
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
BRIEF RESEARCH REPORT article
Front. Mar. Sci., 28 April 2021
Sec. Marine Molecular Biology and Ecology
Volume 8 - 2021 | https://doi.org/10.3389/fmars.2021.650116
 Thomas Brauge1*†
Thomas Brauge1*† Sylvain Trigueros1†
Sylvain Trigueros1† Arnaud Briet1Sabine Debuiche1
Arnaud Briet1Sabine Debuiche1 Guylaine Leleu1Benoit Gassilloud2Amandine Wilhelm2Jean-Sébastien Py2Graziella Midelet1
Guylaine Leleu1Benoit Gassilloud2Amandine Wilhelm2Jean-Sébastien Py2Graziella Midelet1
We evaluated the performance of matrix-assisted laser desorption ionization-time of flight mass spectrometry (MALDI-TOF MS) associated with the Bruker BioTyperTM V7.0.0 database for the identification of 713 bacterial strains isolated from seafood products and sea water samples (ANSES B3PA collection) under culture conditions that may have been significantly different from those used to create the reference spectrum vs. the 16S rDNA sequencing. We identified 78.8% of seafood isolates with 46.7% at the species level (Bruker score above 2) and 21.2% (Bruker score between 1.7 and 2) at the genus level by the two identification methods, except for 3.8% of isolates with a difference of identification between the two methods (Bruker score between 1.7 and 2). There were 41.9% isolates (Bruker score below 1.7) with the identification at the genus level. We identified 94.4% of seafood isolates with 16S rDNA sequencing. The MALDI-TOF allowed a better strain identification to the species level contrary to the 16s rDNA sequencing, which allowed an identification mainly to the genus level. MALDI-TOF MS in association with the Bruker database and 16S rDNA sequencing are powerful tools to identify a wide variety of bacteria from seafood but require further identification by biochemical, molecular technique or other conventional tests.
Seafood is an important source of protein and part of human’s diets (FAO, 2018). However, seafood products may contain spoilage bacteria such as Shewanella spp. (Dehaut et al., 2014), foodborne pathogenic microorganisms such as Salmonella spp., Listeria monocytogenes (Midelet-Bourdin et al., 2007; Bolivar et al., 2016), and Vibrio spp. (Bonnin-Jusserand et al., 2019) that cause a health risk to consumers. Spoilage bacteria quickly deteriorate seafood and make them unfit for consumption. Seafood were also associated with many disease outbreaks in different countries such as the United States (Iwamoto et al., 2010) and Europe (EFSA, 2019). Initial sources of bacterial contamination of seafood are related to microbiological quality of the water from the nursery area to the fishing zone and the environmental conditions. In fact, significant water contamination sources are sewage and agricultural pollutions. Then, other contamination sources of seafood are associated with the preparation, process, handling, packaging, and storage conditions, and they also may affect the microbiological quality of the seafood. Consequently, it is crucial to maintain high quality and safety of seafood products for consumer health, and for this, it is important to have reliable and fast methods to identify and characterize seafood microflora (spoilage and pathogenic bacteria).
In recent years, time-of-flight mass spectrometry analysis by matrix-assisted laser desorption ionization (MALDI-TOF MS) has established itself as a powerful tool for bacteria identification (Wieser et al., 2012). This technique analyzes the proteome of treated cells and emits a protein spectrum specific to the analyzed bacteria. The unknown bacteria is identified by comparing its protein spectrum with databases containing reference spectra of each known bacteria, mainly reference clinical strains and collection strains (ATCC, CIP,…) that do not reflect the strains under stressful environmental conditions. However, many databases are incomplete or contain bacterial reference spectra obtained under specific and stringent culture conditions. Indeed, variability in culture conditions, sample preparation, or strain origin can be problematic because the mass spectrum is influenced by all these factors.
Several studies have investigated the use of this MALDI-TOF MS method to identify the microflora present in different food products such as in seafood (Carrera et al., 2013). There are few studies to identify and classify the seafood spoilage and pathogenic bacteria (Böhme et al., 2011, 2013; Popović et al., 2017), and in these studies, the strains investigated were mainly reference strains and less than 50 strains were isolated from seafood. Databases of MALDI-TOF are rich in patterns of clinical isolates, reference strains but less in patterns of seafood and environmental isolates.
In this study, we evaluated the performance of MALDI-TOF MS associated with the Bruker BioTyper database for the identification of 713 bacterial strains isolated from seafood products and sea water samples vs. the 16S rDNA sequencing. Identification results generated according to each methodology used had been compared, taking into account genera and species.
A total of 713 mesophilic bacterial strains (isolated at 30°C or 37°C depending on the strain) from our ANSES B3PA laboratory collection were studied, including 601 strains isolated from different seafood products (332 from shrimp, 180 from scad, 89 from whiting) and 112 strains from sea water samples. All strains were stored in brain–heart infusion (BHI) medium (Biomerieux, Marcy-l’Étoile, France) supplemented with glycerol (20% v/v) at −80°C in cryotubes. Before each experiment, each strain was grown on Mueller Hinton agar (Bio-Rad, Marne-La-Coquette, France) for 24 h, either at 30°C or 37°C depending on the strain.
For the 16S rDNA analysis, we amplified the total 16S rDNA gene (1,504 pb) by PCR technique. A single colony from Mueller Hinton agar had been collected with sterile loop and added in 200 μl of nuclease-free water (Qiagen, Hilden, Germany). The suspension was incubated for 8 min at 100°C. The suspension was then centrifuged for 5 min at 13,000 × g. PCR was performed in a total volume of 50 μl containing 2.5 μl extracted genomic DNA (supernatant), 0.25 μl of Hot Start Taq DNA Polymerase (Qiagen), 5 μl of PCR buffer at 10×, 1 μl of deoxyribonucleotide at 10 mM (Eurobio, Les Ulis, France), 0.25 μl of forward primer ENV1 at 50 μM (5′-AGA GTT TGA TII TGG CTC AG-3′) (Eurobio) (Olofsson et al., 2007), 0.25 μl of reverse primer ENV2 at 50 μM (5′-CGG ITA CCT TGT TAC GAC TT-3′) (Eurobio), and 40.75 μl of nuclease-free water (Qiagen). PCR was performed in an iCycler thermocycler (Bio-Rad, Marnes La Coquette, France) under the following conditions: 15 min at 95°C followed by 35 cycles of 45 s at 94°C and 45 s at 48°C and 60 s at 72°C. PCR products were sequenced by the Genoscreen private company (Lille, France) using the amplification primers. Alignments of nucleic acid sequences were performed by using Bioedit software. Then, a homology search against the NCBI 16S ribosomal RNA sequences database was performed with the obtained DNA sequences using BLAST algorithm. The identification was considered reliable if the identification rate was greater than or equal to 97% for the genus and 99% for the species (Drancourt et al., 2000). If several species belonging to the same genus match, then the identification was restricted to the genus. If several genera were identified for the same strain, the identification was not determined. Accession numbers of the nucleotide sequence data were referred to Supplementary Table 1.
All isolates were grown on Mueller Hinton agar according to the specific incubation procedures (30 or 37°C depending on the strain) and identified using a Microflex LT mass spectrometer (Bruker Daltonics, Germany), with MALDI BioTyper and FlexControl V3.0 software. Two different sample preparation procedures were used: (i) direct transfer (spotting) of the colony onto a target plate and (ii) formic acid overlay method that consists of depositing 1 μl of 70% formic acid on direct colony spotting. Three colonies per strain and per sample preparation were tested. For the direct spotting, a colony fraction was removed with 200-μl tips and homogeneously spread on a MALDI-TOF target well. Dried deposits were overlaid with 1 μl of 10 mg/ml of α-cyano-4-hydroxycinnamic acid (HCCA) matrix solution (Bruker Daltonics, Germany). The MALDI BioTyperTM system was calibrated with a Bruker Bacterial Test Standard (Escherichia coli DH5α), and the spectra for proteins with masses between 2,000 and 20,000 Da were obtained and matched with the Bruker V7.0.0 database, composed of 8,223 mass spectrometry profiles (MSPs). Analyses were done with a laser frequency of 60 Hz, an acceleration voltage of 20 kV, and extraction delay time of 120 ns. For the isolate identification, the confidence score thresholds were as follows: (1) with a score above 2, there was high confidence identification to genus and species levels; (2) with a score between 1.7 and 2, there was probable identification of genus level and no identification of species level; and (3) with a score below 1.7, there normally was not any reliability of identification (DeMarco and Burnham, 2014).
Seven hundred thirteen bacterial strains were analyzed by MALDI-TOF MS and blasted with the reference strains, mainly clinical isolates, available in the Bruker BioTyperTM V7.0.0 database. Among the 713 strains studied, 78.8% of isolates (562/713) were identified either at genus or species level (Tables 1, 2). For 46.8% of the strains (334/713), identifications were carried out with a Bruker score above 2, which means high confidence identification to the species level (Table 1). We identified 293 strains Gram + and 118 bacteria Gram-. However, for 10.8% of the isolates (77/713), we observed a high Bruker score between 2.091 and 2.403, but the identification was only to the genus level [Ochrobactrum sp. (one strain), Pseudomonas sp. (five strains), and Psychrobacter sp. (71 strains)]. No identification was determined for the strain of Ochrobactrum sp. and for three of five strains of Pseudomonas sp. by 16S rDNA sequencing. In contrast, the identifications were confirmed at the genus level for the two other strains of Pseudomonas sp. and 31 strains of Psychrobacter sp. and at the species level for the other 40 isolates of Psychrobacter celer by 16S rDNA sequencing. And for 21.2% of the isolates (151/713), the identification was probable of the genus level and no identification of the species level with a Bruker score between 1.7 and 2 (Table 2). We identified 95 isolates Gram+ and 56 isolates Gram–.
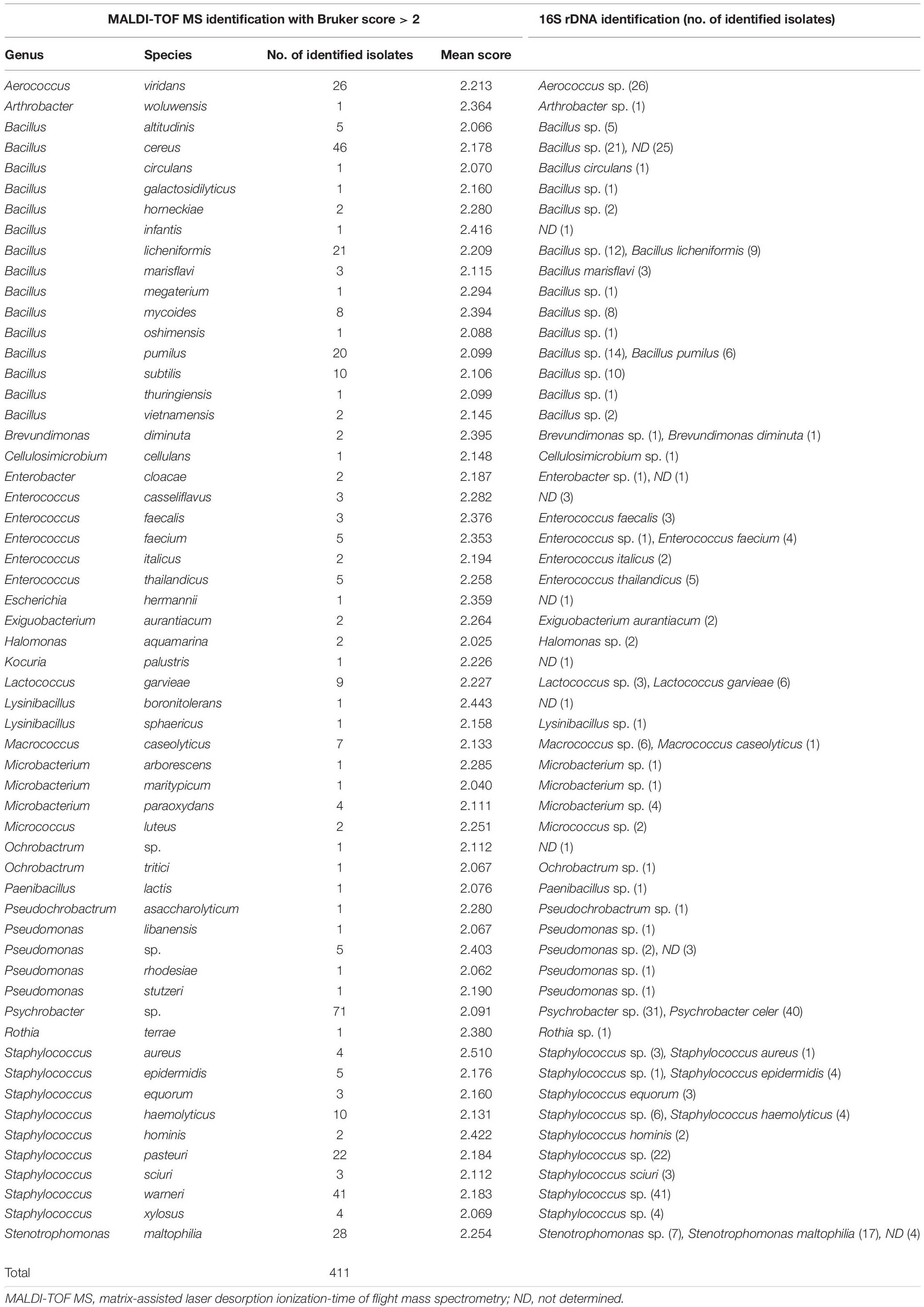
Table 1. Identification of isolates by 16S rDNA technique and by MALDI TOF method with Bruker score > 2.
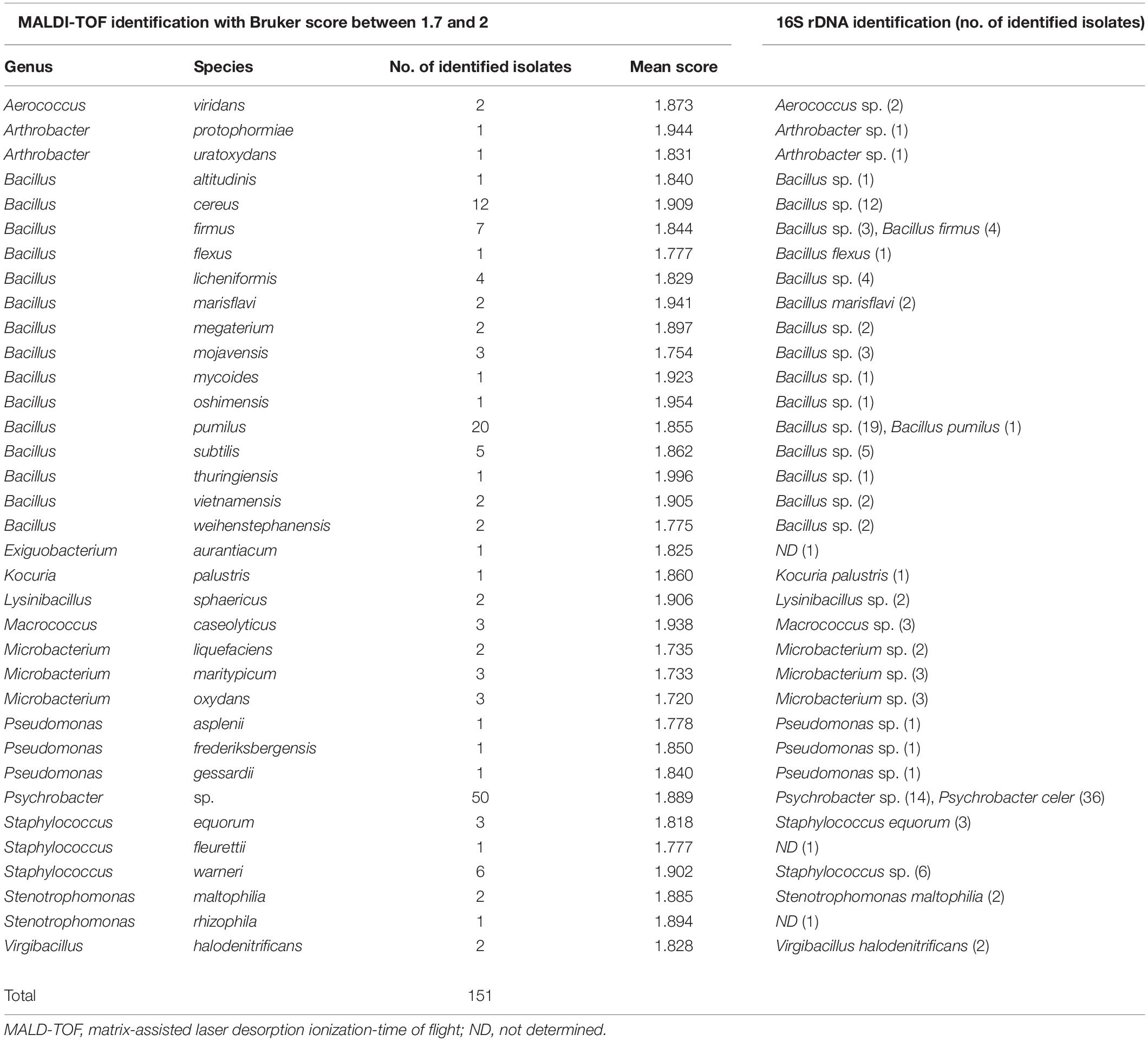
Table 2. Identification of isolates by 16S rDNA technique and by MALDI TOF method with Bruker between 1.7 and 2.
Concerning the 334 strains identified to the species level by MALDI-TOF MS (Bruker score above 2), the identification of 65.8% of the isolates (220/334) was confirmed at the genus level and 23.1% of the strains (77/334) at the species level by 16S rDNA sequencing. But for 11% of the 334 strains, no identification was determined by 16S rDNA sequencing. In the 24 bacterial genera identified, there were five major genera: Aerococcus, Bacillus, Enterococcus, Staphylococcus, and Stenotrophomonas, which matched 86.5% of the strains analyzed (289/334). For 13.5% of the isolates (45/334), 18 other genera were identified that were Arthrobacter, Brevundimonas, Cellulosimicrobium, Enterobacter, Escherichia, Exiguobacterium, Halomonas, Kocuria, Lactococcus, Lysinibacillus, Macrococcus, Microbacterium, Micrococcus, Ochrobactrum, Paenibacillus, Pseudochrobactrum, Pseudomonas, and Rothia. In fact, we showed a high probable species level for 29 strains of 11 different species (Bruker score between 2.353 and 2.510) and a probable species for 305 strains of 43 different species (Bruker score between 2.025 and 2.294). For 61.1% of the isolates (204/334), the main species were Aerococcus viridans, Bacillus cereus, Bacillus licheniformis, Bacillus pumilus, Staphylococcus warneri, Staphylococcus pasteuri, and Stenotrophomonas maltophilia, with 20–46 isolates per species. However, the identification of A. viridans, S. pasteuri, and S. warneri by MALDI-TOF MS was confirmed at the genus level (Aerococcus sp. and Staphylococcus sp.) by 16S rDNA sequencing. For the B. cereus species, 45.7% were confirmed at the genus level and 54.3% were not identified by 16S rDNA sequencing. For the two other species of Bacillus (B. licheniformis and B. pumilus), the identification at the species level was confirmed for 42.9 and 30% of the strains, respectively, and the other strains were confirmed at the genus level by 16S rDNA sequencing. For S. maltophilia species, the identification was confirmed at the species level for 60.7%, at the genus level for 25%, and no identification for 14.3% of the strains by 16S rDNA sequencing. For 151 strains with a Bruker score between 1.7 and 2, 63.6% (96/151) had been identified at the genus level, 34.4% (52/151) at the species level, and no identification for 2% of the strains (3/153) by 16S rDNA sequencing (Table 2). The two main bacterial genera were Psychrobacter sp. (50 strains) and Bacillus sp. (64 strains). For Psychrobacter sp., only the genus level had been determined by MALDI-TOF (Bruker score of 1.889) and was confirmed for 28% of the strains by 16S rDNA sequencing. Nevertheless, the other Psychrobacter sp. isolates (72%) have been identified at the Psychrobacter celer species. For the 12 other genera, 34 species were identified by MALDI-TOF spectroscopy. In these 34 identified species, three species (Exigobacterium aurantacum, Staphylococcus fleurettii, and Stenotrophomonas rhizophila) had no identification by 16S rDNA sequencing, and the identification of four species (Kocuria palustris, Staphylococcus equorum, S. maltophilia, and Virgibacillus halodenitrificans) was confirmed by both identification techniques used.
In the 713 strains analyzed by MALDI-TOF MS, 3.8% of isolates (27/713) had an identification at the species level with a Bruker score between 1.7 and 2, but these results had not been confirmed by 16S rDNA sequencing results (Table 3). In fact, the genus level was validated by the 16S rDNA reference strain sequences but not the species level. Based on species identification levels of the 16S rDNA sequencing results, four species had no reference spectrum in the Bruker BioTyperTM database. In contrast, the results of both identification techniques were identical at the Bacillus genus level for five strains but were different at the species level. In fact, the identified species were B. pumilus by 16S rDNA vs. Bacillus altitudinis by MALDI-TOF with a Bruker score of 1.888 (one strain), Bacillus nealsonii by 16S rDNA vs. Bacillus circulans by MALDI-TOF with a Bruker score of 1.713 (two strains), Bacillus Safensis by 16S rDNA vs. Bacillus pumilus by MALDI-TOF with a Bruker score of 1.725 (one strain) and Bacillus aquimaris by 16S rDNA vs. Bacillus vietnamensis by MALDI-TOF with a Bruker score of 1.865 (one strain). There had been a disagreement of species identification between the two techniques used, although the reference spectrum of these species B. pumilus, B. safensis, B. nealsonii, and B. aquimaris were in the Bruker BioTyperTM database. Finally, 17.4% (124/713 strains) obtained a Bruker score below 1.7 by MALDI-TOF MS, which normally does not allow any reliability of identification (Table 4). Two-sample preparation procedures have been applied to the 124 strains: direct transfer (spotting) and acid extraction. We observed that the best MALDI-TOF MS identification scores (the score nearest 1.7) were obtained with the direct transfer procedure and with the acid extraction procedure for 73 strains and 51 strains, respectively (data not shown). Among these 124 strains, the identification of 41.9% of isolates (52/124) was validated at the genus level by 16S rDNA sequencing (40 Gram+ and 12 Gram−). For three of these 52 strains, the identification had even been down to the species level by both identification techniques (Bacillus firmus, Bacillus vallismortis, and S. rhizophila). In contrast, for two genera (124 strains) and 10 species (50 strains) identified by 16S rDNA sequencing, the reference spectrum of these genera and species was not present in the Bruker database. These genera were Planococcus sp., and Salinicoccus sp., and the species were Bacillus oceanisediminis, Desemzia incerta, Exiguobacterium acetylicum, Exiguobacterium aestuarii, Exiguobacterium profundum, Microbacterium esteraromaticum, Psychrobacter alimentarius, Psychrobacter celer, Psychrobacter faecalis, and Shewanella indica. For 35 of 124 strains, the identification of the genus or species level by 16S rDNA sequencing was different from the results obtained by MALDI-TOF MS despite the presence of the reference spectrum for these genera and species in the Bruker database.
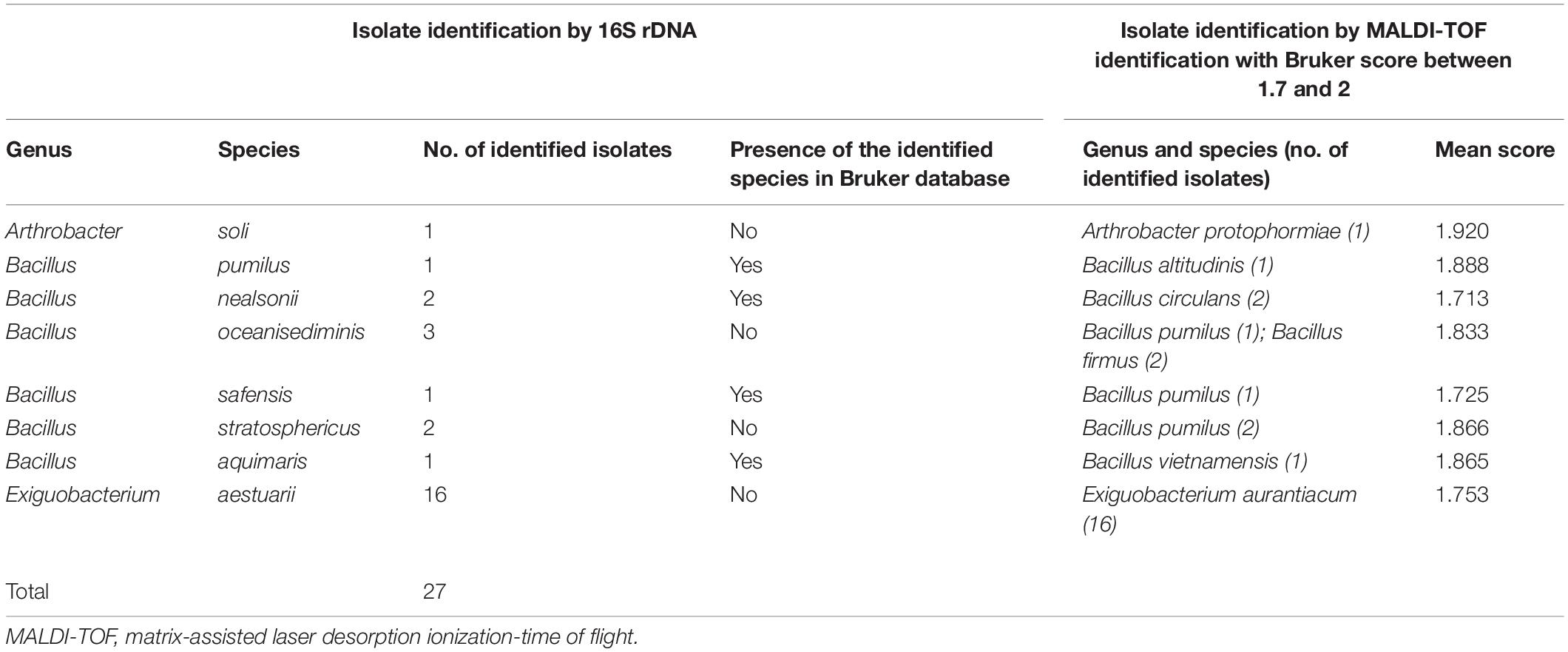
Table 3. Non-conformity of strain identification in 16S rDNA technique and MALDI-TOF method.
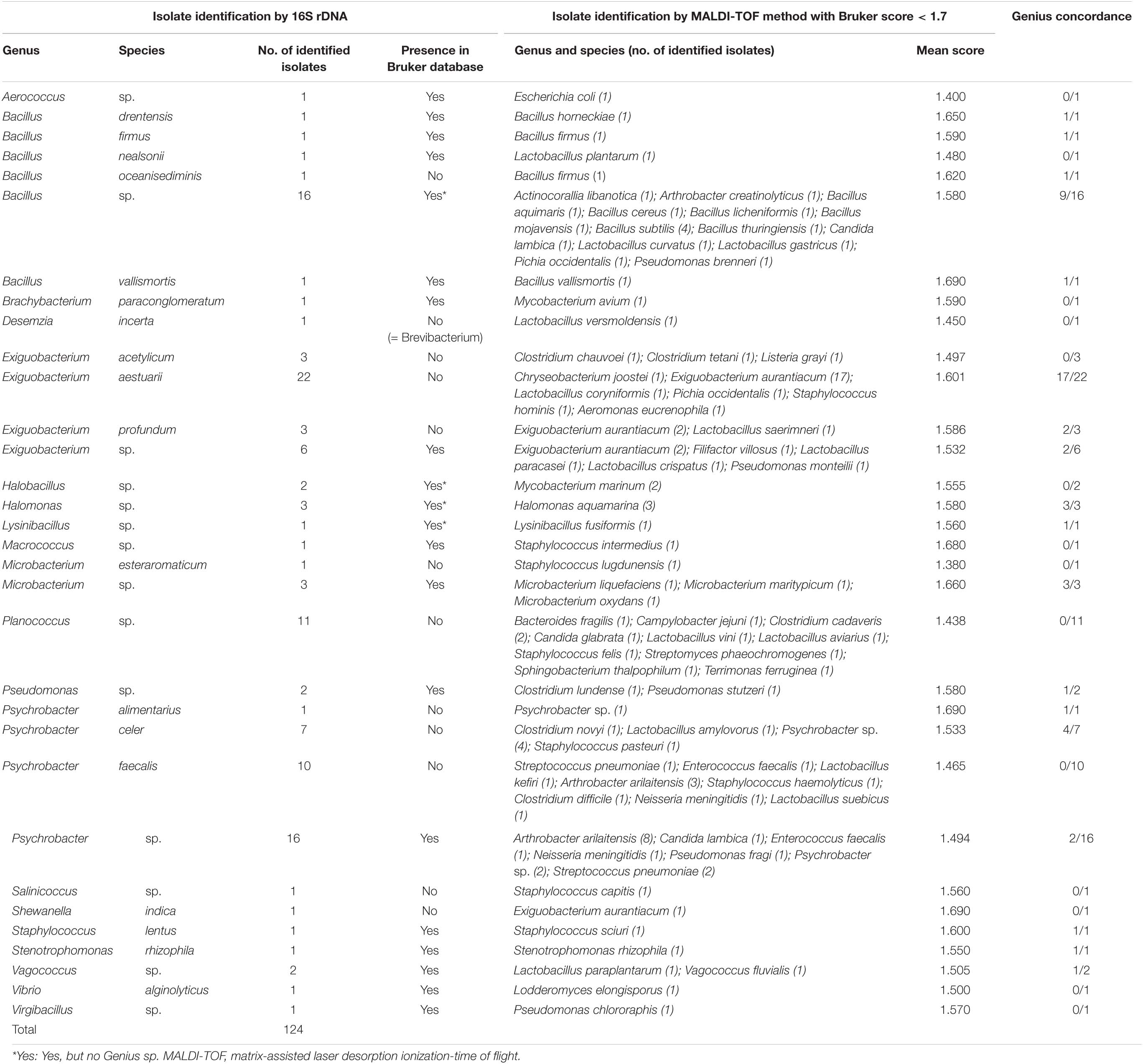
Table 4. Identification of isolates by MALDI-TOF method with Bruker score < 1.7 and 16S rDNA technique.
The aim of our study was to evaluate the ability of MALDI-TOF MS to identify 713 bacterial strains isolated from seafood products and sea water samples under culture conditions that may have been significantly different from those used to create the reference spectrum. It is important to know the microbial biodiversity present in seafood and fish because there could be pathogenic bacteria, which can infect humans or fishes and cause zoonotic or animal diseases. This is a capital step in order to prevent human foodborne illnesses or treat human or animals in aquaculture farms. Moreover, identification of pathogenic and spoilage bacteria in aquatic medium is the basis for further studies, such as epidemiological studies on antibiotic resistance, particularly in the antibiotic resistance genes spreading by horizontal gene transfer through bacterial flora. To our knowledge, this was the first study to identify a great number of seafood-isolated strains (713 isolates) with MALDI-TOF MS in association with the Bruker Biotyper V7.0.0 database vs. 16S rDNA sequencing. In fact, the studies of identification comparison between the conventional microbiological and/or molecular biological methods and the MALDI-TOF MS were mostly for clinically relevant bacteria and/or for the specific species or genera of bacteria (Bacillus, Pseudomonas, Vibrio…) (Popović et al., 2017). MALDI-TOF MS has been shown to be a competent tool for bacterial species differentiation due to the resulting highly specific spectrum, named fingerprints (Giebel et al., 2010). The Bruker BioTyperTM database was mainly composed of human pathogen spectra, and there are few other databases with specific spectra of food or environment isolates. For bacterial identification, the spectrum of a strain of interest is compared to a spectrum library. The problem is that these spectra are usually made under stringent conditions and on reference collection strains (human pathogen mainly causing infectious disease) that do not reflect the strains in their natural environment. The analysis of 16S rDNA sequences is a widely used method to identify bacterial species because it allows to compare 16S rDNA sequences to differentiate the bacteria at the genus/species level. However, the quality of the 16S DNA sequence databases used is an important factor. The sequences present in the databases serve as a reference for taxonomic assignment. The richness and precision of these databases are therefore a determining factor for good bacterial identification. Sometimes, there are several different 16S rDNA databases that have their advantages and disadvantages. In our study, we queried the NCBI 16S ribosomal RNA database as well as RDP database. We had compared the analysis results obtained with the RDP and the NCBI 16S rDNA databases. The RDP database gave equivalent or even less accurate identifications than the NCBI 16S rDNA database. By MALDI-TOF MS technique (Bruker score > 2 and between 1.7 and 2), we identified 58.2% isolates Gram + (415/713) to the species level, 24.4% isolates Gram− (174/713) to the genus level, and 6.6% isolates Gram– (47/713) to the species level. For the results of identification by 16S rDNA sequencing, we identified 39.6% isolates Gram+ (282/713) to the genus level and 10.2% (73/713) to the species level, 9.4% isolates Gram− (67/713) to the genus level, and 13.5% isolates Gram– (96/713) to the species level. No identification was observed for 6.2% of the strains. The identification at the species level was better for Gram + (58.2%) vs. Gram– (6.6%) isolates with MALDI-TOF technique, while it was lower by 16S rDNA sequencing (10.2% for Gram + and 13.5% for Gram−). The identification of more Gram + bacteria could be due to the bacterial biodiversity particularly of nature samples and/or the difficulty of growing viable culturable bacteria from natural habitats (Popovic, 2017). We showed that 57.8% of the strains from seafood were identified with a Bruker score higher than 2 and 21.2% were identified with a Bruker score between 1.7 and 2, which allowed identification to the species level as in the study of Böhme et al. (2013) that identified 76% to the species level (70 reference strains and 50 seafood isolates). We assigned six major genera, Aerococcus, Bacillus, Enterococcus, Psychrobacter, Staphylococcus, and Stenotrophomonas, and seven main species, A. viridans, B. cereus, B. licheniformis, B. pumilus, S. warneri, S. pasteuri, and S. maltophilia in our ANSES B3PA collection of seafood (shrimp, scad, whiting) and sea water isolates. A few studies identified seafood bacterial biodiversity. In fact, Böhme et al. (2010, 2011) have created a database with a collection of pathogenic and spoilage Gram+ and Gram− bacteria potentially present in seafood. They applied their method successfully to identify nine bacterial strains isolated from processed seafood, fish, and seafood (Bacillus, Carnobacterium, Pseudomonas, Serratia, Stenotrophomonas). As in our work, they identified Staphylococcus aureus, Bacillus subtilis, B. licheniformis, Bacillus megaterium, and Stenotrophomans maltophilia. For 141 of 159 isolates of the Psychrobacter genus present in our ANSES B3PA collection of seafood, no identification method was able to determine the species level. The species level was only indicated for 18 other Psychrobacter isolates by 16S rDNA method (Psychrobacter alimentarius, Psychrobacter celer, Psychrobacter faecalis), and Bruker scores were below 1.7 with several genera/species identifications. De Bruker BioTyperTM database was poor in spectra of this Psychrobacter genus vs. the 16S rDNA database. In recent years, several new Psychrobacter species were described based on their phenotypic and biochemical characteristics and from comparative 16S rDNA gene sequences within the genus Psychrobacter but not by MALDI-TOF MS.
However, in 15% (27/180 strains) of the cases, MALDI-TOF MS gives a misidentification of the species with Bruker scores close to 2. The MALDI-TOF MS identification spectrum therefore matched a reference spectrum of a close species for the majority of strains (23/27). The explanation was that reference spectra of bacterial species were not present in the Bruker BioTyperTM database, but the data in 16S rDNA sequence exist. For four others, the identification at the Bacillus genus level was identical but was divergent at the species level, while the spectra of these species were present in the MALDI-TOF BioTyperTM database. The similarity of the protein structures of the species made the discrepancy of differentiation of the quality peak difficult and made it difficult for MALDI-TOF MS to correctly identify the strains. This has been described for both anaerobic bacteria and other genera, such as Bacillus spp. (Takahashi et al., 2020), Streptococcus spp. (Fan et al., 2017), Mycobacterium spp. (Zingue et al., 2016), Enterococcus spp. (Lallemand et al., 2017), and yeast (Ling et al., 2014). Therefore, updating the existing information and perfecting the database of difficultly identified organisms are useful to improve the identification accuracy of MALDI-TOF MS (Li et al., 2019).
We also confirmed the unreliability of MALDI-TOF MS identifications at Bruker scores below 1.7. The primary recommended method for obtaining bacterial IDs from colonies is the cell smear (direct transfer) method. This smear method has shown its efficiency for the identification of numerous bacteria (Schröttner et al., 2014; Tsuchida et al., 2020); however, the probability of correctly identifying certain bacteria can still be quite limited (Tsuchida et al., 2020). If the mass spectrum obtained by the plate method is not sufficient, the acid extraction method can be applied to improve identification. In our study, we tried to improve the identification of 151 strains not identifiable by the MALDI-TOF with the direct transfer procedure. We showed that acid extraction would not improve the identification of these strains, which was surprising in view of the literature (Tsuchida et al., 2020). Among the 124 strains identified at Bruker scores below 1.7, only 41.9% were correctly identified at the genus level, and among these strains, only three strains were correctly identified at the species level by both techniques. Different observations were made previously by Schulthess et al. (2016), which showed that after ethanol-formic acid extraction and analysis of MALDI-TOF MS identifications with scores below the threshold of 1.7, 128 Gram− rods were identified at the genus level among 151 analyzed. Despite this low number of strains and low species diversity, studies by Böhme et al. (2010, 2011, 2013) showed the interest of building a reference spectrum database specific to each pathway/domain/industry.
In conclusion, we have shown that the MALDI-TOF MS in association with the Bruker BioTyperTM V7.0.0 database allowed a better strain identification to the species level contrary to the 16s rDNA sequencing, which allowed an identification mainly to the genus level. MALDI-TOF MS in association with the Bruker database and 16S rDNA sequencing are powerful tools to identify a wide variety of bacteria from seafood but require further identification by biochemical, molecular technique or other conventional tests. However, 16S DNA sequencing remains a powerful tool when combined with next-generation sequencing (NGS). Indeed, each strategy has its advantages. The MALDI-TOF allows good identification of the species but is limited to the identification of a strain previously isolated on culture medium, whereas the NGS based on 16S rDNA allows (despite a less precise identification) to free itself from the culture and to identify bacterial communities directly from a complex sample.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material. The nucleotide sequence data presented in the study are publicly available and are deposited in the Genbank database. The accession numbers can be found in the Supplementary Table S1.
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
This work was supported by a doctoral fellowship from Region Hauts-de-France/ANSES and by the French national effort to reduce antimicrobial resistance in veterinary medicine called “EcoAntibio2017” from the Ministry of Agriculture.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2021.650116/full#supplementary-material
Böhme, K., Fernández-No, I. C., Barros-Velázquez, J., Gallardo, J. M., Cañas, B., and Calo-Mata, P. (2010). Comparative analysis of protein extraction methods for the identification of seafood-borne pathogenic and spoilage bacteria by MALDI-TOF mass spectrometry. Anal. Methods 2, 1941–1947. doi: 10.1039/C0AY00457J
Böhme, K., Fernández-No, I. C., Barros-Velázquez, J., Gallardo, J. M., Cañas, B., and Calo-Mata, P. (2011). Rapid species identification of seafood spoilage and pathogenic Gram-positive bacteria by MALDI-TOF mass fingerprinting. Electrophoresis 32, 2951–2965. doi: 10.1002/elps.201100217
Böhme, K., Fernández-No, I. C., Pazos, M., Gallardo, J. M., Barros-Velázquez, J., Cañas, B., et al. (2013). Identification and classification of seafood-borne pathogenic and spoilage bacteria: 16S rRNA sequencing versus MALDI-TOF MS fingerprinting. Electrophoresis 34, 877–887. doi: 10.1002/elps.201200532
Bolivar, A., Costa, J. C. C. P., Posada-Izquierdo, G. D., Pérez-Rodríguez, F., Bascón, I., Zurera, G., et al. (2016). “Characterization of foodborne pathogens and spoilage bacteria in mediterranean fish species and seafood products,” in Foodborne Pathogens and Antibiotic Resistance, ed. O. V. Singh (John Wiley & Sons, Ltd), 21–39. doi: 10.1002/9781119139188.ch2
Bonnin-Jusserand, M., Copin, S., Bris, C. L., Brauge, T., Gay, M., Brisabois, A., et al. (2019). Vibrio species involved in seafood-borne outbreaks (Vibrio cholerae, V. parahaemolyticus and V. vulnificus): review of microbiological versus recent molecular detection methods in seafood products. Crit. Rev. Food Sci. Nutrit. 59, 597–610. doi: 10.1080/10408398.2017.1384715
Carrera, M., Cañas, B., and Gallardo, J. M. (2013). Proteomics for the assessment of quality and safety of fishery products. Food Res. Int. 54, 972–979. doi: 10.1016/j.foodres.2012.10.027
Dehaut, A., Midelet-Bourdin, G., Brisabois, A., and Duflos, G. (2014). Phenotypic and genotypic characterization of H2S-positive and H2S-negative strains of Shewanella baltica isolated from spoiled whiting (Merlangius merlangus). Lett. Appl. Microbiol. 59, 542–548. doi: 10.1111/lam.12312
DeMarco, M. L., and Burnham, C.-A. D. (2014). Diafiltration MALDI-TOF Mass spectrometry method for culture-independent detection and identification of pathogens directly from urine specimens. Am. J. Clin. Pathol. 141, 204–212. doi: 10.1309/AJCPQYW3B6JLKILC
Drancourt, M., Bollet, C., Carlioz, A., Martelin, R., Gayral, J.-P., and Raoult, D. (2000). 16S Ribosomal DNA sequence analysis of a large collection of environmental and clinical unidentifiable bacterial isolates. J. Clin. Microbiol. 38, 3623–3630. doi: 10.1128/JCM.38.10.3623-3630.2000
EFSA. (2019). Multi-country outbreak of listeria monocytogenes clonal complex 8 infections linked to consumption of cold-smoked fish products. EFSA Support. Public. 16:1665E. doi: 10.2903/sp.efsa.2019.EN-1665
Fan, W.-T., Qin, T.-T., Bi, R.-R., Kang, H.-Q., Ma, P., and Gu, B. (2017). Performance of the matrix-assisted laser desorption ionization time-of-flight mass spectrometry system for rapid identification of streptococci: a review. Eur. J. Clin. Microbiol. Infect. Dis. 6, 1005–1012. doi: 10.1007/s10096-016-2879-2
FAO. (2018). La Situation Mondiale Des Pêches et de L’aquaculture 2018. Atteindre Les Objectifs de Développement Durable. Available online at: http://www.fao.org/3/i9540fr/i9540fr.pdf (accessed December 17, 2020)
Giebel, R., Worden, C., Rust, S. M., Kleinheinz, G. T., Robbins, M., and Sandrin, T. R. (2010). Microbial fingerprinting using matrix-assisted laser desorption ionization time-of-flight mass spectrometry (MALDI-TOF MS) applications and challenges. Adv. Appl. Microbiol. 71, 149–184. doi: 10.1016/S0065-2164(10)71006-6
Iwamoto, M., Ayers, T., Mahon, B. E., and Swerdlow, D. L. (2010). Epidemiology of seafood-associated infections in the United States. Clin. Microbiol. Rev. 23, 399–411. doi: 10.1128/CMR.00059-09
Lallemand, E., Arvieux, C., Coiffier, G., Polard, J.-L., Albert, J.-D., Guggenbuhl, P., et al. (2017). Use of MALDI-TOF mass spectrometry after liquid enrichment (BD BactecTM) for rapid diagnosis of bone and joint infections. Res. Microbiol. 168, 122–129. doi: 10.1016/j.resmic.2016.09.005
Li, Y., Shan, M., Zhu, Z., Mao, X., Yan, M., Chen, Y., et al. (2019). Application of MALDI-TOF MS to rapid identification of anaerobic bacteria. BMC Infect. Dis. 19:941. doi: 10.1186/s12879-019-4584-0
Ling, H., Yuan, Z., Shen, J., Wang, Z., and Xu, Y. (2014). Accuracy of matrix-assisted laser desorption ionization–time of flight mass spectrometry for identification of clinical pathogenic fungi: a meta-analysis. J. Clin. Microbiol. 52, 2573–2582. doi: 10.1128/JCM.00700-14
Midelet-Bourdin, G., Leleu, G., and Malle, P. (2007). Evaluation of the international reference methods NF EN ISO 11290-1 and 11290-2 and an in-house method for the isolation of listeria monocytogenes from retail seafood products in france. J. Food Prot. 70, 891–900. doi: 10.4315/0362-028X-70.4.891
Olofsson, T. C., Ahrné, S., and Molin, G. (2007). The bacterial flora of vacuum-packed cold-smoked salmon stored at 7°C, identified by direct 16S rRNA gene analysis and pure culture technique. J. Appl. Microbiol. 103, 109–119. doi: 10.1111/j.1365-2672.2006.03216.x
Popović, N. T., Kazazić, S. P., Strunjak-Perović, I., and Čož-Rakovac, R. (2017). Differentiation of environmental aquatic bacterial isolates by MALDI-TOF MS. Environ. Res. 152, 7–16. doi: 10.1016/j.envres.2016.09.020
Schröttner, P., Rudolph, W. W., Eing, B. R., Bertram, S., and Gunzer, F. (2014). Comparison of VITEK2, MALDI-TOF MS, and 16S rDNA sequencing for identification of Myroides odoratus and Myroides odoratimimus. Diagnostic Microbiol. Infect. Dis. 79, 155–159. doi: 10.1016/j.diagmicrobio.2014.02.002
Schulthess, B., Bloemberg, G. V., Zbinden, A., Mouttet, F., Zbinden, R., Böttger, E. C., et al. (2016). Evaluation of the Bruker MALDI Biotyper for identification of fastidious gram-negative rods. J. Clin. Microbiol. 54, 543–548. doi: 10.1128/JCM.03107-15
Takahashi, N., Nagai, S., Fujita, A., Ido, Y., Kato, K., Saito, A., et al. (2020). Discrimination of psychrotolerant Bacillus cereus group based on MALDI-TOF MS analysis of ribosomal subunit proteins. Food Microbiol. 91:103542. doi: 10.1016/j.fm.2020.103542
Tsuchida, S., Umemura, H., and Nakayama, T. (2020). Current status of matrix-assisted laser desorption/ionization–time-of-flight mass spectrometry (MALDI-TOF MS) in clinical diagnostic microbiology. Molecules 25:4775. doi: 10.3390/molecules25204775
Wieser, A., Schneider, L., Jung, J., and Schubert, S. (2012). MALDI-TOF MS in microbiological diagnostics—identification of microorganisms and beyond (mini review). Appl. Microbiol. Biotechnol. 93, 965–974. doi: 10.1007/s00253-011-3783-4
Keywords: marine microbiology, seafood, sea water, ecology, bacterial identification, 16s rDNA sequencing, food pathogens and spoilage, MALDI-TOF MS
Citation: Brauge T, Trigueros S, Briet A, Debuiche S, Leleu G, Gassilloud B, Wilhelm A, Py J-S and Midelet G (2021) MALDI-TOF Mass Spectrometry Fingerprinting Performance Versus 16S rDNA Sequencing to Identify Bacterial Microflora From Seafood Products and Sea Water Samples. Front. Mar. Sci. 8:650116. doi: 10.3389/fmars.2021.650116
Received: 07 January 2021; Accepted: 17 March 2021;
Published: 28 April 2021.
Edited by:
Andrew Stanley Mount, Clemson University, United StatesReviewed by:
Karola Böhme, University of Santiago de Compostela, SpainCopyright © 2021 Brauge, Trigueros, Briet, Debuiche, Leleu, Gassilloud, Wilhelm, Py and Midelet. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Thomas Brauge, dGhvbWFzLmJyYXVnZUBhbnNlcy5mcg==
†These authors share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.