 Jesús S. Hernández-Orts1*
Jesús S. Hernández-Orts1* David I. Hernández-Mena2Camila Pantoja1,3
David I. Hernández-Mena2Camila Pantoja1,3 Roman Kuchta1Néstor A. García4Enrique A. Crespo4
Roman Kuchta1Néstor A. García4Enrique A. Crespo4 Rocío Loizaga4
Rocío Loizaga4- 1Institute of Parasitology, Biology Centre, Czech Academy of Sciences, České Budějovice, Czechia
- 2Centro de Investigación y de Estudios Avanzados del Instituto Politécnico Nacional – Unidad Mérida, Mérida, Mexico
- 3Laboratory of Parasitology, Institute of Ecology, Nature Research Centre, Vilnius, Lithuania
- 4Centro Para el Estudio de Sistemas Marinos–Consejo Nacional de Investigaciones Científicas y Técnicas, Puerto Madryn, Argentina
The poorly known Clymene dolphin (Stenella clymene) is a small oceanic cetacean distributed in tropical and subtropical waters of the Atlantic Ocean. In this study, we report, for the first time, a vagrant individual of Clymene dolphin in Argentina (Rada Tilly) that represents the current southern-most record for this cetacean species. We provide a molecular identification of the dolphin, based on partial sequences of the mitochondrial cytochrome b gene, generated from DNA extracted from its metazoan parasites. Three prey species, namely Argentine hake, Patagonian squid and lobster krill were identified from hard pieces collected in the stomach. Seven metazoan parasite taxa (three in adult stage, one immature stage and three in larval stage) were identified using morphological and molecular data, i.e., the digenean Pholeter gastrophilus (new host record), the cestodes Tetrabothrius (Tetrabothrius) forsteri, Clistobothrium grimaldii unidentified phyllobothriid plerocercoids, the nematodes Anisakis pegreffii (new host record), Halocercus delphini and the acanthocephalan Corynosoma australe (new host record). Our study provides the first molecular exploration of the metazoan parasite diversity in delphinids from the southwestern Atlantic, and contributes significantly to understand the poorly known parasite fauna of the Clymene dolphin.
Introduction
The poorly known Clymene dolphin, Stenella clymene (Gray, 1850) (Cetacea: Delphinidae) is a small oceanic dolphin, endemic to tropical and warm-temperate waters of the Atlantic Ocean, Caribbean Sea, and Gulf of Mexico (Fertl et al., 2003; Jefferson, 2018; Briceño et al., 2020 and references therein). Some unusual records of Clymene dolphins have been also registered off the northern coast of Spain (up to 43°N) (Pis-Millán et al., 2019). In the southwestern Atlantic, records of Clymene dolphins have been reported only off the coast of Brazil, where sightings and stranded dolphins have been reported up to 30°S (Simões-Lopes et al., 1994; Fertl et al., 2003; Moreno et al., 2005). However, the occurrence of Clymene dolphins in the Brazilian coast may be considered as unusual (Prado et al., 2016; Costa et al., 2017; Mayorga et al., 2020). Information about the food habits of Clymene dolphin is scarce, with data obtained from very few individuals stranded or incidentally captured off the Gulf of Mexico, the northeaster coast of Brazil or the North Iberian Peninsula (Jefferson and Curry, 2003; Pis-Millán et al., 2019).
Few studies have reported different metazoan parasite species infecting the Clymene dolphin from the Atlantic coasts of North and South America. Jefferson and Curry (2003) reported cestodes (Clistobothrium spp. [identified as Phyllobothrium sp. and Monorygma sp.]), nematodes (Halocercus sp., Pharurus sp. and Nasitrema sp.), whale lice (Syncyamus pseudorcae Bowman, 1955) and barnacles (Xenobalanus sp.). Aguilar-Aguilar et al. (2010) described adults of Halocercus delphini Baylis and Daubney, 1925 (syn. Skrjabinalius guevarai Gallego and Selva, 1979) and larvae of Anisakis sp. from one Clymene dolphin stranded on the coast of Quintana Roo, Mexico. The most comprehensive parasitological study of Clymene dolphins was provided by Carvalho et al. (2010), who reported 11 metazoan parasites, namely Anisakis sp., Bolbosoma sp., Halocercus brasiliensis, Halocercus sp., Clistobothrium grimaldii (Moniez, 1899) [as Monorygma grimaldii (Moniez, 1899)], Clistobothrium delphini (Bosc, 1802) [as Phyllobothrium delphini (Bosc, 1802)], Scolex pleuronectis (Müller, 1788), S. pseudorcae, Strobilocephalus triangularis (Diesing, 1850), Tetrabothrius (Tetrabothrius) forsteri (Krefft, 1871) and Trigonocotyle sp. from 16 dolphins stranded along the northeastern coast of Brazil. Guimarães et al. (2015) described the lesions caused by H. brasiliensis in the lungs of five out of nine dolphins from the northeast region of Brazil. Recently, Azevedo et al. (2017) examined, based on an integrative taxonomy approaches, the diversity of anisakid nematodes from 13 Clymene dolphins stranded on the northeastern coast of Brazil and reported adults and larvae of Anisakis typica (Diesing, 1860) and Anisakis sp.
In this study, we provide the southernmost record of the Clymene dolphin in the Atlantic Ocean. New data on diet composition and helminth fauna, including the first molecular sequences for some parasitic taxa, are provided for this poorly documented dolphin species. Our study also provides a new host record, new parasite localities and comments on the life-cycle of each helminth taxa in the southwestern Atlantic.
Materials and Methods
Stranding Data and Sample Collection
On 23 July 2017 a Clymene dolphin was found stranded in Rada Tilly (45°55′S, 67°55′W), Chubut Province, Argentina (Figure 1A). The dolphin was in fresh condition (Figures 1B–D), transported to a fish factory near Rada Tilly and frozen at −20°C for later examination. The dolphin was transported frozen to the Laboratorio de Mamíferos Marinos (LAMAMA), Centro para el Estudio de Sistemas Marinos (CESIMAR–CONICET) in Puerto Madryn, Argentina for a necropsy to be performed. Thawed dolphin was identified following Jefferson et al. (2015), and measured following standardized methods to the nearest centimeter (Norris, 1961). A complete necropsy was done by the staff of the LAMAMA in order to obtain food habits and parasitological information. The skull and the complete skeleton are deposited in the Colección de Mamíferos Marinos, Centro para el Estudio de Sistemas Marinos (CESIMAR–CONICET), Puerto Madryn, Chubut, Argentina (accession number CNPMAMM 0793).

Figure 1. (A) Distribution of the Clymene dolphin, Stenella clymene (yellow) in the southwestern South Atlantic Ocean according to Fertl et al. (2003). The arrow points the stranding site of the dolphin examined in the present study. (B) Carcase of the female Clymene dolphins stranded in Rada Tilly, showing the beak coloration including the “mustache.” (C,D) Pigmentation pattern showing tripartite flank coloration.
The contents of the four stomach chambers (i.e., forestomach, main and pyloric stomachs, and duodenal ampulla) were collected. The intestine was measured and divided into 30 sections of equal length. The four stomach chambers and each section of the intestine were washed with tap water separately through sieves of either 0.2- or 0.5-mm mesh. Additionally, the wall of the stomach and intestine were carefully examined to detect nodules or attached parasites. The liver and lungs were also examined for parasites. Stomach and intestinal contents were later examined under a dissecting microscope. Food elements (pieces of prey, otoliths and beaks) were preserved in 70% ethanol. Parasites were stored in 70% ethanol or in molecular-grade (99%) ethanol.
Molecular Identification of the Dolphin
Total genomic DNA was extracted from an ethanol-fixed isolate of a pseudaliid nematode (isolate ID: H_delphini_E140) or an anisakid nematode (isolate ID: A_pegreffii_E68), following the procedure described in the section “Molecular data and phylogenetic analyses of parasites.” A partial fragment (ca. 950 nt) of the mitochondrial cytochrome b (cytb) gene was amplified using our designed primers Sten_cox1F (5′-CTC CTC TTG ATG AAA TTT TG-3′) and Sten_cox1R (5′-TTG GCC TAC AAT GAT ATA TG-3′) for molecular identification of the dolphin. The PCR thermocycling profile comprised initial denaturation at 95°C for 3 min, followed by 35 cycles of 94°C for 1 min, 55°C for 1 min and 72°C for 1 min, with a final extension step at 72°C for 7 min. Polymerase chain reaction (PCR) amplifications were carried out in 25 μl reactions, containing 1.5 μl of each primer (10 μM), 8.0 μl PCR water, 12.5 μl of AccuStart II PCR SuperMix (Quanta bio, Beverly, MA, United States) and 1.5 μl of diluted template DNA. PCR amplicons were purified with Exo-SAP-IT Kit (GE Healthcare Life Sciences, United Kingdom). Purified amplicons were Sanger sequenced at SEQme (Dobříš, Czechia).
Sequences obtained were assembled and inspected for errors using Geneious Pro® v.11.0.4 (Biomatters Ltd., Auckland, New Zealand). The identities of our newly generated sequences were verified using the Basic Local Alignment Tool (BLAST) on the NCBI database and deposited in GenBank with the accession number MW714333.
Diet Composition
Food elements were identified using the reference collection of the LAMAMA and from descriptions in available literature (e.g., Clarke, 1986; Boschi et al., 1992; Gosztonyi and Kuba, 1996). The total length (TL) of fish, dorsal mantle length (DML) of squid, maximum abdomen width (MAW) of crustaceans and wet weight (W) of prey were estimated from hard pieces following the allometric regressions detailed in Koen Alonso et al. (2001).
Parasitological Procedures
For parasite identification, flatworms and acanthocephalans were stained with Mayer’s hydrochloric carmine, dehydrated through an ethanol series, cleared in methyl salicylate and mounted in Canada balsam. Nematodes were cleared in glycerine. Mounted or cleared specimens were examined with an Olympus BX51 microscope. Parasites were identified according to Delyamure (1955), Temirova and Skrjabin (1978), Zdzitowiecki (1991), Bray et al. (2008), Anderson et al. (2009), and Jensen and Bullard (2010).
Voucher specimens are deposited in the Helminthological Collection of the Institute of Parasitology (IPCAS), Biology Centre, Czech Academy of Sciences, České Budějovice, Czech Republic, with the following accession numbers: Anisakis pegreffii Campana-Rouget and Biocca, 1955 (IPCAS N-1253), Clistobothrium grimaldii (Moniez, 1899) (IPCAS C-882), Halocercus delphini (IPCAS N-1252), phyllobothriid plerocercoids (IPCAS C-883), Pholeter gastrophilus (Kossack, 1910) (IPCAS D-830) and Tetrabothrius (Tetrabothrius) forsteri (Krefft, 1871) (IPCAS C-884).
Molecular Data and Phylogenetic Analyses of Parasites
A small piece of the dorsal surface of the disc of the acanthocephalan, from the midlevel of the body of the nematodes, the lateral margin of the trematode and the posterior end of the cestodes were used for DNA isolation and sequencing. The remaining parts of the parasites were kept as hologenophores sensu Pleijel et al. (2008). Total genomic DNA was extracted from selected parasitic taxa fixed in 99% ethanol using the REDExtract-N-Amp Tissue PCR kit (Sigma, St. Louis, United States).
Four molecular markers were sequenced: (i) the large subunit of the ribosomal RNA (28S rDNA) gene (domains D1–D3) for phyllobothriid tapeworms and pseudaliid nematodes; (ii) the second internal transcribed spacer (ITS2) gene for pseudaliid nematodes; (iii) the mitochondrial cytochrome c oxidase subunit 1 (cox1) gene for heterophyid trematodes and polymorphid acanthocephalans; and (iv) the mitochondrial cytochrome c oxidase subunit 2 (cox2) gene for anisakid nematodes. Primers and PCR amplification profiles for each molecular marker are described in Table 1. PCR amplification reactions, amplicon purification and sequencing methods follow the procedures described above.
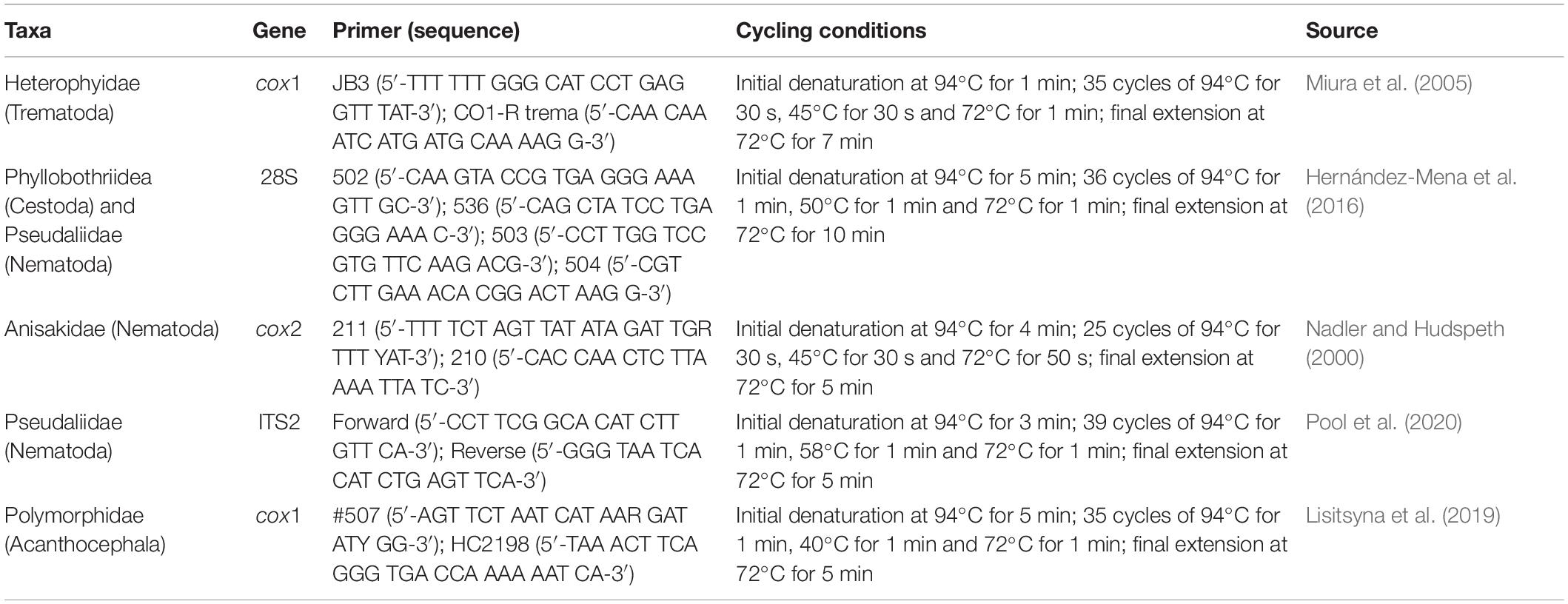
Table 1. List of primers and thermocycling profiles used in the molecular analyses of the helminth parasites from the Clymene dolphin in Argentina.
Sequences obtained from each set of primers were assembled and inspected for errors using Geneious. The identity of our newly generated sequences was verified using BLAST Newly generated sequences were deposited in GenBank with the accession numbers MW692916, MW692937–MW692943, MW711662–MW711669, MW724482, MW724483.
Newly generated sequences of each parasitic taxa were aligned independently with representative sequences for taxa of the same or proximal families (see Supplementary Table 1). The outgroup of each dataset was selected following previous analyses on the phylogeny of each parasitic taxa (e.g., Carreno and Nadler, 2003; Caira et al., 2020; Pool et al., 2020; García-Varela et al., 2021). Sequences included in each dataset were aligned with the MAFFT algorithm (Katoh and Standley, 2013) implemented in Geneious. Phylogenetic analyses were conducted under maximum likelihood (ML) criteria. The best nucleotide substitution model for each dataset was selected with jModelTest v2 (Darriba et al., 2012) under the Akaike Information Criterion. Based on the results of the jModelTest analyses, the following models of evolution were used: (i) GTR for the 28S rDNA dataset for metastrongylid nematodes; (ii) GTR + G for the cox1 dataset for heterophyid trematodes and the 28S dataset for the phyllobothriid cestodes; (iii) GTR + G + I for the cox1 dataset for polymorphid acanthocephalans; and (iv) HKY + G for the cox2 dataset for anisakid nematodes and the ITS2 dataset for pseudaliid nematodes.
Maximum likelihood analyses were constructed in RAxML v.7.0.4 (Stamatakis, 2006), with 1,000 bootstrap replicates to estimate support values, and the resulting trees were visualized in FigTree v.1.4.4 (Rambaut, 2018). Genetic divergences (uncorrected p-distance) were estimated with MEGA v.10.1.8 (Kumar et al., 2018).
Results
Biological Data, Molecular Identification, and Prey Species
The Clymene dolphin stranded in Rada Tilly was a young immature female (Figure 1B) with a total body length of 184 cm and a weight of 57 kg. The stranded Clymene dolphin had the distinctive morphological features described for this species, i.e., a dark stripe or “mustache” on the dorsal surface of the beak (Figure 1B), a three-part color pattern, with a dark gray cape, light gray sides, and white bell (Figures 1C,D), and a dipped dorsal cape (Jefferson et al., 2015). Morphometric data of the Clymene dolphin is provided in Table 2. The dolphin presented 42–43 (upper) and 43–46 (lower) small teeth, which correspond to the tooth counts reported for Clymene dolphins (Jefferson et al., 2015). Morphometric data of the skull (Figure 2) is provided in Supplementary Table 2.
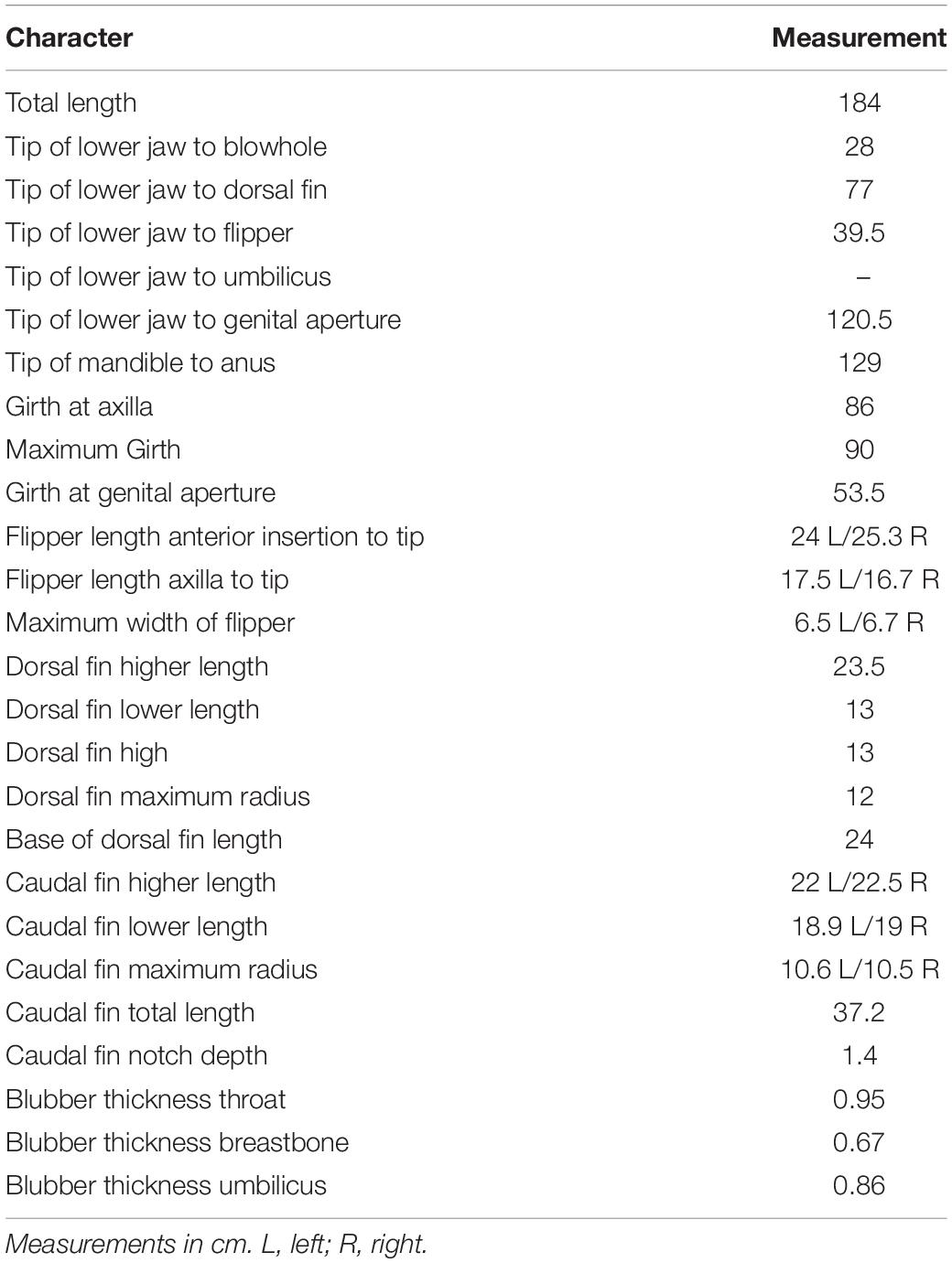
Table 2. External morphological measurements of the Clymene dolphin, Stenella clymene stranded in Rada Tilly, Chubut, Argentina.
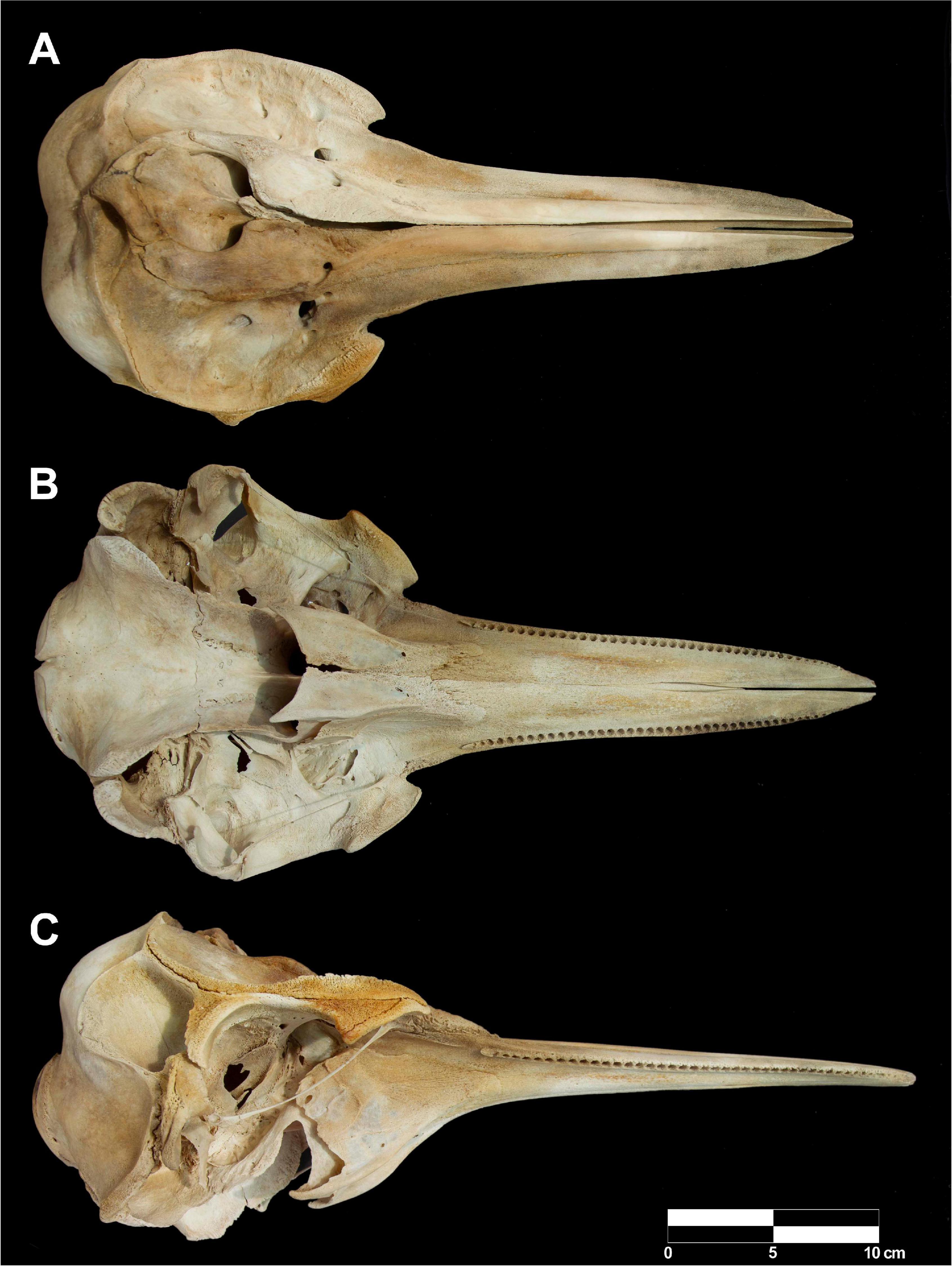
Figure 2. Skull of the young immature female Clymene dolphin, Stenella clymene stranded in Rada Tilly, Chubut, Argentina. (A) Ventral view. (B) Dorsal view. (C) Lateral view.
Our designed primers for dolphins generated two identical cytb sequences (918 nt) from two isolates of parasitic nematodes collected from the Clymene dolphin. BLAST search showed 100% identity with 97% query cover with an isolate of S. clymene from Brazil (KX346591) of Nara et al. (2017).
Hard pieces of three prey species were collected only in the stomachs: Argentine hake, Merluccius hubbsi Marini, 1933 (otoliths of three individuals: TL = 16.8–19.5 mm; W = 26.8–42.4 g), Patagonian squid, Doryteuthis (Amerigo) gahi (d’Orbigny, 1835) (beaks of two individuals: DML = 10.3–12.1 mm; W = 5.1–16.1 g) and lobster krill, Munida gregaria (Fabricius, 1793) (one individual: MAW = 8.9 mm; W = 1.5 g).
Parasite Composition
Seven helminth taxa belonging to the Digenea (n = 1), Cestoda (n = 3), Nematoda (n = 2) and Acanthocephala (n = 1) were collected. Three helminth taxa were adults, one was an immature form and three were found as larval stages. A single nodule with one gravid heterophid trematode Pholeter gastrophilus (Kossack, 1910) (Supplementary Figure 1A) was observed in the duodenal ampulla. Poorly preserved strongylid nematodes, morphologically identified as Halocercus sp. were collected from the airways (n = 159) (Supplementary Figure 1C). Fourth-stage larvae (L4) of nematodes identified based on parasite morphology as Anisakis sp. were found in the forestomach (n = 4), main stomach (n = 32), intestine (n = 1) and rectum (n = 13). In the stomach, we did not observed ulcers associated with these nematodes. Mature individuals of Tetrabothrius (Tetrabothrius) forsteri (Krefft, 1871) were collected in the first sections of the intestine (n = 7) (Supplementary Figures 1D,E), while Type I or II onchoproteocephalid plerocercoids sensu Jensen and Bullard (2010) were found free in the rectum (n = 13) (Supplementary Figure 1F). A single immature male of the polymorphid acanthocephalan Corynosoma australe Johnston, 1937 was found attached to the rectum. Finally, encysted larval Type XV merocercoids sensu Jensen and Bullard (2010) were collected from the anal canal (n = 12) (Supplementary Figure 1B).
Molecular Identification of Metazoan Parasites
A total of 18 partial sequences were generated in this study: one cox1 sequence (825 nt) for P. gastrophilus, one 28S sequence (950 nt) for Type XV merocercoid, eight cox2 sequences (579 nt) for Anisakis sp., one ITS2 (454 nt) and six 28S sequences (999–1,001 nt) for Halocercus sp., and one cox1 sequence (645 nt) for C. australe. We attempted to generate sequences for isolates of adult T. (T.) forsteri and the onchoproteocephalid plerocercoid Type I/II identified based on morphology, but representative sequence from these isolates were not obtained.
Our phylogenetic analyses of the cox1 dataset for heterophyid trematodes (16 taxa; 20 sequences; 369 nt; Supplementary Table 1A) clustered our newly generated sequence of P. gastrophilus from the Clymene dolphin with published sequences of P. gastrophilus from a bottlenose dolphin, Tursiops truncatus (Montagu, 1821) (KT883857 and KX059405), a striped dolphin, Stenella coeruleoalba (Meyen, 1833) (KT883856) and a short-finned pilot whale, Globicephala macrorhynchus Gray, 1846 (KX059411) (Figure 3). Our novel cox1 sequence for P. gastrophilus was identical to the sequence of the adult of this trematode reported from a bottlenose dolphin (KX059405) from Patagonia by Fraija-Fernández et al. (2017). Genetic divergence between isolates of P. gastrophilus from these odontocete species range between 0.0% and 0.3% (0–1 nt).
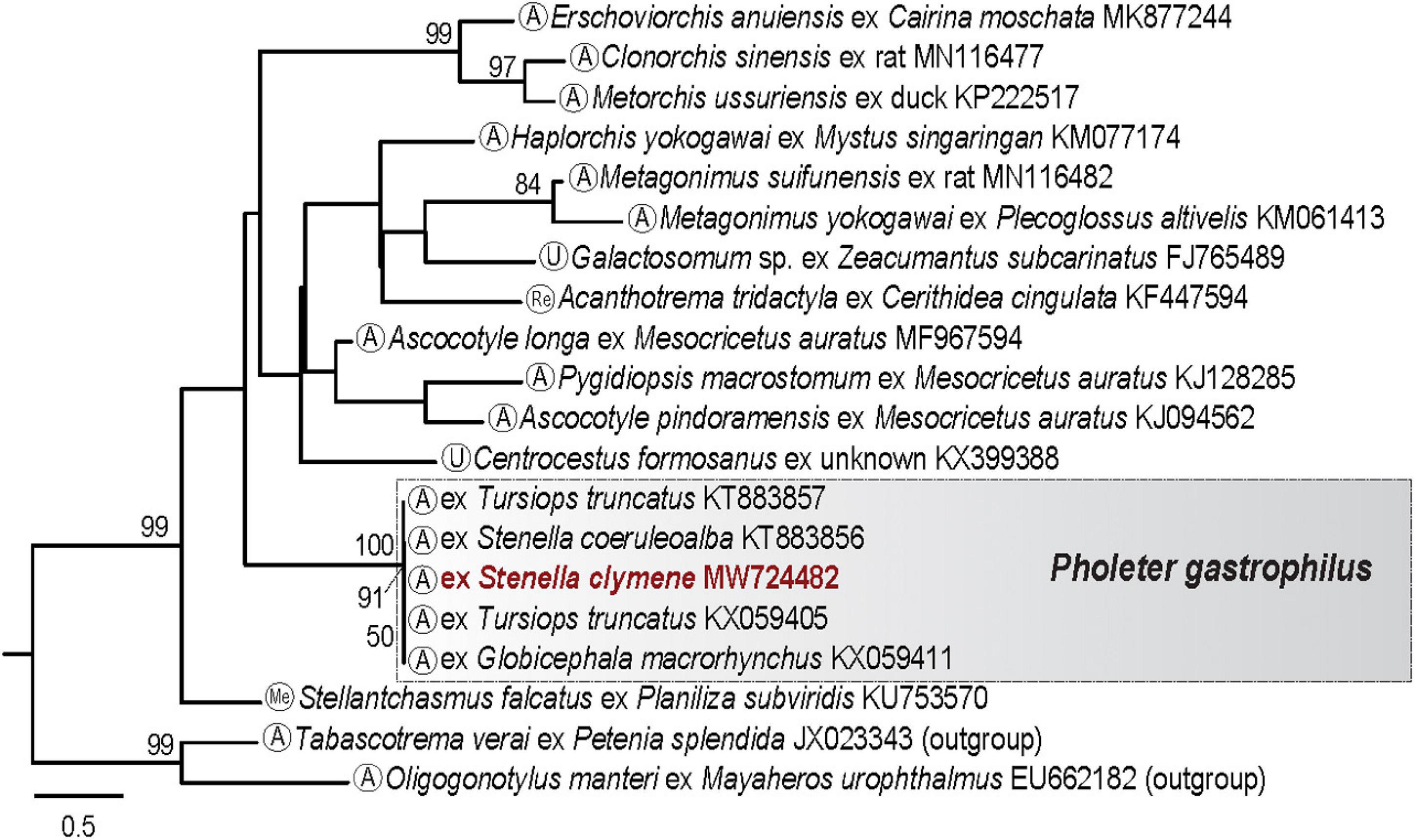
Figure 3. Maximum likelihood (ML) phylogram for the cox1 rDNA dataset for heterophyid trematodes. Numbers indicate nodal supports from ML analysis (bootstrap values >50% shown only). Developmental stage for each taxon is indicated in the circle in terminal taxon labels. The newly generated sequence is indicated in red. The scale-bar indicates the number of substitutions per site. Me, metacercaria; Re, redia; A, adult; U, unknown stage.
Maximum likelihood analysis based on the 28S dataset for phyllobothriid cestodes (15 taxa; 54 sequences; 1,307 nt; Supplementary Table 1B), including our sequence from an isolate identified as a Type XV merocercoid, showed congruent results with Caira et al. (2020). The representative isolate of the Type XV merocercoid clustered, although with a negligible support, with published sequences obtained from merocercoids of Clistobothrium grimaldii (Moniez, 1899) collected from different dolphin species from the Mediterranean Sea and the Pacific coast of South America, and with one isolate from the Cape fur sea, Arctocephalus pusillus pusillus (Schreber) (KU724058) from South Africa (Figure 4). The genetic divergence in this clade ranged between 0 and 0.6% (0–3 nt).
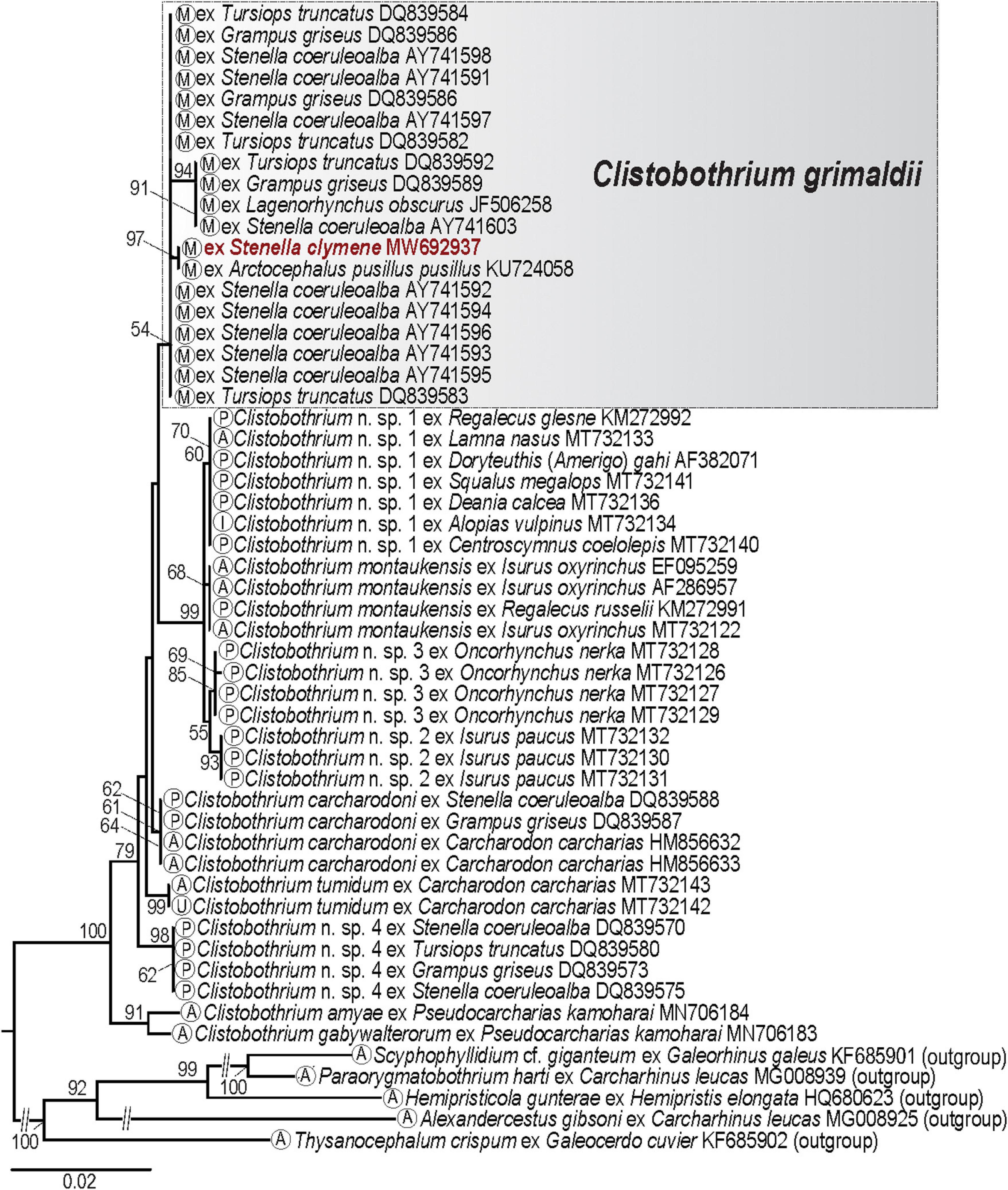
Figure 4. Phylogenetic tree using maximum likelihood for the 28S dataset for phyllobothriid cestodes. Numbers indicate nodal supports from ML analysis (bootstrap values >50% shown only). Developmental stage for each taxon is indicated in the circle in terminal taxon labels. The newly generated sequence is indicated in red. The scale-bar indicates the number of substitutions per site. P, plerocercoid; M, merocercoid; A, adult; U, unknown stage.
A single ITS2 partial sequence was obtained from 1 out of 10 isolates identified based on morphology as Halocercus sp. The ML analysis of the ITS2 dataset (7 taxa; 11 sequences; 699 nt; Supplementary Table 1C) produced a tree topology (Figure 5A) consistent with that in Pool et al. (2020). The newly generated sequence for lungworm nematode falls into a well-supported clade together with isolates identified as Halocercus delphini. Genetic divergence between our new sequence and the published sequences of H. delphini was 0.2–0.9% (19–80 nt). Based on our molecular results we consider the isolate from the Clymene dolphin to be conspecific with H. delphini.
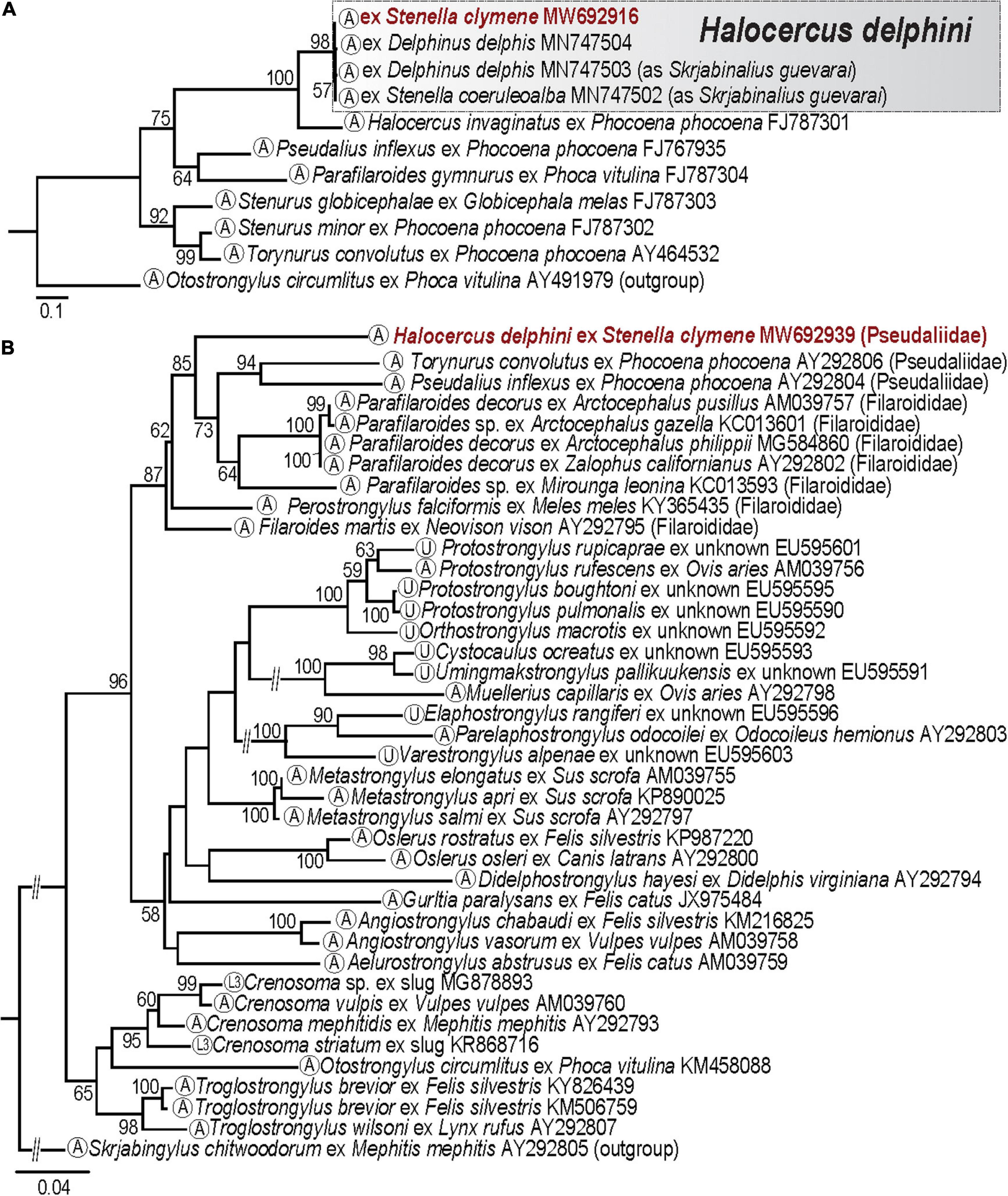
Figure 5. (A) Maximum likelihood (ML) tree constructed using the ITS2 dataset for pseudaliid nematodes. (B) ML phylogram inferred from the 28S dataset for members of the superfamily Metastrongyloidea. Numbers indicate nodal supports from ML analysis (bootstrap values >50% shown only). Developmental stage for each taxon is indicated in the circle in terminal taxon labels. The newly generated sequences are indicated in red. The scale-bar indicates the number of substitutions per site. L3, third-stage larva; A, adult; U, unknown stage.
Novel sequences data for the 28S marker resulted in identical sequences for five lungworms from the Clymene dolphin. Our newly obtained ITS2 (MW692916) and one 28S (MW692939) sequences were generated from the same lungworm isolate (ID: H__delphini_E158). We, therefore, consider the lungworms used in our 28S rDNA analysis as H. delphini. The phylogenetic analysis of the 28S dataset (36 taxa; 44 sequences; 1,200 nt; Supplementary Table 1D) for the superfamily Metastrongyloidea placed our novel partial sequences for H. delphini in a clade together with representative sequences of members of the families Filaroididae Schulz, 1951 (Filaroides van Beneden, 1858, Parafilaroides Dougherty, 1946, Perostrongylus Schlegel, 1934) and Pseudaliidae Railliet and Henry, 1909 (Pseudalius Dujardin, 1845 and Torynurus Baylis and Daubney, 1925) (Figure 5B). Particularly within this clade, H. delphini appeared with strong support as a sister taxon of a group formed by members of the families Filaroididae (Parafilaroides) and Pseudaliidae (Torynurus and Pseudalius) collected from different marine mammal species (Figure 5B).
The analyzed cox2 dataset for the anisakid nematodes (10 taxa; 51 sequences; 584 nt; Supplementary Table 1E), placed our sequences from eight isolates and selected sequences of third-stage larvae of Anisakis pegreffii retrieved from the GenBank dataset in a strongly supported clade (Figure 6). Our phylogenetic analysis provides robust evidence that eight isolates from the Clymene dolphin are conspecific with A. pegreffii. The intraspecific divergence within the A. pegreffii clade ranged between 0% to 2.9% (0–17 nt). The interspecific divergence between the eight isolates collected from the Clymene dolphins also ranged between 0% and 2.9% (0–17 nt).
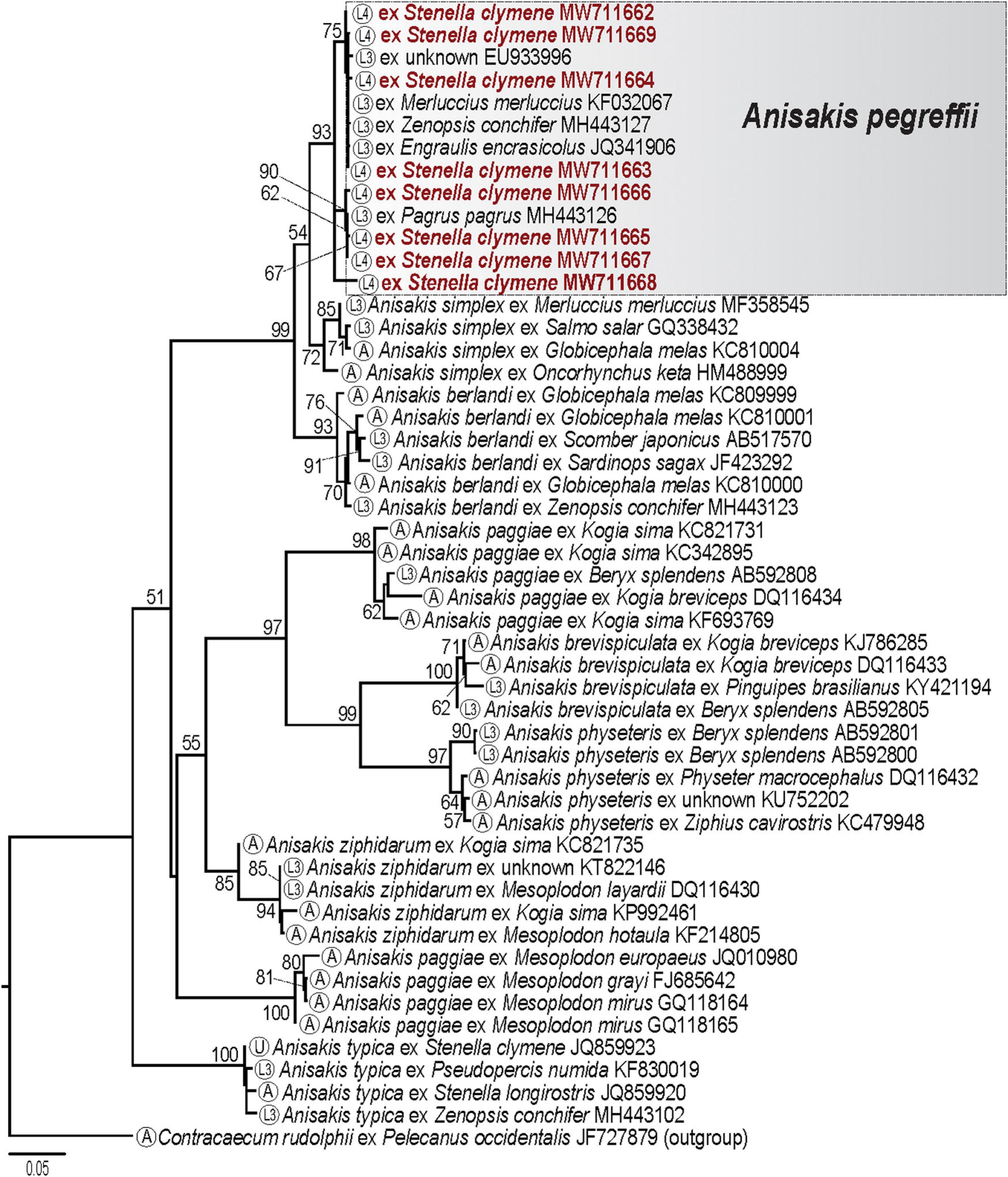
Figure 6. Maximum likelihood tree from phylogenetic analysis of the cox2 dataset for anisakid nematodes. Numbers indicate nodal supports from ML analysis (bootstrap values >50% shown only). Developmental stage for each taxon is indicated in the circle in terminal taxon labels. The newly generated sequences are indicated in red. The scale-bar indicates the number of substitutions per site. L3, third-stage larva; L4, fourth-stage larva; A, adult; U, unknown stage.
Maximum likelihood analysis of the cox1 dataset for the polymorphid acanthocephalans (16 taxa; 23 sequences; 655 nt long; Supplementary Table 1F) resulted in a tree (Figure 7) congruent with the results of García-Varela et al. (2021). Our newly generated sequence of Corynosoma australe formed a well-supported clade with sequences of C. australe from fish, penguins and sea lions from the Pacific coast of North America and the Atlantic coast of South America. The genetic divergence in the cox1 sequence between the isolate of C. australe from the Clymene dolphin and sequences from other marine hosts range between 0.4% and 1.0% (2–6 nt).
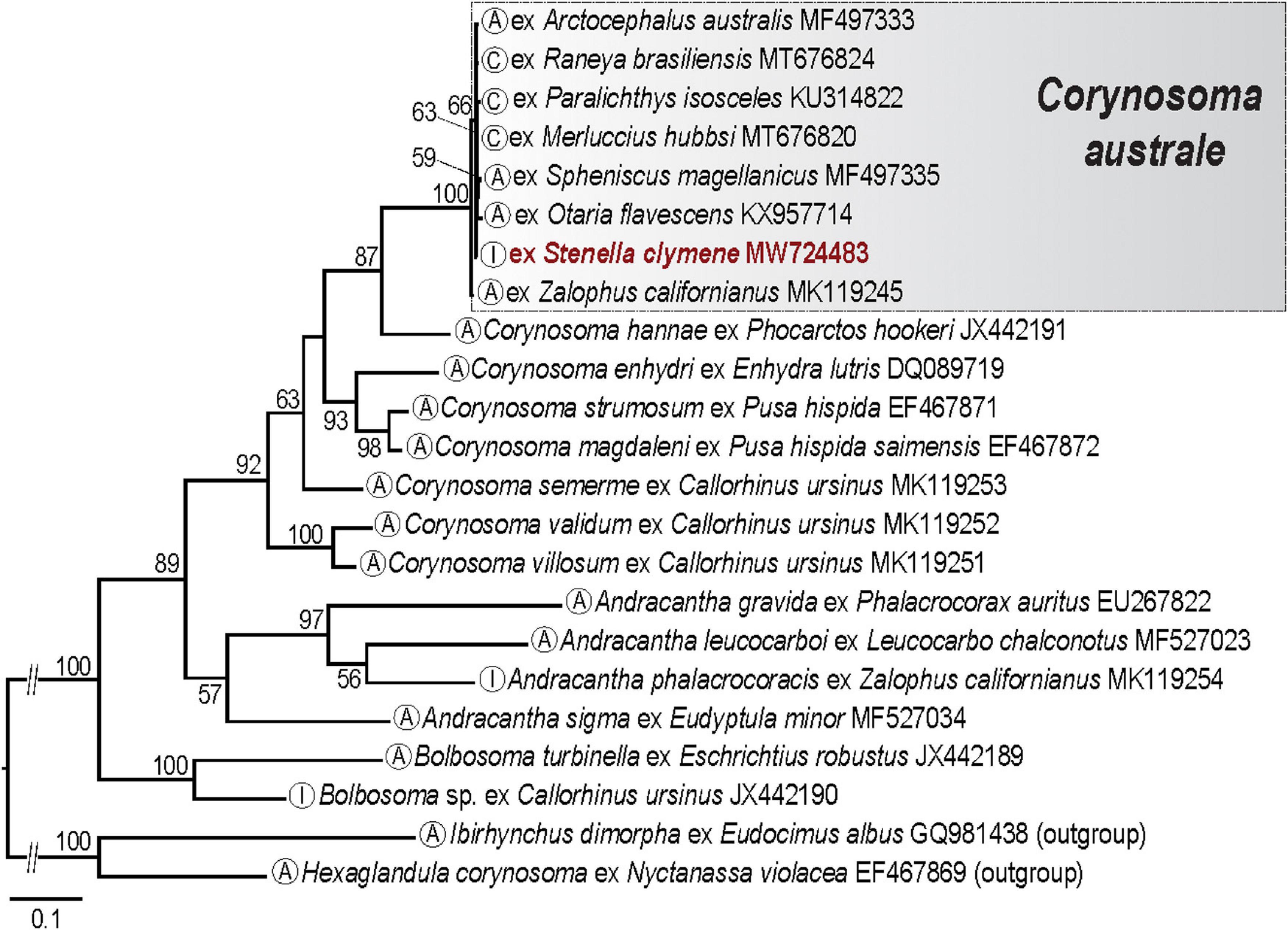
Figure 7. Maximum likelihood (ML) phylogram reconstructed for the cox1 dataset for polymorphid acanthocephalans. Numbers indicate nodal supports from ML analysis (bootstrap values >50% shown only). Developmental stage for each taxon is indicated in the circle in terminal taxon labels. The newly generated sequence is indicated in red. The scale-bar indicates the number of substitutions per site. C, cystacanth; I, immature; A, adult.
Discussion
Molecular Identification, Distribution, and Diet Composition
Our study demonstrated that dolphin DNA can be extracted and successfully amplified from its metazoan parasites. To our knowledge, we provided the first molecular identification of a cetacean using partial sequences obtained from its helminths. Gastrointestinal parasites can be collected in fecal samples of free-living cetaceans (e.g., Hermosilla et al., 2015), therefore, DNA extracted from parasites represents a potential non-invasive approach to obtain molecular data of marine mammals for identification or genetic studies.
In the Southern Hemisphere, Clymene dolphins have been sighted off the Atlantic coast of Angola, Brazil and the Democratic Republic of the Congo, between ca. 3°S and ca. 30°S (Moreno et al., 2005; Weir, 2006). Clymene dolphins were also recorded off Ascension Island (3°40′S, 18°5′W) in the Southeast Atlantic (Fertl et al., 2003). The verified southern-most records for this species were reported by Simões-Lopes et al. (1994) in Torres (29°18′S, 49°42′W), Santa Catarina and by Fertl et al. (2003) in Tramandaí (29°59′S, 50°07′W), Rio Grande do Sul in Brazil. Our record from Rada Tilly (45°55′S, 67°55′W) in Patagonia represents the current southern-most record for this species. However, the occurrence of Clymene dolphin off the Patagonian coast should be considered as unusual.
Little else is known of their ecology, and this species remains one of the most poorly known of all delphinids (Weir et al., 2014). Furthermore, the trophic ecology of Clymene dolphin is scantily explored, very few stomachs have been currently examined and even fewer observations of feeding behavior are reported in the literature (Jefferson, 2018). Jefferson and Curry (2003) reported squid beaks and fish otoliths from the families Myctophidae Gill, 1893, Argentinidae Bonaparte, 1846 and Bregmacerotidae Gill, 1872 in the stomach of a single dolphin stranded in New Jersey, United States. A recent study by Pis-Millán et al. (2019) found a beak of a decapodiform cephalopod and otoliths of European hake, Merluccius merluccius (Linnaeus, 1758) (TL = ca. 10–13 cm) in two Clymene dolphins stranded in the Cantabrian coast of Spain.
The analyses of the stomach contents of the Clymene dolphin examined in this study revealed that demersal-pelagic species, i.e., Argentine hake, Patagonian squid and lobster krill, were consumed. Known ranges of prey show that the Clymene dolphin consumed juvenile Argentine hake and immature Patagonian squid (Hatfield et al., 1990; Pájaro et al., 2005). The ecosystem of central Patagonia constitutes an important nursery area for several species and is dominated by Argentine hake but the relative abundance depends on depth and the time of the year. Among cephalopods, the Patagonian squid is an amphioceanic species which is especially abundant in cold waters of the Patagonian continental shelf up to 350 m depth (Barón, 2001). Hence, it is probable that the vagrant Clymene dolphin from Patagonia just fed on the most abundant available local resources as it has been reported for other dolphin species in the area (e.g., Koen Alonso et al., 1998; Loizaga de Castro et al., 2016).
Considering the oceanic habits of the species its preference for water depths ranging between 437 and 5000 m (Jefferson, 2018), it’s likely that currents carried the vagrant animal toward the shallow waters of central Patagonia. This is the southern-most record of the species and documents the Clymene dolphin as part of the cetacean biodiversity of Argentina. Erratic individuals of other dolphin species have been recorded along the Patagonian coast in the last decades, including two hourglass dolphins, Lagenorhynchus cruciger (Quoy and Gaimard, 1824) a striped dolphin and twelve Fraser’s dolphin, Lagenodelphis hosei (Fraser, 1956) (Fernández et al., 2003; Loizaga de Castro et al., 2011; LAMAMA unpublished data). It is highly possible that tropical or subtropical species, like striped or Fraser’s dolphins, move south with subtropical currents, while subarctic species, like hourglass dolphins, move northward with subantarctic currents. In any case all of these findings correspond to erratic individuals, most of the time juvenile dolphins. However, we cannot rule out that the presence of tropical or subtropical species in Patagonia could be related to climate change. In this scenario, these species will likely extend their distribution poleward if temperature increase as a result of global warming, and will likely induce long-term changes in the local cetacean community (MacLeod, 2009; Poloczanska et al., 2013). The mechanisms causing those changes remain unknown, especially because for some species, it is difficult to differentiate between short-term responses to regional resource variability and longer-term ones that could be driven by climate change (Evans and Bjørge, 2013). Therefore, any changes in the local cetacean community should be fully researched to assess their implications for current and future conservation plans.
Parasite Diversity and Life-Cycle
Pholeter gastrophilus is a generalist digenea with a cosmopolitan distribution infecting coastal and oceanic cetaceans (Aznar et al., 2006). This parasite lives within fibrotic nodules in the submucosa of the stomach of odontocetes (Geraci and Aubin, 1987). In Argentina, six odontocete species have been reported as hosts for this gastric parasite (Supplementary Table 3). The single specimen of P. gastrophilus collected in this study, represent the first record of this trematode in the Clymene dolphin.
Species of Clistobothrium Dailey and Vogelbein, 1990 use sharks of the families Lamnidae Bonaparte, 1835 and Pseudocarchariidae Taylor, Compagno and Struhsaker, 1983 as definitive hosts (see Caira et al., 2020). Pinnipeds and cetaceans have been commonly reported as intermediate hosts for two species now included in Clistobothrium by Caria et al. (2020), but referred to in the past as P. delphini and Monorygma grimaldii (Moniez, 1899) (e.g., Aznar et al., 2007; Klotz et al., 2018). These merocercoids encyst in the liver, subcutaneous blubber, peritoneum of the abdominal cavity, liver and anal canal of cetaceans (Delyamure, 1955; Agustí et al., 2005). The complete life-cycles of Clistobothrium delphini and C. grimaldii are still unknown, but predatory and/or scavenger pelagic sharks may be their definitive hosts (Aznar et al., 2007). Based on morphological evidence, Carvalho et al. (2010) identified merocercoids of these two species of Clistobothrium in the blubber and abdominal cavity of Clymene dolphins from the northeastern coast of Brazil. In this study we molecularly confirmed for the first time the Clymene dolphin as an intermediate hosts of C. grimaldii.
Larval cestodes, collectively referred to as S. pleuronectis have been reported in the digestive tract and hepatopancreatic ducts of several marine mammals (Agustí et al., 2005; Aznar et al., 2007), including Clymene dolphins off Brazil (Carvalho et al., 2010). Molecular data generated from plerocercoids collected in cetaceans revealed clear affinities with members of Clistobothrium (Aznar et al., 2007; Caira et al., 2020). Recently, Caira et al. (2020) suggested that both plerocercoids and merocercoids in marine mammals may belong to different species of Clistobothrium. Following the key of larval type by Jensen and Bullard (2010), the plerocercoids collected in the rectum of the Clymene dolphin were assigned as being Type I or II, which are comprised of onchoproteocephalid cestodes of the genera Acanthobothrium Blanchard, 1848, Phoreiobothrium Linton, 1889, and Triloculatum Caira and Jensen, 2009. However, the larvae collected in our study (Supplementary Figures 1B,C) had a morphology similar to those described from cetaceans by Agustí et al. (2005), which were later assigned to Clistobothrium using molecular methods (Aznar et al., 2007; Caira et al., 2020). In fact, plerocercoids from the Clymene dolphin falls within the range provided by Agustí et al. (2005) for the body size of the large plerocercoid form (referred to as Clistobothrium n. sp. 4. by Caira et al., 2020): 1,213–2,176 μm × 299–357 μm vs. 821–4,261 μm × 200–780 μm. Thus, we cannot rule out that our specimens may belong to Clistobothrium. We therefore prefer to identify these larvae from the Clymene dolphin as “phyllobothriid plerocercoids.” Detailed morphological and molecular characterization of plerocercoids infecting cetaceans in the southwestern Atlantic is necessary for reliable identification of these larvae in this region.
Tetrabothrius (T.) forsteri was the only mature parasites found in the intestine of the Clymene dolphin in the present study. This cestode has been reported worldwide in cetaceans, mainly delphinids (Fraija-Fernández et al., 2016; Mariaux et al., 2017). In the southwestern Atlantic, Carvalho et al. (2010) reported, based on morphological data, specimens of T. (T.) forsteri in the intestine of the Clymene dolphin and the spinner dolphin, Stenella longirostris (Gray, 1828) off Brazil. In Argentina, tetrabothriids have rarely been reported in delphinids (Supplementary Table 3), and the specific identity of most specimens remains unknown.
Lungworm nematodes H. delphini (identified as Skrjabinalius guevarai Gallego and Selva, 1979; see Pool et al., 2020) were reported in the airways of a Clymene dolphin found stranded on the Caribbean coast of Mexico by Aguilar-Aguilar et al. (2010). Carvalho et al. (2010) reported H. brasiliensis Almeida, 1933 and Halocercus sp. in the lungs of a Clymene dolphin from Brazil. However, the specific identity of these two lungworm nematodes, using molecular approaches, is necessary for suitable identification of lungworm nematodes infecting dolphins in Brazil.
Anderson et al. (2009) allocated the genus Halocercus to the subfamily Halocercinae Delamure, 1952 in the Pseudaliidae and classified them mostly based on morphological features of the bursa. However, our phylogenetic analysis of the 28S dataset revealed that Halocercus appears as a distinct family-level clade within the clusters comprising the Pseudaliidae (Pseudalius and Torynurus) and the Filaroididae (Parafilaroides) (Figure 5B). Our result suggests that the systematic position of Halocercus may deserve a detailed morphological and molecular analyses.
Larvae of Anisakis sp. were reported from the main stomach of a Clymene dolphin from the Caribbean coast of Mexico (Aguilar-Aguilar et al., 2010). Anisakis typica was identified using morphological and molecular tools from the stomach of Clymene dolphins in Brazil (Azevedo et al., 2017). Our molecular identification confirmed the presences of A. pegreffii in the Clymene dolphin (new host record). Interestingly, all anisakid nematodes collected in this study were classified as L4 larvae of A. pegreffii. We suggest that the Clymene dolphin may acts as a definitive host for this species, because A. pegreffii mature in the stomach of other dolphin species (e.g., Mladineo et al., 2019; Pons-Bordas et al., 2020). We believe that these nematodes were likely recruited by the Clymene dolphin in Patagonia. In fact, Lanfranchi et al. (2018) reported that A. pegreffii was the dominant species in the Argentine hake from Patagonia and the most common species in the southern Argentine Sea.
Corynosoma australe is a typical acanthocephalan of pinnipeds that can also mature in marine birds (Hernández-Orts et al., 2017). Immature specimens of C. australe have been reported in four species of odontocetes from Argentina (Hernández-Orts et al., 2019). However, this acanthocephalan species is apparently unable to mature in cetaceans (Aznar et al., 2012). Our finding of an immature male in the rectum, suggest that the specimen was passing thorough the gut. We consider that the Clymene dolphin most probably acts as a euparatenic transit hosts for this marine acanthocephalan. Previously, only Carvalho et al. (2010) reported Bolbosoma sp. in the stomach and intestine of Clymene dolphins from Brazil. Therefore, the Clymene dolphin is therefore considered a new host record for C. australe.
The metazoan parasite community of the Clymene dolphin is depauperate, and somewhat similar to that of other delphinids from Argentina (Supplementary Table 3). With the exception of C. grimaldii, T. (T.) forsteri and H. delphini which represent a new locality records, most of the parasites have already been reported in Argentina (Hernández-Orts et al., 2015). Interestingly, two main groups of intestinal helminths that could be predicted for cetaceans worldwide, i.e., cestode species of Tetrabothrius and acanthocephalan species of Bolbosoma (see Aznar et al., 2001), are apparently scarce or absent in delphinids of Argentine waters. For example, T. (T.) forsteri which exhibits high specificity mainly for delphinids (Mateu et al., 2014; Fraija-Fernández et al., 2016), has never been reported in the local dolphin populations off the Argentine coast. It seems plausible that this tapeworm species may not be present in this region of the southwestern Atlantic Ocean. In contrast, Bolbosoma spp. seems to be restricted to coastal waters of the northern region of Argentina, because cystacanths of these acanthocephalans have only been reported in demersal fish from this area (Hernández-Orts et al., 2019). The absence of record of Bolbosoma in odontocetes from Argentina may be related to the fact that very few dolphins from the northern region have been examined for parasites in the past (Supplementary Table 3).
Conclusion
The present study represents the southernmost record for the Clymene dolphin in the Atlantic Ocean, the vagrant dolphin corresponded to an immature female. The current record is very unusual but very valuable, and our investigation has contributed to knowledge of its distribution range, diet and host–parasite relationships. We provided the first molecular identification of a cetacean using molecular data extracted from its metazoan parasites. Amplification of dolphin DNA extracted from its parasites represents a potential non-invasive approach to obtain molecular data for genetic studies of marine mammals. Finally, we provided the first molecular exploration of the metazoan parasite diversity in cetaceans from the southwestern Atlantic.
Data Availability Statement
The skull and the complete skeleton of the Clymene dolphin are deposited in the Colección de Mamíferos Marinos, Centro para el Estudio de Sistemas Marinos (CESIMAR–CONICET), Puerto Madryn, Chubut, Argentina (accession no. CNPMAMM 0793). Helminths are deposited in the Helminthological Collection of the Institute of Parasitology, České Budějovice, Czech Republic (accession nos. IPCAS C-882, C-883, C-884, D-830, N-1252, and N-1253). DNA sequences generated in this study were deposited in the GenBank database (accession nos. MW692916, MW692937–MW692943, MW711662–MW711669, MW714333, MW724482, and MW724483). Further inquiries can be directed to the corresponding author.
Ethics Statement
Ethical review and approval was not required for the animal study because the study was performed using a dead stranded Clymene dolphin submitted for necropsy to the Marine Mammal Laboratory (LAMAMA), Centro para el Estudio de Sistemas Marinos (CESIMAR–CONICET). The collection of the dolphin was approved by the Dirección de Flora, Fauna Silvestre (No. 25/2017-DFyFS) of Chubut, Argentina.
Author Contributions
JH-O, NG, and RL necropsied the dolphin and collected the samples. NG identified the food elements and measured the skull. JH-O and RK performed the morphological identification of the parasites. JH-O, DH-M, and CP contributed to the sequencing and the phylogenetic analyses. JH-O wrote the manuscript. RK, EC, and RL critically reviewed the manuscript. All authors discussed the results, read and approved the final manuscript.
Funding
This study was partially supported by the following projects: Evaluación de efectos denso-dependientes en predadores de alto nivel trófico como consecuencia de cambios ocurridos en el ecosistema marino patagónico: explotación, recuperación y cambio climático (FONCyT 240/16); Reconstruyendo la respuesta histórica a los cambios ambientales en predadores marinos de alto nivel trófico que reproducen en tierra (CONICET 0111/2016); Czech Science Foundation (no. 19-28399X); and the Institute of Parasitology, Biology Center, Czech Academy of Sciences (no. 60077344). JH-O benefited from a postdoctoral fellowship (no. 177603) of the National Council of Science and Technology (CONACYT) of Mexico.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank the Centro para el Estudio de Sistemas Marinos (CESIMAR–CONICET), especially Laboratorio de Mam feros Marinos for logistic and institutional support. We also like to thank to people working in Chubut Coastal Stranding Network who gave notice of the stranding, especially to Leonardo Barattini from the Rada Tilly Conservation Office, Chubut, Argentina for his collaboration in the field and to transport the dolphin to Puerto Madryn. We also thank to Dennis N. Landete for his assistance with parasite collection. Photographs of the skull were made thanks to Cristian Durante. We are in debt with Pesquera Mar del Sur, especially to Javier Tolosano, who kindly froze the dolphin until the necropsy.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2021.658975/full#supplementary-material
Supplementary Figure 1 | Microphotographs of the metazoan parasites of the Clymene dolphin, Stenella clymene. (A) Pholeter gastrophilus (Digenea), stomach; (B) Metacestode of Clistobothrium grimaldii (Cestoda), anal canal; (C) Halocercus delphini (Nematoda), airways; (D) scolex and (E) immature proglottids of Tetrabothrius (Tetrabothrius) forsteri (Cestoda), intestine; (F) phyllobothriid plerocercoid (Cestoda), rectum.
Supplementary Table 1A | Summary data for the isolates used in the molecular analyses.
Supplementary Table 1B | List of phyllobothriid cestodes used in the phylogenetic analysis with data on the host species, developmental stage, locality and GenBank accession number (28S).
Supplementary Table 1C | List of pseudaliid nematodes used in the phylogenetic analysis of the ITS2 dataset with data on the host species, developmental stage, locality and GenBank accession number.
Supplementary Table 1D | List of nematodes of the superfamily Metastrongyloidea used in the phylogenetic analysis with data on the host species, locality, GenBank accession number (28S rDNA) and developmental stage. Abbreviation: L3, third-stage larvae.
Supplementary Table 1E | List of anisakid nematodes used in the phylogenetic analysis with data on the host species, developmental stage, locality and GenBank accession number (cox2). Abbreviation: L3, third-stage larvae; L4, fourth-stage larvae.
Supplementary Table 1F | List of polymorphid acanthocephalans used in the phylogenetic analysis with data on the host species, locality, GenBank accession number (cox1) and developmental stage.
Supplementary Table 2 | Measurements for the skull of the young immature female Clymene dolphin, Stenella clymene stranded in Rada Tilly, Chubut, Argentina. Measurements in mm.
Supplementary Table 3 | Helminth fauna of delphinids off the Argentinean coasts.
References
Aguilar-Aguilar, R., Delgado-Estrella, A., and Moreno-Navarrete, R. G. (2010). New host report for nematodes in a stranded short-snouted spinner dolphin Stenella clymene (Cetacea: Delphinidae) from the Mexican Caribbean coast. Helminthologia 47, 136–138. doi: 10.2478/s11687-010-0020-0
Agustí, C., Aznar, F. J., and Raga, J. A. (2005). Tetraphyllidean plerocercoids from Western Mediterranean cetaceans and other marine mammals around the world: a comprehensive morphological analysis. J. Parasitol. 91, 83–92. doi: 10.1645/GE-372R
Anderson, R. C., Chabaud, A. G., and Willmott, S. (2009). Keys to the Nematode Parasites of Vertebrates: Archival volume. Wallingford: CAB International.
Azevedo, M. I. N., Carvalho, V. L., and Iniguez, A. M. (2017). Integrative taxonomy of anisakid nematodes in stranded cetaceans from Brazilian waters: an update on parasite’s hosts and geographical records. Parasitol. Res. 108, 3105–3116. doi: 10.1007/s00436-017-5622-8
Aznar, F. J., Agustí, C., Littlewood, D. T. J., Raga, J. A., and Olson, P. D. (2007). Insight into the role of cetaceans in the life cycle of the tetraphyllideans (Platyhelminthes: Cestoda). Int. J. Parasitol. 37, 243–255. doi: 10.1016/j.ijpara.2006.10.010
Aznar, F. J., Balbuena, J. A., Fernández, M., and Raga, J. A. (2001). “Living together: the parasites of marine mammals,” in Marine Mammals: Biology and Conservation, eds P. G. H. Evans and J. A. Raga (New York, NY: Kluwer Academic/Plenum Publishers), 385–421.
Aznar, F. J., Fognani, P., Balbuena, J. A., Pietrobelli, M., and Raga, J. A. (2006). Distribution of Pholeter gastrophilus (Digenea) within the stomach of four odontocete species: the role of the diet and digestive physiology of hosts. Parasitology 133, 369–380. doi: 10.1017/S0031182006000321
Aznar, F. J., Hernández-Orts, J., Suárez, A. A., García-Varela, M., Raga, J. A., and Cappozzo, H. L. (2012). Assessing host-parasite specificity through coprological analysis: a case study with species of Corynosoma (Acanthocephala: Polymorphidae) from marine mammals. J. Helminthol. 86, 156–164. doi: 10.1017/S0022149X11000149
Barón, P. J. (2001). First description and survey of the egg masses of Loligo gahi D’Orbigny, 1835, and Loligo sanpaulensis Brakoniecki, 1984, from coastal waters of Patagonia. J. Shellfish Res. 20, 289–295.
Boschi, E. E., Fischbach, C. E., and Iorio, M. I. (1992). Catálogo ilustrado de los crustáceos estomatópodos y decápodos marinos de Argentina. Frente Marítimo 10, 7–94.
Bray, R. A., Gibson, D. I., and Jones, A. (2008). Key to the Trematoda, Vol. 3. Wallingford: CAB International and Natural History Museum, London.
Briceño, Y., Ramírez, S., Bolaños-Jimenez, J., Sánchez, L., Bermúdez-Villapol, L., González, M., et al. (2020). A live-stranding of a Clymene dolphin (Stenella clymene, Gray, 1850) in the Gulf of Venezuela: first record for the southern Caribbean. Lat. Am. J. Aquat. Mamm. 15, 33–36. doi: 10.5597/lajam00260
Caira, J. N., Jensen, K., Pickering, M., Ruhnke, T. R., and Gallagher, K. A. (2020). Intrigue surrounding the life-cycles of species of Clistobothrium (Cestoda: Phyllobothriidea) parasitising large pelagic sharks. Int. J. Parasitol. 50, 1043–1055. doi: 10.1016/j.ijpara.2020.08.002
Carreno, R. A., and Nadler, S. A. (2003). Phylogenetic analysis of the Metastrongyloidea (Nematoda: Strongylida) inferred from ribosomal RNA gene sequences. J. Parasitol. 89, 965–973. doi: 10.1645/GE-76R
Carvalho, V. L., Bevilaqua, C. M. L., Iñiguez, A. M., Mathews-Cascon, H., Ribeiro, F. B., Pessoa, L. M. B., et al. (2010). Metazoan parasites of cetaceans off the northeastern coast of Brazil. Vet. Parasitol. 173, 116–122. doi: 10.1016/j.vetpar.2010.06.023
Clarke, M. R. (1986). A handbook for the identification of cephalopods beaks. Oxford: Clarendon Press.
Costa, A. F., Siciliano, S., Emin-Lima, R., Martins, B. M. L., Sousa, M. E. M., Giarrizzo, T., et al. (2017). Stranding survey as a framework to investigate rare cetacean records of the north and north-eastern Brazilian coasts. ZooKeys 688, 111–134. doi: 10.3897/zookeys.688.12636
Darriba, D., Taboada, G. L., Doallo, R., and Posada, D. (2012). jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 9:772. doi: 10.1038/nmeth.2109
Delyamure, S. L. (1955). Helminthofauna of marine mammals: ecology and phylogeny. Moscow: Akademiya Nauk SSSR.
Evans, P. G., and Bjørge, A. (2013). Impacts of climate change on marine mammals. MCCIP Sci. Rev. 2013, 134–148. doi: 10.14465/2013.arc15.134-148
Fernández, M., Berón−Vera, B., García, N. A., Raga, J. A., and Crespo, E. A. (2003). Food and parasites from two hourglass dolphins, Lagenorhynchus cruciger (Quoy and Gaimard, 1824), from Patagonian waters. Mar. Mam. Sci. 19, 832–836. doi: 10.1111/j.1748-7692.2003.tb01133.x
Fertl, D., Jefferson, T. A., Moreno, I. B., Zerbini, A. N., and Mullin, K. D. (2003). Distribution of the Clymene dolphin Stenella clymene. Mammal Rev. 33, 253–271. doi: 10.1046/j.1365-2907.2003.00033.x
Fraija-Fernández, N., Fernández, M., Lehnert, K., Raga, J. A., Siebert, U., and Aznar, F. J. (2017). Long-distance travellers: phylogeography of a generalist parasite, Pholeter gastrophilus, from cetaceans. PLoS One 12:e0170184. doi: 10.1371/journal.pone.0170184
Fraija-Fernández, N., Fernández, M., Raga, J. A., and Aznar, F. J. (2016). “Helminth diversity of cetaceans: an update,” in Advances in Marine Biology, eds A. Kovács and P. Nagy (Hauppauge: Nova Science Publishers), 29–100.
García-Varela, M., Masper, A., Crespo, E. A., and Hernández-Orts, J. S. (2021). Genetic diversity and phylogeography of Corynosoma australe Johnston, 1937 (Acanthocephala: Polymorphidae), an endoparasite of otariids from the Americas in the northern and southern hemispheres. Parasitol. Int. 80:102205. doi: 10.1016/j.parint.2020.102205
Geraci, J. R., and Aubin, D. J. (1987). Effects of parasites on marine mammals. Int. J. Parasitol. 17, 407–414. doi: 10.1016/0020-7519(87)90116-0
Gosztonyi, A., and Kuba, L. (1996). Atlas de huesos craneales y de la cintura escapular de peces costeros patagónicos. Informes Técnicos del Plan de Manejo Integrado de la Zona Costera Patagónica – Fund. Patagonia Nat. 4, 1–29.
Guimarães, J. P., Febronio, A. M. B., Vergara-Parente, J. E., and Werneck, M. R. (2015). Lesions associated with Halocercus brasiliensis Lins de Almeida, 1933 in the lungs of dolphins stranded in the Northeast of Brazil. J. Parasitol. 101, 248–251. doi: 10.1645/14-513.1
Hatfield, E. M. C., Rodhouse, P. G., and Porebski, J. (1990). Demography and distribution of the Patagonian squid (Loligo gahi d’Orbigny) during the austral winter. ICES J. Mar. Sci. 46, 306–312. doi: 10.1093/icesjms/46.3.306
Hermosilla, C., Silva, L. M. R., Prieto, R., Kleinertz, S., Taubert, A., and Silva, M. A. (2015). Endo- and ectoparasites of large whales (Cetartiodactyla: Balaenopteridae, Physeteridae): Overcoming difficulties in obtaining appropriate samples by non- and minimally-invasive methods. Int. J. Parasitol. Parasites Wildl. 4, 414–420. doi: 10.1016/j.ijppaw.2015.11.002
Hernández-Mena, D., Lynggaard, C., Mendoza-Garfias, B., and Pérez-Ponce de León, G. (2016). A new species of Auriculostoma (Trematoda: Allocreadiidae) from the intestine of Brycon guatemalensis (Characiformes: Bryconidae) from the Usumacinta River basin, Mexico, based on morphology and 28S rDNA sequences, with a key to species of the genus. Zootaxa 4196, 261–277. doi: 10.11646/zootaxa.4196.2.5
Hernández-Orts, J. S., Brandão, M., Georgieva, S., Raga, J. A., Crespo, E. A., Luque, J. L., et al. (2017). From mammals back to birds: Host-switch of the acanthocephalan Corynosoma australe from pinnipeds to the Magellanic penguin Spheniscus magellanicus. PLoS One 12:e0183809. doi: 10.1371/journal.pone.0183809
Hernández-Orts, J. S., Montero, F. E., García, N. A., Crespo, E. A., Raga, J. A., García-Varela, M., et al. (2019). Transmission of Corynosoma australe (Acanthocephala: Polymorphidae) from fishes to South American sea lions Otaria flavescens in Patagonia, Argentina. Parasitol. Res. 118, 433–440. doi: 10.1007/s00436-018-6177-z
Hernández-Orts, J. S., Paso Viola, M. N., García, N. A., Crespo, E. A., González, R., et al. (2015). A checklist of the helminth parasites of marine mammals from Argentina. Zootaxa 3936, 301–334. doi: 10.11646/zootaxa.3936.3.1
Jefferson, T. A. (2018). “Clymene dolphin – Stenella clymene,” in Encyclopedia of Marine Mammals, 3rd Edn, eds B. Würsig, J. G. M. Thewissen, and K. Kovacs (Burlington: Academic Press), 197–200. doi: 10.1016/B978-0-12-804327-1.00093-5
Jefferson, T. A., and Curry, B. E. (2003). Stenella clymene. Mamm. Species 726, 1–5. doi: 10.1644/726
Jefferson, T. A., Webber, M. A., and Pitman, R. L. (2015). Marine mammals of the World. A comprehensive guide to their identification. London: Academic Press.
Jensen, K., and Bullard, S. A. (2010). Characterization of a diversity of tetraphyllidean and rhinebothriidean cestode larval types, with comments on host associations and life-cycles. Int. J. Parasitol. 40, 889–910. doi: 10.1016/j.ijpara.2009.11.015
Katoh, K., and Standley, D. M. (2013). MAFFT Multiple Sequence Alignment Software Version 7: Improvements in performance and usability. Mol. Biol. Evol. 30, 772–780. doi: 10.1093/molbev/mst010
Klotz, D., Hirzmann, J., Bauer, C., Schöne, J., Iseringhausen, M., Wohlsein, P., et al. (2018). Subcutaneous merocercoids of Clistobothrium sp. in two Cape fur seals (Arctocephalus pusillus pusillus). Int. J. Parasitol. Parasites Wildl. 7, 99–105. doi: 10.1016/j.ijppaw.2018.02.003
Koen Alonso, M., Crespo, E. A., García, N. A., Pedraza, S. N., and Coscarella, M. A. (1998). Diet of dusky dolphins, Lagenorhynchus obscurus, in waters off Patagonia, Argentina. Fish. Bull. 96, 366–374.
Koen Alonso, M., Crespo, E. A., García, N. A., Pedraza, S. N., Mariotti, P. A., Berón Vera, B., et al. (2001). Food habits of Dipturus chilensis (Pisces: Rajidae) off Patagonia, Argentina. J. Mar. Sci. 58, 288–297. doi: 10.1006/jmsc.2000.1010
Kumar, S., Stecher, G., Li, M., Knyaz, C., and Tamura, K. (2018). MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 35, 1547–1549. doi: 10.1093/molbev/msy096
Lanfranchi, A. L., Braicovich, P. E., Cantatore, D. M. P., Irigoitia, M. M., Farber, M. D., Taglioretti, V., et al. (2018). Influence of confluent marine currents in an ecotonal region of the South-West Atlantic on the distribution of larval anisakids (Nematoda: Anisakidae). Parasit. Vectors 8:583. doi: 10.1186/s13071-018-3119-7
Lisitsyna, O. I., Kudlai, O., Cribb, T. H., and Smit, N. J. (2019). Three new species of acanthocephalans (Palaeacanthocephala) from marine fishes collected off the East Coast of South Africa. Folia Parasitol. 66:012. doi: 10.14411/fp.2019.012
Loizaga de Castro, R., Leonardi, M. S., Grandi, M. F., García, N. A., and Crespo, E. A. (2011). Far from home: Record of a vagrant striped dolphin in Patagonia with notes on diet, parasites and age determination. Mamm. Biol. 76, 521–524. doi: 10.1016/j.mambio.2011.02.004
Loizaga de Castro, R., Saporiti, F., Vales, D. G., García, N. A., Cardona, L., et al. (2016). What are you eating? A stable isotope insight into the trophic ecology of short-beaked common dolphins in the Southwestern Atlantic Ocean. Mamm. Biol. 81, 571–578. doi: 10.1016/j.mambio.2016.07.003
MacLeod, C. D. (2009). Global climate change, range changes and potential implications for the conservation of marine cetaceans: a review and synthesis. Endanger. Species Res. 7, 125–136. doi: 10.3354/esr00197
Mariaux, J., Kuchta, R., and Hoberg, E. P. (2017). “Tetrabothriidea Baer, 1954” in Planetary biodiversity inventory (2008–2017): Tapeworms from vertebrate bowels of the Earth, eds J. N. Caira, and K Jensen K (Lawrence: University of Kansas, Natural History Museum, Special Publication No. 25). Lawrence: University of Kansas. 357–370.
Mateu, P., Raga, J. A., Fernández, M., and Aznar, F. J. (2014). Intestinal helminth fauna of striped dolphins (Stenella coeruleoalba) in the western Mediterranean: No effects of host body length, age and sex. Mar. Mam. Sci. 30, 961–977. doi: 10.1111/mms.12096
Mayorga, L. F. S. P., Vanstreels, R. E. T., Bhering, R. C. C., Mamede, N., Costa, L. M. B., Pinheiro, F. C. F., et al. (2020). Strandings of cetaceans on the Espírito Santo coast, southeast Brazil, 1975–2015. ZooKeys 948, 129–152. doi: 10.3897/zookeys.948.50468
Miura, O., Kuris, A. M., Torchin, M. E., Hechinger, R. F., Dunham, E. J., and Chiba, S. (2005). Molecular-genetic analyses reveal cryptic species of trematodes in the intertidal gastropod, Batillaria cumingi (Crosse). Int. J. Parasitol. 35, 793–801. doi: 10.1016/j.ijpara.2005.02.014
Mladineo, I., Hrabar, J., Vrbatović, A., Duvnjak, S., Gomerčić, T., and Ðuras, M. (2019). Microbiota and gut ultrastructure of Anisakis pegreffii isolated from stranded cetaceans in the Adriatic Sea. Parasit. Vectors 12:381. doi: 10.1186/s13071-019-3636-z
Moreno, B. I., Zerbini, A. N., Danilewicz, D., de Oliveira Santos, M. C., Simões-Lopes, P. C., and Lailson-Brito, J. Jr., et al. (2005). Distribution and habitat characteristics of dolphins of the genus Stenella (Cetacea: Delphinidae) in the southwest Atlantic Ocean. Mar. Ecol. Prog. Ser. 300, 229–240. doi: 10.3354/meps300229
Nadler, S. A., and Hudspeth, D. S. S. (2000). Phylogeny of the Ascaridoidea (Nematoda: Ascaridida) based on three genes and morphology: hypotheses of structural and sequence evolution. J. Parasitol. 86, 380–393. doi: 10.1645/0022-33952000086[0380:POTANA]2.0.CO;2
Nara, L., Meirelles, A. C. O., Souto, L. R. A., Silva, J. M. J., and Farro, A. P. C. (2017). An initial population structure and genetic diversity analysis for Stenella clymene (Gray, 1850): evidence of differentiation between the North and South Atlantic Ocean. Aquat. Mamm. 43, 507–516. doi: 10.1578/AM.43.5.2017.507
Norris, K. S. (1961). Standardized methods for measuring and recording data on the smaller cetaceans. J. Mammal. 42, 471–476. doi: 10.2307/1377364
Pájaro, M., Macchi, G. J., and Martos, P. (2005). Reproductive pattern of the Patagonian stock of Argentine hake (Merluccius hubbsi). Fish. Res. 72, 97–108. doi: 10.1016/j.fishres.2004.09.006
Pis-Millán, J. A., Roselló, E., Morales-Muñiz, A., and Nores, C. (2019). First record of Clymene dolphin (Stenella clymene Gray, 1846) in European waters. Galemys 31, 83–88. doi: 10.7325/Galemys.2019.N3
Pleijel, F., Jondelius, U., Norlinder, E., Nygren, A., Oxelman, B., Schande, C., et al. (2008). Phylogenies without roots? A plea for the use of vouchers in molecular phylogenetic studies. Mol. Phylogenet. Evol. 48, 369–271. doi: 10.1016/j.ympev.2008.03.024
Poloczanska, E. S., Brown, C. J., Sydeman, W. J., Kiessling, W., Schoeman, D. S., Moore, P. J., et al. (2013). Global imprint of climate change on marine life. Nat. Clim. Change 3, 919–925. doi: 10.1038/nclimate1958
Pons-Bordas, C., Hazenberg, A., Hernandez-Gonzalez, A., Pool, R. V., Covelo, P., Sánchez-Hermosin, P., et al. (2020). Recent increase of ulcerative lesions caused by Anisakis spp. in cetaceans from the north-east Atlantic. J. Helminthol. 94:e127. doi: 10.1017/S0022149X20000115
Pool, R., Fernández, M., Chandradeva, N., Raga, J. A., and Aznar, F. J. (2020). The taxonomic status of Skrjabinalius guevarai Gallego & Selva, 1979 (Nematoda: Pseudaliidae) and the synonymy of Skrjabinalius Delyamure, 1942 and Halocercus Baylis & Daubney, 1925. Syst. Parasitol. 97, 389–401. doi: 10.1007/s11230-020-09921-9
Prado, J. H. F., Mattos, P. H., Silva, K. G., and Secchi, E. R. (2016). Long-term seasonal and interannual patterns of marine mammal strandings in subtropical western South Atlantic. PLoS One 11:e0146339. doi: 10.1371/journal.pone.0146339
Rambaut, A. (2018). Figtree (Version 1.4.4). Available online at https://github.com/rambaut/figtree/releases (accessed date 15 January 2021).
Simões-Lopes, P. C., Praderi, P., and Paula, G. S. (1994). The Clymene dolphin, Stenella clymene (Gray, 1846), in the southwestern South Atlantic Ocean. Mar. Mammal Sci. 10, 213–217. doi: 10.1111/j.1748-7692.1994.tb00263.x
Stamatakis, A. (2006). RAxML-VI-HPC: Maximum likelihood based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 22, 2688–2690. doi: 10.1093/bioinformatics/btl446
Temirova, S. I., and Skrjabin, A. S. (1978). The suborder Tetrabothriata (Ariola, 1899) Skrjabin, 1940. Osnovy Tsestodologii 9, 7–117.
Weir, C. R. (2006). First confirmed records of Clymene dolphin, Stenella clymene (Gray, 1850), from Angola and Congo, south-east Atlantic Ocean. Afr. Zool. 41, 297–300. doi: 10.1080/15627020.2006.11407366
Weir, C. R., Coles, P., Ferguson, A., May, D., Baines, M., Figueirdo, I., et al. (2014). Clymene dolphins (Stenella clymene) in the eastern tropical Atlantic: distribution, group size, and pigmentation pattern. J. Mammal. 95, 1289–1298. doi: 10.1644/14-MAMM-A-115
Keywords: Delphinidae, southwestern Atlantic, Merluccius hubbsi, Trematoda, Cestoda, Nematoda, Acanthocephala, life-cycle
Citation: Hernández-Orts JS, Hernández-Mena DI, Pantoja C, Kuchta R, García NA, Crespo EA and Loizaga R (2021) A Visitor of Tropical Waters: First Record of a Clymene Dolphin (Stenella clymene) Off the Patagonian Coast of Argentina, With Comments on Diet and Metazoan Parasites. Front. Mar. Sci. 8:658975. doi: 10.3389/fmars.2021.658975
Received: 26 January 2021; Accepted: 10 March 2021;
Published: 15 April 2021.
Edited by:
Salvatore Siciliano, Fundação Oswaldo Cruz (Fiocruz), BrazilReviewed by:
Juan Antonio Raga, University of Valencia, SpainPaulo Simões-Lopes, Federal University of Santa Catarina, Brazil
Copyright © 2021 Hernández-Orts, Hernández-Mena, Pantoja, Kuchta, García, Crespo and Loizaga. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jesús S. Hernández-Orts, amVzdXMuaGVybmFuZGV6Lm9ydHNAZ21haWwuY29t