 Camilo Cortés-Useche1,2*Williams Reyes-Gamboa1José Luís Cabrera-Pérez1Johanna Calle-Triviño2Ana Cerón-Flores3Rodolfo Raigoza-Figueras3Roshni Yathiraj4Jesús Ernesto Arias-González1*
Camilo Cortés-Useche1,2*Williams Reyes-Gamboa1José Luís Cabrera-Pérez1Johanna Calle-Triviño2Ana Cerón-Flores3Rodolfo Raigoza-Figueras3Roshni Yathiraj4Jesús Ernesto Arias-González1*- 1Laboratorio de Ecología de Ecosistemas de Arrecifes Coralinos, Departamento de Recursos del Mar, Centro de Investigación y de Estudios Avanzados del IPN, Unidad Mérida, Mérida, Mexico
- 2Wave of Change Iberostar Hotels & Resorts, Mexico City, Mexico
- 3Acuario XCARET, Playa del Carmen, Mexico
- 4Nature Seychelles, Centre for Environment & Education, Mahe, Seychelles
The changing world presents negative impacts on marine ecosystems and has led to the development of diversified tools to support reef restoration. Harnessing restoration to achieve success needs innovative techniques that also address the restoration of reef fish assemblages, contributing to the conservation of biodiversity and ecosystem functions and also tackle the cost-effectiveness through impact-driven solutions. Here, we propose a proof-of-concept for enhancing fish populations on reefs using: (1) postlarvae capture, (2) aquarium culture, and (3) release to reef sites. We conducted field studies in the Mexican Caribbean to analyze for the first time, the possibility of using the capture and aquarium culture of postlarvae fish species and release of juveniles as a tool for the potential recovery of reef biodiversity resilience. We tested the potential of postlarvae capture using two distinct night light traps (BOX and collect by artificial reef ecofriendly traps, C.A.R.E.) in three sampling sites with different distances from shoreline and depth. We collected 748 postlarvae reef fishes from eight orders, 20 families, and 40 species. Acanthuridae, Pomacentridae, Monacanthidae, and Tetraodontidae comprised the highest species number of postlarvae families. We also set up a pilot release experiment with Stegastes partitus using two trials (32 and 1 day after capture) and propose analysis to determine appropriate reef sites to release the cultured juveniles and to aid ecological planning. We present the results of the pilot release experiment with S. partitus, showing that there is a positive effect in survivorship during the capture (80%) and release (76–100%) procedures into suitable habitat and good chance that more studies will bring novelty to the field. Although trials carried out with more species relevant to restoration will be needed. The use of these techniques can be a great opportunity to improve the research of restoration efforts in the Caribbean region with fish-depleted coral reefs with vulnerable food webs, especially at local scales and supporting other management strategies.
Introduction
Active strategies approach has been proposed (Schmidt-Roach et al., 2020) to promote reef restoration in areas with severely decreased reef fish communities (Abelson et al., 2016). In the Caribbean, restoration efforts have been focused on the recovery of corals (Bayraktarov et al., 2020; Calle-Triviño et al., 2020). Although social and ecological outcomes target various benefits (Calle-Triviño et al., 2018), current challenges of these efforts include long-term implementation, identifying the feasibility of this implementation, and ecological processes restoration (Ladd et al., 2018; Duarte et al., 2020).
Efforts for broadening the reef restoration from coral species to fishes remain limited and have been rarely used as a method (Obolski et al., 2016). Nevertheless, reef fishes have a pelagic larval phase during their lifecycle that allows them to disperse spatially and a demersal juvenile phase that leads to the colonization of these species on a reef (Sale, 2015). The former is one of the most critical stages in their cycle, which determines the characteristics of the populations such as distribution, abundance, and population dynamics (Victor and Wellington, 2000; Simpson et al., 2013). It is a phase where they experience high-mortality rates of ~60% (Doherty et al., 2004; Almany and Webster, 2006), and survival is often related to early life-history traits (Sponaugle et al., 2011). This postlarval phase continues to be studied to understand the settlement processes (Dufour, 1994; Hendriks et al., 2001; McCormick et al., 2002; Lecaillon, 2017). Several studies have used settlement stage reef fishes to realize small-scale fisheries based on postlarval capture and culture (Bell et al., 2009). This method has also been integrated into experimental protocols as a potential tool for restoration of fish assemblages as a proof-of-concept (Heenan et al., 2009; Abelson et al., 2016). The aforementioned is based on the concept of removing extremely high-mortality rates that occur during settlement, and post-settlement of the first few weeks, and take advantage of this process to significantly increase survivorship (Vallès et al., 2008).
Several techniques have been developed to capture coral reef fish in the early life stages (Choat et al., 1983), including light traps (Lecaillon, 2004; Moana Initiative, 2007), “hoa” nets that are used in shallow passes (termed hoa in French Polynesia) that allow water to enter “closed” and “semiclosed” atoll lagoons (Lecaillon and Lourié, 2007) and crest nets in French Polynesia (Dufour and Galzin, 1993), Australia (Doherty and McIlwain, 1996), Solomon Islands (Hair et al., 2002), and La Reunion (Durville et al., 2003). In addition, extensive light trap work has been carried out in the Florida Keys, focused on examining and measuring the processes of larval fish supply (Sponaugle et al., 2006; D'Alessandro et al., 2007) and capture during the settlement stage of Stegastes partitus larvae (Rankin, 2010). Remarkably, there have been very few studies on the postlarvae and early stages of ichthyofauna within the Caribbean coral reefs. Vásquez-Yeomans et al. (1998, 2003, 2011) and Álvarez-Cadena et al. (2007) carried out studies on ichthyoplankton obtained by surface trawling, and channel nets. Recently, Ayala-Campos (2014) used a light trap technique for sampling.
Despite these research efforts, little data have been collected on postlarvae of the fish in the Caribbean region, some of which have not been published. In addition, implementing these low-environmental impact techniques as an effective tool for biodiversity monitoring and conservation activities are absent. Given this situation, we performed a proof-of-concept to enhance fish populations on reefs by using techniques for postlarvae capture, aquarium culture, and release. In addition, the goal of this study was to supply primary information to address the current interest in coral reef restoration. To do so, we described temporal variations in postlarvae abundance of reef fishes during the high-settlement season by testing two different kinds of night light traps. We addressed the aquarium-culture factors and release of cultured juveniles, using a pilot release experiment with S. partitus and a landscape analysis. We use these results and analysis to test feasibility and comment on the potential ecological application of this method into Caribbean restoration efforts.
Materials and Methods
Study Approach and Location
A primary criterion for applying our approach and address the study was to consider a multidisciplinary engage approach. It was necessary to establish a strong partnership among the science, public (marine-protected area; MPA), and private sectors (Xcaret and Wave of Change) (Abelson et al., 2016; Doropoulos et al., 2019). In this sense, operationalizing the study, the choice of study site, technical process setup, and field collection were conducted based on the requirements for feasible applications in the Caribbean region (Figure 1).
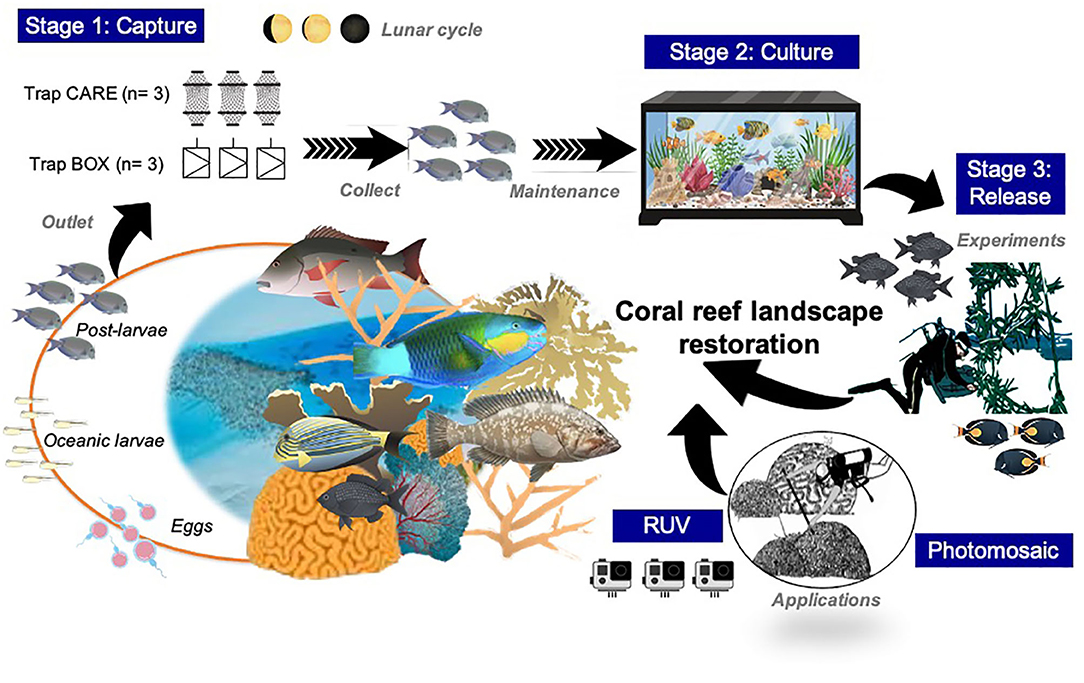
Figure 1. General outline process for scaling up restoration of coral reef using oostlarval reef fishes: capture, culture, and release.
The study was performed within an MPA in the northern part of the State of Quintana “Reserva de la Biosfera del Caribe Mexicano.” This location was chosen because: (1) the area is managed by the authorities (Comisión Nacional de Áreas Naturales Protegidas; CONANP); (2) the staff of Xcaret control the recreational activities including protection and fishing regulations; and (3) it has proximity to logistic facilities to support the study in all the stages. Sampling was conducted in the three sites: Site 1 (Punta INAH reef) is closer to the shore (100 m) with a depth of 10 m. This area is characterized by more complex coral reef structures in comparison with the other sites, the reef crest is conspicuous, and the area is covered with larger seagrasses. At Site 2 (Punta Venado reef), the distance to the coast is 200 m, with an average depth of 12 m, the bottom is mainly characterized by low-seagrass coverage and some isolated reef patches. At Site 3 (Calica reef), the distance to the shoreline is 300 m, with a depth of 15 m, dominated by a few isolated patches of seagrass and coral reef.
Device for Capturing Postlarvae and Methodology
Between July and October 2018, which are the months for the highest recruitment (Williams and Sale, 1981; Leis and McCormick, 2002; Watson et al., 2002; Ayala-Campos, 2014), monthly catches of postlarvae fish were conducted at all three sites as a part of the first stage method. These catches consisted of the placement of six-night light fish traps in total (n = 3 replicates per treatment) at each site: three Australian design single chamber box (acrylic) traps on each side and three C.A.R.E design traps (Collect by Artificial Reef Ecofriendly patented by Ecocean) (Lecaillon, 2004) (Supplementary Figure S1). The traps were deployed during nights of minimal lunar illumination (new moon), set at sunset, and removed at sunrise (minimum of 10 h functioning) for 7 consecutive days.
Aquarium-Culture of Postlarval Fish
As a second stage, a total of 48 fish tanks (32 tanks × 10 L and 16 tanks × 20 L) were designed and built for the sorting, maintenance, and observation of captured organisms. The larger tanks were used to accommodate species of fish that were collected in larger numbers to prevent overstocking or hyperpredation in the tanks. On capture days, the fish were distributed in fish tanks according to the species or behavior (territorial, aggressive, and passive). The fish tanks were conditioned with shelters made of sheets of PVC and/or raffia fibers. The fish were fed twice a day (at 0,800 and 1,700 h) with nauplius, adult artemia (Artemia sp.), and commercially prepared food. The organisms observed were identified following the Cervigón et al. (1992), Humann and DeLoach (2002) identification guides and systematically sorted according to FishBase criteria (Froese and Pauly, 2019). All the collected postlarvae were monitored for 15 days post-capture (complete metamorphosis) to test for differences in survival (Victor, 1991).
Release of Cultured Juveniles Into Caribbean Reefs
To explore the role of releasing cultured juveniles as a feasible restoration tool, we considered site-specific survival, efficiency of target species behavior, and ecology of reef fishes. Given that habitat variation is a key modeling factor in the early-stage of reef fishes (Paddack and Sponaugle, 2008), we performed a pilot release study in a controlled matrix or artificial structure for the juveniles. As the first step of this stage, we selected individuals of bicolor damselfish (Stegastes partitus) for the experiment based on their prevalence, territorial behavior, and their fidelity to the habitat where they settle (Thiessen and Heath, 2006; Heenan et al., 2009). A total of 42 samples of S. partitus were collected and released using two trials to test the site survival of released fish: (1) postlarvae collected and cultured for a period of 32 days; and (2) juveniles collected and released 1 day after capture. Trials were equally split among 21 individuals, resulting in three replicates placed in six tanks (200 L) with seven individuals per tank, considering a 3 cm size for all the replicates. The tanks had artificial structures (matrix) made of cement, raffia, and ceramic tiles as refuges. Before the release of cultured juveniles, the structures were transported to the selected slopes (12 m depth). The artificial structures were haphazardly arranged on the slopes of reefs separated by 10 m between each structure to prevent migration between them. Release protocol and duration of visual counts were focused on to assess post-release survival. Based on the methodology used by Heenan et al. (2009), site survival of released fish was measured twice on days 1, 2, and 3 using visual census (at 0,800 and 1,700 h) and then once daily (0,800 h) for another 5 days, for a total of 8 days.
Selecting Target Sites for Release Criteria
Environmental requirements such as structural complexity, coral composition, fish community, and benthic reef cover need to be considered to enhance the release approach (Figure 2). We used the Agisoft Metashape Professional Edition software to process preliminary images that we obtained by taking pictures with two cameras (GoPro 8) for the building of the photomosaics in Cozumel (Francesita reef) and Riviera Maya (Manchoncitos reef). In addition, we carried out an assessment corresponding to fish communities already present.
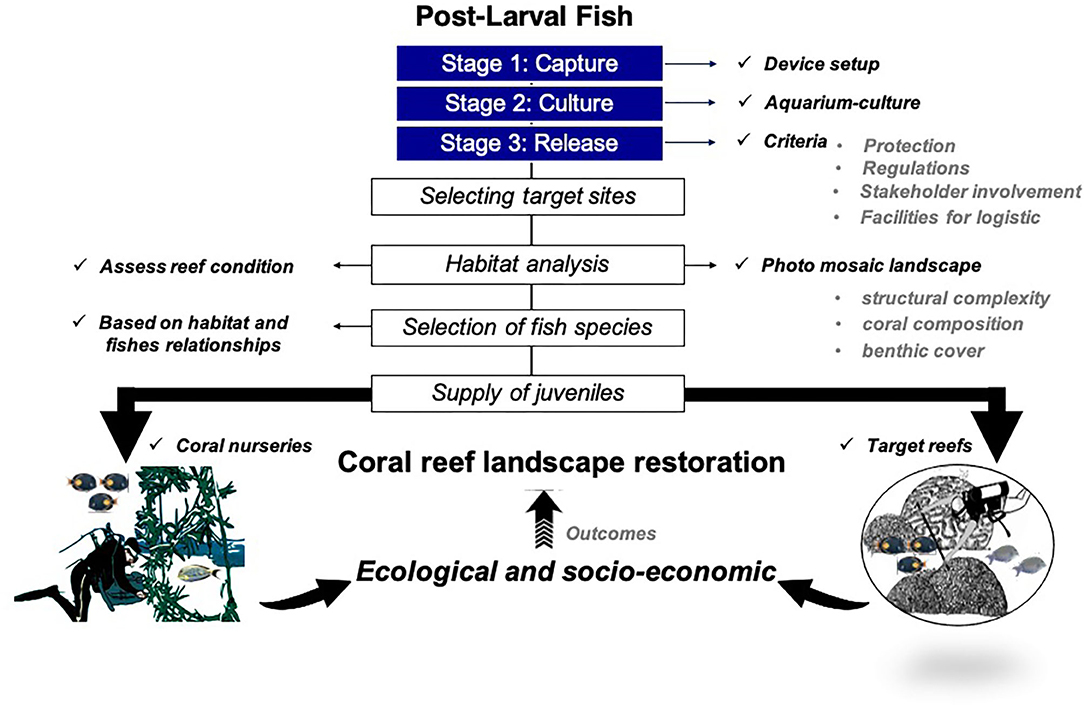
Figure 2. Outline for selecting target sites for release postlarval reef dishes after capture and aquarium culture.
Data Analysis
To recognize the temporal variations through species abundance, we evaluated species richness (number of species present in each sample) and Shannon-Weaver's diversity (H ) (Jost, 2007). An analysis of the local contribution to beta diversity was used to determine the percentage of contribution in each of the sites and identifying the uniqueness based on existing diversity (Legendre and De Cáceres, 2013).
A Multivariate Permutational Analysis of Nested Variance (Nested PERMANOVA) was carried out to determine differences between capture months. The ANOVA one-way was used to determine differences between the catch per unit effort (CPUE) (individual/trap/day) of fish caught by the two different light night traps. A CAP was carried out to visualize the dispersion of the samples through a season with the same data matrix. A heat map (descriptive analysis) of the total abundances of postlarvae species captured by the months of study was made, with cluster analysis (similarity with Euclidean distances).
Differences in site survival of released fish among trials were compared with Kaplan-Meier's survival analysis, with CIs (α = 0.05), followed by Log-Rank (Wilcoxon) pairwise comparisons. All the analyses were performed and plotted using the statistical program R version 3.3.1 (R Core Team, 2017).
Results
A total of 748 postlarval reef fishes from eight orders, 20 families, and 40 species were identified (Table 1). The most abundant species were those belonging to the families Acanthuridae, Pomacentridae, Monacanthidae, and Tetraodontidae. The most dominant species were the bicolor damselfish (Stegastes partitus) with 132 individuals, followed by the sharpnose-puffer fish of the family Tetraodontidae (Canthigaster rostrate) with 121 individuals. The highest abundances and CPUE occurred in October (267 individual and 10 individual/trap/day), followed by August (227 individual and 8.36 individual/trap/day). The highest richness (taxa) and diversity (H') was recorded in August and September, and the lowest was in July (Supplementary Figure S2).
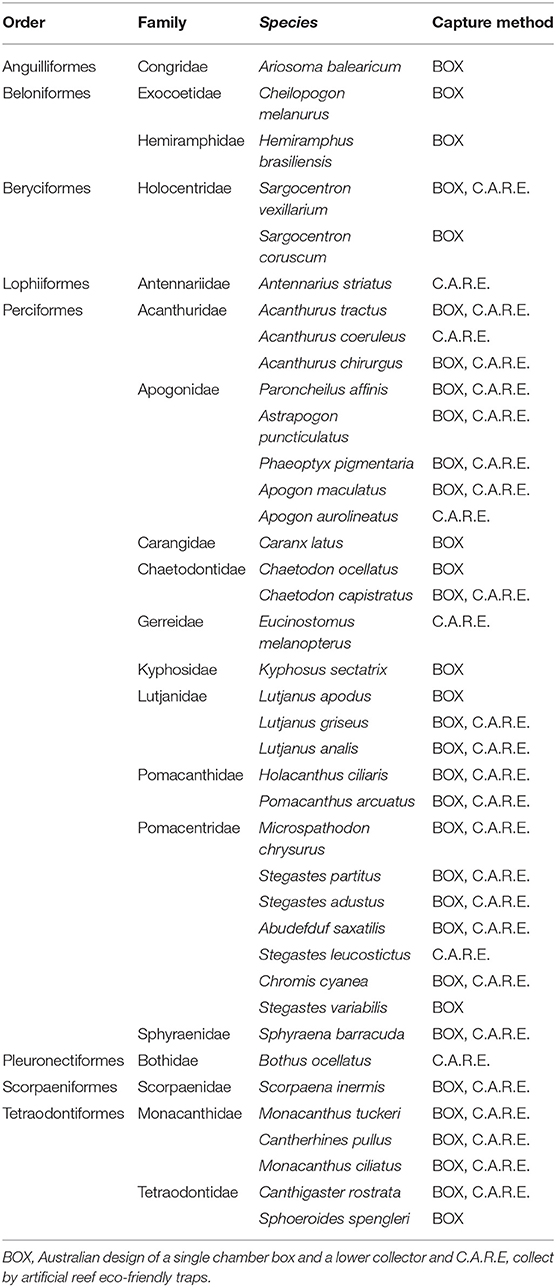
Table 1. Taxonomic list of postlarval fish recorded in the northern Mexican Caribbean.
Species composition by month showed significant differences in terms of species replacement component of beta diversity (p < 0.001) (Supplementary Table S1). Canonical Analysis of Principal Coordinates confirms differences among months (Supplementary Figure S3). July and October were the months with the highest variation of species and least similar in terms of beta diversity, while August and September were more similar (Supplementary Figure S4).
Of the total of 748 individuals, each of the trap types collected approximately 50% of the total fish present (367 individuals in the box design light traps and 381 individuals in the C.A.R.E light traps). The species with the most significant contribution were: S. partitus (Family: Pomacentridae), Canthigaster rostrata (Family: Tetraodontidae), Monacanthus tuckeri, and Cantherhines pullus (Family: Monacanthidae) with a contribution of more than 50% of the total species composition (Figure 3). Species composition between the different months showed the highest abundance and presence of S. partitus and C. rostrata in all the months. However, most of the less abundant species were replaced month by month, for example, of the 23 species recorded in August 9 species were replaced in October.

Figure 3. Some of the most abundant species recorded in the area include the following: (A) Stegastes partitus; (B) Canthigaster rostrata; (C) Monacanthus tuckeri, (D) Acanthurus chirurgus, (E) Acanthurus tractus, and (F) Mycrospathodon chrysurus.
A total of 367 individuals of 34 species were identified from the box light traps. Several rare and unique species were recorded in the net catches in this trap. Of these species, six presented only one individual (Lutjanus apodus, Kyphosus sectatrix, Sargocentron coruscum, Hemiramphus brasiliensis, Ariosoma balearicum, and Caranx latux), and two presented more than one individual (S. variabilis, Sphoeroides spengleri, and Cheilopogon melanurus). The C.A.R.E. light traps captured a total of 381 individuals of 30 species. Five species were unique to this trap and were rare with at least one individual per species (Eucinostomus melanopterus, Bothus ocellatus, Apogon aurolineatus, and Antennarius striatus). The species Acanthurus coeruleus, with an abundance of seven individuals, was caught only by the C.A.R.E. trap. Despite the observed differences in species composition by trap type, the species replacement component of beta diversity was not significant (PERMANOVA, 999 permutations: F1, 53 = 1.23, p = 0.45). However, the one-way ANOVA for CPUE variation between the two types of traps showed a significant difference (P = 0.000676) (Supplementary Table S2). The C.A.R.E. type traps obtained not only higher CPUE of postlarvae type but also more considerable variation in their records (12.4 individual/trap/day ± 5.54 CI) when compared with the box design traps (4.3 individual/trap/day ± 1.26 CI).
Survivorship results based on aquarium culture were analyzed using an overall approach. Survivorship of total postlarvae captured during the aquarium-culture stage was 80% until 15 days of post-capture. Also, we evaluated the survivorship for release stage, after 8 days of the site-specific survival experiment, and 76% of juveniles released within 1 day of capture (n = 16) survived, while juveniles released after 32 days in culture survived 100% (n = 21). A significant difference in survival of juveniles released was evident based on results for the two trials (P = 0.04, X2 = 4.2).
Results based on the preliminary landscape analyses suggest different target sites for release criteria. For example, a seascape dominated by Agaricia agaricites, A. tenuifolia, Porites porites, and P. astreoides were present in Cozumel (Mexico) on a shallow reef (8–10 m). The reef-building corals (e.g., Orbicella complex and Montastrea cavernosa) were present in the Manchoncitos Reef (Riviera Maya) between 9 and 13 m depth range. The highest contributions of fish species in Francesita reef 27.6% were covered by grunts, 11.8% by surgeonfishes, and 10.1% by parrotfishes, while in Manchoncitos reef were 43.6% were covered by grunts and 27.1% by parrotfishes. Haemulon flavolineatum and H. aurolineatum were the most species abundant in Francesita, while Caranx ruber and H. aurolineatum were in the Manchoncitos reef.
Discussion
This study describes the method of capture using two kinds of night light traps, BOX (Doherty, 1987) and C.A.R.E (Ecocean, 2020). Even though this study is preliminary, results show some distribution patterns similar to fish postlarvae in a previous survey (Ayala-Campos, 2014). The species richness (40 spp.) is consistent with most of the representative families: Acanthuridae, Pomacentridae, Monacanthidae, and Tetraodontidae, in addition to the more abundant species S. partitus and C. rostrata (Álvarez-Cadena et al., 2007). Therefore, the species identified in this work constitute 40% of the taxonomic diversity for this region reported by Álvarez-Cadena et al. (2007). In previous studies, 118 species belonging to 53 families and 115 species belonging to 55 families were recorded, respectively (Vásquez-Yeomans et al., 2011). However, most of the research they conducted was not recorded as postlarvae.
The difference in species richness may be related to the method and effort of sampling (Cortés-Useche et al., 2018). In the previous studies conducted in the Mexican Atlantic, a greater number of sites and different sampling methods (i.e., trawls, dredge, and crest nets) were used (Del Moral-Flores et al., 2013). In addition, other studies have focused on the estimation of species richness in tropical communities through the use of various sampling gear and methods (Vásquez-Yeomans et al., 2011). Considering the biology of the species besides their positive phototactic response, capture with light traps has the advantage of efficiency in catching reef species which prefer sheltered locations (tigmotropism), while non-target pelagic fish just swim over the reef (Lecaillon and Lourié, 2007). However, in order to increase efforts in the context of reef restoration, the use of other types of traps such as hoa traps and crest nets may be considered.
The capture of postlarvae during the summer season observed here is consistent with the highest CPUE in French Caribbean islands (Lecaillon, 2017). Our study, focused on captures made, taking into account that most of the recruitment occurs during a relatively short period, during the summer months, and postlarvae have movement during dusk and at night which is more significant on nights with lower luminosity (Dufour and Galzin, 1993; Milicich and Doherty, 1994; Leis and McCormick, 2002; Watson et al., 2002). However, the differences observed between October and July could be due to peaks of temperature during the summer. Villegas-Sánchez et al. (2009) associate the differences in these peaks with variations in sea temperature.
The catches obtained by the two kinds of night light traps showed that the C.A.R.E. trap obtained higher abundances (CPUE), but a lower richness (four fewer species). The effectiveness of this method is because it uses an artificially lit space that takes advantage of the behavior of the recruits to catch them (Lecaillon and Lourié, 2007): (1) the attraction to light (phototropism); (2) the desire to come into contact with a solid object (tigmotropism); and (3) the need to take refuge from predators. These sensory elements are essential for fish postlarvae, which have very acute senses during recruitment (Sweatman, 1988; Kingsford et al., 2002; Lecaillon and Lourié, 2007). The differences observed in CPUE may be associated with the design of the two types of traps, the intensity and type of light (Vadziutsina and Riera, 2020). It is important to highlight the fact there is a risk of the occurrence of hyperpredation in and around the traps during the capture process. For example, in our study, we recorded the presence of predatory individuals of S. barracuda, and we recommend checking the traps continuously to avoid filling them with many individuals and predatory species. In addition, we recommend separating the captures early on in the boat by species and size, thus avoiding the loss of individuals and the loss of benefits of this method of local capture.
Despite the differences between the two types of traps, C.A.R.E and BOX traps can be considered for harvest in a complementary range of species, to catch a greater diversity and larger number of fish (Dufour et al., 1996). In this study, postlarvae collected (Table 1) through the C.A.R.E and BOX traps were effective in terms of both the diversity (40 species) and abundance (748 target postlarvae collected). These results illustrate that capture of postlarvae in the summer season in the context of feasibility is the most appropriate period for collection in the Caribbean region. Moreover, following sampling from three sites, the results of the abundance suggest that further upscaling of locations can provide a more significant number of captured organisms to improve culture and release stages.
In this study, the overall production of juveniles has been achieved successfully with almost 80% survival until 15 days post-capture. For other reef fishes, Durville et al. (2003) cited survival between 60 and 92%, and Moana Initiative (2007) reports survival of target fish grown out were 80% after reaching the juvenile size. Survival can be considered as a crucial criterion for evaluating the optimum-rearing condition. Evidence from experimental studies with reef fishes (Planes and Lecaillon, 2001; Steele and Forrester, 2002; Webster, 2002; Doherty et al., 2004) suggests an advantage in the culture of postlarvae in contrast to postlarvae that settled in the wild. These experience high mortality as a result of increased predation rates (Bailey and Houde, 1989; Doherty, 2002). This result is significant for future directions of scaling up production as it allows for feasibility in its implementation. Overall, technical knowledge of aquarium culture must be expanded, for example, tests are needed to compare survival rates post-capture of specific species and functional groups (i.e., based on trophic level).
To our knowledge, our study is the first to carry out a series of experiments that involves related themes of growth, feeding, and reproduction of reef fish cycle as a potential tool to contribute to restoring Caribbean coral reefs. In Caribbean coral reefs, there is no reviewed literature about restocking experience (Obolski et al., 2016). Our results highlight the need to take advantage of the colonization phase when the postlarvae transform into juveniles and suffer catastrophic mortality rates (>90%) in the week following colonization (Planes et al., 2002; Doherty et al., 2004).
The release stage for future implementation also requires several considerations. As a first step, we addressed the recording of the standard length and total length of individuals on the day of capture and the day of release. This was done to eliminate differences in the sizes of the juveniles released, as several studies indicate that larger individuals may have greater experience with predators, which may result in causing a wary behavior during the experiment (Rankin, 2010). While the pilot experiment with S. partitus showed high survival, damselfishes need to be tested in the context of coral restoration (Heenan et al., 2009). It is crucial to determine if they support or undermine the restoration efforts (Ladd et al., 2018) by algal farming which can lead to tissue mortality (Precht et al., 2010) and/or reduce the presence of other corallivorous by defending their territory (Schopmeyer and Lirman, 2015). It is reasonable to expect that the release of species with different behavioral and ecological characteristics can influence survivorship and migration rates, even in the processes of trophic interactions in coral reefs (Ladd and Andrew, 2020). However, in the context of the experimental setup species with similar life history and behavior (e.g., site fidelity) to that of other damselfishes, may be the starting point for scaling up to other species or groups.
Caribbean reefs have experienced unprecedented declines; they are characterized by coral-algal phase shifts in which coral cover is declining to be replaced by algae (Arias-González et al., 2017). The choice of release habitat should be considered using landscape analysis. For example, based on the relationship between habitat condition and target species, where habitats dominated by macroalgae or algal turfs can be supplied by herbivorous species such as grazers (surgeonfish), scrapers (parrotfish), and browsers (chubs) to provide a top-down control of algae (Green and Bellwood, 2009; Obolski et al., 2016). Caribbean reefs also have been changing in coral composition. Species of genus Agaricia spp. and Porites spp. tend to dominate the seascape (Perry et al., 2018). This scenario (Cozumel—Mexico) can be an opportunity for experiments to test diverse ecological functions.
Another characteristic habitat in the Mexican Caribbean region is the largely acroporid-dominated coral nurseries such as the current ex situ restoration sites of Wave of Change (Cozumel and Riviera Maya). This habitat has the recurrent prevalence of predators such as fireworm Hermodice carunculata that have a highly negative impact on populations of A. cervicornis and A. palmata (Calle-Triviño et al., 2017). These reefscapes can be benefited by the supply of fish such as white grunts (Haemulon plumierii) and sand tilefish (Malacanthus plumieri) (Ladd and Shantz, 2016). Coral growth can be improved by adding fishes such as grunts (Haemulidae) around coral nurseries or outplanting sites via delivery of fish-derived nutrients (Shantz et al., 2015) or via concentrate grazing by Sparisoma sp. and Acathurus sp. (Shantz et al., 2017; Calle-Triviño et al., 2021). Despite, the highest contributions of grunts and herbivorous fishes (surgeonfishes and parrotfishes) in both study sites our results suggest that there is a good chance of using these species to enhance research and may be considered in the context of restoration both structurally and functionally.
The primary method employed here can be used to broaden and predict the taxonomic composition and distribution of postlarval fishes. Also, it can promote the capture, culture, and release of reef fish with a sustainable approach, especially in enforcement and management sites across the Caribbean region. The results obtained are very promising in terms of species richness, diversity, abundance, and CPUE as well as an innovative way to drive restoration of coral reef services and functions. This sampling method provides the benefit of increasing the productivity of target species, for example, commercial and herbivorous fishes (Bell et al., 2009). These efforts can contribute to identifying settlement areas for reef fish, biodiversity monitoring (McLeod and Costello, 2017), managing of MPAs (Obolski et al., 2016), and supporting fisheries control through research and social engagement (Hein et al., 2020; Cortés-Useche et al., 2021). They can also support the restoration efforts of reefs that have suffered a loss in the resilience of their fish biodiversity (Lorenzen et al., 2010) or re-establish the provisioning services delivered by reefs in providing habitat and nursery areas for commercially (Hein et al., 2020) and functionally important species. In this sense, future work could be focused to improve sampling biodiversity and broadening monitoring variables that have an influence on recruitment such as luminosity, pH, algal blooms, wave exposure, rugosity, etc.
The pilot test (S. partitus) showed very promising results on survivorship relative to the settlement stage ex situ. This method tested here can be set up for a variety of fish species and seems to be a feasible restoration tool, to increase the benefits of management through the effective implementation that includes long-term ecological and economic synergies (Lirman and Schopmeyer, 2016; Cortés-Useche et al., 2019). Key aspects, such as fisheries policy and water quality treatment in the context of climate change and managing the connectivity of the tropical coastal reefscapes should be considered (Arias-González et al., 2016; Hein et al., 2020; Schmidt-Roach et al., 2020; Cortés-Useche et al., 2021).
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding authors.
Ethics Statement
The animal study was reviewed and approved by The Secretary of the Environment and Natural Resources approved field collection (PPF/DGOPA-282, DGOPA-DAPA-11811).
Author Contributions
CC-U, JA-G, JC-P, and WR-G conceived and designed the experiments, performed the experiments, analyzed the data, prepared figures and/or tables, authored or reviewed drafts of the paper, and approved the final draft. RY and JC-T analyzed the data, prepared figures and/or tables, authored or reviewed drafts of the paper, and approved the final draft. AC-F conceived and designed the experiments, performed the experiments, authored or reviewed drafts of the paper, and approved the final draft. RR-F performed the experiments, authored or reviewed drafts of the paper, and approved the final draft. All authors contributed to the article and approved the submitted version.
Funding
Part of this research was supported by National Problems project from the National Council for Science and Technology (CONACyT) (PDCPN/2017/5747). CC-U and WR-G were supported by grants provided by CONACyT (29240 and 637330). CC-U was supported with a Post-doctoral fellowship grant provided by CONACyT (PDCPN/2017/5747).
Conflict of Interest
AC-F and RR-F were employed by Acuario XCARET.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
The authors would like to thank the Xcaret Archeo-Ecological Park for the support provided in aquariums, field work and facilities; Coral Reefs Ecosystems Ecology Laboratory of the Center for Research and Advanced Studies - Campus Mérida (CINVESTAV-IPN); and Alex Neufeld, Special Projects Coordinator of Coral Restoration Foundation for his technical support with the Photomosaics. The authors also thank the National Council for Science and Technology (CONACyT) for their valuable contribution (PDCPN/2017/5747) and the National Commission of Natural Protected Areas (CONANP) for permission to work within the MPAs (CNANP-00- 008/00-010). The Secretary of the Environment and Natural Resources approved field collection (PPF/DGOPA-282/17, DGOPA-DAPA-11811). We thank the Iberostar Foundation for covering the purchase of Agisoft Metashape Professional Edition software and the publication fees for free access to this manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2021.718526/full#supplementary-material
References
Abelson, A., Obolski, U., Regoniel, P., and Hadany, L. (2016). Restocking herbivorous fish populations as a social-ecological restoration tool in coral reefs. Front. Mar. Sci. 3:138. doi: 10.3389/fmars.2016.00138
Almany, G. R., and Webster, M. S. (2006). The predation gauntlet: early post-settlement mortality in reef fishes. Coral Reefs 25, 19–22. doi: 10.1007/s00338-005-0044-y
Álvarez-Cadena, J. N., Ordóñez-López, U., Almaral-Mendivil, A. R., Ornelas-Roa, M., and Uicab-Sabido, A. (2007). Larvas de peces del litoral arrecifal del norte de Quintana Roo, Mar Caribe de México. Hidrobiológica 17, 139–150.
Arias-González, J. E., Fung, T., Seymour, R. M., Garza-Pérez, J. R., Acosta-González, G., Bozec, Y.-M., et al. (2017). A coral-algal phase shift in Mesoamerica not driven by changes in herbivorous fish abundance. PLOS ONE 12:e0174855. doi: 10.1371/journal.pone.0174855
Arias-González, J. E., Rivera-Sosa, A., Zaldívar-Rae, J., Alva-Basurto, C., and Cortés-Useche, C. (2016). “The animal forest and its socio-ecological connections to land and coastal ecosystems,” in Marine Animal Forests, eds S. Rossi, L. Bramanti, A. Gori, and C. Orejas (Cham: Springer), 1209–1240. doi: 10.1007/978-3-319-17001-5_33-1
Ayala-Campos, M. (2014). Riqueza específica y abundancia en el reclutamiento de post-larvas de peces arrecifales del Caribe mexicano en zonas aledañas al parque ecológico Xcaret, Quintana Roo, México (Master's thesis). Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México, Mexico City, Mexico.
Bailey, K. M., and Houde, E. D. (1989). Predation on eggs and larvae of marine fishes and the recruitment problem. Adv. Mar. Biol. 25, 1–83. doi: 10.1016/S0065-2881(08)60187-X
Bayraktarov, E., Banaszak, A. T., Maya, P. M., Kleypas, J., Arias-González, J. E., Blanco, M., et al. (2020). Coral reef restoration efforts in Latin American countries and territories. bioRxiv. doi: 10.1101/2020.02.16.950998
Bell, J. D., Clua, E., Hair, C. A., Galzin, R., and Doherty, P. J. (2009). The capture and culture of post-larval fish and invertebrates for the marine ornamental trade. Rev. Fish. Sci. 17, 223–240. doi: 10.1080/10641260802528541
Calle-Triviño, J., Cortés-Useche, C., Sellares, R., and Arias González, J. E. (2017). First record of the fireworm Hermodice carunculata preying on colonies of the threatened staghorn coral Acropora Cervicornis in the southeastern outplanting sites of the Dominican Republic. Novitates Caribaea 11, 97–98. doi: 10.33800/nc.v0i11.17
Calle-Triviño, J., Cortés-Useche, C., Sellares-Blasco, R. I., and Arias-González, J. E. (2018). Assisted fertilization of threatened Staghorn Coral to complement the restoration of nurseries in Southeastern Dominican Republic. Reg. Stud. Mar. Sci. 18, 129–134. doi: 10.1016/j.rsma.2018.02.002
Calle-Triviño, J., Muñiz-Castillo, A. I., Cortés-Useche, C., Morikawa, M., Sellares-Blasco, R., and Arias-González, J. E. (2021). Approach to the functional importance of Acropora cervicornis in outplanting sites in the Dominican Republic. Front. Mar. Sci. 8:668325. doi: 10.3389/fmars.2021.668325
Calle-Triviño, J., Rivera-Madrid, R., León-Pech, M. G., Cortés-Useche, C., Sellares-Blasco, R. I., Aguilar-Espinosa, M., et al. (2020). Assessing and genotyping threatened staghorn coral Acropora cervicornis nurseries during restoration in southeast Dominican Republic. PeerJ 8:e8863. doi: 10.7717/peerj.8863
Cervigón, F., Cipriani, R., Fischer, W., Garibaldi, L., Hendrickx, M., and Lemus, A. J. (1992). “Guía de Campo de las Especies Comerciales Marinas y de Aguas Salobres de la Costa Septentrional de Sur America,” in FAO Species Identification Field Guide for Fishery Purposes (Rome: FAO), 530. Available online at: www.fao.org/docrep/010/t0544e/t0544e00.htm
Choat, J. H., Doherty, P. J., Kerrigan, B. A., and Leis, J. M. (1983). Sampling of larvae and pelagic stages of coral reef fishes: a comparison of towed nets, purse seine, and light aggregation devices. Fish. Bull. U.S. 91, 195–201.
Cortés-Useche, C., Calle-Triviño, J., Sellares-Blasco, R., Luis-Báez, A., and Arias-González, J. E. (2018). An updated checklist of the reef fishes of the Southeastern Reefs Marine sanctuary of the Dominican Republic. Revista mexicana de biodiversidad 89, 382–392. doi: 10.22201/ib.20078706e.2018.2.2149
Cortés-Useche, C., Hernández-Delgado, E. A., Calle-Triviño, J., Sellares Blasco, R., Galván, V., and Arias-González, J. E. (2021). Conservation actions and ecological context: optimizing coral reef local management in the Dominican Republic. PeerJ 9:e10925. doi: 10.7717/peerj.10925
Cortés-Useche, C., Muñiz-Castillo, A. I., Calle-Triviño, J., Yathiraj, R., and Arias-González, J. E. (2019). Reef condition and protection of coral diversity and evolutionary history in the marine protected areas of Southeastern Dominican Republic. Reg. Stud. Mar. Sci. 32:100893. doi: 10.1016/j.rsma.2019.100893
D'Alessandro, E., Sponaugle, S., and Lee, T. (2007). Patterns and processes of larval fish supply to the coral reefs of the upper Florida keys. Mar. Ecol. Prog. Ser. 331, 85–100. doi: 10.3354/meps331085
Del Moral-Flores, L. F., Tello-Musi, J. L., Reyes-Bonilla, H., Pérez-España, H., Martínez-Pérez, J. A., Horta-Puga, G., et al. (2013). Lista sistemática y afinidades zoogeográficas de la ictiofauna del Sistema Arrecifal Veracruzano, Mexico. Revista Mexicana de Biodiversidad 84, 825–846. doi: 10.7550/rmb.34912
Doherty, P., and McIlwain, J. (1996). Monitoring larval fluxes through the surf zones of Australian coral reefs. Mar. Freshw. Res. 47, 383–390. doi: 10.1071/MF9960383
Doherty, P. J. (1987). Light-traps: selective but useful devices for quantifying the distributions and abundances of larval fishes. Bull. Mar. Sci. 41, 423–431.
Doherty, P. J. (2002). “Variable replenishment and the dynamics of reef fish populations,” in Coral Reef Fishes: Dynamics and Diversity in a Complex System, ed P. F. Sale (San Diego: Academic Press), 327–355. doi: 10.1016/B978-012615185-5/50019-0
Doherty, P. J., Dufour, V., Galzin, R., Hixon, M. A., Meekan, M. G., and Planes, S. (2004). High mortality during settlement is a population bottleneck for a tropical surgeonfish. Ecology 85, 2422–2428. doi: 10.1890/04-0366
Doropoulos, C., Vons, F., Elzinga, J., ter Hofstede, R., Salee, K., van Koningsveld, M., et al. (2019). Testing industrial-scale coral restoration techniques: harvesting and culturing wild coral-spawn slicks. Front. Mar. Sci. 6:658. doi: 10.3389/fmars.2019.00658
Duarte, C. M., Agusti, S., Barbier, E., Britten, G. L., Castilla, J. C., Gattuso, J. P., et al. (2020). Rebuilding marine life. Nature 580, 39–51. doi: 10.1038/s41586-020-2146-7
Dufour, V. (1994). Colonization of fish larvae in lagoons of Rangiroa (Tuamotu Archipelago) and Moorea (Society Archipelago). Atoll Res. Bull. 416, 1–8. doi: 10.5479/si.00775630.416.1
Dufour, V., and Galzin, R. (1993). Colonization patterns of reef fish larvae to the lagoon at Moorea Island, French Polynesia. Mar. Ecol. Prog. Ser. 102, 143–152. doi: 10.3354/meps102143
Dufour, V., Riclet, E., and Lo-Yat, A. (1996). Colonization of reef flat fishes at Moorea Island, French Polynesia: temporal and spatial variation of the larval flux. Mar. Freshw. Res. 47, 413–422. doi: 10.1071/MF9960413
Durville, P., Bosc, P., Galzin, R., and Conand, C. (2003). Aquacultural suitability of post-larval coral reef fish. SPC Live Reef Fish Info. Bull. 11, 18–30.
Ecocean (2020). The Post-Larval Capture: A Comprehensive Range of PCC Fishing and Rearing Devices. Available online at: https://www.ecocean.fr/fishing-rearing-devices/ (accessed May 20, 2020).
Froese, R., and Pauly, D. (2019). Fishbase. World Wide Web Electronic Publication. Available online at: http://www.fishbase.org/ (accessed December 13, 2019).
Green, A. L., and Bellwood, D. R. (2009). “Monitoring functional groups of herbivorous reef fishes as indicators of coral reef resilience – A practical guide for coral reef managers in the Asia Pacific region,” in IUCN Working Group on Climate Change and Coral Reefs (Gland: IUCN), 70.
Hair, C., Bell, J., and Doherty, P. (2002). “The use of wild-caught juveniles in coastal aquaculture and its application to coral reef fishes,” in Responsible Marine Aquaculture, eds R. R. Stickney, and J. P. McVey (CABI), 327–353. doi: 10.1079/9780851996042.0327
Heenan, A., Simpson, S. D., Meekan, M. G., Healy, S. D., and Braithwaite, V. A. (2009). Restoring depleted coral-reef fish populations through recruitment enhancement: a proof of concept. Fish Biol. 75, 1857–1867. doi: 10.1111/j.1095-8649.2009.02401.x
Hein, M. Y., McLeod, I. M, Shaver, E. C., Vardi, T., Pioch, S., Boström-Einarsson, L., et al. (2020). Coral Reef Restoration as a Strategy to Improve Ecosystem Services – A Guide to Coral Restoration Methods. Nairobi: United Nations Environment Program.
Hendriks, I., Wilson, D., and Meekan, M. (2001). Vertical distributions of late stage larval fishes in the nearshore waters of the San Blas Archipelago, Caribbean Panama. Coral Reefs 20, 77–84. doi: 10.1007/s003380100139
Humann, P., and DeLoach, N. (2002). Reef Fish Identification: Florida, Caribbean, Bahamas, 3th Edn. Jacksonville: New World Publications.
Jost, L. (2007). Partitioning diversity into independent alpha and beta components. Ecology 88, 2427–2439. doi: 10.1890/06-1736.1
Kingsford, M. J., Leis, J. M., Shanks, A., Lindeman, K. C., Morgan, S. G., and Pineda, J. (2002). Sensory environments, larval abilities and local self-recruitment. Bull. Mar. Sci. 70, 309–340.
Ladd, M. C., and Andrew, A. S. (2020). Trophic interactions in coral reef restoration: a review. Food Webs 4, 2352–2496. doi: 10.1016/j.fooweb.2020.e00149
Ladd, M. C., Miller, M. W., Hunt, J. H., Sharp, W. C., and Burkepile, D. E. (2018). Harnessing ecological processes to facilitate coral restoration. Front. Ecol. Environ. 16, 239–247. doi: 10.1002/fee.1792
Ladd, M. C., and Shantz, A. A. (2016). Novel enemies–previously unknown predators of the bearded fireworm. Front. Ecol. Environ. 14, 342–343. doi: 10.1002/fee.1305
Lecaillon, G. (2004). The “C.A.R.E.” (collect by artificial reef eco-friendly) system as a method of producing farmed marine animals for the aquarium market: An alternative solution to collection in the wild. SPC Live Reef Fish Inf. Bull. 12, 17–20.
Lecaillon, G. (2017). “Post-larval capture and culture of ornamental fishes,” in Marine Ornamental Species Aquaculture, eds R. Calado, I. Olivotto, M. P. Oliver, and G. J. Holt (Hoboken, NJ: John Wiley & Sons). doi: 10.1002/9781119169147.ch18
Lecaillon, G., and Lourié, S. M. (2007). Current status of marine post-larval collection: existing tools, initial results, market opportunities and prospects. SPC Live Reef Fish Inf. Bull. 17, 3–10.
Legendre, P., and De Cáceres, M. (2013). Beta diversity as the variance of community data: dissimilarity coefficients and partitioning. Ecol. Lett. 16, 951–963. doi: 10.1111/ele.12141
Leis, J. M., and McCormick, M. I. (2002). “The biology, behavior, and ecology of the pelagic, larval stage of coral reef fishes,” in Coral Reef Fishes: Dynamic and Diversity in a Complex Ecosystem, ed P. F. Sale (San Diego: Academic Press), 171–199. doi: 10.1016/B978-012615185-5/50011-6
Lirman, D., and Schopmeyer, S. (2016). Ecological solutions to reef degradation: optimizing coral reef restoration in the Caribbean and Western Atlantic. PeerJ 4:e2597. doi: 10.7717/peerj.2597
Lorenzen, K., Leber, K. M., and Blankenship, H. L. (2010). Responsible approach to marine stock enhancement: an update. Rev. Fish. Sci. 18, 189–210. doi: 10.1080/10641262.2010.491564
McCormick, M., Makey, L., and Dufour, V. (2002). Comparative study of meta- morphosis in tropical reef fishes. Mar. Biol. 141, 841–853. doi: 10.1007/s00227-002-0883-9
McLeod, L. E., and Costello, M. J. (2017). Light traps for sampling marine biodiversity. Helgoland Mar. Res. 71:2. doi: 10.1186/s10152-017-0483-1
Milicich, M. J., and Doherty, P. J. (1994). Larval supply of coral reef fish populations: magnitude and synchrony of replenishment to Lizard Island, Great Barrier Reef. Mar. Ecol. Prog. Ser. 110, 121–134. doi: 10.3354/meps110121
Moana Initiative (2007). PCC, a useful method for conserving value and biodiversity. Foundation for Marine Biodiversity. MAB/Unesco and CRISP project. Available online at: http://www.moanainitiative.org
Obolski, U., Hadany, L., and Abelson, A. (2016). Potential contribution of fish restocking to the recovery of deteriorated coral reefs: an alternative restoration method? PeerJ 4:e1732. doi: 10.7717/peerj.1732
Paddack, M. J., and Sponaugle, S. (2008). Recruitment and habitat selection of newly settled Sparisoma viride to reefs with low coral cover. Mar. Ecol. Prog. Ser. 369, 205–212. doi: 10.3354/meps07632
Perry, C. T., Alvarez-Filip, L., Graham, N. A. J., Mumby, P. J., Wilson, S. K., Kench, P. S., et al. (2018). Loss of coral reef growth capacity to track future increases in sea level. Nature 558, 396–400. doi: 10.1038/s41586-018-0194-z
Planes, S., and Lecaillon, G. (2001). Caging experiment to examine mortality during metamorphosis of coral reef fish larvae. Coral Reefs 20, 211–218. doi: 10.1007/s003380100161
Planes, S., Lecaillon, G., Lenfant, P., and Meekan, M. (2002). Genetic and demographic variation in new recruits of Naso unicornis. J. Fish Biol. 61, 1033–1049. doi: 10.1111/j.1095-8649.2002.tb01861.x
Precht, W. F., Aronson, R. B., Moody, R. M., and Kaufman, L. (2010). Changing patterns of microhabitat utilization by the threespot damselfish, Stegastes planifrons, on Caribbean Reefs. PLoS ONE 5:e10835. doi: 10.1371/journal.pone.0010835
R Core Team (2017). R: A language and environment for statistical computing. (3.3.1). Vienna: R Foundation for Statistical Computing. Available online at: https://www.r-project.org/
Rankin, T. L. (2010). The Effects of Early Life History on Recruitment and Early Juvenile Survival of a Coral Reef Fish in the Florida Keys. Coral Gables, FL: University of Miami. Available online at: https://scholarship.miami.edu/discovery/fulldisplay/alma991031447343802976/01UOML_INST:ResearchRepository (accessed May 12, 2010).
Sale, P. (2015). “The future for coral reef fishes,” in Ecology of Fishes on Coral Reefs, ed C. Mora (Cambridge: Cambridge University Press), 283–288. doi: 10.1017/CBO9781316105412.037
Schmidt-Roach, S., Duarte, C. M., Hauser, C. A. E., and Aranda, M. (2020). Beyond reef restoration: next-generation techniques for coral gardening, landscaping, and outreach. Front. Mar. Sci. 7:672. doi: 10.3389/fmars.2020.00672
Schopmeyer, S. A., and Lirman, D. (2015). Occupation dynamics and impacts of damselfish territoriality on recovering populations of the threatened staghorn coral, Acropora cervicornis. PLoS ONE 10:e0141302. doi: 10.1371/journal.pone.0141302
Shantz, A., Mark, C., Ladd, D., and Burkepile, E. (2017). Algal nitrogen and phosphorus content drive inter- and intraspecific differences in herbivore grazing on a Caribbean reef. J. Exp. Mar. Biol. Ecol. 497, 164–171. doi: 10.1016/j.jembe.2017.09.020
Shantz, A. A., Ladd, M. C., Schrack, E., and Burkepile, D. E. (2015). Fish- derived nutrient hotspots shape coral reef benthic communities. Ecol. Appl. 25, 2142–2152. doi: 10.1890/14-2209.1
Simpson, S. D., Piercy, J. J. B., King, J., and Codling, E. A. (2013). Modelling larval dispersal and behaviour of coral reef fishes. Ecol. Complexity 16, 68–76. doi: 10.1016/j.ecocom.2013.08.001
Sponaugle, S., Boulay, J. N., and Rankin, T. L. (2011). Growth- and size-selectivity mortality in pelagic larvae of common reef fish. Aquatic Biol. 13, 263–273. doi: 10.3354/ab00370
Sponaugle, S., Grorud-Colvert, K., and Pinkard, D. (2006). Temperature-mediated variation in early life history traits and recruitment success of the coral reef fish Thalassoma bifasciatum in the Florida Keys. Mar. Ecol. Prog. Ser. 308, 1–15. doi: 10.3354/meps308001
Steele, M. A., and Forrester, G. E. (2002). Early postsettlement predation on three reef fishes: effects on spatial patterns of recruitment. Ecology 83, 1076–1091. doi: 10.1890/0012-9658(2002)083<1076:EPPOTR>2.0.CO;2
Sweatman, H. (1988). Field evidence that settling coral reef fish larvae detect resident fishes using dissolved chemical cues. J. Exp. Mar. Biol. Ecol. 124, 163–174. doi: 10.1016/0022-0981(88)90170-0
Thiessen, R. J., and Heath, D. D. (2006). Characterization of one trinucleotide and six dinucleotide microsatellite markers in bicolor damselfish, Stegastes partitus, a common coral reef fish. Conserv. Genet. 8, 983–985. doi: 10.1007/s10592-006-9207-9
Vadziutsina, M., and Riera, R. (2020). Review of fish trap fisheries from tropical and subtropical reefs: main features, threats and management solutions. Fish. Res. 223:105432. doi: 10.1016/j.fishres.2019.105432
Vallès, H., Kramer, D. L., and Hunte, W. (2008). Temporal and spatial patterns in the recruitment of coral-reef fishes in Barbados. Mar. Ecol. Prog. Ser. 363, 257–272. doi: 10.3354/meps07432
Vásquez-Yeomans, L., Ordoñez-López, U., Quintal-Lizama, C., and Ornelas-Roa, M. (2003). A preliminary fish larvae survey in Banco Chinchorro. Bull. Mar. Sci. 73, 141–152.
Vásquez-Yeomans, L., Ordoñez-López, U., and Sosa-Cordero, E. (1998). Fish larvae to a coral reef in the western Caribbean sea off Mahahual, Mexico. Bull. Mar. Sci. 62, 229–245.
Vásquez-Yeomans, L., Vega-Cendejas, M. E., Montero, J. L., and Sosa-Cordero, E. (2011). High species richness of early stages of fish in a locality of the Mesoamerican Barrier Reef System: a small-scale survey using different sampling gears. Biodivers. Conserv. 20, 2379–2392. doi: 10.1007/s10531-011-9990-6
Victor, B. C. (1991). “Settlement strategies and biogeography of reef fishes,” in The Ecology of Fishes on Coral Reefs, ed P. Sale (San Diego: Academic Press), 231–260. doi: 10.1016/B978-0-08-092551-6.50014-3
Victor, B. C., and Wellington, G. M. (2000). Endemism and the pelagic duration of reef fishes in the eastern Pacific Ocean. Mar. Ecol. Prog. Ser. 205, 241–248. doi: 10.3354/meps205241
Villegas-Sánchez, C. A., Abitia-Cárdenas, L. A., Gutiérrez-Sánchez, F. J., and Galván-Magaña, F. (2009). Asociaciones de peces de arrecifes rocosos en Isla San José, México. Revista Mexicana de Biodiversidad 80, 169–179. doi: 10.22201/ib.20078706e.2009.001.594
Watson, M., Munro, J. L., and Gell, F. R. (2002). Settlement, movement and early juvenile mortality of the yellowtail snapper Ocyurus chrysurus. Mar. Ecol. Prog. Ser. 237, 247–256. doi: 10.3354/meps237247
Webster, M. S. (2002). Role of predators in the early post-settlement demography of coral-reef fishes. Oecologia 131, 52–60. doi: 10.1007/s00442-001-0860-x
Keywords: active restoration, biodiversity, Caribbean, coral reef, fish restocking, recovery
Citation: Cortés-Useche C, Reyes-Gamboa W, Cabrera-Pérez JL, Calle-Triviño J, Cerón-Flores A, Raigoza-Figueras R, Yathiraj R and Arias-González JE (2021) Capture, Culture and Release of Postlarvae Fishes: Proof-of-Concept as a Tool Approach to Support Reef Management. Front. Mar. Sci. 8:718526. doi: 10.3389/fmars.2021.718526
Received: 01 June 2021; Accepted: 20 August 2021;
Published: 20 September 2021.
Edited by:
Wei Jiang, Guangxi University, ChinaReviewed by:
Alex E. Mercado-Molina, Sociedad Ambiente Marino, Puerto RicoEvan D'Alessandro, University of Miami, United States
Copyright © 2021 Cortés-Useche, Reyes-Gamboa, Cabrera-Pérez, Calle-Triviño, Cerón-Flores, Raigoza-Figueras, Yathiraj and Arias-González. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Camilo Cortés-Useche, camilo.cortes@cinvestav.mx; Jesús Ernesto Arias-González, earias@cinvestav.mx