 Manuel Ruiz-Villarreal1*
Manuel Ruiz-Villarreal1* Marc Sourisseau2
Marc Sourisseau2 Phil Anderson3
Phil Anderson3 Caroline Cusack4
Caroline Cusack4 Patricia Neira5
Patricia Neira5 Joe Silke5
Joe Silke5 Francisco Rodriguez6
Francisco Rodriguez6 Begoña Ben-Gigirey6†
Begoña Ben-Gigirey6† Callum Whyte3Solene Giraudeau-Potel3
Callum Whyte3Solene Giraudeau-Potel3 Loic Quemener7
Loic Quemener7 Gregg Arthur8
Gregg Arthur8 Keith Davidson3
Keith Davidson3- 1Centro Oceanográfico de A Coruña, Instituto Español de Oceanografía (IEO, CSIC), A Coruña, Spain
- 2Ifremer, French Research Institute for Exploitation of the Sea, DYNECO PELAGOS, Plouzané, France
- 3Scottish Association for Marine Science (SAMS), Scottish Marine Institute, Oban, United Kingdom
- 4Ocean, Climate and Information Services, Marine Institute, Galway, Ireland
- 5Marine Environment and Food Safety Services, Marine Institute, Galway, Ireland
- 6Centro Oceanográfico de Vigo, Instituto Español de Oceanografía (IEO, CSIC), Vigo, Spain
- 7Ifremer, French Research Institute for Exploitation of the Sea, REM/RDT/DCM, Plouzané, France
- 8Shetland UHI, Shetland, United Kingdom
Harmful algal blooms (HABs) cause harm to human health or hinder sustainable use of the marine environment in Blue Economy sectors. HABs are temporally and spatially variable and hence their mitigation is closely linked to effective early warning. The European Union (EU) Interreg Atlantic Area project “PRIMROSE”, Predicting Risk and Impact of Harmful Events on the Aquaculture Sector, was focused on the joint development of HAB early warning systems in different regions along the European Atlantic Area. Advancement of the existing HAB forecasting systems requires development of forecasting tools, improvements in data flow and processing, but also additional data inputs to assess the distribution of HAB species, especially in areas away from national monitoring stations, usually located near aquaculture sites. In this contribution, we review different novel technologies for acquiring HAB data and report on the experience gained in several novel local data collection exercises performed during the project. Demonstrations include the deployment of autonomous imaging flow cytometry (IFC) sensors near two aquaculture areas: a mooring in the Daoulas estuary in the Bay of Brest and pumping from a bay in the Shetland Islands to an inland IFC; and several drone deployments, both of Unmanned Aerial Vehicles (UAV) and of Autonomous Surface vehicles (ASVs). Additionally, we have reviewed sampling approaches potentially relevant for HAB early warning including protocols for opportunistic water sampling by coastguard agencies. Experiences in the determination of marine biotoxins in non-traditional vectors and how they could complement standard routine HAB monitoring are also considered.
1 Introduction
Harmful algal blooms (HABs) are increases in the density of certain phytoplankton species that cause harm to human health or hinder sustainable use of the marine environment in blue economy sectors (Wenhai et al., 2019). Some HAB species generate “shellfish poisoning syndromes’’ in humans, usually following the consumption of shellfish that have ingested the harmful cells and concentrated the toxin within their flesh (Berdalet et al., 2016). Other HABs can impact the health of farmed fish with significant economic consequences (Davidson et al., 2020). A variety of protists and prokaryotic photosynthetic organisms may be responsible for HAB episodes (Hallegraeff, 2004), including toxic and non-toxic species mainly from dinoflagellates, haptophytes, raphidophyceae, diatoms, pelagophyceans and cyanobacteria, among others, as detailed in the IOC-UNESCO taxonomic reference list of harmful microalgae (Lundholm et al., 2009). Their populations thrive under diverse environmental conditions and display different characteristics (e.g. morphological, chemotaxonomical and genetic), which enables their detection by multiple means depending on the applied technique (Stauffer et al., 2019).
HABs are temporally and spatially variable and hence their mitigation is closely linked to effective early warning. This is primarily achieved by a network of sampling sites located in the vicinity of aquaculture operations. According to the European Union (EU) Regulation No 853/2004 (European Parliament and Council, 2004) and its amendments, typically samples (water and shellfish) are collected on a weekly basis. Water samples are analysed by light microscopy to identify harmful organisms. Shellfish samples are investigated for marine biotoxins by using different instrumental techniques depending on the target analytes. In terms of shellfish safety, should concentration of HABs or their biotoxins in shellfish flesh exceed regulatory thresholds then harvesting restrictions are applied until the biotoxins depurate and the shellfish are safe for human consumption. The spatial and temporal changes of HAB species in the region are reviewed and discussed extensively elsewhere in the literature, for example Trainer et al. (2010), Belin et al. (2021), Bresnan et al. (2021), Fernandes-Salvador et al. (2021), this issue, or Gianella et al. (2021), this issue. In the case of shellfish biotoxin producing species, some monitoring networks consider alert threshold densities to trigger toxin monitoring based on historical data, like e.g. 100 cells l⁻¹ for Dinophysis in Scotland (Swan et al., 2018) or if present whatever the Dinophysis species in France (Belin et al., 2021), while higher threshold densities (1,000-300,000 cells l⁻¹) are defined also in the case of the REPHY network for Alexandrium, Protoceratium, Pseudo-nitzschia and others. Measures to take management actions to minimize damage on farmed fish are possible if ichthyotoxic HAB densities rise significantly, though no threshold densities are established for fish killing HABs (Davidson et al., 2021). In both cases early warning of developing HABs is key to effective mitigation (e.g. Maguire et al., 2016).
High biomass blooms can be carried by advection from offshore areas to the coast. For these near surface bloom events, satellite detection is possible with algorithms developed to discriminate some HAB species from benign phytoplankton (Stumpf et al., 2009; Kurekin et al., 2014; Jordan et al., 2021). Such HAB detection approaches are particularly powerful if coupled to a numerical model that can predict the likely trajectory of the bloom over the following days and hence provide an early warning for at risk aquaculture sites (Davidson et al., 2016; Maguire et al., 2016). Low biomass blooms like those of the shellfish toxin producing dinoflagellates, in the genera Dinophysis, are difficult to monitor since they often present as subsurface blooms that are undetectable in satellite imagery. In addition, they are harmful at low cell densities (<103 cells l⁻¹), which comprise a small proportion of the full microphytoplankton community, and are therefore difficult to detect with conventional monitoring sampling methods (Escalera et al., 2012). Moreover, the frequency of conventional weekly monitoring, usually sampling inshore sites, is insufficient to raise an alert should wind reversals rapidly advect dense shelf populations into aquaculture sites within a matter of days (Escalera et al., 2010; Raine et al., 2010; Whyte et al., 2014). Anyway, hydrodynamical model runs combined with Lagrangian particle tracking simulations can give information on HAB transport along-shore or cross-shore in and out of harvesting areas (Maguire et al., 2016).
Today, the main limitation to the enhancement of HAB warning systems is the availability of high frequency, real or near-real-time, HAB data and relevant parameters in their ecosystem. These data are essential to constrain initiation, movement and growth of the blooms in numerical forecast models, to validate/confirm satellite detections and add value to the human interpretation of how the ecosystem is evolving. Future more sophisticated modelling systems will benefit from “nudging” as new observational in-situ data streams are available to enhance model estimations, a technique known as data assimilation forecasting. The European Union Interreg Atlantic Area project PRIMROSE, Predicting Risk and Impact of Harmful Events on the Aquaculture Sector1, involved partners from the EU Atlantic Area (Spain, Portugal, France, Ireland and the UK) in an effort to improve early warning systems in the partners’ regions building on improved data management and flow from existing HAB and biotoxin monitoring programmes and on coordination and development of HAB sampling and modelling systems (Mateus et al., 2019; Fernandes-Salvador et al., 2021). During PRIMROSE, which ran from 2018 to 2021, we reviewed and demonstrated novel methodologies for HAB sampling through the following activities: 1) deployment of autonomous imaging flow cytometry sensors on moorings near two aquaculture areas: the Daoulas Estuary in French Brittany and the Scottish Shetland Islands to demonstrate a dataflow pipeline, data processing and data sharing, 2) deployment of a suite of cheap, near-shore, autonomous surface vehicles capable of collecting water samples for shore-based assays, 3) evaluating protocols for opportunistic water sampling by coastguard agencies and 4) reviewing the performance of other opportunistic sampling approaches and how they augment standard routine HAB monitoring.
2 Autonomous Flow Cytometer Sensors
Autonomous flow cytometers are a useful tool with fast sample throughput by comparison to traditional optical microscopy. The latter is the reference methodology for most national HAB monitoring programmes (Karlson et al., 2010) but requires time for transport of water samples, for sample preparation (up to 24 hours) and for taxonomic specialists to analyze and report the results (First and Drake, 2012) as well as correct sample preservation methods to prevent changes of morphology for some species during transport. This limits the sample processing throughput rate (Dunker, 2019) and the coupling of these data with information coming from other sensors in a HAB warning system. To address this problem a range of research tools have been developed, with some now being sufficiently mature and available on the market for application within automated mooring systems.
A major development was the emergence of Imaging Flow Cytometry (IFC) (Dashkova et al., 2017), an enhancement of a previous in-flow cytometric system that uses fluorescence and optical cell scatter measurements to discriminate planktonic cells, typically at the “functional group” level (Thyssen et al., 2008). The incorporation of imaging technology along with machine learning identification and classification of the resulting image data library has markedly advanced phytoplankton discrimination and enumeration capabilities in the field. Results from this application are promising (Campbell et al., 2010; Campbell et al., 2013; Dunker et al., 2018). While an IFC is still relatively expensive, cost reductions through product development are making this technology more widely accessible to users, with a recent increase in the number of deployments in a wide range of sensitive marine areas including the east and west coasts of the US (Fischer et al., 2020), a number of locations in Scandinavia (Kraft et al., 2021) and Hong Kong (Guo et al., 2021). Like all instruments, autonomous IFCs have significant cost and time constraints related to training, servicing, data management and subsequent data processing. At present, real-time analysis requires a physical link to land (cables) or a wide bandwidth network connection and access to significant data processing capability. Continuous sampling may require two or three autonomous flow cytometer units, with one deployed while the other units remain in the laboratory undergoing maintenance, calibration, service and training.
During PRIMROSE, the use of autonomous IFCs for HAB assessments have been tested and demonstrated in two sites: one IFC installed on a mooring at the Daoulas Estuary in the Bay of Brest, France and one IFC land based with water being continuously pumped from an adjacent bay in the Scottish Shetland islands.
2.1 Autonomous Flow Cytometer Sensor Installed in a Mooring
A platform with a frame to support a Cytosense flow cytometer (Cytobuoy product) was moored during PRIMROSE (Figure 1) in the Daoulas estuary in the Bay of Brest, an area that experiences regular Alexandrium minutum bloom events since 2012 (Chapelle et al., 2015). Water depth at the mooring site was more than three metres at spring-tide low water to avoid sea-bed contact (station C, Figure 2). The two time periods in 2019 investigated during PRIMROSE were 28th June to 10th July and 14th to 26th August 2019. The site position was 200 m from another established monitoring site (project Alex Breizh) with a mooring measuring temperature, fluorescence, salinity and turbidity (station D) where once per week samples for plankton community characterization are obtained 2 hours around high water (HW) using a kayak or a zodiac. A third station from the national monitoring program REPHY where weekly microscope based phytoplankton enumeration are obtained is located in the Bay of Brest (station B) at a distance of 2 km from the IFC mooring. REPHY samples are also taken during high water and data are available from 2016 (REPHY, 2021). To be in agreement with the microscopic dataset obtained in the existing monitoring in the Bay of Brest and due to the macrotidal characteristic of the Daoulas estuary, the IFC water sampling was also set to occur once during the day (during high water). Our data can also be compared to more oceanic waters analyzed at the entrance of the Bay of Brest (Station A), where a long term environmental monitoring station within the SOMLIT (Service d’Observation en Milieu Littoral) is established2. The environmental variables acquired since 1998 and the availability of phytoplankton samples taken weekly at high tide from 2009 allow the comparison between incoming waters and the Daoulas estuarine waters.

Figure 1 Picture of the deployment of the PRIMROSE mooring platform at Anse du Roz in the Daoulas estuary (A) and scheme (B) showing details of the floating structure frame designed for carrying the autonomous flow cytometer, IFC (C), its associated tools (batteries, etc) and other additional sensors.
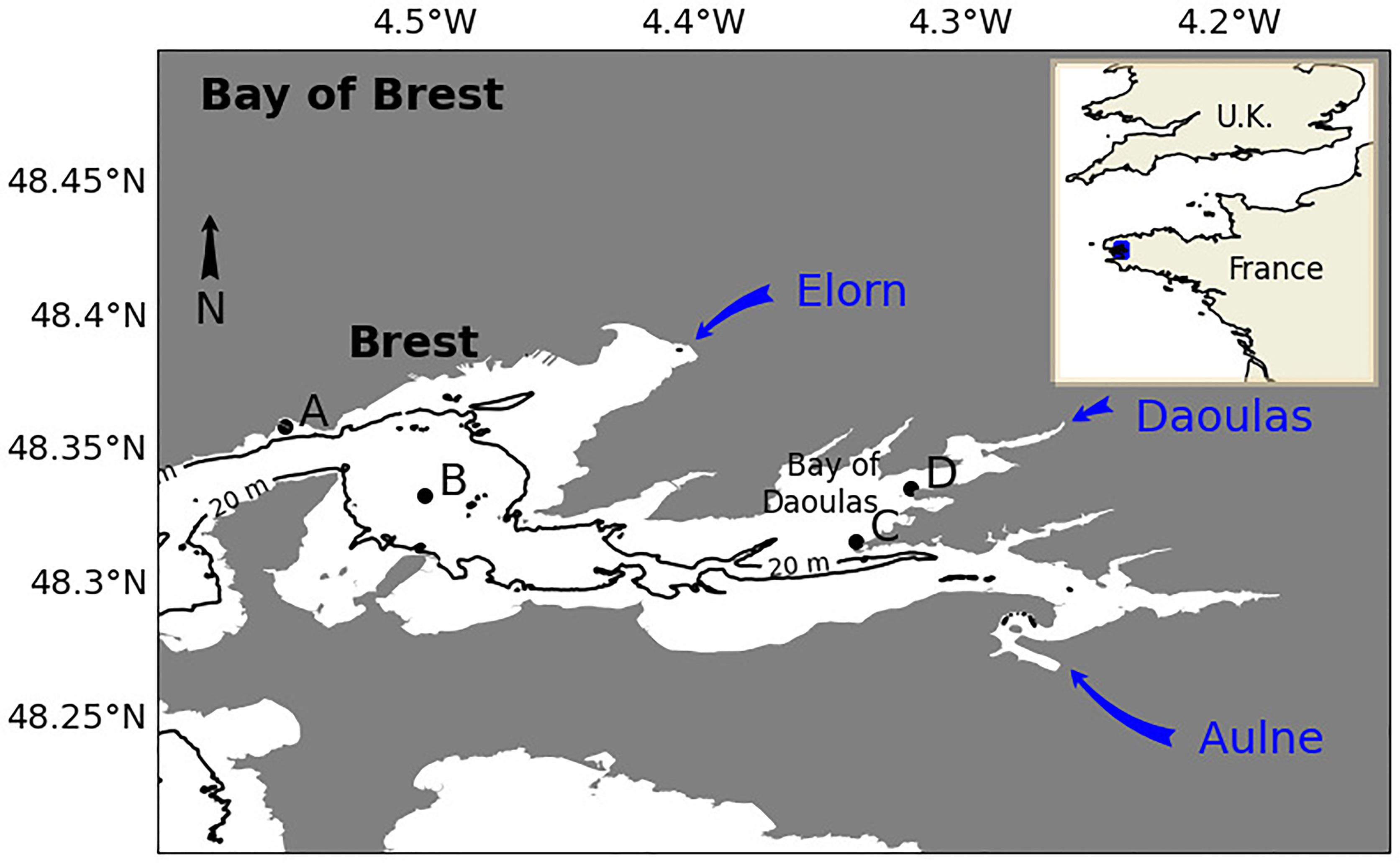
Figure 2 Positions of the monitoring stations in the Bay of Brest: (Station A): Ste Anne du Portzic station from SOMLIT program (Observatory in the coastal environment, http://www.somlit.fr); (Station B): Lanveoc from REPHY national program; (Station C): PRIMROSE IFC mooring at Anse du Roz in the Daoulas estuary; (Station D): Alex Breizh mooring and sampling site at Pointe du Château in the Daoulas estuary.
The ability to classify cells through the identification of collected images and/or the clustering or gating of fluorescence or cell scatter signals is the essential step in the analysis of IFC data. To permit comparisons with other data sets, the clusters were made in agreement with the protocol from other monitoring stations along the French coast (SOMLIT). Based on the size proxy (FSS, Forward Size Scatter) and the red/orange fluorescence ratio, five large clusters were made: Synechococcus-like, Picoeukaryotes, Nanophytoplankton, Cryptophyceae-like and Microphytoplankton. Cluster names, vocabulary and file format agree with the best practice followed in well-established European projects like SeadataCloud and Jerico Next (Artigas et al., 2019) to permit comparison and a larger diffusion of the data set. In this way, the IFC setup and data quality were evaluated in 2018 by comparing estimated abundances from fixed water samples with densities provided by two different flow cytometry platforms (Roscoff laboratory and Brest University). It should be noted that typical flow cytometry platforms do not provide estimates of the microphytoplankton cluster due to the size of their tubes and the volumes analyzed. The comparison was thus limited to the four smallest clusters. The first-round showed that the IFC set-up produced highly correlated abundances for the same clusters. A further quality control was undertaken in 2019 by comparing in-situ time series of autonomous sampling (counting of living cells without calibration beads) with cell densities estimated from chemically fixed samples (with calibration beads) that were manually collected at the same station and time period. Calibration beads were also used to check fluorescence stability and IFC alignment was checked before and after each deployment. The data of both sampling strategies were highly correlated (>0.99) and the data set from our in-situ IFC sampling was deemed sufficiently consistent to be used as a reliable monitoring tool.
The first output from our IFC data analysis was to highlight the strong spatial and temporal variability of the phytoplankton community in the Daoulas Estuary (Figures 3A, B). For such an area, the daily resolution that IFC provides is required to describe the trends of the evolution of cell density whereas the weekly frequency of existing monitoring is clearly inappropriate to make short term forecasts like those required in HAB warning systems. For example, local microphytoplankton blooms occurred at the Daoulas Estuary for 4 to 5 days after some high river runoff events (e.g. in the period 27th June to 1st July, Figure 3A). In the same way, an upstream movement of the maximum phytoplankton concentration occurred during nutrient-limited periods in the middle of the summer; note that in general, due to the estuarine dynamics of the macrotidal Daoulas estuary, HW sampling was associated with low Chl-a concentrations (Figure 3B). This displacement is illustrated by low cell densities at HW from 17th to 27th August 2019 (Figure 3B). The sets of daily microphytoplankton images produced during the IFC measurements are highly relevant to characterize the variability of the phytoplankton community and eventually to provide alerts of the presence of HAB species. An illustration is provided in Figure 4 where IFC images from two days (29th June and 9th July) in the fortnight shown in Figure 3A are plotted. Figure 4A reveals that the microplankton peak coinciding with the chlorophyll maximum on 29th June is composed of chain diatoms (Chaetoceros spp.). After this peak, microplankton concentration is observed to decrease quickly in parallel to salinity increase. Picoplankton, which was high at the beginning of the period, decreases during the microplankton peak and attains a relative maximum on 7th July, when salinities are again higher. The 9th July image (Figure 4B) shows that plankton community is then composed of solitary dinoflagellate species, including Alexandrium spp. and Ceratium spp., which were also detected in the weekly sampling of the Alex Breizh project at Point du Château (station D).
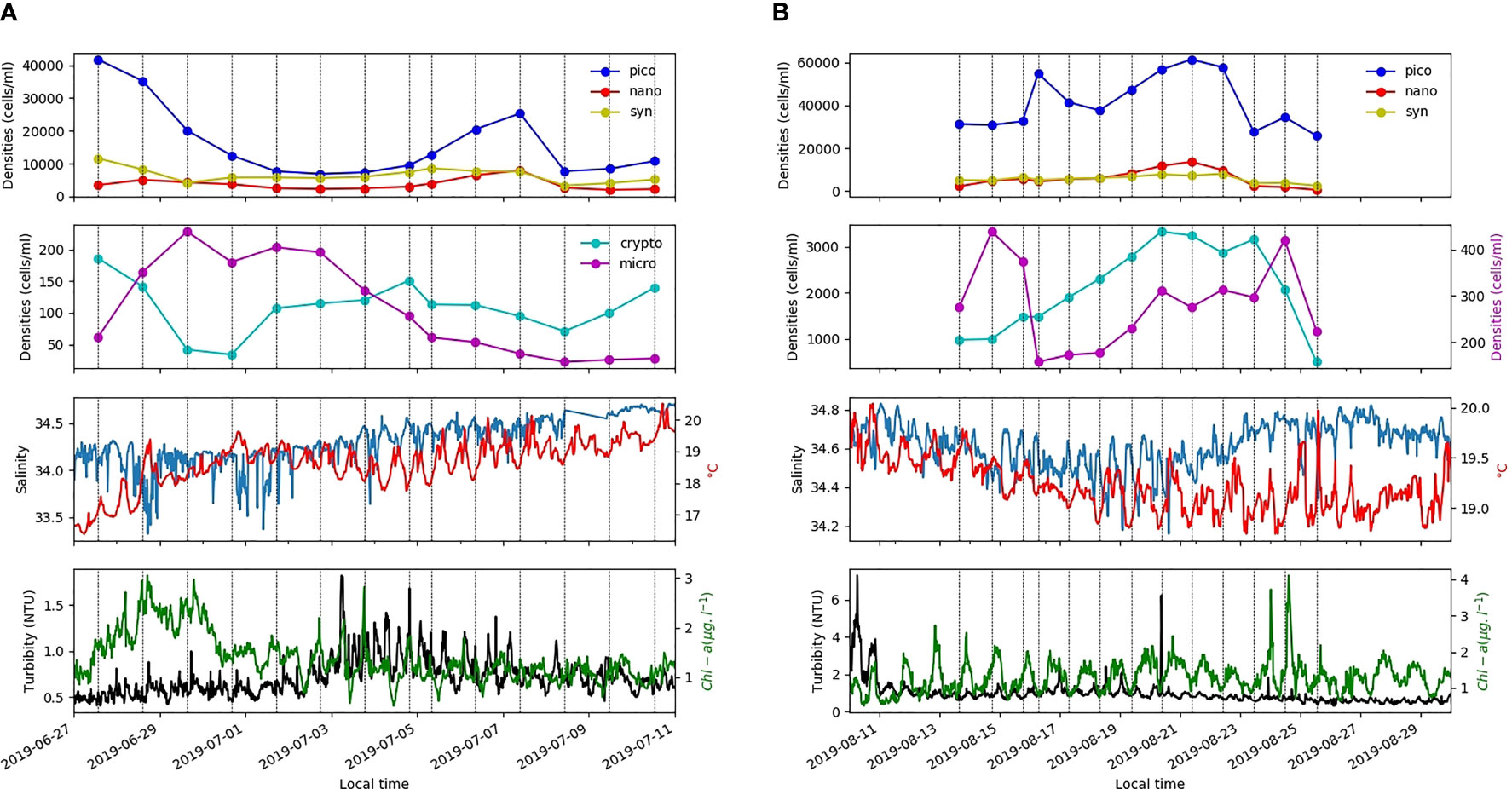
Figure 3 Time series of variables obtained from the IFC and other sensors on the autonomous mooring in the Daoulas estuary (station C) from 28th June to 10th July 2019 (A) and from 13th August to 25th August 2019 (B). Abundances of five regular clusters (Synechococcus, Picoeukaryotes, Nanophytoplankton, Microphytoplankton and Cryptophyceae) obtained from the IFC are plotted. IFC sampling was done at high water during the day (dotted vertical lines).

Figure 4 Microphytoplankton images recorded during daylight high water at the IFC moored in the Daoulas estuary on 29th June (A) and on 9th July 2019 (B).
The methodology for cluster processing of IFC data allowed the comparison of the plankton community at the Daoulas Estuary site (station C) with that at the entrance of the Bay of Brest measured in the weekly SOMLIT sampling (station A). The average seasonal evolution (Figure 5) shows that nanophytoplankton and Cryptophyceae densities are significantly higher in the Daoulas Estuary but that is not the case for picoeukaryotes and Synechococcus. Note that, as pointed out above, the microphytoplankton cluster was not computed at SOMLIT stations. Despite the high tidal mixing and the small residence time of water masses, separate monitoring stations are required to describe local growth and/or accumulation. Therefore, with the IFC mooring data set, we were able to demonstrate the significant difference in the phytoplankton community composition. This observation is based on a global approach to functional groups that remains limited to the macro scale description of the phytoplankton community and cannot yet be directly related to variability of HABs. The nanoflagellate cluster however includes the main part of the Alexandrium spp. populations due to the size limit between nano- and micro-phytoplankton clusters. The threshold was set to 20 µm whereas Alexandrium cell size can vary from 5 to 20 µm. Not all toxic species are included in one cluster (i.e. microphytoplankton).
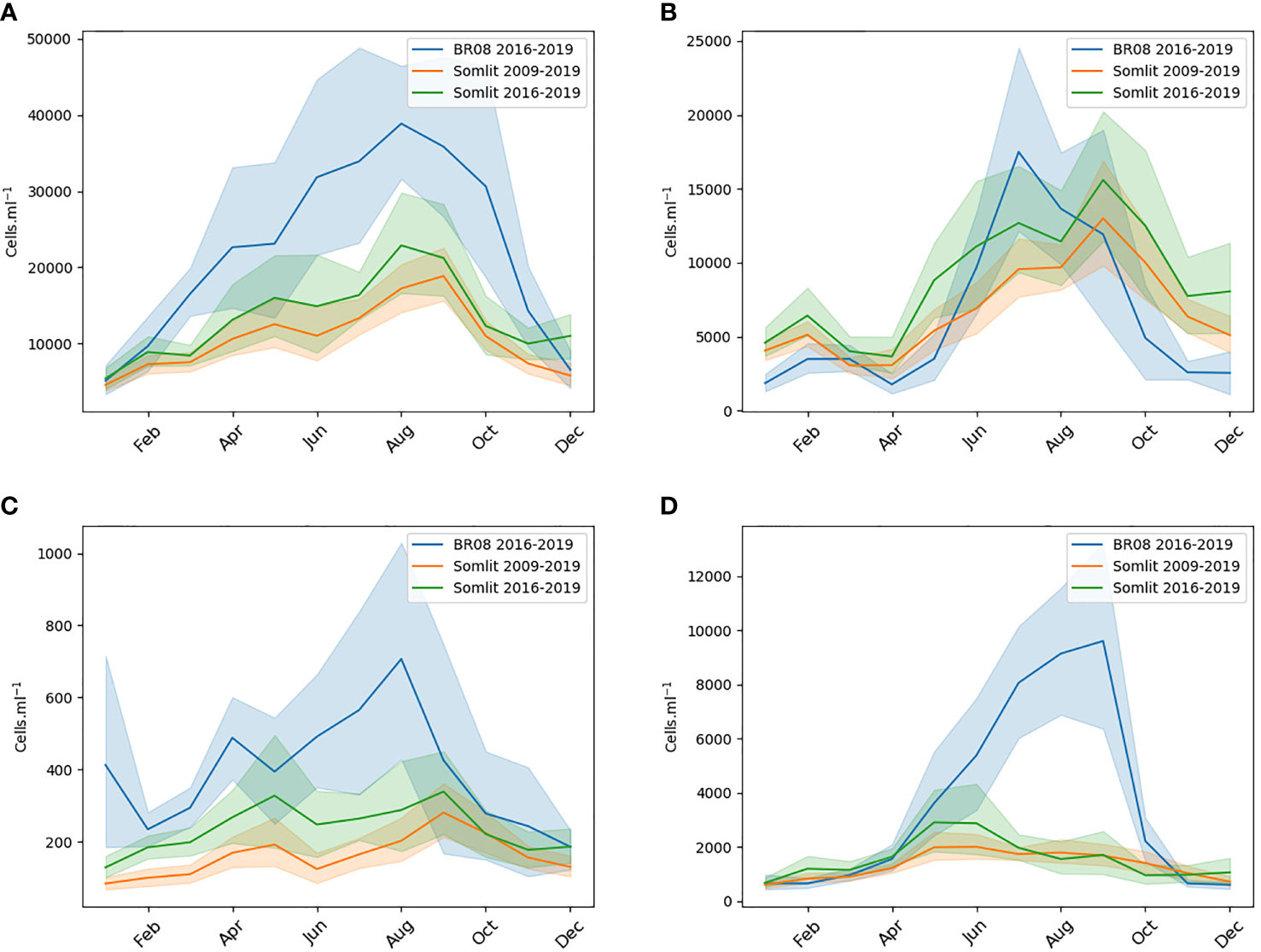
Figure 5 Seasonal patterns of monthly average and standard deviation) observed by the IFC in the Daoulas Estuary (station D, Figure 2, blue line) during 2016-2019 for the Picoeukaryotes, Synechococcus, Cryptophyceae and Nanophytoplankton clusters (A–D). The seasonal pattern at the entrance of the Bay was estimated from the SOMLIT-Brest station (station A) for the same and a longer period (green and red line respectively).
With the settings used during these deployments (the speed of the water, analysis duration, minimal Equivalent Spherical diameter, etc.) and the in-situ cell densities, the number of recorded images were limited to between 100 to 300 per sample in 2019. This prevented a quantitative estimate of several image clusters, but the information gathered was still useful to support identification of the predominant species, as illustrated in Figure 4. Some modifications of the IFC hardware and setting were made to increase the number of images, and it is expected that a quantitative estimate can be achieved. New classifiers for automatic clustering of HAB species were constructed based on specific learning sets of > 1000 images obtained from cultivated strains of two toxic lineages (Alexandrium minutum and Pseudo-nitzschia spp.). Data has been sent in real time by 4G to an internal server since 2020, but the full automatic processing for dissemination is not yet operational.
2.2 Autonomous Flow Cytometer Sensor Installed on Land
Another deployment of an IFC was made in the Scottish Shetland islands in the form of a FlowCytoBot (IFCB). IFCBs have been used previously for HAB monitoring, for example in Texas, where harmful blooms of the shellfish biotoxin producing dinoflagellate Dinophysis were revealed (Campbell et al., 2010). The Shetland Islands are an important centre for Scottish aquaculture, with HAB monitoring by microscopy occuring at a number of aquaculture sites. The relative expense of these instruments prevents their wide deployment, hence, our IFCB has been located at a sentinel location (Scalloway) to provide high temporal frequency regional risk assessment. While not a shellfish harvesting site, the location coincides with the Marine Scotland Science Scottish Coastal Observatory monitoring site that provides parallel environmental information to aid in the understanding of HAB dynamics. In contrast to the French deployment the Scottish instrument is land based, with water being continuously pumped from the adjacent bay (Figure 6). This provides additional monitoring flexibility with the potential for hand collected samples from other aquaculture sites to be manually analyzed. HAB data from Scotland are made publicly available on the web in real time [via www.HABreports.org, (Davidson et al., 2021), this issue]. IFCB images can be displayed in near-real time or in delayed mode in the HABreports web page (Figure 7A). The 31th August 2021 IFC image shows the presence of Pseudo-nitzschia spp. The HABreports website displayed alert levels for week 35 (Aug 30 - Sep 05) and previous weeks at the North Flotta (Weisdale Voe) monitoring site (Figure 7B), indicating an “amber” alert caused by the presence of Pseudo-nitzschia, although Amnesic Shellfish Poisoning (ASP) toxin concentration was below toxic levels. The presence of Alexandrium (rather abundant) and Dinophysis (scarce) was also detected in other IFC images for the same day (not shown). The HABreport bulletin therefore generated an alert that week due to the presence of these harmful taxa. In combination with Okadaic acid (OA)/Dinophysis Toxins (DTXs)/Pectenotoxins (PTXs) that were over regulatory limits an overall “red” alert was generated. There were also a few cells of Karenia mikimotoi in the IFC images although they were not detected in microscope monitoring.

Figure 6 FlowCytoBot (IFCB) deployed in Scalloway in the Shetland Islands during PRIMROSE. The IFCB is land based with seawater being continuously pumped from an adjacent bay.
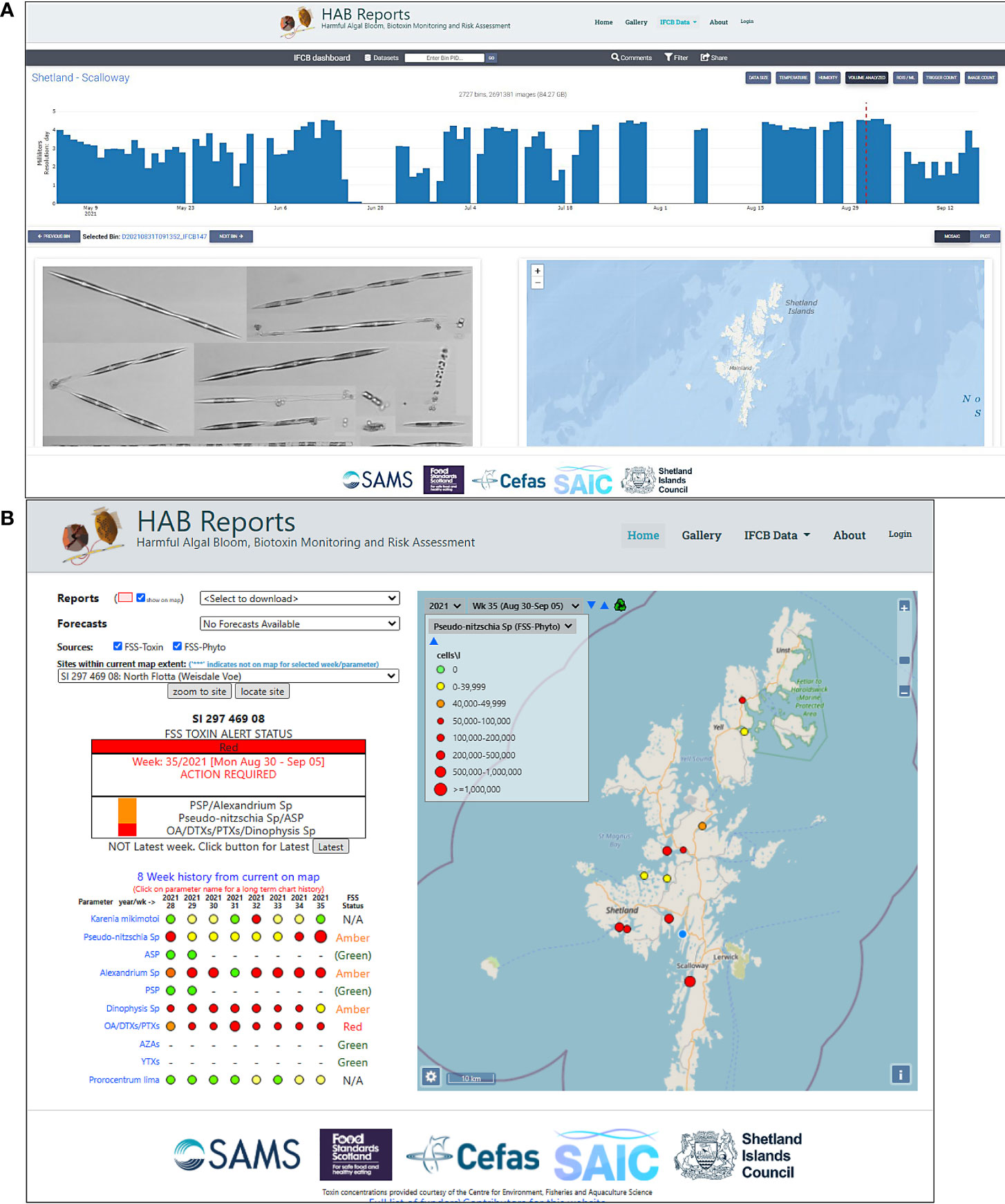
Figure 7 HABreport portal (https://www.habreports.org/) snapshots: (A) interface for viewing and interrogating IFC data from Scalloway, showing the 31st August 2021 IFC image of largest phytoplankton (mainly Pseudo-nitzschia spp.) and (B) HABreports display (see Davidson et al., 2021) showing alert levels based on HAB species concentrations and toxin analysis for the current (week 35, Aug 30 - Sep 05) and previous weeks at the North Flotta HAB monitoring site (blue circle). Green/amber/red are Food Standards Scotland (FSS) “Toxin Traffic Light Guidance” levels that indicate the harvesting action and testing considerations that should follow (N/A, no action).
Similar to the French IFC, operation of the Scottish instrument for plankton community characterisation, and therefore for use in HAB early warning, requires reliable image classification. Typically an IFCB image classifier is built using identifiable characteristics of training images. These characteristics, usually based on shape or texture descriptors, are then input to classifiers like Random Forest Algorithms (RFAs), or Supported Vector Machines (SVMs). Initial operation with a RFA classifier has proved promising in Shetland. However, Deep learning and Convolutional Neural Networks (CNNs) in particular have shown superior performance for image classification tasks (Sharma et al., 2018) and classifiers are currently being developed using these approaches.
3 Robotic Platforms
Fully autonomous vehicles as either instrument platforms or as sample collection platforms for pure science research are becoming commonplace but are still far from a mature technology. Science is willing to take the risk of equipment failing to meet expectations given the returns-benefit of novel and unprecedented data. Forecasting for commerce requires a higher level of Technology Readiness Level (TRL) to give continual and dependable data for the end user to make reliable planning decisions; at present, mobile and static marine robotics are making little impact on aquaculture commerce in general, and HAB detection and analysis in particular. That said, the wish to automate, reduce cost and reduce risk is driving rapid progress in this arena.
We present here an overview of existing technologies that are commercially set to make a significant impact on HAB forecasting, which fall into two broad categories: airborne and surface autonomous vehicles. We review:
● Water landable quadcopter Remotely Piloted Aircraft System (RPAS) for very local sampling
● 100 km range Beyond Visual Line Of Sight (BVLOS) copter-wing RPAS
● 2 m scale Autonomous Surface Vehicle (ASV) for in-shore sampling and ocean color detection
● 300 nautical miles autonomy USV-Mar II ASV for automatic oceanographic sampling in the Galician rias
3.1 Remotely Piloted Aircraft Systems (RPAS) or ‘Drones’
Commercial airborne robotic platforms are becoming commonplace for terrestrial work, predominantly for photography or video capture supporting different applications in an increasing variety of fields. Their use is curtailed to an extent due to the high consequence hazards involved with any aircraft in civil airspace. The term for robotic aircraft preferred by all national aviation authorities is “Remotely Piloted Aircraft System” to reflect that the legal responsibility of the ‘operator’ is identical to that of a pilot in a normal aircraft. The platform, even if fully robotic, must be capable of returning control to the operator at all times, and they must be capable of piloting the aircraft. Most commercial ventures have focused on operating within “Visual Line Of Sight” (VLOS) which legally3 limits the craft to a radius of 500 m from the operator. VLOS limitation has a technical consequence, in that most VLOS operations use multi-copter style RPAS, that is the archetypal four-propeller ‘drone’. The extreme flight inefficiency of this design coupled to the poor energy density of ca. 2020 batteries means flights are of the order of 10 - 20 minutes. The simplicity of operation and lack of need for a runway, coupled to the legal limit to range, however, make this the design of choice for the mass market.
A compromise design to get similar take off/landing simplicity, termed Vertical Take Off and Landing (VTOL) whilst still using efficient wings for lift, is now available. Both are considered below.
3.1.1 Water Landable Quadcopter RPAS
TetraDrone’s TD7 hexacopter platform (Figure 8) was trialled during PRIMROSE for local sampling in Scotland. This aircraft is designed to land upon and to sample the sea surface, and is capable of making airborne measurements of variables such as temperature, humidity and CO2.

Figure 8 TD7 water-landable UAV under trial on a near calm and sheltered embayment at Dunstaffnage Peninsula, Argyll, Scotland.
The TD7 floats act to an extent as a safety net: when operating over water the craft will not sink irretrievably if forced to land. The aircraft is flown within VLOS either from shore or small boats but water-landing capability is limited to fair weather surface conditions. Initial plans to modify the wet payload bay (visible under the aircraft fuselage) for HAB sampling was postponed following results of studies to assess the capability of RPAS-based measurement of ocean color, itself used within the PRIMROSE project to identify HAB signature. An intention was to combine airborne visible spectral data with land-and-sample; however, the airborne ability to detect HAB is limited due to the effects of sunlight ‘glint’ (Weeks, 2019). Further, the limited range and dependency on placid sea-state have put further focus on ASV (see below) as a solution to near-shore detection and sampling of HABs.
3.1.2 Vertical Take-Off and Landing (VTOL) RPAS Operating Beyond Visual Line Of Sight (BLVOS)
The standard commercial quadcopter style of ‘drone’ used in VLOS, with a legal operating radius of 500 m results in a flight time in the tens of minutes, reported in the previous section is quite unsuited to the scales of most marine applications. Fully robotic airborne platforms used as physical transport and delivery infrastructure were demonstrated in Argyll in 2020 and 2021, showing that integration of BVLOS RPAS in civilian airspace is legally possible and VTOL by fixed wing aircraft is technically possible. This UK first of a ‘proof of feasibility’ was conducted in Scotland by the London-based company, Skyports Ltd. Daily for two weeks in Argyll during the Covid-19 crisis medical supplies and test samples were transported, with circa six flights per day between the hospitals in Oban and on the island of Mull.
Full BVLOS, with permission from the UK Civil Aviation Authority (CAA), involves months of preparation and discussion, both with the relevant UK and local authorities, land-owners and other local interested parties such as other, piloted aircraft operators. This is the case even when operating in Argyll with a lower inherent risk compared to air-space above London, it is merely simpler and less complex in Argyll, and this was the main basis for Skyports initial interest in the area.
A more technical issue, but still essential, is the ability for VTOL. The familiar quad (or hexo or octo) -copter can, of course, take off and land vertically and do not need a runway. Such helicopter-style aircraft are far less efficient than those termed ‘fixed-winged’ and with electrical batteries much less energy-dense than fossil fuels, airtime for ‘copters are 15 -30 minutes, even without payload. For BVLOS to be viable, therefore, the aircraft must fly, in the main, like a fixed-wing. An important feature of the Skyports aircraft Wingcopter (Figure 9) is the four swivel-mounted propellers; these rotate with a vertical axis at take-off and landing but rotate to a more familiar horizontal axis for winged flight (the aft propellers cease turning and fold back). This greatly increased payload/flight time, whilst still operating from a small field or (in this case) a helipad.

Figure 9 The Skyports aircraft (Wingcopter VTOL winged aircraft) showing the swivel-mounted forward motors in take-off/landing mode. The under-slung payload ‘pod’ is visible. Reproduced with permission of Wingcopter.
3.2 Autonomous Surface Vehicles (ASV)
3.2.1 ImpYak: A Fully Autonomous Small Surface Vehicle
ImpYak is a prototype impeller driven kayak initially conceived as a student practical demonstrator for the SAMS Marine Robotics course (Figure 10). ImpYak uses the “ArduPilot’’ family of autopilot software, an open-source initiative initiated in 2016 with “ArduRover”, running on specialized Arduino-like microcontroller + sensor hardware. More specifically, ImpYak uses the ArduBoat branch of ArduPilot running on the PixHawk 4 platform. Using ArduPilot and relevant hardware such as PixHawk offers a rapid and economic route to developing autonomous platforms, and there are versions for rovers (land vehicles), fixed-wing aircraft, quad and multi copters and even submersibles such as AUVs and ROVs.

Figure 10 ImpYak ASV in loiter mode on Loch Linnhe in north east Scotland on 9th July 2021 (A), whilst working in conjunction with the FAAM aircraft (Facility for Airborne Atmospheric Measurements, https://www.faam.ac.uk) (B).
ImpYak comprises of:
● A standard river-surfing kayak, which is laterally stable, has no keel.
● Avionics (‘autopilot’) based on the Pixhawk 4 and sensor suite (GPS, compass and accelerometers)
● Off-the-shelf air-cooled motor controllers (Electronic Speed Controllers: ESC)
● A pair of 600W (=1.6 HP) underwater remotely operated vehicle (ROV) impellers
● Battery set (18V): LiPo or lead-acid
Two significant inherent benefits to this system design are, firstly, that all versions have similar ‘front ends’ with mission planning (again via free source software, such as Mission Planner and Q Ground Control), so operating the ImpYak is a simple transition if already familiar with ArduPilot aircraft. Secondly, the communication protocol between mission planning software and platform avionics is via MAVlink, a full-duplex communication protocol specifically designed for UAVs. MAVlink commands and system data are exchanged continually between platform and ground station, but can also be exchanged between vehicles. This already embedded protocol opens the capability of not just swarm behavior (such as multi-kayak fleets), but heterogeneous swarm control, such as an aircraft and boat acting in coordination, and sampling a vertical profile of the ocean atmospheric boundary layer.
Initial trials of these platforms show that they are capable of making pre-planned waypoint surveys or acting as stationary temporary surface mooring, which is a command mode called ‘loiter’. Instrument packages can be included either externally or through-the-hull. The ImpYak has been equipped with a meteorological station and also with a through-the-hull solar spectrometer for zero-glint HAB spectroscopy. A sea-surface sampling system based around an Arduino microcontroller and a suite of peristaltic pumps have been proven using an 80cm boat, and will be installed on an ImpYak system.
3.2.2 The Galician CIVIL UAV Initiative for Automatic Oceanographic Sampling
In 2017 the Galician government launched a public procurement initiative for unmanned vehicles to address a set of technological challenges4. One of the projects was ‘Automatic oceanographic sampling service using unmanned vehicles’ (MAR-2). The contract was financed in 80% with FEDER funds corresponding to the smart growth operational program (POCInt) 2014-2020 and a collaboration agreement signed between the Ministry of Economy and Competitiveness, the Axencia Galega de Innovación (GAIN) and the Axencia para a Modernización Tecnolóxica de Galicia (AMTEGA) for the development of the Civil UAVs Initiative. The main purpose of the MAR-2 project was to find a technological solution using UAVs resulting in oceanographic sampling to significantly reduce costs of water quality control assessments and other monitoring programs, to enable high frequency data collection and to operate in adverse weather conditions. One of the agencies involved in the design of the technical requirements was INTECMAR, the regional government organization in charge of the Galician HAB monitoring service.
In summer 2020, the consortium of companies that won the public procurement tender presented the USV-Mar II prototype5 (Figure 11). The USV-Mar II prototype is an Environmental Surveillance vessel, 10.5 m in length, made of naval steel with a reinforced fibreglass superstructure. It has a double pilot system, with operations possible from the boat or remotely following a predefined route autonomously. On board, it carries a robotic system with instruments to collect samples from water columns at different depths in predefined geographical points and has a system for data transmission to an operation room in land. The autonomy is more than 300 nautical miles. The operation of the prototype was tested in unmanned and manned mode in autumn 2020-winter 2021 and included taking water samples for nutrient analysis and phytoplankton identification and deploying the multiparametric sonde for oceanographic variables used in the weekly Galician HAB monitoring (Conductivity-Temperature-Depth (CTD) profiler equipped with a WETStar fluorescence sensor) in the Galician rias de Pontevedra and Vigo. The sampling system was found to be able to collect, classify, label, store and automatically conserve all samples.

Figure 11 The USV Mar II prototype, developed in the Galician CIVIL UAV Initiative for Automatic oceanographic sampling, operating in the Galician Rias in manned and unmanned mode. Pictures provided by GAIN, Xunta de Galicia, Spain.
4 Opportunistic Sampling
In this section, we review examples of opportunistic approaches to sample collection outside of the usual coastal monitoring at aquaculture sites.
4.1 HAB Data Sampling by Coastguards
During PRIMROSE, we evaluated how the detection of a harmful bloom could be enhanced by opportunistic sampling, in this section through coordination with the Irish Coast Guard and the Irish Naval Service to collect water samples. A protocol was prepared and distributed (see Supplementary Material: Phytoplankton bloom sampling: Quick guide for Coastguard). Some examples of HAB in- situ sampling by coastguards, from selected years (2013 and 2017) when large biomass blooms occurred, are presented below.
On 27th May 2013, a phytoplankton bloom was detected in surface waters of the Irish Sea by the Marine Institute with the help of a Chl-a satellite data product developed by Ifremer/DYNECO and CERSAT in Brest (France) and NASA in the USA. The causative organism was the prymnesiophyte Phaeocystis. The satellite data allowed visualization of the spatial extent of the bloom with continued daily monitoring. The bloom was evident all along the east to southeast coasts of Ireland from Dublin to Wexford and was associated with cooler waters (Figure 12). After detection by satellite, field samples were taken by the Irish Environmental Protection Agency (EPA) who observed globular Phaeocystis colonies in the water, very evident against the Secchi disk at the surface (Figure 12C). Photographic images of the bloom were taken by a commercial airline pilot on 5th June 2013 ~ 18.5 Km off the Irish coast. Beaches on the east coast of Ireland were affected by this algal bloom, and the presence of Phaeocystis pouchetii was confirmed with water samples taken by the Irish Phytoplankton Monitoring Program the Marine Institute operates. Phaeocystis pouchetii is a common species and is known to have caused blooms along the east coast several times in previous years. The species causes water discoloration and foaming on beaches in windy conditions.
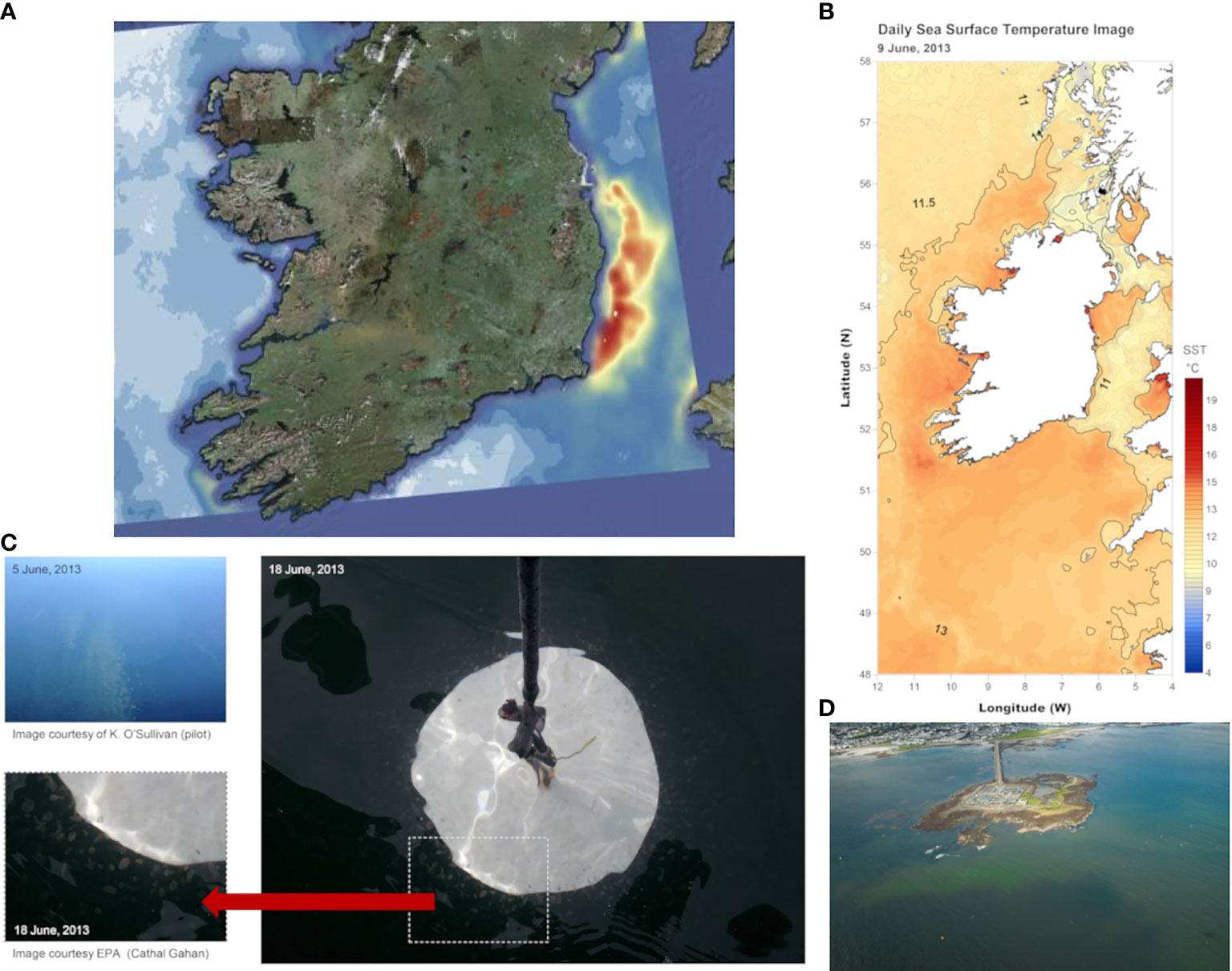
Figure 12 (A) Satellite Chlorophyll image showing the extend of an exceptional Phaeocystis bloom in the Irish Sea, June 2013, (B) Satellite derived sea surface temperature data (CMEMS ODYSSEA SST data product) shows colder water evident at the bloom position (C) photographic images of dense Phaeocystis colonies in surface waters off the north Dublin Irish coastline on 18th June, 2013. The large globular Phaeocystis colonies are visible to the human eye in the water against the Secchi disk at the surface; aerial photograph of the bloom on 5th June taken by a pilot ~ 18.5 Km off the Irish east coast. (D) Bloom of cyanobacteria in Galway Bay, west coast of Ireland, in 2017 taken from a Coastguard Search and Rescue Helicopter revealing the extent of the bloom within the bay.
In September 2017, a blue green algal bloom was reported in bathing areas of Galway Bay, west Ireland. Samples taken were analyzed and cells of the cyanobacteria, genus Anabaena, were predominant and observed in clusters and chains. On 8th September, an opportunity to observe the extent of the bloom was facilitated by the Irish Coastguard Rescue 115 Sikorsky S-92 Helicopter with a number of aerial passes over the affected area in Galway Bay. Photos (Figure 12D) revealed the bloom extended throughout the inner part of the bay and along the north shore; information previously unavailable from shoreline observations.
In 2013, an offshore phytoplankton bloom was detected off the Irish northwest coast in satellite imagery. After detecting the bloom, a report was sent to the Irish Coast Guard on 20th July with information on the geographic extent of the bloom zone. Samples collected by MV Heather Jane II on 26th July were delivered to the Mulroy Coast Guard Unit on return to port. Samples mailed to the Marine Institute were manually analyzed and confirmed a non-harmful coccolithophore bloom that did not require management action. Figure 13 presents the station position and temporal changes of the spatial extent of the bloom over a few days. Coccolithophorids were present at low cell densities (surface = ~6,000 cells l⁻¹; 3.7 m = 3,400 cells l⁻¹) and no toxic or harmful species were present. It is likely that the bloom was at the end stage. Between the 20th and 26th July, the bloom position shifted in an eastward direction away from the targeted sampling region. This may explain the low cell counts in the fixed water samples analyzed.

Figure 13 (A) 20th July 2013 phytoplankton bloom off northwest Ireland NASA (MODIS: Aqua); (B) 26th July 2013 NASA (MODIS Aqua); (C) 20th July 2013 modelled Chlorophyll data and (D) 26th July 2013. Where (C, D) is derived from a data product from Ifremer DYNECO and CERSAT in Brest, France and NASA in the USA. The position of the station where samples were taken by the Irish Coast Guard (IRCG) is indicated in (C, D).
4.2 Coastal Opportunistic Sampling
In summer 2018, an exceptional Paralytic Shellfish Poisoning (PSP) outbreak occurred in the Rías Baixas (Ría de Vigo and Ría de Pontevedra, both southern Galician Rías), northwest Spain (Figure 14). This event was caused by an intense and prolonged (around one month) Alexandrium minutum bloom. The bloom caused prolonged shellfish harvesting closures at mussel rafts and infaunal seabeds due to PSP levels exceeding regulatory limits mainly in Ría de Vigo but also in Pontevedra. Alexandrium minutum has previously been found in Galicia in spring and summer during the upwelling season associated with water stability and stratification, but only in embayments like the Baiona Bay in the Ria de Vigo and the inner part of the Ria de Ares (Bravo et al., 2010). This was the first time that Alexandrium affected a larger area like the Rias de Vigo and Pontevedra. The high concentrations caused water discoloration: at the end of June, red waters appeared in the Ria de Vigo, which were still visible a month later. The Galician monitoring system (INTECMAR) detected the presence of Alexandrium minutum and PSP toxins above regulatory levels and harvesting areas in Vigo and Pontevedra were closed.
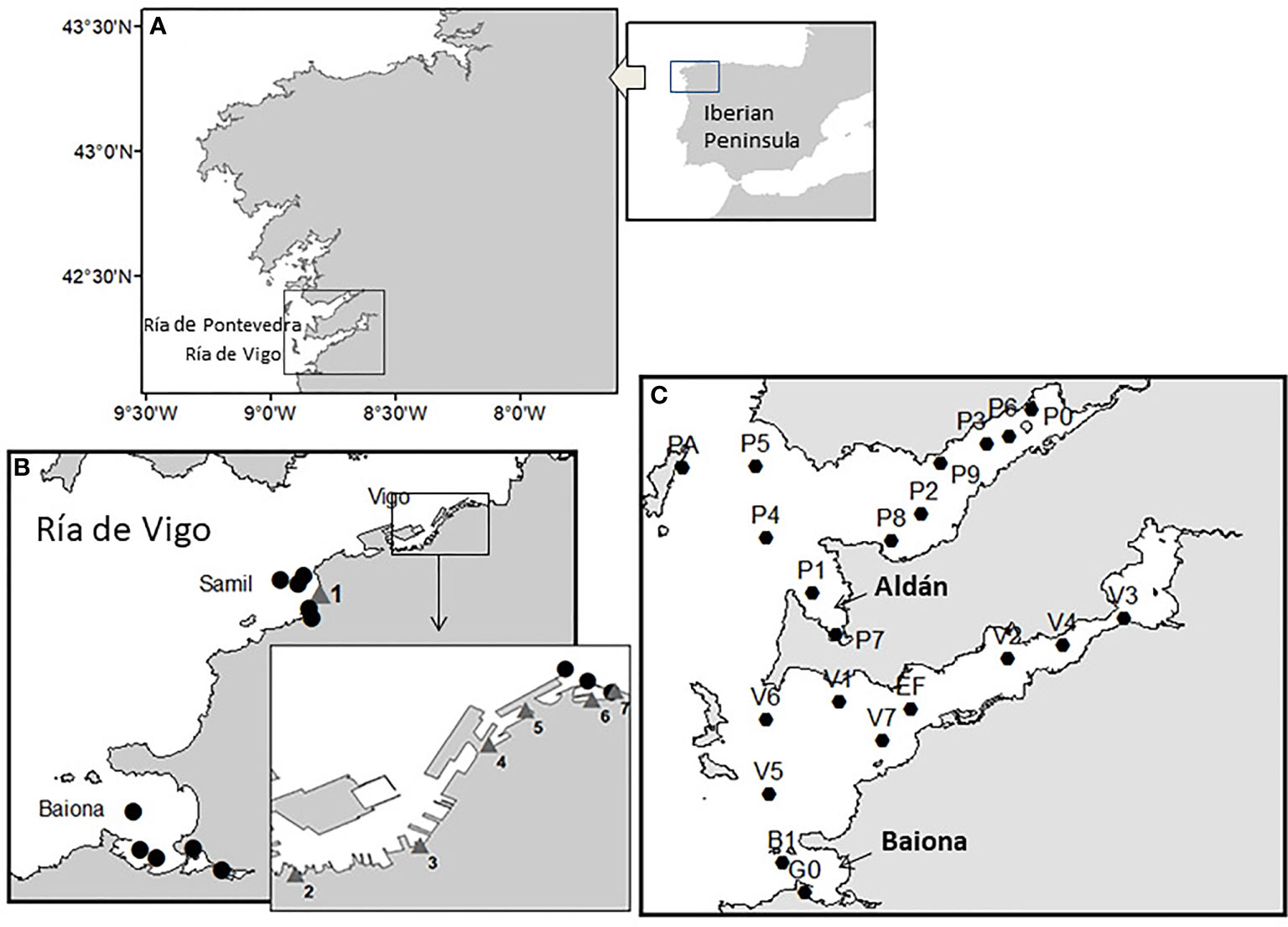
Figure 14 Area of study for the Alexandrium minutum bloom in summer 2018 (Galicia, NW Spain). (A) Rías Baixas. (B) Sampling stations for cyst mapping (circles) and bloom seawater sampling locations (numbered triangles) in Vigo marina and Samil Beach. Fish and invertebrate samples for PSTs analyses were obtained from individuals collected next to 3 and 6. (C) Sampling stations of the Galician monitoring programme (INTECMAR) in the Rías de Vigo and Pontevedra.
The spatial variability of water discoloration outside the monitoring stations and the presence of toxins in marine fauna not sampled in the biotoxin regulatory monitoring in shellfish could be studied with opportunistic sampling during this event (Rodríguez et al., 2018). The location of the sampling stations of INTECMAR and of the places where opportunistic monitoring was performed are provided in Figure 14. On 28th June, a call from the rescue services to the Oceanographic Centre of Vigo (IEO) raised the alarm that Samil Beach waters were discolored, probably due to a red tide. The authorities sought to identify the causative agent and were evaluating whether to restrain bathing until the nature of the phenomenon was clarified. Samil, the most popular urban beach in Vigo, is a few kilometres away from IEO and shore samples were collected rapidly (Figure 15). Reddish-brown patches, likely advected by water currents with the onset of very mild northwesterly winds, formed a bloom strip parallel to the shore visible from the mid-beach to the southern limit of the shore at the mouth of the Lagares River. At the IEO samples were analyzed and Alexandrium cell counts ranged between 30 to 48 x10⁶ cells l⁻¹. Alexandrium became so dominant that the water samples resembled a monospecific culture (Figure 15, see right panel), with only minor presence of other dinoflagellates. The bloom cell densities were somewhat lower (10 x10⁶ cells l⁻¹) in a sample taken 200 m offshore by a rescue boat. Observations of calcofluor-stained samples under the epifluorescence microscope soon identified the species as A. minutum. A detailed description of this summer 2018 A. minutum bloom and an analysis of the environmental factors that triggered this episode is in preparation by Nogueira et al.

Figure 15 PhD student María García-Portela (IEO Vigo) sampling the Alexandrium minutum red tide in Samil Beach, June 28th, 2018. (left). A. minutum: light microscopy micrograph (400X) of the bloom sample from Samil Beach, June 28th (right). Figure reproduced from Rodríguez et al., 2018, Harmful Algae News.
Further opportunistic sampling during the A. minutum bloom event in summer 2018 is reported by Ben-Gigirey et al. (2020). Paralytic Shellfish Toxins (PSTs) were analyzed in marine fauna non-traditional vectors (invertebrates, fish and dolphins), collected at several locations in the Ria de Vigo. Invertebrate and fish samples were collected by scuba divers and local fishermen. In July and August 2018, samples from stranded dolphin (Delphinus delphis) individuals were taken by local research groups (CEMMA, “Coordinadora para o Estudo dos Mamíferos Mariños”, http://www.cemma.org and the Marine Mammals Department at IEO Vigo). Highest PST levels were quantified in bivalve molluscs, however, PSTs were also found in mullet, mackerel, starfish, squids and ascidians. These results highlight the potential for the accumulation of PSTs in marine invertebrates other than shellfish that can potentially act as food web vectors and/or pose a serious risk for human health upon consumption.
5 Discussion
5.1 Use of Drones (RPAS and UAVs) for HAB Monitoring
In the last decade, commercial airborne robotic platforms have become commonplace in the form of UAVs or ‘drones’. Several recent reviews confirm the growing number of applications and developments in aquatic environments (Lally et al., 2019), including experiences in algal bloom monitoring (Kislik et al., 2018; Wu et al., 2019). The flexibility in flight scheduling, the high spatial and temporal resolution of the sampling and the cost effectiveness are the main reported advantages of UAVs, and thus it is expected that UAVs can provide a means of assisting HAB managers and monitoring agencies.
UAVs can collect both images and water samples. The collection of images is promoted by the development of lightweight sensors and allows the acquisition of images in high spatial and temporal resolution (several times per day) with relatively low cost. UAVs in harmful algal research can be more flexible than satellite or airborne sampling and permit adapting to the high spatial and temporal variability of phytoplankton (Kislik et al., 2018). The applications of algal bloom detection with image sensors in UAVs also note that correction and calibration of images can overcome some of the problems of weather, sun glint or aerosols that cause inconsistent optical environments (Kislik et al., 2018; Wu et al., 2019; Cheng et al., 2020), although further testing in real operations is required the generalization of the use of UAVs in algal bloom studies.
Additionally, water samples can be collected by UAVs and drones. Several studies review that water sampling with drones having VTOL capability is feasible (Lally et al., 2019). As described above, during PRIMROSE we were able to prove BVLOS operations of RPAS, opening an opportunity for a step-change in rapid survey and sampling. The technical capability to operate BVLOS has been used in the polar regions for over a decade, but this has relied on the inherent zero risks to air-traffic (as there was no air traffic). The issue with BVLOS when integrating into civilian airspace is to demonstrate that at all times, and during every flight, the aircraft is under complete control, even with intermittent communication or partial system failure; the issue is trust in the aircraft and the operational procedures. These are not yet sufficiently mature for BVLOS to be currently commercially viable.
For the HAB monitoring applications, this dual capability (BVLOS and VTOL) means that aquaculture sites in the Scottish islands and inshore area around the Hebrides are now accessible to the SAMS laboratory at Oban with flight times of under one hour, and a payload capability of 2 -5 kg (depending on flight time required). Delivery of samples from aquaculture sites is essentially identical to a hospital delivery service proven in 2020; water (or other) samples can be transported rapidly from aquaculture sites to the mainland for assay. On the other hand, drones can collect validation data for forecast modelling since they can either fly instrumentation in the pod, rather than deliver physical samples, but also could hover-and-collect sea-surface samples on demand in places outside aquaculture sites, which was the intention of the water-landable quadcopter TD7.
5.2 ASVs in HAB Monitoring
Although not constrained by an equivalent to a Civil Aviation Authority and VLOS rules, the range for ASV operation is limited by the physics of scale. Smaller boats are slower than larger ones, with the hull speed increasing with the square root of the length of the waterline. The volume available for fuel (whether battery or combustion) will also tend to increase with the cube of the waterline length. It is over-simplifying, but a boat four times the length will travel twice as fast for 64x the time (the power requirements will also increase). Boat scale to wave scale also limits the operational limit of upper wind strength. All indicate that for an ASV, bigger is better, and thereby any advantages over existing traditional boats with people on board are diminished.
The Galician development shows how a regional government in the Atlantic Area used public procurement as a tool to advance the TRL of unmanned vehicles towards the demonstration in real conditions of the potential use of this technology in supporting public services. This process requires a high level of preparation effort of the initial procurement documents and concept of the desired development, as well as a high level of financing. A call for tenders was launched in 2017 after a preparation project and a robust prototype is in the demonstration phase. The prototype has been able to collect water samples and operate a CTD in unmanned and manned mode, and further testing is ongoing to ensure that samples can be properly used for phytoplankton identification.
The ImpYak system demonstrated in PRIMROSE benefits from Arduino microcontroller and other technologies affordable to research groups or monitoring programs having limited budget and clearly illustrate the cost-effectiveness of autonomous vehicles. ImpYak can be practically used at the 10 km scale of local aquaculture monitoring, and has been invited to Norway for trials in Svalbard fjords. Although not capable of deep-ocean surveys, these units are proving viable in the intermediate zone between shore-side access with, say, wellington boots or waders, and a large ocean-going vessel: this is the realm of the sea-loch or inner archipelago, which are notoriously difficult to view with satellite or forecast with existing resolution models. Therefore, cost-effective ASVs based on open source solutions like ImpYak are complementary in near coast measurements of HABs, which we have illustrated in this review that are still a necessary complement to HAB monitoring to be able to sample and analyze the presence of harmful algae or toxins.
5.3 Opportunistic and Non-Conventional Sampling
Marine biotoxins are natural compounds mainly produced by marine microalgae, dinoflagellates, and diatoms (Martínez et al., 2015). In terms of their evaluation, it is essential to analyze the toxins in relation to maximum permitted levels defined in the EU Regulation No 853/2004 (European Parliament and Council, 2004) and its amendments: Paralytic Shellfish Toxins, Amnesic Shellfish Toxins, Okadaic acid and Dinophysistoxins, Azaspiracids and Yessotoxins. It is also desirable to evaluate emerging toxins such as Spirolides, Pinnatoxins, Tetrodotoxin, Gymnodimines, etc. In the case of non-traditional vectors, evaluation of these compounds would provide an indication of the presence of marine biotoxins, and their potential accumulation and transfer in the food web, allowing appropriate further monitoring or action to be taken.
The detection of marine biotoxins in non-traditional vectors (rarely covered by the monitoring programs) has been already reported in European countries such as Portugal, UK, Croatia and Spain (Ben-Gigirey et al., 2012; Silva et al., 2013; Silva et al., 2018; Dean et al., 2020). Some of those vectors are consumed by humans and could cause intoxications (Roje-Busatto and Ujevic, 2014). The results from our activity in PRIMROSE demonstrate the need to contemplate non-conventional monitoring to complement traditional monitoring in order to prevent seafood intoxications. For example, the possible presence of PSTs in non-bivalve mollusc species, such as cephalopods, echinoderms and tunicates and the increased interest in the exploitation of marine live resources other than bivalves have promoted a revision of monitoring strategies introducing non-traditional vectors. EU Regulation (EC) No 853/2004 (European Parliament and Council, 2004) sets the maximum PSTs concentrations not only in bivalve molluscs, but also in echinoderms, tunicates and marine gastropods. However, published data relating to these organisms so far are scarce and more studies are needed to evaluate the potential risks they could pose for human health as well as their impacts in the trophic chain. On top of that, more data on the presence of emerging marine toxins in the EU marine invertebrates are also necessary in order to perform risk assessment evaluations studies on these non-traditional vectors.
5.4 IFCs in a HAB Monitoring Context
Traditional light microscopy based HAB monitoring is typically undertaken for shellfish safety on a weekly basis. Given the potential for rapid harmful bloom development under some environmental conditions, this presents a risk to human health. IFCs offer a solution due to their hugely more rapid sample throughput, allowing multiple samples to be analyzed in a day. Finfish farms are not typically supported by such regulatory monitoring and hence often undertake on site microscopy based cell counts on a daily basis. This is labor and time intensive, and while the cost of an IFC is substantial, over the period of a number of years it is likely to be cost effective.
We have shown in our IFC demonstrations that the development of classifiers capable of automatically and reliably identifying and enumerating target phytoplankton cells is critical for use of such instruments in any assessment of phytoplankton communities including HAB species. In the PRIMROSE experience in the Daoulas estuary, the construction of large clusters from daily IFC data in agreement with the protocol for plankton identification from other monitoring stations along the French coast allowed the comparisons with the other data sets, demonstrating that the capacity of IFC for sustained daily monitoring of plankton and an improvement in the assessment of plankton variability. This tool will enlarge the available data set for statistical models [(Fernandes-Salvador et al., 2021), this issue] with new data of primary importance to describe phytoplankton community dynamics. The development of specific classifiers using images is also required but several problems are encountered: while many HAB genera are of concern in multiple countries, the morphological variability of cells requires the development of regional rather than global classifiers. Such developments hinge on the manual identification and annotation of phytoplankton images by trained taxonomists. Ideally many thousand images are required to account for variability in cell size, shape and orientation.
Ongoing improvement of image characterization is the next essential step towards a HAB warning system based on existing classifiers with well-known methods like RFAs, SVMs or CNNs. Typically an IFC image classifier utilizes feature or image characteristic vectors calculated from the labelled images. These characteristics, based on shape or texture descriptors, provide a reduced representation of an image. These vectors are then input to classifiers like RFAs or SVMs that have experienced considerable success in cell classification [e.g. (Sosik and Olson, 2007; Campbell et al., 2010) for SVM (Harred and Campbell, 2014) for RFA]. CNNs are suitable for processing grid-like data such as images and hence may be particularly suitable for IFC data. In fact, deep learning and Convolutional Neural Networks (CNNs) in particular have shown superior performance for image classification tasks (Sharma et al., 2018) and are now being applied to IFC data (Orenstein and Beijbom, 2017; González et al., 2019; Guo et al., 2021). CNNs do not need to specify feature vectors as input because the network itself learns them from the images during the training process called deep features, and may eventually become the method of choice for IFC image classification. For HAB applications, new classifiers for automatic clustering of HAB species need to be specifically constructed based on new data sets like those presented here in the IFC mooring demonstration in the Daoulas estuary for Alexandrium minutum and Pseudo-nitzschia spp. In addition to image recognition for species identification, machine-learning models allow estimation of biovolume from images using distance maps (Moberg and Sosik, 2012). Therefore, IFC have not only a potential to identify HAB species but also to characterize phytoplankton biomass and community composition.
Finally, methods to rapidly disseminate IFC data to stakeholders are required. The IFCB includes a dashboard that is capable of providing data to the user in real time. The interface for discovering and viewing IFC data in the HABreports website (Davidson et al., 2021) is a demonstration of an online user interface system being developed to best characterize and synthesize IFC data for non-expert users to allow them to evaluate HAB risk in their location. The standard clustering and the export flow tested during the deployment of the French IFC mooring follows international standards so that generated data files can be exported in European infrastructures like SeaDataNet, allowing them to be merged with other IFC repositories. Rapid data processing and access, especially of images for clustering, which is the key to an early warning system, remains under construction. The pipelines for data processing and their storage follow the FAIR principles (Findability, Accessibility, Interoperability, and Reusability; Wilkinson et al., 2016).
6 Conclusions and Outlook
The activities undertaken during PRIMROSE clearly show the potential of novel monitoring techniques, especially with autonomous devices, to characterize the high temporal and spatial variability of HABs and open new ways to develop early warning systems that use a larger set of biological data. In the same way, UAVs operating in Beyond Visual Line Of Sight (BVLOS) with Vertical Take-off and Landing (VTOL) and ASVs showed great potential to deliver assay samples and acquire data for model validation.
The examples from France and Scotland demonstrate the potential for IFC based monitoring in locations threatened by HABs, providing markedly enhanced temporal resolution compared to the, typically weekly, regulatory monitoring that currently occurs to ensure shellfish safety. However, the cost of such instruments remains preventative to their wide deployment, at least within the relatively low financial margin shellfish industry. It is therefore important that comparative spatial studies be undertaken to best understand the sphere of influence of regional sentinel sites at which IFCs are deployed. A second IFCB will soon be deployed in Shetland to facilitate such studies in Scottish waters. In the Atlantic Area, a further IFC (Cytobuoy) is also expected to be installed in Galicia by IEO. In France, PRIMROSE partners are now a part of a French national consortium developing an international platform including phytoplankton images. This consortium is supported by the ODATIS Ocean Cluster (Ocean DAta Information and Services, https://www.odatis-ocean.fr/en/). In this way, we envisage participating in developments of pipelines devoted to toxic algae indexes.
Combined monitoring of HABs with the determination of marine biotoxins in non-traditional vectors would enhance the detection of toxic episodes and help to prevent food safety issues. The opportunistic sampling of non-traditional vectors has the advantage of getting very valuable information about the levels of regulated toxins present in those matrices in the production areas. These production areas may be different from those that are currently regularly monitored. Knowing the toxin levels in those vectors, offers the potential to apply measures to prevent human and animal (Dean et al., 2021) intoxications. The information gathered could also guide the design of future monitoring plans. The main drawback is the need for more resources: staff, laboratories equipment, sampling tools, vehicles (i.e. oceanographic vessels), sampling plans in rocky shore areas, etc. Therefore these would add extra-costs to the regular monitoring. The evaluation of emergent toxins in these vectors would additionally provide data for the risk assessment evaluation of those compounds. Therefore they should be reinforced in the near future. Additional studies, supported by PRIMROSE, that aim to evaluate the potential presence of emergent toxins, Cyclic Imines such as Spirolides, Gymnodimine and Pinnatoxins (Villar-González et al., 2006; Davidson et al., 2015; Otero et al., 2019; Lamas et al., 2021) and Tetrodotoxins (Blanco L. et al., 2019) by High Performance Liquid Chromatography coupled to High Resolution Mass Spectrometry in non-traditional vectors from the Galician Rias Baixas remain on-going.
In offshore or non-easily accessible areas, where some HABs initiate, identification relies mainly on satellite observations, limited by cloud cover. However, the potential exists to obtain oceanographic information and plankton samples on the surface from FerryBox devices in commercial vessels (Hartman et al., 2014) or in the water column from autonomous moorings or glider cruises (Stumpf et al., 2010; Seegers et al., 2015). Improving the coordination of oceanographic vessel operators can facilitate the acquisition of samples if the HAB alert happens to coincide with an oceanographic cruise. An illustration of this coordination is reported in Jordan et al., (2021), this issue. Karenia spp. risk was detected by satellite off the south coast of Ireland in July 2019 while an oceanographic cruise was in the area and samples could therefore be taken, confirming the presence of K. mikimotoi. However, most of the time HAB data sampling by coastguards is one of the few methods to acquire HAB in situ data in offshore areas on request. Our experience has shown that elaboration of protocols can improve the capacity of water sampling by coastguards in areas far from monitoring sites, and consequently can complement the HAB detection by satellite, in particular to assess the extent of high biomass blooms in areas of recreation or aquaculture importance. A potential step forward to coordination and protocol sharing initiatives are quick HAB screening approaches like HABscope (Hardison et al., 2019) since it is easy to train non scientists (e.g. fishers, coastguard personnel, aquaculture operators) on its use. This relatively inexpensive, ~$500, robust microscope is attached to an iPod touch and uses Artificial Intelligence (AI) image detection software to isolate the swimming pattern of specific HAB taxa. This facilitates the automatic identification and cell enumeration of the target species with a direct upload and transfer of images via Wi-Fi for verification and integration into HAB early warning systems. Ireland is in the process of testing the NOAA developed HABscope in coastal and shelf waters (Jordan and Cusack, Marine Institute, ongoing work6) to see if Karenia spp. can be detected using the same AI that NOAA uses for K. brevis (Hardison et al., 2019).
Novel monitoring for HABs has the potential to improve the cost-effectiveness of early warning systems. Strategies and obligations vary among countries but HAB monitoring is focused on safeguarding human health rather than on managing aquaculture business risk (Fernandes-Salvador et al., 2021). Since the financial cost of HAB and biotoxin monitoring at all aquaculture sites is prohibitive, the currently regulatory HAB network in a particular country is typically based at a subset of shellfish harvesting sites, with the specific locations and frequency of sampling dictated by local and national risk assessments. Higher frequency autonomous monitoring potentially offers the opportunity to decrease the size of the traditional monitoring network, but will require the identification of sentinel sites that allow the identification of a developing bloom, potentially followed by triggering local sampling at risk aquaculture areas. However, since the current regulatory monitoring of HABs and toxins in regions like the EU is very effective in safeguarding human health (Blanco J. et al., 2019; Belin et al., 2021; Davidson et al., 2021), changes in the monitoring network also require a careful evaluation of the eventual health risk increase associated with the relocation or reduction of sampling sites.
The cost of maintaining HAB monitoring and early warning systems is clearly lower than the cost of HAB induced effects in human health and of socio economic impacts in the aquaculture sector. However, an accurate assessment of the actual effect of short term forecasts mitigation and consequently of their economical impact is very difficult to achieve because the estimation of the real cost of HAB is complex. For fish aquaculture, the economic loss is clear in extreme events causing fish mortality (Karlson et al., 2021) but the losses due to minor fish kills and sub-lethal events require careful evaluation of the mitigation strategies and a cost assessment would involve reviewing aquaculture business planning as well as sharing of monitoring and potentially sensitive commercial information between industry and scientists (Davidson et al., 2020). For shellfish aquaculture, economical evaluation of costs is more difficult since biogeography, seasonal variability, regularity of the bloom, type of shellfish and commercialisation issues can strongly modulate the cost of HAB (Rodriguez et al., 2011; Martino et al., 2020; Guillotreau et al., 2021; Karlson et al., 2021).
Anyway, advancement of existing forecasting approaches requires additional inputs to provide a better understanding and monitoring of the distribution of HAB species and microbiological contamination. We have seen that novel methodologies exist and have potential to complement existing monitoring by improving spatial and temporal coverage of observations. The methodologies we have presented can enhance the capacity of sampling in areas outside the monitoring sites, provide earlier detection of the presence of HAB species and allow higher temporal monitoring and cost-efficient sampling in the period between consecutive samplings in the regulatory monitoring. Thus, the higher temporal frequency provided by these approaches can improve the forecasts as data becomes hourly/daily rather than weekly. Therefore, we believe that novel techniques will lead to improved prediction and early warning of future events resulting in an enhanced mitigation capacity, which will benefit aquaculture producers. The PRIMROSE experience shows that advances can be achieved through cooperation and sharing of knowledge, experiences and demonstrations.
Finally, for all novel monitoring, despite some unavoidable time-consuming constraints associated with sensor servicing or administrative processes like permissions or limitations for flight, the emerging bottleneck appearing now is the data flow. There remains a lot of technical development and further testing is required to obtain these new data sets in real or near real time. Early warning systems for HABs and microbiological risk in aquaculture are expected to be able to make a strong step forward when these pipelines are fully implemented.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors upon request. Any requests for Marine Institute Digital Data, please go to this link: https://www.marine.ie/Home/marine-institute-request-digital-data. The data set from the Somlit monitoring used in section 2.1 has been obtained from a third party. Requests to access these datasets should be directed to Lambert C., Y2hyaXN0b3BoZS5sYW1iZXJ0QHVuaXYtYnJlc3QuZnIu
Author Contributions
All authors participated in the research and discussion of the potential of novel methodologies for providing in situ data to HAB early warning systems. MS and LQ deployed and analysed IFC in the Bay of Brest. KD, CW, SG-P and GA deployed and analysed IFC in Scotland. PA coordinated drone development and demonstrations in Scotland. CC and JS designed HAB data sampling by coastguards. FR and BB-G participated in samplings of harmful algae and marine biotoxins in Galicia. MR-V coordinated research and writing of the manuscript and produced the first draft together with MS, PA, KD, CC and PN and all authors, who approved it for publication.
Funding
This work has been funded by the European Union Interreg Atlantic Area project PRIMROSE EAPA182_2016, an IOC-SCOR GlobalHAB endorsed project. KD and CW were also funded by the UKRI grants NE/T008571/1 and BB/S004246/1 and SG-P was funded by a PhD studentship funded by Marine Scotland Science and the Data Lab. Sampling in Ría de Vigo had support from the Spanish National Project DIANAS (CTM2017-86066-R, MICINN, Spain) and Project IN607A 2019/04 (GRC, Xunta de Galicia, Spain).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank Paola Arce of SAMS for her contribution to the Scottish IFCB development work. We also thank Marie Latimier and Julien Quere for their contributions to the Ifremer flow cytometer deployments and the data analyses. Special thanks to Alex Brown from Wingcopter for providing and allowing the reproduction of pictures in Figure 9, to the Departamento de Xestión de Innovación, Área de Programas of the Axencia Galega de Innovacion for providing pictures of the demonstration of the MAR2 prototype in Figure 11, and to Cathal Gahan (Environmental Protection Agency, Ireland) and Kevin O’Sullivan (Irish pilot) for providing photographic evidence of phytoplankton blooms in Figure 12.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2022.791329/full#supplementary-material
Footnotes
- ^ https://www.shellfish-safety.eu/
- ^ https://www.somlit.fr/brest/
- ^ VLOS is set out in the UK under CAP722 and similar international regulation exists in all International Civil Aviation Organization (ICAO) countries
- ^ https://www.civiluavsinitiative.com/en/programmes/solutions-program/
- ^ https://seadrone.es/mar-2/
- ^ https://www.marine.ie/Home/site-area/news-events/press-releases/irish-scientists-collaborate-noaa-test-new-habscope.
References
Artigas F., Créach V., Houliez E., Karlson B., Lizon F., Seppälä J., et al. (2019). Novel Methods for Automated in Situ Observations of Phytoplankton Diversity and Productivity: Synthesis of Exploration, Inter Comparisons and Improvements. JERICO-NEXT WP3, Deliverable 3.2. Version 5 (Brest, France: IFREMER), 88pp. JERICO-NEXT-WP3-D3.2-120819-V5. doi: 10.25607/OBP-945
Belin C., Soudant D., Amzil Z. (2021). Three Decades of Data on Phytoplankton and Phycotoxins on the French Coast: Lessons From REPHY and REPHYTOX. Harmful Algae 102, 101733. doi: 10.1016/j.hal.2019.101733
Ben-Gigirey B., Rodríguez-Velasco M. L., Otero A., Vieites J. M., Cabado A. G. (2012). A Comparative Study for PSP Toxins Quantification by Using MBA and HPLC Official Methods in Shellfish. Toxicon 60, 864–873. doi: 10.1016/j.toxicon.2012.05.022
Ben-Gigirey B., Rossignoli A. E., Riobó P., Rodríguez F. (2020). First Report of Paralytic Shellfish Toxins in Marine Invertebrates and Fish in Spain. Toxins 12 (11), 723. doi: 10.3390/toxins12110723
Berdalet E., Fleming L. E., Gowen R., Davidson K., Hess P., Backer L. C., et al. (2016). Marine Harmful Algal Blooms, Human Health and Wellbeing: Challenges and Opportunities in the 21st Century. J. Mar. Biol. Assoc. U.K. 2015, 61–91. doi: 10.1017/S0025315415001733
Blanco J., Arévalo F., Correa J., Moroño Á. (2019). Lipophilic Toxins in Galicia (NW Spain) Between 2014 and 2017: Incidence on the Main Molluscan Species and Analysis of the Monitoring Efficiency. Toxins 11 (10), 612. doi: 10.3390/toxins11100612
Blanco L., Lago J., González V., Paz B., Rambla-Alegre M., Cabado A. G. (2019). Occurrence of Tetrodotoxin in Bivalves and Gastropods From Harvesting Areas and Other Natural Spaces in Spain. Toxins (Basel) 11, 1–9. doi: 10.3390/toxins11060331
Bravo I., Fraga S., Figueroa R. I., Pazos Y., Massanet A., Ramilo I. (2010). Bloom Dynamics and Life Cycle Strategies of Two Toxic Dinoflagellates in a Coastal Upwelling System (NW Iberian Peninsula). Deep Sea Res. Part II Top. Stud. Oceanogr. 57, 222–234. doi: 10.1016/j.dsr2.2009.09.004
Bresnan E., Arévalo F., Belin C., Branco M. A., Cembella A. D., Clarke D., et al. (2021). Diversity and Regional Distribution of Harmful Algal Events Along the Atlantic Margin of Europe. Harmful Algae 102, 101976. doi: 10.1016/j.hal.2021.101976
Campbell L., Henrichs D. W., Olson R. J., Sosik H. M. (2013). Continuous Automated Imaging-In-Flow Cytometry for Detection and Early Warning of Karenia Brevis Blooms in the Gulf of Mexico. Environ. Sci. Pollut. Res. 20, 6896–6902. doi: 10.1007/s11356-012-1437-4
Campbell L., Olson R. J., Sosik H. M., Abraham A., Henrichs D. W., Hyatt C. J., et al. (2010). First Harmful Dinophysis (Dinophyceae, Dinophysiales) Bloom In the Us Is Revealed By Automated Imaging Flow Cytometry. J. Phycol. 46, 66–75. doi: 10.1111/j.1529-8817.2009.00791.x
Chapelle A., Le Gac M., Labry C., Siano R., Quere J., Caradec F., et al. (2015). The Bay of Brest (France), a New Risky Site for Toxic Alexandrium Minutum Blooms and PSP Shellfish Contamination. Harmful Algae News 51, 4–5. doi: 10.5281/zenodo.5110019
Cheng K. H., Chan S. N., Lee J. H. W. (2020). Remote Sensing of Coastal Algal Blooms Using Unmanned Aerial Vehicles (UAVs). Mar. Pollut. Bull. 152, 110889. doi: 10.1016/j.marpolbul.2020.110889
Dashkova V., Malashenkov D., Poulton N., Vorobjev I., Barteneva N. S. (2017). Imaging Flow Cytometry for Phytoplankton Analysis. Methods San Diego Calif. 112, 188–200. doi: 10.1016/j.ymeth.2016.05.00
Davidson K., Anderson D. M., Mateus M., Reguera B., Silke J., Sourisseau M., et al. (2016). Forecasting the Risk of Harmful Algal Blooms. Harmful Algae 53, 1–7. doi: 10.1016/j.hal.2015.11.005
Davidson K., Baker C., Higgins C., Higman W., Swan S., Veszelovszki A., et al. (2015). Potential Threats Posed by New or Emerging Marine Biotoxins in UK Waters and Examination of Detection Methodologies Used for Their Control: Cyclic Imines. Mar. Drugs 13, 7087–7112. doi: 10.3390/md13127057
Davidson K., Jardine S., Martino S., Myre G., Peck L., Raymond R., et al. (2020). “The Economic Impacts of Harmful Algal Blooms on Salmon Cage Aquaculture,” in GlobalHAB. Evaluating, Reducing and Mitigating the Cost of Harmful Algal Blooms: A Compendium of Case Studies. Ed. Trainer V. L., (Sidney, BC, Canada: PICES Press) 84–94+.
Davidson K., Whyte C., Aleynik D., Dale A., Gontarek S., Kurekin A. A., et al. (2021). HABreports: Online Early Warning of Harmful Algal and Biotoxin Risk for the Scottish Shellfish and Finfish Aquaculture Industries. Front. Mar. Sci. 8, 631732. doi: 10.3389/fmars.2021.631732
Dean K. J., Alexander R. P., Hatfield R. G., Lewis A. M., Coates L. N., Collin T., et al. (2021). The Common Sunstar Crossaster Papposus—A Neurotoxic Starfish. Mar. Drugs 19, 695. doi: 10.3390/md19120695
Dean K. J., Hatfield R. G., Lee V., Alexander R. P., Lewis A. M., Maskrey B. H., et al. (2020). Multiple New Paralytic Shellfish Toxin Vectors in Onshore North Sea Benthos, a Deep Secret Exposed. Mar. Drugs 18, 400. doi: 10.3390/md18080400
Dunker S. (2019). Hidden Secrets Behind Dots: Improved Phytoplankton Taxonomic Resolution Using High-Throughput Imaging Flow Cytometry. Cytometry A 95, 854–868. doi: 10.1002/cyto.a.23870
Dunker S., Boho D., Waeldchen J., Maeder P. (2018). Combining High-Throughput Imaging Flow Cytometry and Deep Learning for Efficient Species and Life-Cycle Stage Identification of Phytoplankton. BMC Ecol. 18, 51. doi: 10.1186/s12898-018-0209-5
Escalera L., Pazos Y., Dolores Doval M., Reguera B. (2012). A Comparison of Integrated and Discrete Depth Sampling for Monitoring Toxic Species of Dinophysis. Mar. Pollut. Bull. 64 (1), 106–113. doi: 10.1016/j.marpolbul.2011.10.015
Escalera L., Reguera B., Moita T., Pazos Y., Cerejo M., Cabanas J. M., et al. (2010). Bloom Dynamics of Dinophysis Acuta in an Upwelling System: In Situ Growth Versus Transport. Harmful Algae 9 (3), 312–322. doi: 10.1016/j.hal.2009.12.002
European Parliament and Council (2004). Regulation No 853/2004 of 29 April 2004 Laying Down Specific Hygiene Rules for Food of Animal Origin. J. Eur. Union L 139, 55–205. 2004.
Fernandes-Salvador J. A., Davidson K., Sourisseau M., Revilla M., Schmidt W., Clarke D., et al. (2021). Current Status of Forecasting Toxic Harmful Algae for the North-East Atlantic Shellfish Aquaculture Industry. Front. Mar. Sci. 8, 666583. doi: 10.3389/fmars.2021.666583
First M. R., Drake L. A. (2012). Performance of the Human “Counting Machine”: Evaluation of Manual Microscopy for Enumerating Plankton. J. Plankton Res. 34, 1028–1041. doi: 10.1093/plankt/fbs068
Fischer A. D., Hayashi K., McGaraghan A., Kudela R. M. (2020). Return of the “Age of Dinoflagellates” in Monterey Bay: Drivers of Dinoflagellate Dominance Examined Using Automated Imaging Flow Cytometry and Long-Term Time Series Analysis. Limnol. Oceanogr. 65, 2125–2141. doi: 10.1002/lno.11443
Gianella F., Burrows M. T., Swan S. C., Turner A. D., Davidson K. (2021). Temporal and Spatial Patterns of Harmful Algae Affecting Scottish Shellfish Aquaculture. Front. Mar. Sci. 8, 785174. doi: 10.3389/fmars.2021.785174
González P., Castaño A., Peacock E. E., Díez J., Del Coz J. J., Sosik H. M. (2019). Automatic Plankton Quantification Using Deep Features. J. Plankton Res. 41 (4), 449–463. doi: 10.1093/plankt/fbz023
Guillotreau P., Le Bihan V., Morineau B., Pardo S. (2021). The Vulnerability of Shellfish Farmers to HAB Events: An Optimal Matching Analysis of Closure Decrees. Harmful Algae. 101, 101968. doi: 10.1016/j.hal.2020.101968
Guo J., Ma Y., Lee J. H. (2021). Real-Time Automated Identification of Algal Bloom Species for Fisheries Management in Subtropical Coastal Waters. J. Hydro-Environ. Res. 36, 1–32. doi: 10.1016/j.jher.2021.03.002
Hallegraeff G. M. (2004). “Harmful Algal Blooms: A Global Overview,” in Manual on Harmful Marine Microalgae, 2nd revised edition. Eds. Hallegraeff G. M., Anderson D. M., Cembella A. D., Enevoldsen H. O. (Paris, France: UNESCO), 793. (Monographs on Oceanographic Methodology, 11).
Hardison D. R., Holland W. C., Currier R. D., Kirkpatrick B., Stumpf R., Fanara T., et al. (2019). HABscope: A Tool for Use by Citizen Scientists to Facilitate Early Warning of Respiratory Irritation Caused by Toxic Blooms of Karenia Brevis. PloS One 14 (6), e0218489. doi: 10.1371/journal.pone.0218489
Harred L. B., Campbell L. (2014). Predicting Harmful Algal Blooms: A Case Study With Dinophysis ovum in the Gulf of Mexico. J. Plankton Res. 36, 1434–1445. doi: 10.1093/plankt/fbu070
Hartman S. E., Hartman M. C., Hydes D. J., Smythe-Wright D., Gohin F., Lazure P. (2014). The Role of Hydrographic Parameters, Measured From a Ship of Opportunity, in Bloom Formation of Karenia Mikimotoi in the English Channel. J. Mar. Syst. 140, 39–49. doi: 10.1016/j.jmarsys.2014.07.001
Jordan C., Cusack C., Tomlinson M. C., Meredith A., McGeady R., Salas R., et al. (2021). Using the Red Band Difference Algorithm to Detect and Monitor a Karenia Spp. Bloom Off the South Coast of Ireland, June 2019. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.638889
Karlson B., Andersen P., Arneborg L., Cembella A., Eikrem W., John U., et al. (2021). Harmful Algal Blooms and Their Effects in Coastal Seas of Northern Europe. Harmful Algae 102, 101989. doi: 10.1016/j.hal.2021.101989
Karlson B., Cusack C., Bresnan E. (Eds.) (2010). Microscopic and Molecular Methods for Quantitative Phytoplankton Analysis (Intergovernmental Oceanographic Commission Manuals and Guides; 55). (Paris, France: UNESCO). doi: 10.25607/OBP-1371
Kislik C., Dronova I., Kelly M. (2018). UAVs in Support of Algal Bloom Research: A Review of Current Applications and Future Opportunities. Drones 2, 35. doi: 10.3390/drones2040035
Kraft K., Seppälä J., Hällfors H., Suikkanen S., Ylöstalo P., Anglès S., et al. (2021). First Application of IFCB High-Frequency Imaging-In-Flow Cytometry to Investigate Bloom-Forming Filamentous Cyanobacteria in the Baltic Sea. Front. Mar. Sci. 8, 594144. doi: 10.3389/fmars.2021.594144
Kurekin A. A., Miller P. I., van der Woerd H. J. (2014). Satellite Discrimination of Karenia mikimotoi and Phaeocystis Harmful Algal Blooms in European Coastal Waters: Merged Classification of Ocean Colour Data. Harmful Algae 31, 163–176. doi: 10.1016/j.hal.2013.11.003
Lally H. T., O’Connor I., Jensen O. P., Graham C. T. (2019). Can Drones be Used to Conduct Water Sampling in Aquatic Environments? A Review. Sci. Total Environ. 670, 569–575. doi: 10.1016/j.scitotenv.2019.03.252
Lamas J. P., Arévalo F., Moroño Á., Correa J., Rossignoli A. E., Blanco J. (2021). Gymnodimine A in Mollusks From the North Atlantic Coast of Spain: Prevalence, Concentration, and Relationship With Spirolides. Environ. Pollut. 279, 116919. doi: 10.1016/j.envpol.2021.116919
Lundholm N., Churro C., Fraga S., Hoppenrath M., Iwataki M., Larsen J., et al. (Eds.) (2009). IOC-UNESCO Taxonomic Reference List of Harmful Micro Algae. (Paris, France: UNESCO). Available at: https://www.marinespecies.org/hab on 2022-03-14. doi: 10.14284/362
Maguire J., Cusack C., Ruiz-Villarreal M., Silke J., McElligott D., Davidson K. (2016). Applied Simulations and Integrated Modelling for the Understanding of Toxic and Harmful Algal Blooms (ASIMUTH): Integrated HAB Forecast Systems for Europe’s Atlantic Arc. Harmful Algae 53, 160–166. doi: 10.1016/j.hal.2015.11.006
Martínez A., Garrido-Maestu A., Ben-Gigirey B., Chapela M. J., González V., Vieites J. M., et al. (2015). “Marine Biotoxins,” in Springer Handbook of Marine Biotechnology. Ed. Kim S.-K. (Heidelberg, Germany: Springer), 869–904. doi: 10.1007/978-3-642-53971-8_37
Martino S., Gianella F., Davidson K. (2020). An Approach for Evaluating the Economic Impacts of Harmful Algal Blooms: The Effects of Blooms of Toxic Dinophysis spp. on the Productivity of Scottish Shellfish Farms. Harmful Algae. 99, 101912. doi: 10.1016/j.hal.2020.101912
Mateus M., Fernandes J., Revilla M., Ferrer L., Villarreal M. R., Miller P., et al. (2019). Early Warning Systems for Shellfish Safety: The Pivotal Role of Computational Science, in Proceedings of the International Conference on Computational Science (Cham: Springer) 361–375. doi: 10.1007/978-3-030-22747-0_28
Moberg E. A., Sosik H. M. (2012). Distance Maps to Estimate Cell Volume From Two-Dimensional Plankton Images. Limnol. Oceanogr. Methods 10, 278–288. doi: 10.4319/lom.2012.10.278
Orenstein E. C., Beijbom O. (2017). “Transfer Learning and Deep Feature Extraction for Planktonic Image Data Sets,” in IEEE Winter Conference on Applications of Computer Vision (WACV),, 2017. 1082–1088. doi: 10.1109/WACV.2017.125
Otero P., Miguéns N., Rodríguez I., Botana L. M. (2019). LC–MS/MS Analysis of the Emerging Toxin Pinnatoxin-G and High Levels of Esterified OA Group Toxins in Galician Commercial Mussels. Toxins (Basel) 11, 394. doi: 10.3390/toxins11070394
Raine R.McDermott G.Silke J.Lyons K.Nolan G.Cusack C. (2010). A Simple Short Range Model for the Prediction of Harmful Algal Events in the Bays of Southwestern Ireland. J. Mar. Syst. 83 (3–4), 150–157. doi: 10.1016/j.jmarsys.2010.05.001
REPHY – French Observation and Monitoring program for Phytoplankton and Hydrology in coastal waters (2021). REPHY Dataset - French Observation and Monitoring Program for Phytoplankton and Hydrology in Coastal Waters. Metropolitan data (SEANOE). doi: 10.17882/47248
Rodríguez F., Escudeiro A., Reguera B. (2018). Red Waters in Ría De Vigo (NW Spain). Harmful Algae News 61, 1–4. doi: 10.5281/zenodo.5109858
Rodríguez G. R., Villasante S., García-Negro M. C. (2011). Are Red Tides Affecting Economically the Commercialization of the Galician (NW Spain) Mussel Farming? Mar. Policy 35 (2), 252–257. doi: 10.1016/j.marpol.2010.08.008
Roje-Busatto R., Ujevic I. (2014). PSP Toxins Profile in Ascidian Microcosmus Vulgaris (Heller 1877) After Human Poisoning in Croatia (Adriatic Sea). Toxicon 79, 28–36. doi: 10.1016/j.toxicon.2013.12.014
Seegers B. N., Birch J. M., Marin R. III, Scholin C. A., Caron D. A., Seubert E. L., et al. (2015). Subsurface Seeding of Surface Harmful Algal Blooms Observed Through the Integration of Autonomous Gliders, Moored Environmental Sample Processors, and Satellite Remote Sensing in Southern California. Limnol. Oceanogr. 60, 754–764. doi: 10.1002/lno.10082
Sharma N., Jain V., Mishra A. (2018). An Analysis of Convolutional Neural Networks for Image Classification. Proc. Comput. Sci. 132, 377–384. doi: 10.1016/j.procs.2018.05.198
Silva M., Barreiro A., Rodriguez P., Otero P., Azevedo J., Alfonso A., et al. (2013). New Invertebrate Vectors for PST, Spirolides and Okadaic Acid in the North Atlantic. Mar. Drugs 11, 1936–1960. doi: 10.3390/md11061936
Silva M., Rey V., Barreiro A., Kaufmann M., Neto A. I., Hassouani M., et al. (2018). Paralytic Shellfish Toxins Occurrence in non-Traditional Invertebrate Vectors From North Atlantic Waters (Azores, Madeira, and Morocco). Toxins 10, 362. doi: 10.3390/toxins10090362
Sosik H. M., Olson R. J. (2007). Automated Taxonomic Classification of Phytoplankton Sampled With Imaging-in-Flow Cytometry. Limnol. Oceanogr. Methods 5, 204–216. doi: 10.4319/lom.2007.5.204
Stauffer B. A., Bowers H. A., Earle B., Davis T. W., Johengen T. H., Kudela R., et al. (2019). Considerations in Harmful Algal Bloom Research and Monitoring: Perspectives From a Consensus-Building Workshop and Technology Testing. Front. Mar. Sci. 6, 399. doi: 10.3389/fmars.2019.00399
Stumpf S., Fleming-Lehtinen V., Granéli E. (2010). “Integration of Data for Nowcasting of Harmful Algal Blooms,” In: Hall J., Harrison D. E., Stammer D., editors. Proceedings of OceanObs’09: Sustained Ocean Observations and Information for Society, 21–25 September 2009 (Venice, Italy: ESA Publication WPP-306). doi: 10.5270/OceanObs09.36
Stumpf R. P., Tomlinson M. C., Calkins J. A., Kirkpatrick B., Fisher K., Nierenberg K., et al. (2009). Skill Assessment for an Operational Algal Bloom Forecast System. J. Mar. Syst. 76 (1-2), 151–161. doi: 10.1016/j.jmarsys.2008.05.016
Swan S. C., Turner A. D., Bresnan E., Whyte C., Paterson R. F., McNeill S., et al. (2018). Dinophysis Acuta in Scottish Coastal Waters and Its Influence on Diarrhetic Shellfish Toxin Profiles. Toxins 10, 399. doi: 10.3390/toxins10100399
Thyssen M., Tarran G. A., Zubkov M. V., Holland R. J., Gregori G., Burkill P. H., et al, et al. (2008). The Emergence of Automated High-Frequency Flow Cytometry: Revealing Temporal and Spatial Phytoplankton Variability. J. Plankton Res. 30, 333–343. doi: 10.1093/plankt/fbn005
Trainer V. L., Pitcher G. C., Reguera B., Smayda T. J. (2010). The Distribution and Impacts of Harmful Algal Bloom Species in Eastern Boundary Upwelling Systems. Prog. Oceanogr. 85, 33–52. doi: 10.1016/j.pocean.2010.02.003
Villar-González A., Rodríguez-Velasco M. L., Ben-Gigirey B., Botana L. M. (2006). First Evidence of Spirolides in Spanish Shellfish. Toxicon 48, 1068–1074. doi: 10.1016/j.toxicon.2006.09.001
Weeks R. (2019). Use of Multi- and Hyper-Spectral Techniques for Examining Ocean Reflectance From Remotely Piloted Aircraft Platforms (Aberdeen University).
Wenhai L., Cusack C., Baker M., Tao W., Mingbao C., Paige K., et al. (2019). Successful Blue Economy Examples With an Emphasis on International Perspectives. Front. Mar. Sci. 6. doi: 10.3389/fmars.2019.00261
Whyte C., Swan S., Davidson K. (2014). Changing Wind Patterns Linked to Unusually High Dinophysis Blooms Around the Shetland Islands, Scotland. Harmful Algae 39, 365–373. doi: 10.1016/j.hal.2014.09.006
Wilkinson M. D., Dumontier M., Aalbersberg I. J., Appleton G., Axton M., Baak A., et al. (2016). The FAIR Guiding Principles for Scientific Data Management and Stewardship. Sci. Data 3, 160018. doi: 10.1038/sdata.2016.18
Keywords: harmful algal blooms (HABs), HAB early warning, HAB observing system, autonomous imaging flow cytometry (IFC), drones, remotely piloted aircraft systems (RPAS), unmanned aerial vehicles (UAV), autonomous surface vehicles (ASVs)
Citation: Ruiz-Villarreal M, Sourisseau M, Anderson P, Cusack C, Neira P, Silke J, Rodriguez F, Ben-Gigirey B, Whyte C, Giraudeau-Potel S, Quemener L, Arthur G and Davidson K (2022) Novel Methodologies for Providing In Situ Data to HAB Early Warning Systems in the European Atlantic Area: The PRIMROSE Experience. Front. Mar. Sci. 9:791329. doi: 10.3389/fmars.2022.791329
Received: 08 October 2021; Accepted: 01 March 2022;
Published: 14 April 2022.
Edited by:
Tomaso Fortibuoni, Istituto Superiore per la Protezione e la Ricerca Ambientale (ISPRA), ItalyReviewed by:
Johan Robbens, Institute for Agricultural, Fisheries and Food Research (ILVO), BelgiumAletta T. Yñiguez, University of the Philippines Diliman, Philippines
Holly Bowers, Moss Landing Marine Laboratories, United States
Raphael M. Kudela, University of California, Santa Cruz, United States
Copyright © 2022 Ruiz-Villarreal, Sourisseau, Anderson, Cusack, Neira, Silke, Rodriguez, Ben-Gigirey, Whyte, Giraudeau-Potel, Quemener, Arthur and Davidson. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Manuel Ruiz-Villarreal, bWFudWVsLnJ1aXpAaWVvLmVz; bWFudWVsLnJ1aXpAaWVvLmNzaWMuZXM=
†Present address: Begoña Ben-Gigirey, European Union Reference Laboratory for Monitoring of Marine Biotoxins, Vigo, Spain