 Melita Peharda1*
Melita Peharda1* David P. Gillikin2
David P. Gillikin2 Bernd R. Schöne3Anouk Verheyden2
Bernd R. Schöne3Anouk Verheyden2 Hana Uvanović1
Hana Uvanović1 Krešimir Markulin4Tomislav Šarić5
Krešimir Markulin4Tomislav Šarić5 Ivica Janeković6Ivan Župan5
Ivica Janeković6Ivan Župan5
- 1Laboratory of Fisheries Science and Management, Institute of Oceanography and Fisheries, Split, Croatia
- 2Geosciences Department, Union College, Schenectady, NY, United States
- 3Institute of Geosciences, University of Mainz, Mainz, Germany
- 4Laboratory of Chemical Oceanography and Sedimentology of the Sea, Institute of Oceanography and Fisheries, Split, Croatia
- 5Department of Ecology, Agronomy and Aquaculture, University of Zadar, Zadar, Croatia
- 6Oceans Institute, The University of Western Australia, Perth, WA, Australia
Pinna nobilis is a large bivalve endemic to the Mediterranean Sea that lives in shallow coastal areas. Due to its size and relatively fast shell growth rates, it is an interesting taxon for high-resolution study of nitrogen isotopes of carbonate bound organic matter (δ15NCBOM). In this study we tested if P. nobilis shells can be used as an indicator of the nitrogen isotope baseline of the system, if it can provide high-resolution data on environmental δ15N variability, and if the chemical properties of the shell and biomineralization process change in response to mass mortality events spreading in the Mediterranean. Shells were opportunistically collected during 2019 and 2020 by skin diving, as a part of a project on mortality monitoring, from four shallow coastal localities in the eastern Adriatic. Shell powder for δ15NCBOM analysis was collected by milling sample swaths from the internal (low-resolution) and external (high resolution) shell surface. Significant differences in δ15NCBOM, obtained from the internal shell surface, were observed between sampling localities with different anthropogenic influences, with lowest values (∼3–4‰) recorded for shells obtained from Pag Bay, and highest (∼6–8‰) for shells sampled in Lim and Kaštela Bays. High-resolution samples from the external shell surface of Pinna nobilis showed spatial and temporal variations in δ15NCBOM values, with temporal resolution of 1–3 weeks. High-resolution δ15NCBOM data obtained from the shell Kas1 corresponded to a time interval from spring 2018 to summer 2019 and had a pronounced increase of δ15NCBOM values closest to the shell margin coupled with a decrease in δ13Cshell values, indicating that this animal was experiencing stressful conditions several months prior to its death. According to our findings, δ15NCBOM values from P. nobilis shells can serve as an indicator of the isotopic baseline of the ecosystem potentially as a powerful tool to study bivalve physiology.
Introduction
Pinna nobilis is a large bivalve endemic to the Mediterranean Sea. In early autumn of 2016, P. nobilis mass mortality event was detected in the Spanish part of the Mediterranean Sea (Vázquez-Luis et al., 2017), which soon spread to other parts of this enclosed sea (Cabanellas-Reboredo et al., 2019), including the Adriatic Sea (Čižmek et al., 2020; Šarić et al., 2020). By the end of 2020, all P. nobilis populations in the Croatian part of the Adriatic Sea have been affected by the mass mortality and very few individuals survived (Šarić, personal observation). At the first occurrence of a mass mortality event in Spain, the parasite Haplosporidium pinnae was considered. as the likely cause (Darriba, 2017; Catanese et al., 2018). However, further research has indicated that other pathogens, such as bacteria from the genus Mycobacterium and Vibrio, were also involved (Carella et al., 2019; Šarić et al., 2020; Scarpa et al., 2020; Lattos et al., 2021). Moreover, according to these reports, the mass mortality events were presumably caused by polymicrobial disease associated with various abiotic factors, and the pathogenesis has not yet been fully elucidated.
Conservation efforts require comprehensive knowledge on the biology and ecology of species as well as environmental variations, including those related to nitrogen in coastal marine habitats. Pinna nobilis was listed as an endangered and protected species under the European Council Directive 92/43/EEC (EEC, 1992), is recorded in the ANNEX II of the Barcelona Convention, and is under local law protection in all European Union Mediterranean countries. Concerns for the status of P. nobilis were raised even before 2016 and the start of mass mortality event, because it declined due to exposure to a number of cumulative stressors including habitat degradation, food web alteration and contaminant burden (Basso et al., 2015). Due to the devastating effects of mass mortality event on the population of P. nobilis, IUCN changed the status of this species to “critically endangered” (Kersting et al., 2019).
Over the past decade, methods for the analysis of nitrogen stable isotopes in carbonate-bound organic matter (δ15NCBOM) using direct combustion of carbonates have been developed and optimized enabling the investigation of δ15NCBOM in bivalve shells from freshwater, estuarine, and marine habitats (e.g., Versteegh et al., 2011; Darrow et al., 2017; Gillikin et al., 2017; Graniero et al., 2021; Kukolich and Dettman, 2021; Schöne and Huang, 2021). Nitrogen isotopes measured in carbonate-bound organic matter are a function of δ15N values of dissolved nitrogen in the ambient water as well as the physiology and diet of the bivalve (e.g., Gillikin et al., 2017; Whitney et al., 2019; Das et al., 2021; Graniero et al., 2021). Bivalves are considered as low-level consumers, and as such, they can provide information on the nitrogen isotope baseline of the environment (e.g., Jennings and Warr, 2003; Thibault et al., 2020). In many previous studies, δ15NCBOM was measured in marine bivalve species including Mercenaria sp. (O’Donnell et al., 2003), Mercenaria mercenaria (Carmichael et al., 2008; Oczkowski et al., 2016), Ruditapes philippinarum (Watanabe et al., 2009), Mytilus edulis (Versteegh et al., 2011), Crassostrea virginica (Kovacs et al., 2010; Oczkowski et al., 2016; Black et al., 2017; Darrow et al., 2017), Pecten maximus (Gillikin et al., 2017), Rangia cuneata (Graniero et al., 2021), Spisula solidissima (Das et al., 2021), and Arctica islandica (Whitney et al., 2019; Schöne and Huang, 2021). An increase in δ15NCBOM has been related to nutrient enrichment reflecting anthropogenic perturbation (e.g., Carmichael et al., 2008; Kovacs et al., 2010; Oczkowski et al., 2016). As yet, only a limited number of bivalves were sampled at high-resolution to generate δ15NCBOM time-series, e.g., Pecten maximus (Gillikin et al., 2017), Spisula solidissima (Das et al., 2021), and Arctica islandica (Schöne and Huang, 2021). Due to its size (>1 m, Zavodnik et al., 1991) and very fast shell growth rate (Richardson et al., 2004), Pinna nobilis presents an interesting taxon for high-resolution δ15NCBOM research. High-resolution δ15NCBOM can provide an important insight into temporal variation in bivalve diet, physiology (including ontogenetic changes), as well as background nitrogen isotopic signatures (Gillikin et al., 2017; Das et al., 2021; Schöne and Huang, 2021).
Previous studies on the chemical properties of P. nobilis shells include research on oxygen (δ18Oshell) and carbon (δ13Cshell) stable isotopes (Richardson et al., 1999, 2004; Kennedy et al., 2001a; Freitas et al., 2005; García-March et al., 2011), as well as element ratios including Mg/Ca and Sr/Ca (Richardson et al., 2004; Freitas et al., 2005). García-March et al. (2011) suggested that P. nobilis can be used to reconstruct environmental, ecological and climate changes in the Mediterranean Sea. Following previous analyses of δ18O and δ13C in P. nobilis shells, the present study focuses on δ15NCBOM values of this species. The main objectives were to test if P. nobilis shells (i) can be used as an indicator of the nitrogen isotope baseline of the system, and (ii) provides high-resolution data on environmental δ15N variability. Due to the multiple mass mortality events spreading throughout the Mediterranean, including the Adriatic Sea, we also tested if (iii) the chemical properties of P. nobilis and shell deposition process changed in response to the diseases.
Materials and Methods
A preliminary study was conducted in 2018 to test the feasibility to obtain δ15NCBOM data from shells of several bivalve species from the Adriatic Sea using the standard combustion technique on an elemental analyzer (e.g., Versteegh et al., 2011).
Sample Collection
For the purpose of this study, P. nobilis shells were opportunistically collected by skin diving from four shallow coastal localities in the eastern Adriatic (Figure 1), as a part of a project on mortality monitoring. More information on sample collection and epidemiological status of live-collected specimens is available in Šarić et al. (2020). Three specimens from Lim Bay (October 2019; 45°07′55′′N, 13°43′55′′E), Kaštela Bay (January 2020, 43°33′01′′N, 16°21′36′′E), and Mali Ston Bay (November 2019, 42°52′01′′N, 17°41′51′′E) were collected alive, while in Pag Bay (44°26′35′′N, 15°03′01′′E), shells of three recently dead specimens were collected in September 2020. In addition, one dead empty shell collected from Kaštela Bay in January 2020 was also analyzed. Lim Bay and Mali Ston Bay are important bivalve aquaculture areas. Lim Bay is located in the North of the eastern Adriatic and Mali Ston Bay in the South. Kaštela Bay is semi-enclosed bay that is under strong anthropogenic influence including agriculture as well as municipal and industrial effluents (Anđelić et al., 2020). Of the four localities, Pag Bay is the most pristine and least polluted area (Kušpilić, personal communication).
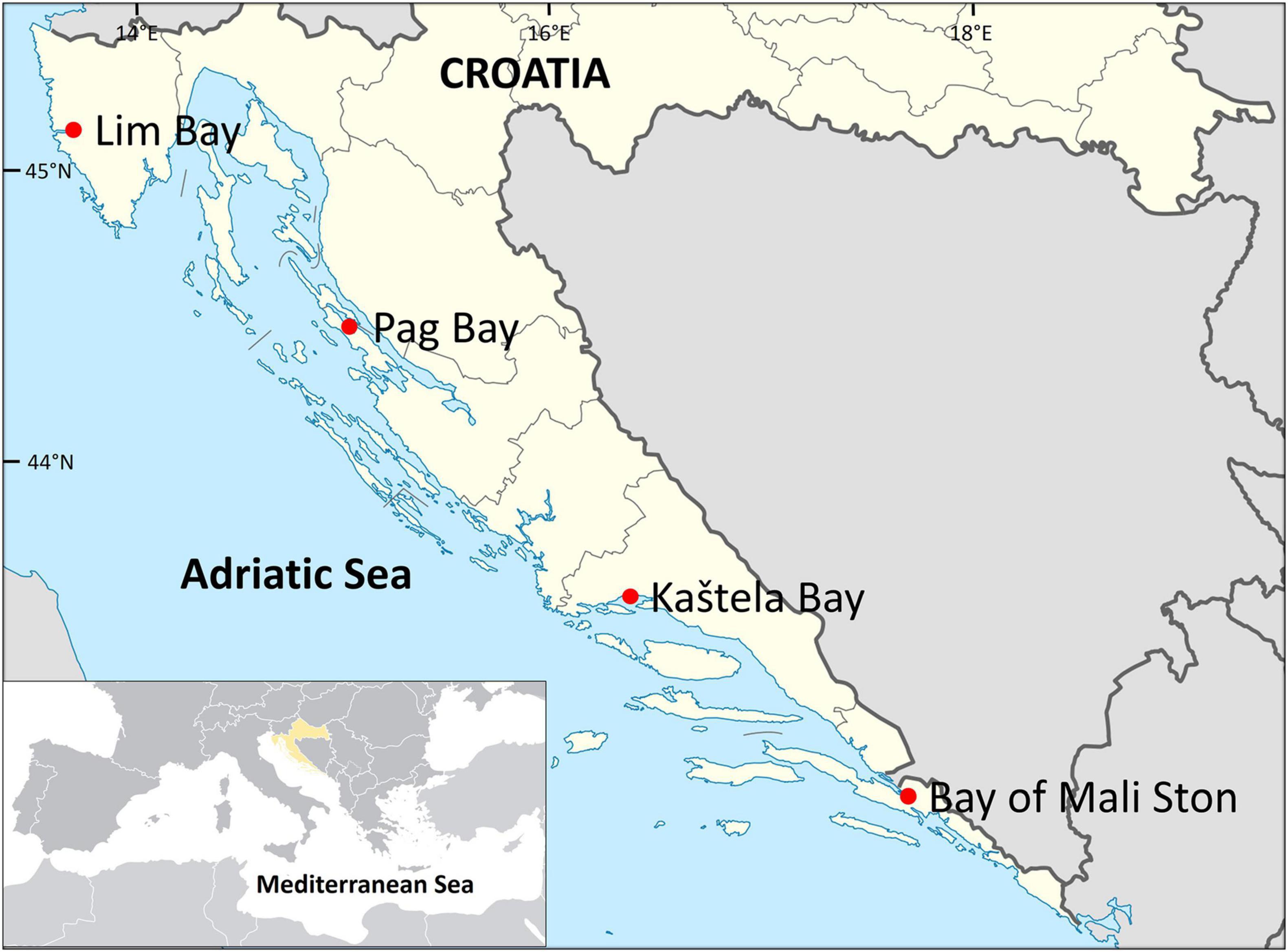
Figure 1. Map of the Adriatic Sea showing sample localities (red dots).
Sample Preparation and Stable Isotope Analysis
In the laboratory, tissue and epibionts were physically removed and shells carefully cleaned and air-dried, and shell heights were measured. Due to the P. nobilis mass mortality event and opportunistic sampling strategy it was not possible to target specific shell sizes. The height of the studied P. nobilis shells ranged from 15 cm (Pag Bay, Pag3) to 58 cm (Mali Ston Bay, Sto1) (Table 1). The average shell height was 50.7 ± 4.0, 30.3 ± 14.6, 39.8 ± 7.9, and 56.7 ± 1.5 cm for Lim Bay, Pag Bay, Kaštela Bay, and Mali Ston Bay, respectively. Muscle tissue samples from Lim Bay and Kaštela Bay were frozen for later isotope analysis.

Table 1. Overview of Pinna nobilis specimens and number of samples per shell used in this study.
Shell powder (∼20–30 mg) was collected by milling sample swaths by hand using a DREMEL Fortiflex drill equipped with a 300 μm tungsten carbide drill bit. For the analysis of spatial δ15NCBOM variations, three samples were collected from each of these shells by milling three shallow lines parallel to the growth axis from the internal shell surface (Figure 2A and Table 1). Sample tracks measured 130 ± 37 mm in length. These sample swaths started, on average, 40 ± 11 mm from the shell edge. For the analysis of temporal variability, high-resolution δ15NCBOM data were collected for the last deposited shell material from two shells collected at three localities including Lim Bay, Kaštela Bay, and Mali Ston Bay. Milling swaths were arranged perpendicular to the major growth axis on the external shell surface (Figure 2B). For shell Kas1 from Kaštela Bay, a total of 40 samples were collected, while between 14 and 17 samples were obtained from each of the five other shells. Shell powder (∼80 μg) was also used for δ18Oshell and δ13Cshell analysis. An overview of shells and samples is given in Table 1. Gillikin et al. (2017) previously established that vigorous chemical cleaning of bivalve shell carbonate is not required for CBOM analysis in bivalves due to the dense structure of the shell.
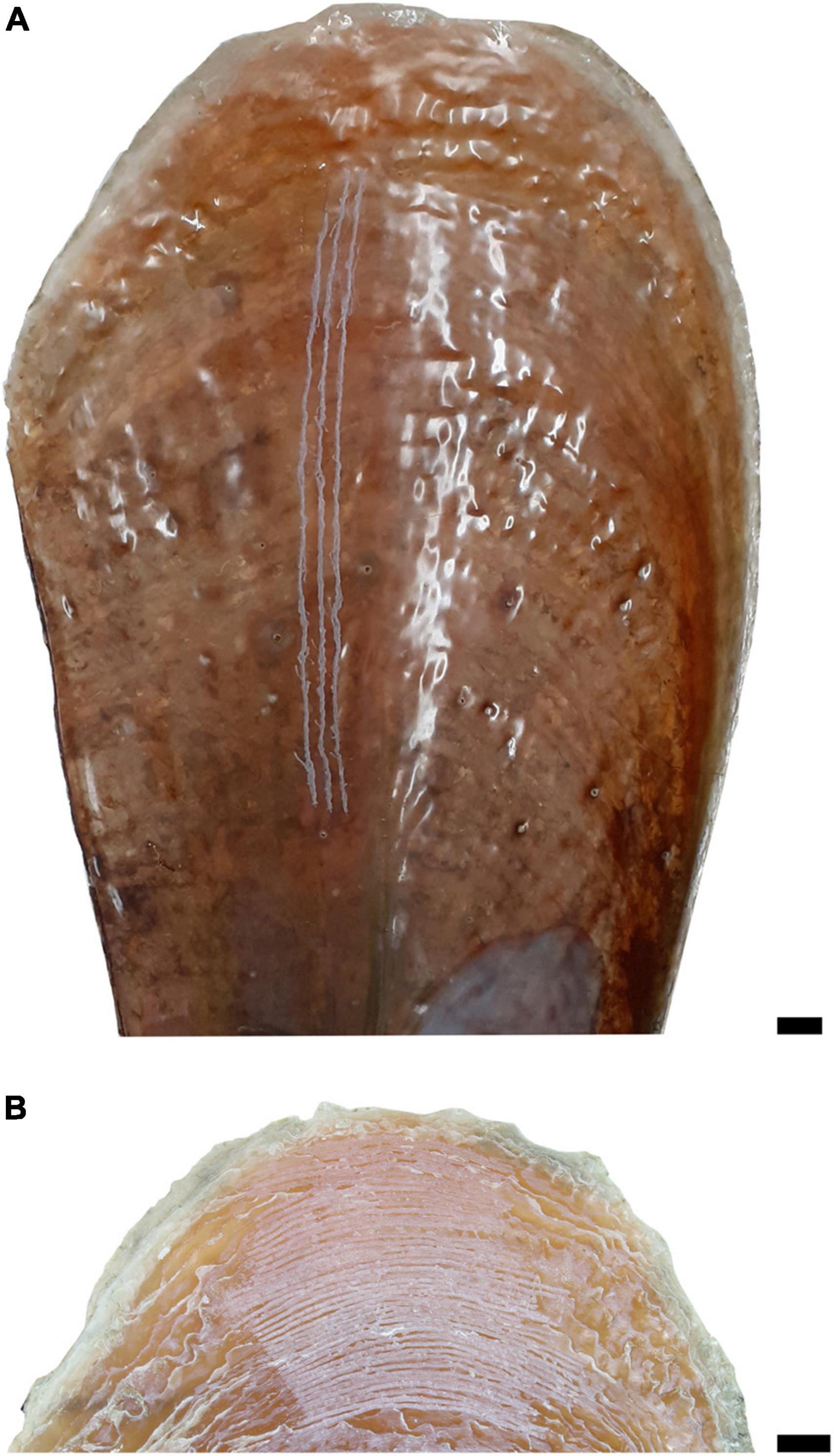
Figure 2. Illustration of milling lines on (A) inner shell surface and (B) external shell surface of Pinna nobilis Kas1 collected at Kaštela Bay. Scale bar 1 cm.
Soft tissues were dried for ∼24 h at 55°C, then ground and homogenized using a mortar and pestle. ∼1 mg powdered samples were packed into tin capsules. 20–30 mg of shell carbonate were packed into tin capsules (signal intensities on the mass spectrometer were similar between soft tissues and shell). Samples were analyzed on a Thermo Delta V Advantage isotope ratio mass spectrometer (IRMS) in continuous flow mode connected to a Costech Elemental Analyzer (EA) via ConFlo IV at Union College (Schenectady, NY) following methods in previous studies (Versteegh et al., 2011; Graniero et al., 2016, 2021; Black et al., 2017; Darrow et al., 2017; Gillikin et al., 2017; Whitney et al., 2019). An in-house acetanilide (δ15N = –0.96‰), ammonium sulfate (IAEA-N-2) (δ15N = + 20.3‰), and caffeine (IAEA-600) (δ15N = + 1.0‰) reference standards were used for regression-based corrections, and to assign the data to the appropriate isotopic scale. Percent N was calculated using additional acetanilide standards of varying mass. Corrections were done using a regression-based method. The reproducibility for δ15N (AIR) was ± 0.04‰, based on 12 acetanilide standards over three analytical sessions. Stable carbon isotope analysis used similar techniques, see Graniero et al. (2021) for complete details.
Oxygen and carbon isotopes of carbonate powders (∼80 μg) were analyzed on a Gas Bench II coupled to the aforementioned mass-spectrometer. Powders were reacted with > 100% phosphoric acid in helium-flushed Exetainer vials at 55°C for at least 3 h. Carbonate isotope results are reported in δ notation (‰) relative to the VPDB carbonate standard. NBS-18 and IAEA 603 were used for regression-based corrections; LSVEC was used as a check standard. The following values (VPDB) were used for δ13C and δ18O, respectively: IAEA 603: + 2.46‰, −2.37‰, NBS-18: −5.014‰, −23.2‰, and LSVEC: −46.6‰, −26.7‰. The reproducibility (1σ) for δ13Cshell and δ18Oshell was ± 0.03 and ± 0.04‰, respectively, based on eight IAEA 603 standards over two analytical sessions.
Environmental Data and Statistical Analysis
In this study we used results based on Regional Ocean Modeling System (ROMS), a 3D numerical ocean model for the Adriatic Sea with a spatial resolution of 2 km and 20 vertical levels in terrain following coordinates (Janeković et al., 2014). The model was previously validated in studies using available observations including CTD profiles and satellite SST data (e.g., Janeković et al., 2010, 2014; Vilibić et al., 2016) and was used to estimate daily bottom salinity and temperature data—corresponding to the first vertical level in the model. These data were used to calculate predicted δ18Oshell values and temporally align the measured δ18Oshell data of Kaštela Bay. An earlier study by García-March et al. (2011) on Pinna nobilis used Friedman and O’Neil (1977) paleothermometry equation for calcite and water and observed ∼1.4‰ offset relative to measured seawater temperature. To enable aligning of data obtained from shell with expected δ18Ocalcite, in this study we used the paleothermometry equation by Killingley and Newman (1982) (Eq. 1).
δ18Owater values were estimated from the modeled salinity values using the equation of Purroy et al. (2018) for the coastal areas in the eastern Adriatic Sea:
including a VPDB-SMOW scale correction for δ18Owater of -0.27‰ (Gonfiantini et al., 1995). Calculated δ18O values were converted from SMOW to VPDB scale using the equation from Coplen et al. (1983) to temporally align δ18Oshell values with predicted shell oxygen isotope values obtained from the ocean model:
Differences in δ15NCBOM between localities were tested using one-way ANOVA. A Shapiro-Wilk test was used to test for normality, and the Brown-Forsythe test for equal variances. The Holm-Sidak method was applied for pairwise multiple comparison. Kruskal-Wallis test was applied for the analysis of percent nitrogen. Spearman correlation coefficient was applied for testing correlation between different variables.
Results
A preliminary study of seven bivalve species from the Adriatic Sea, showed pronounced interspecies variations in %Nshell (Table 2). The lowest values (<0.03%) were noted for Venus verrucosa, and Glycymeris pilosa specimens. Shells of the commercially important European flat oyster (Ostrea edulis) and Mediterranean mussel (Mytilus galloprovincialis), had among highest values, 0.22 and 0.16%, respectively. However, the highest value (0.47%) was obtained for a Pinna nobilis shell, clearly indicating potential of this species for the analysis of δ15NCBOM data.
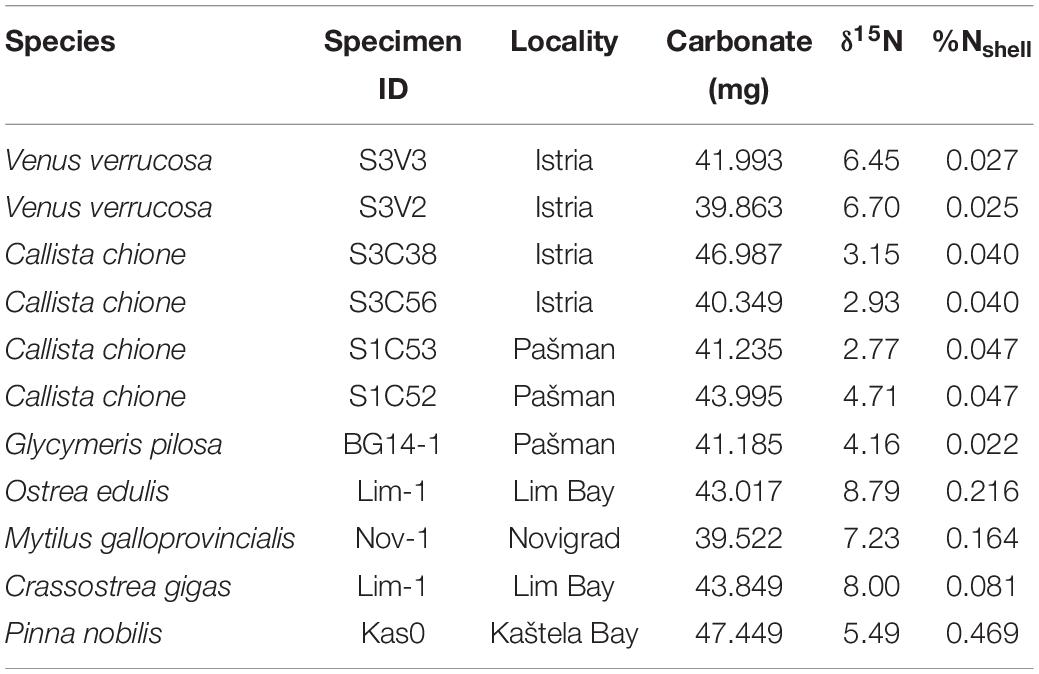
Table 2. Results of preliminary study of δ15NCBOM on bivalve species from the Adriatic Sea.
Spatial Variations in δ15NCBOM Data
Values of δ15NCBOM in samples milled from the internal shell surface of Pinna nobilis shells had pronounced spatial variations (Figure 3 and Supplementary Material 1). Lowest values were obtained for shells from Pag Bay, with a range of 3.06–.47‰ (average ± 1σ = 3.72 ± 0.41‰). Samples from Mali Ston Bay had intermediate values ranging from 5.43 to 5.96‰ (5.72 ± 0.22‰), while values between 6 and 8‰ were characteristic for P. nobilis from Lim Bay and Kaštela Bay. Observed differences were statistically significant between localities (one-way Anova F = 27.36, p < 0.001). According to pairwise comparison tests between locations, differences were not statistically significant, except for Lim Bay and Kaštela Bay. Percent N ranged from 0.2 to 0.7%. Percent nitrogen was higher in shells from Lim Bay (0.43 ± 0.16%), while Pag Bay shells had intermediate amounts (0.35 ± 0.06%), and Mali Ston Bay (0.30 ± 0.03%) and Kaštela Bay (0.28 ± 0.05%) had lower amounts. Observed differences were statistically significant between localities [Kruskal-Wallis test, H (chi2): 13.3, p < 0.005]. According to pairwise comparison tests, samples from Lim Bay had significantly higher percent nitrogen than samples from Mali Ston Bay and Kaštela Bay. Statistically significant difference was also noted between Kaštela Bay and Pag Bay. The localities with highest δ15NCBOM had both the highest (Lim Bay) and lowest percent N (Kaštela Bay).
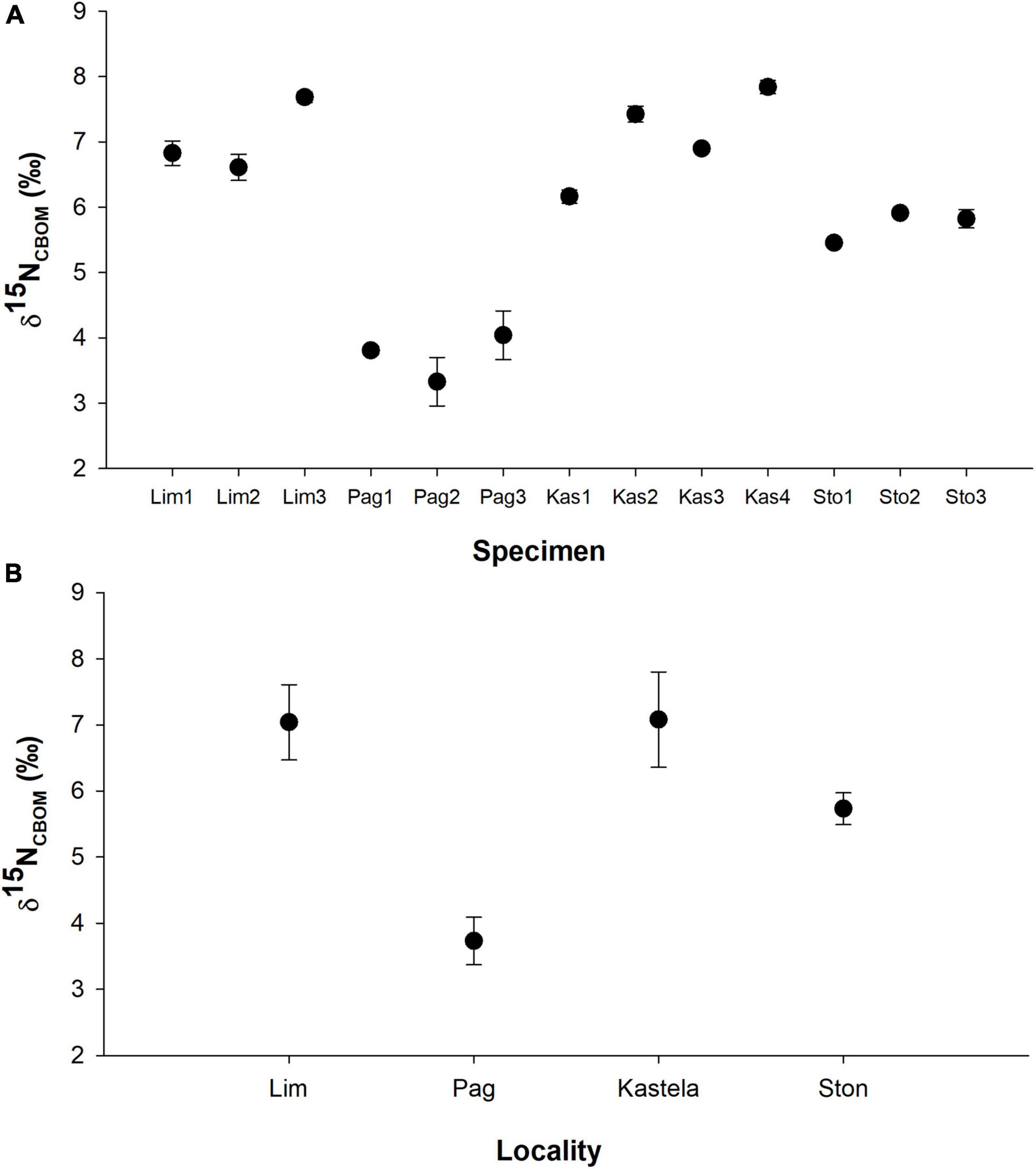
Figure 3. δ15NCBOM values of the inner shell surface of Pinna nobilis. (A) Data of the three milling lines per specimen, (B) differences between locality averages. Mean values ± 1 standard deviation (often smaller than the symbol).
High-Resolution δ15NCBOM Data
Samples from the external shell surface of Pinna nobilis show spatial and temporal variations in δ15NCBOM values. The largest number of samples came from the shell Kas1 (Kaštela Bay) and corresponded to the last 6 cm of shell growth (Figure 4). Earlier deposited shell material had δ15NCBOM values between approx. 4.5 and 5.5‰ and showed cyclic variations, while an increase in δ15NCBOM values (up to approx. 10‰) was identified closer to the shell margin. This increase in δ15NCBOM was accompanied by a pronounced δ13Cshell decrease. The δ18Oshell provided an insight into the timing and rate of shell growth. Accordingly, the sampled shell portion was formed between spring 2018 and summer 2019 (Figure 5). As this shell was collected alive in January 2020 and the last sample was milled 1.6 mm from the shell margin, these results indicate that the shell Kas1 slowed down its growth or even ceased to grow for several months prior to collection. The increase in δ15NCBOM values occurred in spring and summer of 2019. Temporal resolution of δ15NCBOM data was ∼10 days, with seasonal variations ranging between 1 and 3 weeks.
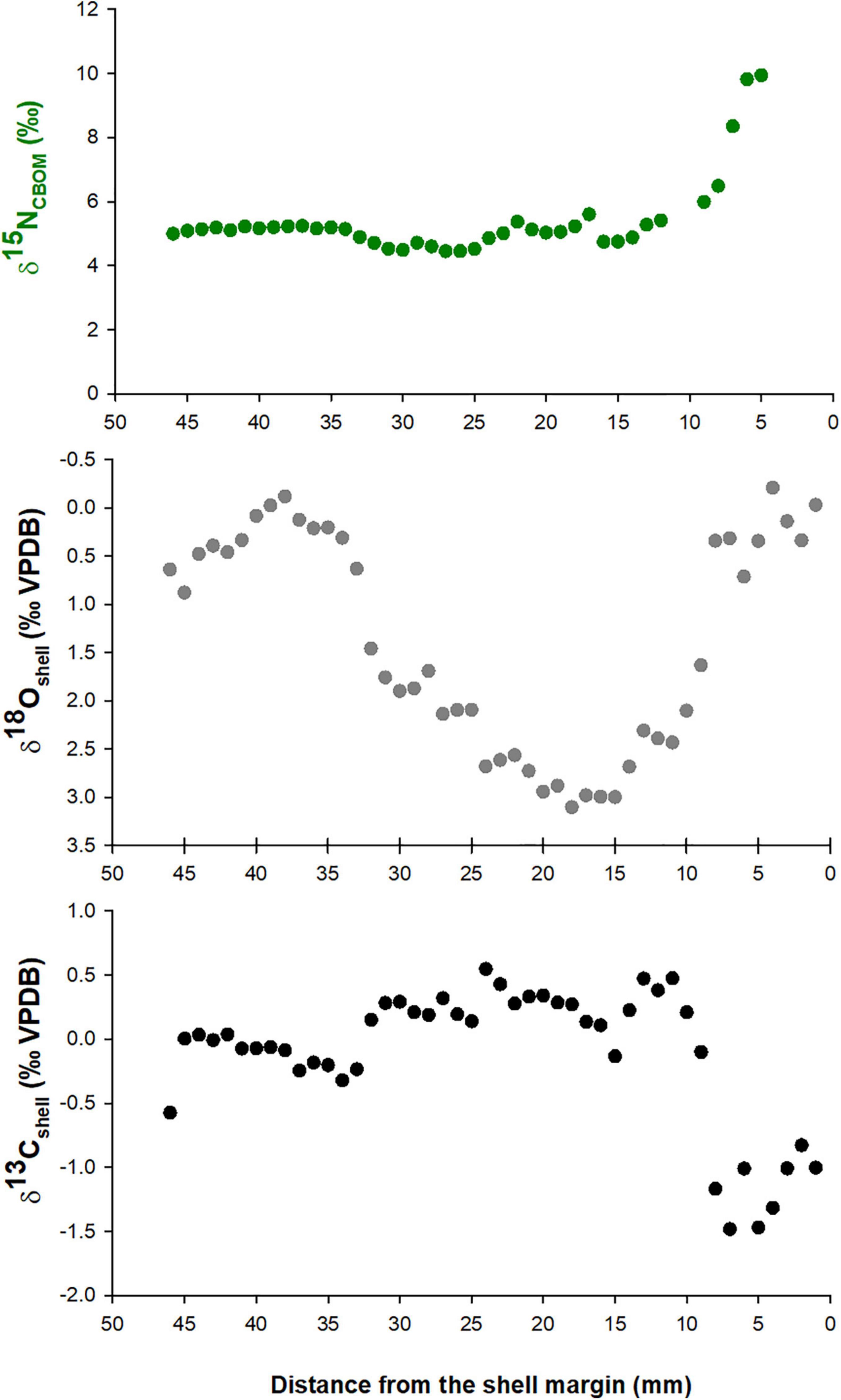
Figure 4. High resolution δ15NCBOM, δ18Oshell, and δ13Cshell data of Pinna nobilis specimen Kas1 collected alive at Kaštela Bay in January 2020.
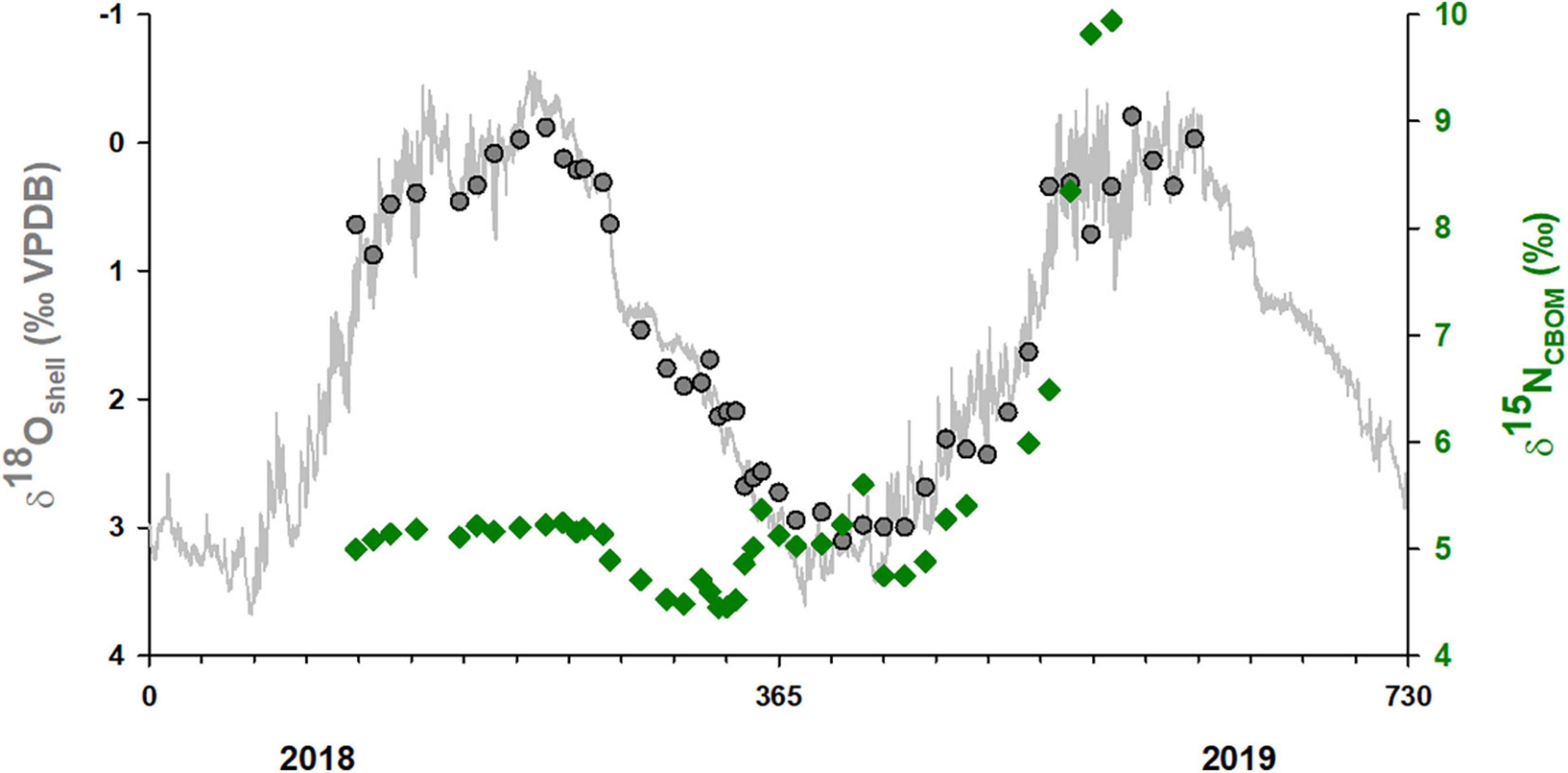
Figure 5. Predicted shell oxygen isotope values (gray line) obtained from the temperature (ROMS model) and δ18Owater estimates based on salinity (ocean model) using the paleothermometry equation by Killingley and Newman (1982). Gray circles represent temporally aligned δ18Oshell values of Pinna nobilis specimen Kas1 collected alive at Kaštela Bay in January 2020; green diamonds represent corresponding δ15NCBOM values. Scale on y1-axes is reversed.
The δ18Oshell values from shell Kas1 ranged between −0.21 and 3.10‰, while those for δ13Cshell values varied between −1.48 and 0.55‰ (N = 46). There was a statistically significant positive correlation between δ18Oshell and δ13Cshell (Spearman r = 0.74, p < 0.001). Shell δ15N values were negatively correlated to both δ18Oshell (r = −0.344, p = 0.03) and δ13Cshell (r = −0.438, p = 0.005). However, this relationship was largely driven by the last eight samples. Without these data points, the only statistically significant correlation exists between δ18Oshell and δ15NCBOM values (r = 0.667, p < 0.001).
We also completed a high-resolution analysis of five additional P. nobilis shells, two live-collected specimens from Lim Bay (Lim1 and Lim2) and Ston Bay (Sto1 and Sto3), and one dead, empty shell from Kaštela Bay (Kas4). In these shells, 15 mm (Lim2) to 23 mm (Sto3) transects were sampled, with the last samples coming from approx. 10 mm before the shell margin. In these specimens, sampling closer to the shell margin was not possible because of contamination: foreign particles, mostly sediment, were trapped in the shell.
δ15NCBOM values ranged from 3.43 to 7.84‰ (Figure 6). Lowest values were characteristic for shells from Mali Ston Bay, with an average value of 4.30 ± 0.42‰ for shell Sto1 4.09 ± 0.52‰ and for shell Sto3. Shells from Lim Bay had highest mean values, i.e., 7.00 ± 0.57‰ (Lim1) and 6.24 ± 0.36‰ (Lim2). These spatial variations are consistent with δ15NCBOM values of the inner shell surface. Shell Kas4 from Kaštela Bay showed intermediate average values for samples obtained from the outer shell (5.46 ± 0.16‰), and the least variable range of 5.22–5.74‰. In the case of this shell, average δ15NCBOM values obtained from samples milled from the outer shell were lower than those obtained from the inner shell (7.84 ± 0.10‰).
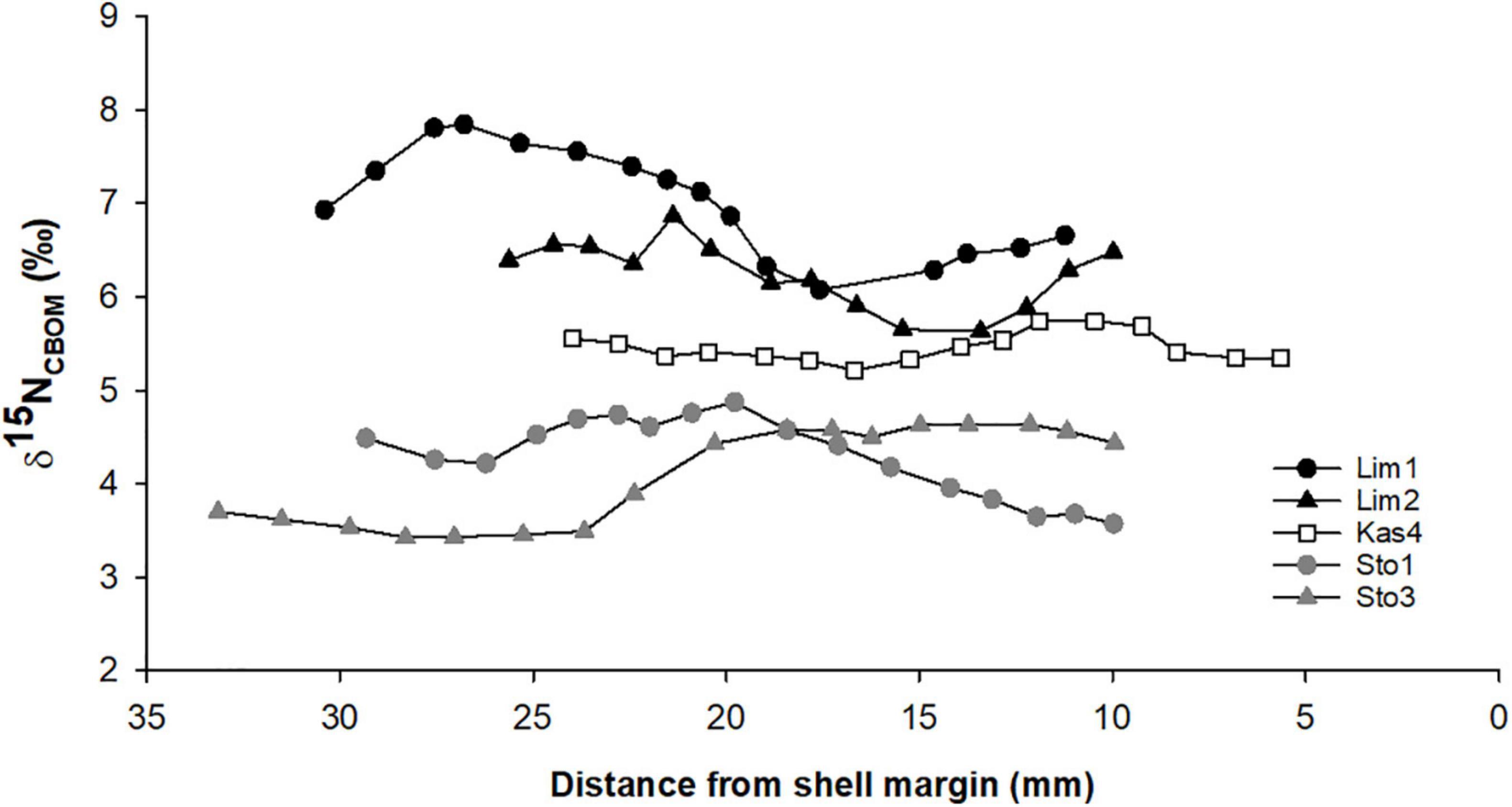
Figure 6. High-resolution δ15NCBOM values of Pinna nobilis shells collected at Lim Bay (Lim1 and Lim2), Kaštela Bay (Kas4), and Mali Ston Bay (Sto1 and Sto3).
As noted above, muscle tissues were only available for shells from Lim Bay and Kaštela Bay (Figure 7). The δ15Ntissue data of these two localities varied from each other, with higher values for Lim Bay (range: 7.22–7.28‰) and lower values for Kaštela Bay (5.74–6.19‰). In the present study, δ13Ctissue data had a very narrow range at both localities (Lim Bay: −19.88 to −19.70‰; Kaštela Bay: −20.06 to −19.57‰).
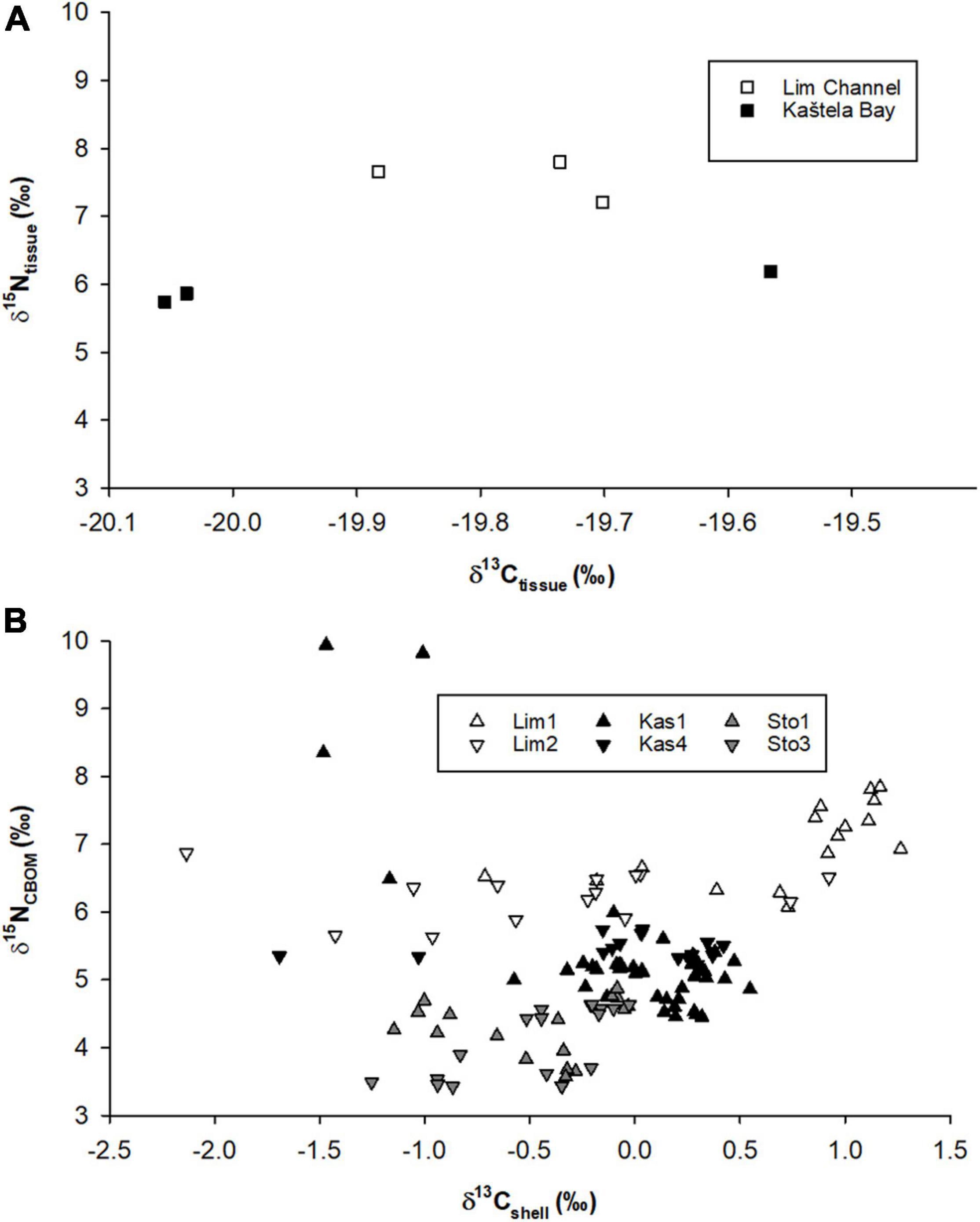
Figure 7. Relationship between δ13C and δ15N in (A) muscle tissue and (B) shell of Pinna nobilis.
Discussion
Shell δ15N Values
Results of this study clearly indicate that Pinna nobilis shells contain a sufficient amount of N-bearing organic matter to enable reliable analyses of δ15NCBOM values using direct combustion. For the analysis of site-specific/temporal δ15NCBOM variations of P. nobilis, carbonate material can be milled from the inner or outer shell surface (Figures 3, 6). Pronounced site-specific δ15NCBOM variations were observed in specimens collected in the eastern Adriatic Sea. δ15NCBOM values from the inner shell surface of specimens from Lim Bay and Kaštela Bay were almost two times higher than those from P. nobilis from the Pag Bay (Figure 3). Similar site-specific δ15NCBOM differences existed in samples from the external shell surface of the specimens collected from three localities.
Kaštela is a semi enclosed coastal bay that was identified as one of the most polluted coastal areas in the eastern Adriatic in the late 20th century (Barić et al., 1992). The situation has improved since 1990 onward when most chemical industries located in the area were closed and again after mid-2000 when a modern wastewater treatment plant was completed (Kušilić et al., 2009). However, due to large urban population in the area, including cities of Split, Kaštela and Trogir, Kaštela bay is still one of the areas with the highest anthropogenic impacts in the eastern Adriatic. Due to the number of freshwater springs, Lim Bay, an important bivalve aquaculture site, is characterized with high nutrient input (Bosak et al., 2009). Other than impacts associated with aquaculture activities, anthropogenic influences in this area are limited. A similar situation exists in Mali Ston Bay, as it is also relatively sparsely populated area. The Mali Ston Bay may be classified as a moderate natural eutrophicated system based on the phytoplankton community structure (Viličić et al., 1998). The least data are available for Pag Bay. The Island of Pag, including the Pag Bay area, is characterized by bare rocks, and scarce terrestrial vegetation. There is no industrial activity in the area, and agriculture is very limited. Previous studies (e.g., Carmichael et al., 2008; Kovacs et al., 2010; Gillikin et al., 2017) showed that δ15NCBOM data relate to nitrogen in the environment and can be used for reconstructing environmental biochemical conditions. Our results are consistent with this, as samples from the more pristine Pag Bay had the lowest δ15NCBOM values.
According to Gillikin et al. (2017) bivalve shells can provide high-resolution δ15NCBOM data because they are capable of recording the δ15N signal of metabolically-acquired N with suitably short time-averaging. The target species of the present study, P. nobilis is the largest marine bivalve in the Mediterranean Sea (Vicente, 1990; Zavodnik et al., 1991) and is characterized by rapid shell growth during early ontogeny. It attains a shell height of 20–30 cm during first 2 years of life (Richardson et al., 2004; Kožul et al., 2012; García-March et al., 2020). In this study, average temporal resolution of δ15NCBOM from the external shell samples was 10 days, and this was obtained by milling ∼20–30 mg of carbonate material. Resolution was estimated from temporal alignment of measured δ18Oshell values with predicted δ18Oshell values obtained from the ocean model. The sample mass of milled carbonate in this study was rather large, as we targeted an easy to analyze mass with a robust signal to noise ratio. The sample mass could be smaller (e.g. Gillikin et al., 2017; Graniero et al., 2021) in future studies, which would greatly increase temporal resolution; down to a few days in early ontogeny and during seasons of faster shell growth. Such extremely highly resolved δ15NCBOM has great potential for reconstructing environmental changes in coastal marine ecosystems. Gillikin et al. (2017) reported a resolution of δ15NCBOM down to 2 days in Pecten maximus shells, and P. nobilis has more than three times the N content and faster growth rates, so daily resolved δ15NCBOM should be attainable using this technique.
In this study, we conducted high-resolution analysis of δ15NCBOM on specimens ranging in size from 42 to 58 cm. Due to the mass mortality event it was not possible to collect more shells and select animals of similar size and age limiting comparison and temporal alignment of geochemical data. As Pinna nobilis does not show periodic growth marks on its shell margin, adductor muscle scars have been used to estimate its age (Richardson et al., 1999). However, later studies pointed out bias and limitation of this method including the difficulty in determining first and second scar and obscuring of scars by nacre in older shells (Richardson et al., 2004; Garcia-March and Márquez-Aliaga, 2007). Furthermore, shells from Lim Bay and Mali Ston Bay did not have distinct muscle scars on the inner shell surface (Peharda, personal observation). Hence, estimation of their age was beyond the scope of the present study. However, it is interesting to note that largest shells came from the Mali Ston Bay, and they appear to have a larger number of muscle scars than those sampled from Kaštela Bay (see Supplementary Material 3). Furthermore, shell Lim1 from Lim Bay had a similar size (47 cm) as shell Kas1 from Kaštela Bay (49 cm), but based on muscle scars visible on the internal shell surface, this shell was older and slower growing thereby likely leading to more time averaging in data from stable nitrogen isotope analysis this shells. In addition, shell layering near the shell margin was the most fragile and friable part of the shell, which resulted in trapping of particles from the environment preventing proper sampling for isotope analysis.
Taking into account the limitations described above, high-resolution δ15NCBOM data were presented according to sample distance from the shell margin (Figure 6). These data revealed strong temporal variations in specimens from same site (Lim2: Δ = 1.24‰; Lim 1: Δ = 1.76‰). In a recent study, Das et al. (2021) also observed inter- and intra- specimen variations in S. solidisima shells. Gillikin et al. (2017) analyzed Pecten maximus and noted differences of up to 2.5‰ between shells growing at the same site and time. Despite variations between specimens, data presented here and in previous studies seem to reflect that the nitrogen isotope baseline changes seasonally. This change is most likely caused by seasonal variation in nutrients and food sources available to bivalves, including P. nobilis, in shallow coastal marine habitats. According to Gillikin et al. (2017) and Das et al. (2021), bivalve shells have a high potential to reveal short-term nitrogen cycle dynamics and this is confirmed in our study.
Precise temporal alignment was only performed for one specimen (Kas1) collected from Kaštela Bay. In this specimen, it was possible to collect carbonate samples from very close to the shell margin, i.e., the last formed shell portion prior to collection. Based on modeled temperature and salinity it was possible to calculate expected δ18Oshell values. In January 2020, when sampling was conducted, Pinna nobilis population in Kaštela Bay has been impacted by a mass mortality event, and strong infiltrative inflammation that included Haplosporidium pinnae and Mycrobacterium was observed in Kas1 specimen (Šarić et al., 2020). The δ18Oshell data indicated that shell material was deposited until late summer (see Figure 5). According to results of previous studies (Kennedy et al., 2001a; Richardson et al., 2004; Freitas et al., 2005; García-March et al., 2020), P. nobilis deposits shell material in late summer and autumn period, what was not the case with our shell Kas1. As this specimen was collected in January, it is possible that it was experiencing starvation and/or infection for several months.
An increase in δ15NCBOM by up to approx. 10‰ most likely occurred in early summer and was coupled with a decrease in δ13Cshell likely indicating a stressed animal and an increase in the amount of metabolic-sourced carbon in the shell (see Lorrain et al., 2004; McConnaughey and Gillikin, 2008). Changes in δ15N have been linked to variations in diet and physiological state of the animals, and according to a recent experimental study by Prado et al. (2021), on feeding of P. nobilis. The increase in δ15N can be a consequence of starvation. As the nitrogen balance of bivalves drops below nitrogen demands, they start to break down their own tissues, thereby preferentially losing 14N and retaining 15N (see Fry, 2006 for examples). Both δ13Cshell and 15NCBOM suggest the animal was sick. This is further supported by the lack of shell growth in the second half of 2019 (Figure 5).
δ15N in Pinna nobilis Tissue
Previous studies conducted on other bivalve species found that bulk δ15NCBOM correlates with δ15N of soft tissues (e.g., O’Donnell et al., 2003; Carmichael et al., 2008; Kovacs et al., 2010; Gillikin et al., 2017; Graniero et al., 2021). In the present study, only a limited number of samples was available for tissue analysis, and due to mass mortality event and conservation status of the species, it was not possible to collect additional samples. Hence, a direct comparison of shell and tissue δ15N was not possible, and the tissue data need to be interpreted with care. Yet, the tissue δ15N values were much higher than such determined in the few previous studies on P. nobilis from relatively undisturbed settings suggesting increased anthropogenic and/or natural inputs at the study localities.
To the best of our knowledge, Kennedy et al. (2001b) provided the first stable isotope data of Pinna nobilis muscle tissue. Respective specimens came from the southeastern Spanish Mediterranean coast. These authors reported a mean value of −18.3‰ for δ13Ctissue and 3.3‰ for δ15Ntissue. In a study by Cabanellas-Reboredo et al. (2009), average stable isotope values for P. nobilis muscle tissue samples were −19.30‰ for δ13Ctissue and 3.51‰ for δ15Ntissue. According to data presented in their study, δ15Ntissue values higher than 5‰ were found only at one sampling site. Similar results were found in a third study conducted in Spain by Alomar et al. (2015). According to their results, P. nobilis tissue samples from non-Marine Protected Areas were characterized by higher δ15Ntissue and δ13Ctissue, than those from Marine Protected Areas. Deudero et al. (2017) analyzed muscle tissue of monthly collected P. nobilis and found a 0.93‰ annual valuation in δ15Ntissue, with minima recorded in March (on average, 2.54‰) and maxima in May (3.47‰). All the above studies were conducted prior to mass mortality events and their values are lower than those obtained for δ15Ntissue in the present study (5.74–7.28‰). Possible explanations for observed differences are variations in anthropogenic perturbations that suggest eutrophication and/or physiological response of animal to the mass mortality event. From the conservation perspective, it would be informative if future studies are conducted on the disease spread involving live-collected bivalves and analysis of stable isotopes in different P. nobilis tissues.
Pinnindae and Shell Chemistry
According to Huber (2010), the cosmopolitan family Pinnidae comprises more than 50 species, inhabiting tropical and temperate seas. Pinnids likewise inhabit shallow coastal regions (e.g., P. nobilis, Zavodnik et al., 1991) and deep waters (e.g., Pinna cellophana, Matsukuma and Okutani, 1986). Despite their characteristic shape and size, new species are still being discovered, e.g., in 2016, Araya and Osorio (2016) described Pinna rapanui from Easter Island, South Pacific Ocean.
Results of this study as well as earlier works conducted on the shell chemistry of P. nobilis (e.g., Richardson et al., 1999, 2004; García-March et al., 2011; Gilbert et al., 2017), clearly indicate the great potential of P. nobilis as well as related taxa for sclerochronological research. Over the past decade, research on shell chemistry has been conducted on several species of Pinnidae including Atrina fragilis (Valentine et al., 2011), Atrina rigida (Gilbert et al., 2017), and Pinna carnea (Gilbert et al., 2017). Respective studies have been conducted on modern and fossil specimens presenting possibilities to reconstruct past environmental conditions. High-resolution analysis of δ15NCBOM in different pinnids from coastal environments would provide powerful insights into anthropogenic caused perturbations of the environment, especially in areas with limited instrumental data.
Conclusion and Recommendations
Results of this study clearly indicate the potential of Pinna nobilis to provide reliable high-resolution δ15NCBOM time-series. Such analyses can be conducted on museum stored specimens for which the collection date is known, thereby extending valuable δ15N reconstructions further into the past. Furthermore, P. nobilis shells collected in the mid twentieth century at different locations within the Adriatic Sea, as well as probably in other parts of the Mediterranean, are available from private collections. Although for many of such specimens exact date of collection is not known, data on approximate year of collection is often available. Challenges of high-resolution δ15NCBOM analysis of P. nobilis shells include the crowding of growth patterns in shell material deposited during late ontogeny that can result in large time averaging, trapping of different particles from the environment (organic and inorganic) within the shell, and difficulties associated with milling samples from the margin where shell is very thin and fragile. Spatial δ15NCBOM variations can be a valuable indicator of the nitrogen isotope baseline. However, due to the current P. nobilis mass mortality event in the Mediterranean, further research on this species is very restricted and attempts should be made to identify other bivalve taxa that are relatively fast growing and whose shells contain sufficient amounts of N-bearing organic matter to enable high-resolution δ15NCBOM analyses. Results of our preliminary study indicate that two commercially important and relatively fast growing bivalves, the European flat oyster (Ostrea edulis) and Mediterranean mussel (Mytilus galloprovincialis), have high shell nitrogen content, making them interesting taxa for high resolution δ15NCBOM analysis in the Mediterranean. At a global level, other species of the Pinnidae likely serve as valuable δ15NCBOM archives as well.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
Author Contributions
MP: obtained the funding, designed the experiment, analyzed the data, and wrote the manuscript. DG: designed the experiment, conducted laboratory analysis of stable isotopes in soft tissues and shell carbonate material, analyzed the data, and wrote the manuscript. BS: designed the experiment, analyzed the data, and wrote the manuscript. AV: conducted laboratory analysis of stable isotopes in soft tissues and shell carbonate material, helped with writing, and revision of the manuscript. HU: milling of carbonate material, help with data analysis, and writing and revision of the manuscript. KM: milling of carbonate material, help with writing and revision of the manuscript. TŠ and IŽ: field work, help with writing and revision of the manuscript. IJ: 3D numerical ocean model data, help with writing and revision of the manuscript. All authors gave final approval of the version to be published.
Funding
Research has been funded by the Croatian Science Foundation under the project BivACME (IP-2019-04-8542). The U.S. National Science Foundation funded Union College’s isotope ratio mass spectrometer and peripherals (NSF-MRI #1229258).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We are grateful to Ivica Matijaca for his help with collection of samples and Madelyn Miller for assistance in the isotope lab (Union College). Sampling was performed under permission from the Croatian Ministry of Environmental Protection and Energy (CLASS UP/I-612-07/18-48/145; N°517-05-1-1-18-3 and CLASS UP/I-612-07/19-48/205; N° 517-05-1-1-19-3).
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2021.816879/full#supplementary-material
References
Alomar, C., Vázquez-Luis, M., Magraner, K., Lozano, L., and Deudero, S. (2015). Evaluating stable isotopic signals in bivalve Pinna nobilis under different human pressures. J. Exp. Mar. Biol. Ecol. 467, 77–86. doi: 10.1016/j.jembe.2015.03.006
Anđelić, I., Roje-Busatto, R., Ujević, I., Vuletić, N., and Matijević, S. (2020). Distribution of bisphenol a in sediment and suspended matter and its possible impact on marine life in Kaštela Bay. Adriatic Sea, Croatia. J. Mar. Sci. Eng. 8:480. doi: 10.3390/jmse8070480
Araya, J. F., and Osorio, C. (2016). Pinna rapanui n. sp. (Bivalvia: Pinnidae): the largest bivalve species from Easter Island. South Pacific Ocean, Chile. Pac. Sci. 70, 83–90. doi: 10.2984/70.1.7
Barić, A., Marasović, I., and Gačić, M. (1992). Eutrophication phenomena with special reference to the Kaštela Bay. Chem. Ecol. 6, 51–68. doi: 10.1080/02757549208035262
Basso, L., Vázquez-Luis, M., García-March, J. R., Deudero, S., Alvarez, E., Vicente, N., et al. (2015). The pen shell, Pinna nobilis: a review of population status and recommended research priorities in the Mediterranean Sea. Adv. Mar. Biol. 71, 109–160. doi: 10.1016/bs.amb.2015.06.002
Black, H. D., Andrus, C. F. T., Lambert, W. J., Rick, T. C., and Gillikin, D. P. (2017). δ15N values in Crassostrea virginica shells provides early direct evidence for nitrogen loading to Chesapeake Bay. Sci. Rep. 7:44241. doi: 10.1038/srep44241
Bosak, S., Burić, Z., Djakovac, T., and Viličić, D. (2009). Seasonal distribution of plankton diatoms in Lim Bay, northeastern Adriatic Sea. Acta Bot. Croat. 68, 351–365.
Cabanellas-Reboredo, M., Deudero, S., and Blanco, A. (2009). Stable-isotope signatures (δ13C and δ15N) of different tissues of Pinna nobilis Linnaeus, 1758 (Bivalvia): isotopic variations among tissues and between seasons. J. Molluscan Stud. 75, 343–349. doi: 10.1093/mollus/eyp021
Cabanellas-Reboredo, M., Vázquez-Luis, M., Mourre, B., Álvarez, E., Deudero, S., Amores, Á, et al. (2019). Tracking a mass mortality outbreak of pen shell Pinna nobilis populations: a collaborative effort of scientists and citizens. Sci. Rep. 9:13355. doi: 10.1038/s41598-019-49808-4
Carella, F., Aceto, S., Pollaro, F., Miccio, A., Iaria, C., Carrasco, N., et al. (2019). A mycobacterial disease is associated with the silent mass mortality of the pen shell Pinna nobilis along the Tyrrhenian coastline of Italy. Sci. Rep. 9:2725. doi: 10.1038/s41598-018-37217-y
Carmichael, R. H., Hattenrath, T., Valiela, I., and Michener, R. H. (2008). Nitrogen stable isotopes in the shell of Mercenaria mercenaria trace wastewater inputs from watersheds to estuarine ecosystems. Aquat. Biol. 4, 99–111. doi: 10.3354/ab00106
Catanese, G., Grau, A., Valencia, J. M., Garcia-March, J. R., Vázquez-Luis, M., Alvarez, E., et al. (2018). Haplosporidium pinnae sp. nov., a haplosporidan parasite associated with mass mortalities of the fan mussel, Pinna nobilis, in the Western Mediterranean Sea. J. Invertebr. Pathol. 157, 9–24. doi: 10.1016/j.jip.2018.07.006
Čižmek, H., Čolić, B., Gračan, R., Grau, A., and Catanese, G. (2020). An emergency situation for pen shells in the Mediterranean: the Adriatic Sea, one of the last Pinna nobilis shelters, is now affected by a mass mortality event. J. Invertebr. Pathol. 173:107388. doi: 10.1016/j.jip.2020.107388
Coplen, T. B., Kendall, C., and Hopple, J. (1983). Comparison of stable isotope reference samples. Nature 302, 236–238. doi: 10.1038/302236a0
Darriba, S. (2017). First haplosporidan parasite reported infecting a member of the Superfamily Pinnoidea (Pinna nobilis) during a mortality event in Alicante (Spain, Western Mediterranean). J. Invertebr. Pathol. 148, 14–19. doi: 10.1016/j.jip.2017.05.006
Darrow, E. S., Carmichael, R. H., Andrus, C. F. T., and Jackson, H. E. (2017). From middens to modern estuaries, oyster shells sequester source-specific nitrogen. Geochim. Cosmochim. Acta 202, 39–56. doi: 10.1016/j.gca.2016.12.023
Das, S., Judd, E. J., Uveges, B. T., Ivany, L. C., and Junium, C. K. (2021). Variation in δ15N from shell-associated organic matter in bivalves: implications for studies of modern and fossil ecosystems. Palaeogeogr. Palaeoclimatol. Palaeoecol. 562:110076. doi: 10.1016/j.palaeo.2020.110076
Deudero, S., Grau, A., Vázquez-Luis, M., Álvarez, E., Alomar, C., and Hendriks, I. E. (2017). Reproductive investment of the pen shell Pinna nobilis Linnaeus, 1758 in Cabrera National Park (Spain). Mediterr. Mar. Sci. 18, 271–284. doi: 10.12681/mms.1645
Freitas, P., Clarke, L. J., Kennedy, H., Richardson, C., and Abrantes, F. (2005). Mg/Ca, Sr/Ca, and stable-isotope (δ18O and δ13C) ratio profiles from the fan mussel Pinna nobilis: seasonal records and temperature relationships. Geochem. Geophys. Geosyst. 6:Q04D14. doi: 10.1029/2004GC000872
Friedman, I., and O’Neil, J. R. (1977). “Compilation of stable isotope fractionation factors of geochemical interest,” in Data of Geochemistry, ed. M. Fleischer (Washington D.C.: United States Government Printing Office), doi: 10.3133/pp440KK Professional Paper 440-KK.
Garcia-March, J. R., and Márquez-Aliaga, A. (2007). Pinna nobilis L., 1758 age determination by internal shell register. Mar. Biol. 151, 1077–1085. doi: 10.1007/s00227-006-0547-2
García-March, J. R., Hernandis, S., Vázquez-Luis, M., Prado, P., Deudero, S., Vincente, N., et al. (2020). Age and growth of the endangered fan mussel Pinna nobilis in the western Mediterranean Sea. Mar. Environ. Res. 153:104795. doi: 10.1016/j.marenvres.2019.104795
García-March, J. R., Surge, D., Lees, J. M., and Kersting, D. K. (2011). Ecological information and water mass properties in the Mediterranean recorded by stable isotope ratios in Pinna nobilis shells. J. Geophys. Res. Biogeosci. 116:G02009. doi: 10.1029/2010JG001461
Gilbert, P. U., Bergmann, K. D., Myers, C. E., Marcus, M. A., DeVol, R. T., Sun, C. Y., et al. (2017). Nacre tablet thickness records formation temperature in modern and fossil shells. Earth Planet Sci. Lett. 460, 281–292. doi: 10.1016/j.epsl.2016.11.012
Gillikin, D. P., Lorrain, A., Jolivet, A., Kelemen, Z., Chauvaud, L., and Bouillon, S. (2017). High-resolution nitrogen stable isotope sclerochronology of bivalve shell carbonate-bound organics. Geochim. Cosmochim. Acta 200, 55–66. doi: 10.1016/j.gca.2016.12.008
Gonfiantini, R., Sticher, W., and Rozanski, K. (1995). Standards and Intercomparison Materials Distributed by the International Atomic Energy Agency for Stable Isotope Measurements (IAEA-TECDOC–825). Vienna: International Atomic Energy Agency (IAEA).
Graniero, L. E., Grossman, E. L., and O’Dea, A. (2016). Stable isotopes in bivalves as indicators of nutrient source in coastal waters in the Bocas del Toro Archipelago. Panama. PeerJ 4:e2278. doi: 10.7717/peerj.2278
Graniero, L., Gillikin, D., Surge, D., Kelemen, Z., and Bouillon, S. (2021). Assessing δ15N values in the carbonate-bound organic matrix and periostracum of bivalve shells as environmental archives. Palaeogeogr. Palaeoclimatol. Palaeoecol. 564:110108. doi: 10.1016/j.palaeo.2020.110108
Janeković, I., Mihanović, H., Vilibić, I., and Tudor, M. (2014). Extreme cooling and dense water formation estimates in open and coastal regions of the Adriatic Sea during the winter of 2012. J. Geophys. Res. Ocean 119, 3200–3218. doi: 10.1002/2014JC009865
Janeković, I., Sikirić, M. D., Tomažić, I., and Kuzmić, M. (2010). Hindcasting the Adriatic Sea surface temperature and salinity: a recent modeling experience. Geofizika 27, 85–100.
Jennings, S., and Warr, K. J. (2003). Environmental correlates of large-scale spatial variation in the δ15N of marine animals. Mar. Biol. 142, 1131–1140. doi: 10.1007/s00227-003-1020-0
Kennedy, H., Richardson, C., Duarte, C., and Kennedy, D. (2001a). Oxygen and carbon stable isotopic profiles of the fan mussel, Pinna nobilis, and reconstruction of sea surface temperatures in the Mediterranean. Mar. Biol. 139, 1115–1124. doi: 10.1007/s002270100673
Kennedy, H., Richardson, C. A., Duarte, C. M., and Kennedy, D. P. (2001b). Diet and association of Pontonia pinnophylax occurring in Pinna nobilis: insights from stable isotope analysis. J. Mar. Biol. Assoc. U.K 81, 177–178. doi: 10.1017/S0025315401003575
Kersting, D., Benabdi, M., Čižmek, H., Grau, A., Jimenez, C., Katsanevakis, S., et al. (2019). Fan Mussel Pinna nobilis. London: IUCN. The IUCN Red List of Threatened Species.
Killingley, J., and Newman, W. A. (1982). 18O fractionation in barnacle calcite: a barnacle paleo temperature equation. J. Mar. Res. 40, 893–902.
Kovacs, C. J., Daskin, J. H., Patterson, H., and Carmichael, R. H. (2010). Crassostrea virginica shells record local variation in wastewater inputs to a coastal estuary. Aquat. Biol. 9, 77–84. doi: 10.3354/ab00228
Kožul, V., Glavić, N., Bolotin, J., and Antolović, N. (2012). Growth of the fan mussel Pinna nobilis (Linnaeus, 1758) (Mollusca: Bivalvia) in experimental cages in the South Adriatic Sea. Aquac. Res. 44, 31–40. doi: 10.1111/j.1365-2109.2011.03003.x
Kukolich, S., and Dettman, D. (2021). Reconstructing seasonal and baseline nitrogen isotope ratios in riverine particulate matter using freshwater mussel shells. Geochem. Geophys. Geosyst. 22:e2020GC009239. doi: 10.1029/2020GC009239
Kušilić, G., Marasović, I., Krstulović, N., Šolić, M., Ninčević-Gladan, Ž, Bojanić, N., et al. (2009). “Restoration potential of eutrophic waters adjacent to large coastal cities: lessons from the coastal zone of Croatia,” in Proceedings of the Workshop Proceedings “Impact of Large Coastal Mediterranean Cities and Maritime Ecosystems (Alexandria).
Lattos, A., Bitchava, K., Giantsis, I. A., Theodorou, J. A., Batargias, C., and Michaelidis, B. (2021). The implication of Vibrio bacteria in the winter mortalities of the critically endangered Pinna nobilis. Microorganisms 9:922. doi: 10.3390/microorganisms9050922
Lorrain, A., Paulet, Y. M., Chauvaud, L., Dunbar, R., Mucciarone, D., and Fontugne, M. (2004). δ13C variation in scallop shells: increasing metabolic carbon contribution with body size? Geochim. Cosmochim. Acta 68, 3509–3519. doi: 10.1016/j.gca.2004.01.025
Matsukuma, A., and Okutani, T. (1986). Studies on the Kawamura collection (Mollusca) in the National Science Museum, Tokyo- II. Catalogue of type specimens, with description of Pinna cellophana n. sp. (Bivalvia). Jap. J. Malac. 45, 1–10.
McConnaughey, T. A., and Gillikin, D. P. (2008). Carbon isotopes in mollusk shell carbonates. Geo-Mar. Lett. 28, 287–299. doi: 10.1007/s00367-008-0116-4
O’Donnell, T. H., Macko, S. A., Chou, J., Davis-Hartten, K. L., and Weshmiller, J. F. (2003). Analysis of δ13C, δ15N, and δ34S in organic matter from the biominerals of modern. Org. Geochem. 34, 165–183. doi: 10.1016/S0146-6380(02)00160-2
Oczkowski, A., Gumbley, T., Carter, B., Carmichael, R., and Humphries, A. (2016). Establishing an anthropogenic nitrogen baseline using Native American shell middens. Front. Mar. Sci. 3:79. doi: 10.3389/fmars.2016.00079
Prado, P., Cabanes, P., Hernandis, S., García-March, R., and Tena, J. (2021). Stable isotope analyses reveal major nutritional deficiencies in captive vs. field juvenile individuals of Pinna nobilis. Mar. Environ. Res. 168:105304. doi: 10.1016/j.marenvres.2021.105304
Purroy, A., Milano, S., Schöne, B. R., Thébault, J., and Peharda, M. (2018). Drivers of shell growth of the bivalve, Callista chione (L. 1758) – combined environmental and biological factors. Mar. Environ. Res. 134, 138–149. doi: 10.1016/j.marenvres.2018.01.011
Richardson, C. A., Peharda, M., Kennedy, H., Kennedy, P., and Onofri, V. (2004). Age, growth rate and season of recruitment of Pinna nobilis (L) in the Croatian Adriatic determined from Mg:Ca and Sr:Ca shell profiles. J. Exp. Mar. Biol. Ecol. 299, 1–16. doi: 10.1016/j.jembe.2003.08.012
Richardson, C., Kennedy, H., Duarte, C., Kennedy, D. P., and Proud, S. (1999). Age and growth of the fan mussel Pinna nobilis from south-east Spanish Mediterranean seagrass (Posidonia oceanica) meadows. Mar. Biol. 133, 205–212. doi: 10.1007/s002270050459
Šarić, T., Župan, I., Aceto, S., Villari, G., Palić, D., De Vico, G., et al. (2020). Epidemiology of noble pen shell (Pinna nobilis L. 1758) mass mortality events in Adriatic Sea is characterised with rapid spreading and acute disease progression. Pathogens 9:776. doi: 10.3390/pathogens9100776
Scarpa, F., Sanna, D., Azzena, I., Mugetti, D., Cerruti, F., Hosseini, S., et al. (2020). Multiple non-species-specific pathogens possibly triggered the mass mortality in Pinna nobilis. Life 10:238. doi: 10.3390/life10100238
Schöne, B. R., and Huang, Q. (2021). Ontogenic δ15N trends and multidecadal variability in shells of bivalve Mollusc. Arctica islandica. Front. Mar. Sci. 8:748593. doi: 10.3389/fmars.2021.748593
Thibault, M., Duprey, N., Gillikin, D. P., Thébault, J., Douillet, P., Chauvaud, L., et al. (2020). Bivalve δ15N isoscapes provide a baseline for urban nitrogen footprint at the edge of a World Heritage coral reef. Mar. Pollut. Bull. 152:110870. doi: 10.1016/j.marpolbul.2019.110870
Valentine, A., Johnson, A. L. A., Leng, M. J., Sloane, H. J., and Balson, P. S. (2011). Isotopic evidence of cool winter conditions in the mid-Piacenzian (Pliocene) of the southern North Sea Basin. Palaeogeogr. Palaeoclimatol. Palaeoecol. 309, 9–16. doi: 10.1016/j.palaeo.2011.05.015
Vázquez-Luis, M., Álvarez, E., Barrajón, A., García-March, J., Grau, A., Henriks, I. E., et al. (2017). S.O.S. Pinna nobilis: a mass mortality event in Western Mediterranean Sea. Front. Mar. Sci. 4:220. doi: 10.3389/fmars.2017.00220
Versteegh, E. A. A., Gillikin, D. P., and Dehairs, F. (2011). Analysis of δ15N values in mollusk shell organic matrix by elemental analysis/isotope ratio mass spectrometry without acidification: an evaluation and effects of long-term preservation. Rapid Commun. Mass Spectrom. 25, 675–680. doi: 10.1002/rcm.4905
Vicente, N. (1990). Estudio ecológico y protección del molusco lamelibranquio Pinna nobilis L. 1758 en la costa Mediterránea. Iberus 9, 269–279.
Vilibić, I., Mihanović, H., Janeković, I., and Šepić, J. (2016). Modelling the formation of dense water in the northern Adriatic: sensitivity studies. Ocean Model. 101, 17–29. doi: 10.1016/j.ocemod.2016.03.001
Viličić, D., Jasprica, N., Carić, M., and Burić, Z. (1998). Taxonomic composition and seasonal distribution of microphytoplankton in Mali Ston Bay (eastern Adriatic). Acta Bot. Croat. 57, 29–48.
Watanabe, S., Kodama, M., and Fukuda, M. (2009). Nitrogen stable isotope ratio in the manila clam, Ruditapes philippinarum, reflects eutrophication levels in tidal flats. Mar. Pollut. Bull. 58, 1447–1453. doi: 10.1016/j.marpolbul.2009.06.018
Whitney, N. M., Johnson, B. J., Dostie, P. T., Luzier, K., and Wanamaker, A. D. (2019). Paired bulk organic and individual amino acid δ15N analyses of bivalve shell periostracum: a paleoceanographic proxy for water source variability and nitrogen cycling processes. Geochim. Cosmochim. Acta 254, 67–85. doi: 10.1016/j.gca.2019.03.019
Keywords: bivalve, stable isotope, shell, Mediterranean, Adriatic
Citation: Peharda M, Gillikin DP, Schöne BR, Verheyden A, Uvanović H, Markulin K, Šarić T, Janeković I and Župan I (2022) Nitrogen Isotope Sclerochronology—Insights Into Coastal Environmental Conditions and Pinna nobilis Ecology. Front. Mar. Sci. 8:816879. doi: 10.3389/fmars.2021.816879
Received: 17 November 2021; Accepted: 20 December 2021;
Published: 27 January 2022.
Edited by:
Liqiang Zhao, Guangdong Ocean University, ChinaReviewed by:
Nina Whitney, Woods Hole Oceanographic Institution, United StatesFrank Dehairs, Vrije University Brussel, Belgium
Copyright © 2022 Peharda, Gillikin, Schöne, Verheyden, Uvanović, Markulin, Šarić, Janeković and Župan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Melita Peharda, bWVsaXRhQGl6b3IuaHI=