 Sergio Ramírez-Amaro
Sergio Ramírez-Amaro Marta Bassitta1
Marta Bassitta1 Antònia Picornell
Antònia Picornell Cori Ramon
Cori Ramon- 1Laboratori de Genètica, Universitat de les Illes Balears, Palma, Spain
- 2Instituto Español de Oceanografía (IEO-CSIC), Centre Oceanogràfic de les Balears, Palma, Spain
Fisheries management involves a broad and complex set of tasks that are necessary to prevent overfishing and to help the recovery of overfished stock. Monitoring fishing activities based on two main sources, landings data and scientific surveys, is a challenging task. Fisheries collection data is often limited, which compromises the accuracy of the results obtained. Therefore, several emerging applications of molecular methods have the potential to provide unique understanding of ecological processes in marine environments and to build stronger empirical underpinnings for the Ecosystem-Based Fisheries Management. Environmental DNA (eDNA) is a complex mixture of genetic material shed by those organisms that inhabit a given environment, whereby DNA is extracted from an environmental sample without accessing the target organism. eDNA studies can be categorized into two main approaches, i) eDNA metabarcoding or semi-targeted (community) approaches and ii) species-specific or targeted approaches (single). Although both categories are often discussed, they differ drastically in their methodology, interpretations and accuracy. Both approaches involve a series of steps that include eDNA capture, preservation, extraction and amplification. This detection will depend on the affinity to the targeted taxa sequences and completeness and accuracy of DNA reference collection databases. The eDNA method applied in marine environments are probably the most challenging aquatic environments for applying this technique. This is because of the extreme relationship between water-volume to biomass, dynamics and the physical and chemical properties of seawater that affect dispersion, dilution and preservation. Here, we review the present application of this novel method in fishery assessment in marine environments. To date, many studies suggest that this method offers the potential to revolutionize fisheries monitoring, which will contribute to improving the range of tasks involved in fisheries management. The compelling conclusion is that the methodological steps including in eDNA surveys should be standardized and that research efforts should focus on developing appropriately validated tests to address environmental and sampling factors that may affect eDNA detection in marine environments in order to draw reliable conclusions. This bioassessment tool can assist fisheries professionals in achieve their research, management, and conservation objectives, but not as a replacement for time-proven assessment methods.
Fisheries
Marine fisheries
Fisheries management involves a broad and complex set of tasks (e.g., gathering, analyses, decision-making, resources allocation, implementation), which have the common goal of ensuring the continued productivity of the resources and the accomplishment of other fisheries objectives (Cochrane and Garcia, 2009). Knowledge and management of fisheries depends on accurate and precise data on the distribution and population status of exploited species. According to Cochrane (2000), there are key principles of fisheries management include i) fish stocks and communities, ii) biological production of a stock, iii) demand for human consumption, iv) multispecies fisheries, v) uncertainty of data collection, vi) the short-term dependence on society, vii) ownership of the resource and viii) appropriate communication.
These principles cannot be considered in isolation, in line with the integrated nature of fisheries ecosystems. Because of this, the concept of Ecosystem-Based Fisheries Management (EBFM) was incorporated. It is a holistic concept for managing fisheries and marine resources by considering the entire ecosystem of the species under management. The main objective of EBFM is to maintain the ecosystem in a healthy, productive and resilient condition so that it can provide the services that humans need (Pikitch et al., 2004). Since this concept was proposed, regular monitoring of the marine environment and its living resources has become increasingly necessary. Monitoring of fishing activities is based on two main sources of information: landings data (fishery-dependent) and scientific surveys (fishery-independent) (Dennis et al., 2015). The former type of data is often limited, compromising the accuracy of the results obtained, and the latter depends on the huge operational costs of the vessels (Dennis et al., 2015). Tools to monitor the interaction between fisheries and the environment are diverse, complex and difficult to characterize accurately. Thus, several emerging applications of molecular methods have the potential to provide unique insights into ecological processes in marine environments and to build stronger empirical underpinnings for EBFM (Ovenden et al., 2015).
What is eDNA?
Environmental DNA (eDNA) is defined as DNA captured from an environmental sample without first isolating any target organisms (Taberlet et al., 2012a; Deiner et al., 2017). This refers to any DNA that is collected from the environment rather than directly from an organism, originating from body cells or waste products of organisms. This DNA is released from organisms into a variety of environmental samples such as soil, seawater, snow, or even air (Ficetola et al., 2008; Epp et al., 2012; Turner et al., 2015). Environmental monitoring through DNA is a rapidly growing field largely driven by novel technological developments such as microarray analysis, high-throughput sequencing (HTS), quantitative Polymerase Chain Reaction (qPCR)/droplet digital PCR (ddPCR) and improved bioinformatics capabilities. For eDNA two approaches can be considered, i) based on DNA metabarcoding employing HTS to detect multi-species of mixed and large complex communities, and ii) generally based on qPCR and ddPCR, where the aim is to determine the presence or absence of a single species (e.g., invasive, rare, elusive or endangered species).
The eDNA surveys have recently been proposed as a tool to improve fisheries assessments, and thus conservation success by reducing systematic errors in species richness inference resulting from low detection probabilities and species misidentifications (Evans and Lamberti, 2018). The eDNA concept began in the 90s, with the development of a new method for the isolation of DNA from a variety of sediments (Ogram et al., 1987) and the use of barcoding to phylogenetically analyse clone libraries of eubacterial 16S ribosomal RNA genes amplified from natural populations of the Sargasso Sea (Giovannoni et al., 1990; Figure 1). At the beginning of the 21st century, there was an increase in eDNA-based studies for the detection of microorganisms (Handelsman et al., 1998) and macroorganism species from water and soil samples (e.g., Ficetola et al., 2008). In the early 2000s, next-generation platforms emerged, leading to new techniques such as metabarcoding and metagenomics.
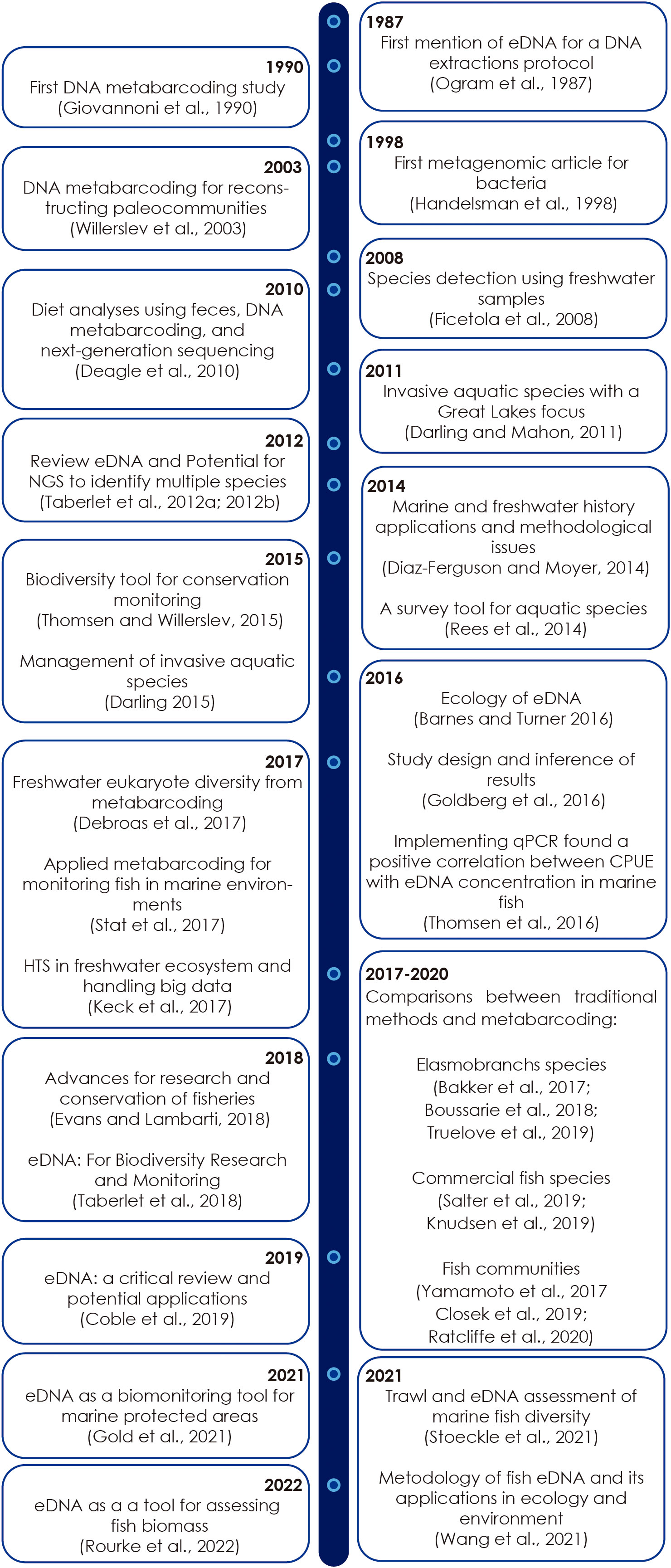
Figure 1 Timeline of representative eDNA literature between 1987 and 2022. The last five years only consider studies of biological monitoring in marine environment.
The first reviews related to eDNA were published in the early 2010s (e.g., Lodge et al., 2012; Taberlet et al., 2012a; Bohmann et al., 2014; Figure 1). The foremost uses of eDNA detection in the marine environments were conducted by Foote et al. (2012) for small-scale genetic monitoring of marine mammals, and by Thomsen et al. (2012a) for analyses of marine macrofauna and meiofauna using seawater samples. In addition, Kelly et al. (2014) published an aquatic eDNA analysis method and also developed a process to monitor the census of marine fishes in a large mesocosm. Furthermore, studies focusing on the comparison of traditional monitoring methods and eDNA (e.g., Boussarie et al., 2018; Knudsen et al., 2019; Stoeckle et al., 2021; Truelove et al., 2019; Figure 1) have helped to demonstrate the high utility of this novel method.
The number of scientific articles published on marine eDNA is lower than the rest of articles focused in the detection of freshwater organisms. However, a search in Google Scholar with both terms “eDNA and ocean”, yielded over 6500 existing articles until the year 2017. And in the last five years (2017-2021) almost 9500 articles have been published, of which 3000 articles are related with fisheries topics, which demonstrate that eDNA is currently widely used in this field.
Applications in fisheries management
eDNA offers a potentially powerful method to improve different task related to fisheries assessment in marine environments, mainly in regards to marine biomonitoring by significantly increasing spatial and temporal biological monitoring in aquatic ecosystems due to the ease of water samples collection (Thomsen and Willerslev, 2015; Sassoubre et al., 2016), and by reducing labor-intensive routine taxonomic identification.
eDNA is an efficient tool for the accurate species identification, which is a key aspect of fisheries management. Determining the species compositions of local assemblages is a prerequisite for understanding how anthropogenic disturbances affect biodiversity. eDNA analysis has the potential to detect changes in the biological composition of communities in different ocean regions, which are much more challenging to measure because most biological monitoring methods focus on a limited taxonomic or size range, and is capable of identifying a huge phylogenetic range of organisms down to the species level. Several studies have demonstrated the utility of eDNA metabarcoding for assessing fish diversity (Supplementary Table S1). Biodiversity baselines can therefore be compiled through eDNA, which can help to understand short or long-term changes through comparison with future collections (Jarman et al., 2018; Djurhuus et al., 2020).
The eDNA method also has the potential to estimate abundance or biomass (see references in Supplementary Table S2), a key aspect for its application in stock assessments. The ability to estimate abundance based on concentrations of eDNA relies in part on the assumption that the release of eDNA from feces, secretions, or tissues correlates with the abundance or standing biomass of the respective individuals. Previous eDNA experiments using discrete static mesocosms (aquaria or ponds) showed an association between density and amplification rate (Ficetola et al., 2008), density and DNA concentration (Thomsen et al., 2012b), and biomass and DNA concentration (Takahara et al., 2012). Although several studies have found a positive relationship between eDNA concentration and abundance or biomass in lentic and lotic systems (Doi et al., 2017; Lacoursière-Roussel et al., 2016a; Takahara et al., 2012), there remains substantial variability around this relationship (Goldberg et al., 2015; Jerde and Mahon, 2015). In particular, the relationship between biomass and eDNA in flowing waters remains unclear due to contrasting results (Laramie et al., 2015; Spear et al., 2015; Doi et al., 2017; Hinlo et al., 2017). The variation in the relationship between eDNA and density could be due to differences in movement and retention of eDNA in the systems, for ecample., while eDNA in lentic systems is contained, transport of eDNA in lotic environments is complicated by flow and stream morphologies (Goldberg et al., 2011; Jerde et al., 2011; Olson et al., 2012).
The patchy distribution of fish DNA and the large variation in copy numbers in samples from the same location currently preclude making inferences about fish abundance (Hinlo et al., 2017). Some difficulties with the use of copy numbers are also related to variation in DNA source (e.g., multiple copies of mtDNA). Copy numbers may indicate some relationship with biomass, but this is also confounded with diversity in size distribution and life history stage of individuals. The correlation between eDNA copy number and catch per unit effort in flowing systems requires further study. Some publications suggest that temporal factors such as breeding and migration seasons, should also be considered in eDNA studies to increase the probability of detection (Thomsen and Willerslev, 2015; Barnes and Turner, 2016). Future studies on the temporal and spatial aspects of eDNA in various species and habitats should be carried out to evaluate the eDNA method in terms of conservation issues (Thomsen and Willerslev, 2015; Furlan et al., 2016). Some studies argue that results obtained with this method should be taken with caution, mainly regarding the inference of abundance through eDNA concentration, especially in natural flowing water bodies. The nature of each marine species, such as habitat preference, could affect the dispersal of eDNA in the water column and thus compromise the success of eDNA (Hinlo et al., 2017). The use of eDNA as a detection tool holds great promise, but as it entails additional cost and effort, studies comparing the performance of eDNA with conventional tools during routine monitoring are needed for its adoption by management.
Using eDNA as a tool for generating population genetic data can be a preferred alternative to sampling biological tissues, which is often expensive and invasive. Knowledge of genetic structure helps conservation management to delineate Management Units and/or stocks and organize actions to preserve genetic diversity (Palsbøll et al., 2007; Abdul-Muneer, 2014). Population characteristics of large whale shark aggregations inferred from seawater eDNA showed that HTS of seawater eDNA can provide useful estimates of genetic diversity, thus extending the applications of eDNA to encompass population genetics of marine organisms (Sigsgaard et al., 2016).
Thomsen et al. (2016) reported eDNA metabarcoding of seawater samples and compare eDNA sequence reads with parallel trawl catch data. The two sampling methods showed generally good overlap. They presented results that suggest a correspondence between fish density (abundance and biomass) and marine eDNA sequence reads produced from Illumina HTS and show a possibility to assess marine fish stock using water samples. Calibration of this new technology against traditional methods is complex due to the fact that all marine monitoring techniques have “catchability” biases (Arreguín-Sánchez, 1996; Fraser et al., 2007). In this sense, Stoeckle et al. (2021) describe an example of bottom trawl fishery. These authors highlighted that bottom trawl catches are influenced by aspects of equipment such as net type, mesh size, and towing speed, and by biological factors such as patchy distribution and habitat preference. They suggested that eDNA surveys should be calibrated and compared with established methods in diverse habitats under a variety of hydrographic conditions.
eDNA analysis in fisheries science has also focused on the application of the method to the detection and monitoring of invasive fish and at-risk species. The sensitivity of eDNA-based methods makes them ideal for detecting the presence of species when efforts to detect low-density species would be unmanageable, such as the presence and distribution of low-density invasive, elusive, or threatened species (Dejean et al., 2012; Takahara et al., 2013; Rees et al., 2014). The non-invasive nature of eDNA analysis may provide advantages over traditional capture-based sampling by allowing the presence or absence of species to be determined without disturbing the fish or their environment. This approach could be particularly beneficial in endangered species situations, where there is a significant risk of injury to fish or damage to critical habitat (Evans and Lamberti, 2018). For endangered species in particular, a non-invasive genetic sampling technique that could provide information on absence/presence data and even estimates of population size, would therefore be of great use for the conservation and management of these species. In addition, eDNA approaches may offer a cost-effective way of obtaining basic distribution and abundance data, and allow limited conservation resources and taxonomic knowledge to be efficiently deployed to maximise returns (Rees et al., 2014). These points have highlighted the potential of eDNA analysis to improve assessments of rare species, as well as to assist in routine fisheries sampling. Several studies have illustrated that eDNA analysis can reliably detect the presence of fish in marine ecosystems where they are known to occur through catch-based sampling (see references in Supplementary Table S1).
The effectiveness of eDNA has been evaluated in comparison with traditional monitoring techniques (e.g., Knudsen and McDonald, 2019; Russo et al., 2021; Stoeckle et al., 2021). The results of most of these studies indicate that eDNA has higher detection rates and has a higher cost-effectiveness or higher catch per unit effort than traditional sampling methods. Scaling up long-term biomonitoring programs with eDNA could improve taxon detection and resolve long-term patterns or changes in species of interest (Berry et al., 2019). eDNA assessment can be used for biomonitoring of pelagic and benthic ecosystems targeting fish (Russo et al., 2021; Stoeckle et al., 2021; Valsecchi et al., 2021), mammals, seabirds (Ushio et al., 2018), and sea turtles (Kelly et al., 2014). Despite the advantages of eDNA detection, there is key ecological information, such as fish recruitment and size/age classes, that can only be obtained through traditional monitoring.
A virtual workshop was held on 28 May 2020, bringing together a broad cross-section of experts from the genetics and fisheries assessment communities to discuss the state-of-the-art and identify barriers and advantages for the application of HTS techniques on stocks developed under the FishGenome contract “Improving Cost-Efficiency of Fisheries Research Surveys and Fish Stocks Assessments using Next-Generation Genetic Sequencing Methods” (Figure 2). In particular, there was a section discussing the use of eDNA approaches in fisheries assessment. One of the main concerns with the use of eDNA that was pointed out is that, to date, the spatial distribution of eDNA in seawater column is unknown. The origin of this genetic material cannot be easily determined and its impact can usually be corrected and buffered by repetition and proper sampling design. The poor quality of existing public databases was also identified as a problem, as they contain significant errors that can mislead in the interpretation of analyses. For this reason, the construction of a specific database, containing the species that inhabit the studied environment, can be considered a recommendable alternative.
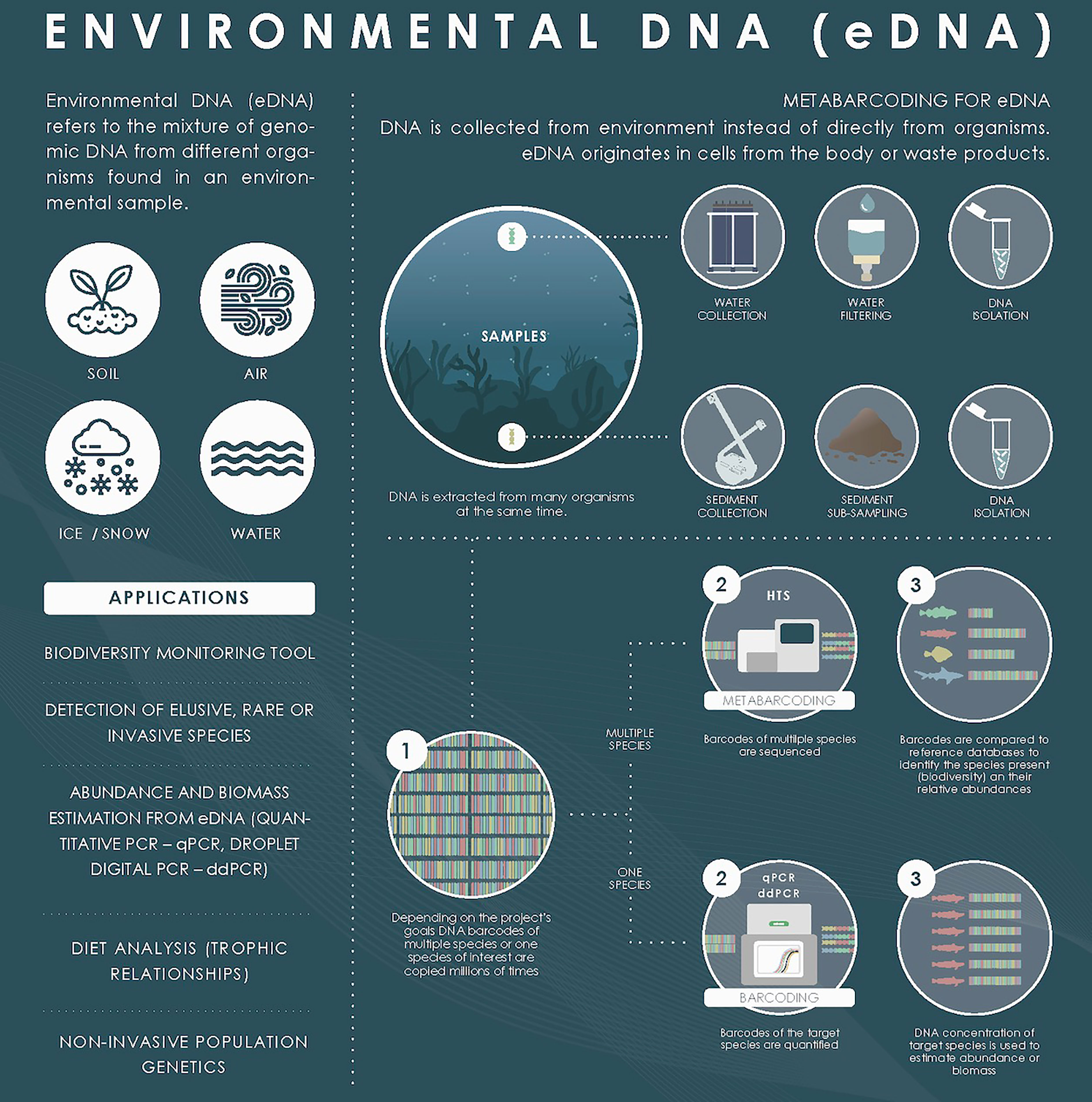
Figure 2 Scheme of eDNA used in the FishGenome contract virtual workshop “Improving Cost-Efficiency of Fisheries Research Surveys and Fish Stocks Assessments using Next-Generation Genetic Sequencing Methods”. The definition, applicability and methodology of eDNA from samples collected from sediment and seawater are indicated.
Regarding the possibility of using eDNA to estimate biomass abundance by qPCR, it seems that further research is needed to calibrate the tool. Once the amount of DNA representing a given fluorescence signal is determined, the biomass of that resource must be calibrated with the amount of DNA, which may depend on many factors. However, some relationships have been found in metabarcoding, indicating that qPCR may work even better for certain species. If the objective is to quantify a species and continuous monitoring is performed, there is a possibility that qPCR can provide some relevant information (there is experience in this regard for detecting presence peaks). It is not yet known how close or how far we are from the goal of real biomass quantification that can be integrated into assessment models by metabarcoding.
As final notes during the workshop, experts suggested that beyond the measurement of species presence and distribution, the implementation of eDNA could provide information on species co-occurrence and relationships across ecological networks, with environmental parameters. This would improve quantifiable ecological information of great interest for biodiversity management and assessment.
eDNA behavior
Marine environments are probably the most difficult and challenging aquatic samples to apply the eDNA method. This is due to the extreme ratio of water-volume to biomass, the effects of sea currents and wave action on eDNA dispersion and dilution, the impact of salinity on eDNA preservation and extraction (Thomsen et al., 2012b). To date, studies of eDNA have mainly focused on proof-of-concept, and further research is needed on the ‘ecology’ of eDNA —release and concentration, degradation, and transport—and its influence on detection, quantification, analysis, and application of eDNA to assessment and conservation (Barnes and Turner, 2016; Thomsen et al., 2016; Hansen et al., 2018; Figure 3). Understanding the origin of eDNA and the physical, chemical, and biological factors that affect eDNA concentration and influence its production, persistence, and transport in marine ecosystems can contribute to our knowledge of the taxa and environments for which eDNA represents an effective tool (Barnes and Turner, 2016).
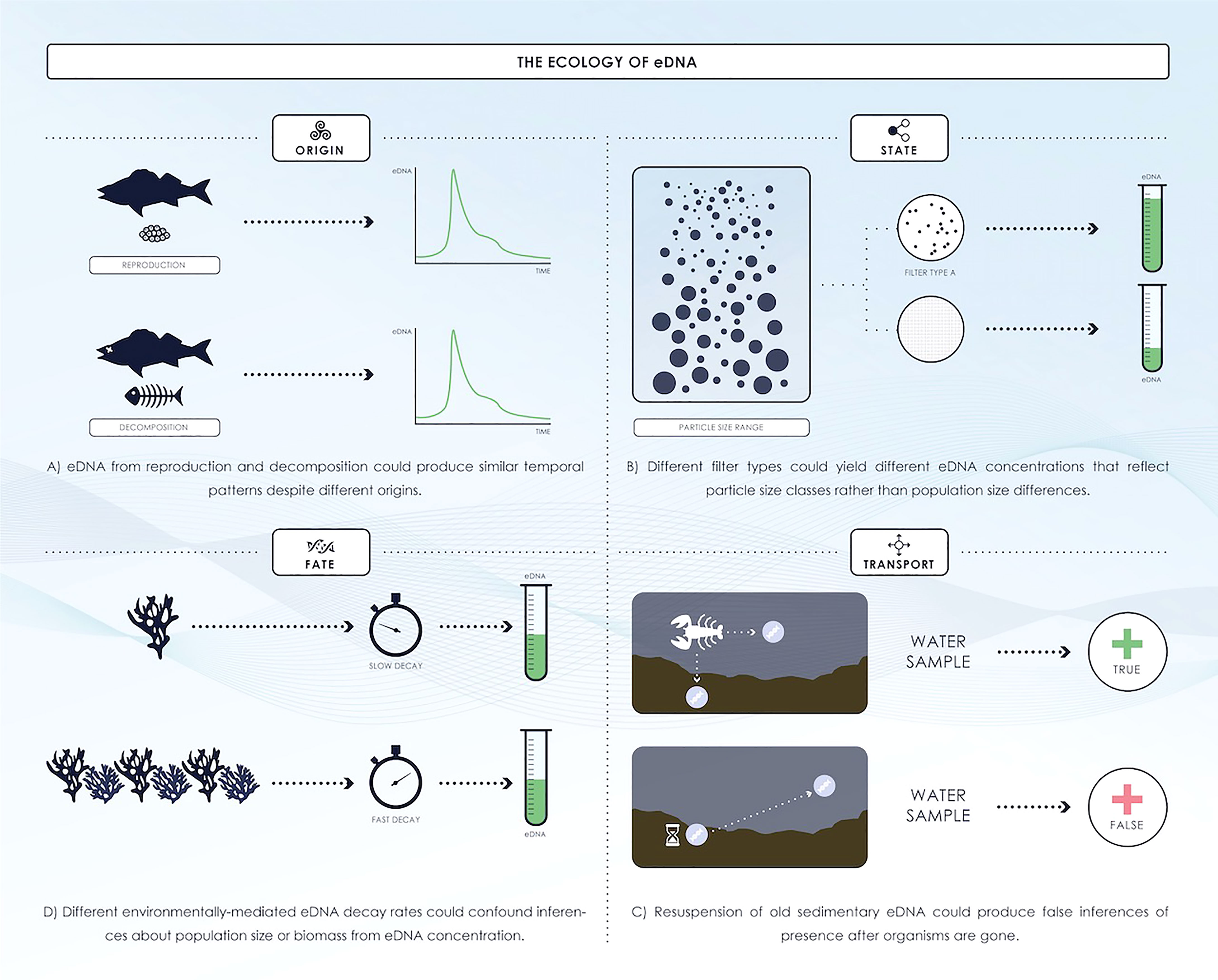
Figure 3 eDNA ecology affects population inferences: Origin, State, Fate and Transport (redrawing from Barnes and Turner, 2016).
eDNA release and concentration
The concentration of eDNA varies by several orders of magnitude between different environmental samples, reflecting the environmental conditions in which DNA is preserved. This concentration in the environment limits the scope of eDNA studies, as often only small segments of genetic material remain. The rate of release and degradation of eDNA, which can be strongly altered by environmental conditions, determine the concentration of eDNA in seawater samples (Lacoursière-Roussel et al., 2016b).
The production of eDNA depends on the biomass, age, and feeding activity of organisms, as well as physiology, life history, and space use (Barnes and Turner, 2016; Goldberg et al., 2016). Different experiments indicate that eDNA concentration is positively correlated with individual biomass (e.g., Pilliod et al., 2014; Evans et al., 2016; Piggott, 2016). However, a point of caution is suggested because differences in size between species (biological differences) and within a species (e.g., age structure or morphometrics) are likely to influence eDNA production and biomass interpretations (Stewart, 2019). Differences due to life history stage have been recurrently observed. For example, incongruities in eDNA production between juveniles and adults have been demonstrated in fish (Maruyama et al., 2014), amphibians (Goldberg et al., 2011; Thomsen et al., 2012b), and invertebrates (Tréguier et al., 2014). In aquaria-based experiments with bluegill sunfish (Lepomis macrochirus), juveniles were observed to have slightly higher excretion rate than adults, possibly due to ontogenetic reduction of metabolic activity in adults (Maruyama et al., 2014). The risk of using measures of eDNA abundance in wild populations without accounting for age-structure would lead to incorrect estimates of population abundance, especially if populations are dominated by one or another age class (Maruyama et al., 2014), such as in dwindling populations with low birth rates, or in newly founded populations monopolized by juveniles.
Species-specific differences may also affect the quantity and quality of eDNA production, which is strongly influenced not only by size but also by the ecology of the target taxa (Stewart, 2019). Indeed, studies have reported disparities in eDNA sources between and within taxonomic groups (e.g., Goldberg et al., 2011; Thomsen et al., 2012b; Sassoubre et al., 2016). For example, eDNA detection was shown to vary between both cownose ray (Rhinoptera bonasus) and Brazilian cownose ray (R. brasiliensis) suggesting that disparities between species are related to their migration seasons (Stoeckle et al., 2020). Thomsen et al. (2012b) also found that eDNA production rates differed between two juvenile amphibian species and two fish species.
On the other hand, some studies found increases in the abundance of eDNA signals during breeding seasons (e.g., Bista et al., 2017; Stoeckle et al., 2017). For example, eDNA signals have been shown to be triggered during fish spawning, suggesting strong seasonal influences on eDNA detection (e.g., Laramie et al., 2015; Erickson et al., 2016). Similar results have also been observed in amphibians and reptiles, presumably reflecting species-specific behaviour (de Souza et al., 2016) such as male–male combat and mass release of gametes (e.g., Eastern Hellbender, Cryptobranchus alleganiensis; Spear et al., 2015). Genetic material such as gametes, blood, and other reproductive tissues (e.g., placenta, lactation, etc.) combine to make breeding events optimal for eDNA detection in wild populations. However, this overproduction of DNA at breeding times may also overestimate the relative presence of a species both temporally and spatially (Stewart, 2019).
The response of organisms to the presence of others is another factor to consider. For instance, it is well-known that predators exert physiological impacts on their prey, including reduced food intake, increased metabolic rate, and elevated stress (Boonstra, 2013; Van Dievel et al., 2016). Although the exact impact of this interaction on eDNA production rates is unknown to date, it is likely to affect these processes. It has been suggested that acute perturbations of physiological homeostasis, such as stress, may have effects on sources of genomic material (Pilliod et al., 2014; Klymus et al., 2015). Several studies have observed an increase in eDNA production due to osmotic changes or following the handling of animals (Maruyama et al., 2014; Pilliod et al., 2014). Increased density (McKenzie et al., 2012), social stress (Sloman et al., 2000), and shelter availability (Milidine et al., 1995) are other examples of stresses affecting the metabolic activity of aquatic organisms, with potential impacts on eDNA sources, that warrant further investigation.
eDNA degradation
eDNA degradation must be considered in eDNA studies, as it can reduce the detectability of species over time. The degradation rate can vary from hours to weeks, depending on the environment and target species (Thomsen et al., 2012b; Maruyama et al., 2014; Balasingham et al., 2017; Barnes et al., 2014; Sassoubre et al., 2016). In sediments and terrestrial soils, a very low proportion of DNA can persist for long periods, adsorbed to organic or inorganic particles that protect it from several potential degradation agents. Dell'Anno and Corinaldesi (2004) demonstrated that in marine sediments the turnover of extracellular DNA is ca. 200 times slower than in sea water (up to 93 days in sediments versus 10 h in seawater). DNA persistence is strictly dependent on the nature of the sediment, e.g., in loamy sediment, persistence time may be similar to persistence in the water column (Deere et al., 1996). However, under certain conditions, DNA can be preserved for hundreds of thousands of years, as observed by Coolen and Overmann (2007) who were able to analyse DNA in 217,000-year-old anoxic sediments. In marine environments, eDNA degrades faster than in freshwater environments, and can be used to obtain a “snapshot” of the species present in that particular environment at the time of sampling, or a few days or weeks earlier.
On the other hand, in marine and freshwater environments, different studies have evidenced that eDNA persistence can vary throughout the water column. In this sense, Matsui et al. (2001) reported a greater degradation of eDNA in the epilimnion (the upper, warmer layer of a thermally stratified lake and more exposed to UV radiation) than in the hypolimnion (the lower, colder layer of a thermally stratified lake). However, this difference in detection time appears to be due to current flow and related dilution, rather than DNA degradation, which is an important variable when dealing with a dynamic system such as the ocean.
eDNA transport
Once released into the environment, eDNA is transported away from organisms and begins to degrade. To better understand the distribution of eDNA in relation to species distribution, studies have begun to examine how this complex DNA signal is transported horizontally and vertically in aquatic environments, as the process of eDNA transport or diffusion is fundamental to sampling design and spatial inference. Understanding this transport is essential to relate the detected eDNA to the presence of species both in space (i.e. how close a species was to the location of eDNA detection) and in time (i.e. how recent the presence of the detected species was). For important conservation applications, being able to connect a positive eDNA detection within specific spatial and temporal boundaries is essential for drawing robust conclusions (Barnes and Turner, 2016).
In marine environments, where long-distance transport is possible, eDNA transport remains a problem (Thomsen et al., 2012a). Vertical transport (i.e. settling) of fish eDNA accumulation in sediments has also been described (Turner et al., 2015). Resuspension of sedimented eDNA within water, considering the high concentrations of sedimented fish eDNA that have been observed, could represent an important element of eDNA ecology (Turner et al., 2015). As eDNA is currently a tool used for contemporary biodiversity monitoring and conservation, it is crucial that the results reflect the current state of an ecosystem. In this regard, it is important to consider the fact that eDNA in soil appears to be able to persist for decades and centuries (Andersen et al., 2012; Yoccoz, 2012) and the potential release of “ancient” eDNA from bottom sediments into water column could complicate the use of aquatic eDNA as strict contemporary biodiversity surveys.
Factors influencing the detectability of eDNA
The factors that influence the persistence of eDNA in aquatic environments have been the subject of different studies, concluding that they fall into two broad categories: (i the abiotic environment (i.e. temperature; UV radiation, salinity); and (ii the biotic environment (i.e. composition and activity of the microbial community and extracellular enzymes) (Barnes and Turner, 2016). These factors can affect both production and degradation of eDNA.
Abiotic environment
This factor plays a major role in the rate of DNA release and degradation in marine environments. Water temperature can affect the release of DNA from organisms and thus the availability of eDNA for detection. The effect of temperature on DNA release can be due to different reasons. For example, fish metabolism, growth, physiology, and immune function are influenced by water temperature (Engelsma et al., 2003; Person-Le Ruyet et al., 2004; Takahara et al., 2011). As a by-product of metabolic influences, evidence suggests that temperature additionally affects the production of feces and urine in fish (Selong et al., 2001; Gale et al., 2013), presumably the main component of eDNA sources. Fish mobility increases with water temperature (Petty et al., 2012), so genetic signals may also be more homogenised and/or spatially dispersed. The immune response can also facilitate mucus excretion and epithelial cells shedding. Studies to date have found conflicting results; in mesocosm experiments, no effect was found in two studies (common carp, Cyprinus carpio, Takahara et al., 2012; bighead carp, Hypophthalmichthys spp., Klymus et al., 2015), while there was a significant increase in production rates in Mozambique tilapia (Oreochromis mossambicus; Robson et al., 2016). In a field study, high water temperature significantly increased the amount of brook charr (Salvelinus fontinalis) eDNA within the water column and, moreover, biomass and thus predictability of population abundance increased at higher temperatures (Lacoursière-Roussel et al., 2016a, Lacoursière-Roussel et al., 2016b). In addition, temperature itself may affect the excretion of genetic material into the environment when phenologies simultaneously affect other physiological (e.g., metabolic regulation) or behavioural responses (e.g., temporal avoidance) of the organism.
Another such factor is the effect of UV radiation on detectability of eDNA. It was shown that eDNA was no longer detectable in samples exposed to full-sun after 8 days, but it was possible to detect eDNA in samples that were stored in the dark after 11 and 18 days, demonstrating the direct effect of UV radiation on eDNA (Pilliod et al., 2014).
Adaptation to saline environments also requires physiological compensation and acclimatization. In most marine fish species, egg fertilization and incubation, early embryogenesis, swim bladder inflation, and larval growth are salinity-dependent (Boeuf and Payan, 2001). In fact, studies have shown that up to 50% of the total energy of fish can be devoted to osmoregulation (Bushnell and Brill, 1992), and that food intake, feed conversion, and hormones associated with growth regulation depend on environmental salinity (Boeuf and Payan, 2001). Smoltification of salmon, for instance, demonstrated a drastic physiological adaptation to seawater, resulting in a significantly different metabolism than their freshwater counterparts (e.g., McCormick et al., 1989).
Widespread links between salinity and fish growth have been demonstrated for both marine and freshwater species, with general patterns suggesting that growth rates of marine species increase in slightly more saline environments, while development of freshwater species shows the opposite relationship (Boeuf and Payan, 2001). While assessment of marine species richness and approximate abundance is a relatively new foray for eDNA (e.g., Günther et al., 2018; Knudsen and McDonald, 2019), it has proven successful for accurate detection, but read abundance has failed to find to be easily correlated with DNA ratios (Günther et al., 2018) or traditional visual measures of biomass, such as trawling (Knudsen and McDonald, 2019).
Biotic environment
eDNA studies showed that DNA persistence in aquaria experiments, where water temperature and sunlight conditions were the same, ranged from one week to one month (Dejean et al., 2011; Thomsen et al., 2012b; Piaggio et al., 2014). Differences in DNA persistence time observed in these studies could be explained by differences in animal density. Endogenous nucleases are another factor influencing the amount of eDNA in the environment (Hebsgaard et al., 2005). Furthermore, disruption of the cell structure releases DNA and cellular fluids into the environment. This, in turn, stimulates the growth of microorganisms and leads to further degradation of DNA by their exogenous DNases (Hebsgaard et al., 2005; Willerslev and Cooper, 2005). Temperature also has an important influence on the action of endonucleases and microorganisms, as at low temperatures these activities can be slowed down or even inactivated (Hofreiter et al., 2001; Zhu, 2006).
Hydrolysis and interstrand crosslinks are another source of DNA damage (Herder et al., 2014). They influence the accessibility to DNA-polymerases and prevent DNA strand cleavage, which blocks DNA replication (Noll et al., 2006). Consequently, amplification of DNA extracted from an environmental sample (e.g., water, soil, sediment) is prevented (Hansen et al., 2006), and the species will not be detected. Decreased DNA detection in the water column could also be due to uptake of DNA by sediments and organic matter present in the water (Deere et al., 1996). Corinaldesi et al. (2008) investigated which environmental factors (temperature, salinity, organic matter loads, and redox potentials) could affect extracellular DNA damage and degradation rates in various marine sediments, showing that extracellular DNA damage rates do not depend on a single factor (e.g., temperature) but on a complex interaction of different factors. In addition, fish physiology such as stress (Pilliod et al., 2014), breeding readiness (Spear et al., 2015), feeding behaviour (Klymus et al., 2015), and metabolic rate (Maruyama et al., 2014) may also play a role in eDNA degradation.
eDNA methodology notes
The widespread implementation of eDNA methods in species monitoring within fisheries assessments is currently prevented by the lack of rigorous standards for both sample collection and bioinformatics analysis.
Metabarcoding
This approach commonly uses universal primers that amplify DNA from a group of target species (e.g., fishes, crustaceans, echinoderms). The amplified fragments are then sequenced using an HTS platform. These new technologies allow to sequence DNA molecules present in the mixture and the simultaneous sequencing of millions or billions of molecules. Furthermore, several independent samples can be multiplexed in a single run. The workflow of the DNA metabarcoding approach is generally based on a double indexing and two-step PCR pipeline. In order to allow the clustering of several samples in the same sequencing run, double indexing is used, where sample-identification barcodes are incorporated in the Illumina adapter sequences, both forward and reverse. Barcodes should not be included in the region-specific primer in the first PCR. These barcodes are short sequences, 3 to 14 nucleotides in length, that should be distinct from each other and can have error-correcting properties to protect against the sequence alterations introduced during synthesis, amplification or sequencing (Krishnan et al., 2011). The first PCR is carried out with primers targeting the region of interest, but these primers already include the overhang adapters for the barcodes and the sequencing adapters. In the second PCR, the sequencing index adapters and barcodes are incorporated into the amplicons. For example, when Illumina indexes are used, a small multiplex identifier is added to the overhang to allow additional pooling of samples for sequencing. The final library should be quantified with qPCR or TapeStation or Bioanalyzer, otherwise the sequencing run will fail due to the inaccurate quantitative data.
The analysis of millions of sequences produced by the eDNA metabarcoding method requires efficient, automated and yet flexible analysis pipelines to translate the raw sequences into a statistically exploitable contingent matrix containing (Dufresne et al., 2019). Sequence clustering can be reference-based if sequences are assigned to a cluster because they are sufficiently similar to a sequence in a specific reference database. Therefore, taxa that are not included in the database will not be clustered, with the consequent loss of biological variation. The most commonly used clustering algorithms are based on the generation of clusters of sequences that differ by less than a fixed sequence dissimilarity threshold. The representative sequences of these clusters are referred to as molecular Operational Taxonomic Units (OTUs). However, the similarity thresholds used to delimit OTUs are arbitrary and depend on the variability of the genomic region and the targeted taxonomic groups and, more importantly, they have been shown to strongly affect molecular biodiversity inventories (Brown et al., 2015; Tapolczai et al., 2019). Recently, new methods have been developed to generate Amplicon Sequence Variants (ASVs), also known as Exact Sequence Variants (ESVs), Zero-radius OTUs (ZOTUs) or an OTU defined by 100% sequence similarity. The imposition of arbitrary dissimilarity thresholds is avoided by using a de novo process that control the errors contained in the dataset sufficiently such that the ASVs sequences only differ by as little as a single nucleotide (Eren et al., 2013; Callahan et al., 2016).
Different algorithms and software have been developed to perform single or multiple processing steps. New applications have even been developed on website platforms that help users to create and execute their own metabarcoding pipelines, such as OBITools (Boyer et al., 2016), DADA2 (Callahan et al., 2016), MiFish (Sato et al., 2018), Anacapa (Curd et al., 2019), Barque (Mathon et al., 2021), metaBEAT (metaBarcoding and eDNA Analysis Tool) v0.8 (https://github.com/HullUni-bioinformatics/metaBEAT), Charybdis (https://github.com/cbirdlab/charybdis)). These pipelines can be modified according to the user’s needs and in order to achieve more accurate and reliable results (Antich et al., 2021; Kim et al., 2021).
Once the HTS output data have been bioinformatically analysed, they can be compared with a reference database, although this can be a source of bias. When using public databases (e.g., GenBank, Ensembl, BOLD systems) as a reference database, the high number of sequencing errors (Harris, 2003) and mislabeled species (Santos and Branco, 2012) must be considered. One solution, to overcome this problem, is the construction of a private databases in which sequences, species labeling, and geographic origin are carefully verified, as in Meta-Fish-Lib reference library hosted at https://github.com/genner-lab/meta-fish-lib, or MetaZooGene Barcode Atlas and Database (https://metazoogene.org/MZGdb), or use methods to identify mislabel species (e.g., SATIVA, https://github.com/amkozlov/sativa).
Quantitative PCR
This approach is commonly used to assess biomass and abundance of marine communities, including fish (Salter et al., 2019; Knudsen et al., 2019). Two types of chemistries are commonly used to detect PCR products using real-time PCR instruments: SYBR® Green and TaqMan®. Initially, intercalator dyes were used to measure real-time PCR products, with the main drawback that they detected the accumulation of both specific and non-specific PCR products. Currently, SYBR® Green method has two requirements for a DNA binding dye for real-time PCR detection: increased fluorescence when bound to double-stranded DNA, and no inhibition of PCR. On the other hand, the TaqMan® method uses a fluorogenic probe that allows the detection of a specific PCR product as it accumulates during PCR.
In addition, droplet digital PCR (ddPCR) is also recently being used for eDNA. This to nucleic acid detection and quantification method offers an alternative method to conventional real-time qPCR for absolute quantification and detection of rare alleles. Digital PCR works by partitioning DNA or cDNA sample into many individuals and performing parallel PCR reactions; some of these reactions contain the target molecule (positive) while others do not (negative). A single molecule can be amplified a million times or more. Nowadays, the most widely used method for quantification of abundance or biomass in aquatic environments is TaqMan chemistry (e.g., Sassoubre et al., 2016; Doi et al., 2017; Atkinson et al., 2018; Levi et al., 2019).
Workflow
In general, both eDNA approaches involve a series of steps that include eDNA capture, preservation, extraction, amplification, and sequencing to ensure detection of target species (Figure 4). Efficiency at each step is expected to affect DNA recovery and, consequently, detection. Researchers often choose methods based on cost, ease of sampling, and availability of materials and equipment. Since eDNA typically exhibits high levels of degradation and is often of poor quality, DNA recovery could vary depending on the quality of the protocol used, so methods that maximise eDNA recovery in a cost-efficient manner are ideal (Hinlo et al., 2017). Several research groups have developed a great variety of protocols for eDNA detection from different marine species and environments. Currently, there is a wide variety of protocols for sampling and analysis of eDNA data, which must be optimized according to the objective of each study. This is a characteristic of emerging scientific fields, and we believe it is essential now, at this juncture, to develop minimum standards of quality assurance.
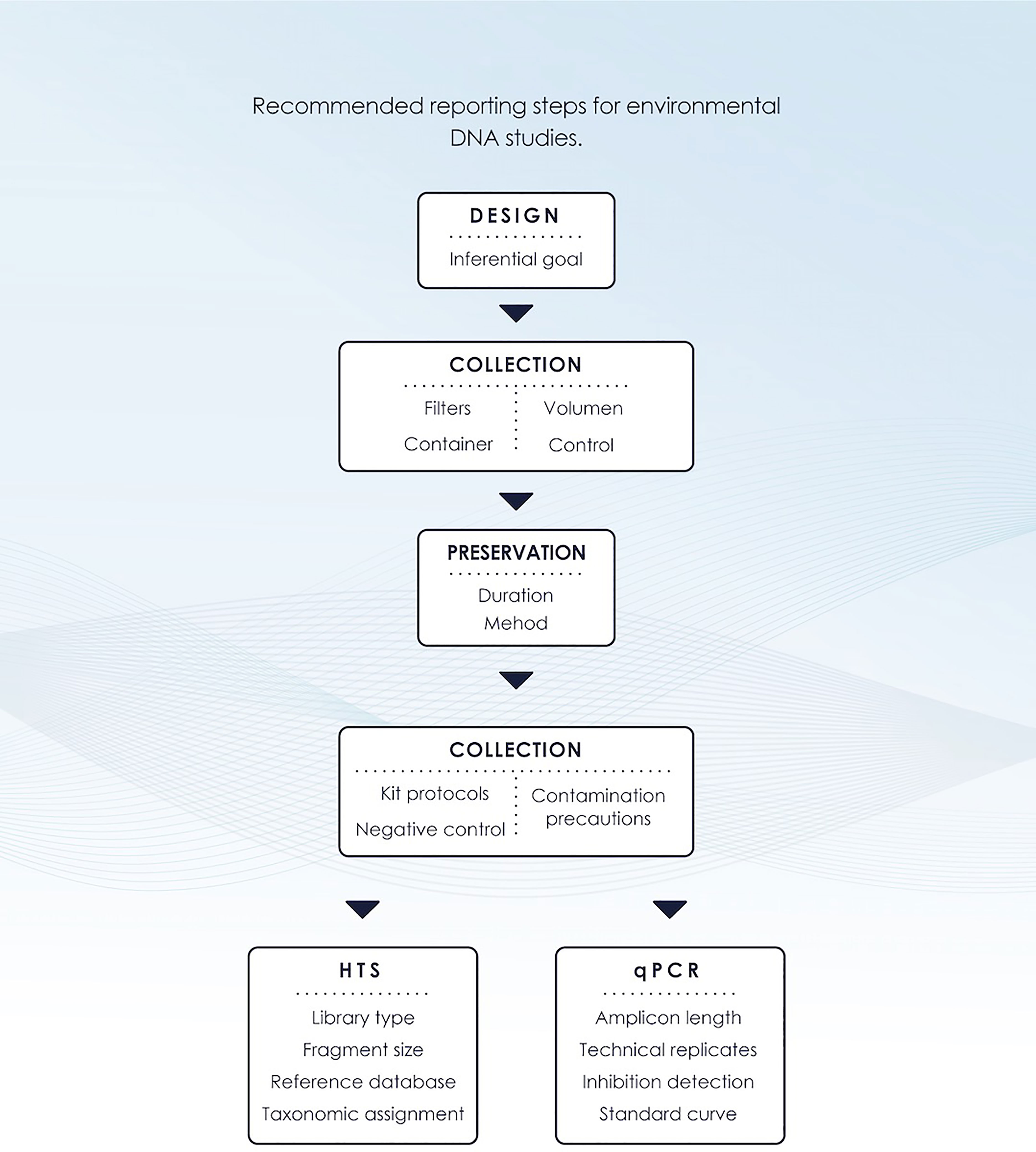
Figure 4 Schematic workflow designed for environmental DNA studies. In general, eDNA studies include the following steps: experimental design, collection, preservation, extraction, and analyses that include High-Throughput Sequencing (HTS) and/or quantitative PCR (qPCR).
Samples
Aquatic samples: In general, two methods are used to capture eDNA from aquatic environments, filtration and precipitation. Filtration requires passing water samples through a filter to trap DNA, and allows larger volumes of water (commonly 250 ml - 5 L; Supplementary Tables S1, S2) to be processed. Water samples are filtered on-site or stored on ice for travel and then filtered in the laboratory. Meanwhile, the precipitation method uses ethanol to precipitate nucleic acids in water (Jerde et al., 2011; Hinlo et al., 2017). Although both methods have shown variable success rates in comparative studies, it is recognized that the filtration method recovers more eDNA from water samples than the precipitation method (e.g., Deiner et al., 2015; Eichmiller et al., 2016; Hinlo et al., 2017), and immediate preservation is generally recommended (Supplementary Tables S1, S2).
In addition to this, the filter material, such as pore size, filter material, and DNA extraction method, could affect the quality and quantity of eDNA, resulting in different final detection rates (Deiner et al., 2015; Renshaw et al., 2015). eDNA studies on marine environments use frequently pore sizes ranging from 0.2μm – 0.45 μm, while the filtration volume commonly ranges from 250 ml to 2 L (Supplementary Table S1, S2). Otherwise, eDNA samples have been effectively collected with different filter material such as cellulose nitrate, glass fiber, polycarbonate, nylon, polyethersulfone and cellulose acetate (Supplementary Tables S1, S2). Hinlo et al. (2017) conducted experiments to compare the recovery of eDNA at different stages of the analysis to determine which methods are most cost-effective, concluding that the most recommended filtration process was the use of cellulose nitrate filters. Finally, filters (and collected materials) can be preserved by freezing (Jerde et al., 2011), immersion in ethanol (Goldberg et al., 2011), drying, or immersion in cell lysis buffer (Renshaw et al., 2015). It is recommended to filter seawater samples within 24 hours, but if this cannot be done, short-term refrigeration (72 hours at 4°C) could be performed (Hinlo et al., 2017).
Sediment or soil samples: Recent studies have shown that, in general, extra membranous eDNA is found in higher concentration in sediments than in the overlying water column, as DNA from water columns can progressively accumulate in sediments (Corinaldesi et al., 2008; Turner et al., 2015). Marine sediments supposedly harbor one of the richest species reservoirs on Earth, but logistics and the scarcity of taxonomic specialist make it difficult to understand their biodiversity, and more so in the case of deep-sea sediments (Grassle and Maciolek, 1992; Snelgrove, 1999). The persistence of eDNA is strictly dependent on the nature of the sediment (Deere et al., 1996). In marine environments, DNA molecules degrade faster than in freshwater environments. Because of this, it can be used to give a “snapshot” of the species present in this particular environment at the time of sampling, or a few days or weeks earlier (Collins et al., 2018). eDNA preserved in sediment or soil samples can be used to obtain an integrative picture of present or past biodiversity (Herder et al., 2014). The high concentration and long persistence of fish eDNA in sediments can help to know the temporal and spatial scales from aquatic eDNA (Bloesch, 1995; Douville et al., 2007). According to Turner et al. (2015) fish eDNA in aquatic sediments may be a promising source of historical genetic materials.
In several studies, samples are collected from the sediment surface, which contains suspended material within the water column, including whole cells and extracellular DNA (Levy-Booth et al., 2007; Guardiola et al., 2015; Holman et al., 2019). Marine sediments are known to have key ecological functions and ecosystem services, and are sensitive to anthropogenic disturbances. However, they have been poorly studied by traditional means, and are not well understood because taxonomic work to describe species found in marine sediments is difficult and rarely undertaken (Guardiola et al., 2015). The potentially extensive persistence of DNA bound to sediments is very valuable, but can be difficult to identify when the target species was present at the sampling site. In fact, detection of eDNA also varies according to sediment texture, so the amount to process is also variable. Typically, the volume of sediment samples processed is between 10-50 g of sediment surface (Guardiola et al., 2015; Holman et al., 2019).
To obtain accurate results from seawater and sediment samples, strict and clean collection protocols must be followed. Decontamination of collecting equipment is essential to maintain sample independence and samples should be preserved in sterile containers. In fact, single use supplies for eDNA collection can significantly reduce the risk of contamination (Goldberg et al., 2016). Bottles for eDNA sample collection should be cleaned with 10% bleach and washed with DNA-free distilled water.
Equipment requirements
Environmental DNA samples present the same contamination challenges as other low-quantity DNA samples, such as ancient, forensic, and non-invasive genetic samples (Herder et al., 2014). eDNA samples should be handled and stored in a dedicated room that is physically separate from rooms where high quantity DNA extraction and PCR products are handled (Taberlet et al., 1999). Laboratories should be organized in order to avoid contamination as much as possible. All eDNA extractions must be carried out in an isolated room ideally equipped with positive air pressure, overnight UV treatment, and air renewal. Pre-amplification and post-amplification work should be performed in separate rooms, distant from each other, ideally in different rooms. DNA extraction and PCR mix preparation should be carried out in the pre-amplification rooms, while PCRs will be performed and the PCR results analysed in the post-amplification room. Control samples without DNA should be extracted at the same time and used as negative controls. Positive PCR controls and qPCR standards should ideally be added in a third room assigned as an intermediate DNA level room, between the pre-amplification and the post-amplification room (Herder et al., 2014). For all laboratory procedures, filter pipette tips and clean gloves should be used.
eDNA extraction
Several DNA extraction protocols have been optimized and applied in eDNA studies. The choice of protocols may affect species detectability and sample diversity, and different approaches may be required, depending on the objective of the study. Different aspects should be considered when selecting the best strategy and protocol for DNA extraction: i) the proportion of sample used for extraction; ii) the sampling or subsampling strategy for DNA extraction; and iii) the detection of the presence of inhibitors. The most frequently used protocols for eDNA studies are: 1) Qiagen DNeasy Blood & Tissue Kit (Qiagen, Germany); 2) MO BIO’s Qiagen PowerWater DNA Isolation Kit (Qiagen, Germany), 3) CTAB (Cetyltrimethylammonium bromide)-chloroform and phenol-chloroform, 4) Qiagen DNeasy Powermax Soil (Qiagen, Germany), and 5) Qiagen DNeasy Powersoil (Qiagen, Germany). Phase separation and precipitation methods for DNA extraction (e.g., CTAB protocol) typically yield more DNA than silica column methods (e.g., Qiagen kits). An initial step is recommended for sediment samples, which consists of mixing the sediment sample with an equivalent volume of phosphate buffer (0.12 M Na2HPO4; pH=8) and then homogenizing the mixture in a shaker for 15 minutes (Holman et al., 2019).
Final product of eDNA extraction is commonly quantified using the NanoDrop (Thermo Scientific) or Qubit (ThermoFisher) system in order to determine DNA concentration. With a Qubit fluorometer, eDNA studies are enhanced by more accurate measurements, as it detects fluorescent dyes that are specific to the target of interest (e.g., DNA, RNA, or protein) in the sample, even at low concentration.
Marker selection: DNA mitochondrial vs nuclear
Mitochondrial DNA (mtDNA) is often targeted because of its high number of copies compared to nuclear DNA, its efficiency in identifying organism to species level by DNA barcoding and its accessibility through universal sequence databases on public servers (e.g., GenBank and BOLD systems). Amplified mitochondrial eDNA can come from extracellular DNA fragments, mitochondria, cells, excretion, or eggs, and the amount of quantified eDNA is likely to vary depending on the target genetic material collected (Herder et al., 2014; Goldberg et al., 2016). Although there is a clear preference for the use of markers based on mtDNA in metabarcoding studies, other types of eDNA were found to be available for this purpose, as multi-copy nuclear eDNA (e.g., ribosomal RNA genes, microsatellites; Günther et al., 2018; Andres et al., 2021; Jo et al., 2022) even though nuclear genes evolve slowly and may diminish diversity (Hillis and Dixon, 1991; Castro et al., 1998). According to Jo et al. (2021) copies of nu-eDNA may provide a more recent estimation of species abundance if its production and degradation rates are higher than those of mtDNA. In addition, using nu-eDNA may enable the evaluation of genetic diversity in a population with a higher resolution than mt-eDNA (Sigsgaard et al., 2020).
The goal of marker selection is to use a portion of the mitochondrial genome of the target species that is species-specific, with the appropriate fragment size, and amenable to accurate primer binding (Bohmann et al., 2014; Rees et al., 2014). Therefore, one of the most important considerations in eDNA studies is the design of PCR primers. Different primers and regions differ in coverage, resolution, and bias between taxa. Short DNA fragments (around 150 bp) degrade slowly and are easier to recover from environmental samples (Herder et al., 2014), so primers for eDNA studies need to amplify a short fragment. This fragment should be variable to amplify a variety of species without sacrificing the specificity of the target group (Epp et al., 2012).
For both metabarcoding and qPCR, species-specific and general primers can be used, depending on the goal of the study and the available budget. Species-specific primers may be needed to ensure detection of specific species (endangered, invasive, elusive, or rare taxa). In contrast, general primers allow detection of a wide range of species, but may not detect less abundant taxa (Thomsen et al., 2012a). The detection power of general primers will depend on i) the affinity to the sequences of target taxa, and ii) the availability of databases of DNA reference collection necessary for species identification. In addition, the choice of primers may bias the results by preferentially amplifying some target sequences more than others, as well as amplifying non-target groups (Cristescu, 2014). One potential solution to this issue is the use of multiple primer sets, in particular evolutionarily independent primer sets that match standardised barcodes for the target taxonomic groups (Drummond et al., 2015). In this sense, many universal primers have been designed for a short fragment containing sufficient sequence variation to correctly assign fish communities (Supplementary Table S3).
Another important factor in PCR and primer design is the use of appropriate “replicates” since they increase species detection and decrease the likelihood of false negatives. The number of replicates used often differs between studies depending on detection probabilities, research objectives, sequencing depth, primer choice, cost constraints, and sequencing platform (Ficetola et al., 2015; Alberdi et al., 2018). In PCR-based amplifications for HTS, primers can be labeled with short nucleotide sequences to uniquely identify their origin in a process commonly referred to as multiplexing. These tags, while useful, also have the potential to bias results, particularly when located at the 5’ end, and therefore, also require rigorous testing prior to implementation (Binladen et al., 2007; Berry et al., 2012). Because of this, the number of cycles in the indexing PCR is typically kept low to minimize PCR errors (Bohmann et al., 2022).
On the other hand, the qPCR approach is usually performed for species detection and involves the use of species-specific primer sets. Detection of a wide range of species by qPCR would involve the use of a high number of primers sets and increased costs (Lodge et al., 2012; Thomsen et al., 2012a; Lacoursière-Roussel et al., 2016a). For qPCR, short primers that recognize and flank the sequence of interest should be used. The resulting amplicons for qPCR assays are usually short (typically 50-150 bp; Supplementary Table S4), even shorter than those used in HTS platforms. DNA sequence databases for species-specific assay design are also far from complete (Kwong et al., 2012); but, qPCR assays require the development of additional sequence databases.
Quality control
In any DNA metabarcoding and qPCR experiments, it is crucial to include appropriate multiple controls in both sample collection and laboratory procedures in order to monitor for potential contamination and interpret the results correctly (Goldberg et al., 2016). The number of negative controls required at each stage should be determined according to the number of samples and the confidence required in the inference (Sepulveda et al., 2020). In this sense, different types of control can be performed, such as those suggested below:
a) Negative collection control: it is recommended to incorporate an “equipment blank” as a negative control for each filtering and sampling step. In the case of the water sampler, DNA-free distilled water, as a negative control, should be treated identically to the sampled water bottles. The DNA from this negative control shall be analysed with the sample filters, which will allow the identification of any field/transport, filter equipment, or background contamination.
b) Negative extraction controls: a negative control corresponds to a mock DNA sample, a mixture of nucleic acid molecules created in vitro to simulate the composition of a nucleic acid isolated therefrom, which are carried out at the same time and using the same consumables as a normal extraction, except that the sample is omitted.
c) Negative PCR controls: a PCR reaction in which the addition of template DNA is replaced by the addition of water (the same DNA-free water used to dilute the PCR reagents).
d) Positive PCR controls: the ideal positive control should be comparable to the samples analysed (similar concentration and complexity). They can be used to detect contaminants, and artifacts.
e) Tagging system controls: due to the considerable number of samples involved in DNA metabarcoding studies, it is generally necessary to implement a tagging system in which each sample shows a unique combination of forward and reverse tags.
f) Internal controls: an internal control is a template DNA that is added to the PCR mix at a low concentration to produce a small percentage of the final PCR product to act as a internal positive control (IPC). Internal controls are especially useful for estimating the relative amount of target DNA in different samples. They are also suitable for identifying PCR inhibition, even if the amount of PCR inhibitors varies between samples. In the case of eDNA samples, the use of a low amount of IPC that matches the expected concentrations of eDNA (e.g., 100 copies) may better reflect the degree of inhibition affecting the samples, as these will result in non-amplification with approximately the same level of PCR inhibition.
Advantages and limitations
As stated above, the eDNA approaches offer distinct advantages and limitations over traditional monitoring methods.
Advantages
In general, eDNA is a non-invasive method, so it does not damage or alter the species or habitats under study (Thomsen and Willerslev, 2015; Senapati et al., 2019) and is a sensitive method that can detect the target species in its environment. These advantages make this method particularly useful for detecting cryptic and sibling species that are often difficult to identify (Senapati et al., 2019). eDNA-based methods are probably, on a medium to large scale, more cost-effective than traditional methods (Dejean et al., 2012; Herder et al., 2014). However, it will depend on the target species or the community assemblage being studied.
Several studies report shorter handling time and lower cost using eDNA compared to traditional monitoring techniques (Jerde et al., 2011; Biggs et al., 2015; Sigsgaard et al., 2015). In fact, sampling can be performed by one or two persons, thus reducing the cost of sampling. Environmental sample collection is relatively simple compared to traditional monitoring methods, which could simplify a standardised sampling scheme, which can be used in different areas (Thomsen and Willerslev, 2015). Standardisation is a requirement for all monitoring methods in order to compare different sampling areas.
In addition, the high-throughput nature of recent sequencing platforms allows multiplexing of hundreds of samples, thereby providing the means to increase new species records in a given environment (Piper et al., 2019).
Limitations
One of the main limitations of eDNA methods is the high rate of DNA degradation that clearly limits the success of these methods. In some cases, the resulting short fragments may not have sufficient genetic information to allow discrimination between species (Herder et al., 2014). Another important limitation, mainly for metabarcoding technique, is that public genetic databases are incomplete. Therefore, species-level assignment of some fish groups is of low reliability. In the case of qPCR, although the relationship between the density of a species and the amount of eDNA it releases into its environment has been proven in several experiments (e.g., Takahara et al., 2012; Thomsen et al., 2012a), little is known about how external factors (e.g., temperature, depth) influence the persistence and dilution of eDNA, making difficult to have a realistic quantification. Another factor to consider is contamination, which is a serious pitfall of eDNA methods, since it raises the possibility of false positive results. Contamination of samples can occur from sample collection in the field to each step of analysis in the laboratory. The use and sensitivity of HTS has further complicated the contamination issue, as it produces a very high yields of DNA sequences that are likely to reveal tiny amounts of lab-source PCR products (Thomsen and Willerslev, 2015; Ficetola et al., 2016).
Similar to contaminations, erroneous DNA sequences can also lead to biased results. These errors can occur prior to sampling in long-term conserved DNA, during PCR (mutations and formation of chimeric molecules), or during sequencing process (Acinas et al., 2005; Hansen et al., 2006; Thomsen and Willerslev, 2015). Because of this, raw sequence data must be carefully filtered to limit false positives and generate a reliable taxon list. There are multiple sources of error for all detection technologies and it is possible to distinguish between errors attributable specifically to the DNA-based method employed (method errors), and errors that arise during the monitoring process despite the effectiveness of that method (process errors) (Darling and Mahon, 2011). In addition, environmental samples may also contain PCR inhibitors, which can be co-extracted with eDNA. In marine environments, suspended particles (e.g., organic matter and sediment) can clog filters and increase the concentrations of PCR inhibitors (Tsai and Olson, 1992). These external substances could interact with PCR by binding to DNA or prohibiting DNA-polymerase binding, Opel et al., 2010). High concentrations of non-target DNA in marine environmental samples could strongly inhibit enzymes such as Taq Polymerase used in PCR reactions (Matheson et al., 2010; McKee et al., 2015), resulting in failed or delayed amplification of DNA from target species. Both false positive and false negative results can have consequences for the subsequent conservation effort, leading to overestimation or underestimation of the presence of a species, respectively (Goldberg et al., 2016). Furthermore, since eDNA is often distributed in its environment in patches, the sampling strategy can also strongly influence the amount of DNA found in the samples. Traditional monitoring methods provide valuable biological data such as population structure, fecundity and fish condition, this type of data, for now, cannot be provided by eDNA (Herder et al., 2014; Evans and Lamberti, 2018).
Future challenges
At present, there is no consensus on eDNA preservation and isolation protocols, nor on the choice of DNA barcodes and PCR primers, not to mention the debate concerning the parameters for clustering molecular operational taxonomic units and their taxonomic assignment. Standardization of molecular protocols is an urgent needed given the constant evolution and parallel development of new biotechnological tools for DNA data acquisition and analysis. Research efforts should focus on the development of appropriately validated tests to address environmental and sampling factors that may affect eDNA detection, develop competency and proficiency testing for laboratory accreditation, promote inter-laboratory comparisons, and improve and enhance reference databases for DNA analysis. In addition, and given the dynamic nature of marine ecosystems novel studies are required that focus on better understanding the temporal and spatial distribution of eDNA in different marine habitats, to know exactly the relationship between eDNA concentration and species abundance, as well as to understand how biotic and abiotic factors influence the persistence and dispersal of eDNA in different environments. Furthermore, it is important to note that it cannot be stressed enough that the interpretation of eDNA results must go through well-trained taxonomists and ecologists to meaningfully interpret the results and recommend subsequent actions.
It should also be noted that the reference database of bioindicator taxa is far from complete, despite the continuing efforts of numerous national barcoding initiatives. Most existing metabarcoding data are only available locally and are geographically dispersed, which is hampering the development of useful tools at the global level. Considerable effort is still needed to ensure coverage of a range of stressor values at least as wide as that of the development of the traditional methods. In this respect, we believe that eDNA approaches will complement, rather than replace, traditional monitoring. This bioassessment tool can assist fisheries professionals achieve their research, management, and conservation objectives, but not replace time-tested assessment methods. If the costs of eDNA analysis continues to fall, and experience becomes commonplace, eDNA will become an increasingly viable option to complement fisheries monitoring and conservation programs.
It is appealing to imagine the possibilities that eDNA could open up in fisheries ecology and monitoring if advances in molecular ecology, bioinformatics, and sequencing technologies continue to accelerate. Several of the studies mentioned in this work suggest that it will one day be possible to accurately quantify the relative abundance of fish using eDNA analysis (qPCR). In addition to these promising results from eDNA analysis of target fish species, advances in eDNA metabarcoding are now making possible the simultaneous detection of multiple species and the estimation of total species richness from seawater samples. The eDNA methodology, which has already been successfully applied, is likely to be further developed in the near future, leading to increased detection probabilities and reliability (Ruppert et al., 2019).
Third-generation sequencing is likely to provide us with new opportunities to improve the performance of eDNA methods. New generations of powerful technologies, such as single molecular real-time sequencing (SMRT) (from Pacific Bioscience), carbon nanotube chips, and real-time laser transmission spectroscopy, are waiting to be fully tested for their promising potential in eDNA approaches (Lee et al., 2022). The new technology is also being applied in the eDNA collection process. Researchers at NOAA’s Atlantic Oceanographic and Meteorological Laboratory have designed a new instrument that will provide valuable data on biodiversity in marine environments (Formel et al., 2021). These authors have designed a low-cost automated subsurface sampler for eDNA (SASe), which is submersible to 55 m and can filter a programmable volume of seawater and store the eDNA at a collection site.
In the future, if the technology to transmit live data is combined with currently developed technology, the sequence information of interest could be transmitted remotely, with additional overlaid that could help identify how long the eDNA has been in the environment and where it probably originated. In the short term, there is the possibility of using eDNA in population genetics, with, for example, applications for conservation genetics and phylogeography.
Author contributions
SR-A, BT, AP, and CR conceived ideas and obtained specific funding. SR-A and MB searched and collected information related to metabarcoding and qPCR, respectively. All authors contributed to the article and approved the submitted version.
Funding
The information and views set out in this publication are based on scientific data and information collected under Service Contract “Improving cost-efficiency of fisheries research surveys and fish stocks assessments using next-generation genetic sequencing methods [EMFF/2018/015]” signed with the European Climate, Infrastructure and Environment Executive Agency (CINEA) and funded by the European Union. The information and views set out in this publication are those of the author(s) and do not necessarily reflect the official opinion of CINEA or of the European Commission. Neither CINEA nor the European Commission can guarantee the accuracy of the scientific data/information collected under the above Specific Contract or the data/information included in this publication. Neither CINEA nor the European Commission or any person acting on their behalf may be held responsible for the use which may be made of the information contained therein.” SRA (Vicenç Mut Estabilidad) and MB (ForInDoc) were supported by postdoctoral contracts co-funded by the Regional Government of the Balearic Islands and the European Social Fund.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2022.1004674/full#supplementary-material
References
Abdul-Muneer P. M. (2014). Application of microsatellite markers in conservation genetics and fisheries management: Recent advances in population structure analysis and conservation strategies. Genet. Res. Int., 691759. doi: 10.1155/2014/691759
Acinas S. G., Sarma-Rupavtarm R., Klepac-Ceraj V., Polz M. F. (2005) PCR-induced sequence artifacts and bias: Insights from comparison of two 16S rRNA clone libraries constructed from the same sample. Appl. Environ. Microbiol. 71 (12), 8966–8969. doi: 10.1128/AEM.71.12.8966-8969.2005
Alberdi A., Aizupura O., Bohmann K., Gopalakrishnan. S., Lynggard C., Nielsen M., et al. (2018). promises and pitfalls of using high-throughput sequencing for diet analysis. Mol. Ecol. Resour. 19, 327–348. doi: 10.1111/1755-0998.12960
Andersen K., Bird K. L., Rasmussen M., Haile J., Breuning-Madsen H., Kjær K. H., et al. (2012). Metabarcoding of ‘dirt’DNA from soil reflects vertebrate biodiversity. Mol. Ecol. 21 (8), 1966–1979. doi: 10.1111/j.1365-294X.2011.05261.x
Andres K. J., Sethi S. A., Lodge D. M., Andrés J. (2021). Nuclear eDNA estimates population allele frequencies and abundance in experimental mesocosms and field samples. Mol. Ecol. 30, 685–697. doi: 10.1111/mec.15765
Antich A., Palacin C., Wangensteen O. S., Turon X. (2021). To denoise or to cluster, that is not the question: optimizing pipelines for COI metabarcoding and metaphylogeography. BMC Bioinf. 22 (1), 1–24. doi: 10.1186/s12859-021-04115-6
Arreguín-Sánchez F. (1996). Catchability: a key parameter for fish stock assessment. Rev. Fish. Biol. Fish. 6, 221–242. doi: 10.1007/BF00182344
Atkinson S., Carlsson J. E. L., Ball B., Egan. D., Kelly-Quinn M., Whelan K., et al. (2018). . a quantitative PCR-based environmental DNA assay for detecting Atlantic salmon (Salmo salar L.). Aquatic. Conserv.: Mar. Freshw. Ecosyst. 28, 1238–1243. doi: 10.1002/aqc.2931
Bakker J., Wangensteen O. S., Chapman D. D., Boussarie G., Buddo D., Guttridge T.L., et al. (2017). Environmental DNA reveals tropical shark diversity in contrasting levels of anthropogenic impact. Sci. Rep. 7, 16886. doi: 10.1038/s41598-017-17150-2
Balasingham K. D., Walter R. P., Heath D. D. (2017). Residual eDNA detection sensitivity assessed by quantitative real-time PCR in a river ecosystem. Mol. Ecol. Resour. 17 (3), 523–532. doi: 10.1111/1755-0998.12598
Barnes M. A., Turner C. R. (2016). The ecology of environmental DNA and implications for conservation genetics. Conserv. Genet. 17 (1), 1–17. doi: 10.1007/s10592-015-0775-4
Barnes M. A., Turner C. R., Jerde C. L., Renshaw M. A., Chadderton W. L., Boussarie G., et al. (2014). . environmental conditions influence eDNA persistence in aquatic systems. Environ. Sci. Technol. 48, 1819–1827. doi: 10.1021/es404734p
Berry D., Mahfoudh B., Wagner M., Loy A. (2012). Barcode primers used in multiplex amplicon pyrosequencing bias amplification. Appl. Environ. Microbiol. 77, 7846–7849. doi: 10.1128/AEM.05220-11
Berry T. E., Saunders B. J., Coghlan M. L., Stat M., Jarman S., Richardson A. J., et al. (2019). Marine environmental DNA biomonitoring reveals seasonal patterns in biodiversity and identifies ecosystem responses to anomalous climatic events. PloS Genet. 15 (2), e1007943. doi: 10.1371/journal.pgen.1007943
Biggs J., Ewald N., Valentini A., Gaboriaud C., Dejean T., Griffiths R. A., et al. (2015). Using eDNA to develop a national citizen science-based monitoring program for the great crested newt (Triturus cristatus). Biol. Conserv. 183, 19–28. doi: 10.1016/j.biocon.2014.11.029
Binladen J., Gilbert M. T. P., Blolback J. P., Panitz F., Bendixen C., Nielsen R., et al. (2007). The use of code PCR primers enables high-throughput sequencing of multiple homolog amplification products by 454 parallel sequencing. PloS One 2, e197. doi: 10.1371/journal.pone.0000197
Bista I., Carvalho G., Walsh K., Seymour M., Hajibabaei M., Lallias D., et al. (2017). Annual time-series analysis of aqueous eDNA reveals ecologically relevant dynamics of lake ecosystem biodiversity. Nat. Commun. 8, 14087. doi: 10.1038/ncomms14087
Bloesch J. (1995). Mechanisms, measurement and importance of sediment resuspension in lakes. Mar. Freshw. Res. 46 (1), 295–304. doi: 10.1071/MF9950295
Boeuf G., Payan P. (2001). How should salinity influence fish growth? comp. biochem. Physiol. Part C. 130 (4), 411–423. doi: 10.1016/s1532-0456(01)00268-x
Bohmann K., Elbrecht V., Carøe C., Bista I., Leese F., Bunce M., et al. (2022). Strategies for sample labelling and library preparation in DNA metabarcoding studies. Mol. Ecol. Resour. 22 (4), 1231–1246. doi: 10.1111/1755-0998.13512
Bohmann K., Evans A., Gilbert M. T. P., Carvalho G. R., Creer S., Knapp M., et al. (2014). Environmental DNA for wildlife biology and biodiversity monitoring. Trends Ecol. Evol. 29 (6), 358–367. doi: 10.1016/j.tree.2014.04.003
Boonstra R. (2013). Reality as the leading cause of stress: rethinking the impact of chronic stress in nature. Funct. Ecol. 27 (1), 11–23. doi: 10.1111/1365-2435.12008
Boussarie G., Bakker J., Wangensteen O. S., Mariani S., Bonnin L., Juhel J. N., et al. (2018). Environmental DNA illuminates the dark diversity of sharks. Sci. Adv. 4 (5), eaap9661. doi: 10.1126/sciadv.aap9661
Boyer F., Mercier C., Bonin A., Le Bras Y., Taberlet P., Coissac E. (2016). obitools: a unix-inspired software package for DNA metabarcoding mol. Ecol. Resour. 16 (20), 176–182. doi: 10.1111/1755-0998.12428
Brown E. A., Chain F. J. J., Crease T. J., MacIsaac H. J., Cristescu M. E. (2015). Divergence thresholds and divergent biodiversity estimates: can metabarcoding reliably describe zooplankton communities? Ecol. Evol. 5 (11), 2234–2251. doi: 10.1002/ece3.1485
Bushnell P. G., Brill R. W. (1992). Oxygen transport and cardiovascular responses in skipjack tuna (Katsuwonus pelamis) and yellowfin tuna (Thunnus albacares) exposed to acute hypoxia. J. Comp. Physiol. B. 162 (2), 131–143. doi: 10.1007/BF00398338
Callahan B. J., McMurdie P. J., Rosen M. J., Han A. W., Johnson A. J., Holmes S. P. (2016). DADA2: High resolution sample inference from amplicon data. Nat. Methods 13, 581–583. doi: 10.1038/nmeth.3869
Castro J. A., Picornell A., Ramon M. (1998). Mitochondrial DNA: a tool for populational genetics studies. Int. Microbiol. 1 (4), 327–332.
Closek C. J., Santora J. A., Starks H. A., Schroeder I. D., Andruszkiewicz E. A., Sakuma K. M., et al. (2019). Marine vertebrate biodiversity and distribution within the central California current using environmental DNA (eDNA) metabarcoding and ecosystem surveys. Front. Mar. Sci. 6, 732. doi: 10.3389/fmars.2019.00732
Coble A. A., Flinders C. A., Homyack J. A., Penaluna B. E., Cronn R. C., Weitemier K. (2019). eDNA as a tool for identifying freshwater species in sustainable forestry: a critical review and potential future applications. Sci. Total Environ. 649, 1157–1170. doi: 10.1016/j.scitotenv.2018.08.370
Cochrane K. L. (2000). Reconciling sustainability, economic efficiency and equity in fisheries: the one that got away? Fish. Fish. 1, 3–21. doi: 10.1046/j.1467-2979.2000.00003.x
Cochrane K. L., Garcia S. M. (2009). “Introduction – fisheries management,” in A fishery manager's guidebook. Eds. Cochrane K. L., Garcia S. M. (West Sussex, United Kingdom: FAO). doi: 10.1002/9781444316315.ch1
Collins R. A., Wangensteen O. S., O’Gorman E. J., Mariani S., Sims D. W., Genner J., et al. (2018). Persistence of environmental DNA in marine systems. Commun. Biol. 1, 185. doi: 10.1038/s42003-018-0192-6
Coolen M. J., Overmann J. (2007). 217.000-year-old DNA sequences of green sulfur bacteria in Mediterranean sapropels and their implications for the reconstruction of the paleoenvironment. Environ. Microbiol. 9 (1), 238–249. doi: 10.1111/j.1462-2920.2006.01134.x
Corinaldesi C., Beolchini F., Dell’Anno A. (2008). Damage and degradation rates of extracellular DNA in marine sediments: implications for the preservation of gene sequences. Mol. Ecol. 17 (17), 3939–3951. doi: 10.1111/j.1365-294X.2008.03880.x
Cristescu M. E. (2014). From barcoding single individuals to metabarcoding biological communities: towards an integrative approach to the study of global biodiversity. Trends Ecol. Evol. 29, 556–571. doi: 10.1016/j.tree.2014.08.001
Curd E. E., Gold Z., Kandlikar G. S., Gomer J., Ogden M., O’Conell T., et al. (2019). Anacapa toolkit: An environmental DNA toolkit for processing multilocus metabarcode datasets. Methods Ecol. Evol. 10, 1469 –1475. doi: 10.1111/2041-210X.13214
Darling J. A. (2015). Genetic studies of aquatic biological invasions: closing the gap between research and management. Biol. Invasions 17, 951–971. doi: 10.1007/s10530-014-0726-x
Darling J. A., Mahon A. R. (2011). From molecules to management: adopting DNA-based methods for monitoring biological invasions in aquatic environments. Environ. Res. 111 (7), 978–988. doi: 10.1016/j.envres.2011.02.001
Deagle B. E., Chiaradia A., McInnes J., Jarman S. N. (2010). Pyrosequencing faecal DNA to determine diet of little penguins: is what goes in what comes out? Conserv. Genet. 11, 2039–2048. doi: 10.1007/s10592-010-0096-6
Debroas D., Domaizon I., Humbert J. F., Jardillier L., Lepère C., Oudart A., et al. (2017). Overview of freshwater microbial eukaryotes diversity: a first analysis of publicly available metabarcoding data. FEMS Microbiol. 93, (4). doi: 10.1093/femsec/fix023
Deere D., Porter J., Pickup R. W., Edwards C. (1996). Survival of cells and DNA of aeromonas salmonicida released into aquatic microcosms. J. Appl. Bacteriol. 81 (3), 309–318. doi: 10.1111/j.1365-2672.1996.tb04333.x
Deiner K., Bik H. M., Mächler E., Seymour M., Lacoursière-Roussel A., Altermatt F., et al. (2017). Environmental DNA metabarcoding: Transforming how we survey animal and plant communities. Mol. Ecol. 26 (21), 5872–5895. doi: 10.1111/mec.14350
Deiner K., Walser J. C., Mächler E., Altermatt F. (2015). Choice of capture and extraction methods affect detection of freshwater biodiversity from environmental DNA. Biol. Conserv. 183, 53–63. doi: 10.1016/j.biocon.2014.11.018
Dejean T., Valentini A., Duparc A., Pellier-Cuit S., Pompanon F., Taberlet P., et al. (2011). Persistence of environmental DNA in freshwater ecosystems. PloS One 6 (8), e23398. doi: 10.1371/journal.pone.0023398
Dejean T., Valentini A., Miquel C., Taberlet P., Bellemain E., Miaud C., et al. (2012). Improved detection of an alien invasive species through environmental DNA barcoding: the example of the American bullfrog Lithobates catesbeianus. J. Appl. Ecol. 49 (4), 953–959. doi: 10.1111/j.1365-2664.2012.02171.x
Dell'Anno A., Corinaldesi C. (2004). Degradation and turnover of extracellular DNA in marine sediments: ecological and methodological considerations. Appl. Environ. Microbiol. 70 (7), 4384–4386. doi: 10.1128/AEM.70.7.4384-4386.2004
Dennis D., Plagányi E., Van Putten I., Hutton T., Pascoe S. (2015). Cost benefit of fishery-independent surveys: Are they worth the money? Mar. Policy 58, 108–115. doi: 10.1016/j.marpol.2015.04.016
de Souza L. S., Godwin J. C., Renshaw M. A., Larson E. (2016). Environmental DNA (eDNA) detection probability is influenced by seasonal activity of organisms. PloS One 11, e0165273. doi: 10.1371/journal.pone.0165273
Díaz-Ferguson E. E., Moyer G. R. (2014). History, applications, methodological issues and perspectives for the use of environmental DNA (eDNA) in marine and freshwater environments. Rev. Biol. Trop. 62 (4), 1273–1284. doi: 10.15517/rbt.v62i4.13231
Djurhuus A., Closek C. J., Kelly R. P., Pitz K. J., Michisaki R. P., Starks H. A., et al. (2020). Environmental DNA reveals seasonal shifts and potential interactions in a marine community. Nat. Commun. 11, 254. doi: 10.1038/s41467-019-14105-1
Doi H., Inui R., Akamatsu Y., Kanno K., Yamanaka H., Takahara T., et al. (2017). Environmental DNA analysis for estimating the abundance and biomass of stream fish. Freshw. Biol. 62, 30–39. doi: 10.1111/fwb.12846
Douville M., Gagné F., Blaise C., André C. (2007). Occurrence and persistence of bacillus thuringiensis (Bt) and transgenic bt corn cry1Ab gene from an aquatic environment. ecotoxicol. Environ. Saf. 66, 195–203. doi: 10.1016/j.ecoenv.2006.01.002
Drummond A. J., Newcomb R. D., Buckley T. R., Xie D., Dopheide A., Potter B. C. M., et al. (2015). Evaluating a multigene environmental DNA approach for biodiversity assessment. GigaScience 4, 46. doi: 10.1186/s13742-015-0086-1
Dufresne Y., Lejzerowicz F., Perret-Gentil L. A., Pawlowski J., Cordier T. (2019). SLIM: a flexible web application for the reproducible processing of environmental DNA metabarcoding data. BMC Bioinf. 20, 88. doi: 10.1186/s12859-019-2663-2
Eichmiller J. J., Miller L. M., Sorensen P. W. (2016). Optimizing techniques to capture and extract environmental DNA for detection and quantification of fish. Mol. Ecol. Resour. 16 (1), 56–68. doi: 10.1111/1755-0998.12421
Engelsma M. Y., Hougee S., Nap D., Hofenk M., Rombout J. H., van Muiswinkel W. B., et al. (2003). Multiple acute temperature stress affects leucocyte populations and antibody responses in common carp, Cyprinus carpio L. Fish. Shellfish Immun. 15 (5), 397–410. doi: 10.1016/S1050-4648(03)00006-8
Epp L. S., Boessenkool S., Bellemain E. P., Haile J., Esposito A., Riaz T., et al. (2012). New environmental metabarcodes for analysing soil DNA: potential for studying past and present ecosystems. Mol. Ecol. 21, 1821–1833. doi: 10.1111/j.1365-294X.2012.05537.x
Eren A. M., Maignien L., Sul W. J., Murphy L. G., Grim S. L., Morrison H. G., et al. (2013). Oligotyping: differentiating between closely related microbial taxa using 16S rRNA gene data. Methods Ecol. Evol. 4, 1111–1119. doi: 10.1111/2041-210X.12114
Erickson R. A., Rees C. B., Coulter A. A., Merkes C. M., McCalla S. G., Touzinsky K. F., et al. (2016). Detecting the movement and spawning activity of bigheaded carps with environmental DNA. Mol. Ecol. Resour. 16 (4), 957–965. doi: 10.1111/1755-0998.12533
Evans N. T., Lamberti G. A. (2018). Freshwater fisheries assessment using environmental DNA: A primer on the method, its potential, and shortcomings as a conservation tool. Fish. Res. 197, 60–66. doi: 10.1016/j.fishres.2017.09.013
Evans N. T., Olds B. P., Renshaw M. A., Turner C. R., Li Y., Jerde C. L., et al. (2016). Quantification of mesocosm fish and amphibian species diversity via environmental DNA metabarcoding. Mol. Ecol. Resour. 16 (1), 29–41. doi: 10.1111/1755-0998.12433
Ficetola G. F., Miaud C., Pompanon F and Taberlet P. (2008). Species detection using environmental DNA from water samples. Biol. Lett. 4 (4), 423–425. doi: 10.1098/rsbl.2008.0118
Ficetola G. F., Pansu J., Bonin A., Coissac E., Giguet-Covex C., De Barba M., et al. (2015). Replication levels, false presences and the estimation of the presence/absence from eDNA metabarcoding data. Mol. Ecol. Resour. 15, 543–556. doi: 10.1111/1755-0998.12338
Ficetola G. F., Taberlet P., Coissac E. (2016). How to limit false positives in environmental DNA and metabarcoding? Mol. Ecol. Resour. 16 (3), 604–607. doi: 10.1111/1755-0998.12508
Foote A. D., Thomsen P. F., Sveegaard S., Wahlberg M., Kielgast J. (2012). Investigating the potential use ofof Environmental DNA (eDNA) for genetic monitoring of marine mammals. PloS One 7 (8), e41781. doi: 10.1371/journal.pone.0041781
Formel N., Enochs I. C., Sinigalliano C., Anderson S. R., Thompson L. R. (2021). Subsurface automated samplers for eDNA (SASe) for biological monitoring and research. HardwareX 10, e00239. doi: 10.1016/j.ohx.2021.e00239
Fraser H. M., Greenstreet S. P. R., Piet G. J. (2007). Taking account of catchability in groundfish survey trawls: implications for estimating demersal fish biomass. J. Mar. Sci. 64 (9), 1800–1819. doi: 10.1093/icesjms/fsm145
Furlan E. M., Gleeson D., Hardy C. M., Duncan R. P. (2016). A framework for estimating the sensitivity of eDNA surveys. Mol. Ecol. Resour. 16, 641–654. doi: 10.1111/1755-0998.12483
Gale N. K., Heath G., Cameron E., Rashid S., Redwood S. (2013). Using the framework method for the analysis of qualitative data in multi-disciplinary health research. BMC Med. Res. Methodol. 13 (1), 117. doi: 10.1186/1471-2288-13-117
Giovannoni S. J., Britschgi T. B., Moyer C. L., Field K. G. (1990). Genetic diversity in Sargasso Sea bacterioplankton. Nature 345(6270), 60–63. doi: 10.1038/345060a0
Goldberg C. S., Pilliod D. S., Arkle R. S., Waits L. P. (2011). Molecular detection of vertebrates in stream water: a demonstration using rocky mountain tailed frogs and Idaho giant salamanders. PloS One 6 (7), e22746. doi: 10.1371/journal.pone.0022746
Goldberg C. S., Strickler K. M., Pilliod D. S. (2015). Moving environmental DNA methods from concept to practice for monitoring aquatic macroorganisms. Biol. Conserv. 183, 1–3. doi: 10.1016/j.biocon.2014.11.040
Goldberg C. S., Turner C. R., Deiner K., Klymus K. E., Thomsen P. F., Murphy M. A., et al. (2016). Critical considerations for the application of environmental DNA methods to detect 0aquatic species. Methods Ecol. Evol. 7 (11), 1299–1307. doi: 10.1111/2041-210X.12595
Gold Z., Sprague J., Kushner D. J., Zerecero M. E., Barber P. H. (2021). eDNA metabarcoding as a biomonitoring tool for marine protected areas. PloS One 16 (2), e0238557. doi: 10.1371/journal.pone.0238557
Grassle J. F., Maciolek N. J. (1992). Deep-sea species richness: regional and local diversity estimates from quantitative bottom samples. Am. Nat. 139 (2), 313–341. Available at: https://www.jstor.org/stable/2462414
Guardiola M., Uriz M. J., Taberlet P., Coissac E., Wangensteen O. S., Turon X. (2015). Deep-Sea, deep-sequencing: Metabarcoding extracellular DNA from sediments of marine canyons. PloS One 10 (10), e0139633. doi: 10.1371/journal.pone.0139633
Günther T., Malmström H., Svensson E. M., Omrak A., Sánchez-Quinto F., Kılınç G. M., et al. (2018). Population genomics of Mesolithic Scandinavia: Investigating early postglacial migration routes and high-latitude adaptation. PloS Biol. 16 (1), e2003703. doi: 10.1371/journal.pbio.2003703
Handelsman J., Rondon M. R., Brady S. F., Clardy J., Goodman R. M. (1998). Molecular biological access to the chemistry of unknown soil microbes: a new frontier for natural products. Chem. Biol. 5 (10), R245–R249. doi: 10.1016/s1074-5521(98)90108-9
Hansen B. K., Bekkevold D., Clausen L. W., Nielsen E. E. (2018). The sceptical optimist: challenges and perspectives for the application of environmental DNA in marine fisheries. Fish. Fish. 19 (5), 751–768. doi: 10.1111/faf.12286
Hansen A. J., Mitchell D. L., Wiuf C., Paniker L., Brand T. B. (2006). Crosslinks rather than strand breaks determine access to ancient DNA sequences from frozen sediments. Genetics 173 (2), 1175–1179. doi: 10.1534/genetics.106.057349
Harris J. D. (2003). Can you bank on GenBank? Trends Ecol. Evol. 18, 317–319. doi: 10.1016/S0169-5347(03)00150-2
Hebsgaard M. B., Phillips M. J., Willerslev E. (2005). Geologically ancient DNA: fact or artefact? Trends Microbiol. 13 (5), 212–220. doi: 10.1016/j.tim.2005.03.010
Herder J., Valentini A., Bellemain E., Dejean T., Van Delft J. J. C. W., Thomsen P. F., et al. (2014). Environmental DNA. a review of the possible applications for the detection of (invasive) species (Stichting RAVON, Nijmegen: Netherlands Food and Consumer Product Safety Authority). Report 2013-104. doi: 10.13140/RG.2.1.4002.1208
Hillis D. M., Dixon M. T. (1991). Ribosomal DNA: molecular evolution and phylogenetic inference. Q Rev. Biol. 66, 411–453. doi: 10.1086/417338
Hinlo R., Gleeson D., Lintermans M., Furlan E. (2017). Methods to maximise recovery of environmental DNA from water samples. PloS One 12 (6), e0179251. doi: 10.1371/journal.pone.0179251
Hofreiter M., Serre D., Poinar H. N., Kuch M., Pääbo S. (2001). Ancient DNA. Nat. Rev. Genet. 2 (5), 353. doi: 10.1038/35072071
Holman L. E., Bruyn M., Creer S., Carvalho G., Robidart J., Rius M. (2019). Detection of introduced and resident marine species using environmental DNA metabarcoding of sediment and water. Sci. Rep. 9, 911559. doi: 10.1038/s41598-019-47899-7
Jarman S. N., Berry O., Bunce M. (2018). The value of environmental DNA biobanking for long-term biomonitoring. Nat. Ecol. Evol. 2, 1192–1193. doi: 10.1038/s41559-018-0614-3
Jerde C. L., Mahon A. R. (2015). Improving confidence in environmental DNA species detection. Mol. Ecol. Resour. 15, 461–463. doi: 10.1111/1755-0998.12377
Jerde C. L., Mahon A. R., Chadderton W. L., Lodge D. M. (2011). “Sight-unseen” detection of rare aquatic species using environmental DNA. Conserv. Lett. 4 (2), 150–157. doi: 10.1111/j.1755-263X.2010.00158.x
Jo T., Sakata M. K., Murakami H., Masuda R., Minamoto T. (2021). Universal performance of benzalkonium chloride for the preservation of environmental DNA in seawater samples. Limnol. Oceanogr-Methods 19, 758–768. doi: 10.1002/lom3.10459
Jo T., Takao K., Minamoto T. (2022). Linking the state of environmental DNA to its application for biomonitoring and stock assessment: Targeting mitochondrial/nuclear genes, and different DNA fragment lengths and particle sizes. Environ. DNA 4, 271–283. doi: 10.1002/edn3.253
Keck F., Vasselon V., Tapolczai K., Rimet F., Bouchez A. (2017). Freshwater biomonitoring in the information age. Front. Ecol. Environ. 15 (5), 266–274. doi: 10.1002/fee.1490
Kelly R. P., Port J. A., Yamahara K. M., Crowder L. B. (2014). Using environmental DNA to census marine fishes in a large mesocosm. PloS One 9 (1), e86175. doi: 10.1371/journal.pone.0086175
Kim A. R., Yoon T. H., Lee C. I., Kang C. K., Kim H. W. (2021). Metabarcoding analysis of ichthyoplankton in the East/Japan Sea using the novel fish-specific universal primer set. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.614394
Klymus K. E., Richter C. A., Chapman D. C., Paukert C. (2015). Quantification of eDNA shedding rates from invasive bighead carp Hypophthalmichthys nobilis and silver carp Hypophthalmichthys molitrix. Biol. Conserv. 183, 77–84. doi: 10.1016/j.biocon.2014.11.020
Knudsen S. W., Ebert R. B., Hesselsøe M., Kuntke F., Hassingboe J., Bondgaard P., et al. (2019). Species-specific detection and quantification of environmental DNA from marine fishes in the Baltic sea. J. Exp. Mar. Biol. Ecol. 510, 31–45. doi: 10.1016/j.jembe.2018.09.004
Knudsen E. E., McDonald D. (2019). Sustainable fisheries management (Boca Ratón, United States: CRC Press), ISBN: 9781566704809.
Krishnan A., Sweeney M., Vasic J., Galbraith D., Vasic B. (2011). Barcodes for DNA sequencing with guaranteed error-correction capability. Electron. Lett. 47 (4), 236–237. doi: 10.1049/el.2010.3546
Kwong S., Srivathsan A., Meier R. (2012). An update on DNA barcoding: low species coverage and numerous unidentified sequences. Cladistics 28, 639–644. doi: 10.1111/j.1096-0031.2012.00408.x
Lacoursière-Roussel A., Côté G., Leclerc V., Bernatchez L. (2016a). Quantifying relative fish abundance with eDNA: A promising tool for fisheries management. J. Appl. Ecol. 53, 1148–1157. doi: 10.1111/1365-2664.12598
Lacoursière-Roussel A., Rosabal M., Bernatchez L. (2016b). Estimating fish abundance and biomass from eDNA concentrations: Variability among capture methods and environmental conditions. Mol. Ecol. Resour. 16, 1401–1414. doi: 10.1111/1755-0998.12522
Laramie M. B., Pilliodm D. S., Goldberg C. S. (2015). Characterizing the distribution of an endangered salmonid using environmental DNA analysis. Biol. Conserv. 183, 29–37. doi: 10.1016/j.biocon.2014.11.025
Lee J. H., La H. S., Kim J. H., Son W., Park H., Kim Y. M., et al. (2022). Application of dual metabarcoding platforms for the meso- and macrozooplankton taxa in the Ross sea. Genes 13, 922. doi: 10.3390/genes13050922
Levi T., Allen J. M., Bell D., Joyce J., Russuell J. R., Tallmon D. A., et al. (2019). Environmental DNA for the enumeration and management of pacific salmon. Mol. Ecol. Resour. 19, 597–608. doi: 10.1111/1755-0998.12987
Levy-Booth D. J., Campbell R. G., Gulden R. H., Hart M. M., Powell, Klironomos J. N., et al. (2007). Cycling of extracellular DNA in the soil environment. Soil Biol. Biochem. 39 (12), 2977–2991. doi: 10.1016/j.soilbio.2007.06.020
Lodge D. M., Turner C. R., Jerde C. L., Barnes M. A., Chadderton L., Egan S. P., et al. (2012). Conservation in a cup of water: estimating biodiversity and population abundance from environmental DNA. Mol. Ecol. 21 (11), 2555–2558. doi: 10.1111/j.1365-294X.2012.05600.x
Maruyama A., Nakamura K., Yamanaka H., Kondoh M., Minamoto T. (2014). The release rate of environmental DNA from juvenile and adult fish. PloS One 9 (12), e114639. doi: 10.1371/journal.pone.0114639
Matheson C. D., Gurney C., Esau N., Lehto R. (2010). Assessing PCR inhibition from humic substances. Open Enzym. Inhid. J. 3 (1), 38–45. doi: 10.2174/1874940201003010038
Mathon L., Guérin P. E., Normandeau E., Valentini A., Noel C., Lionnet C., et al. (2021). Benchmarking bioinformatic tools for fast and accurate eDNA metabarcoding species identification. Mol. Ecol. Resour. 21 (7), 2565–2579. doi: 10.1111/1755-0998.13430
Matsui K., Honjo M., Kawabata Z. (2001). Estimation of the fate of dissolved DNA in thermally stratified lake water from the stability of exogenous plasmid DNA. Aquat. Microb. Ecol. 26 (1), 95–102. doi: 10.3354/ame026095
McCormick S. T., Pinedo M. L., Shenker S., Wolf B. (1989). Sequencing in an assembly line with blocking to minimize cycle time. Oper. Res. 37 (6), 925–935. doi: 10.1287/opre.37.6.925
McKee A. M., Spear S. F., Pierson T. W. (2015). The effect of dilution and the use of a post-extraction nucleic acid purification column on the accuracy, precision, and inhibition of environmental DNA samples. Biol. Conserv. 183, 70–76. doi: 10.1016/j.biocon.2014.11.031
McKenzie D. J., Höglund E., Dupont-Prinet A., Larsen B. K., Skov P. V., Pedersen P. B., et al. (2012). Effects of stocking density and sustained aerobic exercise on growth, energetics and welfare of rainbow trout. Aquaculture 338, 216–222. doi: 10.1016/j.aquaculture.2012.01.020
Milidine K., Armstrong J., Metcalfe N. (1995). Presence of shelter reduces maintenance metabolism of juvenile salmon. Funct. Ecol. 20 (5), 839–845. doi: 10.1111/j.1365-2435.2006.01166.x
Noll D. M., Mason T. M., Miller P. S. (2006). Formation and repair of interstrand cross-links in DNA. Chem. Rev. 106 (2), 277–301. doi: 10.1021/cr040478b
Ogram A., Sayler G. S., Barkay T. (1987). The extraction and purification of microbial DNA from sediments. J. Microbiol. Methods 7 (2-3): 57-66. doi: 10.1016/0167-7012(87)90025-X
Olson Z. H., Briggler J. T., Williams R. N. (2012). An eDNA approach to detect eastern hellbenders (Cryptobranchus a. alleganiensis) using samples of water. Wildl. Res. 39 (7), 629–636. doi: 10.1071/WR12114
Opel K. L., Chung D., McCord B. R. (2010). A study of PCR inhibition mechanisms using real time PCR. J. Forensic Sci. 55 (1), 25–33. doi: 10.1111/j.1556-4029.2009.01245.x
Ovenden J. R., Berry O., Welch D. J., Buckworth R., Dichmont M. (2015). Ocean’s eleven: a critical evaluation of the role of population, evolutionary and molecular genetics in the management of wild fisheries. Fish. Fish. 16, 125–159. doi: 10.1111/faf.12052
Palsbøll P., Berube M., Allendorf F. (2007). . identification of management units using population genetic data. Trends Ecol. Evol. 22, 11–16. doi: 10.1016/j.tree.2006.09.003
Person-Le Ruyet J., Mahé K., Le Bayon N., Le, Delliou H. (2004). Effects of temperature on growth and metabolism in a Mediterranean population of European sea bass, Dicentrarchus labrax. Aquaculture 237 (1-4), 269–280. doi: 10.1016/j.aquaculture.2004.04.021
Petty J. T., Hansbarger J. L., Huntsman B. M., Mazik P. M. (2012). Brook trout movement in response to temperature, flow, and thermal refugia within a complex Appalachian riverscape. Trans. Am. Fish. Soc 141 (4), 1060–1073. doi: 10.1080/00028487.2012.681102
Piaggio A. J., Engeman R. M., Hopken M. W., Humphrey J. S., Keacher K. L., Bruce W. E., et al. (2014). Detecting an elusive invasive species: a diagnostic PCR to detect Burmese python in Florida waters and an assessment of persistence of environmental DNA. Mol. Ecol. Resour. 14 (2), 374–380. doi: 10.1111/1755-0998.12180
Piggott M. P. (2016). Evaluating the effects of laboratory protocols on eDNA detection probability for an endangered freshwater fish. Ecol. Evol. 6 (9), 2739–2750. doi: 10.1002/ece3.2083
Pikitch E. K., Santora C., Babcock E. A., Bakun A., Bonfil R., Dayton P., et al. (2004). Ecology. ecosystem-based fishery management. Science 305 (5682), 346–347. doi: 10.1126/science.1098222
Pilliod D. S., Goldberg C. S., Arkle R. S., Waits L. P. (2014). Factors influencing detection of eDNA from a stream-dwelling amphibian. Mol. Ecol. Resour. 14 (1), 109–116. doi: 10.1111/1755-0998.12159
Piper A. M., Batovska J., Cogan N. O. I., Weiss J., Cunningham J. P., Rodoni B. C., et al. (2019). Prospects and challenges of implementing DNA metabarcoding for high-throughput insect surveillance. GigaScience 8 (8), giz092. doi: 10.1093/gigascience/giz092
Ratcliffe F. C., Uren Webster T. M., Garcia de Leaniz C., Consuegra S. (2020). A drop in the ocean: Monitoring fish communities in spawning areas using environmental DNA. Environ. DNA 3, 43–54. doi: 10.1002/edn3.87
Rees H. C., Maddison B. C., Middleditch D. J., Patmore J. R., Gough K. C. (2014). The detection of aquatic animal species using environmental DNA–a review of eDNA as a survey tool in ecology. J. Appl. Ecol. 51 (5), 1450–1459. doi: 10.1111/1365-2664.12306
Renshaw M. A., Olds B. P., Jerde C. L., McVeigh M. M., Lodge. D. M. (2015). The room temperature preservation of filtered environmental DNA samples and assimilation into a phenol–chloroform–isoamyl alcohol DNA extraction. Mol. Ecol. Resour. 15 (1), 168–176. doi: 10.1111/1755-0998.12281
Robson H. L., Noble T. H., Saunders R. J., Robson S. K., Burrows D. W., Jerry D. R., et al. (2016). Fine-tuning for the tropics: application of eDNA technology for invasive fish detection in tropical freshwater ecosystems. Mol. Ecol. Resour. 16 (4), 922–932. doi: 10.1111/1755-0998.12505
Rourke M. L., Fowler A. M., Hughes J. M., Broadhurst M., DiBattista J., Fielder S., et al. (2022). Environmental DNA (eDNA) as a tool for assessing fish biomass: A review of approaches and future considerations for resource surveys. Environ. DNA. 4, 9–33. doi: 10.1002/edn3.185
Ruppert K. M., Kline R. J., Rahman M. S. (2019). . past, present, and future perspectives of environmental DNA (eDNA) metabarcoding: A systematic review in methods, monitoring, and applications of global eDNA. Glob. Ecol. Conserv. 17, e00547. doi: 10.1016/j.gecco.2019.e00547
Russo T., Maiello G., Talarico L., Baillie C., Colosimo G., D’Andrea L., et al. (2021). All is fish that comes to the net: metabarcoding for rapid fisheries catch assessment. Ecol. Appl. 31 (2), e02273. doi: 10.1002/eap.2273
Salter I., Joensen M., Kristiansen R., Steingrund P., Vestergaard P. (2019). Environmental DNA concentrations are correlated with regional biomass of Atlantic cod in oceanic waters. Commun. Biol. 2, 461. doi: 10.1038/s42003-019-0696-8
Santos A. M., Branco M. (2012). The quality of name-based species records in databases. Trends Ecol. Evol. 27, 6–7. doi: 10.1016/j.tree.2011.10.004
Sassoubre L. M., Yamahara K. M., Gardner L. D., Block B. A., Boehm A. B. (2016). Quantification of environmental DNA (eDNA) shedding and decay rates for three marine fish. Environ. Sci. Technol. 50 (19), 10456–10464. doi: 10.1021/acs.est.6b03114
Sato Y., Miya M., Fukunaga T., Sado T., Iwasaki W. (2018). MitoFish and MiFish pipeline: A mitochondrial genome database of fish with an analysis pipeline for environmental DNA metabarcoding. Mol. Biol. Evol. 35, 1553–1555. doi: 10.1093/molbev/msy074
Selong J. H., McMahon T. E., Zale A. V., Barrows F. T. (2001). Effect of temperature on growth and survival of bull trout, with application of an improved method for determining thermal tolerance in fishes. Trans. Am. Fish. Soc 130 (6), 1026–1037. doi: 10.1577/1548-8659(2001)130<1026:EOTOGA>2.0.CO;2
Senapati D., Bhattacharya M., Kar A., Chini D. S., Das B. K., Patra B. C. (2019). Environmental DNA (eDNA): A promising biological survey tool for aquatic species detection. Proc. Zool. Soc 72 (3), 211–228. doi: 10.1007/s12595-018-0268-9
Sepulveda A. J., Hutchins P. R., Forstchen M., Mckeefry M. N., Swigris A. M. (2020). The elephant in the Lab (and field): Contamination in aquatic environmental DNA studies. Front. Ecol. Evol. 8. doi: 10.3389/fevo.2020.609973
Sigsgaard E. E., Jensen M. R., Winkelmann I. E., Møller P. R., Hansen M. M., Thomsen P. F. (2020). Population-level inferences from environmental DNA–current status and future perspectives. Evol. Appl. 13, 245 –262. doi: 10.1111/eva.12882
Sigsgaard E. E., Carl H., Møller P. R., Thomsen P. F. (2015). Monitoring the near-extinct European weather loach in Denmark based on environmental DNA from water samples. Biol. Conserv. 183, 46–52. doi: 10.1016/j.biocon.2014.11.023
Sigsgaard E. E., Nielsen I. B., Bach S. S., Lorenzen E. D., Robinson D. P., Knudsen S. W., et al. (2016). Population characteristics of a large whale shark aggregation inferred from seawater environmental DNA. Nat. Ecol. Evol. 1 (1), 4. doi: 10.1038/s41559-016-0004
Sloman K. A., Motherwell G., O'connor K., Taylor A. C. (2000). The effect of social stress on the standard metabolic rate (SMR) of brown trout, salmo trutta. fish physiol. Biochem 23 (1), 49–53. doi: 10.1023/A:1007855100185
Snelgrove P. V. (1999). Getting to the bottom of marine biodiversity: sedimentary habitats: ocean bottoms are the most widespread habitat on earth and support high biodiversity and key ecosystem services. BioScience 49 (2), 129–138. doi: 10.2307/1313538
Spear S. F., Groves J. D., Williams L. A., Waits L. P. (2015). Using environmental DNA methods to improve detectability in a hellbender (Cryptobranchus alleganiensis) monitoring program. Biol. Conserv. 183, 38–45. doi: 10.1016/j.biocon.2014.11.016
Stat M., Huggett M. J., Bernasconi R., DiBattista J. D., Berry T. E., Newman S. J., et al. (2017). Ecosystem biomonitoring with eDNA: Metabarcoding across the tree of life in a tropical marine environment. Sci. Rep. 7, 12240. doi: 10.1038/s41598-017-12501-5
Stewart K. A. (2019). Understanding the effects of biotic and abiotic factors on sources of aquatic environmental DNA. Biodivers. Conserv. 28 (5), 983–1001. doi: 10.1007/s10531-019-01709-8
Stoeckle M. Y., Adolf J., Charlop-Powers Z., Dunton K. J., Hinks G., VanMorter S. M., et al. (2021). Trawl and eDNA assessment of marine fish diversity, seasonality, and relative abundance in coastal new Jersey, USA. ICES. J. Mar. Sci. 78, (1) 293–304. doi: 10.1093/icesjms/fsaa225
Stoeckle M. Y., Das M. M., Charlop-Powers Z. (2020). Improved environmental DNA reference library detects overlooked marine fishes in new Jersey, united states. Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.00226
Stoeckle M. Y., Soboleva L., Charlop-Powers Z. (2017). Aquatic environmental DNA detects seasonal fish abundance and habitat preference in an urban estuary. PloS One 12 (4), e0175186. doi: 10.1371/journal.pone.0175186
Taberlet P., Bonin A., Zinger L., Coissac E. (2018). Environmental DNA (UK: Oxford University Press), ISBN: 9780198767282. doi: 10.1093/oso/9780198767220.001.0001
Taberlet P., Coissac E., Hajibabaei M., Rieseberg L. H. (2012a). Environmental DNA. Mol. Ecol. 21 (8), 1789–1793. doi: 10.1111/j.1365-294X.2012.05542.x
Taberlet P., Prud’Homme S. M., Campione E., Roy J., Miquel C., Shehzad W., et al. (2012b). Soil sampling and isolation of extracellular DNA from large amount of starting material suitable for metabarcoding studies. Mol. Ecol. 21 (8), 1816–1820. doi: 10.1111/j.1365-294X.2011.05317.x
Taberlet P., Waits L. P., Luikart G. (1999). Noninvasive genetic sampling: look before you leap. Trends Ecol. Evol. 14, 323–327. doi: 10.1016/s0169-5347(99)01637-7
Takahara T., Minamoto T., Doi H. (2013). Using environmental DNA to estimate the distribution of an invasive fish species in ponds. PloS One 8 (2), e56584. doi: 10.1371/journal.pone.0056584
Takahara T., Minamoto T., Yamanaka H., Doi H., Kawabata Z. I. (2012). Estimation of fish biomass using environmental DNA. PloS One 7 (4), e35868. doi: 10.1371/journal.pone.0035868
Takahara T., Yamanaka H., Suzuki A. A., Honjo M. N., Minamoto T., Yonekura R., et al. (2011). Stress response to daily temperature fluctuations in common carp, Cyprinus carpio l. Hydrobiologia 675 (1), 65. doi: 10.1007/s10750-011-0796-z
Tapolczai K., Vasselon V., Bouchez A., Stenger-Kovács C., Padisák J., Rimet F. (2019). The impact of OTU sequence similarity threshold on diatom-based bioassessment: a case study of the rivers of Mayotte (France, Indian ocean). Ecol. Evol. 9, 166–179. doi: 10.1002/ece3.4701
Thomsen P. F., Kielgast J., Iversen L. L., Møller P. R., Rasmussen M., Willerslev E. (2012a). Detection of a diverse marine fish fauna using environmental DNA from seawater samples. PloS One 7 (8), e41732. doi: 10.1371/journal.pone.0041732
Thomsen P. F., Kielgast J. O. S., Iversen L. L., Wiuf C., Rasmussen M., Gilbert M. T. P., et al. (2012b). Monitoring endangered freshwater biodiversity using environmental DNA. Mol. Ecol. 21 (11), 2565–2573. doi: 10.1111/j.1365-294X.2011.05418.x
Thomsen P. F., Møller P. R., Sigsgaard E. E., Knudsen S. W., Jørgensen O. A., Willerslev E. (2016). Environmental DNA from seawater samples correlate with trawl catches of subarctic, deepwater fishes. PloS One 11 (11), e0165252. doi: 10.1371/journal.pone.0165252
Thomsen P. F., Willerslev E. (2015). Environmental DNA–an emerging tool in conservation for monitoring past and present biodiversity. Biol. Conserv. 183, 4–18. doi: 10.1016/j.biocon.2014.11.019
Tréguier A., Paillisson J. M., Dejean T., Valentini A., Schlaepfer M. A., Roussel J. M. (2014). Environmental DNA surveillance for invertebrate species: advantages and technical limitations to detect invasive crayfish Procambarus clarkii in freshwater ponds. J. Appl. Ecol. 51 (4), 871–879. doi: 10.1111/1365-2664.12262
Truelove N. K., Andruszkiewicz E. A., Block B. A. (2019). A rapid environmental DNA method for detecting white sharks in the open ocean. Methods Ecol. Evol. 10, 1128–1135. doi: 10.1111/2041-210X.13201
Tsai Y. L., Olson B. H. (1992). Rapid method for separation of bacterial DNA from humic substances in sediments for polymerase chain reaction. Appl. Environ. Microbiol. 58 (7), 2292–2295. doi: 10.1128/aem.58.7.2292-2295.1992
Turner C. R., Uy K. L., Everhart R. C. (2015). Fish environmental DNA is more concentrated in aquatic sediments than surface water. Biol. Conserv. 183, 93–102. doi: 10.1016/j.biocon.2014.11.017
Ushio M., Murakami H., Masuda R., Sado T., Miya M., Sakurai S., et al. (2018). Quantitative monitoring of multispecies fish environmental DNA using high-throughput sequencing. Metabarcoding Metagenom. 2, e23297. doi: 10.3897/mbmg.2.23297
Valsecchi E., Arcangeli A., Lombardi R., Boyse E., Carr I. M., Galli P., et al. (2021). Ferries and environmental DNA: underway sampling from commercial vessels provides new opportunities for systematic genetic surveys of marine biodiversity. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.704786
Van Dievel M., Janssens L., Stoks R. (2016). Short-and long-term behavioural, physiological and stoichiometric responses to predation risk indicate chronic stress and compensatory mechanisms. Oecologia 181 (2), 347–357. doi: 10.1007/s00442-015-3440-1
Wang Y., Chen K., Gao J., Wang M., Dong J., Xie Y., et al. (2021). Environmental DNA of preservative ethanol performed better than water samples in detecting macroinvertebrate diversity using metabarcoding. Divers. Distrib. 27, 1989–2002. doi: 10.1111/ddi.13284
Willerslev E., Cooper A. (2005). Ancient DNA. Proc. R. Soc B. Sci. 272 (1558), 3–16. doi: 10.1098/rspb.2004.2813
Willerslev E., Hansen A. J., Binladen J., Brand T. B., Gilbert M. T. P., Shapiro B., et al. (2003). Diverse plant and animal genetic records from Holocene and pleistocene sediments. Science 300 (5620), 791–795. doi: 10.1126/science.1084114
Yamamoto S., Masuda R., Sato Y., Sado T., Araki H., Kondoh M., et al. (2017). Environmental DNA metabarcoding reveals local fish communities in a species-rich coastal sea. Sci. Rep. 7, 40368. doi: 10.1038/srep40368
Yoccoz N. G. (2012). The future of environmental DNA in ecology. Mol. Ecol. 21 (8), 2031–2038. doi: 10.1111/j.1365-294X.2012.05505.x
Keywords: eDNA, fisheries, experimental design, marine environment, metabarcoding, monitoring, standardization, biodiversity
Citation: Ramírez-Amaro S, Bassitta M, Picornell A, Ramon C and Terrasa B (2022) Environmental DNA: State-of-the-art of its application for fisheries assessment in marine environments. Front. Mar. Sci. 9:1004674. doi: 10.3389/fmars.2022.1004674
Received: 27 July 2022; Accepted: 31 October 2022;
Published: 05 December 2022.
Edited by:
Fran Saborido-Rey, Spanish National Research Council (CSIC), SpainCopyright © 2022 Ramírez-Amaro, Bassitta, Picornell, Ramon and Terrasa. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sergio Ramírez-Amaro, c3JncmFtaXJlejA4QGdtYWlsLmNvbQ==