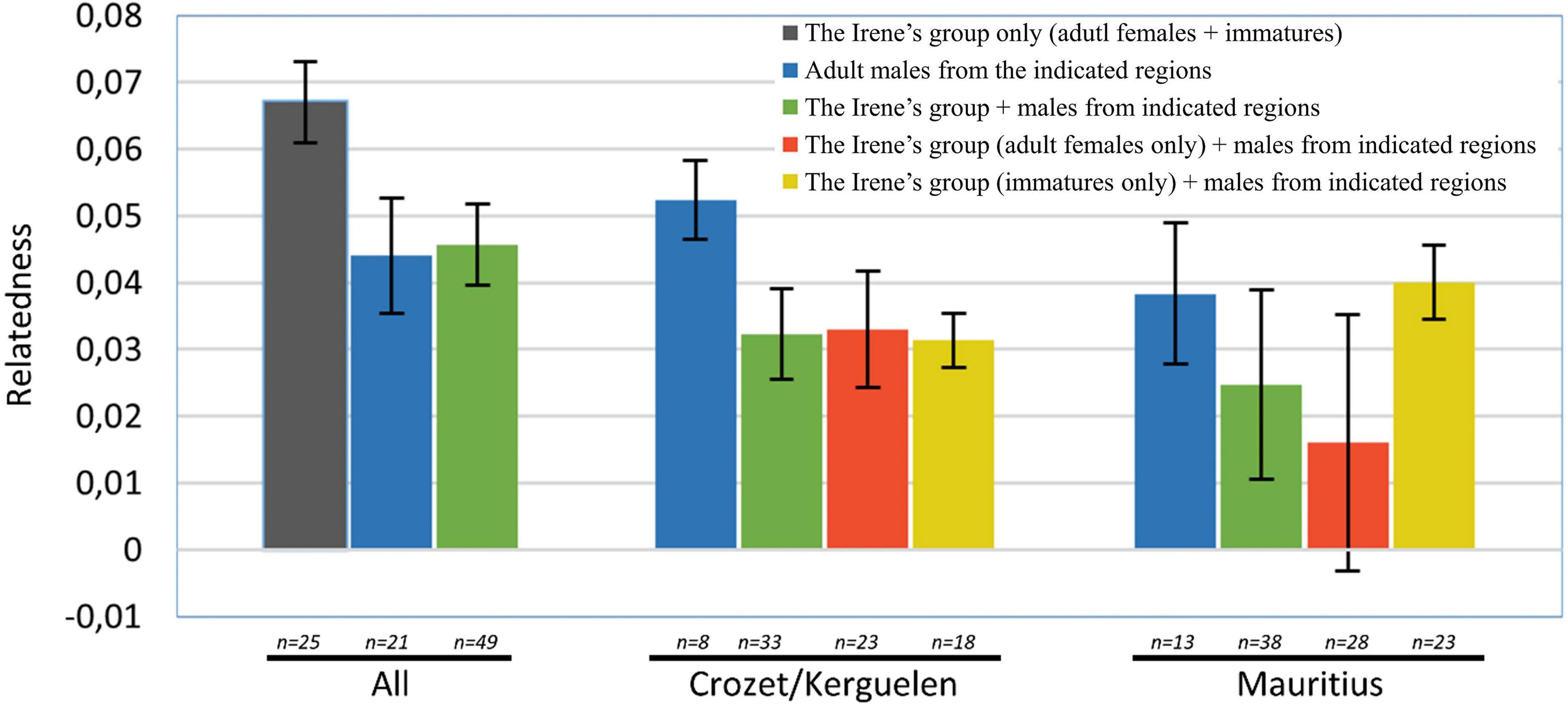
Figure 4. Differences of average relatedness coefficients in groups and subgroups. Relatedness coefficients computed as the average of rk (Kalinowski et al., 2006), rW (Wang, 2002) and rL (Li et al., 1993) calculated through ML Relate and through Relate. The relatedness coefficients are first represented for all members of the Irène’s group, for all adult males and for all individuals. Note that the values of the Irène’s group are the higher (shown in dark gray, see Supplementary Table 8 for detail). Different combinations of individuals were then formed, and the relatedness coefficients calculated. The partitioning of the Irène’s group between adult females and immatures had a strong impact on the r calculated with adult males sampled off Mauritius, but not with those from Crozet/Kerguelen.
Average relatedness values were higher among males sampled in Crozet/Kerguelen (r = 0.052) than between these males and members of the Irène’s group, whether the Irène group was partitioned into adult females only, immatures only, or the entire group (r = 0.031–0.033, Figure 4 and Supplementary Table 8).
In contrast, and despite variation among the different metrics of relatedness, all of the metrics show the same overall pattern (Supplementary Table 8): The partitioning of the Irène group had an impact on the relatedness values in comparison to males sampled in Mauritius. Average relatedness was higher between juveniles from the Irene’s group and males sampled in Mauritius (r = 0.040), than between adult females from the Irene’s group and the same group of males (r = 0.016, Figure 4 and Supplementary Table 8).
Currently, our knowledge of behavior, ecology and genetic diversity of iconic marine megafauna still suffers from holes. An outstanding example concerns male sperm whales, the largest toothed creature on Earth (Cantor et al., 2019). Sperm whales are steeped in our culture, from the star of one of the most-read novels (Melville, 1851) to the use of their spermaceti oil during the industrial revolution (e.g., Whitehead, 2002). However, social and breeding behaviors of male sperm whales remain largely unclear, especially in terms of geographical and social fidelity. Here, we studied sperm whales off Mauritius under the auspices of the Maubydick project (Sarano et al., 2021, 2022) and off Crozet/Kerguelen (Janc et al., 2018; Labadie et al., 2018; Richard et al., 2020). This allowed us (1) to document the presence of different males visiting the focal mostly matrilineal sperm whale social unit, the Irène group; (2) to identify several recaptures of males with the Irène group over years; (3) to decipher some paternal kinships as well as (4) to capture a diagram of kin relationships at a larger geographic scale. Based on this, we infer that adult males can show social and geographical fidelities to breeding and feeding areas within the Indian Ocean.
Natal philopatry can be defined as fidelity to birthplace and has been evidenced in different species of marine mammals (e.g., Baker et al., 2013; Rendell et al., 2019). Among the 13 adult males sampled in Mauritian waters, 12 did not share the SW_M1 haplotype characteristic of the Irène social group (Sarano et al., 2021), and can therefore not have been born in this group. Only one, Herman, had the SW_M1 MCR haplotype, but mitogenome sequencing revealed seven mutations between Herman’s and the predominant Irène’s group mitogenome (Justine Girardet, Agnès Dettaï and Jean-Luc Jung, Supplementary Data 2). Nuclear DNA analysis is consistent with this statement (i.e., an absence of natal philopatry): The lowest average r calculated for any combination of individuals in our study, was between the adult female members of the Irène’s group and males sampled off Mauritius (Figure 4 and Supplementary Table 8).
In contrast to the lack of natal philopatry of adult males demonstrated by our analyses, we found seven instances (three confirmed by resightings over multiple years, and four correlated to gametic recaptures) of males coming back several times to the same area and to the same social unit to breed and/or socialize. These are strong indications that adult male sperm whales may show social fidelity to particular female-dominated social groups, not based on kin relationships with adult females in the group, and that, in turn, they could be well known by the members of these female-dominated social groups.
Nuclear DNA analysis revealed two father-offspring relationships between adult males sampled off Mauritius and immature members of the Irène’s group. One paternity has been attributed to Jonas (father of Daren born in 2018), and one to Noé (father of Lana born in 2019), both sampled in 2018 (Supplementary Table 4). These “gametic” recaptures (Garrigue et al., 2004) proved that some of the males observed in Mauritian waters are reproductive. This reproductive status is supported by the value of the average r calculated between males from Mauritius and members of the Irène group, which is nearly tripled if immatures of the Irène’s group alone are considered as compared to adult females of the group (Figure 4 and Supplementary Table 8). The presence of Jonas in the Irène’s group was highlighted over at least three different years (its presence was inferred in 2017 for mating as proved by the “gametic” recapture, and by 2 years of direct observation, in 2018 and 2019).
In addition, nuclear DNA analysis revealed three potential full sibling relations. Two were detected between immatures (Alexander and Zoé born in 2019 and 2013, Chesna and Tache Blanche born in 2018 and 2011). The other one was between adult females (Adélie and Emy) whose years of birth are unknown. Sperm whale twins are very rare. They were recorded in about 0.5% of cases in three different studies (3854 fetuses analyzed in total, reviewed in Best et al., 1984), so it is very likely that Adélie and Emy were not born in the same year. Thus, the fathers of each of these three pairs came back at least in two different years to the same group—and to the same specific receptive female—to mate. The father of Chesna and Tache blanche could in addition be the father of Eliot, supposed half-brother of Tache Blanche (Supplementary Table 6). Despite these gametic recaptures being based on relatedness estimate calculations, and therefore subject to uncertainties, these findings provide powerful evidence in support of enduring relationships between adult males and specific female-dominated social groups.
Most fieldwork was conducted between February and May (Supplementary Table 1). Nevertheless, it is of note that the three males recaptured between years were seen at the same period of the year (Hugues in October 2013 and October-November 2019, Navin in July 2015 and June 2018, Jonas in July 2018 and May-June 2019), mainly during months when fewer observations were made; therefore the non-uniform observation effort over the year cannot be at the origin of these possible privileged periods of presence. This could indicate either a certain degree of seasonality specific to each individual, or, if they are visiting different female-dominated social groups, a difference in the order that each social group is visited between males. The case of Jonas stands out: Jonas was observed in 2018 and 2019, he is the likely father of Daren, born in 2018, and his arrival, with another male Aman, coincided with the gathering of tens of females and immatures in 2018. Jonas has therefore a marked and repeated social fidelity for the Irène’s group. It cannot be excluded that he is in turn well known to the group members. As suggested by Gero et al. (2014), spectacular gatherings could also support the hypothesis that females play a role in mating choice. Sexual selection could be based on body size or vocalization as described (Cranford, 1999). Given the familiarity of juveniles (both males and females, Figure 1) with some adult males, this alternative hypothesis alone could not explain the large gathering observed.
Male sperm whales were present in the Irène’s social group most of the year with a peak of occurrence in April and May during the austral autumn, which could represent the breeding season. Janc et al. (2018) and Labadie et al. (2018) highlighted a seasonality in occurrence of sperm whales in the high latitude feeding area of the Indian Ocean, with increased sightings in spring and summer. However, observations in Mauritius are only conducted on a regular basis from February to May, thus the number of males identified in each month could be biased in other months by lower observation effort, therefore reproduction throughout the year cannot be excluded. Residency of males off Mauritius appears to be on the scale of a few days to few weeks with an average stay (8.25 days), twice as high as that previously reported off Dominica, for example (3.76 days) (Gero et al., 2014). Recurrent interactions between adult males and members of the social unit have been observed, confirming previous observations (e.g., Gordon et al., 1998; Gero et al., 2014). Interactions between adult males and adult females and/or immatures have already been reported, for example in Northern Chile and off Dominica (Coakes and Whitehead, 2004; Gero et al., 2014). Here, the males identified were often observed in proximity (i.e., less than 100 m) of members of the Irène’s social group and several types of interactions (e.g., physical contact, vocal interactions) were recorded with both adult females and immatures. The exceptional gathering of tens of individuals,—which probably represent a substantial proportion of the local population -, after the arrival of two adult males in the Mauritian waters (MMCO, Field report of July 18 2018) seems not to be restricted to the Indian Ocean: Gero et al. (2014) observed a similar aggregation of several tens of individuals near an adult male in the Atlantic. Although there may be other explanations (i.e., a form of sexual selection from the female), some males appear therefore to be well known to particular stable social groups. This assumption is reinforced by the numerous interactions observed between adult males and members of Irene’s group (adult females and juveniles), and by the several potential full sibling relationships identified.
While all members of the Irène’s group except one harbored the same MCR haplotype (Sarano et al., 2021), adult male sperm whales showed a mtDNA diversity in the same range of what was calculated by Alexander et al. (2016) for the broader Indian Ocean (Haplotype diversities around H = 0.8, nucleotide diversities around π = 0.0028). The haplotypes identified in this study near Mauritius and matching to Alexander et al. (2016) haplotypes all corresponded to minor and major haplotypes of the Indian Ocean. In Crozet/Kerguelen, mtDNA haplotypes suggest a widespread geographic origin of adult male sperm whales: they match to North Indian Ocean haplotypes identified from the west to the east of the Ocean (Figure 2). Even though we sampled only limited numbers of male sperm whales, tests of differentiation based on mtDNA detected some levels of genetic differentiation between Mauritius and Crozet/Kerguelen (ΦST and FST significant or nearly so), which reflect divergent distribution of mtDNA haplotypes between the two sites, although a high number of second- and third-degree relationships were inferred between males sampled in the two areas (although, as noted earlier, it must be kept in mind that false positives and false negatives may exist when inferring relationships).
While the mtDNA results likely reflect the widespread origin of males at specific geographic locations, nuDNA polymorphisms support male-mediated gene flow at large scales, and highlight the reproductive status of males sampled off Mauritius. The average relatedness r calculation revealed similar and informative patterns. Between males sampled in Crozet/Kerguelen and members of the Irène’s group, the average r is similar when subsetting to adults or immatures of the Irène’s group. Therefore, males sampled in Crozet/Kerguelen do not appear to breed preferentially with the Irène social group. This situation is strongly contrasting with the pattern observed for males sampled in Mauritius, where their role of as paternal relatives was demonstrated by a three times higher average relatedness with immatures than with adult females (Figure 4). Differences between the different estimators used appear to stem from differences in the methods of calculation. For example, the estimator of Wang (2002), which can be considered as an improved version of that of Li et al. (1993) and valuable for small sample sizes, showed greater variance (Supplementary Table 8).
Male recaptures and social interactions between males and members of social groups have already been observed and suggest some level of male social fidelity in breeding areas in the Pacific (Rendell et al., 2005) and in the West Indies (Gero et al., 2014). Here, we confirm and extend these observations in the Indian Ocean. The level of this male social fidelity (e.g., for social units, for vocal clans, defined in Konrad et al., 2018) is still to be evaluated.
Our results suggest that this fidelity is not due to natal social philopatry, i.e., fidelity for the social group of birth. It appears that natal social philopatry is exclusive to female sperm whales. Therefore, males must acquire their fidelity for places and groups other than that of their birth and based on the diversity of mtDNA haplotypes observed in males, this might occur across large geographical scales.
The high mtDNA diversity found in male sperm whales (as compared to the almost complete absence of diversity found in the group of Irène) is likely to reflect disparities in their respective birth places. Alexander et al. (2016) found that, in the Indian Ocean, 44.4% of the variance in mtDNA frequencies was explained by regions, and 12.3% by social groups. If the mostly matrilineal nature of the Irène’s group (Sarano et al., 2021) is a more or less general rule for sperm whale social units in the Indian Ocean, the geographical patterns of mtDNA distributions found by Alexander et al. (2016) may well correspond to discrete regional partitions of social units, more than to different proportions of mtDNA haplotypes in different populations, found for instance in humpback whales (e.g., Baker et al., 2013; Richard et al., 2018). This would be explained by the strong natal social philopatry of females (more than by a natal geographical philopatry). Interestingly, the situation could well be different in the Pacific, where sperm whale social groups could be of larger size and aggregate more often (Whitehead and Kahn, 1992), and where partitioning of variance in mtDNA has been explained by social groups and not by regional differences (Alexander et al., 2016).
The number of adult male sperm whales sampled off Mauritius is relatively low (n = 13), but it is nevertheless notable that their mtDNA haplotypes are frequent in different regions of the Indian Ocean neighboring Mauritius. In contrast, sperm whales sampled in the Crozet/Kerguelen (n = 8) have haplotypes found in a much broader area covering all the north of the Indian Ocean, from west to east (this study; Alexander et al., 2016; Day et al., 2021). This is reflected by significant or nearly significant ΦST and FST values between Mauritian and Crozet/Kerguelen males. Mesnick et al. (2011) suggested that, in the North Pacific, male sperm whales from different regions mix in feeding grounds and exhibit some degree of geographical philopatry for the region of their birth when breeding. Our results highlight a lack of natal philopatry of male sperm whales at the social unit scale but they could well fit into the Mesnick et al. (2011) hypothesis, with a certain degree of philopatry at a larger geographic scale (here, an area corresponding more or less to the north west of the Indian Ocean). As in the North Pacific (Mesnick et al., 2011), and still remaining cautious because of the low number of samples in our study, the high latitude feeding areas in the Southern Indian Ocean could host mixed groups of male sperm whales with a widespread geographic origin, larger than in the breeding areas. These observations are in agreement with previous population genetic studies, highlighting a strong female philopatry and male-mediated gene flow (Lyrholm et al., 1999; Engelhaupt et al., 2009; Alexander et al., 2016).
Our results strongly suggest that a double fidelity of adult male sperm whales for breeding and feeding grounds exists in the Indian Ocean: (i) a certain level of male fidelity has been detected in feeding grounds of the Indian Ocean (Labadie et al., 2018, and our results, a same male has been sampled in 2011 and 2017 off Crozet), and (ii) our study highlights the existence of a social and geographical fidelity in a sperm whale breeding area of the south west of the Indian Ocean.
Until now, sperm whales were not believed to follow defined migration routes (Cantor et al., 2019), but, at least in the Indian Ocean, as some degree of fidelity is now proved both for breeding and feeding areas, male sperm whales could well take similar routes to migrate on successive years, also supported by the similar time of year distinct males were observed when resighted between years. Estimating the strength of both fidelities as well as long-term satellite tags could help to test this hypothesis.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: NCBI (accession: MK907146-MK907148, MK907159, MK907163, MK907172, MW854724-MW854731 and MW929445- MW929452).
Ethical review and approval was not required for the animal study because permission to conduct the Maubydick project, including the taking of sloughed skin fragments, was granted by the Department for Continental Shelf, Maritime Zones Administration and Exploration of the Mauritius Prime Minister Office, on the 21 February 2017. Skin samples were sent to Brest (France) under the CITES agreement FR1702900025-I. The sampling of sperm whales at Crozet/Kerguelen was approved by the Comité de l’Environnement Polaire and the French Ministry of Research (04040.03).
FS and J-LJ designed the study. FS, JG, GR, PT, CG, AA, and J-LJ contributed variously to the conception of the project. FS, VS, RH, AP, GR, PT, CG, and HV performed the field experiments and identification of individual sperm whales. JG, J-LJ, and AG-C conducted the genetic analysis (laboratory procedures). JG and J-LJ analyzed, interpreted the genetic data, and wrote the manuscript. PT, AA, FS, VS, BM, AG-C, GR, CG, HV, and OA critically revised the manuscript. All authors contributed to the article and approved the submitted version.
The work in the sub-Antarctic region was supported by the French Polar Institute (IPEV program ORNITHOECO-109). The Lush Foundation partly supported the genetics lab work costs.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We thank Navin Boodhonee and the Blue Water Diving Center (Trou aux Biches, Mauritius) for their all years round field work. We thank to Carole Decker (Brest, France) for her help with the lab experiments, and to Marine Pensec and Cédric Le Maréchal (Laboratoire de génétique moléculaire et d’histocompatibilité, CHRU, Brest France) for their help with microsatellite analysis. We are very grateful to Marion Sarano (https://marionsarano.fr/graphiste-freelance/) who made the drawing of Figure 2. We also thank the Terres Australes et Antarctiques Françaises, the Muséum National d’Histoire Naturelle de Paris, the toothfish fishing companies (SARPC), the fishers and fishery observers aboard toothfish longliners of Crozet and Kerguelen for their support in collecting data on sperm whales over the years. Mauritian public authorities greatly helped the Maubydick project, in particular the Mauritian Prime Minister Office, the Marine Continental Shelf Exploration and Administration (MCSEA; Dr. Réza Badal and his team), the Albion Fisheries Research Center (AFRC; Chief Scientific Officer Mr. Satish Kadhun), the Mauritius Film Development Corporation (MFDC; Mr. Sachin Jootun et Miss Eliana Timol), and the Tourism Authority (TA; Miss. Khoudijah Boodoo, Ex-Director).
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2022.815684/full#supplementary-material
Alexander, A., Steel, D., Hoekzema, K., Mesnick, S. L., Engelhaupt, D., Kerr, I., et al. (2016). What influences the worldwide genetic structure of sperm whales (Physeter macrocephalus)? Mol. Ecol. 25, 2754–2772. doi: 10.1111/mec.13638
Alfonsi, E., Hassani, S., Carpentier, F.-G., Clec’h, J.-Y. L., Dabin, W., Canneyt, O. V., et al. (2012). A european melting pot of harbour porpoise in the French Atlantic coasts inferred from mitochondrial and nuclear data. PLoS One 7:e44425. doi: 10.1371/journal.pone.0044425
Baker, C. S., Steel, D., Calambokidis, J., Falcone, E., González-Peral, U., Barlow, J., et al. (2013). Strong maternal fidelity and natal philopatry shape genetic structure in North Pacific humpback whales. Mar. Ecol. Prog. Ser. 494, 291–306. doi: 10.3354/meps10508
Beerman, A., Ashe, E., Preedy, K., and Williams, R. (2016). Sexual segregation when foraging in an extremely social killer whale population. Behav. Ecol. Sociobiol. 70, 189–198. doi: 10.1007/s00265-015-2038-2
Best, P. B. (1979). “Social organization in sperm whales, Physeter macrocephalus,” in Behavior of Marine Animals, eds H. E. Winn and B. L. Olla (Boston, MA: Springer), 227–289. doi: 10.1007/978-1-4684-2985-5_7
Best, P. B., Canham, P., and McLeod, N. (1984). Patterns of reproduction in sperm whales, Physeter macrocephalus. Rep. Int. Whal. Comm. 6, 51–79.
Blouin, M. S. (2003). DNA-based methods for pedigree reconstruction and kinship analysis in natural populations. Trends Ecol. Evol. 18, 503–511. doi: 10.1016/S0169-5347(03)00225-8
Cantor, M., Gero, S., Whitehead, H., and Rendell, L. (2019). “Sperm whale: the largest toothed creature on Earth,” in Ethology and Behavioral Ecology of Odontocetes. Ethology and Behavioral Ecology of Marine Mammals, ed. B. Würsig (Cham: Springer), 261–280. doi: 10.1007/978-3-030-16663-2_12
Chambault, P., Fossette, S., Heide-Jørgensen, M. P., Jouannet, D., and Vély, M. (2021). Predicting seasonal movements and distribution of the sperm whale using machine learning algorithms. Ecol. Evol. 11, 1432–1445. doi: 10.1002/ece3.7154
Christal, J. (1998). An Analysis of Sperm Whale Social Structure: Patterns of Association and Genetic Relatedness. Ph. D. thesis. Halifax, NS: Dalhousie University, 1–212.
Christal, J., and Whitehead, H. (1997). Aggregations of mature male sperm whales on the Galapagos islands breeding grounds. Mar. Mamm. Sci. 13, 59–69. doi: 10.1111/j.1748-7692.1997.tb00612.x
Christal, J., and Whitehead, H. (1999). Sperm whale social units: variation and change. Can. J. Zool. 76, 1431–1440. doi: 10.1139/z98-087
Coakes, A. K., and Whitehead, H. (2004). Social structure and mating system of sperm whales off northern Chile. Can. J. Zool. 82, 1360–1369. doi: 10.1139/z04-115
Cranford, T. W. (1999). The sperm whale’s nose: sexual selection on a grand scale? Mar. Mamm. Sci. 15, 1133–1157. doi: 10.1111/j.1748-7692.1999.tb00882.x
Curé, C., Antunes, R., Alves, A. C., Visser, F., Kvadsheim, P. H., and Miller, P. J. O. (2013). Responses of male sperm whales (Physeter macrocephalus) to killer whale sounds: implications for anti-predator strategies. Sci. Rep. 3:1579. doi: 10.1038/srep01579
Day, J., Power, D., Gales, R., Bannister, J., Piggott, M. P., Bilgmann, K., et al. (2021). Australian sperm whales from different whaling stocks belong to the same population. Aquat. Conserv. Mar. Freshw. Ecosyst. 31, 1452–1465. doi: 10.1002/aqc.3494
Decker, C., Hassani, S., Jezequel, M., Rault, C., Dumas, C., Méheust, E., et al. (2017). Mitochondrial DNA reveals historical maternal lineages and a postglacial expansion of the grey seal in European waters. Mar. Ecol. Prog. Ser. 566, 217–227. doi: 10.3354/meps12003
Engelhaupt, D., Hoelzel, A. R., NicholsonI, C., Frantizs, A., Mesnick, S., Gero, S., et al. (2009). Female philopatry in coastal basins and male dispersion across the North Atlantic in a highly mobile marine species, the sperm whale (Physeter macrocephalus). Mol. Ecol. 18, 4193–4205. doi: 10.1111/j.1365-294X.2009.04355.x
Excoffier, L., Laval, G., and Schneider, S. (2005). Arlequin (version 3.0): an integrated software package for population genetics data analysis. Evol. Bioinform. Online 1, 47–50. doi: 10.1177/117693430500100003
Galezo, A. A., Krzyszczyk, E., and Mann, J. (2017). Sexual segregation in Indo-Pacific bottlenose dolphins is driven by female avoidance of males. Behav. Ecol. 29, 377–386. doi: 10.1093/beheco/arx177
Garrigue, C., Dodemont, R., Steel, D., and Baker, S. C. (2004). Organismal and ‘gametic’ capture-recapture using microsatellite genotyping confirm low abundance and reproductive autonomy of humpback whales on the wintering grounds of New Caledonia. Mar. Ecol. Prog. Ser. 274, 251–262. doi: 10.3354/meps274251
Gero, S., Milligan, M., Rinaldi, C., Francis, P., Gordon, J., Carlson, C., et al. (2014). Behavior and social structure of the sperm whales of Dominica, West Indies. Mar. Mamm. Sci. 30, 905–922. doi: 10.1111/mms.12086
Gerson, H., and Hickie, J. (1985). Head scarring on male narwhals (Monodon monoceros): evidence for aggressive tusk use. Can. J. Zool. 63, 2083–2087. doi: 10.1139/z85-306
Gordon, J. (1987). Sperm whale groups and social behaviour observed off Sri Lanka. Rep. Int. Whal. Comm. 37, 205–217.
Gordon, J., Moscrop, A., Carlson, C., Ingram, S., Leaper, R., Matthews, J., et al. (1998). Distribution, movements and residency of sperm whales off the commonwealth of Dominica, Eastern Caribbean: implications for the development and regulation of the local whale watching industry. Rep. Int. Whal. Comm. 48, 551–557.
Huijser, L. A. E., Estrade, V., Webster, I., Mouysset, L., Cadinouche, A., and Dulau-Drouot, V. (2020). Vocal repertoires and insights into social structure of sperm whales (Physeter macrocephalus) in Mauritius, southwestern Indian ocean. Mar. Mamm. Sci. 36, 638–657. doi: 10.1111/mms.12673
Janc, A., Richard, G., Guinet, C., Arnould, J. P. Y., Villanueva, M. C., Duhamel, G., et al. (2018). How do fishing practices influence sperm whale (Physeter macrocephalus) depredation on demersal longline fisheries? Fish. Res. 206, 14–26. doi: 10.1016/j.fishres.2018.04.019
Jaquet, N., Dawson, S., and Slooten, E. (2000). Seasonal distribution and diving behaviour of male sperm whales off Kaikoura: foraging implications. Can. J. Zool. 78, 407–419. doi: 10.1139/z99-208
Kalinowski, S. T., Taper, M. L., and Marshall, T. C. (2007). Revising how the computer program cervus accommodates genotyping error increases success in paternity assignment. Mol. Ecol. 16, 1099–1106. doi: 10.1111/j.1365-294X.2007.03089.x
Kalinowski, S. T., Wagner, A. P., and Taper, M. L. (2006). ML-relate: a computer program for maximum likelihood estimation of relatedness and relationship. Mol. Ecol. Notes 6, 576–579. doi: 10.1111/j.1471-8286.2006.01256.x
Kobayashi, H., and Amano, M. (2020). Residency and abundance of sperm whales (Physeter macrocephalus) in Nemuro Strait, Hokkaido, Japan. Mar. Mamm. Sci. 36, 612–622. doi: 10.1111/mms.12662
Kobayashi, H., Whitehead, H., and Amano, M. (2020). Long-term associations among male sperm whales (Physeter macrocephalus). PLoS One 15:e0244204. doi: 10.1371/journal.pone.0244204
Konrad, C. M., Gero, S., Frasier, T., and Whitehead, H. (2018). Kinship influences sperm whale social organization within, but generally not among, social units. R. Soc. Open Sci. 5, 180914–180918. doi: 10.1098/rsos.180914
Labadie, G., Tixier, P., Barbraud, C., Fay, R., Gasco, N., Duhamel, G., et al. (2018). First demographic insights on historically harvested and poorly known male sperm whale populations off the Crozet and Kerguelen islands (Southern ocean). Mar. Mamm. Sci. 34, 595–615. doi: 10.1111/mms.12469
Lambersten, R. H. (1987). A biopsy system for large whales and its use for cytogenetics. J. Mammal. 68, 443–445. doi: 10.2307/1381495
Le Boeuf, B., and Laws, R. (1994). Elephant Seals: Population Ecology, Behavior, and Physiology. Berkeley, CA: University of California Press. doi: 10.1525/9780520328150
Lettevall, E., Richter, C., Jaquet, N., Slooten, E., Dawson, S., Whitehead, H., et al. (2002). Social structure and residency in aggregations of male sperm whales. Can. J. Zool. 80, 1189–1196. doi: 10.1139/z02-102
Li, C. C., Weeks, D. E., and Chakravarti, A. (1993). Similarity of DNA fingerprints due to chance and relatedness. Hum. Hered. 43, 45–52. doi: 10.1159/000154113
Librado, P., and Rozas, J. (2009). DnaSP v5: a software for comprehensive analysis of DNA polymorphism data. Bioinformatics 25, 1451–1452. doi: 10.1093/bioinformatics/btp187
Lyrholm, T., Leimar, O., Johanneson, B., and Gyllensten, U. (1999). Sexbiased dispersal in sperm whales: contrasting mitochondrial and nuclear genetic structure of global populations. Proc. R. Soc. Lond. Ser. B. Biol. Sci. 266, 347–354.
Madsen, P., Wahlberg, M., and Møhl, B. (2002). Male sperm whale (Physeter macrocephalus) acoustics in a high-latitude habitat: implications for echolocation and communication. Behav. Ecol. Sociobiol. 53, 31–41. doi: 10.1007/s00265-002-0548-1
Melville, H. (1851). Moby Dick or the Whale. New York, NY: Harper & Brothers. doi: 10.5962/bhl.title.62077
Mesnick, S., and Ralls, K. (2018). “Sexual dimorphism,” in Encyclopedia of Marine Mammals, 3rd Edn, eds B. Würsig, J. G. M. Thewissen, and K. M. Kovacs (London: Elsevier), 848–853. doi: 10.1016/B978-0-12-804327-1.00226-0
Mesnick, S. L., Taylor, B., Archer, F., Martien, K., Trevino, S., Hancock-Hanser, B., et al. (2011). Sperm whale population structure in the eastern and central North Pacific inferred by the use of single-nucleotide polymorphisms, microsatellites and mitochondrial DNA. Mol. Ecol. Res. 11, 278–298. doi: 10.1111/j.1755-0998.2010.02973.x
Mizroch, S. A., and Rice, D. W. (2012). Ocean nomads: distribution and movements of sperm whales in the North Pacific shown by whaling data and discovery marks. Mar. Mamm. Sci. 29, E136–E165. doi: 10.1111/j.1748-7692.2012.00601.x
Morin, P. A., Foote, A. D., Baker, C. S., Hancock-Hanser, B. L., Kaschner, K., Mate, B. R., et al. (2018). Demography or selection on linked cultural traits or genes? Investigating the driver of low mtDNA diversity in the sperm whale using complementary mitochondrial and nuclear genome analyses. Mol. Ecol. 27, 2604–2619. doi: 10.1111/mec.14698
Pew, J., Muir, P. H., Wang, J., and Frasier, T. R. (2014). Related: an R package for analysing pairwise relatedness from codominant molecular markers. Mol. Ecol. Res. 15, 557–561. doi: 10.1111/1755-0998.12323
Pinela, A. M., Quérouil, S., Magalhães, S., Silva, M. A., Prieto, R., Matos, J. A., et al. (2009). Population genetics and social organization of the sperm whale (Physeter macrocephalus) in the Azores inferred by microsatellite analyses. Can. J. Zool. 87, 802–813. doi: 10.1139/Z09-066
Rendell, L., Cantor, M., Gero, S., Whitehead, H., and Mann, J. (2019). Causes and consequences of female centrality in cetacean societies. Philos. Trans. R. Soc. B. Biol. Sci. 374:20180066. doi: 10.1098/rstb.2018.0066
Rendell, L., Whitehead, H., and Coakes, A. (2005). Do breeding male sperm whales show preferences among vocal clans of females. Mar Mamm. Sci. 21, 317–322. doi: 10.1111/j.1748-7692.2005.tb01231.x
Rice, D. W. (1989). “Sperm whales physeter macrocephalus Linnaeus, 1758,” in Handbook of Marine Mammals, Vol. 4, eds S. H. Ridgeway and R. Harrison (London: Academic Press), 177–233.
Richard, G., Bonnel, J., Tixier, P., Arnould, J. P. Y., Janc, A., and Guinet, C. (2020). Evidence of deep-sea interactions between toothed whales and longlines. Ambio 49, 173–186. doi: 10.1007/s13280-019-01182-1
Richard, G., Titova, O. V., Fedutin, I. D., Steel, D., Meschersky, I. G., Hautin, M., et al. (2018). Cultural transmission of fine-scale fidelity to feeding sites may shape humpback whale genetic diversity in Russian Pacific waters. J. Hered. 109, 724–734. doi: 10.1093/jhered/esy033
Richard, K. R., McKarrey, S., and Wright, J. M. (1994). DNA sequence from the SRY gene of sperm whale (Physeter macrocephalus) for use in molecular sexing. Can. J. Zool. 72, 873–877. doi: 10.1139/z94-118
Roche, C., and Guinet, C. (2007). Marine mammals and demersal longline fishery interactions in Crozet and Kerguelen exclusive economic zones: an assessment of depredation levels. CCALMR Sci. 14, 67–82.
Rødland, E. S., and Bjørge, A. (2015). Residency and abundance of sperm whales (Physeter macrocephalus) in the Bleik Canyon, Norway. Mar. Biol. Res. 11, 974–982. doi: 10.1080/17451000.2015.1031800
Sarano, F., Girardet, J., Sarano, V., Vitry, H., Preud’homme, A., Heuzey, R., et al. (2021). Kin relationships in cultural species of the marine realm: case study of a matrilineal social group of sperm whales off Mauritius island, Indian ocean. R. Soc. Open Sci. 8:201794. doi: 10.1098/rsos.201794
Sarano, V., Sarano, F., Giradet, J., Preudhomme, A., Vitry, H., Heuzey, R., et al. (2022). Underwater Photo-Identification of Sperm Whales (Physeter macrocephalus) Off Mauritius Marine Biology Research. (in Press). doi: 10.1080/17451000.2022.2040737
Schakner, Z. A., Lunsford, C., Straley, J., Eguchi, T., and Mesnick, S. L. (2014). Using models of social transmission to examine the spread of longline depredation behavior among sperm whales in the Gulf of Alaska. PLoS One 9:e109079. doi: 10.1371/journal.pone.0109079
Shine, R. (1989). Ecological causes for the evolution of sexual dimorphism: a review of the evidence. Q. Rev. Biol. 64, 419–461. doi: 10.1086/416458
Smolker, R., Richards, A., Connors, R., and Pepper, J. (1992). Sex differences in patterns of association among Indian ocean bottlenose dolphins. Behaviour 123, 38–69. doi: 10.1163/156853992X00101
Steiner, L., Lamoni, L., Plata, M. A., Jensen, S.-K., Letteval, E., and Gordon, J. (2012). A link between male sperm whales, Physeter macrocephalus, of the Azores and Norway. J. Mar. Biol. Assoc. U. K. 92, 1751–1756. doi: 10.1017/S0025315412000793
Teloni, V., Mark, J. P., Patrick, M. J. O., and Peter, M. T. (2008). Shallow food for deep divers: dynamic foraging behavior of male sperm whales in a high latitude habitat. J. Exp. Mar. Biol. Ecol. 354, 119–131. doi: 10.1016/j.jembe.2007.10.010
Tixier, P., Gasco, N., Duhamel, G., Viviant, M., Authier, M., and Guinet, C. (2010). Interactions of patagonian toothfish fisheries with killer and sperm whales In the Crozet islands exclusive economic zone: an assessment of depredation levels and insights on possible mitigation strategies. CCAMLR Sci. 17, 179–195.
Tixier, P., Giménez, J., Reisinger, R., Méndez-Fernandez, P., Arnould, J., Cherel, Y., et al. (2019). Importance of toothfish in the diet of generalist subantarctic killer whales: implications for fisheries interactions. Mar. Ecol. Prog. Ser. 613, 197–210. doi: 10.3354/meps12894
Van der Linde, M. L., and Eriksson, I. K. (2020). An assessment of sperm whale occurrence and social structure off São Miguel Island, Azores using fluke and dorsal identification photographs. Mar. Mamm. Sci. 36, 47–65. doi: 10.1111/mms.12617
Wang, J. (2002). An estimator for pairwise relatedness using molecular markers. Genetics 160, 1203–1215. doi: 10.1093/genetics/160.3.1203
Whitehead, H. (2002). Estimates of the current global population size and historical trajectory for sperm whales. Mar. Ecol. Prog. Ser. 242, 295–304. doi: 10.3354/meps242295
Whitehead, H., Coakes, A., Jaquet, N., and Lusseau, S. (2008). Movements of sperm whales in the tropical Pacific. Mar. Ecol. Prog. Ser. 361, 291–300. doi: 10.3354/meps07412
Keywords: marine megafauna, genetic diversity, cultural species, sperm whales, Indian Ocean, male-social fidelity, kin relationships
Citation: Girardet J, Sarano F, Richard G, Tixier P, Guinet C, Alexander A, Sarano V, Vitry H, Preud’homme A, Heuzey R, Garcia-Cegarra AM, Adam O, Madon B and Jung J-L (2022) Long Distance Runners in the Marine Realm: New Insights Into Genetic Diversity, Kin Relationships and Social Fidelity of Indian Ocean Male Sperm Whales. Front. Mar. Sci. 9:815684. doi: 10.3389/fmars.2022.815684
Received: 15 November 2021; Accepted: 15 February 2022;
Published: 16 March 2022.
Edited by:
Lars Bejder, University of Hawai’i at Mānoa, United StatesReviewed by:
Rob Harcourt, Macquarie University, AustraliaCopyright © 2022 Girardet, Sarano, Richard, Tixier, Guinet, Alexander, Sarano, Vitry, Preud’homme, Heuzey, Garcia-Cegarra, Adam, Madon and Jung. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jean-Luc Jung, amVhbi1sdWMuanVuZ0BtbmhuLmZy, anVuZy5qZWFubHVjQGdtYWlsLmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.