
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Mar. Sci., 02 June 2021
Sec. Marine Conservation and Sustainability
Volume 8 - 2021 | https://doi.org/10.3389/fmars.2021.659579
This article is part of the Research TopicImpacts on Neotropical Diversity: A Call for More Integrative Coastal ManagementView all 4 articles
 Ryan Andrades1,2,3*
Ryan Andrades1,2,3* Jean-Christophe Joyeux1
Jean-Christophe Joyeux1 Raphael M. Macieira1Bruno Spacek Godoy4
Raphael M. Macieira1Bruno Spacek Godoy4 José Amorim Reis-Filho3Andrew L. Jackson2
José Amorim Reis-Filho3Andrew L. Jackson2 Tommaso Giarrizzo3
Tommaso Giarrizzo3Niche-related processes (e.g., density or niche-breadth compensation and competition) are fundamental to a broad understanding of community ecology and ecosystem functioning. Most evidences of competition are from controlled indoor trials with few species, and it remains a challenge to estimate competition among multiple species in the field. Here, we analyze stable isotopes and distributional data from 51 fish taxa in six locations in the southwestern Atlantic to predict intraspecific trophic pressure (ITP) and the potential competitive strength among species in a trophic-based framework. We used two proxies built upon 2-dimensional isotopic space (δ13C vs. δ15N), its predicted overlap, and fish density to calculate winner and loser taxa in potential paired interspecific competitive interactions. The intraspecific proxy indicated that cryptobenthic fishes are under high among-individual trophic pressure (high densities and small niche sizes). Also, cryptobenthic behavior together with feeding specialization and extremely small-sizes were the most important traits related to low success in interspecific simulations. Although cryptobenthic fishes face strong competitive pressures, there are some known inherent trade-offs to cryptobenthic life such as trophic and habitat use specializations. These seem to compensate and ensure coexistence among cryptobenthic fishes and non-cryptobenthic species. Habitat loss/degradation via urbanization, invasive species and climate-change-driven sea-level rise can reduce the suitability of habitat and increase competition on cryptobenthic species, especially in shallow reefs and intertidal shores.
Competition is one of the most important driving forces shaping life forms, from microbes to large-sized plants and animals (Gause, 1934; Simberloff, 1982; Connell, 1983; Xavier and Foster, 2007; Bonin et al., 2015). The understanding of how the struggle for space and resources occurs drew Darwin’s attention on the mechanisms involved in the coexistence and contests within and among species (Darwin, 1859). In his seminal book Darwin dedicated one chapter to the discussion of the mechanisms behind one species becoming victorious over another and on the role of natural selection in a competition spectrum (Darwin, 1859). Since then, several studies on the issue have argued over ways and means to assess competitive interactions (Lotka, 1925; Volterra, 1926; Gause, 1934; Hutchinson, 1961; MacArthur and Levins, 1967; Connell, 1983; Schoener, 1983; Simberloff, 1983; Huisman and Weissing, 1994; Tilman, 1994) but these processes remain laborious to measure, especially in a speciose framework.
Competition is directly influenced by life-history modes in plants, invertebrates and vertebrates. In many cases, complex life-history traits (e.g., pelagic larval duration, food acquisition modes, size of early recruits) may define the settlement and adaptation success of species facing potential competition (Schoener, 1968; Osenberg, 1989; Olden et al., 2006; Andrades et al., 2019a). Historically, the search for estimates of niche overlap (on a spatial or trophic basis) have shed light on competition measurements and supported the majority of ecological inferences regarding potential competition assessments (Hutchinson, 1961, 1978; MacArthur, 1968; Bonin et al., 2015).
More recently, stable isotopes-based Bayesian estimates have allowed the measurement of the Hutchinsonian niche components in an isotopic space (mainly δ13C vs. δ15N) called the isotopic niche and ultimately represented by ellipses (Newsome et al., 2007; Jackson et al., 2011). Since then, isotopic models have been used to explore niche-relationships in terrestrial, freshwater and marine environments through the quantification of the overlap, or partitioning, of trophic niches among species. For instance, isotopic niche size and overlap among individuals of small mammals in the Queen Elizabeth national park (Uganda) lead to the conclusion that individual trophic specialization emerges from increased intragroup competition (Sheppard et al., 2018). Similarly, ichthyologists evaluate that on Caribbean reefs the invasive lionfishes (Pterois spp.) compete more directly with a native endangered grouper (O’Farrell et al., 2014). In intertidal ecosystems, stable isotope analysis has been useful to detect the intraspecific partitioning of the dietary niche in fish species and has clarified the niche expansion phenomenon driven by competition release in island communities (Andrades et al., 2019a,b). Thus, stable isotope models provide reliable assessments of niche-relationships, including potential competition, mainly for those cases hard to observe or measure in nature.
An historically good model to investigate resource sharing and competition in nature is species occupancy in intertidal shores (rocky and biogenic reefs). The zonation under different pressure gradients (e.g., exposure to air, predation and temperature) and limiting living space revealed strong competition strategies and complex coexistence modes for gastropods, echinoderms and algae (Connell, 1961; Paine, 1969, 1974; Underwood, 1984; Metaxas and Scheibling, 1993; Vaz-Pinto et al., 2014) and inspired Robert Paine to coin the “keystone species” concept that has enlightened the role of predators in the maintenance of species diversity balance (Paine, 1966). However, most studies focused on sessile or slow-moving species (Metaxas and Scheibling, 1993) and, in the case of fishes, were mostly based on indoor experiments using one or few species simultaneously (Szabo, 2002; Arakaki and Tokeshi, 2011, 2012; Paijmans and Wong, 2017; Colella et al., 2019). Intertidal reef pools host fishes that exhibit diverse physiological, morphological and behavioral adaptations to the stressful life “between the tides” (Gibson and Yoshiyama, 1999; Martin and Bridges, 1999; Zander, 2011). Cryptobenthic fishes are a remarkably abundant fish-group that is typically composed of small-sized species strongly associated to the bottom and that present rich taxonomic and functional aspects (Brandl et al., 2018). In space-limited tidepools, high populational and specific density and elevated energetic demands would put these fishes into a particularly high intra- and interspecific competitive framework. Because pools are isolated from the sea during low tides they offer the possibility to acquire robust density data to be merged with resource and habitat use information. Thus, the appraisal of winner and loser species in potential competitive interactions and the relationships among species intrinsic individual-traits can allow the detection of vulnerable taxa (potentially inferior competitors) in a multi-species framework built upon scenario of increased competition intermediated by habitat loss/degradation through human impacts.
Here, we examine six fish assemblages in oceanic and coastal intertidal reefs to investigate which species traits are associated with competitive success under distinct scenarios. For this, we use stable isotopes δ13C and δ15N signatures and fish density data to assess intraspecific trophic pressure (ITP) and simulate interspecific competition. Further, we determine what species are the competitors most prone to deleterious effect in pairwise simulated competition.
Fifteen tidepools were sampled in each of six tropical intertidal reef sites (N = 90) within Brazilian waters. Three sites were located in oceanic islands: (i) Rocas Atoll (03°51′ S; 33°49′ W), the only atoll in the South Atlantic (266 km off coast) built primarily by coralline algae, vermetid gastropods and hermatypic corals (Gherardi and Bosence, 2001); (ii) Fernando de Noronha Archipelago (03°50′ S; 32°25′ W), 345 km from mainland, where rocky intertidal reefs are often associated with biogenic carbonate substrate composed by encrusting coralline algae; and (iii) Trindade Island (20°30′ S; 29°20′ W), the most isolated insular environment within Brazilian territory (1,160 km off coast) sheltering biogenic and phonolitic rocky reefs. Trindade has the highest reef fish endemism rate (9.6%) among the Brazilian islands (Pinheiro et al., 2015). In these sites, tidepool water temperature and salinity often varies from 27°C to 35°C and from 35 to 41, respectively (Andrades et al., 2018b). The coastal intertidal reefs in our study were: (i) Salinópolis (Farol Beach; 00°36′ S; 47°21′ W) located in northern region of Brazil where coastline are dominated by mangrove forests and estuaries, despite the occurrence of rocky outcrops. Farol Beach comprises a sandy beach with sparse lateritic and biogenic intertidal reefs with tidepool water temperature and salinity varying from 31°C to 35°C and 36 to 41, respectively (Andrades et al., 2018b); (ii) Jericoacoara (02°47′ S; 40°30′ W) located in northeastern Brazil where quartzite beachrocks makes of the intertidal reefs. Water temperature and salinity in tidepools varied from 24°C to 32°C and 35 to 41, respectively (Machado et al., 2015; Andrades et al., 2018b); and (iii) Anchieta (Castelhanos Beach; 20°49′ S; 40°36′ W) located in the southeastern region of Brazil with a flat intertidal reef composed mainly by encrusting coralline algae and stony coral skeletons with tidepool water temperature and salinity varying from 24°C to 35°C and 36 to 41, respectively (Macieira and Joyeux, 2011; Andrades et al., 2018b).
Fish were sampled in field expeditions performed during the dry seasons of 2014 (Rocas Atoll) and 2015 (the other sites). At each site, fifteen tidepools were chosen at random during the ebb tide providing there was no connectivity to the sea or to other pools. Specimens were caught using hand nets after application of the anesthetic clove oil (40 mg⋅l–1 in ethanol), an efficient and selective method not inducing mortality in non-target fauna (Griffiths, 2000). Concomitantly to fish sampling, the topography and bathymetry of the tidepools were surveyed using a 10 cm × 10 cm grid for the measurement of the depth at each point of tidepool. Then, the tidepool volume (m3) was calculated by kriging. At each site, non-systematic (extensive) fish collections in tidepools other than those surveyed were carried out to increase the sample size for stable isotope analysis (see below) and to collect species not encountered during systematic sampling. After collection, individuals were frozen at −20°C in the laboratory, where abundance, total length and wet weight measurements were taken. For stable isotopes, muscle tissue samples were removed from the anterio-lateral portion of the trunk, except for small-sized fish. Individuals with a total length under 30 mm (e.g., Tomicodon sp.), were used whole after removal of viscera, scales, fins, and head. All samples were dried in a standard laboratory oven at 60°C during 48 h, and then homogenized using a pestle and mortar.
Fish samples (ca. 0.5 mg ground tissues) from systematic and extensive collections were analyzed for stable isotope ratios of carbon and nitrogen using continuous-flow isotope ratio mass spectrometry in a Costech elemental analyzer coupled to a Micromass Isoprime isotope ratio mass spectrometer (EA/IRMS). The laboratory data were calibrated using NIST (National Institute of Standards and Technology) reference materials, PDB (Pee Dee Belemnite) for carbon and airborne N2 for nitrogen. Stable isotope values are expressed using δ (delta) notation and parts per thousand (‰) as follows:
where X is 13C or 15N and R = 13C/12C for carbon and 15N/14N for nitrogen.
In order to avoid any bias resulting from chemical interference in the isotopic signatures, we did not extract lipids using chemical solvents, such as methanol-chloroform (Post et al., 2007; Boecklen et al., 2011). Given this, consumers δ13C signatures were corrected for lipid-rich samples (C:N > 3.5), following Post et al. (2007). Also, in order to avoid bias caused by ontogeny (Andrades et al., 2019a), whenever possible variation in body size was constrained among sites both within species and congeneric species (Andrades et al., 2019b).
Isotopic niche spaces (95% prediction ellipse area) occupied by species were estimated from stable isotope ratios (δ13C and δ15N) using the multivariate ellipse-based model “SIBER” (Stable Isotope Bayesian Ellipses in R; Jackson et al., 2011). Mean values of δ13C and δ15N, as well as niche areas are available in Supplementary Table 1. To verify potential niche overlap among species we calculated the overlap of the maximum likelihood fitted standard ellipses using the function maxLikOverlap based on 95% prediction ellipse area.
We set seven traits for intertidal reef fishes based on information provided in literature for intertidal reef fish ecology (Gibson, 1982; Gibson and Yoshiyama, 1999; Griffiths, 2003; Andrades et al., 2018a,b), these were endemism, residency level, trophic group, level of competition within trophic group, habitat association on tidepool, cryptobenthic behavior, and body size. All species and trait categorization and information are in Supplementary Table 2.
A proxy of the intensity of intraspecific trophic interaction pressure was determined by dividing the density of each fish species by its isotopic niche width, as follows:
where FDi is the transformed (log+1) mean fish density (ind.m–3) of species i and SEAc is the small sample size cored (i.e., core isotopic niche ellipses) of species i, containing the 95% prediction ellipse interval. This simple approach represents the density of a population in relation to the size of its isotopic niche size, with higher ITP values representing a greater degree of ITP in the species population. Higher ITP implies an abundant species with limited resource use, which a scenario of habitat loss and limited resource availability may pose a threat to the maintenance of population energetic needs. For these analyses we included all fish taxa that had their density estimated during the systematic tidepool surveys and had, at least, three individuals for isotopic niche width estimates.
We quantified species competition proxy (SCP) considering the amount of overlap in the isotopic niche between two species in relation to the density of the focal species. Specifically, we calculated:
where Ovij is the overlap of the area of the corrected standard ellipse between species i and j that corresponds to the overlap of ellipse area. We were thus able to measure the effect of species i on species j and vice versa through:
where αij is the effect of species j on i. We only evaluated potential competition effects for pairs of species that presented some degree of overlap of their small sample size cored 95% prediction ellipses, otherwise, SCPi would be equal to FDi, which is meaningless. Based on the competitive interactions between species and the number of potential competitors, we identified the most successful competitors in each studied assemblage, that is, the species that had the greatest net potential effect in the pairwise comparisons, and, in turn, the species most vulnerable to potential competitive interactions (i.e., weak competitors). Only fish species collected in systematic samplings were included in the analysis since stable isotope signatures and niche area were estimated. Species that were only collected during the extensive surveys (i.e., not sampled in systematic surveys, and thus not considered in density estimates) were a priori considered losers when in potential competitive interaction with those (which overlapped niche areas) from systematic surveys.
Then, we computed the competitive success rate (%) of each species of the six studied sites and classified taxa as strong competitors (>66% success in paired simulated scenarios), regular competitors (33–66% success) and weak competitors (<33% success). Thus, we built a classification tree using log-linear models from the function loglin and “rpart” package (Therneau and Atkinson, 2018) to identify the best response variables associated to the predictor, where the competitive success rate was the dependent variable and the species traits defined a priori were the independent variables. The tree was pruned using the complexity parameter value to minimize the cross-validation error (Breiman et al., 1984; De’ath and Fabricius, 2000; Borcard et al., 2018). Also, a confusion matrix was extracted to inform on the classification accuracy and the Kappa Index of Agreement was used to evaluate the proportion of correct classification predicted at a rate higher than expected (Cohen, 1960). The statistical analyses were ran in R software (R Core Team, 2017).
Altogether, 51 fish taxa from the six sites were evaluated for ITP. The magnitude of ITP is represented as the percentage of each taxon ITP proxy in relation to sympatric taxa in each site (Figure 1). Overall, weak competitors (red-colored) were small-sized taxa belonging to typical cryptobenthic families (Blenniidae, Gobiidae, Labrisomidae, and Gobiesocidae).
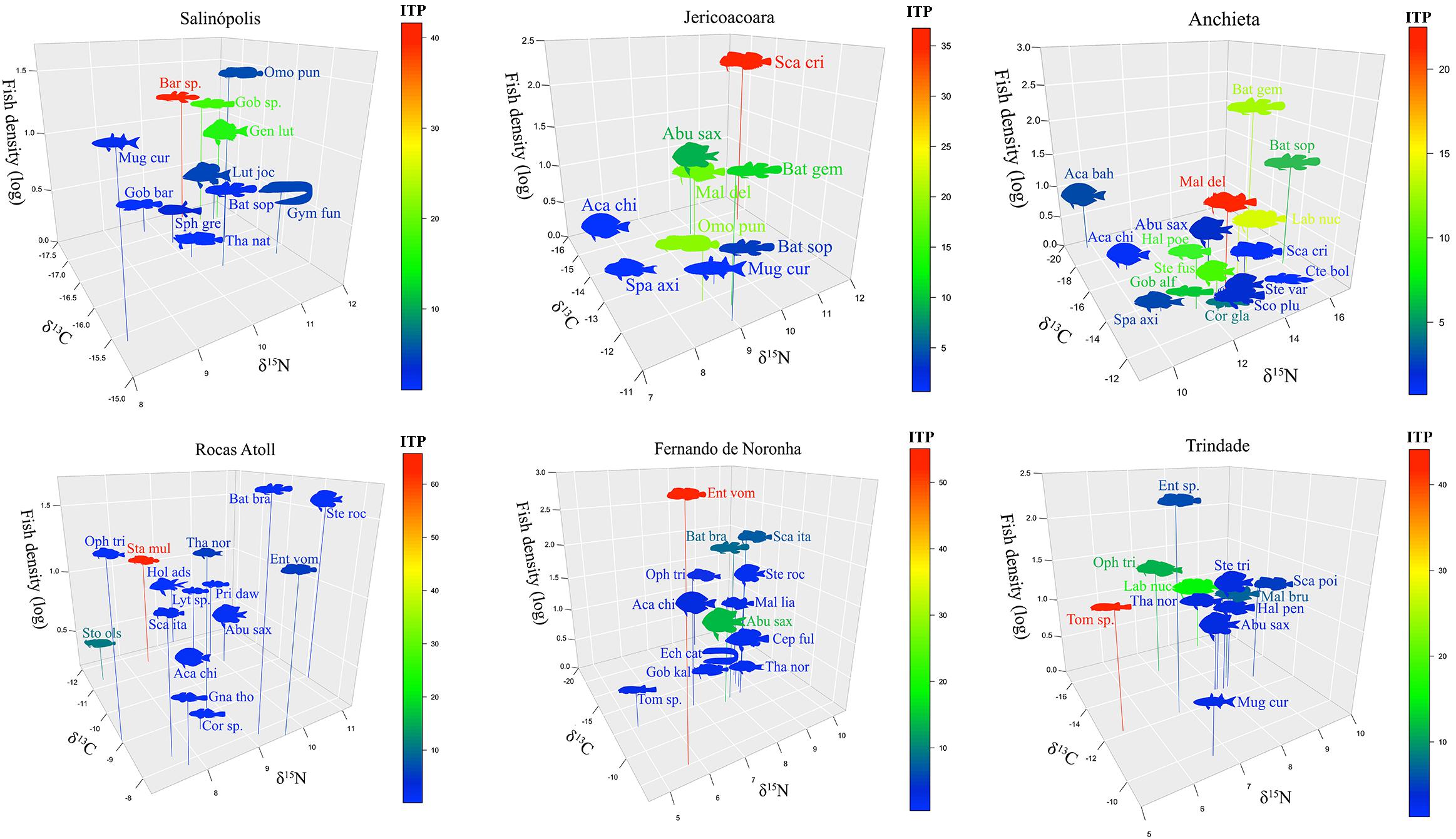
Figure 1. Intraspecific trophic pressure (ITP; fish silhouette color) in 3-D space defined from isotopic niche (δ13C and δ15N signatures on the x and y axes, respectively) and density (ind.m–3; z-axis) in six intertidal environments. The color scale indicates the relative ITP intensity at each site (low intensity = coldest color and high intensity = warmest color). Taxa acronyms are given Supplementary Table 1.
The evaluation of interspecific interactions using the SCP indicated that the potentially inferior competitors were Coryphopterus sp., Starksia multilepis, and Storrsia olsoni at Rocas, Abudefduf saxatilis, Gobioclinus kalisherae, and Entomacrodus vomerinus at Noronha, Malacoctenus brunoi, Tomicodon sp. and Mugil curvidens at Trindade (Supplementary Figure 1), Gobiesox barbatulus, Sphoeroides testudineus and Thalassophryne nattereri at Salinópolis, Omobranchus punctatus, Acanthurus chirurgus and Sparisoma axillare at Jericoacoara, and Sparisoma axillare, Stegastes variabilis, and Coryphopterus glaucofraenum at Anchieta (Supplementary Figure 2).
Best-explanation tree model for SCP data included three traits: trophic group, body size and cryptobenthic behavior (R2 = 0.22; root node error 0.60, Figure 2). The importance of these predictors in defining the tree structure was 38, 23, and 14%, respectively. Based on the confusion matrix (Supplementary Table 3), the percentage of the predicted response was 53.42 % (46.58 % of confusion) and the average kappa index was 0.27, an acceptable agreement. The primary split (1) separated the strongly competitive omnivore species from weakly competitive specialist species (carnivores, herbivores and micro-carnivores). The second node (3) splitted species in respect to their size and the subsequent split (6) in relation to their cryptic habit. Weak competitors (node 13) were cryptobenthic species of small (e.g., Gobiosoma alfiei and Tomicodon sp.) and large (e.g., Labrisomus nuchipinnis) sizes.
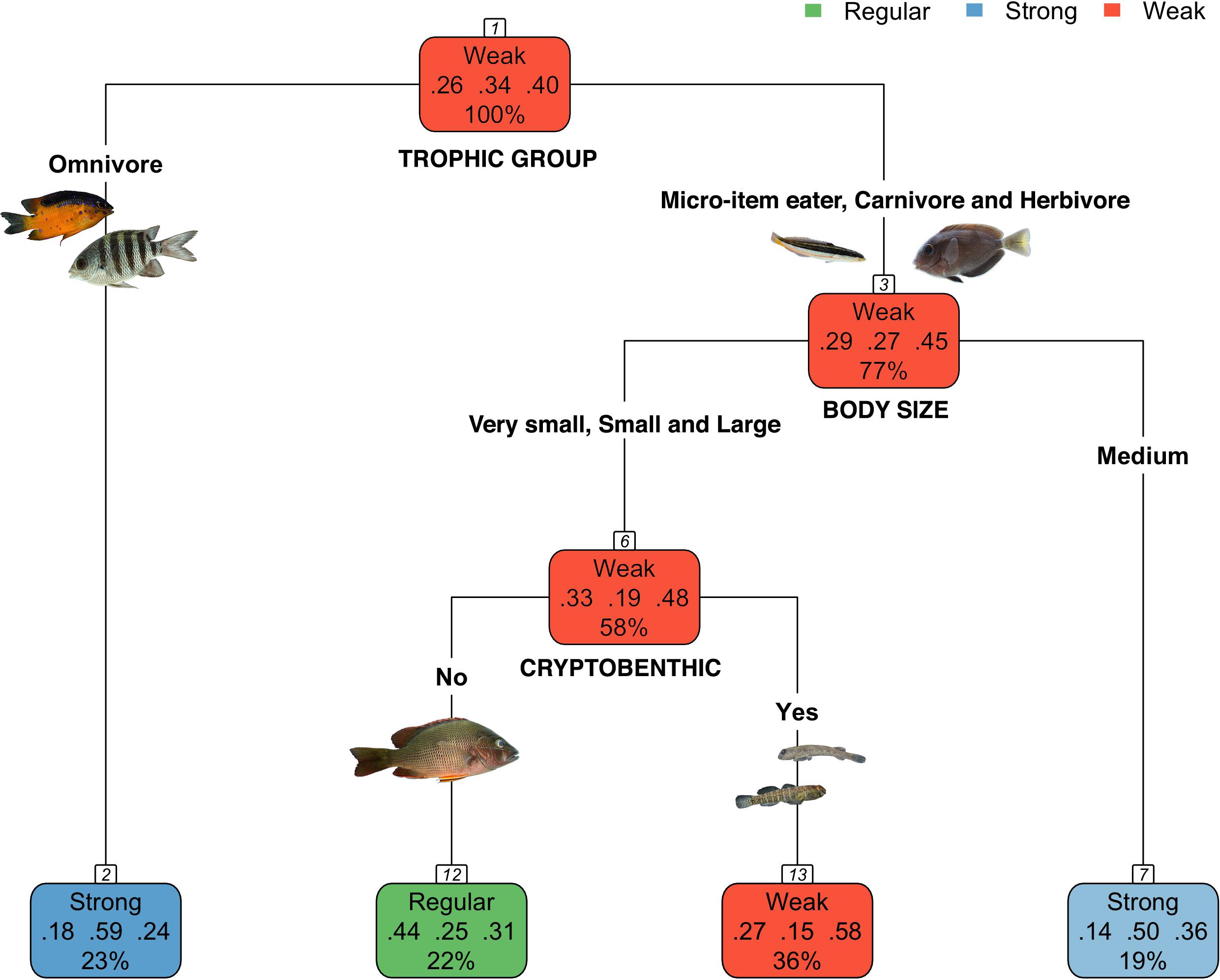
Figure 2. The four-leaf tree predicting the traits related to interspecific competition success of fish taxa. Each node (color boxes) depicts the predicted class (regular, strong, or weak competition classes), its probabilities (from left to right: regular, strong, and weak) and the percentage of observations in the node. Error was 0.77 and cross-validation error was 0.90.
We have shown that a diverse group of abundant species, the feeding-specialists of cryptobenthic behavior, is extremely vulnerable to interspecific competition and presents high rates of relative ITP for resources. Here, isotopic niche overlap between species was used to infer on potential interspecific competition. Although classical and contemporary approaches of niche overlap agree with this interpretation (Hutchinson, 1978; Eckrich et al., 2020), stable isotopes have limitations and how much niche overlap means to competition per se needs to be cautiously considered. Consumers with similar δ15N signatures can feed upon different preys and similar δ13C does not reflect obligatorily that both consumers are relying on the same resources. In fact, one of the main stable isotope analysis constraints is the lack of taxonomic refinement in potential prey identification (Newsome et al., 2007). On the other hand, competition can also exist between species whose realized niches do not overlap (Holt, 1987). Yet, in a space-limited habitat, such as intertidal reefs, potential competition among species cannot be underestimated, even among species belonging to different trophic guilds. For instance, typical territorial herbivorous reef fishes (e.g., Stegastes spp.) are able to compete for food with several other species acting as omnivorous (Zamprogno, 1989; Pimentel et al., 2018), whereas resident gobies (e.g., Bathygobius spp.) expected to be carnivorous often consume also large amounts of algae (Zamprogno, 1989; Feitosa and Araújo, 2002). Despite its caveats, stable isotopes analysis overcomes the complexity of determining all competitive interactions and the accurate quantification of all axes of the niches hypervolumes. Our study is underpinned by evidences and case studies where isotopic niche overlap was considered a reliable and robust estimate of potential resource sharing or partitioning through the realized niches of species or individuals (Jackson et al., 2012; Fleming et al., 2015; Storero et al., 2020). Thus, isotopic niche overlap is closely linked to competition and furnishes plausible competition estimates (Colwell and Futuyma, 1971; Pianka, 2000; Sexton et al., 2017; Pastore et al., 2021).
The intrinsic biological resilience of the cryptobenthic species is associated with competition trade-offs and demographic dynamics. Very shallow reefs (<5 m depth) are stressful environments in which conditions widely fluctuate within the physicochemical (e.g., hydrodynamic and water salinity) and biological (e.g., primary and secondary production) spectra. It is expected that under high environmental variability rough competition scenarios may be relieved by competition trade-offs (Hutchinson, 1961; Duthie et al., 2014). In fact, such mechanisms can be observed from our intertidal assemblages. Rockpool blennies Entomacrodus spp. under high intra- and interspecific competitions in oceanic islands (mainly Noronha – Figure 1) persist as key-structuring species by occupying almost exclusively isolated low-complexity tidepools of the high-shore (Andrades et al., 2018b). This displacement is so marked that very dense groups of Entomacrodus (Figure 3A) are common, even in small-sized pools. Possibly, Entomacrodus spp., a few small invertebrates and Tomicodon spp. converged toward this underutilized niche as it supports basal-resource exploitation while – essentially – persisting during the island existence (Macieira et al., 2015; Pinheiro et al., 2017; Andrades et al., 2019b). The stronger trophic pressure rates recorded for Noronha than Trindade endemic Entomacrodus can be explained by the abundant and speciose fish communities (Andrades et al., 2018b). These features are probably consequence of the larger intertidal area (ca. 60 km vs. 22 km total perimeter) and older origin of Noronha (∼12.5 Ma vs. ∼3.9 Ma; see Mohriak, 2020).

Figure 3. Cryptobenthic intertidal reef fish of the South Atlantic Ocean. A dozen of Entomacrodus vomerinus lumped in a small tidepool (A). A solitary Tomicodon sp. in a rare moment out of a sea urchin crevice (B). Starksia multilepis within macroalgal intertices in an intertidal reef (C) and detail of the small mouth of 2 cm sized individual (D). Juvenile and adult of Labrisomus nuchipinnis (E) often inhabit intertidal reefs, but presented high niche partitioning of habitat and resource uses. The predator Echidna catenata (F).
The only other sympatric fish that densely occupies the high-shore are Tomicodon spp. (Figure 3B; the species on the mainland, Trindade and Noronha/Rocas are distinct and only the first has been described, see Briggs, 1955; Macieira et al., 2015). These miniature taxa are often found in the surge zone, in small crevices or dead bivalve shells, under boulders or sea urchins (Karplus, 2014). Microhabitat specialization and competition release effects (Andrades et al., 2018b) allow higher abundance of Entomacrodus spp. and Tomicodon spp. in islands compared to mainland (Andrades et al., 2018a), agreeing to the MacArthur et al. (1972) “density compensation” hypothesis. Yet, as intraspecific competition increases in response to density compensation (Crowell, 1962; Pafilis et al., 2009), a certain balance is probably maintained by species’ capability to occupy micro-niches.
Other species under intense intra- and interspecific competition is the manyscaled blenny S. multilepis (Figure 3C), mainly in Rocas. This algae and anemone dweller inhabits rocky interstices. Its small mouth (Figure 3D) enables it to pick micro-items from algal turfs and fronds and its cryptic behavior favors avoidance of direct competition with stronger competitors. In fact, Starksia species presents high species richness at the regional-scale in the Caribbean (Baldwin et al., 2011), which is probably sustained by high levels of microhabitat partitioning among conspecifics through ecological speciation at small spatial scale (Rocha et al., 2005; Rocha and Bowen, 2008).
In relatively large-bodied intertidal reef specialists, ontogenetic niche shifts can drive microhabitat and dietary segregation to facilitate coexistence. The juvenile and adult of the cryptobenthic hairy blenny L. nuchipinnis (Figure 3E) act as two ecologically distinct species: they display distinct habitat occupancy, feeding modes and functional traits in order to maximize their food intake and reduce predation, including cannibalism by adults (Andrades et al., 2019a). Also, it is imperative to note that though the conventional definition of cryptobenthic species takes into account the small-sized nature of the species (Brandl et al., 2018), morays like the chain moray E. catenata (Figure 3F) have a cryptobenthic nature while also being a top predator (see Andrades et al., 2019b). Such species perform highly vulnerable functions in reef environments due to their rarity and specific functional traits (nocturnal-feeding, sedentary) (Mouillot et al., 2013), but for few species daytime foraging also occurs (Sazima and Sazima, 2004). This justifies and brings attention to the presence of large vulnerable species in intertidal reefs (Figure 2).
Overall, cryptobenthic micro-specialists compensate their restricted niche, short-life span and high mortality with high reproductive rate and high larval input (Depczynski and Bellwood, 2006; Lefèvre et al., 2016). Brandl et al. (2019) estimate that this input is responsible to fueling reef environments with almost 60% of the fish biomass consumed, illustrating both this group vulnerability and its resilience to density-dependent processes such as intra- and interspecific competition.
Although cryptobenthic fish species can be found from the intertidal to mesophotic depths (<150 m) (Tornabene et al., 2016; Brandl et al., 2018), in our study many vulnerable taxa were restricted to the intertidal layer of only a few centimeters to depths of less than 2 m. For instance Tomicodon sp. (Trindade), S. multilepis and S. olsoni (Rocas), E. vomerinus (Noronha) and Barbulifer sp. (Salinópolis) (see Figure 1 and Supplementary Figures 1, 2) are under high selective competition pressures, despite their importance to the intertidal food chain (Andrades et al., 2019b). In a general competitive framework, an increase in the use of resources by populations under high intraspecific pressure is expected (Giller, 1984; Svanbäck and Bolnick, 2007), a feature that would be especially relevant in species with limited capacity for niche adaptation. Therefore, intertidal cryptobenthic species having ecological traits (e.g., small body size or a particular habitat) that a priori limit trophic flexibility (e.g., exploitation of larger prey) or habitat change (e.g., toward greater depth) would be under strong competition. It should be noted, however, that miniaturization facilitates singular spatial and trophic micro-niches occupancy among fishes (Miller, 1996; Brandl et al., 2018).
The idea that competitive interactions are responsible for biodiversity maintenance is well recognized. For the tropical marine fauna competition and its consequences may be the most important speciation processes in space-limited reefs (Schemske et al., 2009; Brown, 2014). Presumably, over the evolutionary time many tropical lineages have arisen, or at least presented high net diversification, to use low-quality diets instead of outcompeting strong competitors over high-quality resources (Harmelin-Vivien, 2002; Lobato et al., 2014). However, strong competition does not allow inferring detrimental effects over (expected) inferior competitors. Actually, niche overlap per se does not necessarily translate into interference effects but only in shared requirements over resources. No evidence for interference process limiting some species was detected in the present study but a general rule was clearly manifested: more-efficient specialists probably emerge to compensate the overpressure of generalists. In fact, while fine-grained experiment studies can demonstrate the immediate effects of competitive interactions they often are limited to a few species and local patterns. The model chosen for the regression tree presented a satisfactory result to estimate the competitive success of fish. However, other elements than those we used to build the model may have high relevancy and could have contributed to increase the observed fit. Some functional traits not included, because not sampled or considered secondary, may not be irrelevant to interactions. Further studies are needed to ascertain whether the insertion of other life-history traits (e.g., size classes and resource availability for each taxa or fish-group) can improve the fit of theoretical models. Finally, we used data from six locations, so regional differences in environmental characteristics may drive the importance of interactions between species in the communities. Further studies must consider, in addition to isotopic niche overlap, quantify prey-specific contribution through mixing models to best infer on potential overlap or partitioning of resources among consumers.
The costs of living under high competition pressure involve many deleterious effects such as low reproductive fitness and high energetic allocation in food searching, contest behavior and predation avoidance (Briffa and Sneddon, 2010; Bonin et al., 2015; Forrester, 2015). Here, using stable isotopes allied to density data of more than 50 reef fish taxa, we demonstrated that high intra- and interspecific competition levels are centered over small-sized specialists, mostly belonging to cryptobenthic families. We also discussed a number of mechanisms that facilitate the coexistence of cryptobenthic fishes in an extremely fluctuating and competitive environment such as intertidal reefs. Nevertheless, anthropogenic impacts (e.g., biological invasion, urbanization, sea-level rise) can propel these taxa into risky scenarios of competition increase. Further studies must focus on fine-scale niche partitioning among taxa in order to enlighten the trophic plasticity and selective process upon target resources.
The data analyzed in this study is subject to the following licenses/restrictions: The dataset will be published as part of a data paper soon. Requests to access these datasets should be directed to RA, cnlhbmFuZHJhZGVzQGdtYWlsLmNvbQ==.
The animal study was reviewed and approved by the UFES’ Ethics Committee (CEUA-UFES; Protocol 29/2016). All specimen collection was licensed by Instituto Chico Mendes de Conservação da Biodiversidade/MMA – Brazilian Ministry of the Environment, through the SISBIO system.
RA, J-CJ, RM, and TG designed the study. RA, J-CJ, JR-F, RM, and TG collected the data. RA, BG, AJ, and TG developed the analysis. RA analyzed the data and wrote the manuscript. All authors revised and contributed to the final version.
This project was funded by the Espírito Santo State Research and Innovation Foundation (FAPES, 67504876) and Trindade Island Research Program (CNPq 405426/2012-7). RA received a Ph.D. scholarship from CAPES-DS and CAPES-PDSE (Edital 19/2016, process 88881.132520/2016-01). This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) – Finance Code 001. J-CJ was supported by FAPES (J-CJ – Edital 007/2014, process 67504876) and CNPQ (441243/2016-9). TG and J-CJ received productivity grants from the National Council for Scientific and Technological Development (CNPq 311078/2019-2 and 304274/2016-0).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We are grateful to T. Simon (in memoriam), H. Guabiroba, G. Cardozo-Ferreira, M. E. Alves, L. Mendonça, R. F. Martins, J. Andrade, M. Bolzan, F. Machado, C. Sales, A. Porrua, L. Karvat, H. Entringer, C. Albuquerque, J. Dantas, M. Brito-Silva, and R. G. Santos for field and laboratory assistance. The field surveys were supported logistically by Maurizélia Brito Silva, ICMBio, SOS Mata Atlântica, Wagner Cardoso, the Brazilian Navy (Marinha do Brasil), and S. Costa-Abrantes.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2021.659579/full#supplementary-material
Andrades, R., Andrade, J. M., Jesus-Junior, P. S., Macieira, R. M., Bernardino, A. F., Giarrizzo, T., et al. (2019a). Multiple niche-based analyses reveal the dual life of an intertidal reef predator. Mar. Ecol. Prog. Ser. 624, 131–141. doi: 10.3354/meps13027
Andrades, R., Jackson, A. L., Macieira, R. M., Reis-filho, J. A., Bernardino, A. F., Joyeux, J.-C., et al. (2019b). Niche-related processes in island intertidal communities inferred from stable isotopes data. Ecol. Indic. 104, 648–658. doi: 10.1016/j.ecolind.2019.05.039
Andrades, R., Machado, F. S., Reis-Filho, J. A., Macieira, R. M., and Giarrizzo, T. (2018a). Intertidal biogeographic subprovinces: local and regional factors shaping fish assemblages. Front. Mar. Sci. 5:412. doi: 10.3389/fmars.2018.00412
Andrades, R., Reis-Filho, J. A., Macieira, R. M., Giarrizzo, T., and Joyeux, J.-C. (2018b). Endemic fish species structuring oceanic intertidal reef assemblages. Sci. Rep. 8:10791. doi: 10.1038/s41598-018-29088-0
Arakaki, S., and Tokeshi, M. (2011). Analysis of spatial niche structure in coexisting tidepool fishes: null models based on multi-scale experiments. J. Anim. Ecol. 80, 137–147. doi: 10.1111/j.1365-2656.2010.01749.x
Arakaki, S., and Tokeshi, M. (2012). Species and size matter: an experimental study of microhabitat use under the influence of competitive interactions in intertidal gobiids. J. Exp. Mar. Bio. Ecol. 418–419, 59–68. doi: 10.1016/j.jembe.2012.03.011
Baldwin, C., Castillo, C., Weigt, L., and Victor, B. (2011). Seven new species within western Atlantic Starksia atlantica, S. lepicoelia, and S. sluiteri (Teleostei, Labrisomidae), with comments on congruence of DNA barcodes and species. Zookeys 79, 21–72. doi: 10.3897/zookeys.79.1045
Boecklen, W. J., Yarnes, C. T., Cook, B. A., and James, A. C. (2011). On the use of stable isotopes in trophic ecology. Annu. Rev. Ecol. Evol. Syst. 42, 411–440. doi: 10.1146/annurev-ecolsys-102209-144726
Bonin, M. C., Boström-Einarsson, L., Munday, P. L., and Jones, G. P. (2015). The prevalence and importance of competition among coral reef fishes. Annu. Rev. Ecol. Evol. Syst. 46, 169–190. doi: 10.1146/annurev-ecolsys-112414-054413
Borcard, D., Gillet, F., and Legendre, P. (2018). Numerical Ecology with R. Cham: Springer International Publishing, doi: 10.1007/978-3-319-71404-2
Brandl, S. J., Goatley, C. H. R., Bellwood, D. R., and Tornabene, L. (2018). The hidden half: ecology and evolution of cryptobenthic fishes on coral reefs. Biol. Rev. 93, 1846–1873. doi: 10.1111/brv.12423
Brandl, S. J., Tornabene, L., Goatley, C. H. R., Casey, J. M., Morais, R. A., Côté, I. M., et al. (2019). Demographic dynamics of the smallest marine vertebrates fuel coral reef ecosystem functioning. Science 364, 1189–1192. doi: 10.1126/science.aav3384
Breiman, L., Friedman, J. H., Olshen, R. A., and Stone, C. J. (1984). Classification and Regression Trees. Boca Raton, FL: Chapman & Hall/CRC.
Briffa, M., and Sneddon, L. U. (2010). “Contest behavior,” in Evolutionary Behavioral Ecology, eds D. F. Westneat and C. W. Fox (Oxford: Oxford University Press), 246–265.
Briggs, J. C. (1955). A monograph of the clingfishes (Order Xenopterygii). Stanford Ichthyol. Bull. 6, 1–224.
Brown, J. H. (2014). Why are there so many species in the tropics? J. Biogeogr. 41, 8–22. doi: 10.1111/jbi.12228
Cohen, J. (1960). A coefficient of agreement for nominal scales. Educ. Psychol. Meas. 20, 37–46. doi: 10.1177/001316446002000104
Colella, D. J., Paijmans, K. C., and Wong, M. Y. L. (2019). Size, sex and social experience: experimental tests of multiple factors mediating contest behaviour in a rockpool fish. Ethology 125, 369–379. doi: 10.1111/eth.12861
Colwell, R. K., and Futuyma, D. J. (1971). On the measurement of niche breadth and overlap. Ecology 52, 567–576. doi: 10.2307/1934144
Connell, J. H. (1961). The influence of interspecific competition and other factors on the distribution of the barnacle Chthamalus stellatus. Ecology 42, 710–723. doi: 10.2307/1933500
Connell, J. H. (1983). On the prevalence and relative importance of interspecific competition: evidence from field experiments. Am. Nat. 122, 661–696. doi: 10.1086/284165
Crowell, K. L. (1962). Reduced interspecific competition among the birds of Bermuda. Ecology 43, 75–88. doi: 10.2307/1932042
Darwin, C. (1859). On the Origin of Species by Means of Natural Selection, or, the Preservation of Favoured Races in the Struggle for Life. London: John Murray.
De’ath, G., and Fabricius, K. E. (2000). Classification and regression trees: a powerful yet simple technique for ecological data analysis. Ecology 81, 3178–3192. doi: 10.1890/0012-9658(2000)081[3178:cartap]2.0.co;2
Depczynski, M., and Bellwood, D. R. (2006). Extremes, plasticity, and invariance in vertebrate life history traits: insights from coral reef fishes. Ecology 87, 3119–3127. doi: 10.1890/0012-9658(2006)87[3119:epaiiv]2.0.co;2
Duthie, A. B., Abbott, K. C., and Nason, J. D. (2014). Trade-offs and coexistence: a lottery model applied to fig wasp communities. Am. Nat. 183, 826–841. doi: 10.1086/675897
Eckrich, C. A., Albeke, S. E., Flaherty, E. A., Bowyer, R. T., and Ben-David, M. (2020). rKIN: kernel-based method for estimating isotopic niche size and overlap. J. Anim. Ecol. 89, 757–771. doi: 10.1111/1365-2656.13159
Feitosa, C. V., and Araújo, M. E. (2002). Hábito alimentar e morfologia do trato digestivo de alguns peixes de poças de maré, no estado do Ceará, Brasil. Arq. Ciências do Mar. 35, 97–105. doi: 10.32360/acmar.v35i1-2.30911
Fleming, N. E. C., Harrod, C., Newton, J., and Houghton, J. D. R. (2015). Not all jellyfish are equal: isotopic evidence for inter- and intraspecific variation in jellyfish trophic ecology. PeerJ 2015, 1–21. doi: 10.7717/peerj.1110
Forrester, G. E. (2015). “Competition in reef fishes,” in Ecology of Fishes on Coral Reefs, ed. C. Mora (Cambridge: Cambridge University Press), 34–40. doi: 10.1017/CBO9781316105412.006
Gherardi, D. F. M., and Bosence, D. W. J. (2001). Composition and community structure of the coralline algal reefs from Atol das Rocas, South Atlantic, Brazil. Coral Reefs 19, 205–219. doi: 10.1007/s003380000100
Gibson, R. N. (1982). Recent studies on the biology of intertidal fishes. Oceanogr. Mar. Biol. 20, 363–414.
Gibson, R. N., and Yoshiyama, R. M. (1999). “Intertidal fish communities,” in Intertidal Fishes: Life in Two Worlds, eds M. M. Horn, K. L. M. Martin, and M. A. Chotkowski (Cambridge, MA: Academic Press), 264–296. doi: 10.1016/b978-012356040-7/50014-7
Griffiths, S. P. (2000). The use of clove oil as an anaesthetic and method for sampling intertidal rockpool fishes. J. Fish Biol. 57, 1453–1464. doi: 10.1006/jfbi.2000.1406
Griffiths, S. P. (2003). Rockpool ichthyofaunas of temperate Australia: species composition, residency and biogeographic patterns. Estuar. Coast. Shelf Sci. 58, 173–186. doi: 10.1016/S0272-7714(03)00073-8
Harmelin-Vivien, M. L. (2002). “Energetics and fish diversity on coral reefs,” in Coral Reef Fishes: Dynamics and Diversity on a Complex Ecosystem, ed. P. F. Sale (San Diego, CA: Academic Press), 265–274. doi: 10.1016/b978-012615185-5/50016-5
Holt, R. D. (1987). On the relation between niche overlap and competition: the effect of incommensurable niche dimensions. Oikos 48:110. doi: 10.2307/3565696
Huisman, J., and Weissing, F. J. (1994). Light-limited growth and competition for light in well-mixed aquatic environments: an elementary model. Ecology 75, 507–520. doi: 10.2307/1939554
Hutchinson, G. E. (1978). An Introduction to Population Ecology. New Haven, CT: Yale University Press.
Jackson, A. L., Inger, R., Parnell, A. C., and Bearhop, S. (2011). Comparing isotopic niche widths among and within communities: SIBER–stable isotope Bayesian Ellipses in R. J. Anim. Ecol. 80, 595–602. doi: 10.1111/j.1365-2656.2011.01806.x
Jackson, M. C., Donohue, I., Jackson, A. L., Britton, J. R., Harper, D. M., and Grey, J. (2012). Population-level metrics of trophic structure based on stable isotopes and their application to invasion ecology. PLoS One 7:e31757. doi: 10.1371/journal.pone.0031757
Lefèvre, C. D., Nash, K. L., González-Cabello, A., and Bellwood, D. R. (2016). Consequences of extreme life history traits on population persistence: do short-lived gobies face demographic bottlenecks? Coral Reefs 35, 399–409. doi: 10.1007/s00338-016-1406-3
Lobato, F. L., Barneche, D. R., Siqueira, A. C., Liedke, A. M. R., Lindner, A., Pie, M. R., et al. (2014). Diet and diversification in the evolution of coral reef fishes. PLoS One 9:e102094. doi: 10.1371/journal.pone.0102094
MacArthur, R., and Levins, R. (1967). The limiting similarity, convergence, and divergence of coexisting species. Am. Nat. 101, 377–385. doi: 10.1086/282505
MacArthur, R. H., Diamond, J. M., and Karr, J. R. (1972). Density compensation in island faunas. Ecology 53, 330–342. doi: 10.2307/1934090
Machado, F. S., Macieira, R. M., Gómez, M. A. Z., Costa, A. F., Mesquita, E. M. C., and Giarrizzo, T. (2015). Checklist of tidepool fishes from Jericoacoara National Park, southwestern Atlantic, with additional ecological information. Biota Neotrop. 15, 1–9.
Macieira, R. M., and Joyeux, J.-C. (2011). Distribution patterns of tidepool fishes on a tropical flat reef. Fish. Bull. 109, 305–315.
Macieira, R. M., Simon, T., Pimentel, C. R., and Joyeux, J.-C. (2015). Isolation and speciation of tidepool fishes as a consequence of Quaternary sea-level fluctuations. Environ. Biol. Fishes 98, 385–393. doi: 10.1007/s10641-014-0269-0
Martin, K. L. M., and Bridges, C. R. (1999). “Respiration in water an air,” in Intertidal Fishes: Life in Two Worlds, eds M. M. Horn, K. L. M. Martin, and M. A. Chotkowski (Cambridge, MA: Academic Press), 54–78. doi: 10.1016/b978-012356040-7/50005-6
Metaxas, A., and Scheibling, R. E. (1993). Community structure and organization of tidepools. Mar. Ecol. Prog. Ser. 98, 187–198. doi: 10.3354/meps098187
Miller, P. J. (1996). “The functional ecology of small fish: some opportunities and consequences,” in Miniature Vertebrates: the Implications of Small Body Size, ed. P. J. Miller (Oxford: Oxford University Press), 175–199.
Mohriak, W. (2020). Genesis and evolution of the South Atlantic volcanic islands offshore Brazil. Geo-Mar. Lett. 40, 1–33. doi: 10.1007/s00367-019-00631-w
Mouillot, D., Bellwood, D. R., Baraloto, C., Chave, J., Galzin, R., Harmelin-Vivien, M., et al. (2013). Rare species support vulnerable functions in high-diversity ecosystems. PLoS Biol. 11:e1001569. doi: 10.1371/journal.pbio.1001569
Newsome, S. D., Martínez del Rio, C., Bearhop, S., and Phillips, D. L. (2007). A niche for isotope ecology. Front. Ecol. Environ. 5:429–436. doi: 10.1890/060150.01
O’Farrell, S., Bearhop, S., McGill, R. A. R., Dahlgren, C. P., Brumbaugh, D. R., and Mumby, P. J. (2014). Habitat and body size effects on the isotopic niche space of invasive lionfish and endangered Nassau grouper. Ecosphere 5:art123. doi: 10.1890/ES14-00126.1
Olden, J. D., Leroy Poff, N., and Bestgen, K. R. (2006). Life-history strategies predict fish invasions and extirpations in the Colorado River Basin. Ecol. Monogr. 76, 25–40. doi: 10.1890/05-0330
Osenberg, C. W. (1989). Resource limitation, competition and the influence of life history in a freshwater snail community. Oecologia 79, 512–519. doi: 10.1007/BF00378669
Pafilis, P., Meiri, S., Foufopoulos, J., and Valakos, E. (2009). Intraspecific competition and high food availability are associated with insular gigantism in a lizard. Naturwissenschaften 96, 1107–1113. doi: 10.1007/s00114-009-0564-3
Paijmans, K. C., and Wong, M. Y. L. (2017). Linking animal contests and community structure using rockpool fishes as a model system. Funct. Ecol. 31, 1612–1623. doi: 10.1111/1365-2435.12859
Paine, R. T. (1966). Food web complexity and species diversity. Am. Nat. 100, 65–75. doi: 10.1086/282400
Paine, R. T. (1969). The Pisaster-Tegula interaction: prey patches, predator food preference, and intertidal community structure. Ecology 50, 950–961. doi: 10.2307/1936888
Paine, R. T. (1974). Intertidal community structure: experimental studies on the relantionship between a dominant competitor and its principal predator. Oecologia 15, 93–120. doi: 10.1007/BF00345739
Pastore, A. I., Barabás, G., Bimler, M. D., Mayfield, M. M., and Miller, T. E. (2021). The evolution of niche overlap and competitive differences. Nat. Ecol. Evol. 5, 330–337. doi: 10.1038/s41559-020-01383-y
Pimentel, C. R., Soares, L. S. H., Macieira, R. M., and Joyeux, J.-C. (2018). Trophic relationships in tidepool fish assemblages of the tropical Southwestern Atlantic. Mar. Ecol. 39:e12496. doi: 10.1111/maec.12496
Pinheiro, H. T., Bernardi, G., Simon, T., Joyeux, J.-C., Macieira, R. M., Gasparini, J. L., et al. (2017). Island biogeography of marine organisms. Nature 549, 82–85. doi: 10.1038/nature23680
Pinheiro, H. T., Mazzei, E., Moura, R. L., Amado-Filho, G. M., Carvalho-Filho, A., Braga, A. C., et al. (2015). Fish biodiversity of the Vitória-Trindade seamount chain, southwestern Atlantic: an updated database. PLoS One 10:e0118180. doi: 10.1371/journal
Post, D. M., Layman, C. A., Arrington, D. A., Takimoto, G., Quattrochi, J., and Montaña, C. G. (2007). Getting to the fat of the matter: models, methods and assumptions for dealing with lipids in stable isotope analyses. Oecologia 152, 179–189. doi: 10.1007/s00442-006-0630-x
R Core Team. (2017). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Rocha, L. A., and Bowen, B. W. (2008). Speciation in coral-reef fishes. J. Fish Biol. 72, 1101–1121. doi: 10.1111/j.1095-8649.2007.01770.x
Rocha, L. A., Robertson, D. R., Roman, J., and Bowen, B. W. (2005). Ecological speciation in tropical reef fishes. Proc. R. Soc. B Biol. Sci. 272, 573–579. doi: 10.1098/2004.3005
Sazima, I., and Sazima, C. (2004). Daytime hunting behaviour of Echidna catenata (Muraenidae): why chain morays foraging at ebb tide have no followers. Aqua, J. Ichthyol. Aquat. Biol. 8, 1–8.
Schemske, D. W., Mittelbach, G. G., Cornell, H. V., Sobel, J. M., and Roy, K. (2009). Is there a latitudinal gradient in the importance of biotic interactions? Annu. Rev. Ecol. Evol. Syst. 40, 245–269. doi: 10.1146/annurev.ecolsys.39.110707.173430
Schoener, T. W. (1968). The Anolis lizards of Bimini: resource partitioning in a complex fauna. Ecology 49, 704–726. doi: 10.2307/1935534
Schoener, T. W. (1983). Field experiments on interspecific competition. Am. Nat. 122, 240–285. doi: 10.1086/284133
Sexton, J. P., Montiel, J., Shay, J. E., Stephens, M. R., and Slatyer, R. A. (2017). Evolution of ecological niche breadth. Annu. Rev. Ecol. Evol. Syst. 48, 183–206. doi: 10.1146/annurev-ecolsys-110316-023003
Sheppard, C. E., Inger, R., McDonald, R. A., Barker, S., Jackson, A. L., Thompson, F. J., et al. (2018). Intragroup competition predicts individual foraging specialisation in a group-living mammal. Ecol. Lett. 21, 665–673. doi: 10.1111/ele.12933
Simberloff, D. S. (1983). Competition theory, hypothesis-testing, and other community ecological buzzwords. Am. Nat. 122, 626–635. doi: 10.1086/284163
Storero, L. P., Ocampo Reinaldo, M., Narvarte, M., Iribarne, O., and Botto, F. (2020). Trophic interactions and isotopic niche of octopuses and sea stars in North Patagonia. Mar. Biol. 167, 35. doi: 10.1007/s00227-019-3642-x
Svanbäck, R., and Bolnick, D. I. (2007). Intraspecific competition drives increased resource use diversity within a natural population. Proc. R. Soc. B Biol. Sci. 274, 839–844. doi: 10.1098/rspb.2006.0198
Szabo, A. R. (2002). Experimental tests of intercohort competition for food and cover in the tidepool sculpin (Oligocottus maculosus Girard). Can. J. Zool. 80, 137–144. doi: 10.1139/z01-218
Therneau, T. M., and Atkinson, B. (2018). rpart: Recursive Partitioning and Regression Trees. R Packag. version 4.1-13. Available online at: https//CRAN.R-project.org/package=rpart (accessed March 06, 2021).
Tilman, D. (1994). Competition and biodiversity in spatially structured habitats. Ecology 75, 2–16. doi: 10.2307/1939377
Tornabene, L., Van Tassell, J. L., Robertson, D. R., and Baldwin, C. C. (2016). Repeated invasions into the twilight zone: evolutionary origins of a novel assemblage of fishes from deep Caribbean reefs. Mol. Ecol. 25, 3662–3682. doi: 10.1111/mec.13704
Underwood, A. J. (1984). “Paradigms, explanations and generalizations in models for the structure of intertidal communities on rocky shore,” in Ecological Communities: Conceptual Issues and the Evidence, eds D. R. Strong, D. Simberloff, L. G. Abele, and A. B. Thistle (Princeton, NJ: Princeton University Press), 151–180. doi: 10.1515/9781400857081.151
Vaz-Pinto, F., Martínez, B., Olabarria, C., and Arenas, F. (2014). Neighbourhood competition in coexisting species: the native Cystoseira humilis vs the invasive Sargassum muticum. J. Exp. Mar. Bio. Ecol. 454, 32–41. doi: 10.1016/j.jembe.2014.02.001
Volterra, V. (1926). Variazioni e fluttuazioni del numero d’individui in specie animali conviventi. Memorie della R. Accad. dei Lincei 2, 31–113.
Xavier, J. B., and Foster, K. R. (2007). Cooperation and conflict in microbial biofilms. Proc. Natl. Acad. Sci. U.S.A. 104, 876–881. doi: 10.1073/pnas.0607651104
Zamprogno, C. (1989). Distribuição e Hábitos Alimentares dos Peixes da Zona Entremarés de Recifes Rochosos da Praia de Manguinhos, Espírito Santo. Master’s thesis. Campinas, SP: Universidade Estadual de Campinas.
Keywords: tidepool, stable isotopes, rockpool ecology, reef fish, competition
Citation: Andrades R, Joyeux J-C, Macieira RM, Godoy BS, Reis-Filho JA, Jackson AL and Giarrizzo T (2021) Niche-Relationships Within and Among Intertidal Reef Fish Species. Front. Mar. Sci. 8:659579. doi: 10.3389/fmars.2021.659579
Received: 27 January 2021; Accepted: 26 April 2021;
Published: 02 June 2021.
Edited by:
Trevor Willis, Stazione Zoologica Anton Dohrn, ItalyReviewed by:
Eric Diaz Delgado, Anton Dohrn Zoological Station, ItalyCopyright © 2021 Andrades, Joyeux, Macieira, Godoy, Reis-Filho, Jackson and Giarrizzo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ryan Andrades, cnlhbmFuZHJhZGVzQGdtYWlsLmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.