
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
BRIEF RESEARCH REPORT article
Front. Mar. Sci. , 11 October 2022
Sec. Marine Fisheries, Aquaculture and Living Resources
Volume 9 - 2022 | https://doi.org/10.3389/fmars.2022.1021046
This article is part of the Research Topic Current Knowledge and Recent Advances in Aquatic Animal Health Management View all 4 articles
 Eva Vallejos-Vidal1
Eva Vallejos-Vidal1 Ali Reza Khansari2
Ali Reza Khansari2 Mariana Teles2,3
Mariana Teles2,3 Sebastián Reyes-Cerpa4,5
Sebastián Reyes-Cerpa4,5 Juan Miguel Mancera6
Juan Miguel Mancera6 Lluis Tort2*
Lluis Tort2* Felipe E. Reyes-López7*
Felipe E. Reyes-López7*Cortisol carries out its physiological mechanism of action through the recognition by the mineralocorticoid receptor (MR) and the glucocorticoid receptor (GR) 1 (GR1) and GR2. Previous studies reported that the main difference between gr1 and gr2 nucleotide sequences resides in a 27-nucleotide insertion/deletion in the DNA-binding region, respectively. However, in gilthead sea bream (Sparus aurata) the annotation for gr1 and gr2 seems contradictory. The gr2 sequence possesses the characteristic 27-nucleotide insertion that, in fact, is associated with the gr1 nucleotide sequence. Thus, this study aimed to elucidate the nucleotide sequences for the gr1 and gr2 in gilthead sea bream. The Clustal Omega alignment for different fish species corroborated the presence of such 27-nucleotide insertion/deletion in the DNA-binding region for gr1 and gr2, respectively. Then, we design specific primers set for the amplification of the gilthead sea bream gr1 by polymerase chain reaction (PCR). Importantly, the gr1 nucleotide partial sequence has a high similarity with other gr1 sequences already published for other fish species, being present in all of them the 27-nucleotide insertion in the DNA-binding region. We also detected that in European sea bass the gr1 and gr2 sequences had not been named according to the 27-nucleotide insertion/deletion criteria in the DNA-binding region. Thus, our study makes an urgent call to the scientific community to discuss the establishment of an updated agreement that allows homogenizing the criteria for the nomenclature defining the gr1 and gr2 nucleotide sequences for a better understanding of the stress response in teleost fish species.
Stress is a situation provoked by exposure to a stressor in which the organism may suffer real or symbolic damage to its integrity (Tort, 2011). When a stressful situation has sufficient intensity and duration, it causes the activation of a coordinated system at the physiological networked level (Balasch and Tort, 2019). The HPI axis (Hypothalamus-Pituitary-Interrenal axis) is responsible for producing cortisol by the interrenal cells. In teleosts, cortisol is the main corticosteroid and plays an important role in physiology including growth, immunoregulation, energy maintenance, and immune modulation (Reyes-López et al., 2018; Tsalafouta et al., 2018).
Cortisol carries out its effect at the molecular level through its recognition by the mineralocorticoid receptor (MR) and the glucocorticoid receptors (GRs) (Guo and Dixon, 2021). These receptors are present in the hypothalamus, the pituitary, and the interrenal cells, suggesting the participation of both receptors in the regulation of the HPI axis (Teles et al., 2013; Liu et al., 2019; Martorell Ribera et al., 2020). It has been suggested that the MR signaling pathway is involved in brain-dependent visual and behavioral responses, stress responses, and osmoregulatory functions (Kiilerich et al., 2017). Two types of GRs (GR1 and GR2) have been identified and the modulation of their mRNA levels have been evaluated in several fish species, including rainbow trout (Oncorhynchus mykiss) (Bury et al., 2003; Peterson et al., 2019) and European sea bass (Dicentrarchus labrax) (Vazzana et al., 2010; Azeredo et al., 2022). Such receptors are proposed to have separate functions, thus responding to different levels of cortisol concentration (Stolte et al., 2006). It has been reported that the main difference between gr1 and gr2 nucleotide sequences resides in an insertion/deletion in the DNA-binding region. Thus, the gr1 sequence has nine extra amino acids (corresponding to 27 nucleotides) compared to the gr2 sequence in teleost fish (Bury et al., 2003; Stolte et al., 2006). In 2007, our research team identified one gr nucleotide sequence in gilthead sea bream (Acerete et al., 2007). Then, Tsalafouta et al. (2018) identified and characterized another GR in gilthead sea bream, that the authors named gr2. Contrary to the reports of Bury et al. (2003) and Stolte et al. (2006), such a gr2 sequence possesses the characteristic 27-nucleotide insertion that, in fact, is associated with the gr1 nucleotide sequence. Therefore, it is clear that the nomenclature for GR1 and GR2 is controversial and is not followed in the same way by the scientific community. Thus, the objective of this study is to elucidate the nucleotide sequences corresponding to gr1 and gr2 in gilthead sea bream. At the same time, our goal is also to emphasize the urgency of standardizing the nomenclature used to name glucocorticoid receptors for a better understanding of the stress response in teleost fish species.
Gilthead sea bream (S. aurata, Linnaeus, 1758) were obtained from a local fish farm (Culmarex, Puerto de Sagunto, Valencia, Spain). The fish were acclimatized to the laboratory conditions in the same tank for 30 days. Fish were kept in a closed recirculating seawater system at 21-22°C, on a 12-h light/12-h dark cycle, and fed a commercial diet at 1.5% of total body weight per day (Skretting). They were kept in 300-liter tanks at a density of 2 kg/m3. The fish were fasted 24 h before the experimental procedures. Dissolved oxygen (7.1 ± 0.1 mg/l), pH (7.1 ± 0.1), nitrite (<0.5 mg/l), nitrate (<10 mg/l), and total ammonia (≤ 0.5 mg/l) were analyzed periodically.
All procedures were carried out following the ethical guidelines for animal experimentation (EEC regulation 86/609), authorized and supervised by the ethics committee of the “Universitat Autònoma de Barcelona” (Ref. OH4218_4219).
We obtained biological samples for the analysis of glucocorticoid receptors 1 and 2 amplification by real-time Polymerase Chain Reaction (RT-qPCR). Gilthead sea bream were subjected for one minute to acute stress by air exposure using a net due to its effectiveness as a model of stress response induction (Arends et al., 1999; Khansari et al., 2018; Skrzynska et al., 2018; Khansari et al., 2019; Liu et al., 2019). Fish were fasted 24 h before the experimental procedure. Then, fish were placed in a 300-liter tank at a culture density of 2 kg/m3. The tank had a closed recirculation system consisting of a water pump, solid filter, and a mature biofilter. Fish were sampled at 1-hour post-stress (hps) and sacrificed by anesthetic overdose (MS222 (Sigma), 200 mg/l). Then, the body cavity was opened laterally and the head kidney (HK) was sampled and immediately frozen in liquid nitrogen and stored at -80°C for further processing.
Total RNA was extracted from each HK sample using the TriReagent reagent (Sigma). Thus, 100 mg of HK was homogenized in 1 ml Tri Reagent® (Sigma) with a Polytron, and then incubated at room temperature for 5 min. Subsequently, 0.1 mL of BCP (Sigma) was added to each sample, vortexed, and incubated at room temperature for 15 min. The samples were centrifuged at 12,000 x g for 15 min at 4°C to separate the phases. After transferring the aqueous phase to a new tube, the total RNA was precipitated by adding 0.5 ml of 2-propanol (Sigma). The samples were incubated at -20°C overnight and subsequently centrifuged at 12,000 x g for 10 min at 4°C. Finally, the total RNA pellet was washed twice with 1 ml of 75% ethanol (prepared with DEPC-water), air-dried for 10 min, resuspended in DEPC-water, and then incubated in a thermoblock at 55°C for 10 min. The RNA sample was quantified with a Nanodrop ND-1000 (Thermo Scientific) and the RNA quality was measured by Bioanalyzer (Agilent Technologies). All the samples had an RNA integrity number (RIN) value > 7.0, being all of them suitable for further analysis. Then, cDNA was synthesized from 1,000 ng of total RNA, using the iScript cDNA kit (Bio-Rad Laboratories) according to the manufacturer’s instructions.
We first aligned by Clustal Omega (Sievers et al., 2011) the glucocorticoid receptor sequences for different fish species. We included the rainbow trout (Oncorhynchus mykiss) gr1 (NM_001124730.1), rainbow trout gr2 (AY495372.1), European sea bass (Dicentrarchus labrax) gr (AY549305.1, AY619996.1), and the available sequence for gilthead sea bream gr (DQ486890.1) obtained from Acerete et al. (2007). We focused our comparative sequence analysis on the DNA-binding region because of the 27-nucleotide insertion/deletion difference between the gr1 and gr2 sequences, respectively (Bury et al., 2003; Stolte et al., 2006).
Based on the insertion/deletion conservative pattern confirmed by our analysis, we designed a primer set for gr1 (Forward: GAAGGATGGAGAGCACGACAAAA; Reverse: TGCAGTGCTCCTGGCTCTTCCT; expected PCR product size: 418 bp). The structure and specificity of the primers were checked with Primer-Blast. PCR reactions were carried out using Platinum™ Pfx DNA Polymerase (Invitrogen) following the manufacturer’s instructions. The template consisted of 200 ng of cDNA obtained from gilthead sea bream head kidneys subjected to acute stress by air exposure (1 min), as described above. The thermal conditions used were 5 min at 94°C (pre-denaturation), followed by 35 cycles at 95°C for 15s (denaturation), 55°C for 30s (annealing), and extension at 68°C for 1 min. The PCR products were analyzed by electrophoresis (80V; 1.5 h; 1X TAE buffer) in 1.5% agarose gel (Sigma). The band that matched the amplicon size was excised and purified from the agarose gel using the NucleoSpin® Gel and PCR Clean-up kit (Macherey-Nagel) according to the manufacturer’s instructions. The purified PCR product was sequenced at the Servei de Genòmica i Bioinformàtica (Universitat Autonoma de Barcelona) using the forward and reverse primer described above. The sequence obtained was subjected to a Blast analysis (NCBI) to check the identity of the sequence.
The phylogenetic analysis was conducted by MEGA 11. The analysis consisted of a multiple alignments by Clustal W, followed by a Neighbor-Joining method (Saitou and Nei, 1987; Kumar et al., 2004) using the Tamura-Nei model for drawing the trees. A total of fifteen mRNA sequences were obtained from Nucleotide database (NCBI), including Sparus aurata (DQ486890.1; XM_030437675.1; GR2-Sparus aurata TR96646|c2_g1_i11 [from (Tsalafouta et al., 2018)]; Dicentrarchus labrax (AY619996.1; AY549305.1); Oncorhynchus mykiss (AY495372.1; NM_001124730.1); Salmo salar (XM_014136782.2); Paralichthys olivaceus (AB013444.1); Astatotilapia burtoni (AF263740.1; AF263738.1); Lates calcarifer (XM_018675716.1); Oreochromis niloticus (XM_013271702.3). We also included, as an outside group, the nucleotide sequence for Mus musculus (DQ504162.1) and Homo sapiens (NM_000176.3). The reliability of the tree was assessed using 1,000 bootstraps replicates.
We first determined if the nucleotide sequence reported as glucocorticoid receptor by Acerete et al. (2007) corresponds to gr1 or gr2 in gilthead sea bream. Thus, we carried out a Clustal Omega analysis including the gr1 and gr2 for rainbow trout, European sea bass, and the gr sequence for gilthead sea bream. The multiple alignment analysis of the five sequences determined that two of them contained the characteristic conserved 27-nucleotide insertion at the DNA-binding region corresponding to the gr1 sequence. By contrast, we noted that the gilthead sea bream gr sequence described by Acerete et al. (2007) has no 27-nucleotide insertion (Figure 1; the full alignment is displayed in Supplementary Figure 1). Therefore, and according to Bury et al. (2003) and Stolte et al. (2006), this sequence corresponds to the gilthead sea bream gr2 but not the gr1, and not as it was stated in a previous study (Tsalafouta et al., 2018).
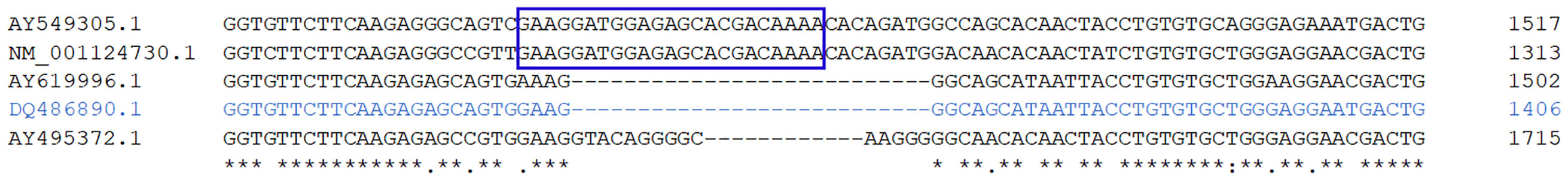
Figure 1 Multiple alignment for the comparison of the glucocorticoid receptors’ nucleotide sequences at the DNA-binding domain. The Clustal Omega analysis includes the glucocorticoid receptors sequences for rainbow trout (Oncorhynchus mykiss) [NM_001124730.1: defined by several reports as glucocorticoid receptor (Ducouret et al., 1995), glucocorticoid receptor with insert (Takeo et al., 1996), as well as glucocorticoid receptor 1 (Becker et al., 2008), supported by nucleotide database (NCBI)]; and the AY495372.1: defined by several reports as glucocorticoid receptor 2 (Bury et al., 2003; Becker et al., 2008), supported by nucleotide database (NCBI)]; sea bass (Dicentrarchus labrax) [AY549305.1: defined by Terova et al. (Terova et al., 2005) as glucocorticoid receptor; AY619996.1: defined by Vizzini et al. (Vizzini et al., 2007) as “Dicentrarchus labrax glucocorticoid receptor 1 (DlGR1)]; and sea bream [DQ486890.1: defined by Acerete et al. (2007) as glucocorticoid receptor, or glucocorticoid receptor 1 by Tsalafouta et al. (Tsalafouta et al., 2018); sequence colored in blue]. The Nucleotide accession number (NCBI) is detailed on the left of each sequence. The blue box represents the forward primer designed into the conserved 27-nucleotide insertion at the DNA-binding region for identifying the sea bream glucocorticoid receptor 1 by PCR. The "*" (asterisk) indicates positions which have a single, fully conserved residue. The ":" (colon) indicates conservation between groups of strongly similar properties. The "." (period) indicates conservation between groups of weakly similar properties.
Importantly, the AY619996.1 sequence defined by Vizzini et al. (2007) as “Dicentrarchus labrax glucocorticoid receptor 1 (DlGR1)” does not contain the 27-nucleotide insertion at the DNA-binding region. Thus, for the European sea bass, this sequence is probably the gr2 but not the gr1 nucleotide sequence.
In order to identify the partial nucleotide sequence of gr1 in gilthead sea bream, a PCR was conducted from the head kidney of fish stressed by air exposure. Notably, the PCR fragment sequenced contains the 27-nucleotide insertion in the DNA-binding region and characteristic of the gr1 in teleost species (Figure 2). This newly identified gr1 partial nucleotide sequence was analyzed by Blast (NCBI) to check for any match with other sequences in the repository. Thus, this identified gr1 showed 99% identity with the XM_030437675.1 sequence (PREDICTED: Sparus aurata nuclear receptor subfamily 3 group C member 1 (nr3c1), transcript variant X1, mRNA) (Figure 2). Moreover, both sequences shared the 27-nucleotide insertion in the DNA-binding region sequence (Figure 2).

Figure 2 Multiple alignment for the sequence obtained from our purified PCR product and the sequence with the highest match obtained from BLAST analysis. The alignment for the DNA-binding region is presented for the PCR product (381-bp) and the sequence XM_030437675.1 (from 1955- to 2336-bp). A 99% identity is obtained (378/383-bp) between both sequences. In red is represented the 27-nucleotide insertion in the DNA-binding region and characteristic of the gr1 sequence. The "*" (asterisk) indicates positions which have a single, fully conserved residue. The "." (period) indicates conservation between groups of weakly similar properties.
We confirmed by multiple alignments, including the gr1 and gr2 from several teleost fish species, that the sequences that contain the 27-nucleotide insertion in the DNA-binding region correspond to glucocorticoid receptor 1 (Figure 3). On the other hand, the sequences that do not contain such an insertion correspond to the gr2 sequence (Figure 3). Thus, we confirm that the sequence obtained from the sequencing of the purified PCR product and the subsequent match with the BLAST sequence (XM_030437675.1) corresponds to gr1, while the glucocorticoid receptor from Acerete et al. (2007) is the gr2. This is reinforced by the phylogenetic analysis, grouping the gr1 and gr2 sequences into clearly different clusters (Figure 4). The full alignment of the sequences contained in Figures 3, 4 is detailed in Supplementary Figure 2.
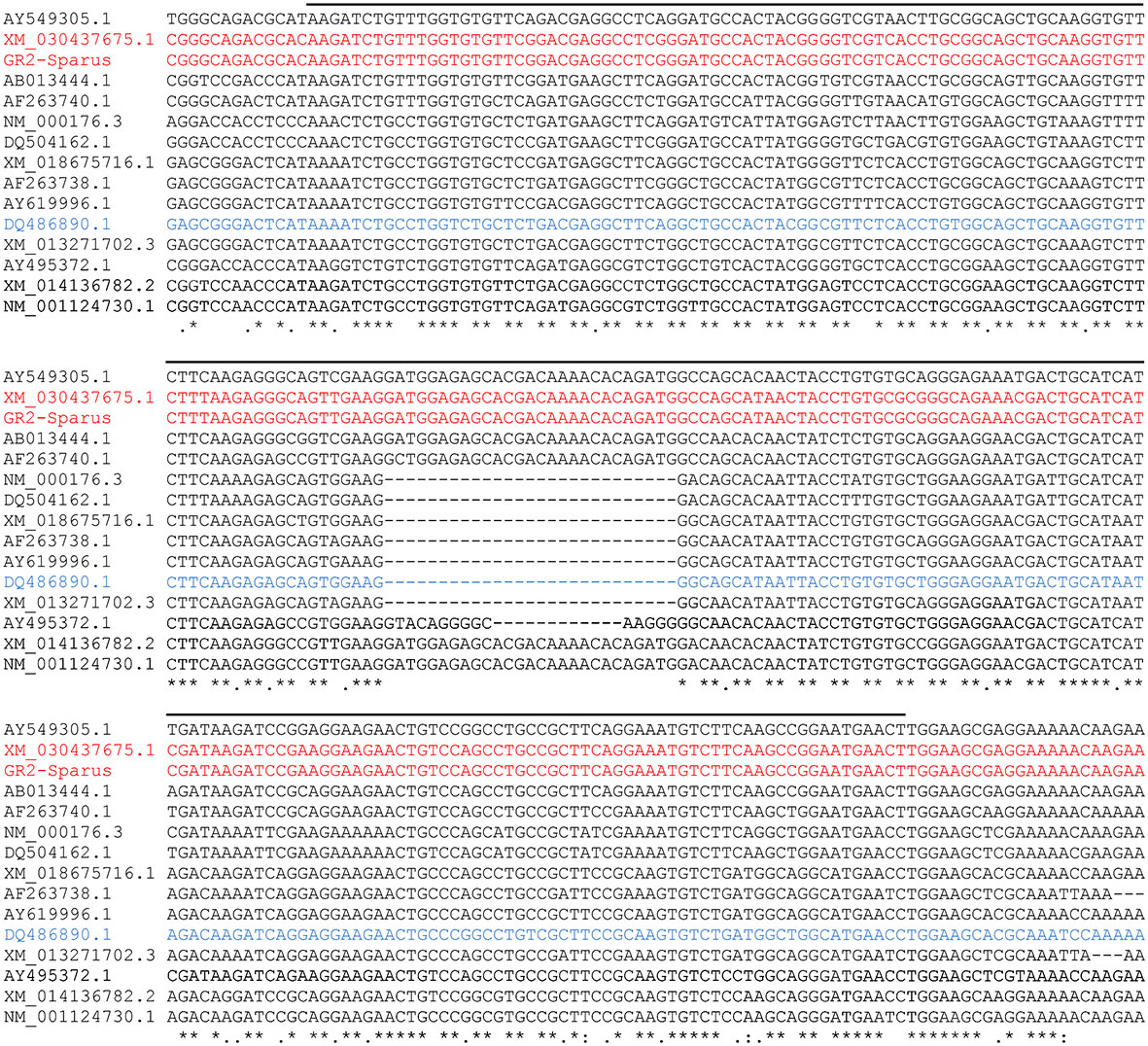
Figure 3 Multiple alignment for the comparison of the glucocorticoid receptors’ nucleotide sequences at the DNA-binding region. The Clustal Omega analysis includes the glucocorticoid receptors (gr) sequences for several fish species. The DNA-binding region is denoted by the upper horizontal line throughout the alignment. The gap observed in the central band corresponds to the characteristic 27-nucleotide deletion for the identification of gr2. The sequences associated to gr1 are detailed below according to the order of presentation (from top to bottom) in the alignment, and indicating on the left column of alignment the fish species and the Nucleotide accession number (NCBI), represented: AY549305.1 for European sea bass (Dicentrarchus labrax); XM_030437675.1 (99% identity with our sequenced PCR product; indicated in red) and GR2-Sparus aurata TR96646|c2_g1_i11 for gilthead sea bream [S. aurata; indicated in red and described in Tsalafouta et al. (2018)]; AB013444.1 for Japanese flounder (Paralichthys olivaceus); AF263740.1 for Burton’s mouth-brooder (Astatotilapia burtoni); XM_014136782.2 for Atlantic salmon (Salmo salar); and NM_001124730.1 for rainbow trout (Oncorhynchus mykiss). The sequences for the outgroup were (from top to bottom) NM_000176.3 for humans (Homo sapiens), and DQ504162.1 for mice (Mus musculus). The sequences associated with gr2 were (from top to bottom) the XM_018675716.1 for Asian sea bass (Lates calcarifer); AF263738.1 for Burton’s mouth-brooder (A. burtoni); AY619996.1 for European sea bass (D. labrax); DQ486890.1 for gilthead sea bream (S. aurata; defined by Acerete et al. (2007) as glucocorticoid receptor; indicated in blue); XM_013271702.3 for Nile tilapia (Oreochromis niloticus); and AY495372.1 for rainbow trout (O. mykiss). The "*" (asterisk) indicates positions which have a single, fully conserved residue. The ":" (colon) indicates conservation between groups of strongly similar properties. The "." (period) indicates conservation between groups of weakly similar properties.

Figure 4 Phylogenetic analysis of gr1 and gr2 sequences for different species of teleost fish. The image represents the clades for teleost glucocorticoid receptor 2 (gr2; orange), teleost gr1 (gr1; light blue) that includes the 27-nucleotide (27-nt) insertion, and the outgroup (green). The sequences associated with gr2 are detailed below according to the order of presentation (from top to bottom) in the alignment, and indicating the fish species and the Nucleotide accession number [(NCBI), represented on the left column of alignment]: DQ486890.1 for gilthead sea bream (Sparus aurata; defined by Acerete et al. (2007) as glucocorticoid receptor); AY619996.1 for European sea bass (Dicentrarchus labrax); XM_018675716.1 for Asian sea bass (Lates calcarifer); XM_013271702.3 for Nile tilapia (Oreochromis niloticus); AF263738.1 for Burton’s mouth-brooder (Astatotilapia burtoni); and AY495372.1 for rainbow trout (Oncorhynchus mykiss). The sequences associated with gr1 (from top to bottom) were the AF263740.1 for Burton’s mouth-brooder (A. burtoni); AB013444.1 for Japanese flounder (Paralichthys olivaceus); AY549305.1 for European sea bass (D. labrax); XM_030437675.1 (99% identity with our sequenced PCR product) and GR2-Sparus aurata TR96646|c2_g1_i11 [described in Tsalafouta et al. (2018)] for gilthead sea bream (S. aurata); NM_001124730.1 for rainbow trout (O. mykiss); and XM_014136782.2 for Atlantic salmon (Salmo salar). The values at branch nodes indicate the percentage of trees in which the associated taxa clustered together.
This study aimed to elucidate the nucleotide sequences corresponding to gr1 and gr2 in gilthead sea bream. We demonstrate that the glucocorticoid receptor reported by Acerete et al. (2007) is the gr2 sequence because it possesses the 27-nucleotide deletion in the DNA-binding region. On the other hand, we found a new glucocorticoid nucleotide sequence that contained the 27-nucleotide insertion in the DNA-binding region, calling it glucocorticoid receptor 1 (gr1). Importantly, we have found 100% identity with a sequence already published in the NCBI database. Therefore, we suggest that the sequence XM_030437675.1 corresponds to the full-sequence of gr1. The phylogenetic analysis supports our conclusion. Thus, our study is the first to report the nucleotide sequence for the glucocorticoid receptor 1 in gilthead sea bream. These findings are in agreement with Bury et al. (2003) and Stolte et al. (2006) which indicate that the gr1 contains the 27-nucleotide insertion in the DNA-binding region. By contrast, the gr2 sequence contains the 27-nucleotide deletion in the same region.
The GR1 and GR2 are particularly important for the activation of the different physiological mechanisms to deal with the stressor. It has been postulated that GR2 has a higher affinity for cortisol, making possible the ligand-receptor interaction at a very low concentration of cortisol. By contrast, GR1 would be responsible for the recognition of cortisol at higher concentrations (Vazzana et al., 2010). Hence, glucocorticoid receptors play a predominant role in the stress response (Schreck and Tort, 2016; Jaikumar et al., 2020). Accordingly, previous efforts have been made for unveiling the glucocorticoid receptors sequences in several teleost species, including gilthead sea bream. In the report of Tsalafouta et al. (Tsalafouta et al., 2018), two different glucocorticoid sequences for gilthead sea bream were found by sequencing. One of them showed 100% identity with the gr sequence published by Acerete et al. (Acerete et al., 2007). Such sequence has the 27-nucleotide deletion in the DNA-binding region. Unfortunately, this gene sequence was called gr1 instead of gr2 (Tsalafouta et al., 2018). By contrast, the other sequence containing 27-nucleotide insertion in the DNA-binding region was called gr2 instead of gr1 (Tsalafouta et al., 2018). Thus, the study of Tsalafouta et al. (Tsalafouta et al., 2018) did not follow the nomenclature proposed by Bury et al. (2003) and Stolte et al. (2006). Unfortunately, another study used the information described in Tsalafouta et al. to evaluate the gene expression profile of gr1 and gr2 (Aedo et al., 2019).
The study of Tsalafouta et al. (2018) is not the unique study that has not taken into account the GR nomenclature in teleost species. In fact, a glucocorticoid sequence used by Vizzini et al. (Vizzini et al., 2007) was described as “Dicentrarchus labrax glucocorticoid receptor 1 (DlGR1). This sequence contains the 27-nucleotide deletion in the DNA-binding region. Consequently, and according to Bury et al. (2003) and Stolte et al. (2006), this sequence seems to be the gr2 of the European sea bass. In other Perciformes species, such as European sea bass, the first sequence described by Terova et al. (Terova et al., 2005) was defined as a glucocorticoid receptor. In our analysis, we detected that this sequence contains the 27-nucleotide in the DNA-binding region. Thus, we hypothesize these studies bypassed the foundations established in previous studies for naming glucocorticoid receptors. This generates a lack of precision or an alternative way to name the same receptors, thus generating background noise in the understanding of the mechanisms responsible for the activation of the stress response. Therefore, in agreement with Bury et al. (2003) and Stolte et al. (2006), our study suggests that this sequence should be named as gr1. This apparent confusion or lack of specification between glucocorticoid receptors could even compromise the findings obtained for such GRs. For this reason, we recommend extreme caution when interpreting the scope of the reports that name glucocorticoid receptors with different nomenclature (Vizzini et al., 2007; Tsalafouta et al., 2018; Aedo et al., 2019). In the particular context of gilthead sea bream, it remains to elucidate the effect of the stress response upon the modulation of the gr1 and gr2 at the gene expression level. In this context, a recent study revealed that in mucosal tissues gr1 but not gr2 is modulated in gilthead sea bream exposed to acute air-exposure stress (Vallejos-Vidal et al., 2022).
In summary, our study identified and clarified the nucleotide sequences for gr1 and gr2. In this process, we found that in gilthead sea bream and in the European sea bass these sequences had not been named according to the findings established in the reports of Bury et al. (2003) and Stolte et al. (2006). For this reason, our study makes an urgent call to the scientific community to re-establish an updated agreement that allows homogenizing the criteria for the nomenclature defining the gr1 and gr2 nucleotide sequences in teleosts. In this way, we propose to follow the conclusions of Bury et al. (2003) and Stolte et al. (2006) as a benchmark because they include fish but also mammals species in their multiple alignment and phylogenetic analyses. This consensus will allow us a better understanding of the stress response in the teleost fish species.
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
All procedures were carried out following the ethical guidelines for animal experimentation (EEC regulation 86/609), authorized and supervised by the ethics committee of the “Universitat Autònoma de Barcelona” (Ref. OH4218_4219).
EV-V, JMM, LT, and FER-L conceptualized the study. EV-V, AK, MT, LT, and FER-L carried out the experiments. EV-V, AK, and MT made the sampling processing for the identification of gr1 and gr2. Data sequencing analysis was in charge of EV-V, SR-C, and FER-L. EV-V, AK, and FER-L conducted the sampling processing for PCR. EV-V, JMM, LT, and FER-L performed the data interpretation. EV-V and FER-L conceptualized the figures. EV-V and FER-L wrote the original draft. All authors contributed to the article and approved the submitted version.
This study thanks to the AGL2016-76069-C2-2- R, PID2020-117557RB-C21, PID2020-117557RB-C22 grants (AEI-MINECO; Spain). EV-V thanks the support of Fondecyt iniciación grant (project number 11221308; Agencia Nacional de Investigación y Desarrollo de Chile, Government of Chile). AK was the recipient of a Ministry of Science, Research, and Technology (Iran) fellowship. MT thanks for the support of the post-doctoral fellowship “Ramón y Cajal” (ref. RYC2019-026841-I) (Ministerio de Ciencia e Innovación, Spanish Government). FER-L thanks the support of Fondecyt regular grant (project number: 1211841; Agencia Nacional de Investigación y Desarrollo de Chile, Government of Chile).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The handling Editor (RA) declared a past co-authorship with the author (JMM).
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2022.1021046/full#supplementary-material
Acerete L., Balasch J. C., Castellana B., Redruello B., Roher N., Canario A. V., et al. (2007). Cloning of the glucocorticoid receptor (GR) in gilthead seabream (Sparus aurata). Differential expression of GR and immune genes in gilthead seabream after an immune challenge. Comp. Biochem. Physiol. - B Biochem. Mol. Biol. 148, 32–43. doi: 10.1016/j.cbpb.2007.04.015
Aedo J. E., Ruiz-Jarabo I., Martínez-Rodríguez G., Boltaña S., Molina A., Valdés J. A., et al. (2019). Contribution of non-canonical cortisol actions in the early modulation of glucose metabolism of gilthead Sea bream (Sparus aurata). Front. Endocrinol. (Lausanne) 10. doi: 10.3389/fendo.2019.00779
Arends R. J., Mancera J. M., Muñoz J. L., Wendelaar Bonga S. E., Flik G. (1999). The stress response of the gilthead sea bream (Sparus aurata l.) to air exposure and confinement. J. Endocrinol. 163, 149–157. doi: 10.1677/joe.0.1630149
Azeredo R., Machado M., Pereiro P., Barany A., Mancera J. M., Costas B. (2022). Acute inflammation induces neuroendocrine and opioid receptor genes responses in the seabass dicentrarchus labrax brain. Biol. (Basel) 11, 1–22. doi: 10.3390/biology11030364
Balasch J. C., Tort L. (2019). Netting the stress responses in fish. Front. Endocrinol. (Lausanne) 10. doi: 10.3389/fendo.2019.00062
Becker H., Sturm A., Bron J. E., Schirmer K., Bury N. R. (2008). The A/B domain of the teleost glucocorticoid receptors influences partial nuclear localization in the absence of hormone. Endocrinology 149, 4567–4576. doi: 10.1210/en.2007-1683
Bury N. R., Sturm A., Le Rouzic P., Lethimonier C., Ducouret B., Guiguen Y., et al. (2003). Evidence for two distinct functional glucocorticoid receptors in teleost fish. J. Mol. Endocrinol. 31, 141–156. doi: 10.1677/jme.0.0310141
Ducouret B., Tujaguei M., Ashraf J., Mouchel N., Servel N., Valotaire Y., et al. (1995). Cloning of a teleost fish glucocorticoid receptor that it contains a deoxyribonucleic acid-binding domain different from that of mammals b.from that of mammals *. Endocrinology 136, 3774–3783. doi: 10.1210/endo.136.9.7649084
Guo H., Dixon B. (2021). Understanding acute stress-mediated immunity in teleost fish. Fish Shellfish Immunol. Rep. 2, 100010. doi: 10.1016/j.fsirep.2021.100010
Jaikumar G., Slabbekoorn H., Sireeni J., Schaaf M., Tudorache C. (2020). The role of the glucocorticoid receptor in the regulation of diel rhythmicity. Physiol. Behav. 223, 112991. doi: 10.1016/j.physbeh.2020.112991
Khansari A. R., Balasch J. C., Vallejos-Vidal E., Parra D., Reyes-López F. E., Tort L. (2018). Comparative immune- and stress-related transcript response induced by air exposure and vibrio anguillarum bacterin in rainbow trout (Oncorhynchus mykiss) and gilthead seabream (Sparus aurata) mucosal surfaces. Front. Immunol. 9. doi: 10.3389/fimmu.2018.00856
Khansari A. R., Balasch J. C., Vallejos-Vidal E., Teles M., Fierro-Castro C., Tort L., et al. (2019). Comparative study of stress and immune-related transcript outcomes triggered by vibrio anguillarum bacterin and air exposure stress in liver and spleen of gilthead seabream (Sparus aurata), zebrafish (Danio rerio) and rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 86, 436–448. doi: 10.1016/j.fsi.2018.11.063
Kiilerich P., Servili A., Péron S., Valotaire C., Goardon L., Leguen I., et al. (2017). Regulation of the corticosteroid signalling system in rainbow trout HPI axis during confinement stress. Gen. Comp. Endocrinol 258, 184–193. doi: 10.1016/j.ygcen.2017.08.013
Kumar S., Tamura K., Nei M. (2004). MEGA3: Integrated software for molecular evolutionary genetics analysis and sequence alignment. Brief. Bioinform. 5, 150–163. doi: 10.1093/bib/5.2.150
Liu X. H., Khansari A. R., Teles M., Martínez-Rodríguez G., Zhang Y. G., Mancera J. M., et al. (2019). Brain and pituitary response to vaccination in gilthead seabream (Sparus aurata l.). Front. Physiol. 10. doi: 10.3389/fphys.2019.00717
Martorell Ribera J., Nipkow M., Viergutz T., Brunner R. M., Bochert R., Koll R., et al. (2020). Early response of salmonid head-kidney cells to stress hormones and toll-like receptor ligands. Fish Shellfish Immunol. 98, 950–961. doi: 10.1016/j.fsi.2019.11.058
Peterson B. C., Chatakondi N. G., Small B. C. (2019). Ontogeny of the cortisol stress response and glucocorticoid receptor expression during early development in channel catfish, ictalurus punctatus. Comp. Biochem. Physiol. -Part A Mol. Integr. Physiol. 231, 119–123. doi: 10.1016/j.cbpa.2019.02.003
Reyes-López F. E., Aerts J., Vallejos-Vidal E., Ampe B., Dierckens K., Tort L., et al. (2018). Modulation of innate immune- related genes and glucocorticoid synthesis in gnotobiotic full-sibling European Sea bass (Dicentrarchus labrax) larvae challenged with vibrio anguillarum. Front. Immunol. 9. doi: 10.3389/fimmu.2018.00914
Saitou N., Nei M. (1987). The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 4, 406–425. doi: 10.1093/oxfordjournals.molbev.a040454
Schreck C. B., Tort L. (2016). Biology of stress in fish. 1st ed. Eds. Farrell A. P., Brauner C. J. (London: Academic Press).
Sievers F., Wilm A., Dineen D., Gibson T. J., Karplus K., Li W., et al. (2011). Fast, scalable generation of high-quality protein multiple sequence alignments using clustal omega. Mol. Syst. Biol. 7, 539. doi: 10.1038/msb.2011.75
Skrzynska A. K., Maiorano E., Bastaroli M., Naderi F., Míguez J. M., Martínez-Rodríguez G., et al. (2018). Impact of air exposure on vasotocinergic and isotocinergic systems in gilthead sea bream (Sparus aurata): New insights on fish stress response. Front. Physiol. 9. doi: 10.3389/fphys.2018.00096
Stolte E. H., Van Kemenade B. M. L. V., Savelkoul H. F. J., Flik G. (2006). Evolution of glucocorticoid receptors with different glucocorticoid sensitivity. J. Endocrinol. 190, 17–28. doi: 10.1677/joe.1.06703
Takeo J., Hata J., Segawa C., Toyohara H., Yamashita S. (1996). Fish glucocorticoid receptor with splicing variants in the DNA binding domain. FEBS Lett. 389, 244–248. doi: 10.1016/0014-5793(96)00596-0
Teles M., Boltaña S., Reyes-López F., Santos M. A., Mackenzie S., Tort L. (2013). Effects of chronic cortisol administration on global expression of GR and the liver transcriptome in sparus aurata. Mar. Biotechnol. (NY) 15, 104–114. doi: 10.1007/s10126-012-9467-y
Terova G., Gornati R., Rimoldi S., Bernardini G., Saroglia M. (2005). Quantification of a glucocorticoid receptor in sea bass ( dicentrarchus labrax , l .) reared at high stocking density. Gene 357, 144–151. doi: 10.1016/j.gene.2005.06.016
Tort L. (2011). Stress and immune modulation in fish. Dev. Comp. Immunol. 35, 1366–1375. doi: 10.1016/j.dci.2011.07.002
Tsalafouta A., Sarropoulou E., Papandroulakis N., Pavlidis M. (2018). Characterization and expression dynamics of key genes involved in the gilthead Sea bream (Sparus aurata) cortisol stress response during early ontogeny. Mar. Biotechnol. 20, 611–622. doi: 10.1007/s10126-018-9833-5
Vallejos-Vidal E., Sanz-Milián B., Teles M., Reyes-Cerpa S., Mancera J. M., Tort L. (2022). The gene expression profile of the glucocorticoid receptor 1 (gr1) but not gr2 is modulated in mucosal tissues of gilthead sea bream (Sparus aurata) exposed to acute air-exposure stress. Front. Mar. Sci. 9, 977719. doi: 10.3389/fmars.2022.977719
Vazzana M., Vizzini A., Sanfratello M. A., Celi M., Salerno G., Parrinello N. (2010). Differential expression of two glucocorticoid receptors in seabass (teleost fish) head kidney after exogeneous cortisol inoculation. Comp. Biochem. Physiol. - A Mol. Integr. Physiol. 157, 49–54. doi: 10.1016/j.cbpa.2010.05.003
Vizzini A., Vazzana M., Cammarata M., Parrinello N. (2007). Peritoneal cavity phagocytes from the teleost sea bass express a glucocorticoid receptor (cloned and sequenced) involved in genomic modulation of the in vitro chemiluminescence response to zymosan. Gen. Comp. Endocrinol. 150, 114–123. doi: 10.1016/j.ygcen.2006.07.016
Keywords: stress response, cortisol, glucocorticoid receptors (GRs), glucocorticoid receptor 1 (GR1), glucocorticoid receptor 2 (GR2), perciformes fish species
Citation: Vallejos-Vidal E, Khansari AR, Teles M, Reyes-Cerpa S, Mancera JM, Tort L and Reyes-López FE (2022) The insertion/deletion in the DNA-binding region allows the discrimination and subsequent identification of the glucocorticoid receptor 1 (gr1) and gr2 nucleotide sequences in gilthead sea bream (Sparus aurata): Standardizing the gr nomenclature for a better understanding of the stress response in teleost fish species. Front. Mar. Sci. 9:1021046. doi: 10.3389/fmars.2022.1021046
Received: 16 August 2022; Accepted: 14 September 2022;
Published: 11 October 2022.
Edited by:
Rita Azeredo, University of Porto, PortugalReviewed by:
Zhenhua Ma, South China Sea Fisheries Research Institute (CAFS), ChinaCopyright © 2022 Vallejos-Vidal, Khansari, Teles, Reyes-Cerpa, Mancera, Tort and Reyes-López. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lluis Tort, TGx1aXMuVG9ydEB1YWIuZXM=; Felipe E. Reyes-López, ZmVsaXBlLnJleWVzLmxAdXNhY2guY2w=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.