 Marta Gellert
Marta Gellert Graham Bird
Graham Bird Anna Stȩpień
Anna Stȩpień Maciej Studzian
Maciej Studzian Magdalena Błażewicz
Magdalena Błażewicz- 1Department of Invertebrate Zoology and Hydrobiology, University of Lodz, Lodz, Poland
- 2Independent Researcher, Waikanae, New Zealand
- 3Department of Molecular Biophysics, University of Lodz, Lodz, Poland
- 4Laboratory of Transcriptional Regulation, Institute of Medical Biology, Polish Academy of Sciences (PAS), Lodz, Poland
A new family of paratanaoidean Tanaidacea, the hamatipedids, formerly part of the Typhlotanaidae, is established to accommodate three genera (Hamatipeda and two new). Deep-sea hamatipedids collected from four sites in the Atlantic (Argentine and Guiana basins) and 14 sites from the Southeast coast of Australia were studied using a taxonomic approach combining morphological and morphometric data. Four new species of Hamatipeda and one of a new genus are described from different deep-sea areas of the Atlantic and Pacific oceans. Hamatipeda sima originally classified within Hamatipeda, is transferred to a new genus. We observed that several morphometric characters (i.e., length of the last two pereonites) in different life-stages of one of the new Hamatipeda species (neuter, manca-2, and manca-3) are correlated with the total body length (TBL). Applying a morphometric approach, we aimed: (1) to identify those ontogenetic-dependent characters, and (2) to detect the characters, which can be used in discrimination of Hamatipedidae species, regardless their life-stage.
Introduction
Intensive exploration of the deep sea since the 1960s has uncovered a remarkable diversity of benthic organisms (Sanders et al., 1965; Sanders, 1968). The large number of new taxa discovered during each expedition unequivocally refute the 19th Century concept of the deep sea as unproductive and devoid of life ecosystem. These studies also revealed that many undescribed deep-sea species are often much smaller than their shallow-water counterparts, a reason why these have been overlooked over the decades of deep-ocean exploration (Larsen, 2005; Błażewicz-Paszkowycz et al., 2012; McCallum et al., 2015; Frutos et al., 2016). The number of new species and their enormous diversity discovered during each deep-sea expedition, confirms that oceanic bottom is the last recognized ecosystem of the Earth (Ramirez-Llodra et al., 2011; Frutos and Sorbe, 2014; Costello and Chaudhary, 2017; Jażdżewska et al., 2018, 2021). A paucity of specialists and awareness of the role taxonomy for understanding and protection of the biodiversity has meant that collections of invertebrates from deep-sea expeditions were shelved in museums awaiting the attention of taxonomists and formal description (Brandt et al., 2007; Appeltans et al., 2012).
Among these organisms is the superfamily Paratanaoidea Lang, 1949, a monophyletic group of the crustacean suborder Tanaidomorpha (Kakui et al., 2011). It is represented by relatively small peracarids (<4 mm) of high diversity that is still under-recognized and undescribed (Błażewicz-Paszkowycz and Bamber, 2007; Błażewicz et al., 2019). It is currently represented by 19 recent families.
The Typhlotanaidae Sieg (1984) is one of the most diverse paratanaoidean families in the deep sea, comprising 17 genera and 119 species (Gellert et al., unpublished). Before the first phylogenetic approaches (Larsen and Wilson, 2002), typhlotanaids were grouped within the Leptognathiidae Sieg, 1976, although their morphological distinctiveness was often emphasized (Sieg, 1984). The Typhlotanaidae were characterized by a three-articulated antennule, six-articulated antenna (Sieg, 1984), and absence of eyes, which are considered evidence for a deep-water origin of the family (Błażewicz-Paszkowycz, 2007; Gellert et al., unpublished). The morphological distinctness of the ornamentation of their pereopods and their monophyletic origin is still being resolved, with the “true” typhlotanaids being defined by the presence of a “clinging apparatus” on the carpus of pereopods 4–6, which facilitates movement within their tubicolous domiciles. This apparatus includes several specialized sets of hooks, thorns, and pectinate spines rather than simple “bayonet” spines. Additionally, some genera have rounded and minutely spinulate structures called “prickly tubercles” (Błażewicz-Paszkowycz, 2007). Those structures are apparently absent in some genera such as Aremus Segadilha et al., 2018 and Hamatipeda Błażewicz-Paszkowycz, 2007, leading to their affinity with Typhlotanaidae being questioned (Segadilha et al., 2018).
The basis for this paper is tanaid material collected during the pioneering expedition exploring the abyssal zone of the West Atlantic and slope off southeastern Australia. Those collections were deposited in the Museum of Comparative Zoology (Boston, MA, United States) and in the Melbourne Museum (Australia). The material was initially identified to the genus Hamatipeda, but closer identification has revealed a richer diversity allowing us to distinguish several taxa. Four of the species are formally described (three species from the SW Atlantic and one from SE Australia), two new genera are established, and analysis of morphological characteristics confirms that aspects of the attachment of the cheliped to the cephalothorax, pereopod setation, and the shape of the carpus cheliped enable us to define a new family.
Materials and Methods
Sampling
The 5,832 typhlotanaids specimens for the research were collected during the expeditions completed in the SW Atlantic (5,771 individuals) and SE Australia (61 individuals). From the Atlantic, the material included: 147 individuals from the expedition organized by Woods Hole Oceanographic Institution during 1971–1972 aboard the RV Knorr found in two places, e.g., the Guiana Basin and Argentine Basin at a wide depth range (1,022–3,317 m), and 61 individuals from the slope and abyss off the Australian coasts of New South Wales (off Eden) to Tasmania (off Freycinet Peninsula) at a wide depth range (49–2,900 m) collected during the SLOPE Program during 1979–1988. The distribution of the stations is given in Supplementary Table 1.
The samples were preserved in formalin. Distribution maps were prepared for each species using the QGIS 2.18 software (Szczepanek, 2017). The type-material and other materials studied for this research are deposited at Museums Victoria, Melbourne Museum (NMV) (Melbourne, Australia) and the Museum of Comparative Zoology (MCZ), Harvard University (Cambridge, MA, United States).
Morphological Analyses
Initial species identification was based on morphological observations with a dissecting microscope. The whole collection was sorted to several morpho-groups, and 208 individuals were preliminarily identified as Hamatipeda and were chosen for further comprehensive morphological study. From each group several individuals were designated for thorough morphological analysis and dissected with chemically sharpened tungsten needles. The dissected cephalothorax, pereon, and pleon appendages were mounted on slides using glycerin and sealed with molten paraffin (Błażewicz et al., 2021). Morphological drawings were prepared using a light microscope (Nikon Eclipse 50i) equipped with a camera lucida. Digital pictures were completed using a graphic tablet following Coleman (2003).
Total body length (TBL) was measured along the central axis of symmetry, from the rostrum to the tip of the pleotelson. In contrast, body width was assessed perpendicular to the symmetry axis at the widest point (BW). Body width and length of cephalothorax, pereonites, pleonites, and pleotelson were measured on whole specimens. Hamatipeda mojito n. sp. (see below) was represented by numerous specimens of different ontogenetic stages. Observed variability of morphometric characters between life stages pose the question if the length of appendages changes proportionally to increasing body size during developmental growth (isometric growth) or not (allometric growth). In total, we measured ninety-seven specimens of H. mojito in three life stages: manca-2 (35 individuals), manca-3 (24 individuals) and neuter (38 individuals). For each specimen nine characters i.e., body length, pereonites 1–6 length, uropodal exopod and endopod configuration, were recorded. All measurements were assessed along the axis of symmetry and were made with a camera connected to the microscope (Nikon Eclipse Ci-L) and the NIS-Elements View software1.
Morphological terminology is largely as in Błażewicz-Paszkowycz (2007). The seta types are recognized as: (1) simple setae—without ornamentation, (2) serrate—with serration or denticulation, (3) penicillate—with a tuft of setules located distally and with a small knob on which a seta is fixed to the tegument and (4) rod setae—slightly inflated distally and with a pore followed Jakiel et al. (2020).
Stout setae (L:W < 5.0) are called spines (= spiniform setae), and the robust pereopod 4–6 carpal spines which are curved and extremely robust are called “hooks” (Błażewicz-Paszkowycz, 2007) and some, which are apically blunt are described “molariform”. Unspecified setae in taxonomic description refer to simple setae by default. Tegumental extension on appendages arranged in combs [comb-like scales in terminology by Garm and Watling (2013)] are called microtrichia (e.g., Błażewicz-Paszkowycz, 2007).
Statistical Analysis
The relationships between body size, body segments, and uropod rami measured at different developmental stages were presented as a power function (y = axb) and logarithmically linearized (log y = log a + b log x). All dimensions were log transformed before computing the regression equation. The slope of the regression line (b) represents the relative growth and was used to test the degree of allometry: isometry (b = 1), negative allometry (b < 1) or positive allometry (b > 1) (Hartnoll, 1982) (Supplementary Table 2).
Imaging
The scanning electron microscope work was performed on a Phenom Pro X (Department of Invertebrate Zoology and Hydrobiology, University of Lodz, Poland) to examine fine morphological details in a subset of specimens from the MCZ collection. Specimens were frozen at −10°C and analyzed using a temperature-controlled sample holder. Confocal laser scanning microscopy (CLSM) images were obtained with LSM 780 (Zeiss) microscope equipped with Plan-Apochromat 63x/1.4 objective using InTune tuneable excitation laser system (set to excitation wavelength 555 nm). Specimens were stained for 24 h with equal volume mixture of saturated water solutions of Congo red and acid fuchsin. Before dissection and mounting in 100% glycerol, stained animals were washed thoroughly with 50% aqueous glycerol solution. Fluorescence was registered in single emission channel: 561–695 nm. Images were recorded as Z-stacks with 12.6 μs pixel dwell and two times line averaging with optical cross section of 0.5 μm. Collected data was pseudo-colored in gold and reconstructed into a 3D image stack by maximum intensity projection using ZEN software (Zeiss).
Taxonomy
Suborder: Tanaidomorpha Sieg, 1980 Superfamily Paratanaoidea Lang, 1949 Family Hamatipedidae Błażewicz, Gellert and Bird, n. fam. LSID urn:lsid:zoobank.org:act:1F0F139B-5F63-481C-AED2-0C1CEA2B590E.
Diagnosis: Body long > 10 L:W. Pereonite-1 long (subequal or longer than cephalothorax), without hyposphenium. Pereonites 1–5 longer than wide (pereonites 2–3 over 1.5 L:W). Antennule three-articled. Mandible molar process wide, crushing surface and irregular edge, without tubercles and teeth. Cheliped basis not reaching pereonite-1, posterior lobe enfolded by sclerite. Pereopods 1–3 with seta on coxa; pereopods 4–6 coxa fused with body. Pereopod-1 more slender than others. Pereopods 2–3 often robust, with short setae and small spines. Pereopods 4–6 merus and carpus with two (or three*) hooks and one molariform spine, carpus without prickly tubercles; unguis trifurcate (or bifurcate*). Pleopods small, vestigial; setae always plumose. Uropod endopod one or two-articled; exopod one-articled.2
Male: Unknown.
Type species: Hamatipeda trapezoida Błażewicz-Paszkowycz (2007).
Genera included: Hamatipeda Błażewicz-Paszkowycz (2007); Rakaduta n. gen.; Yarutanais n. gen.
Remarks: The robust carpal hooks of pereopods 4–6 in the absence of a prickly tubercle are an autapomorphy for the new family and the character that distinguishes its members from the Typhlotanaidae. The Hamatipedidae lack both a hyposphenium on the pereonite-1 and the prickly tubercles on the carpus of pereopods 4–6, both characteristic of “true” typhlotanaids. So far, six typhlotanaid genera, i.e., Paratyphlotanais Kudinova-Pasternak and Pasternak, 1978, Meromonakantha Sieg, 1986, Obesutanais Larsen et al., 2006, Targaryenella Błażewicz and Segadilha, 2019; Typhlamia Błażewicz-Paszkowycz (2007) and Aremus lack the prickly tubercles, although they have robust spines or bayonet setae (or “bayonet-like spines,” Bird and Holdich, 1988) on the carpus of pereopods 4–6. There are several other features which distinguish the Hamatipedidae from those genera and these genera can be segregated into two groups based on the setation of pereopods 1–3: the first, Meromonakantha, Paratyphlotanais, and Targaryenella differ from all other typhlotanaid and hamatipedid genera in having simple (or bayonet-like) spines on the carpus (and merus to lesser extent). The second group, Aremus, Obesutanais, and Typhlamia share the general setation pattern of the hamatipedids and typhlotanaids (sensu stricto) but the first lacks pleopods in females and has an articulated spine on the antennule apex, Obesutanais is characterized by a short and compact body (3.0–6.0x L:W) and only a pair of hooks on the carpus of pereopods 4–6, inter alia, and Typhlamia by its long antennule article-3 (about 14x L:W) and long setae, and more elongate merus, carpus and propodus of pereopods 1–3. Hamatipedidae have elongated pereonites and the cheliped basis posterior lobe separated from the pereonite-1 (Figure 1), making it similar to Typhlamia at first glance. However, hamatipedids have a shorter antennule than Typhlamia, and their carpal clinging apparatus of pereopods 4–6 is different. In hamatipedids it is formed by hooks (naked or serrated), which are sometimes apically flattened (molariform). In Typhlamia the spines are small, slender, and only distally flattened (Gellert et al., unpublished). The morphological phylogeny of Typhlamia and Hamatipedidae should be corroborated in future analysis implementing molecular techniques.

Figure 1. Hamatipeda mojito n. sp.: cheliped/cephalothorax details showing sclerite enfolding the posterior lobe of the cheliped basis.
Hamatipedidae is the seventh paratanaoid family whose females have three-articled antennules. It comprises three genera: Hamatipeda that was originally classified in the Typhlotanaidae (Błażewicz-Paszkowycz, 2007), and two newly erected genera (Rakaduta n. gen.; Yarutanais n. gen.). A taxonomic key for identification of the paratanaoidean families with a three-articled antennule, and Hamatipedidae genera are presented below.
Key for identification of Paratanaoidea with three-articled antennule (neuters) and uropod endopod with one or two articles.
1. Antenna present ……………………………………………………………………2
- Antenna with one or two articles …………….. genus Agathotanais
2. Pereonite-1 much shorter than broad; pereopods 2–3 carpus with or without blade-like spine…………………….. Pseudotanaidae
- Pereonite-1 not much shorter than broad; pereopods 2–3 carpus with setae or simple spines ………………………………………..3
3. Eyes present; pereopods 2–3 carpus with two inferodistal spines…………………………………………………………………Nototanaidae
- Eyes absent; pereopods 2–3 carpus with two inferodistal spines……………………………………………………………………………………4
4. Cephalothorax shape narrower anteriorly, cheliped dactylus clearly more slender than fixed finger, often rugose dorsally……………………………………………………………….Tanaissuidae
- Cephalothorax not narrower anteriorly, dactylus slightly narrower than fixed finger, smooth……………………………………….5
5. Pereopods 4–6 carpus with prickly tubercles ………………………….. …………………………………………………………….Typhlotanaidae (part)
- prickly tubercles absent………………………………………………………….6
6. Pereopod 4–6 carpus with distal hooks or molariform spines…………………………………………………..Hamatipedidae n. fam.
- Pereopod 4–6 carpus with distal spines…………………………………7
7. Cephalothorax shape narrower anteriorly, margin straight………………………………………………………….Paratyphlotanais
- Cephalothorax rounded………………………………………………………..8
8. Pleopods absent………………………………………………………… Aremus
- Pleopods present………………………………………………………………….. 9
9. Antennule article-3 at least 10x L:W……………………….Typhlamia
- Antennule article-3 clearly less than 10 × LW……………………..10
10. Body well calcified, cephalothorax rounded; antennule conical, shorter than cephalothorax…………………………Meromonakantha
- Body weakly calcified, cephalothorax not rounded, antennule longer than cephalothorax………………………………………………….. 11
11. Body short (<6.0x L:W); uropodal exopod one- articled ……………………………………………………………………………..Obesutanais
- Body long (>7.0x L:W); uropodal exopod two- articled……………………………………………………………….Targaryenella
Hamatipeda Błażewicz-Paszkowycz (2007)
Diagnosis (after Błażewicz-Paszkowycz, 2007, amended): Pereonite-1 0.7–0.8x pereonite-2. Antennule article-3 long (>3.0x L:W); article-1 ventral microtrichia absent. Antenna articles 2–3 with setae. Cheliped carpal shield absent. Pereopod-1 propodus long (>3.0x L:W). Pereopod-3 carpus with few (2–3) setae. Pereopod-5 propodus seta short.
Type species: Hamatipeda trapezoida Błażewicz-Paszkowycz (2007) (by designation).
Species included: Hamatipeda kohtsukai Kakui and Hiruta (2021); H. longa (Kudinova-Pasternak, 1975); H. trapezoida Błażewicz-Paszkowycz (2007); H. prolata Segadilha et al. (2019); H. caipirinha n. sp.; H. caipiroska n. sp.; H. lelibi n. sp.; H. mojito n. sp.
Remarks: Until this study, Hamatipeda included four species3. The first to be described, Hamatipeda longa, was collected off the Falkland Islands and placed in the genus Typhlotanais (Kudinova-Pasternak, 1975). Later, Błażewicz-Paszkowycz (2007) described Hamatipeda trapezoida from Drake Passage and assigned it to a new genus, Hamatipeda. This genus was supplemented with two other species, Hamatipeda sima by Błażewicz-Paszkowycz and Bamber (2012) recovered in SE Australia (Eastern Bass Strait and Flinders Island), and Hamatipeda prolata from the SE Brazilian coast (Segadilha et al., 2019). Recently, Kakui and Hiruta (2021) has described another species — H. kohtsukai, which was the first record of the genus in the Northern Hemisphere. Because it lacked a trifurcate unguis in the pereopods 4–6 (a key-character for the genus), the definition of the Hamatipeda was extended (Kakui and Hiruta, 2021).
During our examination of the SW Atlantic and SE Australia specimens, we observed a high variety of morphological features (character of the antennule, antennule ornamentation, and character of their pereopods) and decided to extract H. sima from Hamatipeda. It is deposited it in a new genus Yarutanais n. gen. (see below). In addition, Rakaduta n. gen. is established to accommodate a new hamatipedid species from SE Australia.
Key to identification of Hamatipedidae genera (neuter).
1. Antennule article-3 short (1.0x L:W); antennule article-1 with ornamentation; cheliped carpal shield present… Yarutanais n. gen.
- Antennule article-3 long (>3.0x L:W); antennule article-1 ornamentation absent or with weak microtrichia; cheliped carpal shield absent………………………………………………………………………….. 2
2. Antenna articles 2–3 with short and weak setae; pereopod 2–3 propodus with distoventral seta……………………….. Hamatipeda
- Antenna articles 2–3 with robust and long setae; pereopod 2–3 propodus with distoventral spine……………… Rakaduta n. gen.
Hamatipeda caipirinha Gellert and Błażewicz n. sp.
LSID urn:lsid:zoobank.org:act:1AE88054-4248-4020-90B4-
(Figures 2, 3)
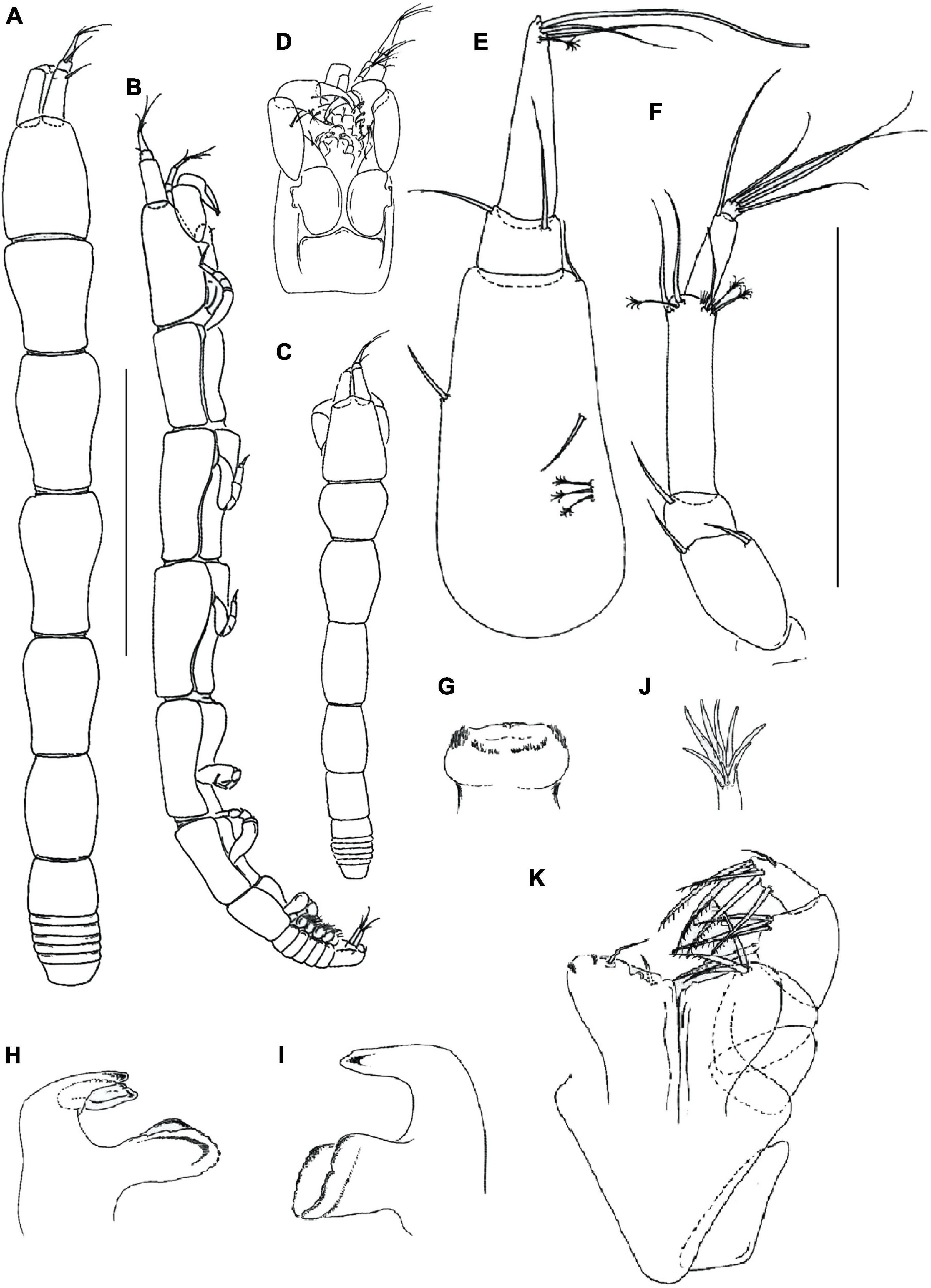
Figure 2. Hamatipeda caipirinha n. sp. (MCZ 48366) (A,B) neuter; (C) manca-3; morphological details: (D) cephalothorax ventral side; (E) antennule; (F) antenna; (G), labrum; (H,I), mandible left and right, respectively; (J), maxillule endite tip; (K), maxilliped and maxilla. Scale: panels (A,B) = 1 mm, panels (E–K) = 0.1 mm.
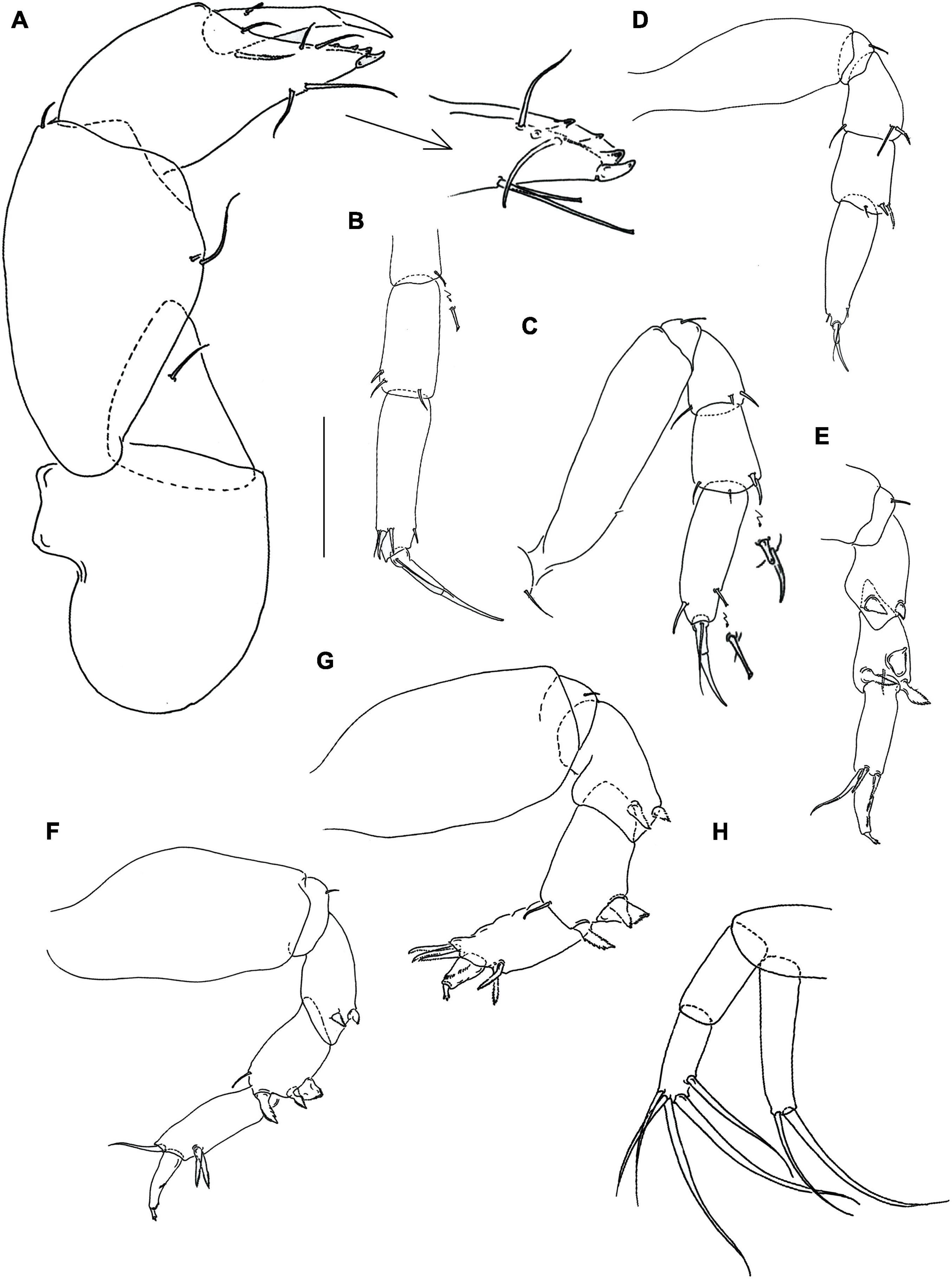
Figure 3. Hamatipeda caipirinha n. sp. (MCZ 48366) (A) cheliped; (B) pereopod-1; (C) pereopod-2; (D) pereopod-3; (E) pereopod-4; (F) pereopod-5; (G) pereopod-6; (H) uropod. Scale: panels (A–H) = 0.1 mm.
Material examined: Holotype: neuter (in tube), MCZ 48366, St. 259A. Paratypes: manca-3 (2.1 mm), MCZ 48350, St. 245A; two neuters (2.6 mm; 2.9 mm—dissected on slide), manca-2 (1.6 mm), one broken (dissected on slide), MCZ 48366, St. 259A.
Diagnosis: Pereonites 1–3 margins narrower posteriorly; pereonites 4–6 margins rounded; pereonites 2–5 proximal margins simple; pereonite-4 long (1.7 L:W). Antennule article-3 long (4.7 L:W). Cheliped carpus 2.2 L:W; fixed finger ventral setae equal length, simple. Pereopod-2 propodus ventrodistal simple seta; pereopod-2 merus with ventrodistal seta. Pereopods 4–6 ischium with seta; carpal molariform spine serrate. Uropod exopod 0.9x endopod; endopod two-articled.
Etymology: The name is given after the popular Brazilian cocktail drink — caipirinha.
Description of neuter with BL 2.9 mm. Paratype. Body (Figures 2A,B), slender 9.8 L:W. Cephalothorax narrow, 1.4 L:W, 1.1x pereonite-1, naked. Pereonites smooth, wider anteriorly, margins gently rounded. Pereonites 1–6: 1.3, 1.7, 1.8, 1.7, 1.5 and 0.9 L:W, respectively. Pereonite-1 0.8x pereonite-2; pereonite-2, 0.9x pereonite-3; pereonite-3, 1.2x pereonite-4; pereonite-4, 1.2x pereonite-5; pereonite-5, 1.9x pereonite-6. Pleon 0.1x total body length; pleonites 1–5 same size, 0.1 L:W. Pleotelson 1.2x pereonite-6.
Antennule (Figure 2E) 0.7x cephalothorax; article-1, 2.0 L:W, 0.6x of TBL, 4.5x article-2, with three penicillate and two simple middle setae, and distal seta; article-2, 1.0 L:W, 0.4x article-3, with two long simple (0.5x article-3) setae distally; article-3, 5.1 L:W, with distal spur, one aesthetasc, one penicillate and two long simple setae distally.
Antenna (Figure 2F) article-1 fused with the cephalothorax; article-2, 1.8 L:W, 2.2x article-3, with two distal setae; article-3, 0.9 L:W, 0.3x article-4, with distal seta; article-4, 4.6 L:W, 2.4x article-5, with three penicillate and three simple distal setae; article-5, 3.1 L:W, 5.0x article-6, with long distal simple seta; article-6 minute, with four simple distal setae.
Mouthparts. Labrum (Figure 2G) rounded and distally setose. Left mandible (Figure 2H) left incisor distally narrow and smooth; lacinia mobilis distally with six rounded projections; molar wide margin irregularly rugose. Right mandible (Figure 2I) incisor distally oblique; molar like in left mandible. Maxillule (Figure 2J) with nine distal spines (one spine located centrally). Labium and epignath lost during the dissection.
Maxilliped (Figure 2K) palp article-1, 1.5 L:W, naked; article-2, 1.3 L:W, with three serrated inner setae (outer seta not seen); article-3, 1.2 L:W, with four serrated inner setae; article-4, 2.6 L:W, with five serrated inner and one outer setae. Basis 0.9 L:W, naked; each endite distal margin almost simple, with two middle setae, 2 minute gustatory cusps, and lateral corners finely setose. Maxilla simple semi-triangular.
Cheliped (Figure 3A) basis separated from pereonite-1, 1.4 L:W (Figure 2D), naked; merus triangular with seta; carpus 1.6 L:W, with two ventral setae and one dorsodistal seta (dorsoproximal seta not seen); chela distally narrower, 2.0 L:W, 0.9x carpus; palm 1.7x fixed finger, with seta on inner side and seta near dactylus insertion; fixed finger with two ventral rod setae (unequal length); cutting edge with three setae and three strong, blunt small teeth, distal tooth relatively small; dactylus with dorsoproximal seta; unguis slender.
Pereopod-1 (Figure 3B) slender; basis and merus broken merus with ventrodistal rod seta; carpus 2.3 L:W, 0.7x propodus, with three short (one fine and two robust) distal setae; propodus 3.8 L:W, with one ventrodistal and three dorsodistal setae; dactylus 5.8 L:W, with seta subequal unguis; dactylus and unguis together 0.8x propodus.
Pereopod-2 (Figure 3C) robust, overall 10 L:W; basis 4.0 L:W, with short dorsodistal seta; ischium with ventral seta; merus 1.6 L:W, 0.9x carpus, with one dorsodistal seta and two ventrodistal setae; carpus 1.6 L:W, 0.7x propodus, with three simple setae and small spine distally; propodus 2.9 L:W, with rod ventrodistal seta and simple dorsodistal seta; dactylus 4.3 L:W, with seta, 0.7x unguis; dactylus and unguis together 0.7x propodus.
Pereopod-3 (Figure 3D) robust, overall 6.0 L:W; basis naked, 2.8 L:W; ischium with ventral seta; merus 1.5 L:W, same length as carpus, with one serrated and two simple setae distally; carpus 1.6 L:W, 0.7x propodus, with spine, one serrated seta, and one simple seta distally; propodus 3.2 L:W, with rod ventrodistal seta and simple dorsodistal seta; dactylus 4.5 L:W, with seta, 0.8x unguis; dactylus and unguis together 0.8x propodus.
Pereopod-4 (Figure 3E) clinging type; basis broken; ischium with ventral seta; merus 2.0 L:W, the same length as carpus, with two serrated ventrodistal spines; carpus 2.4 L:W, 1.1x propodus, with two serrated, slender (distal and ventrodistal) and one serrated molariform hooks, and simple dorsodistal seta; propodus 2.0 L:W, with two serrated ventrodistal spines and simple dorsodistal seta (longer than dactylus); dactylus 3.3 L:W, 3.3x unguis; dactylus and unguis together 0.8x propodus; unguis trifurcate.
Pereopod-5 (Figure 3F) overall 6.0 L:W, as pereonite-4; propodus 2.8 L:W, with two serrated ventrodistal spines and simple dorsodistal seta; dactylus 4.7 L:W, 4.6x unguis; dactylus and unguis together 0.6x propodus; unguis trifurcate.
Pereopod-6 (Figure 3G) overall 4.0 L:W, as pereopod-4; propodus 1.9 L:W, with three dorsodistal setae; dactylus 2.5 L:W, 3.0x unguis; dactylus and unguis together 0.5x propodus; unguis trifurcate.
Pleopods 1–5 small (vestigial) with few short, weak setae, destroyed during dissection.
Uropod (Figure 3H) basal article 1.3 L:W. Endopod two-articled, article-1 naked 2.3 L:W, 1.1x article-2; article-2, 2.8 L:W, with five long distal (two robust) setae. Exopod one-articled, 5.3 L:W, with one robust and one simple distal setae, almost equal to endopod.
Description of manca-2 with BL 1.7 mm; generally similar to neuter. Body (Figure 2C), slender 7.0 L:W. Cephalothorax narrow, 1.4 L:W, 1.5x pereonite-1, naked. Pereonites smooth, wider anteriorly, margins gently rounded. Pereonites 1–6: 0.9, 1.4, 1.7, 1.8, 1.2 and 0.6 L:W, respectively. Pereonite-1 0.6x pereonite-2; pereonite-2, 1.1x pereonite-3; pereonite-3, 1.1x pereonite-4; pereonite-4, 1.5x pereonite-5; pereonite-5, 2.0x pereonite-6. Pleon 0.1x total body length; pleonites 1–5 same size, 0.1 L:W. Pleotelson 1.7x pereonite-6. Pereopods 1–6 similar to neuter, but pereopod-6 absent.
Distribution: Argentine Basin, Atlantic, at depths of 2,707–3,317 m (Figure 14).
Remarks: Hamatipeda caipirinha n. sp., from the SW Atlantic, can be distinguished from other members of the family by the uropod endopod slightly shorter than exopod. Similar equal-length uropod rami are present in H. trapezoida from the Antarctic (Błażewicz-Paszkowycz, 2007), but H. caipirinha has a two-articled endopod (one-articled in H. trapezoida) and ischial seta on pereopods 4–6 (two in H. trapezoida).
H. caipirinha has two rod ventral setae on the cheliped fixed finger (of unequal length) and the ventrodistal part of the pereopod-2 propodus, which are simple in the other members of Hamatipeda (Supplementary Table 1).
Hamatipeda caipiroska Gellert and Błażewicz n. sp.
LSID urn:lsid:zoobank.org:act:85BCDA06-A8DB-48F2-A34A-
(Figures 4, 5)
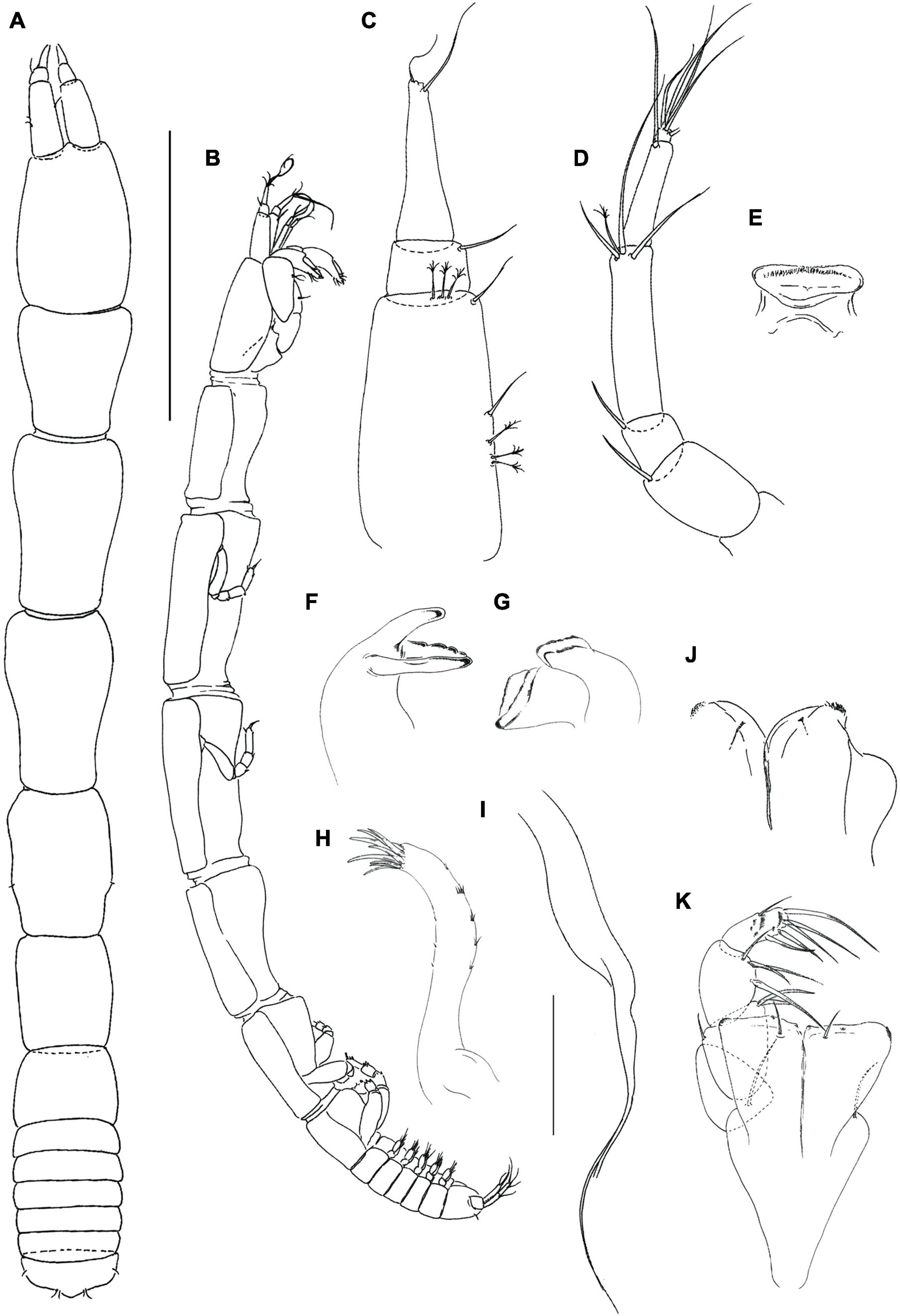
Figure 4. Hamatipeda caipiroska n. sp. (MCZ 48350) (A,B) neuter; (C) antennule; (D) antenna; (E) labrum; (F,G) mandible left and right, respectively; (H,I) maxillule endite and palp; (J), labium; (K) maxilliped. Scale: panels (A,B) = 1 mm, panels (C–J) = 0.1 mm.
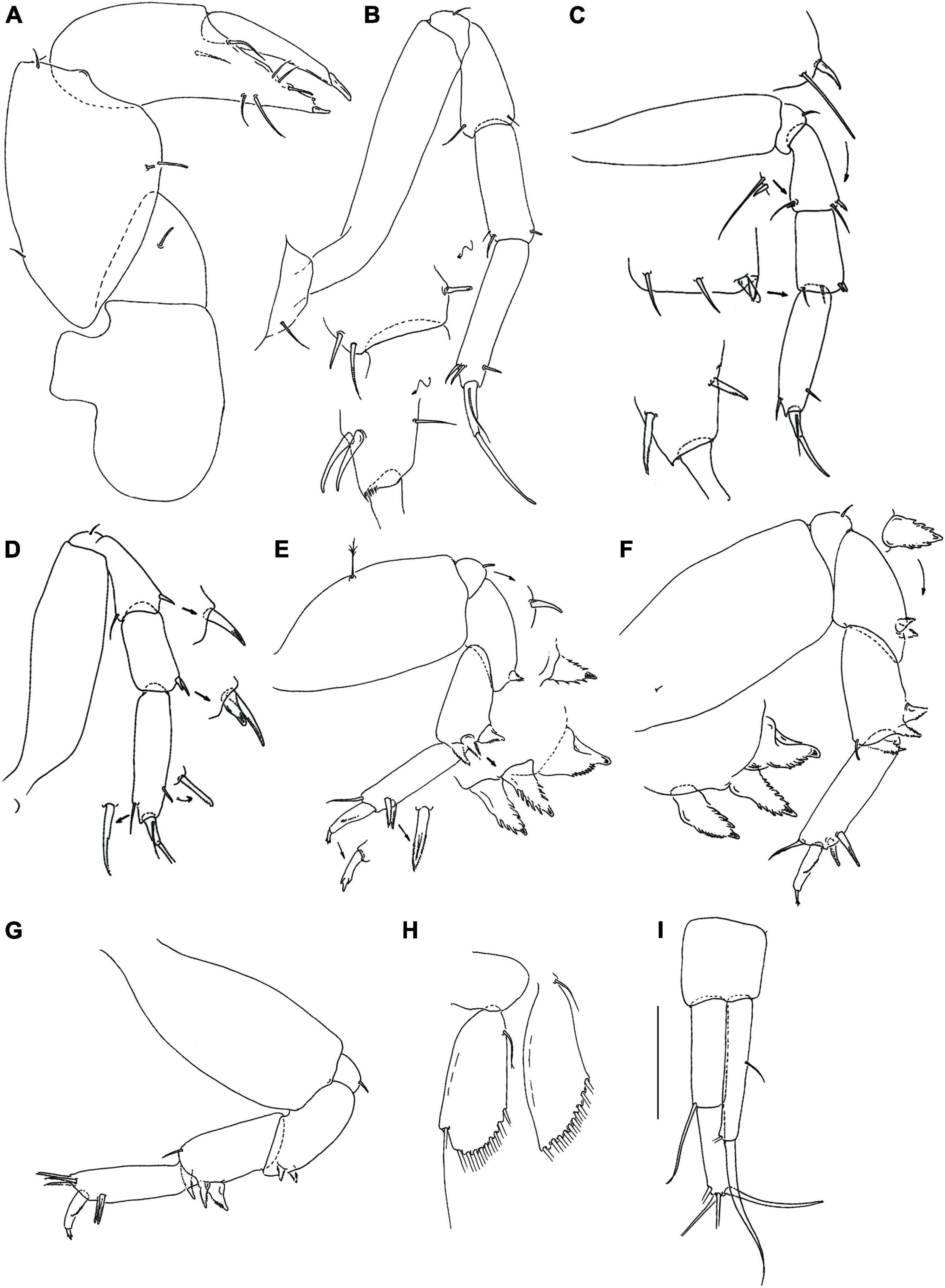
Figure 5. Hamatipeda caipiroska n. sp. (MCZ 48350) (A) cheliped; (B) pereopod-1; (C) pereopod-2; (D) pereopod-3; (E) pereopod-4; (F) pereopod-5; (G) pereopod-6; (H) pleopod; (I) uropod. Scale: panels (A–I) = 0.1 mm.
Material examined: Holotype: neuter (3.7 mm), MCZ 48369, St. 259A. Paratypes: neuter (4.1 mm, dissected on slide), two mancae-3 (2.1 mm, 2.5 mm), manca-2 (1.6 mm), MCZ 48350, St. 245A; manca-3 (2.0 mm), six mancae-2 (1.3–1.8 mm, 1.6 mm — dissected on slide), MCZ 48366, St. 259A; two mancae-3 (1.8, 2.0 mm), manca-2 (1.8 mm), MCZ 48369, St. 259A.
Diagnosis: Pereonites 1–3 margins narrower posteriorly; pereonites 4–6 margins rectangular; pereonites 2–5 proximal margins simple; pereonite-4 short (1.4 L:W). Antennule article-3 long (5.1 L:W). Cheliped carpus 1.6 L:W; fixed finger ventral setae equal length, simple. Pereopod-2 merus with ventrodistal seta; propodus with simple ventrodistal seta. Pereopods 4–6 ischium with seta; carpal molariform spine serrate. Uropod exopod 0.7x endopod; endopod two-articled.
Etymology: The name is given after the well-known Brazilian cocktail drink—caipiroska.
Description of neuter with BL 4.1 mm. Paratype. Body (Figures 4A,B), slender 10.3 L:W. Cephalothorax narrow, 1.5 L:W, 1.3x pereonite-1, naked. Pereonites smooth, wider anteriorly, margins gently rounded. Pereonites 1–6: 1.1, 1.7, 1.8, 1.5, 1.2 and 0.7 L:W, respectively. Pereonite-1 0.7x pereonite-2; pereonite-2 same length as pereonite-3; pereonite-3, 1.3x pereonite-4; pereonite-4, 1.3x pereonite-5; pereonite-5, 1.6x pereonite-6. Pleon 0.1x total body length; pleonites 1–5 same size, 0.3 L:W. Pleotelson 2.5x pereonite-6.
Antennule (Figure 4C) 0.6x cephalothorax; article-1, 2.1 L:W, 0.6 × of TBL, 4.1x article-2, with one simple and three penicillate middle setae, and one simple and three penicillate distal setae; article-2, 0.9 L:W, 0.4x article-3, with moderate length distal seta; article-3, 4.7 L:W, with distal spur and two simple distal setae.
Antenna (Figure 4D) article-1 fused with the cephalothorax; article-2, 1.5 L:W, 1.7x article-3, with distal seta; article-3, 1.3 L:W, 0.3x article-4, with distal seta; article-4, 4.2 L:W, 1.7x article-5, with one penicillate and three simple distal setae; article-5, 4.6 L:W, 8.0x article-6, with long seta; article-6 minute, with five (one broken) distal setae.
Mouthparts. Labrum (Figure 4E) rounded and distally setose. Left mandible (Figure 4F) incisor distally narrow and smooth; lacinia mobilis distally with five rounded projections; molar wide with margin irregularly rugose. Right mandible (Figure 4G) incisor distally oblique; molar like in left mandible. Maxillule (Figures 4H,I) with nine distal spines (one spine located centrally); endite with two distal setae. Labium (Figure 4J) with two lobes, inner lobe disto-outer margin finely setose, outer lobe feeble, smooth. Maxilla not seen.
Maxilliped (Figure 4K) palp article-1, 1.5 L:W, naked; article-2, 1.4 L:W, with three serrated inner setae and one outer seta; article-3, 1.5 L:W, with four serrated inner setae; article-4, 2.8 L:W, with five inner serrated and one outer setae. Basis 1.7 L:W, naked; each endite distal margin almost simple, with middle seta, minute gustatory cusp (second gustatory cusp not seen), and lateral corners finely setose. Epignath not seen.
Cheliped (Figure 5A) basis 1.4 L:W, naked; merus triangular with seta; carpus 1.9 L:W, with two ventral setae, dorsodistal and dorsoproximal setae; chela distally narrower, 1.5 L:W, 1.8x carpus; palm 0.8x fixed finger, with seta on inner side and seta near dactylus insertion; fixed finger with two ventral setae (unequal length); cutting edge with three setae and three weak, blunt small spines, distal spine relatively small; dactylus with dorsoproximal seta; unguis slender.
Pereopod-1 (Figure 5B) overall slender (15 L:W); basis 5.6 L:W, naked; ischium with ventral seta; merus 2.2 L:W, with ventrodistal and dorsodistal simple setae; carpus 3.1 L:W, 0.8x propodus, with two dorsodistal setae and one ventrodistal spine; propodus 4.8 L:W, with one ventrodistal and two robust dorsodistal setae; dactylus 7.0 L:W, with seta, 0.5x unguis; dactylus and unguis together 0.9x propodus.
Pereopod-2 (Figure 5C) robust; overall 9.0 L:W; basis 3.4 L:W, naked; ischium with ventral seta; merus 2.8 L:W, 1.0x carpus, with simple seta and small spine in dorsodistal and ventrodistal corners; carpus 2.1 L:W, 0.7x propodus, with two simple setae and two small spines distally; propodus 4.2 L:W, with one ventrodistal and one dorsodistal setae; dactylus 4.0 L:W, with seta, 0.6x unguis; dactylus and unguis together 0.6x propodus.
Pereopod-3 (Figure 5D) robust; basis naked, 3.5 L:W; ischium with ventral seta; merus 2.4 L:W, same length as carpus, with simple dorsodistal seta and ventrodistal spine; carpus 2.0 L:W, 0.6x propodus, with two distal spines; propodus 3.8 L:W, with one rod ventrodistal and one dorsodistal serrated setae; dactylus 4.0 L:W, with seta; unguis broken.
Pereopod-4 (Figure 5E) overall 5.0 L:W; basis 2.0 L:W, with ventral penicillate seta; ischium with ventral seta; merus 2.2 L:W, 0.9x carpus, with two serrated ventrodistal spines; carpus 2.3 L:W, 0.9x propodus, with two serrated, slender (distal and ventrodistal) spines and one serrated molariform spine (Figure 8B); propodus 3.9 L:W, with two serrated ventrodistal spines and one simple dorsodistal seta (shorter than dactylus); dactylus 3.8 L:W, 3.8x unguis; dactylus and unguis together 0.6x propodus; unguis trifurcate.
Pereopod-5 (Figure 5F) overall 5.0 L:W; as pereonite-4, but carpus with dorsodistal seta.
Pereopod-6 (Figure 5G) overall 5.0 L:W; as pereopod-4, but propodus with three serrated dorsodistal spines.
Pleopods 1–5 (Figure 5H) basal article naked. Endopod 2.3 L:W, with one proximal and nine simple distal setae on outer margin. Exopod 3.1 L:W, with one proximal and ten distal serrated setae on outer margin.
Uropod (Figure 5I) basal article 1.1 L:W. Endopod two-articled, article-1, 3.0 L:W, 0.9x article-2, with seta; article-2, 3.4 L:W, with four long distal setae. Exopod one-articled, 6.1 L:W, with one robust and one simple (broken) distal setae.
Distribution: Argentine Basin, Atlantic, at depths of 2,707–3,317 m (Figure 14).
Remarks: Hamatipeda caipiroska n. sp. is an abyssal species of the SW Atlantic. It can be distinguished from other members of the genus by the presence of a relatively stout cheliped carpus (1.6 L:W) that is more slender in other Hamatipeda species (at least 2.0x L:W). Moreover, H. caipiroska has a two-articled uropodal endopod, as in H. longa, H. prolata, H. caipirinha, and H. mojito, but only H. prolata, known from the Brazilian slope, shares with H. caipirinha parallel margins on pereonites 4–6. These two species are also distinguished by the shape of pereonites 1–3, which are rectangular in H. prolata and trapezoidal in H. caipiroska (Supplementary Table 1).
Hamatipeda caipiroska and H. caipirinha were collected by dredging at the same station in the abyssal of Argentina Basin. They can be distinguished by the length of the uropod rami (almost equal in H. caipirinha, and with exopod clearly shorter 0.7x endopod in H. caipiroska), the length of pereonite-4 (long 1.7 L:W) in H. caipirinha, and short (1.4 L:W in H. caipiroska), and the aspect ratio of the cheliped carpus (slender in H. caipirinha (2.2 L:W), and robust (1.6 L:W) in H. caipiroska).
Hamatipeda mojito Gellert and Błażewicz n. sp.
LSID urn:lsid:zoobank.org:act:E377CF1D-556E-4B8D-85E9-
(Figures 6–8)
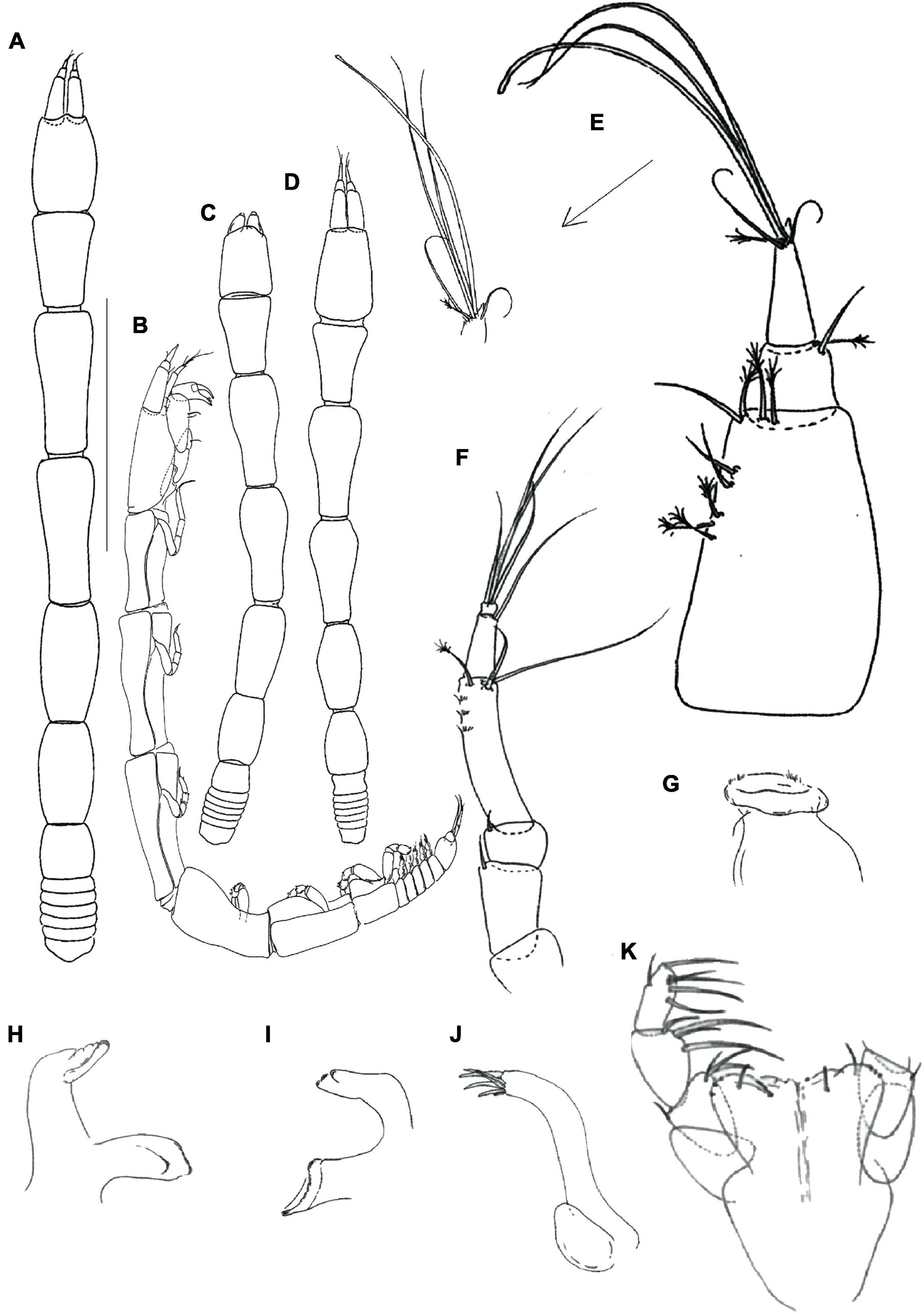
Figure 6. Hamatipeda mojito n. sp. (MCZ 49272) (A,B) neuter; (C,D) manca-2 and 3, respectively; (E) antennule; (F) antenna; (G) labrum; (H,I) mandible left and right, respectively; (J) maxillule endite and maxilla; (K) maxilliped. Scale: panels (A,B) = 1 mm, panels (C–K) = 0.1 mm.
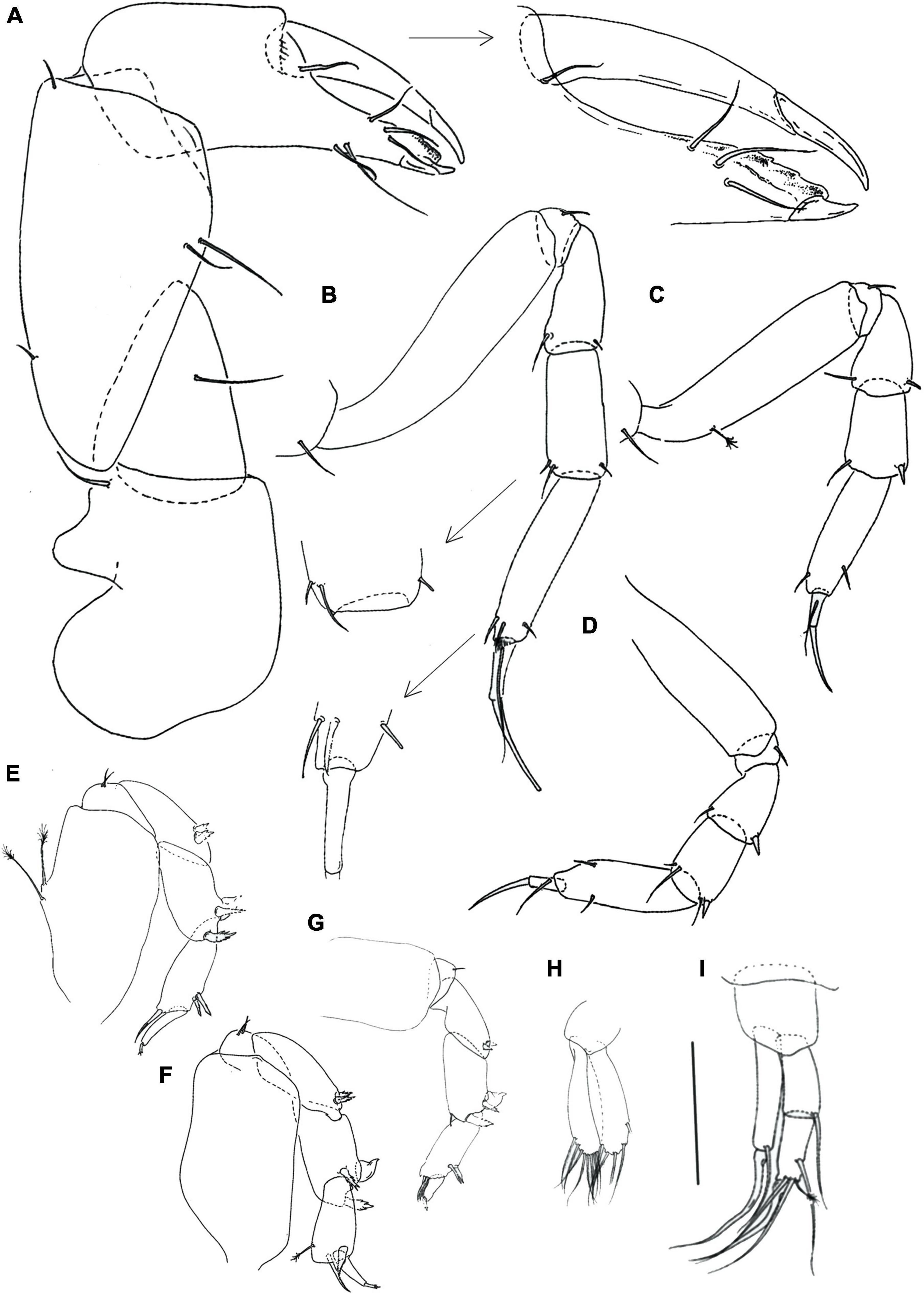
Figure 7. Hamatipeda mojito n. sp. (MCZ 49272) (A) cheliped; (B) pereopod-1; (C) pereopod-2; (D) pereopod-3; (E) pereopod-4; (F) pereopod-5; (G) pereopod-6; (H) pleopod; (I) uropod. Scale: panels (A–I) = 0.1 mm.
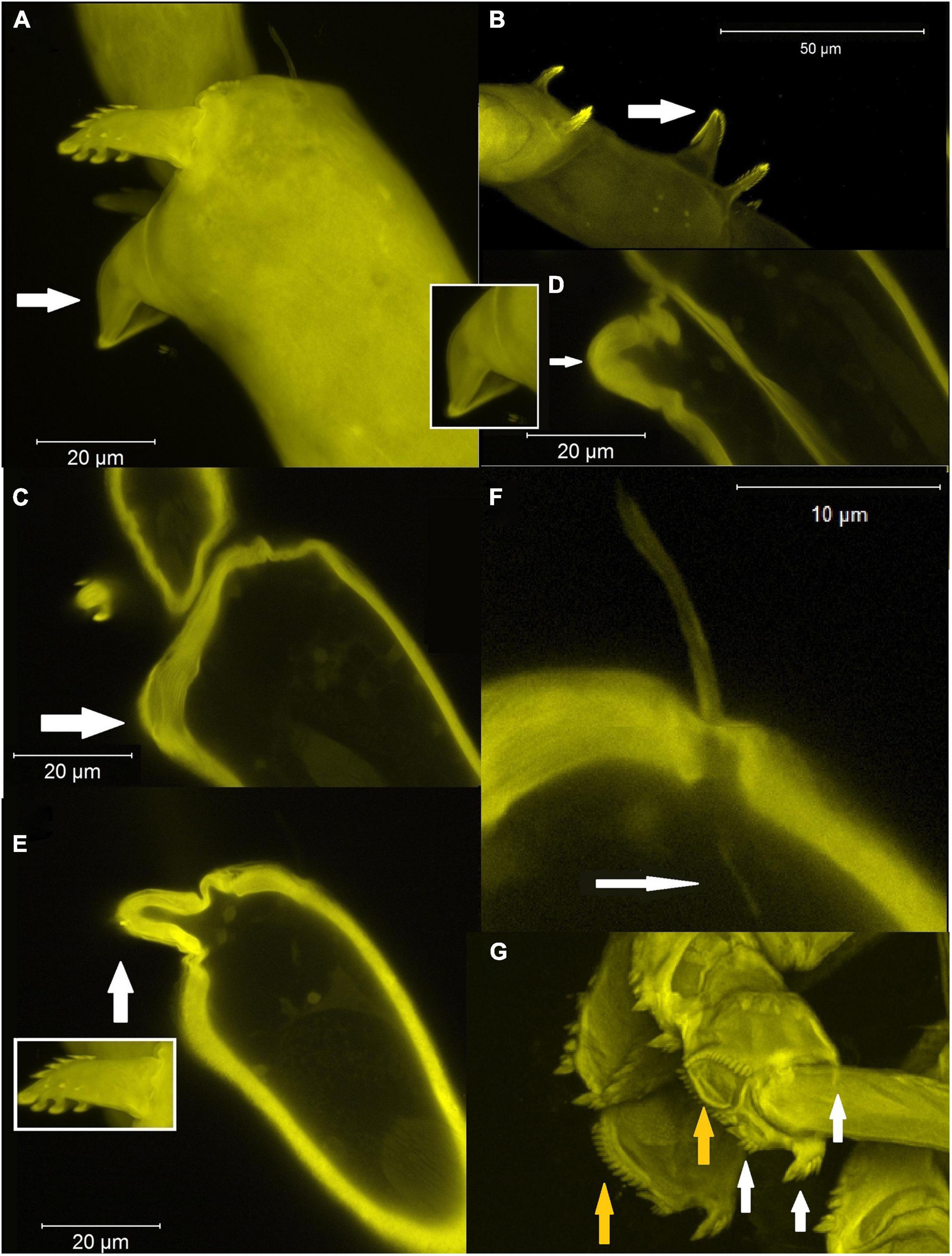
Figure 8. CLSM images of pereopods. Maximum intensity projections [panels (A,B,G) and insets] or single optical cross sections (C–F) are shown. (A–E) Structure and morphology of pereopods 4–6 hooks and setae. (A) Ornamentation of the pereopod-4 carpus in H. mojito n. sp. (arrow indicates a smooth molariform spine); (B) ornamentation of the pereopod-4 carpus in H. caipiroska (arrow indicates a serrated molariform spine); (C–E) Optical cross section through a smooth (C,D) and serrate (E) hooks; (F) optical cross section through seta (arrow indicates innervation); (G) prickly tubercles in Torquella sp.—white arrows indicate hooks (dorsodistal seta obscured), orange arrows the separate prickly tubercles.
Material examined: Holotype: neuter (4.0 mm) (MCZ 49272) St. 293. Paratypes: 35 neuters (2.6–4.56 mm, one broken), neuter (3.44 mm, dissected on slide) 25 mancae-3 (2.1–2.5 mm), 42 mancae-2 (1.6–2.1 mm), six juvenile males (3.9–5.0 mm), 16 individuals broken (two dissected on slide) (MCZ 49272) St. 293; neuter (3 mm), manca-3 (2.1 mm) (MCZ 49288) St. 295.
Diagnosis: Pereonites 1–3 margins narrower posteriorly; pereonites 4–6 margins rounded; pereonites 2–5 proximal margins simple; pereonite-4 very long (2.0 L:W). Antennule article-3 long (4.6 L:W). Cheliped carpus 2.0 L:W; fixed finger ventral setae equal length, simple. Pereopod-2 merus with ventrodistal seta; propodus ventrodistal simple seta. Pereopods 4–6 ischium with two setae; carpal molariform spine smooth. Uropod exopod 0.8x endopod; endopod two-articled.
Etymology: The name is given after the famous Cuban cocktail drink — mojito; as noun in apposition.
Description of neuter with 4.0 mm BL. Paratype. Body (Figures 6A,B), slender 13.6 L:W. Cephalothorax narrow, 1.6 L:W, same length as pereonite-1, naked. Pereonites smooth, wider anteriorly, margins gently rounded. Pereonites 1–6: 1.4, 2.3, 2.5, 1.9, 1.8 and 1.0 L:W, respectively. Pereonite-1 0.6x pereonite-2; pereonite-2 same length as pereonite-3; pereonite-3, 1.2x pereonite-4; pereonite-4, 1.2x pereonite-5; pereonite-5, 1.8x pereonite-6. Pleon 0.1x of TBL; pleonites 1–5: all same size—0.3 L:W. Pleotelson 1.5x pereonite-6.
Antennule (Figure 6E) 0.6x cephalothorax; article-1 2.8 L:W, 1.9x of TBL, 5.8x article-2, with two simple and three penicillate middle setae and three penicillate setae distally; article-2 0.9 L:W, 0.3x article-3, with simple and penicillate distal setae; article-3 5.0 L:W, with distal spur, two short, two long and penicillate setae distally, and one aesthetasc.
Antenna (Figure 6F) article-1 fused with the cephalothorax; article-2 1.9 L:W, 1.9x article-3, with distal seta; article-3 0.9 L:W, 0.3x article-4, with distal seta; article-4 4.0 L:W, 2.4x article-5, with penicillate and two long simple setae and microtrichia distally; article-5 2.5 L:W, 5x article-6, with long simple seta; article-6 minute, with five simple distal setae.
Mouthparts. Labrum (Figure 6G) rounded and distally setose. Left mandible (Figure 6H) incisor distally narrow and smooth; lacinia mobilis molar wide margin distally with projections. Right mandible right (Figure 6I) incisor distally oblique; molar as in left mandible. Maxillule (Figure 6J) with eight distal spines; maxilla subtriangular, naked. Labium not seen.
Maxilliped (Figure 6K) palp article-1, 1.4 L:W, naked; article-2, 1.3 L:W, with three serrated inner setae and outer simple seta; article-3, 1.6 L:W, with four serrated inner setae; article-4, 2.7 L:W, with five serrated inner and one outer setae; each endite distal margin almost simple, with two middle setae, 2 minute gustatory cusps, and lateral corners finely setose. Basis 1.2 L:W, naked. Epignath not seen.
Cheliped (Figure 7A) basis 1.3 L:W, naked; merus triangular with seta; carpus 1.4 L:W, with two ventral setae, dorsodistal and dorsoproximal seta; chela distally narrower, 1.1 L:W, 1.9x carpus; palm 2.1x fixed finger, with seta on inner side near dactylus insertion; fixed finger with two ventral setae; cutting edge with three setae and three subtle, blunt small teeth (distal tooth relatively small); dactylus with dorsoproximal seta; unguis slender.
Pereopod-1 (Figure 7B) overall slender (14 L:W); basis 5.0 L:W, naked; ischium with ventral seta; merus 2.9 L:W, with ventrodistal and dorsodistal simple setae; carpus 2.4 L:W, 0.7x propodus, with ventrodistal and two dorsodistal setae; propodus 4.1 L:W, with one ventrodistal and two dorsodistal setae; dactylus 4.7 L:W, with long seta; unguis 12.5 L:W; dactylus and unguis together 0.9x propodus.
Pereopod-2 (Figure 7C) robust; overall 10 L:W; basis 4.0 L:W, naked; ischium with ventral seta; merus 2.0 L:W, 1.0x carpus, with dorsodistal seta and ventrodistal spine; carpus 1.8 L:W, 0.7x propodus, with two simple setae and two small spines distally; propodus 3.0 L:W, with one ventrodistal and one dorsodistal setae; dactylus 3.3 L:W, with seta; unguis 18.0 L:W; dactylus and unguis together 0.8x propodus.
Pereopod-3 (Figure 7D) robust; overall 9.7 L:W; basis naked, 3.7 L:W; ischium with ventral seta; merus 1.5 L:W, 0.9x carpus, with simple distal seta and spine; carpus 1.6 L:W, 0.6x propodus, with two dorsodistal setae and ventrodistal spine and seta; propodus 3.0 L:W, with one ventrodistal and one dorsodistal setae; dactylus 3.0 L:W, with seta; unguis 8.5 L:W; dactylus and unguis together 0.7x propodus.
Pereopod-4 (Figures 7E, 8A) overall 5.2 L:W; basis 2.0 L:W, with two penicillate setae; ischium with two ventral setae; merus 2.2 L:W, 1.1x carpus, with two serrated ventrodistal spines; carpus (Figure 8A) 2.0 L:W, 0.9x propodus, with two serrated, slender (distal and ventrodistal) spines and one smooth molariform spine; propodus 3.5 L:W, with two serrated ventrodistal spines and one simple dorsodistal seta (shorter than dactylus); dactylus 7.0 L:W, 3.5x unguis; dactylus and unguis together 0.6x propodus; unguis trifurcate.
Pereopod-5 (Figure 7F) overall 5.0 L:W, as pereopod-4, but basis naked; propodus 2.3 L:W, with penicillate dorsal seta; dactylus 3.7 L:W, 2.8x unguis; dactylus and unguis together 0.7x propodus.
Pereopod-6 (Figure 7G) as pereopod-4, but basis naked; propodus 2.8 L:W, with three dorsodistal setae; dactylus 2.7 L:W, 2.0x unguis; dactylus and unguis together 0.5x propodus.
Pleopods 1–5 (Figure 7H) basal article naked. Endopod 3.5 L:W, with eight simple distal setae on outer margin. Exopod 3.4 L:W, with one proximal and four distal plumose setae on outer margin.
Uropod (Figure 7I) basal article 1.1 L:W. Endopod two-articled, article-1, 2.2 L:W, 1.2x article-2, with seta; article-2, 2.3 L:W, with subdistal seta penicillate seta and three long setae. Exopod one-articled, 5.7 L:W, with one robust and one simple distal setae.
Description of manca-3, BL 3.5 mm. Body similar to neuter (Figure 6C), slender 11.4 L:W. Cephalothorax narrow, 1.3 L:W, 0.9x pereonite-1, naked. Pereonites smooth, wider anteriorly, margins gently rounded. Pereonites 1–6: 1.4, 2.1, 2.5, 2.0, 1.5 and 0.8 L:W, respectively. Pereonite-1 1.1x pereonite-2; pereonite-2 same length as pereonite-3; pereonite-3, 1.3x pereonite-4; pereonite-4, 1.3x pereonite-5; pereonite-5, 2.3x pereonite-6. Pleon 0.1x of TBL; pleonites 1–5: all same size—0.2 L:W. Pleotelson 1.8x pereonite-6. Pereopods like neuter, but pereopod-6 undeveloped.
Description of manca-2, BL 3.3 mm. Body similar to neuter (Figure 6D), slender 11.1 L:W. Cephalothorax narrow, 1.6 L:W, 1.1x pereonite-1, naked. Pereonites smooth, wider anteriorly, margins gently rounded. Pereonites 1–6: 1.4, 2.0, 2.3, 1.8, 1.3 and 0.8 L:W, respectively. Pereonite-1 0.7x pereonite-2; pereonite-2 same length as pereonite-3; pereonite-3, 1.3x pereonite-4; pereonite-4, 1.4x pereonite-5; pereonite-5, 2.7x pereonite-6. Pleon 0.1x of TBL; pleonites 1–5: all same size—0.2 L:W. Pleotelson 0.8x pereonite-6. Pereopods similar to neuter, but pereopod-6 absent.
Distribution: Guiana Basin, at depths of 1,000–1,518 m (Figure 14).
Remarks: Hamatipeda mojito n. sp. is the only member of the genus that has a smooth carpal molariform spine on pereopods 4–6 (Supplementary Table 3 and Figure 8A). The other species that has possibly similar smooth molariform spine is H. longa, known from the Falkland Islands, although preservation of the holotype of H. longa and its long-term storage in formalin did not allow for observation of the minute ornamentation. Nevertheless, H. longa has a much shorter uropod exopod (0.5x endopod), that is clearly longer (0.8x endopod) in H. mojito.
Currently, four species of Hamatipeda are known from the coast of South America. Two of them are deep-sea species (present below 2,500 m, H. caipirinha and H. caipiroska) and two bathyal taxa, H. prolata and H. mojito. H. mojito has pereopod 4–6 carpal molariform spines smooth, distinguishing it from other species found in the same area, which have serrate molariform spines (H. caipirinha, H. caipiroska and H. prolata).
Hamatipeda lelibi Gellert and Błażewicz n. sp.
LSID urn:lsid:zoobank.org:act:E4DE8BE7-9C1E-470D-AAA1 -6FA7FD3F8A9F
(Figures 9, 10)
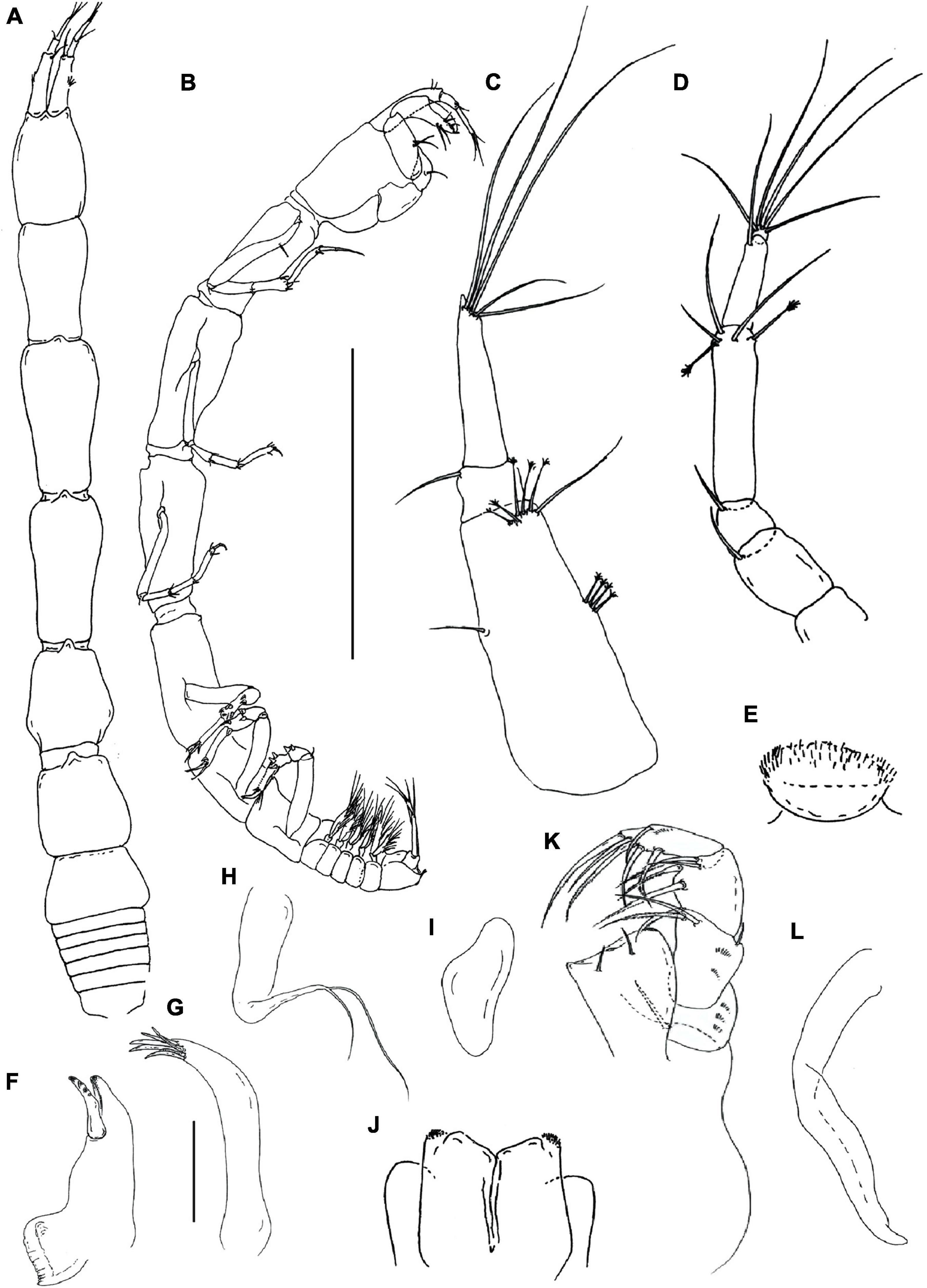
Figure 9. Hamatipeda lelibi n. sp. (J57937) (A,B) neuter; (C) antennule; (D) antenna; (E) labrum; (F) mandible left; (G,H) maxilla endite and palp; (I) maxilla; (J) labium; (K) maxilliped; (L) epignath. Scale: panels (A,B) = 1 mm, panels (C–L) = 0.1 mm.
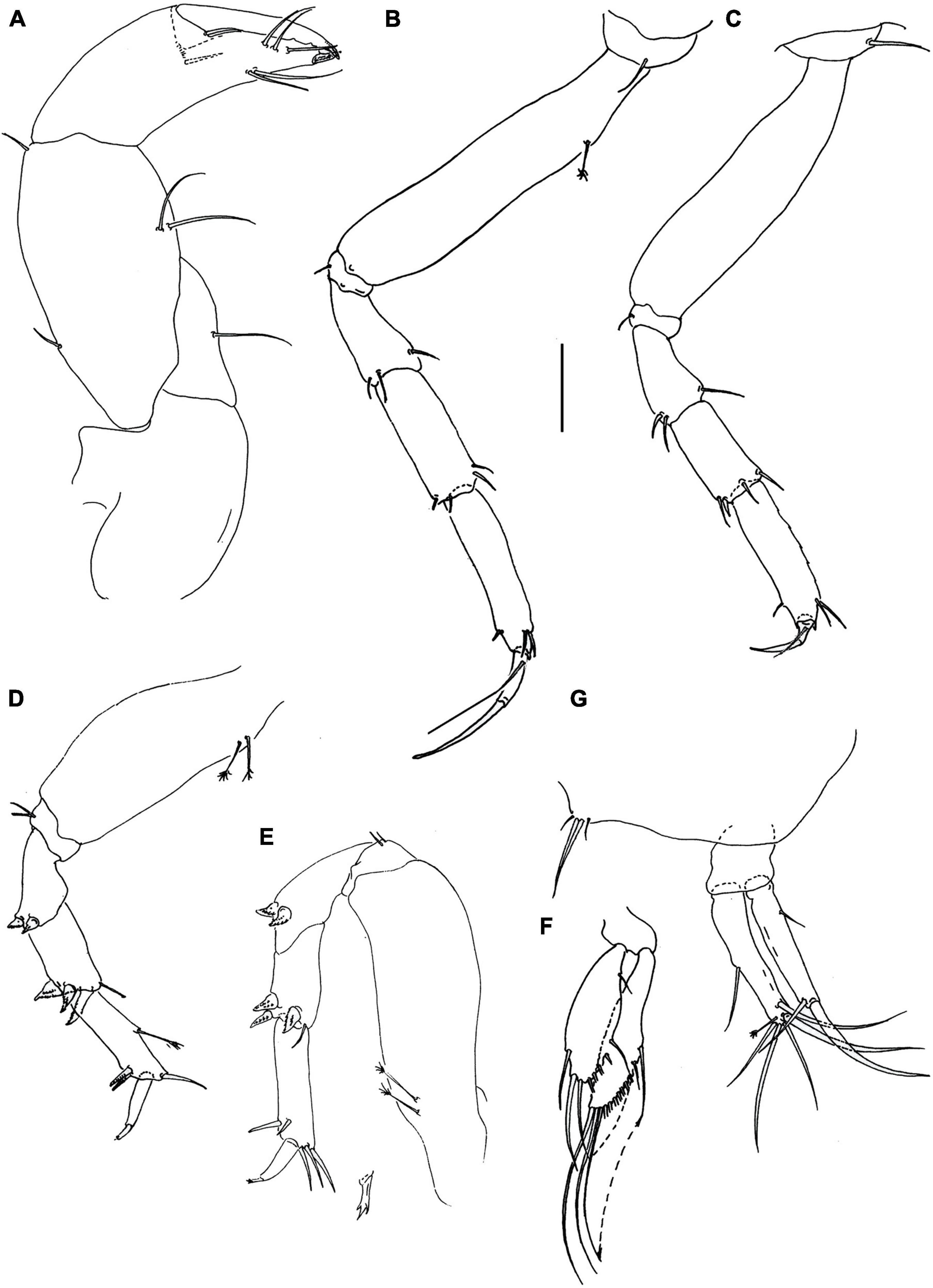
Figure 10. Hamatipeda lelibi n. sp. (J57937) (A) cheliped; (B) pereopod-1; (C) pereopod-2; (D) pereopod-4; (E) pereopod-6; (F) pleopod; (G) uropod. Scale: panels (A–G) = 0.1 mm.
Material examined: Holotype: neuter (6.0 mm) (NMV J37857), SLOPE 54. Paratypes: neuter (5.0 mm) (NMV J59671) SLOPE 67; neuter (6.5 mm) (NMV J57828), SLOPE 25; neuter, dissected on slides (NMV J57937), SLOPE 25.
Diagnosis: Pereonites 1–3 margins narrower posteriorly; pereonites 4–6 margins narrower anteriorly; pereonites 2–5 proximal margins with small process; pereonite-4 very short (1.1 L:W). Antennule article-3 long (5.7 L:W). Cheliped carpus 2.2 L:W; fixed finger ventral setae equal length, simple. Pereopod-2 propodus ventrodistal simple seta; pereopod-2 merus with ventrodistal seta. Pereopods 4–6 ischium with two setae; carpal molariform spine serrate. Uropod exopod 0.8 as endopod; endopod one-articled.
Etymology: The name is given after the Australian drink: Lemon, Lime and Bitters (LLB) which is a combination of clear lemonade, lime cordial, and bitters.
Description of neuter with 2.8 mm BL. Paratype. Body (Figures 9A,B), slender 12.9 L:W. Cephalothorax narrow, 1.4 L:W, same length as pereonite-1, naked. Pereonites smooth, wider anteriorly, margins gently rounded. Pereonites 1–6: 1.6, 2.0, 1.9, 1.3, 1.1, and 0.6 L:W, respectively. Pereonite-1 0.7x pereonite-2; pereonite-2, 1.1x pereonite-3; pereonite-3, 1.4x pereonite-4; pereonite-4, 1.1x pereonite-5; pereonite-5, 1.6x pereonite-6. Pleon 0.1x of TBL; pleonites 1–5: all same size—0.2 L:W. Pleotelson 1.7x pereonite-6, with two longer and two short terminal setae.
Antennule (Figure 9C) 0.9x cephalothorax; article-1 2.7 L:W, 0.6x of TBL, 5.8x article-2, with one simple and four penicillate middle setae and one long and five penicillate setae distally; article-2 0.9 L:W, 0.3x article-3, with simple distal seta; article-3 5.0 L:W, with distal spur, two short and three long setae distally, aesthetasc not seen.
Antenna (Figure 9D) article-1 fused with the cephalothorax; article-2 1.4 L:W, 1.7x article-3, with long (as long as article-3) distal seta; article-3 as long as wide, 0.3x article-4, with distal seta; article-4 5.3 L:W, 1.9x article-5, with two penicillate and three simple setae distally; article-5 3.7 L:W, 5.5x article-6, with long simple seta; article-6 minute, with five simple distal setae.
Mouthparts. Labrum (Figure 9E) rounded and distally setose. Left mandible (Figure 9F) incisor distally narrow and smooth; lacinia mobilis distally with four rounded projections and molar process wide, distally oblique, with a rugose margin. Right mandible not seen. Maxillule (Figure 9G) with at least seven distal spines; palp with two distal setae (Figure 9H). Maxilla (Figure 9I) subtriangular. Labium (Figure 9J) with two lobes, inner lobe disto-outer margin finely setose, outer lobe feeble, smooth.
Maxilliped (Figure 9K) palp article-1, 1.4 L:W, with numerous microtrichia; article-2, 1.9 L:W, with one outer and three serrated inner setae and numerous microtrichia; article-3, 1.7 L:W, with four inner setae; article-4, 3.2 L:W, with five inner and one outer setae. Basis elongate (damaged during dissection), with distal seta reaching half of the endite; each endite distal margin almost simple, with middle seta, two minute gustatory cusps, and lateral corners finely setose. Epignath (Figure 9L) distally narrower, tip rounded.
Cheliped (Figure 10A) basis 1.5 L:W; merus wedge-shaped, with ventral seta; carpus 1.9 L:W, with two long ventral setae, two distal and subproximal short setae on the dorsal margin; chela slender, 3.0 L:W, 0.9x carpus; palm 1.2x fixed finger, with seta on inner side, and seta near dactylus insertion; fixed finger with two ventral setae (one longer than the other); cutting edge with three setae and three weak, blunt teeth distally; seta of dactylus not observed.
Pereopod-1 (Figure 10B) overall slender (13 L:W); coxa with seta; basis 4.9 L:W, with penicillate seta dorsally; ischium with ventral seta; merus 2.8 L:W, 0.9x carpus, with one dorsodistal and two ventrodistal setae; carpus 2.7 L:W, 0.8x propodus, with four distal setae; propodus 5.3 L:W, with one ventrodistal and two dorsodistal setae; dactylus 7.0 L:W, 0.6x unguis, seta not seen; dactylus and unguis together 0.9x propodus.
Pereopod-2 (Figure 10C) overall slender (12 L:W); basis 3.9 L:W, naked; ischium with ventral seta; merus 2.5 L:W, 0.9x carpus, with two ventrodistal and one dorsodistal setae; carpus 2.2 L:W, 0.6x propodus, with three simple setae and spine distally; propodus 4.3 L:W, with ventrodistal seta and two dorsodistal setae; dactylus 3.0 L:W, 0.6x unguis; dactylus and unguis together 0.5x propodus.
Pereopod-3 as pereopod-2 (not illustrated).
Pereopod-4 (Figure 10D) overall 6.0 L:W; basis 2.5 L:W, with two proximal penicillate setae; ischium with two ventral seta; merus 2.8 L:W, same length as carpus, with two serrated ventrodistal spines; carpus 2.2 L:W, 0.8x propodus, with two serrated slender spines, one serrated molariform spine, and simple dorsodistal seta; propodus 3.5 L:W, with two serrated spines ventrodistally, one middle penicillate seta and one simple dorsodistal seta (shorter than dactylus); dactylus 3.0 L:W, 2.0x unguis; dactylus and unguis together 0.6x propodus; unguis trifurcate.
Pereopod-5 as pereopod-4 (not illustrated).
Pereopod-6 (Figure 10E) overall 7.0 L:W; as pereopod-4 but propodus with three dorsodistal spines.
Pleopods 1–5 (Figure 10F) basal article naked. Endopod 4.8 L:W, with one proximal and ten simple distal setae on outer margin. Exopod 3.3 L:W, with one proximal and seven distal simple setae on outer margin.
Uropod (Figure 10G) basal article 0.9 L:W. Endopod one-articled, article-1, 5.7 L:W, 0.9x exopod 3.4 L:W, with one penicillate and five long distal setae. Exopod one-articled, 5.8 L:W, with one robust seta and one simple distal seta.
Distribution: SE Australia, from New South Wales to Victoria, south of Point Hicks, at depths of 1,119–2,600 m (Figure 14).
Remarks: Hamatipeda lelibi n. sp. from the slope off SE Australia can be distinguished from other Hamatipeda species by the distinctive projections on the anterior margins of the tergites of pereonites 2–5 (Supplementary Table 1). These are a unique character in the Paratanaoidea, although a similar process is present on the anterior margin of pleonite-1 of the paratanaid Pseudobathytanais gibberosus Larsen and Heard, 2001 (p.17, Figure 8A).
Hamatipeda lelibi has unarticulated and subequal uropod rami as does the Antarctic H. trapezoida, but the lateral margins of pereonites 4–6 of that species are evenly rounded, whereas in H. lelibi these are trapezoidal (wider posteriorly).
Key for identification of Hamatipeda (neuters). See also Supplementary Table 3.
1. Pereopod 4–6 unguis bifurcate……….. Hamatipeda kohtsukai
(NW Pacific, Sagami Sea, 488 m)
- Pereopod 4–6 unguis trifurcate…………………………………………. 2
2. Uropod endopod 5.8 L:W…………………………….. H. lelibi n. sp.
(SW Pacific, SE Australian slope, 1,119–2,600 m)
- Uropod endopod < 5.8 L:W……………………………………………… 3
3. Cheliped carpus short (< 2.0 L:W)………. H. caipiroska n. sp.
(SW Atlantic, Argentinian Basin, 2,707–3,317 m)
- Cheliped carpus long (>2.0 L:W)……………………………………… 4
4. Antennule article-3 4.8 L:W……………….. .H. caipirinha n. sp.
(SW Atlantic, Argentinian Basin, 2,707–3,317 m)
- Antennule article-3 < 4.8 L:W………………………………………….. 5
5. Uropod endopod one-articled……………………… H. trapezoida
(SW Atlantic, 2372–3876)
- Uropod two-articled…………………………………………………………. 6
6. Pereopod 2–3 merus with ventrodistal seta………….. H. longa
(SW Atlantic, 720 m)
- Pereopod 2–3 merus with ventrodistal seta and spine……….. 7
7. Pereopod 4–6 molariform spine smooth…… H. mojito n. sp.
(W Atlantic, Guiana Basin, 1,000–1,518 m)
- Pereopod 4–6 molariform spine serrate……………… .H. prolata
(W Atlantic, Brazilian Basin, 77–1,310 m)
Genus Rakaduta Gellert and Błażewicz n. gen.
LSID urn:lsid:zoobank.org:act:03B68010-395E-458B-BA5F-939C86FB5A18
Diagnosis: Pereonite-1 0.5x pereonite-2. Antennule article-3 long (4.0 L:W); article-1 with few microtrichia. Antenna articles 2–3 with spines. Cheliped without carpal shield. Pereopod-1 propodus short (2.3 L:W). Pereopods 2–3 propodus with short ventrodistal spine. Pereopod-3 carpus with few (three) setae. Pereopod-5 propodus seta short. Endopod one-articled.
Type species: Rakaduta inexcessis n. sp. (by monotypy).
Etymology: In the Aboriginal language Walpiri, “rakadu” means “deep,” which reflects deeper distribution of the genus in relation to Yarutanais, which occurs on the continental shelf. The ending “ta” are two first letters from Tanaidacea.
Remarks: The genus is defined by a unique robust and long seta on antenna articles 2–3, where other hamatipedids have a rather short and weak seta. Moreover, Rakaduta n. gen. also has a short ventrodistal spine on the propodus of pereopods 2–3 and rather short unguis and dactylus in pereopods 2–3, where all other hamatipedids have a propodal seta (see Supplementary Table 3).
Rakaduta inexcessis Gellert and Błażewicz n. sp.
LSID urn:lsid:zoobank.org:act:0B845327-E37F-4F95-9C08-7559913E9040
(Figures 11, 12)
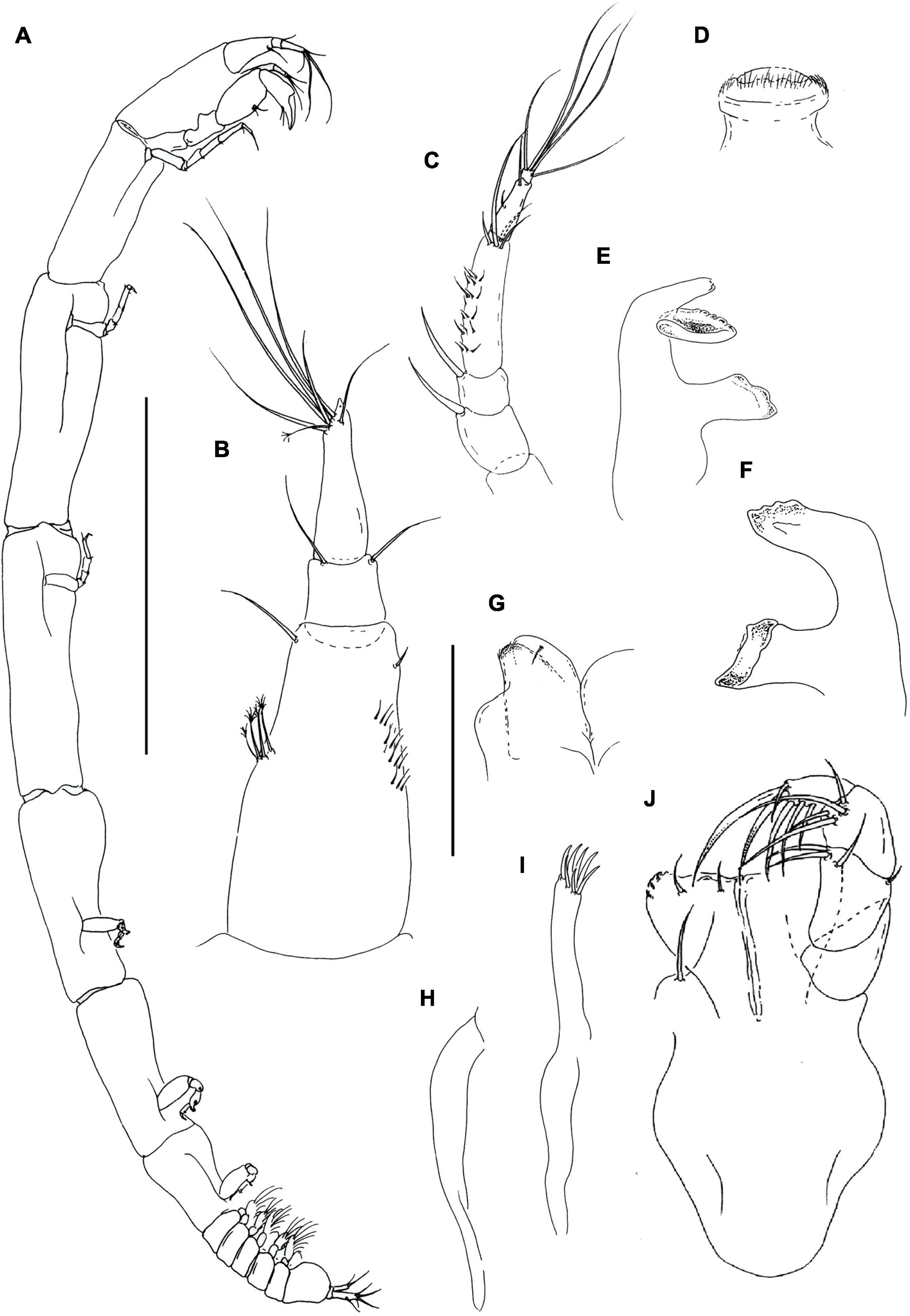
Figure 11. Rakaduta inexcessis n. sp. (J57983) (A) neuter; (B) antennule; (C) antenna; (D) labrum; (E,F) mandible left and right, respectively; (G) labium; (H) epignath; (I) maxillule endite and palp; (J) maxilliped. Scale: panel (A) = 1 mm, panels (B–J) = 0.1 mm.
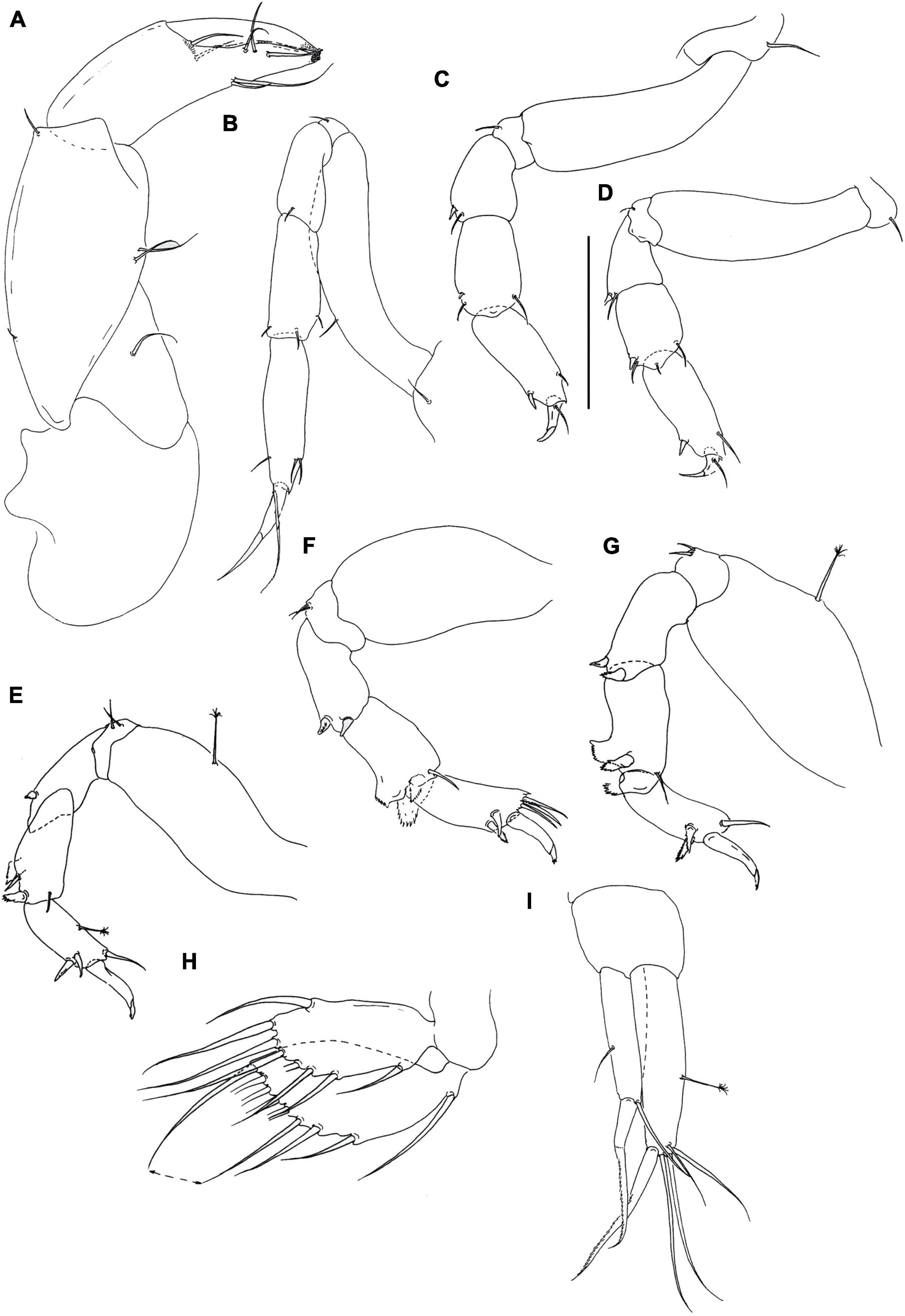
Figure 12. Rakaduta inexcessis n. sp. (J57983) (A) cheliped; (B) pereopod-1; (C) pereopod-2; (D) pereopod-3; (E) pereopod-4; (F) pereopod-5; (G) pereopod-6; (H) pleopod; (I) uropod. Scale: panels (A–I) = 0.1 mm.
Material examined: Holotype: neuter (4.5 mm) NMV J57938, SLOPE 33. Paratypes: four neuters (3.6 mm, 3.7 mm, 4.0 mm and one damaged) NMV J37878, SLOPE 32; neuter (2.7 mm) NMV J62715, SLOPE 2; three post-ovi. females (3.0 mm, 3.4 mm, and 3.8 mm), manca-3, 2.2 mm, J37862, SLOPE 45; neuter (2.5 mm) NMV J62714, SLOPE 40; three neuters (3.5 mm, one in tube) NMV J37866, one dissected on slides NMV J57939, SLOPE 33; neuter (2.5 mm) NVM J57883, SLOPE 40.
Diagnosis: As for genus (monotypy).
Type species: Rakaduta inexcessis n. sp.
Etymology: The name of the species is for the Australian rock band (INXS) formed in Sydney, New South Wales.
Description of neuter with BL 3.9 mm (paratype). Body (Figure 11A) slender, 15.3 L:W. Cephalothorax narrow, 2.4 L:W, 0.8x pereonite-1, naked. Pereonites smooth, wider anteriorly, margins gently rounded. Pereonites 1–6: 2.4, 3.3, 3.3, 2.5, 2.0 and 1.7 L:W, respectively. Pereonite-1, 0.7x pereonite-2; pereonite-2 same length as pereonite-3; pereonite-3, 1.3x pereonite-4; pereonite-4, 1.2x pereonite-5; pereonite-5, 1.5x pereonite-6. Pleon 0.1x of total body length; pleonites 1–5 same size, 0.3 L:W. Pleotelson 1.4x pereonite-6.
Antennule (Figure 11B) 0.6x cephalothorax; article-1 2.3 L:W, 0.6 of antennule length, with four penicillate ventral setae, several fine midlength seta on dorsal margin, and simple distal seta; article-2, 1.3 L:W, 0.6x article-3, with two long distal setae; article-3, 4.0 L:W, with distal spur, one penicillate and six long setae distally.
Antenna (Figure 11C) article-1 fused with the cephalothorax; article-2 1.1 L:W, 1.1x article-3, with dorsodistal long and robust seta (2.0x article-3); article-3, 1.1 L:W, 0.4x article-4, with dorsodistal long and robust setae (0.5x article-4); article-4, 4.0 L:W, 2.0x article-5, with few fine sparsely distributed setae on dorsal margin and four simple distal setae (one longer and three shorter); article-5 4.7 L:W, 7.0x article-6, with distal seta; article-6 minute with distal five setae.
Mouthparts. Labrum (Figure 11D) rounded and distally setose. Left mandible (Figure 11E) incisor distally with two small and blunt processes; lacinia mobilis well developed, distally with five rounded projections; molar process wide, with irregularly undulate margins. Right mandible (Figure 11F) incisor distally truncate, with four blunt processes; molar as in left mandible. Maxillule (Figure 11J) with at least seven distal spines. Maxilla not seen. Labium (Figure 11G) with two lobes, inner lobes finely setose, outer lobe feeble with seta.
Maxilliped (Figure 11I) palp article-1 1.6 L:W, naked; article-2 1.6 L:W, with one outer and three serrated inner setae; article-3 1.5 L:W with four inner setae; article-4, slender 2.0 L:W, with five inner and one outer setae. Basis elongated, 2.6 L:W, with distal seta 0.7x endite; each endite distal margin almost simple, with two middle setae, 2 minute gustatory cusps, and lateral corners finely setose. Epignath (Figure 10H) distally rounded.
Cheliped (Figure 12A) basis naked, 1.5 L:W; merus wedge-shaped, with ventral seta; carpus 2.2 L:W, with two ventral setae and two short dorsal setae (dorsodistal rod and subproximal simple); chela narrow, 2.9 L:W, 0.9x carpus; palm 1.3x fixed finger with seta on inner side and seta near dactylus insertion; fixed finger, with two ventral setae (unequal length); cutting edge weakly calcified, with three setae; dactylus proximal seta not seen.
Pereopod-1 (Figure 12B) relatively slender, overall 13 L:W; coxa with seta; basis 5.4 L:W, with middorsal seta; ischium with ventral seta; merus 2.7 L:W, 0.8x carpus, with distal seta; carpus 2.7 L:W, 0.8x propodus, with three fine distal setae; propodus 4.0 L:W, with one ventrodistal and two dorsodistal setae; dactylus 4.5 L:W, 0.5x unguis; dactylus and unguis together 0.9x propodus.
Pereopod-2 (Figure 12C) robust, overall 10 L:W; coxa with seta; basis naked 3.5 L:W; ischium with ventral seta; merus 1.4 L:W, 0.9x carpus, with simple seta and spine ventrodistally; carpus 1.5 L:W, 0.8x propodus, with two short distal seta and one ventrodistal spine; propodus 2.7 L:W, with ventrodistal spine and two dorsodistal setae; dactylus 2.0 L:W, 0.5x unguis; dactylus and unguis together 0.5x propodus.
Pereopod-3 (Figure 12D) as pereopod-2.
Pereopod-4 (Figure 12E) overall 6.0 L:W; basis 2.7 L:W, with penicillate midventral seta; ischium with two ventral setae; merus 2.7 L:W, 1.1x carpus, with two serrated ventrodistal spines; carpus 3.3 L:W, with two robust serrate and one serrate, molariform spines (semifused with the article); propodus 2.2 L:W, with penicillate middorsal seta; two serrated spines ventrodistally and one seta (0.8x dactylus); dactylus 4.0 L:W, 2.7x unguis; dactylus and unguis together 0.8x propodus; unguis trifurcate.
Pereopod-5 (Figure 12F), as pereopod-4.
Pereopod-6 (Figure 12G) overall 7 L:W; as pereopod-5 but propodus with three setae.
Pleopods 1–5 (Figure 12H) basal article naked. Endopod 2.1 L:W, with one proximal and eight distal setae on outer margin. Exopod 2.0 L:W, with one proximal and six distal setae on outer margin.
Uropod (Figure 12I) basal article 0.9 L:W. Endopod one-articled, 2.9 L:W, with penicillate seta at mid-length, and one robust serrated and four simple setae distally. Exopod one-articled, 3.2 L:W, with one simple midlength seta, and one robust, serrate and one simple distal setae.
Distribution: Australia, from off Nowra, New South Wales to off Freycinet Peninsula, Tasmania, at depths 400–1,000 m (Figure 14).
Remarks: As for the genus because of monotypy.
Genus Yarutanais Gellert and Błażewicz n. gen.
LSID urn:lsid:zoobank.org:act:A3D283EB-6C41-4BFD-8BF3-1FAFCFCFFCA3
(Figure 13)
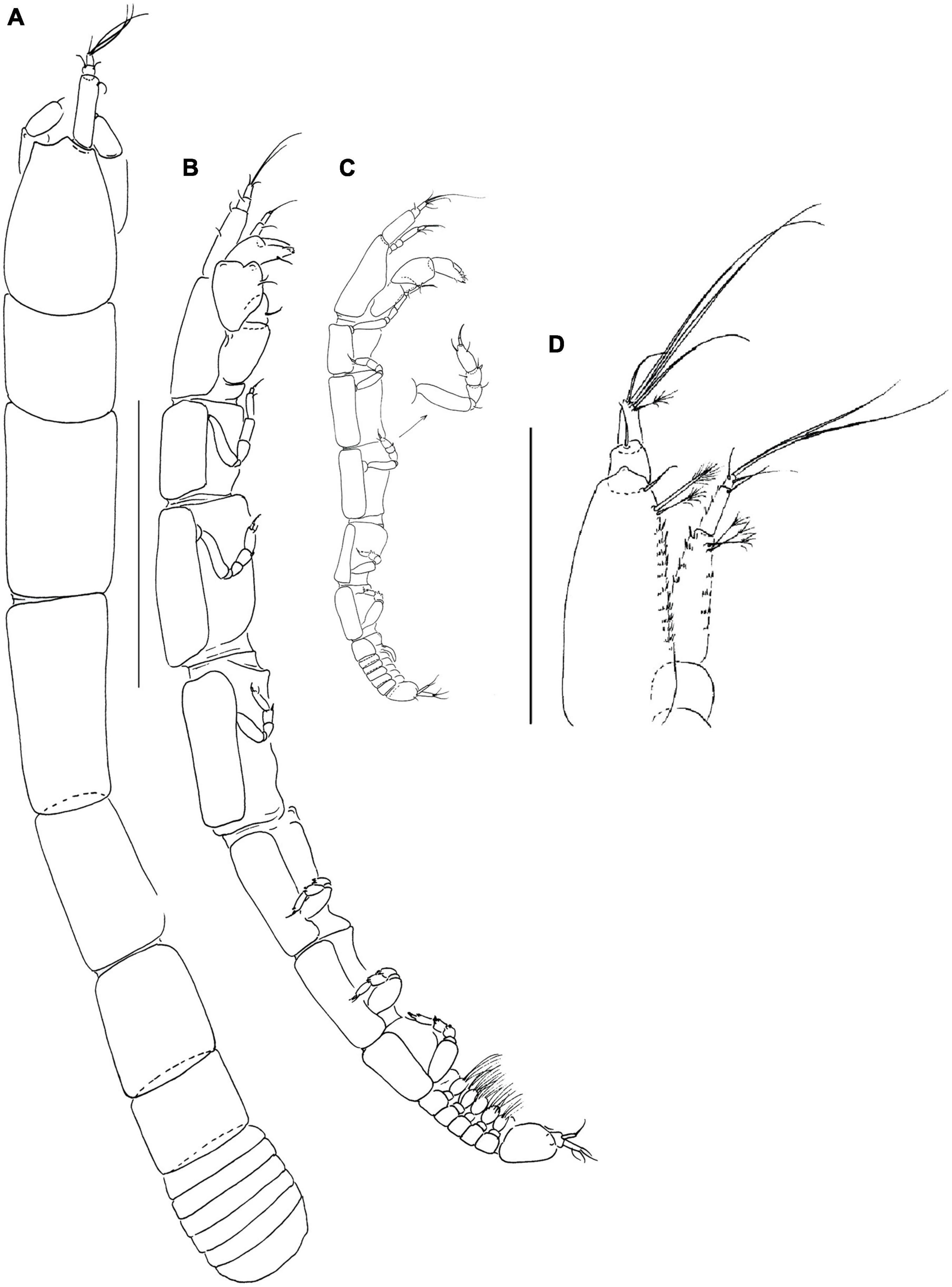
Figure 13. Yarutanais sima (Błażewicz-Paszkowycz and Bamber, 2012) (A,B) neuter (J69711); (C) manca-3; (D) antennule (J58902). Scale: panels (A–C) = 1 mm, panel (D) = 0.1 mm.
Diagnosis: Pereonite-1 little shorter than pereonite-2. Antennule article-3 short (1.0x L:W); article-1 with numerous ventral and ventrolateral microtrichia. Antenna articles 2–3 with setae. Cheliped carpal shield present. Pereopod-1 propodus short (0.8 L:W). Pereopod-3 carpus with several (five) setae. Pereopod-5 propodus seta long. Endopod one-articled.
Type species: Yarutanais sima Błażewicz-Paszkowycz and Bamber (2012) n. comb. (by monotypy).
Synonym: Hamatipeda sima Błażewicz-Paszkowycz and Bamber (2012)
Etymology: In Aboriginal language Walpiri, “yaru” means “shallow,” which reflects its shallow-water distribution.
Distribution: Australia, from off Nowra, New South Wales to Eastern Bass Strait, 50 km NE of Babel Island, Tasmania, at depths 49–1,000 m (Figure 14).
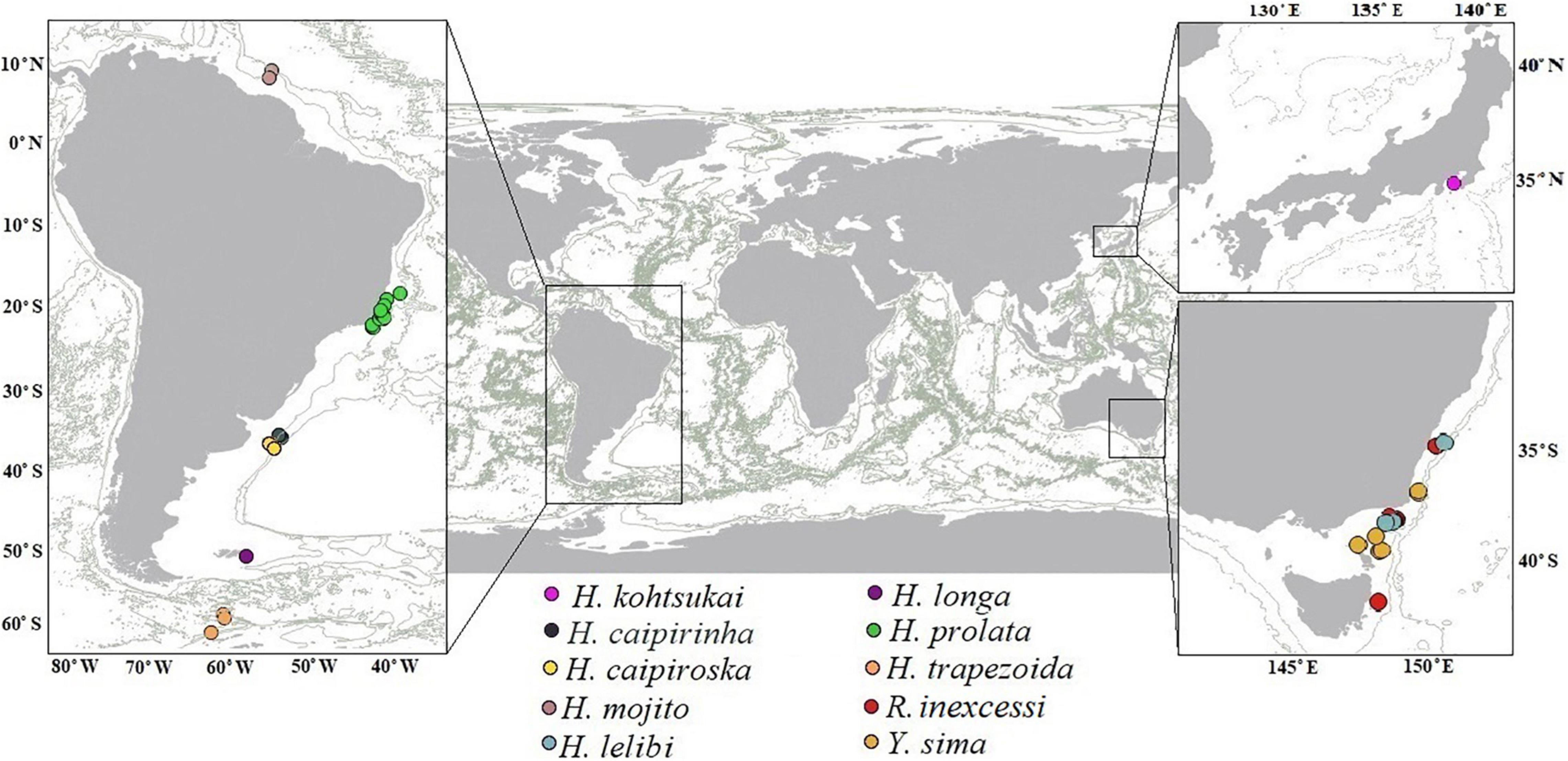
Figure 14. Distribution of Hamatipedidae based on the present study and literature data (Błażewicz-Paszkowycz, 2007; Błażewicz-Paszkowycz and Bamber, 2012; Segadilha et al., 2019; Kakui and Hiruta, 2021).
Remarks: Yarutanais sima is the only Yarutanais species on the shelf and slope off SE Australia. It can be distinguished from other members of the Hamatipedidae by the presence of a cheliped carpal shield and five setae on the carpus pereopods 2–3 (Błażewicz-Paszkowycz and Bamber, 2012: figure 116 A,C–D) and short antennule article-3 (only little longer than article-3; Figure 13E). Moreover, the antennule article-1 has ventral and ventrolateral robust microtrichia that are absent in other hamatipedids (Figure 13D).
Results
Diversity and Distribution
As a result of our study, the Hamatipedidae includes three genera (two new for science) and ten species (five new for science). The most speciose genus is Hamatipeda, with the two other genera Rakaduta and Yarutanais monotypic.
The Hamatipedidae is a wide-spread element of the benthic community in the Southern Hemisphere (Figure 14), and underestimated element of benthic deep-sea communities. In the Atlantic, the most northern record of the family is H. mojito, found off French Guiana. Except for Yarutanais sima that occurs in the shallow Bass Strait, all hamatipedids inhabit greater depths beyond the continental shelf, i.e., on the continental slope (seven species) and only two are known from the abyssal.
Morphometrics
Analysis of the relationships between body size, body segments, and uropod rami measured at different developmental stages of Hamatipeda mojito indicates allometric growth. Positive allometry was calculated for pereonite-6, isometry for pereonites 4–5, and negative allometry for the carapace, pereonites 1–3, and uropod endopod and exopod (Figure 15 and Supplementary Table 2). Moreover, two size stages of neuters were observed.
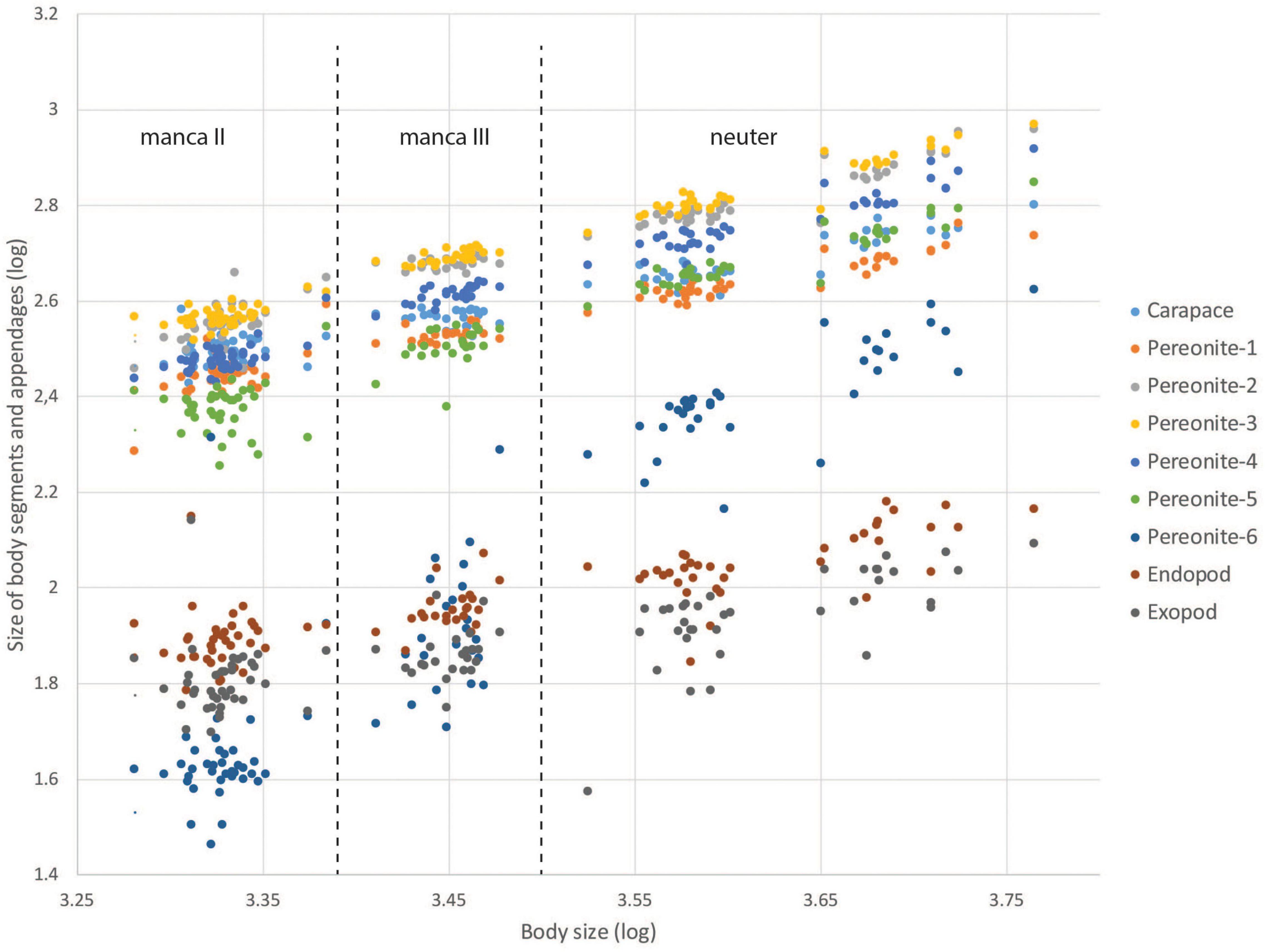
Figure 15. Scatterplot of Spearman correlation of nine morphological characters (body length, pereonites 1–6 length, uropodal exopod and endopod length) of the three developmental stages (manca-2, manca-3, neuter) of Hamatipeda mojito.
Discussion
The proposed new family is the twentieth recent family of the Paratanaoidea and the eleventh that apparently radiated in light-deprived environments such as the deep-sea. The lack of eyes in many paratanaoideans is suggested as evidence of the place of this origin, although in some cases, e.g., the blind Tanaissuidae Bird and Larsen (2009), Bird (2002, 2012) the lack of eyes is considered an adaptation for tubicolous life-style.
The earliest paratanaoidean families to be recognized were relatively straightforward for taxonomic classification. The presence of the multi-articled uropods (Leptocheliidae Lang, 1973), lateral plumose seta on pleonites 1–4 (Paratanaidae Lang, 1949; Teleotanaidae Bamber, 2008), cheliped attached directly/posteriorly to the cephalothorax (Agathotanaidae Lang, 1971 and Anarthruridae Lang, 1971) or one pair of the oostegites (Pseudotanaidae Sieg, 1976), were sufficiently diagnostic for these families. Later classification of the Paratanaoidea and the definitions of the newly established families were less obvious and required use of morphometry, dissection of the mouthparts and having of at least basic experience in tanaidacean taxonomy to capture specific and often fine details in morphological structures. The definition of more recently erected families based on character of pereopod setation (Jóźwiak et al., 2009; Błażewicz et al., 2019), proportion of the uropods rami (Larsen and Wilson, 2002; Bird and Larsen, 2009; Błażewicz-Paszkowycz and Bamber, 2009) or shape of the sclerite that links the cheliped with cephalothorax (Błażewicz et al., 2021).
The application of molecular data in taxonomic studies can promote the validation of morphological data that, although indispensable for taxonomists, can be deficient for proponents of Linnaean taxonomy. Undeniably, integrative taxonomy, in which morphology supplemented with molecular and other data e.g., biology, ecology; (Kaiser et al., 2018; Jakiel et al., 2019, 2020), allows to reliable “group related species into genealogical trees, which represent the evolutionary lineage of modern organisms from common ancestors” (Paterlini, 2007). It serves an ideal way for establishing any new taxa. Nevertheless, decent quality in molecular data is extremely difficult to obtain when studying small deep-sea crustaceans, as they are often represented by single and small-sized specimens, or when historical collections are inappropriate for molecular analysis, i.e., fixed/preserved with formalin. For this reason, only requisite preservation and processing of the collection warrant successful molecular investigations (Riehl et al., 2014).
The decision here to establish the family Hamatipedidae is based solely on morphological observation and focused primarily on a unique setation of the carpus of pereopods 4–6, that has uniquely short, robust and bent spine that was termed a “hook” when the type-genus Hamatipeda was established (Błażewicz-Paszkowycz, 2007) to emphasize their unique character. Here, the three new species, whose ornamentation of these legs clearly indicates close affinity with earlier described species of Hamatipeda, are supplemented with two species, related, but sufficiently different to warrant two new genera (Rakaduta and Yarutanais) and place them in the new family.
Ornamentation and setation of crustacean legs are important components in understanding evolutionary relationships of modern organisms and their ancestors. In the most recent system proposed by Garm and Watling (2013), which simplified earlier setae classification (Garm, 2004; Garm and Watling, 2013), the setae were divided into seven categories depending on the function, ornamentation, articulation and the presence of terminal/subterminal pore that extends to an internal lumen and innervation (Figure 8D). The carpal spines (= hooks) which are diagnostic characters for the Hamatipedidae might be classified as cuspidate setae in Garm and Watling’s classification since they lack a terminal pore and reveal residual articulation (Figures 8B,C,E) unlike setae (Figure 8F). Loss of flexibility and articulation of the setae implies their purely mechanical function related for tube-life. Imaging of prickly tubercles (Figure 8G) shows that they are a separate structure from the three (spinulate) spines on the carpus and are probably derived from microtrichia and associated region of the carpal cuticle.
The low abundance of deep-sea populations often represented by a few individuals in the samples precludes studying a life cycle of deep-sea Tanaidacea. The knowledge we currently have on life history and reproductive strategies of deep water tanaids comes from observations on only a few shallow-water species that we extrapolate to deep-water species (see Esquete et al., 2012; Rumbold et al., 2014; Gellert and Błażewicz, 2018; Stępień et al., 2021). The material we investigated was unique, as one species — H. mojito was represented by 127 individuals at different developmental stages, which allowed us to make a series of measurements of total body length, body segments and uropodal rami. Our results have indicated the presence of two postmarsupial manca stages, as those observed for several shallow-water species (e.g., Bückle Ramírez, 1965; Fonseca and D’Incao, 2003). Although we did not observe females with developed oostegites in our material (neither fully developed nor oostegite buds), some neuters were clearly bigger than others. Without a thorough histological analysis assessing the degree of ovarian development, it is impossible to determine unequivocally the life history of H. mojito; however, it can be assumed that this species may breed at least twice in a lifetime, like other shallow-water tanaidomorphs (e.g., Lang, 1952; Bückle Ramírez, 1965; Johnson and Attramadal, 1982; Błażewicz-Paszkowycz, 2001; Toniollo and Masunari, 2007; Bamber, 2014). Our data also demonstrate that the last undeveloped last thoracomere (pereonite-6) of H. mojito grows about twice as fast as other pereonites. A similar observation was made for the pereonite-5 although it grows slower than in pereonite-6. Nevertheless, relative length of the last two pereonites cannot serve as a reliable diagnostic character. Conversely, the length of the uropod rami of M. mojito is constant during ontogenesis, offering a favorable diagnostic character.
Discovering and understanding the biodiversity of the deepest parts of the ocean is written in the priorities of recent marine biology, that is essential for efficient protection of the fragile deep-sea ecosystems. The pressure to apply modern and sophisticated research methods discourages us from focusing on unworked historical collections. Our results, however, demonstrate that investigation of even a small part of historical materials can substantially increase the knowledge of deep-sea diversity. This serves as a reference point for future analyses crucial for developing conservation strategies, and particularly important in the context of the global warming observed in recent decades, which affects also still unknown deep sea.
Data Availability Statement
This article is registered in ZooBank under 6CA6F1DE-4939-4FEC-9ACF-AEE48310D90A.
Author Contributions
MG: taxonomic identification, statistical, and manuscript writing. MB: concept of the manuscript, taxonomic identification, manuscript writing, and discussion. GB: manuscript writing and discussion. AS: statistical analysis and manuscript writing. MS: confocal imaging. All authors contributed to the article and approved the submitted version.
Funding
The research was financed by NCN OPUS (2018/31/B/NZ8/03198) and NCN PRELUDIUM (2021/41/N/NZ8/02039).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We would like to thank Joanne Taylor and Melanie Mackenzie (Melbourne Museum, Victoria) and Adam Baldinger (Museum of Comparative Zoology, Cambridge, MA, United States) for making the collection available for our research. We would also like to thank Prof. Mirosław Przybylski (University of Lodz) for help in statistical analysis and to Piotr Jóźwiak (University of Lodz) for help with SEM imaging.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2021.773437/full#supplementary-material
Footnotes
- ^ www.nikoninstruments.com
- ^ (*) See remarks (Kakui and Hiruta, 2021) and Supplementary Table 3.
- ^ http://www.marinespecies.org
References
Appeltans, W., Ahyong, S. T., Anderson, G., Angel, M. V., Artois, T., Bailly, N., et al. (2012). The magnitude of global marine species diversity. Curr. Biol. 22, 2189–2202. doi: 10.1016/j.cub.2012.09.036
Bamber, R. N. (2008). Tanaidaceans (Crustacea: Peracarida: Tanaidacea) from Moreton Bay, Queensland. Mem. Queensl. Museum 54, 143–218.
Bamber, R. N. (2014). Two new species of Sinelobus Sieg, 1980 (Crustacea: Tanaidacea: Tanaididae), and a correction to the higher taxonomic nomenclature. J. Nat. Hist. 48, 2049–2068. doi: 10.1080/00222933.2014.897767
Bird, G. J. (2002). A re-evaluation of the genus Tanaissus (Crustacea, Tanaidacea) in British and adjacent waters. Sarsia 87, 152–166. doi: 10.1080/003648202320205238
Bird, G. J. (2012). Stachyops, a new nototanaid genus (Crustacea: Peracarida: Tanaidacea) from New Zealand, with remarks on nototanaid and tanaissuid phylogeny. Zootaxa 10, 1–10. doi: 10.11646/zootaxa.3572.1.1
Bird, G. J., and Holdich, D. M. (1988). Deep-sea Tanaidacea (Crustacea) of the North-East Atlantic: the tribe Agathotanaini. J. Nat. Hist. 22, 1591–1621. doi: 10.1080/00222938800771001
Bird, G. J., and Larsen, K. (2009). Tanaidacean phylogeny – the second step: the basal paratanaoidean families (Crustacea: Malacostraca). Arthropod Syst. Phylogeny 67, 137–158.
Błażewicz, M., Jakiel, A., Bamber, R. N., and Bird, G. J. (2021). Pseudotanaidae Sieg, 1976 (Crustacea: Peracarida) from the Southern Ocean: diversity and bathymetric pattern. Eur. Zool. J. 88, 994–1070. doi: 10.1080/24750263.2021.1960444
Błażewicz, M., Jóźwiak, P., Jennings, R. M., Studzian, M., and Frutos, I. (2019). Integrative systematics and ecology of a new deep-sea family of tanaidacean crustaceans. Sci. Rep. 9:18720. doi: 10.1038/s41598-019-53446-1
Błażewicz-Paszkowycz, M. (2001). Remarks on the population structure of two Antarctic peracarid crustaceans: Eudorella splendida Zimmer, 1902 (Cumacea) and Nototanais antarcticus (Hodgson, 1902) (Tanaidacea). Pol. Polar Res. 22, 35–44.
Błażewicz-Paszkowycz, M. (2007). A revision of the family Typhlotanaidae Sieg, 1984 (Crustacea: Tanaidacea) with the remarks on the Nototanaidae Sieg, 1976. Zootaxa 1598, 1–141. doi: 10.11646/zootaxa.1598.1.1
Błażewicz-Paszkowycz, M., and Bamber, R. N. (2007). New apseudomorph tanaidaceans (Crustacea: Peracarida: Tanaidacea) from eastern Australia: Apseudidae, Whiteleggiidae, Metapseudidae and Pagurapseudidae. Mem. Museum Vic. 64, 107–148. doi: 10.24199/j.mmv.2007.64.11
Błażewicz-Paszkowycz, M., and Bamber, R. N. (2009). A new genus of a new Austral family of paratanaoid tanaidacean (Crustacea: Peracarida: Tanaidacea), with two new species. Mem. Museum Vic. 66, 5–15. doi: 10.24199/j.mmv.2009.66.2
Błażewicz-Paszkowycz, M., and Bamber, R. N. (2012). The shallow-water Tanaidacea (Arthropoda: Malacostraca: Peracarida) of the Bass Strait, Victoria, Australia (other than the Tanaidae). Mem. Museum Vic. 69, 1–235. doi: 10.24199/j.mmv.2012.69.01
Błażewicz-Paszkowycz, M., Bamber, R. N., and Anderson, G. (2012). Diversity of Tanaidacea (Crustacea: Peracarida) in the world’s oceans – how far have we come? PLoS One 7:e33068. doi: 10.1371/journal.pone.0033068
Brandt, A., Gooday, A. J., Brandão, S. N., Brix, S., Brökeland, W., Cedhagen, T., et al. (2007). First insights into the biodiversity and biogeography of the Southern Ocean deep sea. Nature 447, 307–311. doi: 10.1038/nature05827
Bückle Ramírez, L. F. (1965). Untersuchungen über die biologie von Heterotanais oerstendi Kroyer (Crustacea, Tanaidacea). Z. Morphol. Ökol. Tiere 55, 714–782. doi: 10.1007/BF00406235
Coleman, C. O. (2003). “Digital inking”: how to make perfect line drawings on computers. Org. Divers. Evol. 3, 303–304. doi: 10.1078/1439-6092-00081
Costello, M. J., and Chaudhary, C. (2017). Marine biodiversity, biogeography, deep-sea gradients, and conservation. Curr. Biol. 27, 511–527. doi: 10.1016/J.CUB.2017.04.060
Esquete, P., Bamber, R. N., Moreira, J., and Troncoso, J. S. (2012). Apseudopsis adami, a new species of tanaidacean (Crustacea: Peracarida) from the NW Iberian Peninsula: postmarsupial development and remarks on morphological characters. Helgol. Mar. Res. 66, 601–619. doi: 10.1007/s10152-012-0295-2
Fonseca, D. B., and D’Incao, F. (2003). Growth and reproductive parameters of Kalliapseudes schubartii in the estuarine region of the Lagoa dos Patos (southern Brazil). J. Mar. Biol. Assoc. U. K. 83, 931–935. doi: 10.1017/S0025315403008087h
Frutos, I., Brandt, A., and Sorbe, J. C. (2016). “Deep-sea suprabenthic communities: the forgotten biodiversity,” in Marine Animal Forests, eds S. Rossi, L. Bramanti, A. Gori, and C. Orejas Saco del Valle (Cham: Springer International Publishing), 1–29. doi: 10.1007/978-3-319-17001-5_21-1
Frutos, I., and Sorbe, J. C. (2014). Bathyal suprabenthic assemblages from the southern margin of the Capbreton Canyon (“Kostarrenkala” area), SE Bay of Biscay. Deep Sea Res. II Top. Stud. Oceanogr. 104, 291–309. doi: 10.1016/j.dsr2.2013.09.010
Garm, A. (2004). Revising the definition of the crustacean seta and setal classification systems based on examinations of the mouthpart setae of seven species of decapods. Zool. J. Linn. Soc. 142, 233–252. doi: 10.1111/j.1096-3642.2004.00132.x
Garm, A., and Watling, L. (2013). “The crustacean ontegument: setae, setules, and other ornamentation,” in Functional Morphology and Divresity, eds L. Watling and M. Thiel (Oxford: Oxford University Press), 167–198. doi: 10.1093/acprof
Gellert, M., and Błażewicz, M. (2018). New species of Anarthruridae (Tanaidacea: Crustacea) of the western Australian slope. Mar. Biodivers. 49, 583–601. doi: 10.1007/s12526-017-0826-9
Gellert, M., Palero, F., and Błażewicz, M. (unpublished). Deeper diversity exploration: new Typhlotanaidae (Crustacea: Tanaidacea) from the Kuril–Kamchatka Trench and nearby waters. Front. Mar. Sci.
Hartnoll, R. G. (1982). “Growth,” in The Biology of Crustacea, Embryology, Morphology and Genetics, Vol. 2, eds D. E. Bliss and L. G. Abele (New York, NY: Academic Press), 111–196.
Jakiel, A., Palero, F., and Błażewicz, M. (2019). Deep ocean seascape and Pseudotanaidae (Crustacea: Tanaidacea) diversity at the Clarion-Clipperton Fracture Zone. Sci. Rep. 9:17305. doi: 10.1038/s41598-019-51434-z
Jakiel, A., Palero, F., and Błażewicz, M. (2020). Secrets from the deep: Pseudotanaidae (Crustacea: Tanaidacea) diversity from the Kuril–Kamchatka Trench. Prog. Oceanogr. 183:102288. doi: 10.1016/j.pocean.2020.102288
Jażdżewska, A. M., Corbari, L., Driskell, A., Frutos, I., Havermans, C., Hendrycks, E., et al. (2018). A genetic fingerprint of Amphipoda from Icelandic waters-the baseline for further biodiversity and biogeography studies. Zookeys 731, 55–73. doi: 10.3897/zookeys.731.19931
Jażdżewska, A. M., Horton, T., Hendrycks, E., Mamos, T., Driskell, A. C., Brix, S., et al. (2021). Pandora’s box in the deep sea—intraspecific diversity patterns and distribution of two congeneric scavenging amphipods. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.750180
Johnson, S., and Attramadal, Y. G. (1982). Reproductive behaviour and larval development of Tanais cavolirrii (Crustacea: Tanaidacea). Mar. Biol. 71, 11–16. doi: 10.1007/BF00396987
Jóźwiak, P., Stępień, A., and Błażewicz-Paszkowycz, M. (2009). A revision of the genus Paranarthrurella Lang, 1971a (Crustacea: Tanaidacea). Zootaxa 2238, 56–68. doi: 10.11646/zootaxa.2238.1.5
Kaiser, S., Brix, S., Kihara, T. C., Janssen, A., and Jennings, R. M. (2018). Integrative species delimitation in the deep-sea genus Thaumastosoma Hessler, 1970 (Isopoda, Asellota, Nannoniscidae) reveals a new genus and species from the Atlantic and central Pacific abyss. Deep Res. II Top. Stud. Oceanogr. 148, 151–179. doi: 10.1016/j.dsr2.2017.05.006
Kakui, K., and Hiruta, C. (2021). Description of a new Hamatipeda species, with an 18S molecular phylogeny (Crustacea: Tanaidacea: Typhlotanaidae). Zool. Sci. 39, 1–7. doi: 10.2108/zs210065
Kakui, K., Katoh, T., Hiruta, S. F., Kobayashi, N., Kajihara, H., Kakui, K., et al. (2011). Molecular systematics of Tanaidacea (Crustacea: Peracarida) based on 18S sequence data, with an amendment of suborder/superfamily-level classification. Zool. Sci. 28, 749–757. doi: 10.2108/zsj.28.749
Kudinova-Pasternak, R. K. (1975). Tanaidacea, Atlantic sector, Antarctic and subantarctic. Akad. Nauk SSSR 103, 194–229. doi: 10.11646/zootaxa.3630.3.2
Kudinova-Pasternak, R. K., and Pasternak, F. A. (1978). Deep-sea Tanaidacea (Crustacea, Malacostraca) collected in the Caribbean Sea and Puerto-Rico trench during the 16-th cruise of R/V “Akademic Kurchatov” and the resemblance between fauna of deep-sea Tanaidacea of the Caribbean region and the Pacific. Trudy Inst. Okeanol. 113, 178–197.
Lang, K. (1949). Contribution to the systematics and synonymics of the Tanaidacea. Ark. Zool. 42, 1–14.
Lang, K. (1971). Taxonomische und phylogenetische Untersuchungen über die Tanaidaceen 7. Revision der Gattung Strongylura G. 0. Sars, 1882, nebst Beschreibung einer neuen Art dieser Gattung. Ark. Zool. 23, 403–415.
Lang, K. (1973). Taxonomische und phylogenetische Untersuchungen über die Tanaidaceen (Crustacea). Zool. Scr. 2, 197–229. doi: 10.1111/j.1463-6409.1974.tb00752.x
Larsen, K. (2005). Deep-Sea Tanaidacea (Peracarida) from the Gulf of Mexico. Leiden: Brill. doi: 10.1163/9789047416883
Larsen, K. Błażewicz-Paszkowycz, M., and Cunha, M. R. (2006). Tanaidacean (Crustacea: Peracarida) fauna from chemically reduced habitats–the Lucky Strike hydrothermal vent system, Mid-Atlantic Ridge. Zootaxa 1187, 1–36. doi: 10.11646/zootaxa.1187.1.1
Larsen, K., and Heard, R. W. (2001). A new tanaidacean subfamily, Bathytanaidinae (Crustacea: Paratanaididae), from the Australian continental shelf and slope. Zootaxa 19, 1–22. doi: 10.11646/zootaxa.19.1.1
Larsen, K., and Wilson, G. D. F. (2002). Tanaidacean phylogeny, the first step: the superfamily Paratanaidoidea. J. Zool. Syst. Evol. Res. 40, 205–222. doi: 10.1046/j.1439-0469.2002.00193.x
McCallum, A. W., Woolley, S., Błażewicz-Paszkowycz, M., Browne, J., Gerken, S., Kloser, R., et al. (2015). Productivity enhances benthic species richness along an oligotrophic Indian Ocean continental margin. Glob. Ecol. Biogeogr. 24, 462–471. doi: 10.1111/geb.12255
Paterlini, M. (2007). There shall be order. The legacy of Linnaeus in the age of molecular biology. EMBO Rep. 8, 814–816. doi: 10.1038/sj.embor.7401061
Ramirez-Llodra, E., Tyler, P. A., Baker, M. C., Bergstad, O. A., Clark, M. R., Escobar, E., et al. (2011). Man and the last great wilderness: human impact on the deep sea. PLoS One 6:e22588. doi: 10.1371/journal.pone.0022588
Riehl, T., Brenken, N., Brix, S., Driskell, A., Kaiser, S., and Brandt, A. (2014). Field and laboratory methods for DNA studies on deep-sea isopod crustaceans. Pol. Polar Res. 35, 203–224. doi: 10.2478/popore
Rumbold, C. E., Spivak, E. D., and Obenat, S. M. (2014). Morphometry and relative growth of populations of Tanais dulongii (Audoin, 1826) (Tanaidacea: Tanaidae) in pristine and impacted marine environments of the Southwestern Atlantic. J. Crustac. Biol. 34, 581–592. doi: 10.1163/1937240X-00002265
Sanders, H. L. (1968). Marine benthic diversity: a comparative study. Am. Nat. 102, 243–282. doi: 10.1086/282541
Sanders, H. L., Hessler, R. R., and Hampson, G. R. (1965). An introduction to the study of deep-sea benthic faunal assemblages along the Gay Head-Bermuda transect. Deep Res. Oceanogr. Abstr. 12, 845–867. doi: 10.1016/0011-7471(65)90808-9
Segadilha, J. L., Gellert, M., and Błażewicz, M. (2018). A new genus of Tanaidacea (Peracarida, Typhlotanaidae) from the Atlantic slope. Mar. Biodivers. 48, 915–925. doi: 10.1007/s12526-018-0856-y
Segadilha, J. L., Serejo, C. S., and Błażewicz, M. (2019). New species of Typhlotanaidae (Crustacea, Tanaidacea) from the Brazilian coast: genera Hamatipeda, Meromonakantha and Paratyphlotanais, with description of Targaryenella gen. nov. Zootaxa 4661, 309–342. doi: 10.11646/zootaxa.4661.2.4
Sieg, J. (1976). To the natural system of Dikonophora Lang (Crustacea, Tanaidacea). Crustac. Monogr. 35, 119–133.
Sieg, J. (1980). Taxonomische Monographie der Tanaidae Dana, 1849 (Crustacea: Tanaidacea). Frankfurt am Main: W. Kramer.
Sieg, J. (1986). Tanaidacea (Crustacea) von der Antartiks und Subantarktis. II. Tanaidacea gesammelt von Dr. J. W. Wägele während der Deutschen Antarktis expedition 1983. Mitt. Zool. Museum Univ. Kiel 4, 1–80.
Stępień, A., Jóźwiak, P., Jakiel, A., Pełczyńska, A., and Błażewicz, M. (2021). Diversity and abundance of Pacific Agathotanais (Pericarida: Tanaidacea). Front. Mar. Sci. doi: 10.3389/fmars.2021.741536
Szczepanek, R. (2017). Systemy Informacji Przestrzennej z QGIS część I i II. Kraków: Wydawnictwo PK.
Keywords: Paratanaoidea, Hamatipeda, Rakaduta, Yarutanais, taxonomy, slope, abyssal, diversity
Citation: Gellert M, Bird G, Stȩpień, A, Studzian M and Błażewicz M (2022) A Hidden Diversity in the Atlantic and the SE Pacific: Hamatipedidae n. fam. (Crustacea: Tanaidacea). Front. Mar. Sci. 8:773437. doi: 10.3389/fmars.2021.773437
Received: 29 September 2021; Accepted: 06 December 2021;
Published: 08 February 2022.
Edited by:
Marcos Rubal, University of Porto, PortugalReviewed by:
Patricia Esquete Garrote, University of Aveiro, PortugalCatarina Araújo-Silva, Federal University of Pernambuco, Brazil
Copyright © 2022 Gellert, Bird, Stȩpień, Studzian and Błażewicz. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marta Gellert, bWFydGEuZ2VsbGVydEBlZHUudW5pLmxvZHoucGw=