 Christian Díaz1,2*†
Christian Díaz1,2*† Catterina Sobenes
Catterina Sobenes Juan José Cruz Motta
Juan José Cruz Motta Rodrigo Riera
Rodrigo Riera- 1Departamento de Ingeniería Industrial, Facultad de Ingeniería, Universidad Católica de la Santísima Concepción, Concepción, Chile
- 2Centro de Investigación en Biodiversidad y Ambientes Sustentables (CIBAS), Universidad Católica de la Santísima Concepción, Concepción, Chile
- 3Departamento de Ingeniería Civil, Facultad de Ingeniería, Universidad Católica de la Santísima Concepción, Concepción, Chile
- 4Department of Marine Science, University of Puerto Rico, Mayagüez, Puerto Rico
- 5BIOCON, IU-ECOAQUA, Universidad de Las Palmas de Gran Canaria, Las Palmas de Gran Canaria, CI, Spain
- 6Departamento de Ecología, Facultad de Ciencias, Universidad Católica de la Santísima Concepción, Concepción, Chile
Most of the marine bivalve production comes from aquaculture and only 11% comes from the wild fishery. For example, mussel aquaculture still strongly relies on spat - the recently settled juveniles. Gaps of information on the recruitment and settlement of spat are important drawbacks for the development of mussel aquaculture in several geographic regions. We here study these processes using three collectors (Anchovy mesh, Rope and Raschel mesh) throughout one year in two coastal sites of Central Chile (Coliumo and Punta Lobería) at three depths (2, 4 and 6 m). Five spat species were settled in the collectors, i.e. Aulacomya atra, Choromytilus chorus, Mytilus chilensis, Mytilus galloprovincialis and Semimytilus algosus. Recruitment was consistently higher during summer and autumn at both sites. Different seasonal recruitment patterns were observed regarding the spat species. Spat collection of the former mytilid species occurs from spring to autumn with the exception of M. galloprovincialis in Coliumo and M. chilensis in Punta Lobería. M. galloprovincialis is settled mostly in autumn and summer, and M. chilensis during winter. There were also differences when considering the collector type, mostly due to A. atra and S. algosus which peaked in nylon rope. Spat collection of the commercial species M. chilensis should be preferentially carried out in nylon rope during autumn in Coliumo and summer and autumn in Punta Lobería. During summer and autumn, also S. algosus will be settled. These results will enable the implementation of management measures to promote mytilid culture on the coastal zone of the Biobío region.
Introduction
The global production of marine bivalves for human consumption is over 17.7 million tonnes per year during the period 2020, which is about 14.6% of the overall marine global production (FAO, 2022). Mussel production is one of the most important within bivalves aquaculture, steadily increasing to reach 2 million tonnes in 2016 (FAO, 2021). Most of the mussel seeds are collected by means of collectors, deployed adjacent to natural banks. The collectors are materials that act as a substrate for the mitilid larvae to settle, to later be extracted from them and used in the fattening process (Wijsman et al., 2019).
In the European Union, four culture techniques have been developed, namely, raft culture and long-line culture, both suspended, and bottom culture and bouchot culture, the latter two associated with the seabed (Avdelas et al., 2021). In Chile, the materials extensively used are stamped anchovy mesh, knotted anchovy mesh, salmon mesh, raschel mesh and ropes, because the fiber composition and structure of these collectors are important factors for the settlement of mitilid larvae (Çelik et al., 2016). In NW Spain, the seed collection by means of artificial collector ropes has increased (Filgueira et al., 2007). This, due to the fact that their higher growth rate is carried out in longline crops, but, nevertheless, the establishment of the larvae shows a high spatial and temporal variability, due to several biotic and abiotic factors that participate in both dispersal and in colonization (Peteiro et al., 2010). Among the biotic factors, it needs to take into consideration the effects of biofilms in substrates where mitilid larvae of certain species, such as Mytilus coruscus (Yi-Feng et al., 2014), Perna canaliculus (Ganesan et al., 2010) and Argopecten purpuratus (Avendaño-Herrera et al., 2002) can be found. Regarding the abiotic factors, it can be noted that the settlement season for Mytilus galloprovincialis takes place from late autumn to early winter, but there is no information on which are the best areas for laying collectors or which are the optimal depths of settlement, this information is useful to improve seed collection practices and to understand the dynamics of substrate colonization (Curiel-Ramírez and Cáceres-Martínez, 2010).
Pediveliger larvae of mitilids, commonly known as “spat”, are able to discriminate between substrates and are selective in their preferences for settlement (Carl et al., 2012a). These larvae can produce a non-permanent settlement as a way of testing the selected substrate, being able to separate from it and re-settle in an alternative habitat (Carl et al., 2012b). Thus, spat remain highly mobile and can detach several times from their primary settlement site and drift away to resettle, i.e. secondary settlement (Buchanan and Babcock, 1997). This behaviour allows small spat to recruit from their primary settlement sites into adult mytilid beds (Le Corre et al., 2013). This two-stage process has important evolutionary advantages to spat, namely avoid competition and cannibalism by adult mytilids (Porri et al., 2008). However, this stage is problematic for mytilid farmers that expect to maximize the retention of spat on farming structures despite tendency toward natural losses (Skelton and Jeffs, 2021). The operations of mitilid cultivation have traditionally been located in areas where they are readily available each year, since there is not accurate knowledge on the settlement of mitilids (Curiel-Ramírez and Cáceres-Martínez, 2010).
In Chile, the mussel industry is the second largest production aquaculture activity, after salmon farming, where the main cultured species is the Chilean mussel (Mytilus chilensis) (Cursach et al., 2011). Other mitilids of commercial interest extracted in Chile are the Shoe woolly (Choromytilus chorus), Cholga (Aulacomya atra) and Araucano woolly (Mytilus galloprovincialis) (López et al., 2008). The growing demand has made insufficient the quantity of seeds obtained to the market, and the use of technology is urgently required to improve the catch processes and optimize resources, aimed at supplying seeds to producers (Díaz, 2010). The technology that has been firstly developed are specialized culture ropes, made of synthetic filaments and natural fibers, which improve the fixation and settlement of mitilid larvae (Carl et al., 2012a). Unfortunately, the seed shortage of M. chilensis (the main production resource of Chilean mussel farming) occurred in 2013, of which it had consequences in the production of that year, with up to 60% less seed production. At the same time, there is a gap of information regarding the ideal conditions and seasons to install collectors. This gap is mostly due to the fact that the operations of cultivation of mitilids worldwide have traditionally been located in areas where these individuals are easily recruited each year (Curiel-Ramírez and Cáceres-Martínez, 2010). Our main aim was to identify the most suitable substrates for spat collectors of commercial mussel species in Chile. Namely, we used three settlement technologies for mitilid larvae, throughout one year in two coastal localities with contrasting hydrodynamic conditions at different depths.
Material and methods
Study area and experimental design
The spat collector was installed in the Biobío region (Central Chile), in two coastal bays, one protected (Coliumo 36°32’10.52’’S and 72°57’09.31’’O) and one exposed (Punta Lobería 37°09’01.52’’S and 73°34’07.84’’O) (Figure 1). At the time of the study, in Coliumo, the Pacific oyster (Crassostrea gigas) was cultivated with triploid hatchery seeds, harvesting 1 ton in 2014 (SERNAPESCA, 2014). In Punta Lobería, Chorito (M. chilensis) culture was cultivated, harvesting 16 tons in 2014 (SERNAPESCA, 2014). To avoid interference with the experiment at Punta Lobería, the location of the collectors was selected considering the ocean circulation and waves coming from the west to the east (Parada et al., 2001), with the cultivation center following the direction of the current, 0.5 km to the SE of the experiment.
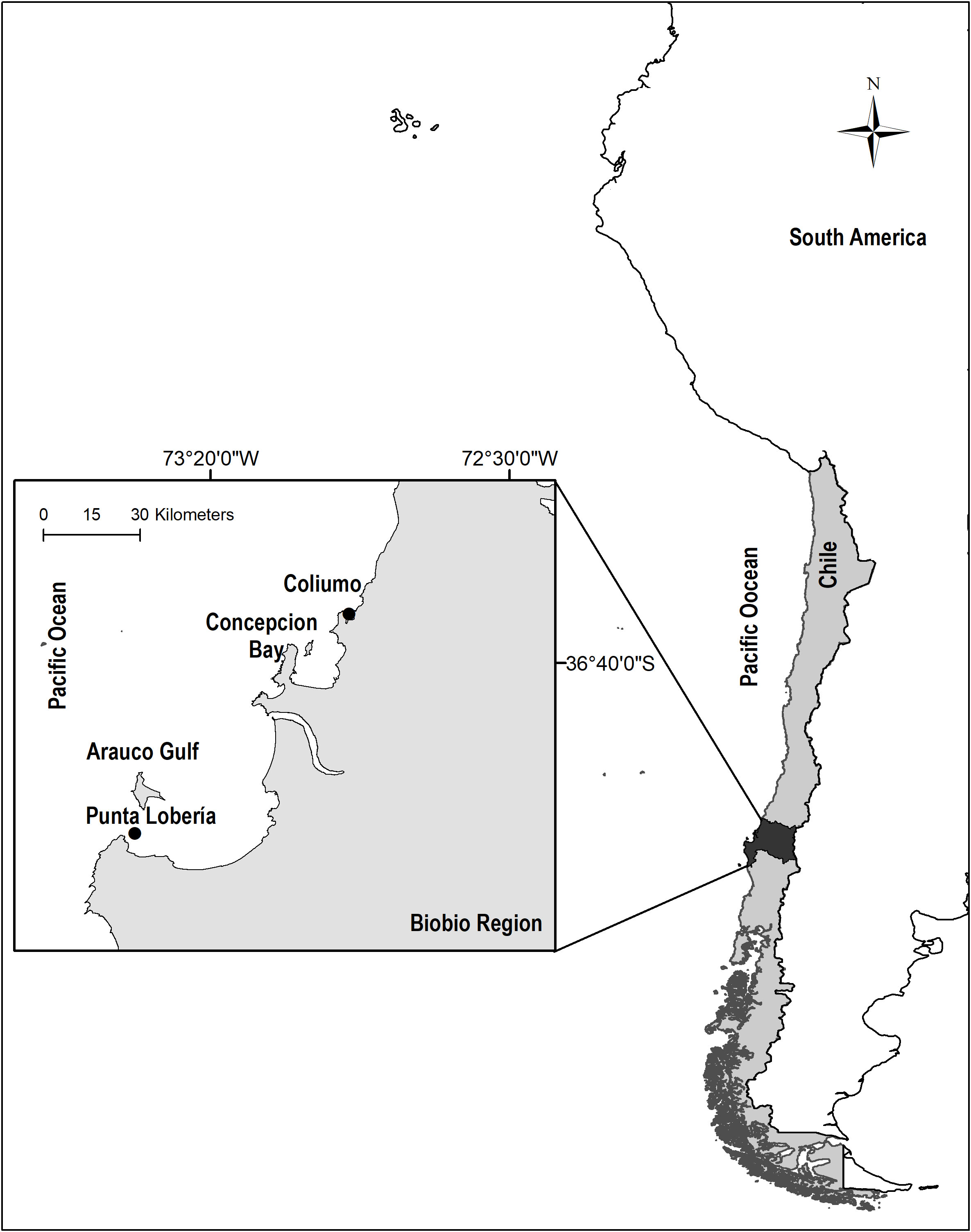
Figure 1 Study coastal zones in Biobío region, Chile.
At each locality, a longline system was installed, made up of a 15-m -long HDPE pipe (330 mm diameter), which served as a floating device for placing collector lines (Figure 2). A 22 mm diameter main line was installed, in which nine collector lines were placed (3 replicates per collector), made up of 1 m long PVC pipes (10 mm diameter), separated by 0.6 m from each other. The collectors were deployed at three depths (2, 4 and 6 m). Each line of collectors was stabilized by weights to maintain the verticality of the system. The longline was secured by two mooring ropes, which were held together with two 2,500 kg concrete blocks. The collectors dimension were variable, i.e. the anchovy nets were 20 x 20 cm (400 cm²), the ropes were 22 mm diameter and 20 cm long (152.05 cm²), the Raschel meshes were 20 x 20 cm (400 cm²) (see Figure 2 for details).
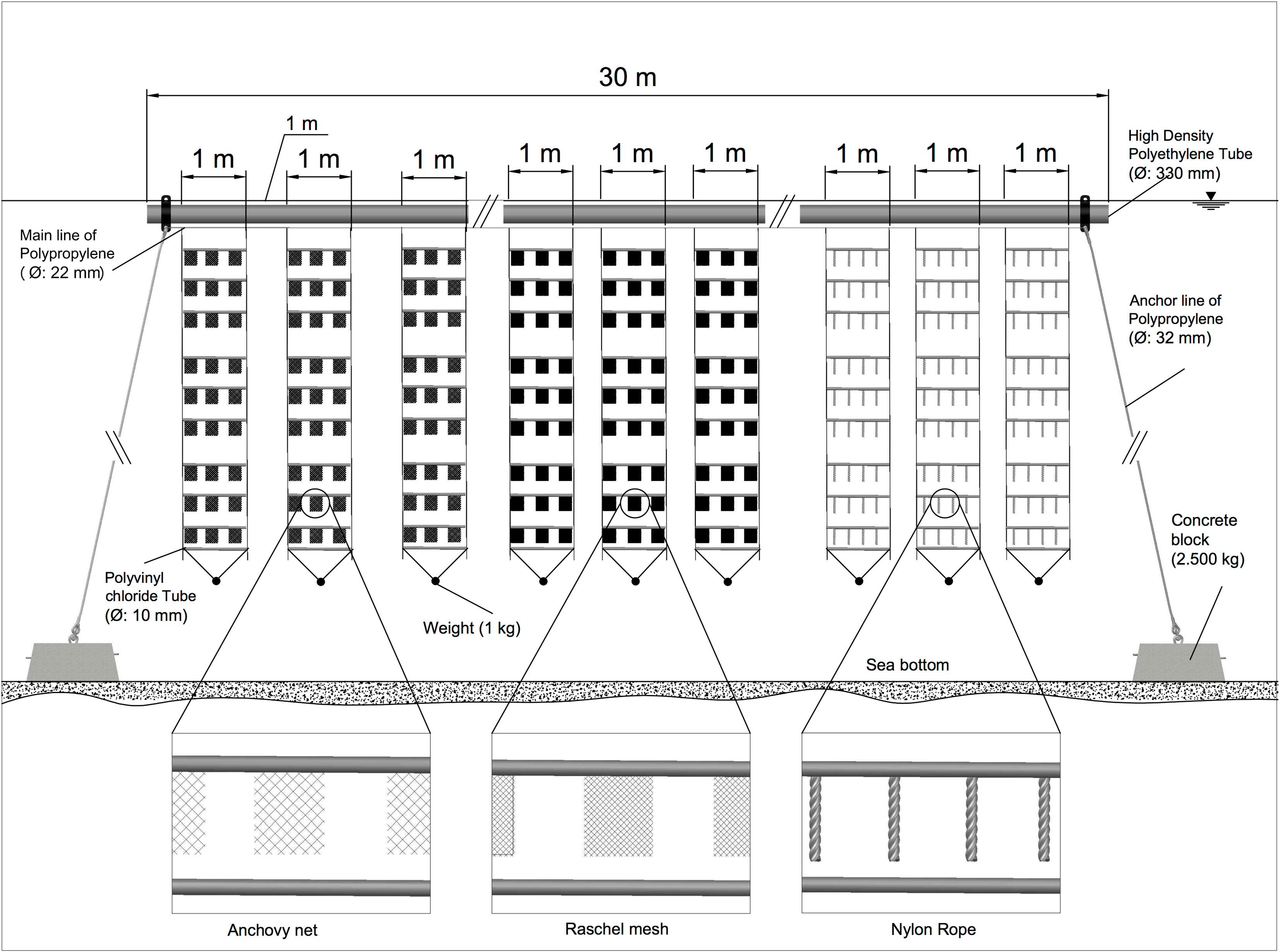
Figure 2 Longline system with different spat collectors.
Sampling processing
The spat collectors were deployed throughout 12 months (May 2014-May 2015), and sampling was carried out on a monthly basis at each season (winter, spring, summer and autumn). At each location, 243 collectors were installed, (81 collectors of the anchovy mesh type, 81 of the rope type and 81 of the Raschel mesh type). Twenty-seven collectors of each type were deployed at 2 m, 27 at 4 m and 27 at 6 m. For each stratum, it was divided into three replicates of 9 collectors each. Every 3 months, all collectors were released and replaced to avoid biofilm accumulation and fouling overlapping. Three collectors were randomly sampled, at each depth, collector type and replica (out of a total of 9 collectors), collecting a total of 54 collectors, composed of 9 anchovy meshes (one per each depth, 3 replicates), 9 nylon ropes (one per each depth, 3 replicates) and 9 raschel meshes (one per each depth, 3 replicates), for the two study sites. Later, the samples were taken to the laboratory, preserved in plastic bags and frozen, and transferred to the Ecohydraulic laboratory of the Universidad Católica de la Santísima Concepción (UCSC-Chile).
For sample preparation, the collectors were first cleaned and mitilid postlarvae and seeds were collected. The collectors, the postlarvae and seeds, were separated, using six sieves, 200, 300, 500, 750, 2000 µm and >2000 µm. Each sample was deployed in a tray, in 350 ml of 3% chlorine (250 ml solution of 10% sodium hypochlorite, diluted in 750 ml of water), for 10 minutes. The organic matter that was attached to the collectors was eliminated by oxidation, to get a “clean” sample. Subsequently, the sodium hypochlorite was drained from the beaker with the collector, by placing the couplets with the 200 and 750 µm sieves in a separate 1-l beaker, which served to obtain two types of sizes of individuals, partially isolated from the collector. Each collector was washed inside each 1-l beaker, which contained the collector that was previously separated from solution, by means of a water flow rate of 17 l min-1. This procedure was repeated four times. Subsequently, using a 500 ml tank with freshwater, the postlarvae from the coupler with the 200 µm sieve were introduced into the 60 mm Petri dish by means of a water flow, and the postlarvae and seeds from the coupler with the sieve of 750 µm in the 120 mm Petri dish. The postlarvae larger than 1 or 2 mm, and the seeds, found on the 750 µm sieve, were separated and placed in the 30 mm Petri dish.
Postlarvae and seeds were identified to species, using a Diprolab magnifying glass, at 1.5x magnification, and then extracted by using a disposable 5 ml pipette at a 30 ml vial with 70% alcohol. Visual identification was carried out following Ramorino and Campos (1983) for the postlarvae and seeds of the commercial mitilids. After preserving the samples in 30 ml vials with 70% alcohol, they were extracted for measurements by species, carried out using a Diprolab microscope and a computer with Motic Images Plus 2.0 software, to measure postlarvae.
Average density was measured as ind. 400 cm², similar to Çelik et al. (2016), which is a unit of measurement equivalent to the calculation of the standard area of 1 m of collector, 20 cm wide on both sides, giving a total area of 400 cm² for anchovy net and raschel mesh. A total of 131.94 cm² was measured for nylon rope.
Data analysis
A multivariate analysis was performed on the spat density at each site throughout the year to explore the environmental factors that influence the spat settlement. BIOENV analysis calculates the Spearman rank correlation coefficient (rho) between the similarity matrix and corresponding environmental data. Namely, it was performed to analyze the relationship between larval density and environmental variables: sea temperature (°C), chlorophyll a (mgm-3), dissolved oxygen (ppm), salinity (PSU), surface of collector (cm2) and depth (m). Temperature, chlorophyll a and salinity were log-transformed and dissolved oxygen was square-root transformed. A total of 4,999 permutations of the raw data were used to build the null hypotheses of no correlation between biological and environmental variables.
The aforementioned environmental variables were measured once monthly at the three depths (2, 4 and 6 m). Sea temperature was taken using Hobo thermometers, where every 30 minutes a data was taken throughout the study year. Dissolved oxygen and average salinity were measured using Hanna 9828 multi-parameter equipment, with a periodicity of 30 days for three months during each season of the year.
For each site, the null hypothesis that there are no differences among the treatments was tested using a three-factor orthogonal test of analysis of variance (season: fixed, four-level, month: fixed, three-level nested in season, depth: fixed with three levels and collector: fixed with three levels). Both sites were compared using a four-factor orthogonal test of analysis of variance, including “site” as a fixed factor with two levels (Coliumo and Punta Lobería). The hypothesis was evaluated with respect to the abundances of each mitilid species. Data were square-root transformed for the analysis of the dissimilarity indices. Subsequently, the transformed matrix was converted into a dissimilarity matrix using the Bray-Curtis index (Clarke, 1993). This matrix was used to perform the multivariate analysis of permutation of variance (PERMANOVA) (Anderson et al., 2008) using the abovementioned factors. The probabilities associated at each F-ratio were obtained with 9,999 permutations of residuals under a reduced model. These analysis were conducted using PRIMERv7 software (Clarke and Gorley, 2015).
Results
Environment variables
The lowest sea surface temperatures were reported in autumn at both sites (11°C) and the maximum ones in summer (20°C in Coliumo Bay and 18°C in Punta Lobería). The chlorophyll a peaked in winter in Coliumo Bay and in summer in Punta Lobería, both sites at a mean depth of 4 m, whilst the minimum concentrations were reported in autumn at both sites. Salinity remained constant during spring, summer and autumn, with an average value of 34 psu at both sites. The lowest salinity was reported in winter (24 psu) in the two sampling locations. Dissolved oxygen showed temporal variations throughout the study year, with the lowest concentrations during summer season (Figure 3).
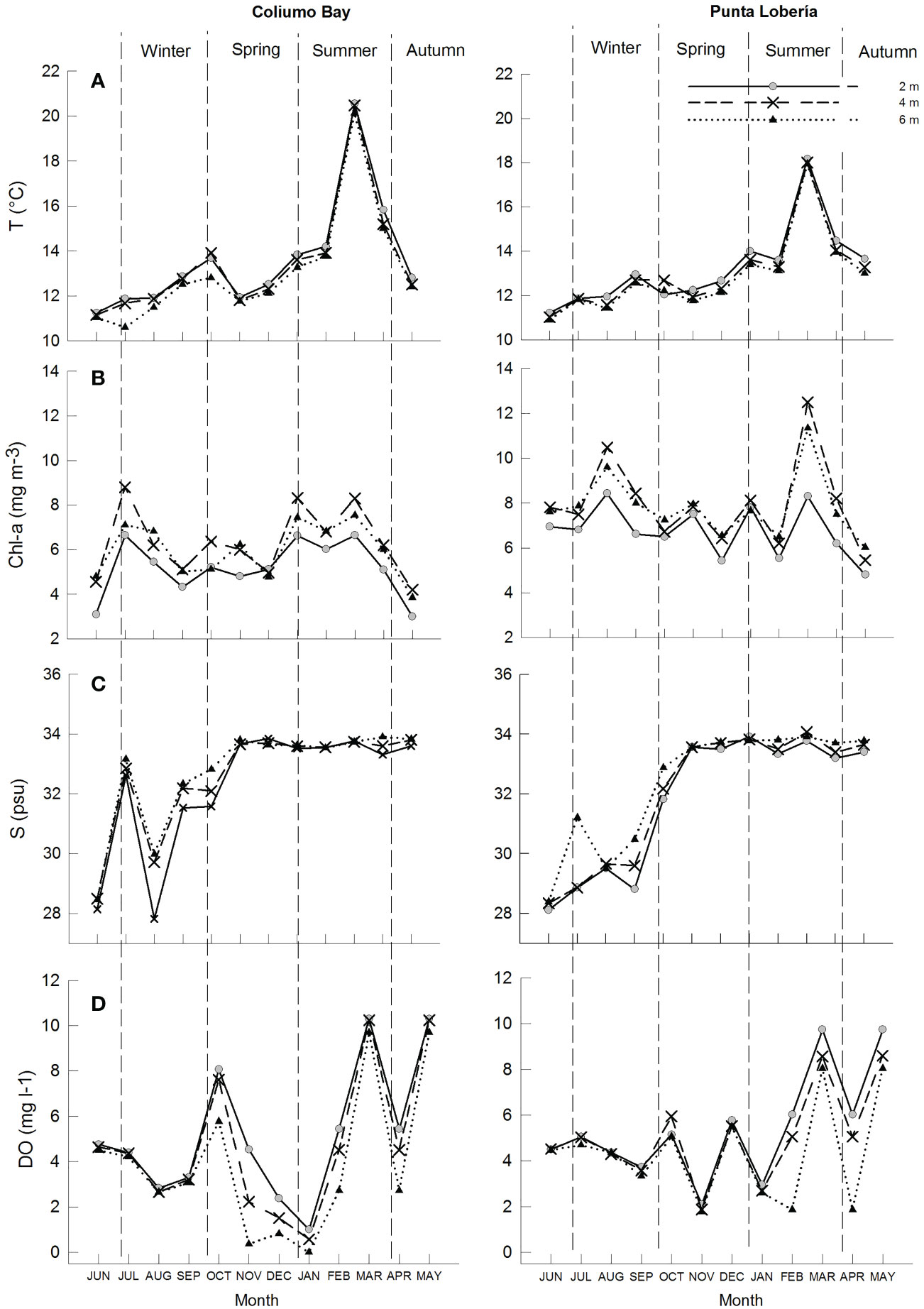
Figure 3 Monthly environmental conditions (A) temperature (T(°C)), (B) chlorophyll a (Chl-a (mg m−3)), (C) salinity (S(psu)) and (D) dissolved oxygen (DO(mg l-1)), by month at 2, 4 and 6 m depth in Coliumo Bay and Punta Lobería.
The spat densities in the collectors showed no significant correlations with the above environmental variables throughout the study period as a whole at both sites, being the temperature and the dissolved oxygen the best related to the spat densities (Rho=0.174, p=0.13). However, if data are analyzed separated by season for each site, different trends are observed. Namely, in Coliumo Bay dissolved oxygen is significantly correlated to spat densities during winter ((Rho=0.76, p<0.01), Chlorophyll a concentration showed no correlation during spring (Rho=0.43, p=0.09), temperature showed significant correlations with spat densities during summer season (Rho=0.78, p<0.01), and during autumn, temperature with salinity (Rho=0.94, p<0.001). In Punta Lobería, the spat densities showed significant trends with environmental variables depending on the season. Salinity and dissolved oxygen showed a significant trend during spring (Rho=0.62, p<0.05) and autumn (Rho=0.8, p<0.05). Chlorophyll a showed significant correlations with spat densities during winter (Rho=0.734, p<0.05).
Spat species
The settled spat species were A. atra, C. chorus, M. chilensis, M. galloprovincialis and S. algosus (see Table 1). The most abundant species was A. atra in Coliumo Bay and Punta Lobería, with peaks during summer (30.39 and 11.97 ind cm-2, respectively). In contrast, M. chilensis showed the minimum abundances (< 5 ind cm-2) at both coastal sites during winter season.
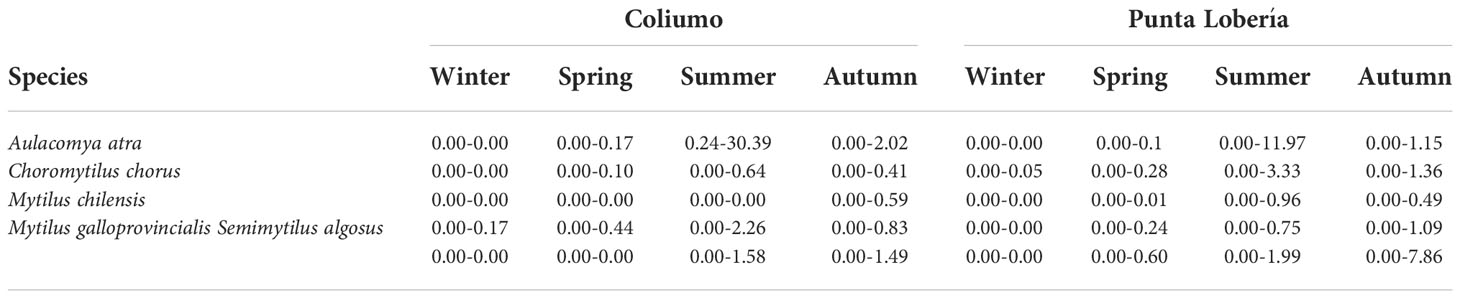
Table 1 Density ranges (ind cm-2) of spat species settled in collectors at both sites throughout the four seasons of the study year.
In Coliumo Bay, PERMANOVA analyses showed a significant second order interaction between Season, Collector and Depth, indicating that patterns of recruitment of mussels across different collectors depended on the season and also the depth (Table 2A). On the other hand, in Punta Lobería, first order interactions involving the nested factor month were significant, indicating that patterns of recruitment between collectors and season varied from month to month (Table 2B). Significant temporal variation due to month was not surprising since it was expected that as the time passed, the number of recruits would have increased. Despite this temporal variation at the month scale, PERMANOVA analyses also showed a significant interaction between Season and Collectors, indicating that differences in collectors were mainly due to the season the collectors were deployed (Table 2B). Despite these differences, multivariate ordinations (PCO) showed similar patterns in terms of seasonal differences (Figure 4). In both locations, all seasons clearly showed differences among them. Furthermore, in both sites differences between collectors and depths were considerably larger during summer and autumn, compared to spring and winter (Figure 4). Seasonal differences showed different recruitment patterns regarding the spat species (Figure 5). Despite clear differences between locations, it can be clearly noted that recruitment rates were non-existent or minimal in both locations during winter and spring (Figure 5). It also can be noted that M. chilensis and S. algosus typically recruit during summer and autumn, with a high correlated pattern of temporal and spatial distribution at both locations (Figure 4) whereas the other three recruit from spring onward (Figure 5). Depending on the location, recruitment of M. galloprovincialis (Coliumo Bay) and C. chorus (Punta Loberia) were observed as early as the latest months of winter, however their densities were considerably lower than those observed during summer and autumn (Figure 5). Highest recruitment rates were observed for A. atra (Coliumo Bay) and S. algosus (Punta Lobería) during summer and autumn, respectively (Figure 5).
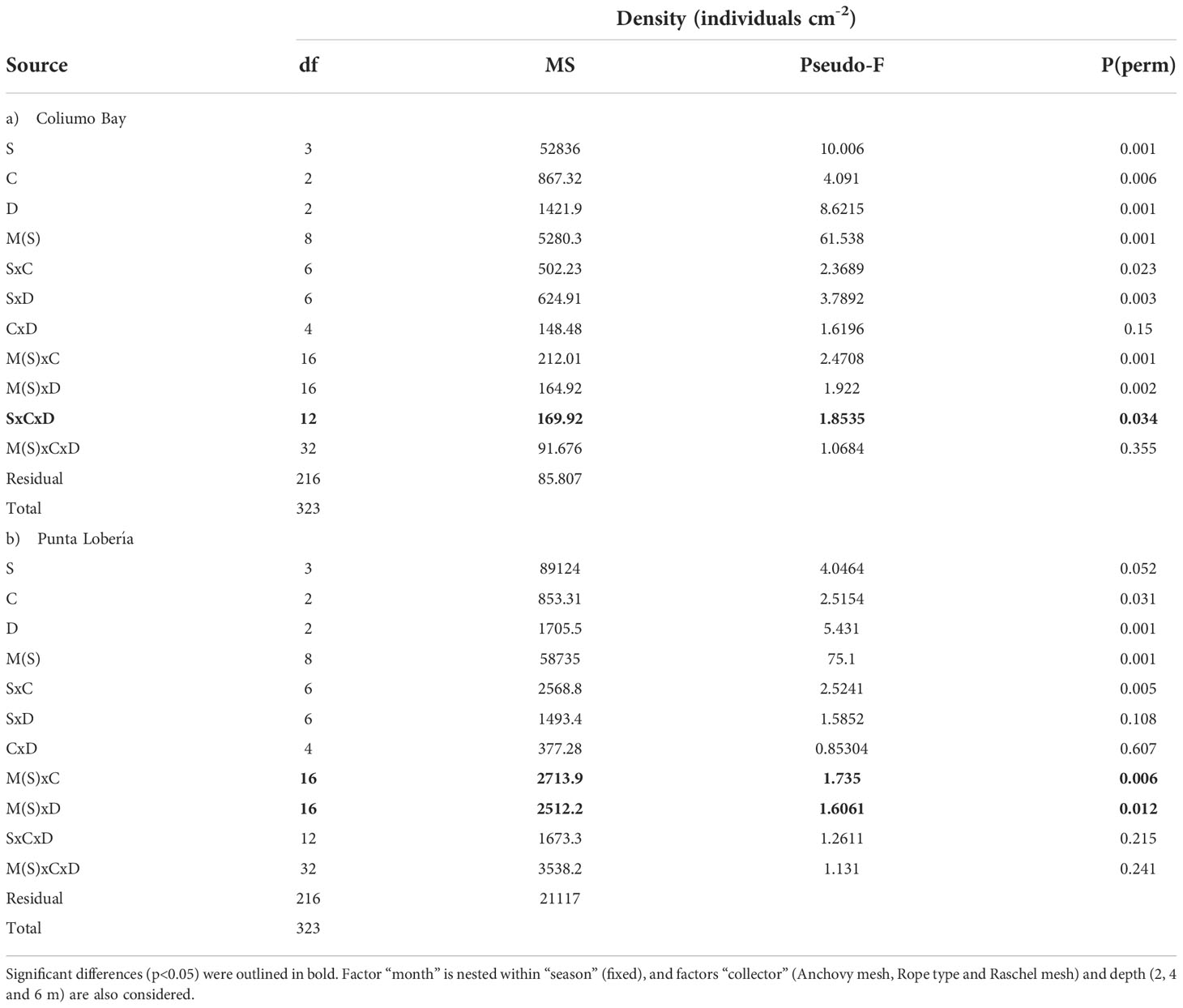
Table 2 PERMANOVA analyses regarding the spat density at both sites a) Coliumo Bay, b) Punta Lobería.
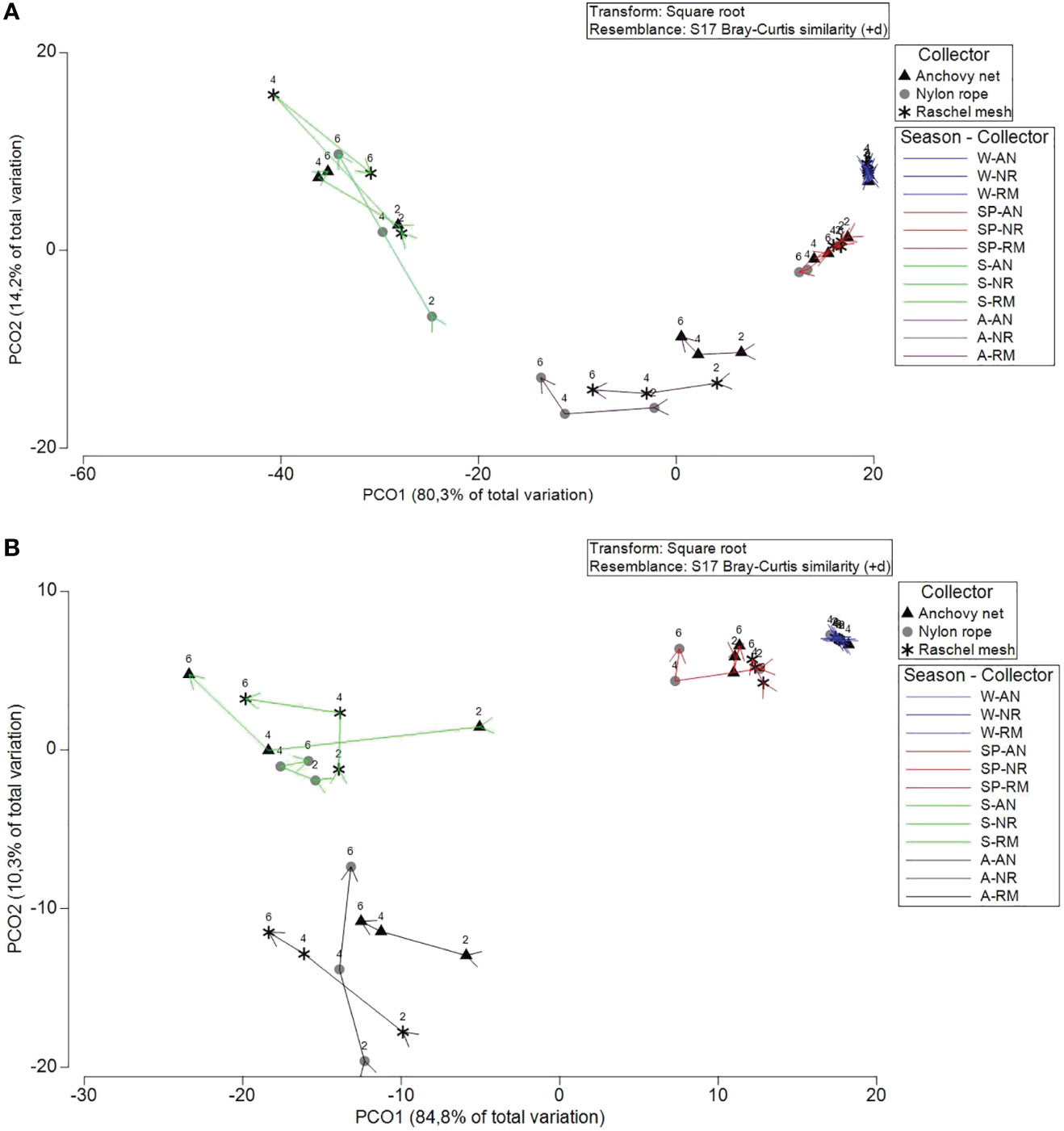
Figure 4 Principal coordinates analysis (PCO) from Euclidean distances among centroids per season (winter=W, spring=Sp, summer=S, Autumn=A), collector (anchovy net=AN, nylon rope=NP, raschel mesh = RM), at 2, 4 and 6 m depth for (A) Coliumo Bay and (B) Punta Lobería.
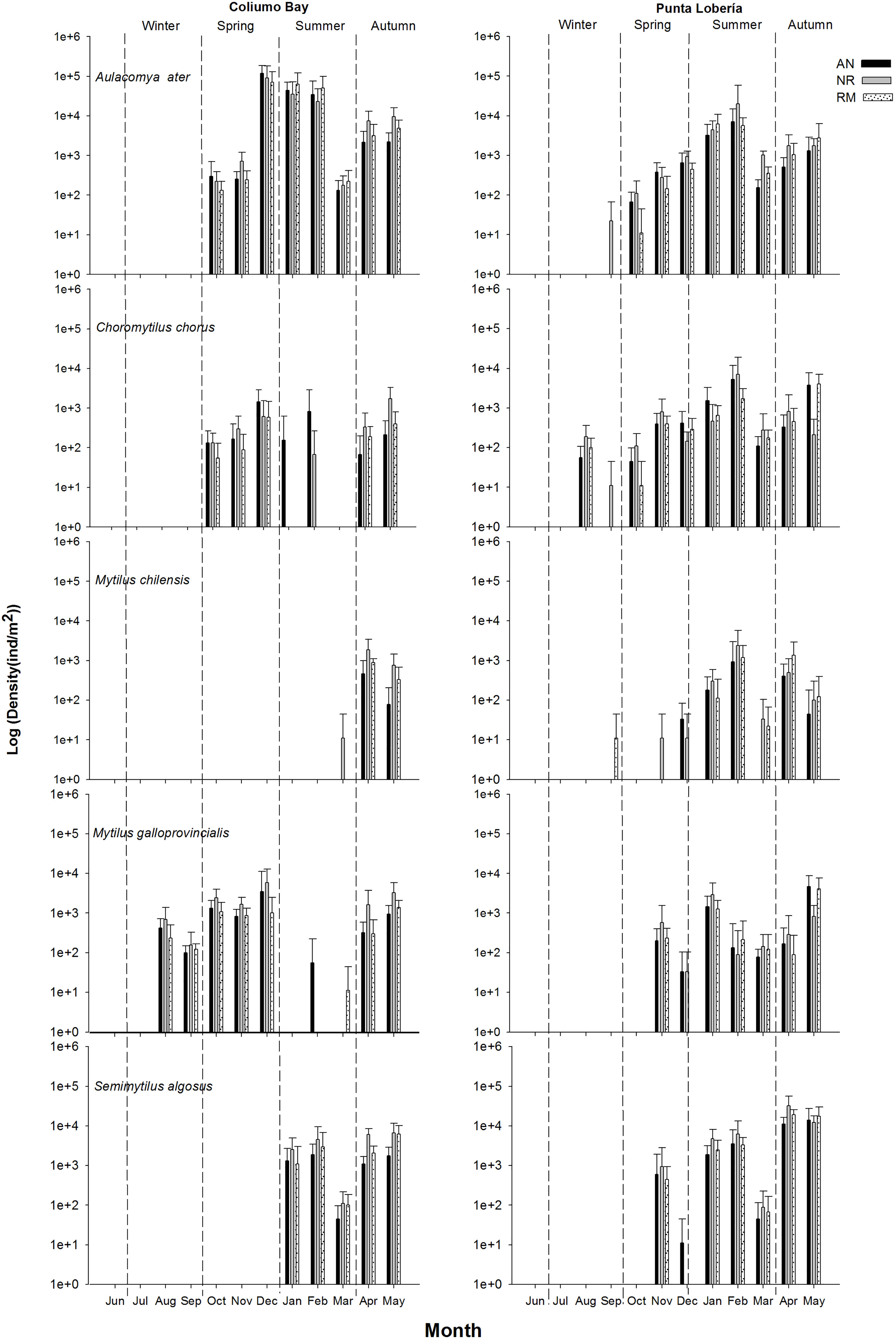
Figure 5 Spat density at logarithmic scale for each species at both coastal sites (AN, anchovy net; NR, nylon rope; RM, raschel mesh). Error bars indicate + 1 standard error.
Differences due to collector type and depth were considerably smaller when compared to seasonality (Figure 4). Consequently, differences between collectors and depth were examined per season (only summer and autumn when densities were higher) and site. Multivariate ordinations (PCO) showed that differences between collectors and depths were significant during the autumn season, but during summer those differences were not obvious (Supplementary Figure 1 and Supplementary Figure 2). These differences were mostly due to A. atra and S. algosus which were more abundant in nylon rope than the other two collectors.
Discussion
In the present study, five bivalves species settled in the collectors throughout the experiment. Four of these are native, i.e. A. atra, C. chorus, M. chilensis, and S. algosus (Uriarte, 2008), and one introduced M. galloprovincialis (Larraín et al., 2018), which is only allowed its experimental cultivation in Benthic Resource Management and Exploitation Areas (AMERB, in Spanish) off the coast of Chile (Decree n° 95/2015). Of the native species, only S. algosus is not of commercial interest, and M. chilensis is the main species of commercial interest in the study area (Kamermans and Capelle, 2019). In the last three decades, mytilid production has been steadily increasing in Chile, with a current production of over 330,000 tonnes yearly (Gonzalez-Poblete et al., 2018). In Chile, the seeding of spat is mainly carried out by organizations of artisanal fishermen within their AMERBs and by independent fishermen and aquaculture farmers who have transitory permits in their aquaculture concessions (Contreras and Godoy, 2021). One of the main bottlenecks of the mitilid aquaculture industry, as occurs in other regions (Skelton and Jeffs, 2021) is the dependence of wild spat. Spat settlement greatly depends on the site and depth, regardless of the collector. Seasonal variability in spat settlement was high throughout the study period, with a higher settlement during summer and autumn relative to winter and spring. These seasonal variations were more pronounced during summer in M. galloprovincialis in Coliumo and C. chorus in both sites. M. chilensis and S. algosus showed a higher settlement in autumn in Coliumo and in summer in Punta Lobería. A. altra and C. chorus showed the highest settlement in summer at both sites. These results are in accordance with the spawning and settlement of mitilids, since temperature is the key driver underpinning these processes (South et al., 2021). Spawning seasons are spring and summer, with a prolonged larval development up to 45 days (Karayücel et al., 2003; Barria et al., 2012; Çelik et al., 2016).
Mytilus chilensis is the most important commercial species in the study region, concentrating over 90% of the overall production in Chile (Molinet et al., 2015). This species showed the highest abundances during autumn at both sites, being negligible the remaining seasons of the year. The settlement patterns of this species are similar to those observed in A. atra, which peaked during summer in Punta Lobería. Zaixso (2004) also observed high abundances of A. atra during summer season in mixed substrates, i.e. rocky-sandy seabeds, from natural banks at San José Gulf (Argentina). A previous study conducted in one of the study sites (Punta Lobería), the settlement of A. atra was observed during spring and summer (Leiva et al., 2007). These results might be explained by the ENSO event that occurred during autumn on the Biobío coasts (central Chile) (CIIFEN, 2017). In terms of depth, this species showed the highest abundances in the deepest level (6 m) throughout the study period, similar to the findings of Ojeda et al. (2014) on spatial distribution patterns of intertidal mollusks from Chile.
Mytilus galloprovincialis showed high abundances in autumn and spring, being almost absent during winter. This species peaked at 2 m deep during autumn, and during winter in Mexico (Curiel-Ramírez and Cáceres-Martínez, 2010), with similar temperatures at both seasons. This suggests a dependence of the spat of this species to sea temperature. The species S. algosus, though with no commercial interest, is one the most abundant myitilids on Chilean coasts; it showed the highest abundances in autumn at both sites. These results are in accordance with a previous study (Belapatiño, 2007) carried out in Peru throughout 8 years (1996-2003). This species has been reported to spawn throughout the year, peaking at 1 m depth in June and July (González et al., 1980), however, no recruitment was observed during the study period at both coastal sites. The highest abundances of S. algosus were observed at 2 m depth in the nylon rope collector. The remaining two mitilid species collected in the collectors throughout the study period were A. atra and C. chorus, that showed peaks during the summer season and also the latter a second peak of abundances in late spring and autumn in one of the study sites (Coliumo). Avendaño and Cantilláne (2011) observed a displacement of A. atra to deeper sites when interacting this species with C. chorus, as it was observed here in Punta Lobería during summer. Winter conditions are not favourable for the settlement of spat regardless of the mitilid species in the study region.
A plethora of studies have been conducted regarding spat settlement, with a wide range of variables involved to identify the most efficient combination of collectors (material, type, etc.) and environmental variables (depth, temperature, etc.) to ensure detachment of spat (see Yildiz and Berber, 2010; Molinet et al., 2017; Molinet et al., 2021; Skelton and Jeffs, 2021; Weston et al., 2021). Also it needs to be taken into consideration that spat settlement is a process that may be affected by multiple factors such as, larval predation or competition for source from mussel crop (Woods et al., 2012; Atalah et al., 2020); water quality (Boukadida et al., 2016) or combination of environmental disturbances (Delorme et al., 2020). Mussel farming is facing challenges due to a substantial decline in spatfall in the last decade in certain regions (Atalah and Forrest, 2019). In New Zealand, the causes of this spat decrease are unknown, but it has been suggested that a widespread depletion of wild parental beds might be a main driver of this trend, since a 97% decline has been observed since the late 1960s due to industrial dredging (Handley, 2015; Toone et al., 2022). This depletion was concomitant with the initial development of the mussel aquaculture industry (Jeffs et al., 1999).
The present findings showed the importance of preserving natural stocks of mitilids (“Parent areas’’) in the study area. As observed in the present study, spat recruitment is greatly conditioned by seasonal variations of water temperature, and spat did not show clear preference on the material that the collector was made of; yet, depth was an important factor to consider. This study constitutes a first step to shed light on the spatial and seasonal spat dynamics that are essential to develop sustainable management of mitilid culture in Chile.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
CD, CS contributed to conception and design of the study, funding acquisition, project administration, resource supervision. CD, CS and JJ contributed to statistical and formal analyses and investigation. CS and RR contributed writing of the original draft and review and editing of the manuscript. All authors contributed to the article and approved the submitted version.
Funding
Centro de Investigación en Biodiversidad y Ambientes Sustentables (CIBAS), Dirección de Investigación of the Universidad Católica de la Santísima Concepción, and for business innovation from Corfo Innova Chile n° 13IDL2-23315.
Acknowledgements
The authors are especially grateful with the leaders of the Llico BRMA fishing organization and to the Cultivos Pacfish S.A, Foodcorp S.A. and Acuimarc S.A. companies, for making available the instalations of the farming center at Punta Loberiá.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2022.1068305/full#supplementary-material
References
Anderson M., Gorley R. N., Clarke R. K. (2008). “PERMANOVA+ for PRIMER: Guide to software and statistical methods”. In Primer-E (Plymouth: PRIMER-E)
Atalah J., Fletcher L. M., Davidson I. C., South P. M., Forrest B. M. (2020). Artificial habitat and biofouling species distributions in an aquaculture seascape. Aquac. Environ. Interact. 12, 495–509. doi: 10.3354/aei00380
Atalah J., Forrest B. M. (2019). Forecasting mussel settlement using historical data and boosted regression trees. Aquac. Environ. Interact. 11, 625–638. doi: 10.3354/aei00337
Avdelas L., Avdic-Mravlje E., Borges A. C., Cano S., Capelle J. J., Carvalho N., et al. (2021). The decline of mussel aquaculture in the European union: causes, economic impacts and opportunities. Rev. Aquac. 13, 91–118. doi: 10.1111/raq.12465
Avendaño M., Cantilláne M. (2011). Restablecimiento de Choromytilus chorus (Molina 1782) (Bivalvia mytilidae) en el norte de Chile. Lat. Am. J. Aquat. Res. 39, 390–396. doi: 10.3856/vol39-issue2-fiilltext-20
Avendaño-Herrera R., Riquelme C., Silva. F. (2002). Utilización de biopelículas bacterianas en el asentamiento de larvas de Argopecten purpuratus (Lamarck 1819) en un hatchery comercial. Rev. Biol. Mar. Oceanogr. 37 (1), 35–41. doi: 10.4067/S0718-19572002000100006
Barria A., Gebauer P., Molinet C. (2012). Variabilidad especial y temporal del suminstro larval de mitílidos en el seno de reloncavía, sur de Chile. Rev. Biol. Mar. Oceanogr. 47, 475–487. doi: 10.4067/S0718-19572012000300009
Belapatiño A. (2007) Variabilidad del asentamiento larval de mitílidos en bahía independencia, pisco – perú, durante el periodo 1996-2003. tesis para optar el título profesional de biólogo con mención en hidrobiología y pesquería. Lima, universidad nacional mayor de San marcos. Available at: https://cybertesis.unmsm.edu.pe/handle/20.500.12672/920 (Accessed October 2021).
Boukadida K., Banni M., Gourves P. Y., Cachot J. (2016). High sensitivity of embryo-larval stage of the Mediterranean mussel, Mytilus galloprovincialis to metal pollution in combination with temperature increase. Mar. Environ. Res. 122, 59–66. doi: 10.1016/j.marenvres.2016.09.007
Buchanan S., Babcock R. (1997). Primary and secondary settlement by the greenshell mussel Perna canalicullus. Oceanogr. Lit. Rev. 12, 1500.
Carl C., Poole A. J., Sexton B. A., Glenn F. L., Vucko M. J., Williams, et al. (2012b). Enhancing the settlement and attachment strength of pediveligers of Mytilus galloprovincialis bychanging surface wettability and microtopography. Biofouling 28, 175–186. doi: 10.1080/08927014.2012.662676
Carl C., Poole A. J., Williams M. R., de Nys R. (2012a). Where to settle–settlement preferences of Mytilus galloprovincialis and choice of habitat at a micro spatial scale. PloS One 7 (12), e52358. doi: 10.1371/journal.pone.0052358
Çelik M. Y., Karayücel S., Karayücel I., Eyüboğlu B., Öztürk R. (2016). Settlement and growth of the mussels (Mytilus galloprovincialis, lamarck 1819) on different collectors suspended from an offshore submerged longline system in the black Sea. Aquac. Res. 47, 3765–3776. doi: 10.1111/are.12827
CIIFEN (2017) El Niño 2015-2016: evolución, vulnerabilidad e impactos en latinoamérica. Available at: http://181.198.20.220/documents/1150#more (Accessed April 1, 2021).
Clarke K. R. (1993). Non-paramteric multivariate analyses of changes in community structure. Aust. J. Ecol. 18, 117–143. doi: 10.1111/j.1442-9993.1993.tb00438.x
Contreras J., Godoy C. (2021) Manual de procedimientos y buenas prácticas para la captación de semillas de chorito (Mytilus chilensis) en Áreas de manejo y explotación de recursos bentónicos. fortalecimiento de la capacidad de adaptación en el sector pesquero y acuícola chileno al cambio climático. Santiago de Chile. Available at: https://www.subpesca.cl/portal/617/articles-97384_manual_procedimientos_choritos.pdf (Accessed January 28, 2022).
Curiel-Ramírez S., Cáceres-Martínez J. (2010). Settlement of Mytilus galloprovincialis on collectors suspended at different depths in bahía de todos santos b. c., Mexico. Aquac 300, 102–106. doi: 10.1016/j.aquaculture.2009.12.019
Cursach J. A., Suazo C. G., Rau J. R., Tobar C. N., Gantz A. (2011). Ensamble de aves en una mitilicultura de chiloé, sur de Chile. Rev. Biol. Mar. Oceanogr. 46, 243–247. doi: 10.4067/S0718-19572011000200013
Delorme N. J., Biessy L., South P. M., Zamora L. N., Ragg N. L. C., Burritt D. J. (2020). Stress-on-stress responses of a marine mussel, Perna canaliculus: food limitation reduces the ability to cope with heat stress in juveniles. Mar. Ecol. Prog. Ser. 644, 105–117. doi: 10.3354/meps13375
Díaz E. (2010). Mitílidos en la región de los Lagos, condiciones de trabajo en la industria del chorito (Santiago de Chile, Chile. Andros Impresores: Departamento de estudios de la Direcció del Trabajo). Available at: http://www.dt.gob.cl/documentacion/1612/articles-98903_recurso_1.pdf (Accessed June 10, 2022).
FAO. (2021). FAO yearbook. fishery and aquaculture statistics 2019/FAO annuaire. Available at: https://doi.org/10.4060/cb7874t.
FAO. (2022). The state of world fisheries and aquaculture 2022. towards blue transformation. Rome, FAO. Available at: https://doi.org/10.4060/cc0461en.
Filgueira R., Peteiro L. G., Labarta U., Fernández-Reiriz M. J. (2007). Assessment of spat collector ropes in Galician mussel farming. Aquac. Eng. 37, 195–201. doi: 10.1016/j.aquaeng.2007.06.001
Ganesan A. M., Alfaro. A. C., Brooks J. D., Higgins C. M. (2010). The role of bacterial biofilms and exudates in the settlement of mussel (Perna canaliculus) larvae. Aquac 306, 1–4. doi: 10.1016/j.aquaculture.2010.05.007
Gonzalez-Poblete E., Hurtado C. F., Rojo C., Norambuena R. (2018). Blue mussel aquaculture in Chile: Small or large scale industry? Aquac 493, 113–122. doi: 10.1016/j.aquaculture.2018.04.026
González J., Villagra J., Hinojosa M., Becerra. R. (1980). Potencialidad del cultivo en balsa de Semimytilus algosus (Gould) 1850, en la bahía de concepción (Mollusca, Bivalvia, Mytilidae). Bol. Inst. Paul. 29, 187–189. doi: 10.1590/S0373-55241980000200038
Handley S. (2015) The history of benthic change in pelorus sound (Te hoiere) marlborough. nelson. national institute of water and atmospheric research. Available at: https://envirolink.govt.nz/assets/Envirolink/1520-MLDC97-The-history-of-benthic-change-in-Pelorus-Sound-Te-Hoiere-Marlborough.pdf (Accessed May 23, 2022).
Jeffs A. G., Holland R. C., Hooker S. H., Hayden B. J. (1999). Overview and bibliography of research on the greenshell mussel, Perna canaliculus, from new Zealand waters. J. Shellfish. Res. 18, 347–360.
Kamermans P., Capelle J. J. (2019). “Provisioning of mussel seed and its efficient use in culture,” in Goods and services of marine bivalves. Eds. Smaal A., Ferreira J., Grant J., Peterson J., Strands Ø. (Cham: Springer), pp 27–pp 49. doi: 10.1007/978-3-319-96776-9_3
Karayücel S., Karayücel I., Erdem M., Saygun S., Uyan O. (2003). Growth and production in long-line cultivated Mediterranean mussel (Mytilus galloprovincialis) in sinop, black Sea. Isr. J. Aquac. 55, 169–178. Available at: https://evols.library.manoa.hawaii.edu/server/api/core/bitstreams/843e69b5-d283-4f69-8da3-a22028008efb/content
Larraín M. A., Zbawicka M., Araneda C., Gardner J. P. A., Wenne R. (2018). Native and invasive taxa on the pacific coast of south America: Impacts on aquaculture, traceability and biodiversity of blue mussels (Mytilus spp.). Evol. Appl. 11, 298–311. doi: 10.1111/eva.12553
Le Corre N., Martel A. L., Guichard F., Johnson L. E. (2013). Variation in recruitment: differentiating the roles of primary and secondary settlement of blue mussels Mytilus spp. Mar. Ecol. Prog. Ser. 481, 133–146. doi: 10.3354/meps10216
Leiva G., Santibáñez C., Bartheld J., Molinet C., Navarro J. (2007) Definición de criterios biológicos, ambientales, sanitarios y operativos para la instalación de colectores de moluscos bivalvos en la X región. litoral austral ltda. informe final proyecto del fondo de investigación pesquera (FIP) (Chile: Subsecreataría de Pesca y Acuicultura). Available at: https://www.subpesca.cl/fipa/613/articles-89100_informe_final.pdf (Accessed May 23, 2022).
López D. A., López B. A., González M. L. (2008). Shellfish culture in Chile. IJEP 33 (4), 401–431. doi: 10.1504/IJEP.2008.02057
Molinet C., Astorga M., Cares L., Díaz M., Hueicha K., Marín, et al. (2021). Vertical distribution patterns of larval supply and spatfall of three species of mytilidae in a Chilean fjord used for mussel farming: Insights for mussel spatfall efficiency. Aquac 535, 736341. doi: 10.1016/j.aquaculture.2021.736341
Molinet C. M., Díaz M. A., Arriagada C. B., Cares L. E., Marín S. L., Astorga M. P., et al. (2015). Spatial distribution pattern of Mytilus chilensis beds in the reloncaví fjord: hypothesis on associated processes. Rev. Chil. Hist. Nat. 88, 1–12. doi: 10.1186/S40693-015-0041-7
Molinet C., Díaz M., Marín S., Astorga M., Ojeda M., Cares L., et al. (2017). Relation of mussel spatfall on natural and artificial substrates: Analysis of ecological implications ensuring long-term success and sustainability for mussel farming. Aquac 20, 211–218. doi: 10.1016/j.aquaculture.2016.09.019
Ojeda J., Rosenfeld S., Marambio J., Rozzi R., Mansilla. A. (2014). Patrones estacionales y espaciales de la diversidad de moluscos intermareales de bahía róbalo, canal beagle, reserva de la biósfera cabo de hornos, Chile. Rev. Biol. Mar. Oceanogr. 49, 493–509. doi: 10.4067/S0718-19572014000300007
Parada C. E., Sobarzo M. A., Figueroa D., Castro L. (2001). Cirulación del golfo de arauco en un período de transición estacional: Un nuevo enfoque. Investig. Mar. 29, 11–23. doi: 10.4067/S0717-71782001000100002
Peteiro L. G., Filgueira R., Labarta U., Fernández-Reiriz M. (2010). The role of fish predation on recruitment of Mytilus galloprovincialis on different artificial mussel collectors. Aquac. Eng. 42, 25–30. doi: 10.1016/j.aquaeng.2009.09.003
Porri F., Jordaan T., McQuaid C. D. (2008). Does cannibalism of larvae by adults affect settlement and connectivity of mussel populations? Estuar. Coast. Shelf. Sci. 79, 687–693. doi: 10.1016/j.ecss.2008.06.010
Ramorino L., Campos B. (1983). Larvas y postlarvas de mytilidae de Chile (Mollusca: Bivalvia). Rev. Biol. Mar. 19 (2), 143–192. Available at: https://rbmo.uv.cl/resumenes/v432/432-327.pdf (Accessed October 12, 2022)
SERNAPESCA (2014) Anuario estadístico de pesca y acuicultura. desembarques y acuicultura. Available at: http://www.sernapesca.cl/informes-datos/resultado-estadisticas/1287?field_anho_tid=44 (Accessed November 8, 2022).
Skelton B. M., Jeffs A. G. (2021). The loss of spat seeding into coastal GreenshellTM mussel (Perna canaliculus) farms. Aquac 544, 737115. doi: 10.1016/j.aquaculture.2021.737115
South P. M., Delorme N. J., Skelton B. M., Floerl O., Jeffs A. G. (2021). The loss of seed mussels in longline aquaculture. Rev. Aquac. 14, 440–455. doi: 10.1111/raq.12608
Toone T. A., Benjamin E. D., Handley S., Jeffs A., Hillman J. R. (2022). Expansion of shellfish aquaculture has no impact on settlement rates. Aquacult. Environ. Interact. 14, 135–145. doi: 10.3354/aei00435
Uriarte I. (2008). “Estado actual del cultivo de moluscos bivalvos en chile. en a. lovatelli,” in Taller regional de la FAO sobre el estado actual del cultivo y manejo de moluscos bivalvos y su proyección futura: Factores que afectan su sustentabilidad en américa latina. actas de pesca de la FAO. no. 12. Eds. Farías A., Uriarte I. (Roma: FAO), 61–75.
Weston K., Jahangard S., Ingram B. A., Miller A. D., Jennings G., Sherman C. D. (2021). Factors affecting settlement, growth and metamorphosis of hatchery-produced Australian blue mussel larvae. Aquac. Int. 29 (5), 1963–1977. doi: 10.1007/s10499-021-00728-z
Wijsman J. W. M., Troost K., Fang J., Rocaranti J. (2019). “Global production of marine bivalves. trends and challenges,” in Goods and services of marine bivalves. Eds. Smaal A., Ferreira J., Grant J., Peterson J., Strands Ø. (Springer International Publishing), pp 7–pp26. doi: 10.1007/978-3-319-96776-9_2
Woods C. M. C., Floerl O., Hayden B. J. (2012). Biofouling on greenshell™ mussel (Perna canaliculus) farms: a preliminary assessment and potential implications for sustainable aquaculture practices. Aquac. Int. 20, 537–557. doi: 10.1007/s10499-011-9484-2
Yi-Feng L., Yu-Ru C., Jin-Long Y., Wei-Yang B., Xing-Pan G., Xiao, et al. (2014). Effects of substratum type on bacterial community structure in biofilms in relation to settlement of plantigrades of the mussel Mytilus coruscus. Int. Biodeterior. Biodegradation. 96, 41–49. doi: 10.1016/j.ibiod.2014.08.012
Yildiz H., Berber S. (2010). Depth and seasonal effects in the settlement density of Mytilus galloprovincialis l. 1819 in the dardanelles. J. Anim. Vet. Adv. 9, 756–759. doi: 10.3923/javaa.2010.756.759
Keywords: bivalve, mussel, aquaculture, postlarvae, settlement preferences, SE Pacific Ocean
Citation: Díaz C, Sobenes C, Cruz Motta JJ and Riera R (2022) Settlement experiment of spat using three different collectors in two contrasting coastal sites from Chile (SE Pacific Ocean). Front. Mar. Sci. 9:1068305. doi: 10.3389/fmars.2022.1068305
Received: 12 October 2022; Accepted: 22 November 2022;
Published: 08 December 2022.
Edited by:
Torstein Pedersen, UiT The Arctic University of Norway, NorwayReviewed by:
Domitilia Coutinha Matias, Instituto Português do Mar e Atmosfera, PortugalLiqiang Zhao, Guangdong Ocean University, China
Copyright © 2022 Díaz, Sobenes, Cruz Motta and Riera. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Christian Díaz, Y2hkaWF6QHVjc2MuY2w=
†These authors have contributed equally to this work and share first authorship