 Lorena Viloria-Gómora
Lorena Viloria-Gómora Jorge Urbán R
Jorge Urbán R Braulio Leon-Lopez
Braulio Leon-Lopez Eduardo Romero-Vivas
Eduardo Romero-Vivas- 1Departamento de Ciencias Marinas y Costeras, Universidad Autónoma de Baja California Sur, La Paz, Mexico
- 2Grupo de Investigación en Acústica y Procesamiento de Señales, Centro de Investigaciones Biológicas del Noroeste, La Paz, Mexico
Although Bryde’s whale (Balaenoptera edeni) has worldwide distribution, it is considered one of the most difficult species of baleen whales to study. Two populations existing in the Gulf of California have been proposed, but difficulties in obtaining biopsies have complicated testing the hypothesis of discrete stock difference between north and south of the Gulf. On the other hand, bioacoustics analyses have been used to differentiate populations from other whale species. Therefore, to provide information for the previously proposed hypothesis, the most common Bryde’s call (Be4) in the region was analyzed. Acoustic data was collected using a High-frequency Acoustic Recording Package (HARP) system placed at two locations in the Gulf of California: one in the north from November 2006 to June 2007 and other one in the south from August to December 2008. Duration and frequency range were measured for calls with a signal-to-noise ratio (SNR) greater than 27 dB. The Mann–Whitney-U test was performed to compare distributions in both geographic areas, and the results were compared with a photo-identification analysis. Of the 903 Be4 calls detected, 103 (north) and 92 (south) met the SNR requirement. Duration showed a significant difference (Mann–Whitney test, U = 11238, z-statistics = 2.91, p = 0.003) with southern calls being longer than the northern ones. The results suggest that the northern population remains resident in the north with some movement toward the south, also verified by photo-identification. A detailed inspection showed a double distribution in the south (Mann–Whitney test, U = 2211, z-statistics = −8.39, p = 4.4776e-17), suggesting the presence at the same time of resident and transient populations. The latter has been suggested to belong to the Eastern Tropical Pacific. The results suggest a geographic variation in B. edeni Be4 call type and support the existence of two populations within the Gulf of California.
Introduction
Bryde’s whale (Balaenoptera edeni) -considered one of the least known Balaenopteridae family members (Cummings, 1985; Kato and Perrin, 2009; International Whale Commission, 2020) and one of the most difficult mysticete species to study (Constantine et al., 2018)- has a worldwide distribution inhabiting mainly temperate and tropical oceans. Furthermore, its taxonomical status has been controversial since no agreement exists whether B. edeni includes two subspecies (Balaenoptera edeni edeni and Balaenoptera edeni brydei) or they are two different species (Kato and Perrin, 2009; Committee on Taxonomy, 2020). Bryde’s whales due to similar coloration, dorsal fin and body shape could be confused at sea with sei whales (Balaenoptera borealis) or young fin whales (Balaenoptera physalus); particularly in the western Pacific, confusion might happen with Omura’s whales (Balaenoptera omurai) (Jefferson et al., 2008; Kato and Perrin, 2009).
Unlike some baleen whales, such as humpback or gray whales, the information about Bryde’s whale ecology and behavior is limited (Jefferson et al., 2008). Migration movements and main breeding season are unknown (Leatherwood et al., 1988; Kato and Perrin, 2009; Reeves et al., 2002). The scarce existing information mentions that Bryde’s whales may congregate in groups of as many as 10–20 individuals when they feed (Jefferson et al., 2008).
Many researchers attest to the difficulties of achieving a close approach at sea to record precise behavioral observations (Cummings et al., 1986). Furthermore, biopsies have been only taken after long follows and continuous efforts to approach the animals (Pers. Obs. LVG, JUR). Therefore, Bryde’s whale can be described as an elusive species.
Bryde’s whales have been visually detected all-year-round in the Gulf of California (Urbán and Flores-Ramírez, 1996). Photo-identification, genetic, and stable isotopes (Breese and Tershy, 1987; Dizon et al., 1995; Niño-Torres et al., 2013) have suggested the existence of discrete stocks. Studies have proposed that two populations can be distinguished, one in the northern part consisting of mainly resident individuals and another one in the south primarily consisting of transient individuals from the Pacific Ocean (Breese and Tershy, 1987; Vidal et al., 1987; Urbán and Flores-Ramírez, 1996). Nevertheless, difficulties in obtaining biopsies and continuous unsuccessful sightings have hindered testing this hypothesis. Therefore, alternative methods are required, and fixed-point passive acoustic monitoring can be a useful technique since it is non-invasive and would not modify whales’ behavior. These systems have shown effectiveness for studying several aspects, such as species presence, abundance, distribution, and population identity (Mellinger and Barlow, 2003; Au and Hastings, 2008). Unlike traditional markers (molecular and photo-identification), vocalization variation can reflect recent and fast changes in whale populations, such as movements or association patterns (McDonald et al., 2006). Some researchers have suggested that using vocalizations could be a more effective tool for identifying population stocks (Mellinger and Barlow, 2003; Delarue et al., 2009; Rendell et al., 2011).
Song geographic variation in whale species has been reported in humpback whales (Megaptera novaeangliae) and used as stock indicator (Payne, 1979). Moreover, call variation patterns have been used to identify populations of fin whales (Balaenoptera physalus) (Hatch and Clark, 2004) and stable songs depending on the geographical region in blue whales (Balaenoptera musculus) (McDonald et al., 2006). Additionally, variations within populations in the same geographical area can be found in odontocetes, such as killer and sperm whales (Baird, 2000; Rendell and Whitehead, 2003).
Currently, there is a lack of information about the acoustic repertoire of Bryde’s whales. Vocal repertoire characteristics are still being studied, and most knowledge remains limited (Constantine et al., 2018). Even though they are vocally active, the low amount of calls and low production rate detected in a previous work in the Gulf of California (Viloria-Gómora et al., 2015) suggest that this species might be less vocally active compared to other baleen whales. For example, while Miller et al. (2014) recorded 1617 Blue whale calls in 130 h recording with visual confirmation (12.7 calls/h), Viloria-Gómora et al. (2015) reported 23 Bryde’s whales calls in 18.2 h recordings with visual confirmation (1.3 calls/h). Moreover, there are reports of events where individuals stopped vocalizing once vessels approached (Cummings et al., 1986) which complicates obtaining acoustic recording.
The first call description for the species comes from opportunistic recordings with visual confirmation in the north and central parts of the Gulf of California (Cummings et al., 1986; Edds et al., 1993). Oleson et al. (2003) described vocalizations recorded in the Eastern Tropical Pacific (including the southwest area of the Baja California peninsula), Caribbean Sea, and Japanese coast. Furthermore, Heimlich et al. (2005) performed passive acoustic monitoring in the Eastern Tropical Pacific and identified Bryde’s whale vocalizations in accordance with spectrograms published by Oleson et al. (2003). Both studies identified a relationship between call characteristics and geographical location. McDonald et al. (2006), identified a seasonal pattern in Bryde’s whale’s vocalizations at the Great Barrier Island, New Zealand and suggested that the variation might be related with the presence of different species from the B. edeni-brydei complex or due to different populations of the same species. In the Atlantic Ocean, a description of just a few calls was recorded during visual confirmation in the Gulf of Mexico (Rice et al., 2014; Širović et al., 2014), as well as other few calls off the coast of Brazil (Figueiredo and Simão, 2014), Tosa Bay, Japan (Chiu, 2009), and in the Gulf of California (Viloria-Gómora et al., 2015). Recent studies on Bryde’s whale acoustics have been conducted in Australia (Erbe et al., 2017), New Zealand (Putland et al., 2018a,b), and South Africa (Ogundile and Versfeld, 2020; Ogundile et al., 2020).
Complications that have arisen while studying Bryde’s whales (evasive species, difficult to distinguish at sea, and probably less vocally active than other species) may explain the scarcity of research projects targeting this species specifically, and why most current information about them comes from opportunistic encounters.
Therefore, the aim of this study was to record calls and identify if a call (Be4) type was different between the north and south populations of the Gulf of California, testing the hypothesis of discrete stocks. The acoustics analysis was compared with a photo-identification analysis between both regions to support the conclusions.
Materials and Methods
Data Collection
Acoustic data was collected using a stationary HARP (High-frequency Acoustic Recording Package) system designed and deployed by Scripps Institution of Oceanography at the University of California, United States consisting of a hydrophone [Transducers AQ-1s and ITC-1042 (10–100 kHz response)] sensitivity −193 dB VRMS/μPa, 40 dB preamp (Wiggins, 2003), batteries, and memory disks, everything contained on polyvinyl chloride (PVC) sealed cylinders. Recordings were obtained at a frequency sample of 192.6 kHz. The equipment was programmed to take time intervals of 5-min recording and 10-min rest (8 h each day), for 6 months (due to battery life and memory capacity).
The recording system was moored at 700 m depth in two locations in the Gulf of California: (1) Punta Pescadero (23° 48.448′ N, 109° 37.687′ W) from December 2006 to June 2007 and (2) Bahía de los Ángeles (29° 01.750′ N, 113° 22.474′ W) from August to December 2008 (Figure 1).
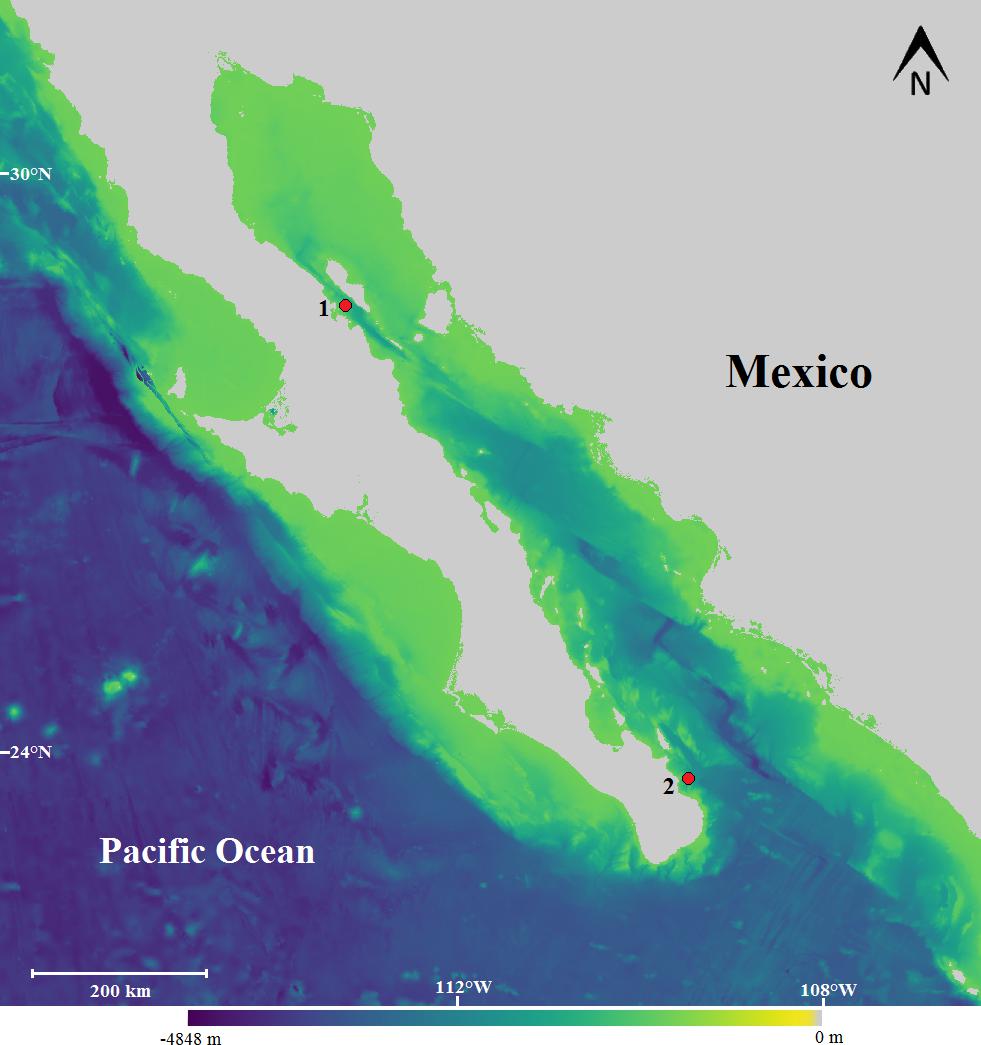
Figure 1. Location of the recording sites of Bryde’s whale calls in the north from November 2006 to June 2007 and south from August to December 2008 within the Gulf of California. (1) Bahía de los Ángeles, Baja California and (2) Punta Pescadero, Baja California Sur, Mexico. Bathymetry is also shown.
Field recordings from small boats were taken at the same sites from 2010 to 2014 to obtain vocalizations with visual confirmation. The effort was similar in both sites (38 surveys in Bahía de Los Ángeles and 43 surveys in Punta Pescadero). Every time a sighting of the species was identified, the engine was switched off, and a hydrophone [transducers AQ-1 and ITC-1042 (10 Hz–100 kHz)] sensitivity −193 dB VRMS/μPa, 40 dB preamp (Wiggins, 2003) -connected to a Fostex FR2 recorder- was submerged 10 m. The frequency sample was the same as in the moored systems (192.6 kHz) (Viloria-Gómora et al., 2015). From 109 audio recordings a total of 23 calls were detected. When those calls were analyzed, four call types were confirmed: three new in the southern region (Be10, Be11, and Be12), described in Viloria-Gómora et al. (2015), and in the northern region, the Be4 call already described by Oleson et al. (2003). The calls obtained during visual confirmation were exclusively used to compare and give confidence when searching for calls from the moored HARP systems (Figure 2). Additionally, spectrograms published (Cummings et al., 1986; Edds et al., 1993; Oleson et al., 2003; Heimlich et al., 2005; Figueiredo et al., 2014; Rice et al., 2014; Širović et al., 2014) were used to identify Bryde’s whale call types.

Figure 2. Call example (type Be4) of Bryde’s whale from the Gulf of California, Mexico. Recorded during visual confirmation (left) and recorded from the HARP (right). Spectrogram: Hamming window size 2048, 85% overlap, and a frame of 4096.
Data Analysis
Only data from the HARP were used for further analyses. The HARP audio files were decimated to 2 kHz sampling frequency. They were used to generate long term spectral average (LTSA) spectrograms, using 5 s of temporal resolution and a frequency band 10–1000 Hz (Wiggins and Hildebrand, 2007). LTSA were revised, searching for Bryde’s whale calls using Triton (v 1.64.20070724, SCRIPPS) running on Matlab® (Mathworks Inc., United States) (Matlab Signal Processing Toolbox v 6.2). All files were revised visually, as using current popular automatic detectors might lead to false positives or negatives (Putland et al., 2018b), especially for poorly understood whale species.
Only two types of calls were confirmed in HARP recordings, Be11 and Be4, consisting of 0.5% (1/196 calls) and 99.5% (195/196 calls) of the amount of calls, respectively. Those calls identified as type Be4 -consisting of a pulse around 60 Hz with harmonic structure (Figure 2; Oleson et al., 2003; Kerosky et al., 2012)- were separated. The analysis focused on this call type as it was the most commonly found in both regions of the Gulf of California. Moreover, this call has also been reported for other areas in the northeast Pacific (Kerosky et al., 2012).
The typical 2D spectrogram representation does not necessarily show clear differences among calls. Therefore, calls were inspected using a 3D visualization, which can provide more information. Spectrogram representation as a surface plot can help to determine amplitude characteristics aiding visualization and recognize important features in a signal (Romero-Vivas et al., 2013; Figure 3).
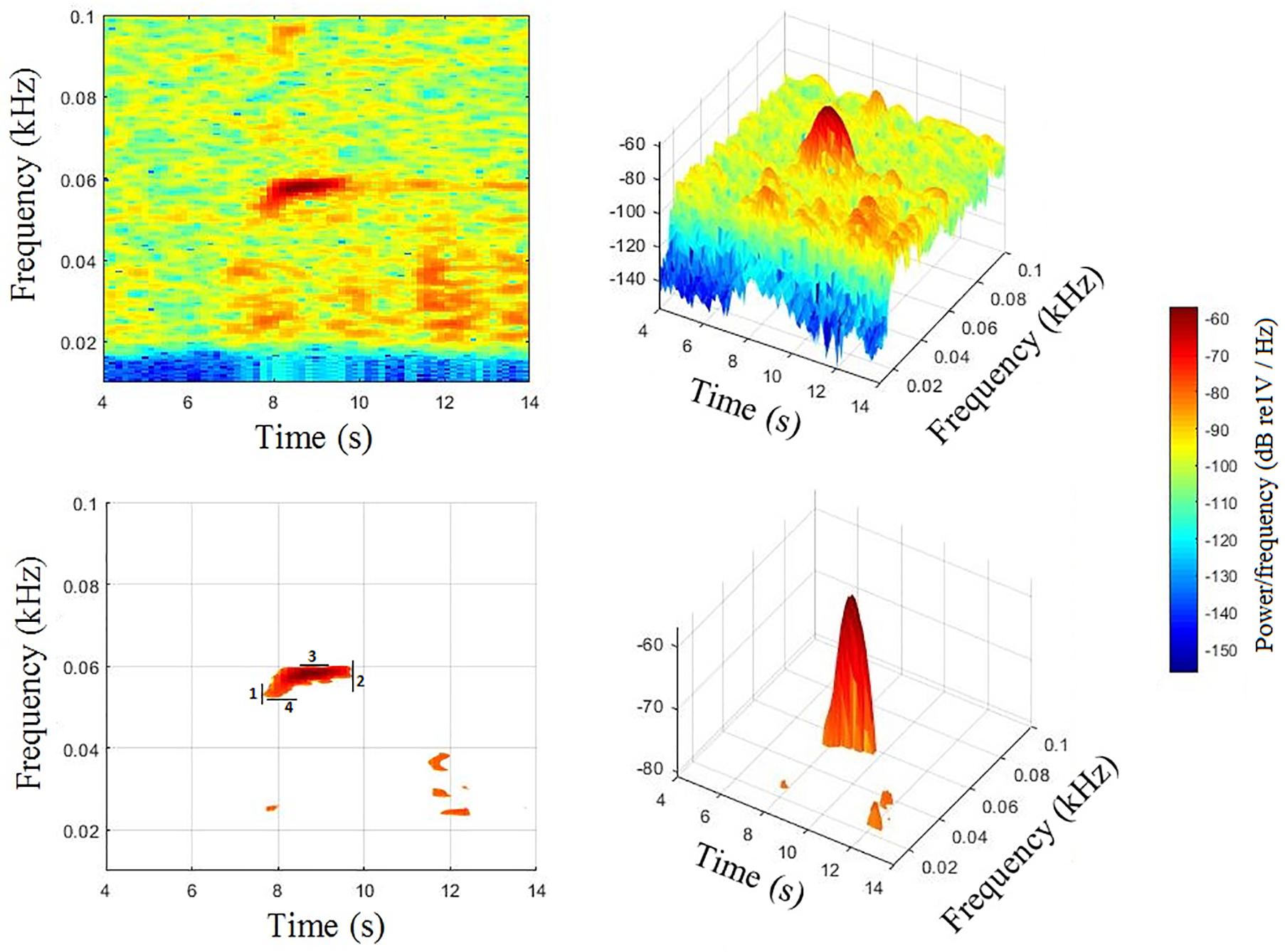
Figure 3. Call example (type Be4) of Bryde’s whale from the Gulf of California, Mexico. Spectrogram 2D (top left), spectrogram 3D (top right), spectrogram 3D threshold at 24 dB from the maximum amplitude value (bottom right), and spectrogram 2D threshold at 24 dB from the maximum amplitude value (bottom left) showing the clear limits of the call: start (1), end (2), maximum (3), and minimum (4). Spectrogram: Butterworth (2nd order) high-pass filter at 40 Hz cut off, Hamming window size 2048, 85% overlap, and a frame of 4096.
The main 60 Hz component -characteristic of Be4- has shown morphological differences when viewed as a 3D representation. Calls from the south showed intensity spread over a longer duration compared to the north calls. However, an apparent spread of intensity over a wider frequency band seemed to be shown in the north calls (Romero-Vivas et al., 2013).
Signals were first classified by their SNR computed in the time domain. There were no high intensity background signals in the same frequency band as the Be4 call in the sites (Figure 4); low signal-to-noise ratios were due to low intensity signals rather than high intensity background noise (Vera-Cuevas, 2016). Range of detection was computed using Bryde’s Source Level [SL = 152 dB ref 1 μPa at 1 m (Cummings, 1985)], and the Noise level shown in Figure 4 (91 dB RMS), using the SONAR equation (SNR = SL -TL -NL). For a signal just detectable (SNR = 1 dB), the maximum Transmission Loss (TL) was 61 dB. Considering geometrical loss and absorption the maximum range was 6.3 km for the sites of deployment (800 m depth). This range, the complex bathymetry, and the sinuous coastline (Figure 1) let us assume that only calls from the surrounding area were considered for the analysis since the distance between sites was more than 700 km. Bearing that a hydrophone frequency response is considered flat within a 3 dB variation, the first 3 dB were considered as the noise floor. A Be4 signal was classified as detectable but of Low quality if its mean intensity was less than double (3 dB) the noise floor (3 dB) yielding a 6 dB SNR threshold. Similarly SNR thresholds were defined following this 3 dB criteria, yielding a range of 6–9 dB for Medium quality signals and >9 dB for High quality signals. The group of High quality signals was band pass filtered (3rd order high-pass Butterworth 45 Hz cut off, 3rd order Butterworth low-pass 70 Hz cut off) to extract the 60 Hz Be4 component. SNR was measured again in the frequency domain, and the signal value with the lowest SNR (in our case 27 dB) was used to set a cutting threshold 3 dB higher, giving the 24 dB cutting threshold applied to all high quality signals for the acoustic parameter measurements (Figure 3). Duration and frequency range were measured directly on Matlab® (Mathworks Inc., United States), calculating each spectrogram with a Hamming window size 2048, 85% overlap, and a frame of 4096.
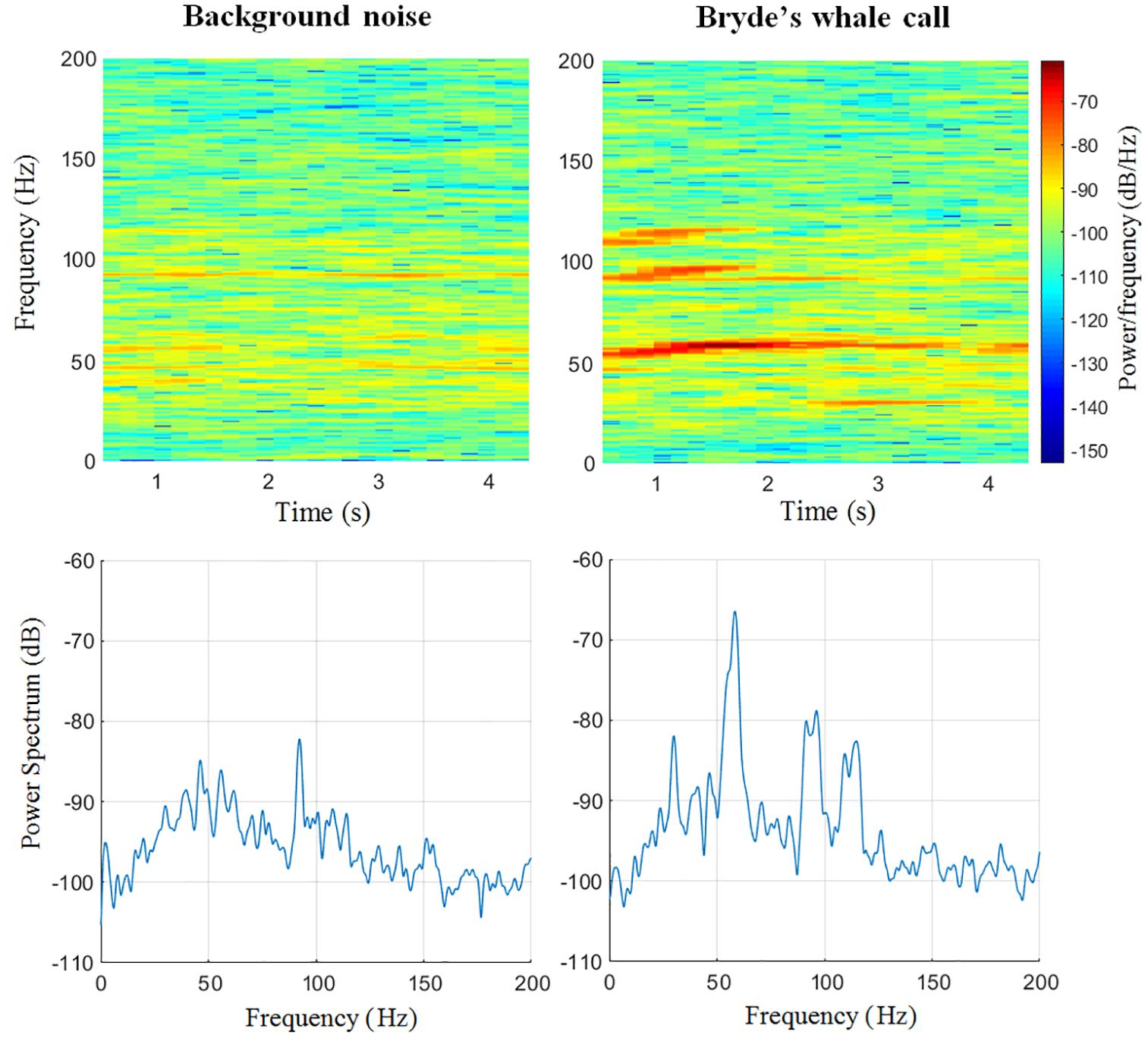
Figure 4. Example of a Bryde’s whale’s Be4 call (right) and its immediate background noise (left) at Punta Pescadero, Baja California Sur. Spectrograms 2D on the top and their respective power spectrum on the bottom. Spectrograms: Hamming window size 2048, 85% overlap, and a frame of 4096. Power spectra: Frequency resolution 1.9536 Hz.
Mann–Whitney-U tests were performed to compare call duration and frequency bandwidth distributions in both geographical areas. Non-parametric statistics were selected since they tend to be more conservative, less sensitive to unequal sample sizes, and do not assume normal data distribution (MacFarland and Yates, 2016).
Photo-Identification
Right-side photographs of dorsal fins taken from 2009 to 2014 in the northern and southern Gulf of California were compared to identify whether or not the same individuals can be found in both regions of the gulf. A recapture is defined as a match between photographs of the same individual taken at different times and different and same sites using natural marks as identification. All photographs were taken with reflex cameras (Nikon D300, 7000, Minato City, Tokyo, Japan) with 70–300 mm lenses, at a distance from 20 to 30 m, sailing at 5–7 knots and parallel to the individual whale. Each photograph was georeferenced using GPS Garmin® 710 (Garmin Ltd., Switzerland) for location.
Results
Call Analysis
From HARP systems, a total of 902 Be4 calls were detected in 932 h revised: 407 from Punta Pescadero and 495 from Bahía de Los Ángeles. Only did 195 Be4 calls fulfill the SNR requirements, considered as good SNR, which were used for further analyses – 103 Be4 calls from Punta Pescadero and 92 Be4 calls from Bahía de Los Ángeles (Table 1). Examples of Be4 calls in both locations can be seen in Figure 5.
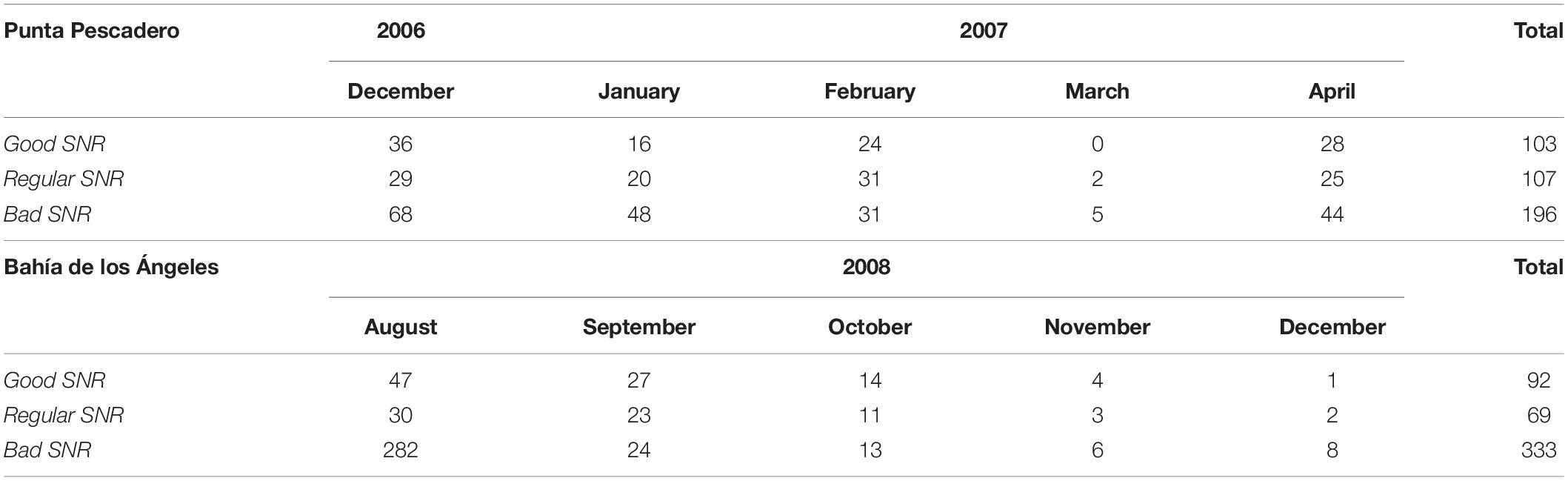
Table 1. Number Be4 calls of Bryde’s whale detected by signal-to-noise ratio (SNR) in recordings by the site.
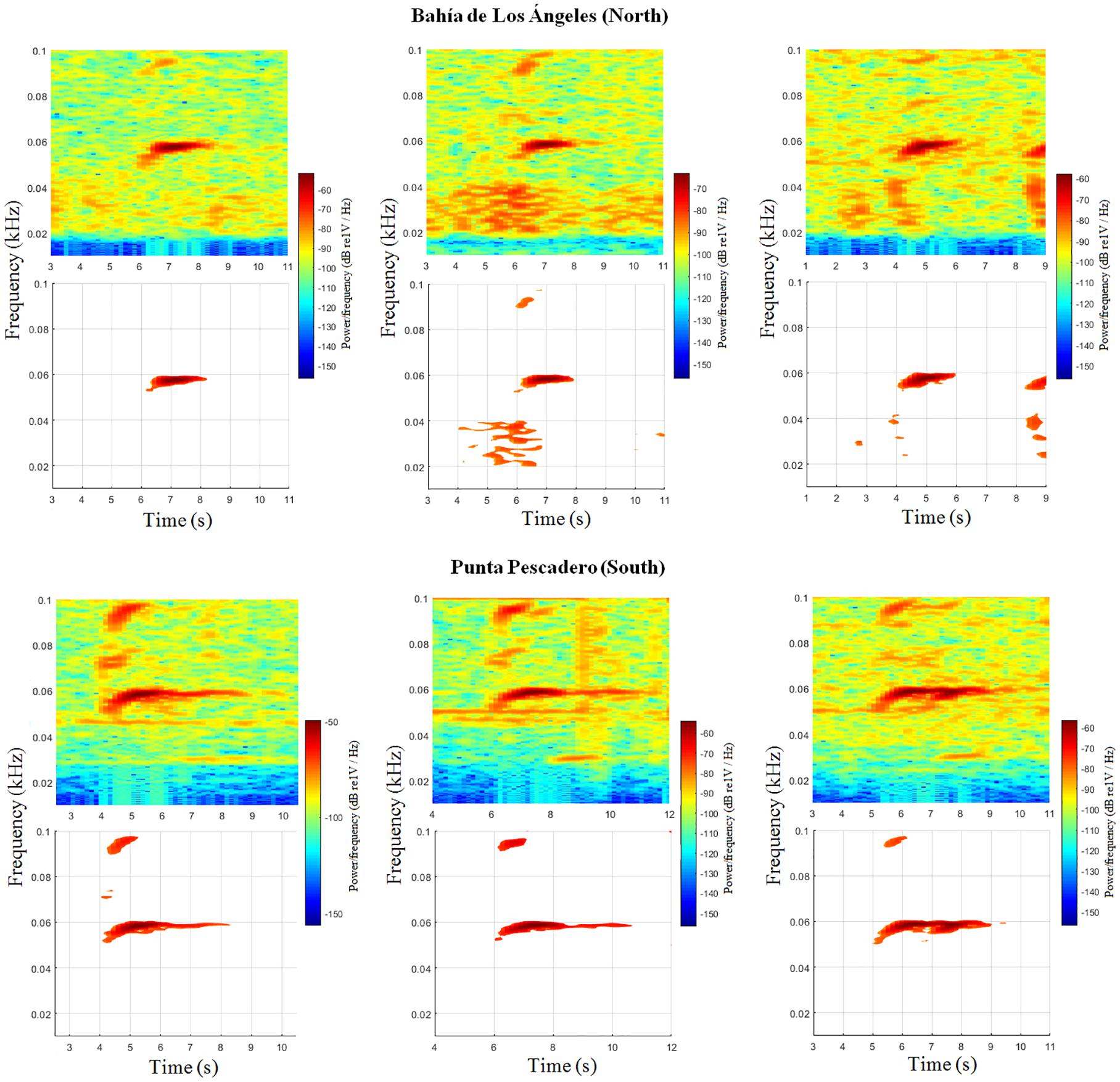
Figure 5. Examples of Be4 calls from the north (Bahía de Los Ángeles, Baja California) and from the south (Punta Pescadero, Baja California Sur). Spectrograms 2D showed for each call without (top) and with (bottom) the threshold cut at 24 dB from the maximum amplitude value. Spectrograms: Butterworth (2nd order) high-pass filter at 40 Hz cut off, Hamming window size 2048, 85% overlap, and a frame of 4096.
The frequency range did not show a significant difference between medians at both sites (Mann–Whitney test, U = 10002, z-statistics = −0.2334, p = 0.82). However, duration showed a significant difference (Mann–Whitney test, U = 11238, z-statistics = 2.91, p = 0.003) (Figure 6).
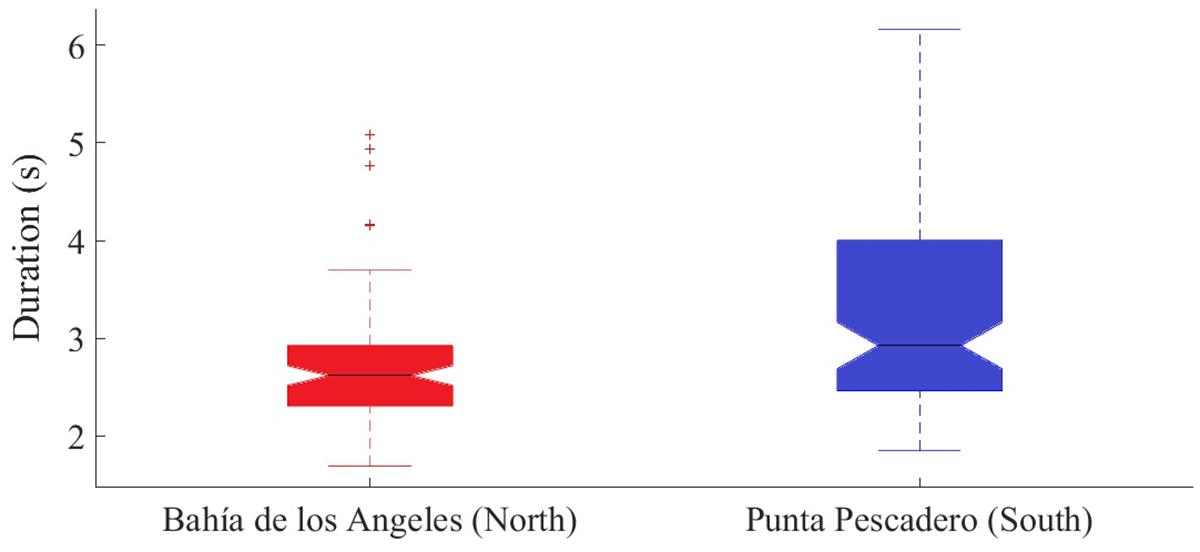
Figure 6. Boxplots showing the first quartile, median, third quartile of the call duration of Bryde’s whale data from the north (Bahía de Los Ángeles, Baja California) from November 2006 to June 2007 and the south (Punta Pescadero, Baja California Sur) from August to December 2008. The calls of both sites are significantly different (p = 0.003). Outliers are represented as (+).
To further investigate how the call duration was represented in both sites, values were plotted in histograms. Those graphs showed that data for Bahía de Los Ángeles centered mainly on one basic distribution. On the other hand, for Punta Pescadero, data seemed to be shown in two distributions. The first one was similar to that observed in Bahía de Los Ángeles, with almost half of the second distribution values.
A percentile analysis allowed a call length threshold of 3.5420 s delineating that 90% of the data from Bahía de Los Ángeles were below that value. A slightly less than half (37%) of the data for Punta Pescadero was above the same threshold (Figure 7).
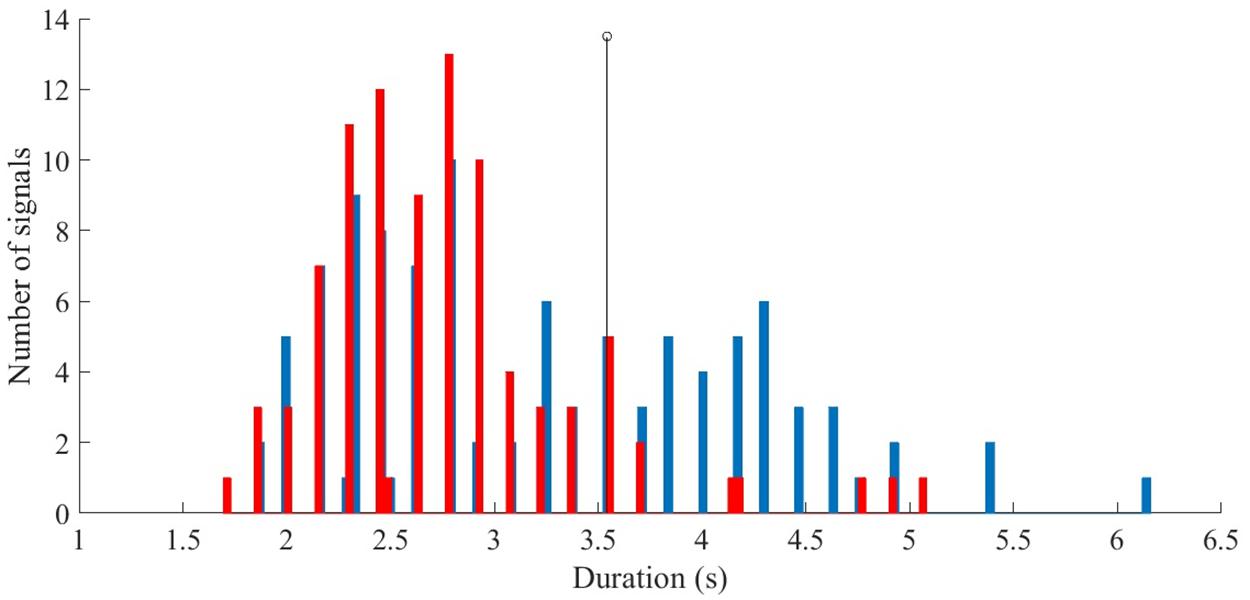
Figure 7. Distribution of duration calls of Bryde’s whale. Blue bars represent calls from the south (Punta Pescadero, Baja California Sur) from August to December 2008, red bars represent calls from the north (Bahía de Los Ángeles, Baja California) from November 2006 to June 2007. The black line represents a call duration threshold level for the population divide (3.5420 s). Distribution from the north overlaps with one of the distributions found in the south.
The difference between two-call duration distributions found in the south was significant (Mann–Whitney test, U = 2211, z-statistics = −8.39, p = 4.4776e-17). The north call duration distribution was not significantly different from the short duration call distribution from the south (Mann–Whitney test, U = 4839.5, z-statistics = −1.44, p = 0.15) (Figure 8).
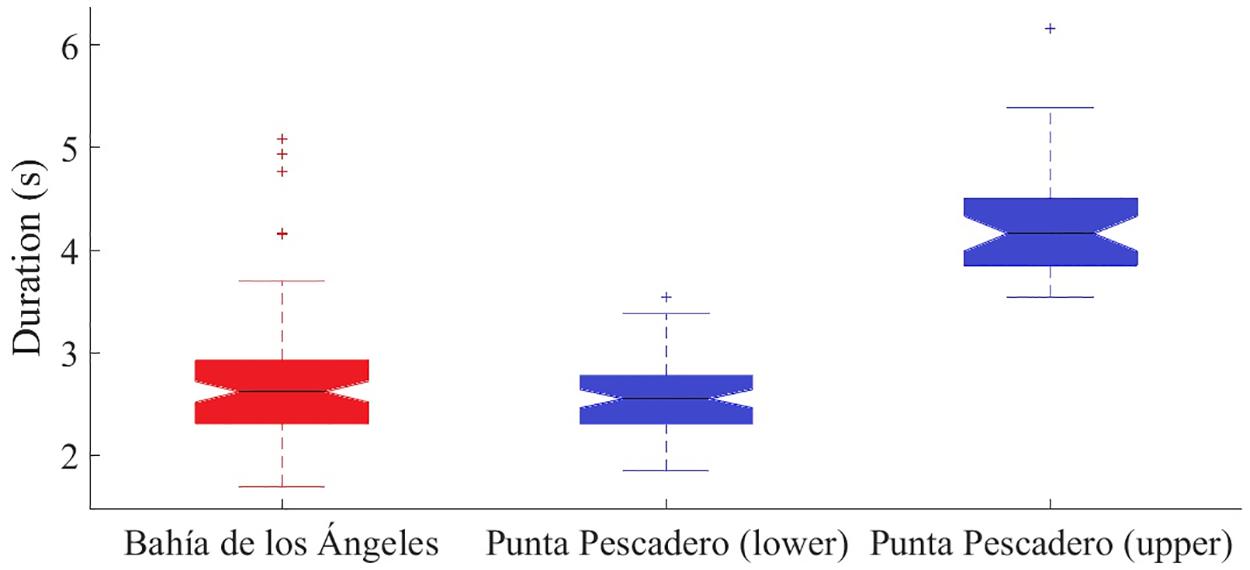
Figure 8. Boxplots showing the first quartile, median, third quartile of the call duration distribution of Bryde’s whale from the north (Bahía de Los Ángeles, Baja California) from November 2006 to June 2007, and the two suggested distributions in the south (Punta Pescadero, Baja California Sur) from August to December 2008. Both distributions in the south are significantly different between each other (p = < 0.00001). Outliers are represented as (+).
Photo-Identification
From the 118 photo-identified individuals, 14 were recaptured. Half of the recaptures were inter-annual, and half were intra-annual. From the inter-annual recaptures, five occurred in the same geographical region (north-north or south-south), and two occurred north-south (Table 2). From the intra-annual recaptures, all happened in the same geographic area (Figure 9).
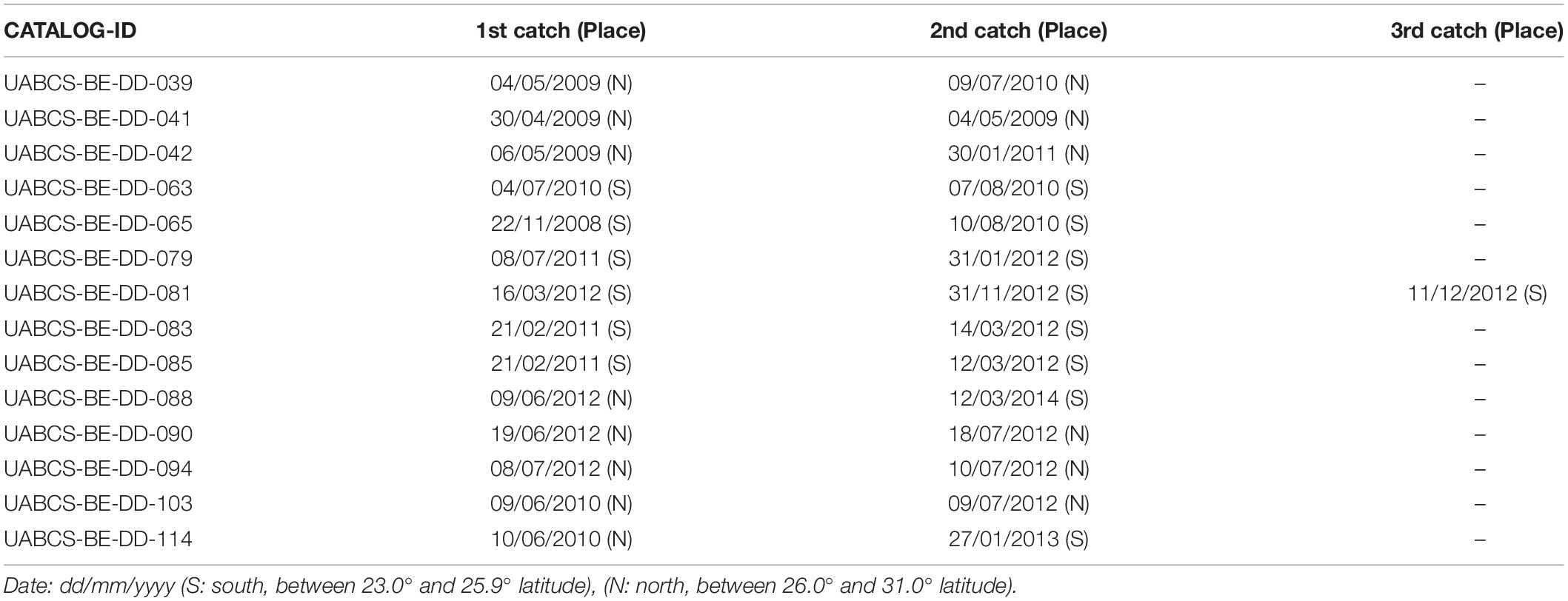
Table 2. Recaptures of photo-identified individuals of Bryde’s whale.

Figure 9. Photographs of Bryde’s whale individual with catalog ID 039, first (2009) and second (2010) catch in the Northern Gulf of California, Mexico.
Discussion
Call Measurements and Comparability
Comparing whale calls among different studies can be a useful exercise to increase understanding of geographical variations. However, the lack of an established method for acoustic parameter measurements or that of detailed information about measurements taken from spectrograms (spectrogram parameters, energy level above ambient noise, level cut to establish limits of call, etc.) can be problematic for future researchers interested in comparing data. It is even more important in species like Bryde’s whales whose reports on acoustic behavior remain scarce. In this study, the detailed procedure used to obtain the duration and frequency range values was described with the aim of facilitating comparison with future studies focusing on calls from this species.
The abundance of Be4 calls found in this study (99.5%) coincide with data reported by Kerosky et al. (2012) at California Bight, who also described Be4 call as the most abundant (97%). However, both contrast with the 5% reported by Oleson et al. (2003) from the equator to parallel 23°N. These differences could be explained by the latitudinal separation of the call types described by Oleson et al. (2003), who reported Be4 calls exclusively for the southern coasts of Baja California Peninsula. Thus, the results in this study combined with what was reported by Oleson et al. (2003) and Kerosky et al. (2012) suggest that Be4 calls are only found from Southern California to both coasts of Baja California Peninsula.
Geographic Differences in Bryde’s Whales Calls
Even though studies started in the 1970s, knowledge about Bryde’s whales in the Gulf of California remains insufficient to explain its dynamics. Past studies have suggested the presence of a resident population in the northern area, based on photo-identification analysis (Breese and Tershy, 1987) and migration of individuals from the Pacific to the southern area, based on seasonal migration (Vidal et al., 1987; Urbán and Flores-Ramírez, 1996; Salvadeo et al., 2011). This study represents the first approach on using call structure to identify population differences and relations of individuals within the Gulf of California. The results showed that from high signal to noise ratio Be4 calls detected in the northern Gulf of California (median duration 2.61 s) differed significantly in duration with those found in the southern area (median duration 2.93 s) (Figure 6).
Geographic differences in calls have been claimed in other species of the family Balenopteridae: Humpback whales (Payne, 1979; Cerchio et al., 2001), fin whales (Hatch and Clark, 2004; Širović et al., 2017), and minke whale (Risch et al., 2014). Vocalization variation is particular to each species. Atlantic minke whales show regional differences in pulse number (around 100 pulses of difference) and pulse train duration (20–30 s difference) (Risch et al., 2014). Similarly, Pacific minke whales also show regional differences in their calls known as “boings” by approximately 1 s difference in duration (Rankin and Barlow, 2005). Geographic differences in fin whale songs consist of inter-pulse duration, as well as number of pulses. These differences were found not only in large geographic scale (California Coast vs. Gulf of California) but also in smaller areas, such as within the coast of California or the Gulf of California (Širović et al., 2017).
Those cases mentioned above show the large diversity of intraspecific vocalization variation that can be found in whale species. Therefore, finding the biological meaning and importance of call geographical variation among populations can be a complex task. Interestingly, Catchpole and Slater (2008) claimed that to explain bird song geographical variation it was necessary to identify the historical processes that led to those variations and the reasons why such variations persisted. This paradigm can be applied to the geographic call variation found in baleen whales.
For Bryde’s whales, the meaning of the variation found is unknown. Similar situation occurs with blue whales, for which developing studies connecting at least four factors have been proposed: historical, ecological, morphological and behavioral to understand function and meaning of regional song variation (McDonald et al., 2006). For fin whales, such regional variation might refer to differential song functionality (Širović et al., 2017). For minke whales, long pulse trains have been associated to reproductive behavior and shorter pulse trains to feeding although authors insist that further research is necessary (Risch et al., 2014).
Knowing the biological function of whale calls can help to interpret the relevance of geographic variation. Nevertheless, associating correctly a function with a call can be a challenge. In cases where the emission of specific call type coincides with breeding season and males can be identified, it could be relatively easy to associate those calls with reproductive behavior, as it happens with humpback whales (Payne and McVay, 1971). However, for species that lack clear specific breeding seasons or area, association call-behavior-function becomes more difficult. Considering that Bryde’s whale breeding season or area remain unknown and surface behavior tends to be limited, associating this function to calls is nearly impossible. Viloria-Gómora et al. (2015) claim that call Be4 was always detected when more than one individual was sighted and most of the time in prey (e.g., sardine) presence (Tershy et al., 1993). According with this idea, Kato and Perrin (2009) described that Bryde’s whale individuals only form groups during feeding, so this grouping represents opportunities for socialization. Moreover, one of the main social events for many species is reproduction (Tyack, 2009), thus, call Be4 possibly is associated with this behavior as it has been described for fin whales (Croll et al., 2002). Nevertheless, to confirm this hypothesis further research with specific methodology is required.
Bryde’s Whale Population Dynamics in the Gulf of California
The results of this study support the idea that in the southern Gulf of California two whale population units of Bryde’s whales are dwelling and mixing (Figure 7). The southern area is considered populated by transient individuals from the Pacific Ocean but could also contain a local population that moves in a wider area within the Gulf. The similarity in genetics (Viloria-Gómora, 2015) and an overlap in the most common contact call duration between north and south could indicate a dynamic population within the Gulf of California. This finding might mean that a population has a strong philopatry in the north, as it has been suggested in the past (Tershy et al., 1990, 1993) but with internal movements to the southern part. The hypothesis of individuals in the south structured in two populations is proposed by the distinct distributions of call duration (Figure 8). This claim is supported by the continuous presence of the species in the southern part of the Gulf, suggestion of existence of a transient and resident populations (Urbán and Flores-Ramírez, 1996), and influence of individuals migrating from the Eastern Tropical Pacific (Salvadeo et al., 2011).
The photo-identification results support the data found in the acoustic analysis. For instance, regional fidelity (north or south) can be noticed for some individuals (n = 7), likely associated with the suggested presence of resident populations (Tershy et al., 1990, 1993; Urbán and Flores-Ramírez, 1996). Two individuals were recaptured in both regions, suggesting some interaction between those stocks (Urbán and Flores-Ramírez, 1996). These movements can be the result of prey availability (Salvadeo et al., 2011), since socialization and feeding behavior seems to be linked in this species. However, due to the number of identified animals (n = 108), these results are still considered preliminary, proposing the need of increasing research effort on this particular species. Further research can be done using a combined effort of photo-identification, telemetry, tagging and passive acoustics to detail the movements of individuals within and outside the Gulf of California. Photo-identification and telemetry have been successfully used for other species in the area (Jiménez-López et al., 2019).
The comparison of Be4 call duration reported by Oleson et al. (2003) and by Kerosky et al. (2012) with the one reported in this study was only possible to a subjective level. The lack of information associated with the measurement procedures and descriptive values for the calls prevented from conducting a numerical comparison. Nonetheless, general visual comparison allowed to notice that Be4 calls registered in the southern California Bight are similar to those recorded in the Gulf of California (S. Kerosky, com. Pers.). A standardized numerical comparison of Be4 call duration among the three studies and explanation of regional differences remain on hold for future research.
This study contributes to a new insight about acoustic variation among populations of this species in the North Pacific, considering that some degree of geographic variation in the Eastern Tropical Pacific has been already suggested (Oleson et al., 2003; Heimlich et al., 2005). It also opens an opportunity for investigating population dynamics of the species more extensively using acoustic analysis as it has been used for other species, such as fin whales (Hatch and Clark, 2004).
Conclusion
Bryde’s whales are an elusive and poorly understood cetacean species. Even though their distribution is worldwide, the cryptic nature of the species greatly limits visual research opportunities. The Gulf of California is one of the few areas where a population has been partially monitored throughout several years. The presence of a resident population in the northern Gulf of California has been suggested in past works, and this study adds evidence with acoustic data to the hypothesis of the existence of at least two population stocks in the area.
The northern population seems to remain resident in the north with some movement to the south, proven by photo-identification and call detections. The stock in the south is likely composed by resident individuals plus migrants entering from the Pacific Ocean.
Passive acoustic methods have shown an excellent complement to whale population studies in addition to photo-identification and tissue samples. However, the importance of standardizing call measurements cannot be ignored to compare repertoires efficiently and accurately. Long-term monitoring can help to understand the movements of Bryde’s whales and increase knowledge about their acoustic repertoire.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation. Requests to access the datasets should be directed to LV-G, bHZpbG9yaWFAdWFiY3MubXg=.
Ethics Statement
Ethical review and approval was not required for the animal study because the study involves only non-invasive techniques: passive acoustics and pictures.
Author Contributions
LV-G and JUR conceived the study and acquired funding. LV-G and ER-V planned methodology. LV-G coordinated field work. LV-G and BL-L performed the data analysis and wrote original draft. ER-V and JUR revised and edited the draft. All authors contributed to the article and approved the submitted version.
Funding
This work was financially supported by the Consejo Nacional de Ciencia y Tecnología (National Council of Science and Technology) (SEP-CONACyT #134251), the WWF-TELCEL Alliance, Ecology Project International. LV-G would like to thank CONACYT Mexico for her scholarship (number: 362054).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank all the staff of the Marine Mammal Research Program for their support during field trips, in particular Sergio Martinez, field supervisor; to Universidad Autónoma de Baja California Sur for support in administering the draft; to John Hildebrand from Scripps Institution of Oceanography for his collaboration in acquiring stationary recordings; to Diana Fischer for editorial services in English, and to the editor and reviewers for their comments and suggestions.
References
Au, W. W., and Hastings, M. C. (2008). Principles of Marine Bioacoustics, Vol. 510. New York: Springer.
Baird, R. W. (2000). Status of killer whales, Orcinus orca, in Canada. Can. Field Nat. 115, 676–701.
Breese, D., and Tershy, B. (1987). “Residency patterns in female Bryde’s (Balaenoptera edeni) whales in the Gulf of California, Mexico,” in Proceedings of the Seventh Biennial Conference on the Biology of Marine Mammals, 5–7 December, Miami, FL.
Catchpole, C. K., and Slater, P. J. (2008). Bird Song: Biological Themes and Variations. London: Cambridge University Press.
Cerchio, S., Jacobsen, J. K., and Norris, T. N. (2001). Temporal and geographical variation in songs of humpback whales, Megaptera novaeangliae: synchronous change in Hawaiian and Mexican breeding assemblages. Anim. Behav. 62, 313–329. doi: 10.1006/anbe.2001.1747
Chiu, P. H. (2009). Studies on the Population and Acoustic Signals of Bryde’s Whales (Balaenoptera edeni) in Tosa Bay, Japan. Kaohsiung: National Sun Yat-sen University.
Committee on Taxonomy (2020). List of Marine Mammal Species and Subspecies. Lawrence: Society for Marine Mammalogy.
Constantine, R., Iwata, T., Nieukirk, S. L., and Penry, G. S. (2018). Future directions in research on Bryde’s whales. Front. Mar. Sci. 5:333. doi: 10.3389/fmars.2018.00333
Croll, D. A., Clark, C. W., Acevedo, A., Tershy, B., Flores, S., Gedamke, J., et al. (2002). Only male fin whales sing loud songs. Nature 417, 809–809. doi: 10.1038/417809a
Cummings, W. C. (1985). “Bryde’s whale balaenoptera edeni Anderson, 1878,” in Handbook of Marine Mammals, Vol. 3, eds S, H. y. Ridgway and R. Harrison (London: Academia Press), 137–154.
Cummings, W. C., Thompson, P. O., and Ha, S. J. (1986). Sounds from Bryde, Balaenoptera edeni, and finback, Balaenoptera physalus, whales in the Gulf of California. Fish. Bull. 84, 359–370.
Delarue, J., Todd, S. K., Van Parijs, S. M., and Di Iorio, L. (2009). Geographic variation in Northwest Atlantic fin whale (Balaenoptera physalus) song: implications for stock structure assessment. J. Acous. Soc. Am. 125, 1774–1782. doi: 10.1121/1.3068454
Dizon, A., Lux, C. A., LeDuc, R. G., Urban, J., Henshaw, M., Baker, C. S., et al. (1995). An Interim Phylogenetic Analysis of Sei and Bryde’s Whales; Whale Mitochondrial DNA Control Region Sequences. SC/47/NP23 IWC Scientific Committee. Schaffhausen: IWC.
Edds, P. L., Odell, D. K., and Tershy, B. R. (1993). Vocalizations of a captive juvenile and free-ranging adult-calf pairs of Bryde’s whales, Balaenoptera edeni. Mar. Mamm. Sci. 9, 269–284. doi: 10.1111/j.1748-7692.1993.tb00455.x
Erbe, C., Dunlop, R., Jenner, K. C. S., Jenner, M. N., McCauley, R. D., Parnum, I., et al. (2017). Review of underwater and in-air sounds emitted by Australian and Antarctic marine mammals. Acoust. Aust. 45, 179–241. doi: 10.1007/s40857-017-0101-z
Figueiredo, L. D., and Simão, S. M. (2014). Bryde’s whale (Balaenoptera edeni) vocalizations from Southeast Brazil. Aquat. Mamm. 40, 225–231. doi: 10.1578/am.40.3.2014.225
Figueiredo, L. D., Tardin, R. H., Lodi, L., Maciel, I. S., Alves, M. A. S., and Simão, S. M. (2014). Site fidelity of Bryde’s whales (Balaenoptera edeni) in Cabo Frio region, southeastern Brazil, through photoidentification technique. Braz. J. Aquat. Sci. Tech. 18, 59–64. doi: 10.14210/bjast.v18n2.p59-64
Hatch, L. T., and Clark, C. W. (2004). Acoustic Differentiation between Fin Whales in Both the North Atlantic and North Pacific Oceans, and Integration with Genetic Estimates of Divergence. SC/56/SD6 IWC Scientific Committee. Schaffhausen: IWC.
Heimlich, S. L., Mellinger, D. K., Nieukirk, S. L., and Fox, C. G. (2005). Types, distribution, and seasonal occurrence of sounds attributed to Bryde’s whales (Balaenoptera edeni) recorded in the eastern tropical Pacific, 1999–2001. J. Acoust. Soc. Am. 118, 1830–1837. doi: 10.1121/1.1992674
International Whale Commission (2020). Bryde’s Whale-Balaenoptera Edeni. Available online at: https://iwc.int/brydes-whale (accessed June 24, 2020).
Jefferson, T. A., Weber, M. A., and Pitman, R. L. (2008). Marine Mammals of the World, A Comprehensive Guide to Their Identification. San Diego, CA: Academic Press.
Jiménez-López, M. E., Palacios, D. M., Jaramillo-Legorreta, A., Urbán, R. J., and Mate, R. B. (2019). Fin whale movements in the Gulf of California, Mexico, from satellite telemetry. PLoS One 14:e0209324. doi: 10.1371/journal.pone.0209324
Kato, H., and Perrin, W. F. (2009). Bryde’s whales: Balaenoptera edeni/brydei. In: Encyclopedia of Marine Mammals. London: Academic Press, 158–163.
Kerosky, M. S., Široviæ, A., Roche, L. K., Baumann-Pickering, B., Wiggins, S. M., and Hildebrand, A. J. (2012). Bryde’s whale seasonal range expansion and increasing presence in the Southern California Bight from 2000 to 2010. Deep Sea Res. 1, 125–132. doi: 10.1016/j.dsr.2012.03.013
Leatherwood, S., Reeves, R. R. W., Perrin, F., and Evans, W. E. (1988). Whales, Dolphins, and Porpoises of the Eastern North Pacific and Adjacent Arctic Waters, A Guide to Their Identification. NOAA Technical Report NMFS Circular 444. Washington, DC: NOAA.
MacFarland, T. W., and Yates, J. M. (2016). “Mann–Whitney U Test”, in: Introduction to Nonparametric Statistics for the Biological Sciences Using R. Cham: Springer.
McDonald, M., Mesnick, S. L., and Hildebrand, J. (2006). Biogeographic characterization of blue whale song worldwide: using song to identify populations. J. Cetacean Res. Manag. 8, 55–65.
Mellinger, D., and Barlow, J. (2003). Future Directions for Acoustic Marine Mammal Surveys: Stock Assessment and Habitat Use. NOAA OAR Special Report, NOAA/PMEL Contribution 2557. Washington, DC: NOAA.
Miller, B. S., Collins, K., Barlow, J., Calderan, S., Leaper, R., McDonald, M., et al. (2014). Blue whale vocalizations recorded around New Zealand: 1964–2013. J. Acoust. Soc. of Am. 135, 1616–1623. doi: 10.1121/1.4863647
Niño-Torres, C., Urban, R. J., Olavarrieta, T., Blanco-Parra, M., and Hobson, K. A. (2013). Dietary preferences of Bryde’s whales (Balaenoptera edeni) from the Gulf of California: A δ13C, δ15N analysis. Mar. Mam. Sci. 30, 1140–1148.
Ogundile, O. O., Usman, A. M., Babalola, O. P., and Versfeld, D. J. (2020). A hidden Markov model with selective time domain feature extraction to detect inshore Bryde’s whale short pulse calls. Ecol. Inform. 57:101087. doi: 10.1016/j.ecoinf.2020.101087
Ogundile, O. O., and Versfeld, D. J. (2020). Analysis of template-based detection algorithms for inshore Bryde’s whale short pulse calls. IEEE Access. 8, 14377–14385. doi: 10.1109/access.2020.2966254
Oleson, E. M., Barlow, J., Gordon, J., Rankin, S., and Hildebrand, J. A. (2003). Low frequency calls of Bryde’s whales. Mar. Mamm. Sci. 19, 407–419. doi: 10.1111/j.1748-7692.2003.tb01119.x
Payne, R. S. (1979). “Humpback whale songs as an indicator of “stocks”,” in Proceedings of the Third Biennial Conference on the Biology of Marine Mammals, Seattle, WA.
Payne, R. S., and McVay, S. (1971). Songs of humpback whales. Science 173, 585–597. doi: 10.1126/science.173.3997.585
Putland, R. L., Merchant, N. D., Farcas, A., and Radford, C. A. (2018a). Vessel noise cuts down communication space for vocalizing fish and marine mammals. Glob. Chang. Biol. 24, 1708–1721. doi: 10.1111/gcb.13996
Putland, R. L., Ranjard, L., Constantine, R., and Radford, C. A. (2018b). A hidden Markov model approach to indicate Bryde’s whale acoustics. Ecol. Indic. 84, 479–487. doi: 10.1016/j.ecolind.2017.09.025
Rankin, S., and Barlow, J. (2005). Source of the North Pacific “boing” sound attributed to minke whales. J. Acoust. Soc. Am. 118, 3346–3351. doi: 10.1121/1.2046747
Reeves, R. R., Steward, B., Clapham, P. J. y., and Powell, J. A. (2002). Guide to marine mammals or the world. Natl. Audub. Soc. U.S.A. 527.
Rendell, L., Mesnick, S. L., Dalebout, M. L., Burtenshaw, J., and Whitehead, H. (2011). Can genetic differences explain vocal dialect variation in sperm Whales, Physeter macrocephalus? Behav. Genet. 42, 332–343. doi: 10.1007/s10519-011-9513-y
Rendell, L. E., and Whitehead, H. (2003). Vocal clans in sperm whales (Physeter macrocephalus). Proc. R. Soc. B. 270, 225–231. doi: 10.1098/rspb.2002.2239
Rice, A. N., Palmer, K. J., Tielens, J. T., Muirhead, C. A., and Clark, C. W. (2014). Potential Bryde’s whale (Balaenoptera edeni) calls recorded in the northern Gulf of Mexico. J. Acoust. Soc. Am. 135, 3066–3076. doi: 10.1121/1.4870057
Risch, D., Castellote, M., Clark, C. W., Davis, G. E., Dugan, P. J., Hodge, L. E. W., et al. (2014). Seasonal migrations of North Atlantic minke whales: novel insights from large-scale passive acoustic monitoring networks. Mov. Ecol. 2:24. doi: 10.1186/s40462-014-0024-3
Romero-Vivas, E., Viloria-Gómora, L., and Urbán, J. (2013). “Morphological and gradient power spectrum representations for Bryde’s Whale (Balaenoptera edeni) geographical call differentiation,” in Proceedings of the Conference: 1er Coloquio Nacional de Acústica, ESIME-IMA, New Delhi.
Salvadeo, C., Flores-Ramírez, S., Gómez-Gallardo, A., MacLeod, C., Llunch-Belda, D., Jaume-Schinkel, S., et al. (2011). Bryde’s whale (Balaenoptera edeni) in the southwestern Gulf of California Relationship with ENSO variability and prey availability. Cienc. Mar. 37, 215–225. doi: 10.7773/cm.v37i2.1840
Širović, A., Bassett, H. R., Johnson, S. C., Wiggins, S. M., and Hildebrand, J. A. (2014). Bryde’s whale calls recorded in the Gulf of Mexico. Mar. Mamm. Sci. 30, 399–409. doi: 10.1111/mms.12036
Širović, A., Oleson, E. M., Buccowich, J., Rice, A., and Bayless, A. R. (2017). Fin whale song variability in southern California and the Gulf of California. Sci. Rep. 7, 1–11.
Tershy, B. R., Acevedo-Gutiérrez, A., Breese, D., and Strong, C. (1993). Diet and feeding behavior of fin and Bryde’s whales in the central Gulf of California, Mexico. Rev. Investig. Científica 1, 31–37.
Tershy, B. R., Breese, D., and Strong, C. S. (1990). Abundance, seasonal distribution and population composition of balaenopterid whales in the Canal de Ballenas, Gulf of California, Mexico. Report Int. Whaling Commiss. 12, 369–375.
Tyack, P. (2009). “Behavior, overview. 101–108,” in Encyclopedia Marine Mammals, 2a Edn, eds F. P. William, B. y. Würsig, and J. G. M. Thewissen (Amsterdam: Elservier), 157–162. doi: 10.1080/010503998420720
Urbán, J., and Flores-Ramírez, S. (1996). A note on Bryde’s whales (Balaenoptera edeni) in the Gulf of California, Mexico. Rep. Int. Whaling Commiss. 46, 453–458.
Vera-Cuevas, V. C. (2016). Análisis de Variaciones Geográficas de Vocalizaciones del Rorcual Tropical (Balaenoptera edeni) en el Golfo de California. dissertation/master’s thesis. La Paz: Centro de Investigaciones Biologicas del Noroeste.
Vidal, O., Aguayo, A., Findley, L., Robles, A., Bourillón, L., Vomend, I., et al. (1987). “Avistamientos de mamíferos marinos durante el crucero “Guaymas I” en la región superior del Golfo de California, primavera de 1984,” in Proceedings of the Memorias de la X Reunión Internacional sobre Mamíferos Marinos, La Paz.
Viloria-Gómora, L. (2015). Estructura Poblacional del Rorcual tropical (Balaenoptera edeni brydei), en el Golfo de California Con Base en Técnicas Acústicas y Moleculares. dissertation/PhD thesis. La Paz: Universidad Autonoma de Baja Califoria Sur.
Viloria-Gómora, L., Romero-Vivas, E., and Urbán, J. R. (2015). Calls of Bryde’s whale (Balaenoptera edeni) recorded in the Gulf of California. J. Acoust. Soc. Am. 138, 2722–2725. doi: 10.1121/1.4932032
Wiggins, S. (2003). Autonomous acoustic recording packages (ARPs) for long-term monitoring of whale sounds. Mar. Technol. Soc. J. 37, 13–22. doi: 10.4031/002533203787537375
Wiggins, S. M., and Hildebrand, J. A. (2007). “High-frequency Acoustic Recording Package (HARP) for broad-band, long-term marine mammal monitoring,” in Proceedings of the 2007 Symposium on Underwater Technology and Workshop on Scientific Use of Submarine Cables and Related Technologies, (Piscataway, NJ: IEEE), 551–557.
Keywords: cetacean, Balaenoptera, vocalization, Baja California, Mexico
Citation: Viloria-Gómora L, Urbán R J, Leon-Lopez B and Romero-Vivas E (2021) Geographic Variation in Bryde’s Whale Be4 Calls in the Gulf of California: An Insight to Population Dynamics. Front. Mar. Sci. 8:651469. doi: 10.3389/fmars.2021.651469
Received: 09 January 2021; Accepted: 16 June 2021;
Published: 09 July 2021.
Edited by:
Rosalyn Putland, Centre for Environment, Fisheries and Aquaculture Science (CEFAS), United KingdomReviewed by:
Robert McCauley, Curtin University, AustraliaLouise Wilson, The University of Auckland, New Zealand
Copyright © 2021 Viloria-Gómora, Urbán R, Leon-Lopez and Romero-Vivas. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Eduardo Romero-Vivas, ZXZpdmFzQGNpYm5vci5teA==