
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Mar. Sci., 08 June 2021
Sec. Marine Megafauna
Volume 8 - 2021 | https://doi.org/10.3389/fmars.2021.646021
 Anmari Alvarez-Alemán1,2,3*†
Anmari Alvarez-Alemán1,2,3*† Eddy García Alfonso4James A. Powell2
Eddy García Alfonso4James A. Powell2 Charles A. Jacoby5
Charles A. Jacoby5 James D. Austin1,6†
James D. Austin1,6† Thomas K. Frazer1,7
Thomas K. Frazer1,7The Antillean manatee is categorized as endangered by the International Union for the Conservation of Nature primarily due to its low abundance. Further declines in abundance are expected due to a group of human related threats that include illegal hunting, fatal interactions with fishing gear, increasing watercraft traffic, and pollution. The nature of current threats is not well described for every critical population of manatees. In particular, threats to manatees have not been documented or evaluated in a systematic manner in coastal waters surrounding Cuba, which is a major reason why plans specifically addressing conservation of manatees in Cuba have not been implemented. To address this lack of information, we assessed patterns in various threats by reviewing reports of stranded (alive or dead) and captured manatees, with some reports being historical and others consistently available since 2001. For 94% of the contemporary cases, carcasses of dead animals were reported; the remainder (6%) involved live manatees and subsequent efforts to rescue and rehabilitate them. Documented causes of mortality included poaching (38%), entanglement in fishing gear (10%), deaths of dependent calves (2.5%), and boat collisions (2.5%). The cause of death for the remaining 47% of reports could not be determined. Mortalities were highest in areas with intensive commercial fishing effort, and mortalities in marine protected areas indicated a need for improved management. These initial findings provide the first estimates of the magnitude of mortality of manatees in Cuba and yield valuable insights into the causes of deaths. The existence of intense and poorly regulated fishing in certain areas, including protected areas, represents a potentially significant threat to manatees. The effects and management of this activity, especially in habitats that are critical for manatees, merit further attention.
Megafauna (large-bodied terrestrial, freshwater, and marine vertebrates) are 2.75 times more likely to be threatened by extinction than other vertebrate species that have been assessed by the International Union for the Conservation of Nature (IUCN, 2012; Ripple et al., 2019). Marine megafauna, including large mammals (>45 kg), play important ecological roles that include transporting nutrients through long distance migrations and dispersal and modifying habitats via feeding, mortality, and locomotion (Pimiento et al., 2020). Marine megafauna, like their terrestrial counterparts, face a number of threats related to human exploitation, global warming, and habitat loss (Ripple et al., 2019; Pimiento et al., 2020).
Many of the threats that currently affect marine megafauna have impacted sirenians (manatees and dugongs). The vulnerability of this group was demonstrated when the Steller sea cow (Hydrodamalis gigas) was harvested to extinction in the Bering Sea and local populations of manatees were extirpated in parts of their geographic range (Marsh et al., 2011). The West Indian manatee (Trichechus manatus, Linnaeus 1758) is one of the four extant sirenian species, with two subspecies (T. m. manatus and T. m. latirostris) declared as endangered by the IUCN (Deutsch, 2008; Self-Sullivan and Mignucci-Giannoni, 2008; Marsh et al., 2011). The distribution of West Indian manatees is fragmented, with total numbers of T. m. latirostris along the east and west coasts of Florida estimated at 7,520–10,280 individuals and the more dispersed populations of T. m. manatus thought to range from 50 to 1,250 individuals (Lefebvre et al., 2001; Quintana-Rizzo and Reynolds, 2010; Castelblanco-Martínez et al., 2012; Self-Sullivan and Mignucci-Giannoni, 2012; Martin et al., 2015; Hostetler et al., 2018). Reduced population sizes and insufficient resources for effective conservation threaten the remaining populations of T. m. manatus distributed among 13 island nations, 12 continental nations, and 13 territories throughout the Caribbean (Marsh et al., 2011).
Mortality of T. manatus has been linked to several anthropogenic and natural factors (Bossart et al., 1998, 2002a,b; Langtimm et al., 1998, 2004; Lightsey et al., 2006; Marsh et al., 2011). Anthropogenic threats are associated with hunting, interaction with fisheries, collisions with watercraft, and habitat loss, whereas natural threats are related to the incidence of diseases, parasites, predation, and stress from adverse climatic conditions (Bossart et al., 1998, 2002a,b; Langtimm et al., 1998, 2004; Mignucci-Giannoni et al., 2000; Parente et al., 2004; Lightsey et al., 2006; Marsh et al., 2011; Borges et al., 2017; Wyrosdick et al., 2018).
In addition, it has been predicted that climate change will have detrimental effects on manatees. For example, the near-term effects of climate change (e.g., increased red tides, extreme cold weather, and increases in the frequency and intensity of storms), coupled with changes in habitats due to loss of warm water refuges and increased boat traffic could affect manatees detrimentally (Edwards, 2013). In fact, the cumulative effects of climate change on habitats and many animals are projected to surpass the effects of human-related land use (Dawson et al., 2011); therefore, reducing manageable threats now is essential because the future promises even greater challenges from increasing human populations and a rapidly changing climate (Marsh et al., 2011).
Overall, information on causes of mortality is available primarily for manatees in Florida (Lightsey et al., 2006; FWC, 2021). Mortality in the Caribbean and South American populations of Antillean manatees has been examined in five countries (e.g., Puerto Rico, Colombia, Brazil, Mexico, and Belize; Mignucci-Giannoni et al., 2000; Montoya-Ospina et al., 2001; Parente et al., 2004; Meirelles, 2008; Castelblanco-Martínez et al., 2009, 2018; Balensiefer et al., 2017); however, basic information about key threats is lacking for the remaining populations. Such information is important because identifying the key causes of preventable mortality is a critical step toward effective conservation.
In particular, Cuba is the largest island in the Caribbean, and it has large expanses of healthy coastal habitats that could potentially sustain considerable numbers of manatees (Alcolado, 2006). However, the population size on the Cuban archipelago is believed to be small (Alvarez-Alemán et al., 2018), and the proximate causes of mortality for manatees have not been systematically evaluated or well documented. As a consequence of the poor understanding of the status of the local population, appropriate management plans have not been developed.
Here, we consolidate empirical information for manatees in Cuba to evaluate patterns in strandings and mortalities. This assessment of threats to manatees is the first for Cuba, and we place it in the context of existing environmental policies, with the goal of recommending positive steps toward protecting manatees and promote the survival of this endangered population.
The events evaluated herein were historical and contemporary in nature. Historical reports from 1950 to 1988 were obtained from museums, university collections, and the literature. Contemporary reports included events from 2001 to 2018, and they were collected opportunistically through a multi-institutional network of biologists, locals, fishers, and government officials from coastal communities and marine protected areas (Alvarez-Alemán et al., 2014). The records were verified by trained staff (usually biologist and park rangers), with the main national institutions involved in this network being the Center for Marine Research from the University of Havana, the National Enterprise of Flora and Fauna from The Ministry of Agriculture, and the National Center for Protected areas from the Ministry of Science, Technology and Environment (CNAP-CITMA).
Whenever possible, entries in the database included date, latitude and longitude, corresponding fishery zone and subzone, presence or absence of protection for the site, condition of the carcass, sex, length (cm), size class (visual estimates to classify manatees as possible calves or adults), and cause of death (Bonde et al., 1983; Mignucci-Giannoni et al., 2000).
When length was available, we classified the manatees based on a scheme similar to that applied to manatees in Florida and Belize (calf < 235 cm, sub-adult 235–264 cm, or adult > 264 cm; Bonde et al., 2012a; Allen et al., 2018; Galves and Powell, pers. comm.). For comparative purposes we also provide classifications based on the scheme used by Mignucci-Giannoni et al. (2000) for manatees from Puerto Rico (calves < 175 cm, subadults 176–225 cm, and adults > 225). Size categories for both subspecies were included because both have been applied in the Caribbean and Florida and there are no data to guide the selection of a scheme for Cuba.
In some cases, a physical examination was performed on the animal (dead or alive), and samples were collected from dead animals following the protocol for necropsy and assessing mortality described by Bonde et al. (1983). Reports lacking a physical examination or verification by trained observers also were incorporated when they had reliable species identifications, photographs, dates, and locations. Some reports were for stranded live manatees, and, in these cases, the animals were returned to the water. Sex was assessed by morphological examination or through a genetic test (see methodology in Tringali et al., 2008). Periotic bones from museum specimens (n = 10, these specimens were not included in other analyses of the historic or contemporary data due to incomplete records) and some of the carcasses (n = 33) were analyzed by counting growth layer groups (GLGs) to yield age-at-death (see methodology in Brill et al., 2016). The estimated average age at death was calculated and compared with other populations.
We investigated spatial patterns in contemporary events around Cuba and evaluated the occurrence of human-related mortality. We coded six parameters to characterize each event (Table 1; Bonde et al., 1983). The first parameter, cause of stranding or mortality, included unknown, being a dependent calf (if the manatees were smaller than 150 cm), entanglement in fishing gear (a sighting in a trawl or marks, particularly on the head and flippers, that were consistent with swimming into and being abraded by a net or trawl), injury compatible with an impact from a watercraft (observation of a collision or an epidermal wound consistent with a collision or contact with a propellor), and poaching (dismembered carcasses or carcasses with meat removed). Additionally, locations of strandings and mortalities were characterized as being within or outside of a protected area; being within an area where the number of state-owned fishing vessels was either less than 50 or at least 50; being within an area with low (<100 boats), medium (100–200 boats), or high (>200 boats) numbers of private fishing vessels; being within an area where fishing targeted either lobster or finfish; and whether or not the stranding or mortality was in an area with an organized network that reported such events. The resulting information related to 85 events was used to perform a hierarchical clustering based on Bray–Curtis resemblance measures using the software PRIMER 6 (Anderson et al., 2008). Groups of events with similar characteristics were identified with a SIMPROF analysis (similarities profile), and the events were given codes for the appropriate group to accompany their latitudes and longitudes. The matrix generated in the SIMPROF analysis was evaluated with a SIMPER analysis (similarities percentages) to identify the parameters primarily responsible for differentiating the groups.
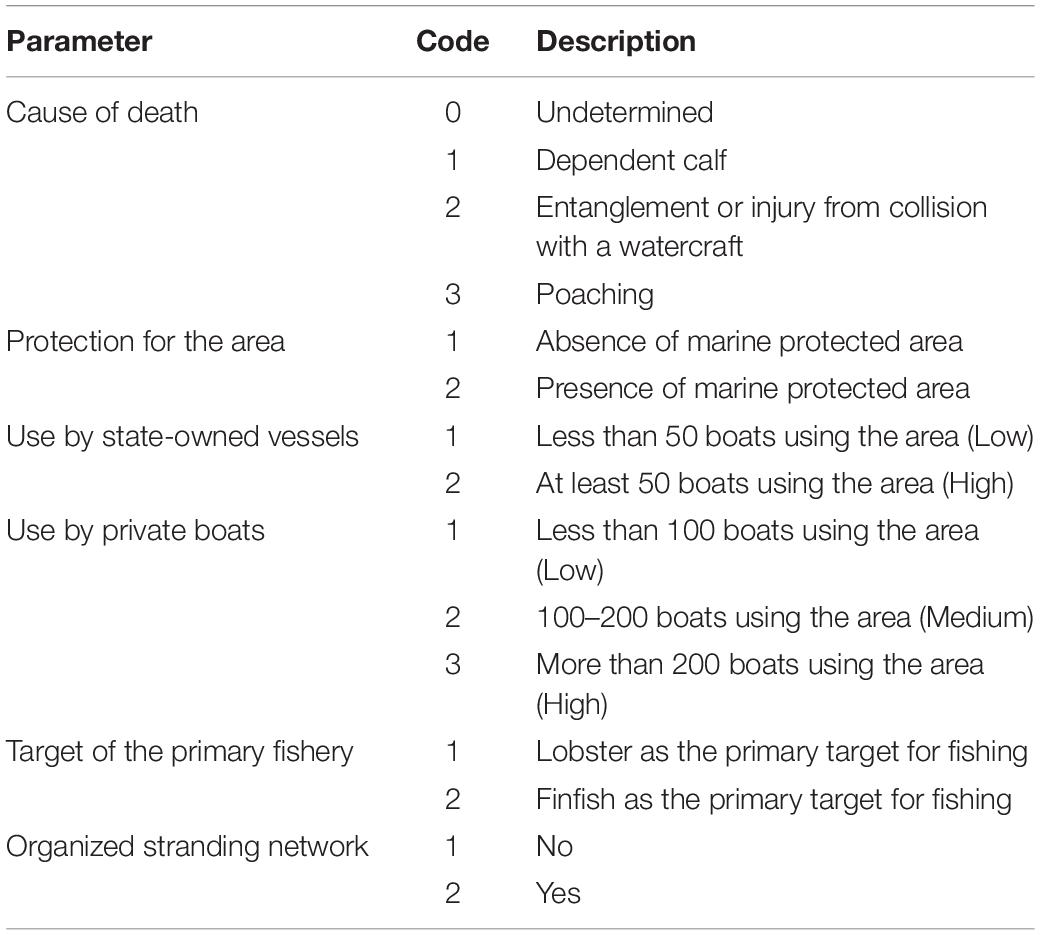
Table 1. Codes created for six parameters that characterize contemporary events affecting manatees in Cuba, with the numeric codes forming the basis for hierarchical clustering of the events.
Thirteen historical events between 1950 and 1988 were drawn from museum and university records, published literature, and local sources of information (Table 2). Five of these individuals were captured for exhibition in the National Aquarium, Zoological Park of Havana and the museum “Ignacio Agramonte” in Camaguey, so they were not included in further analyses (Table 2). Three manatees died due to entanglement in fishing nets, and five were found dead from unknown causes (Table 2). Additionally, osteological samples from eight specimens were located in the Felipe Poey’s museum at the University of Havana, the Museum of Sagua in Villa Clara, and the Archeology Institute (Gabinete de Arqueología), but lack of information on their origins and dates of collection meant they were used to estimate age-at-death but they were not included in the database.
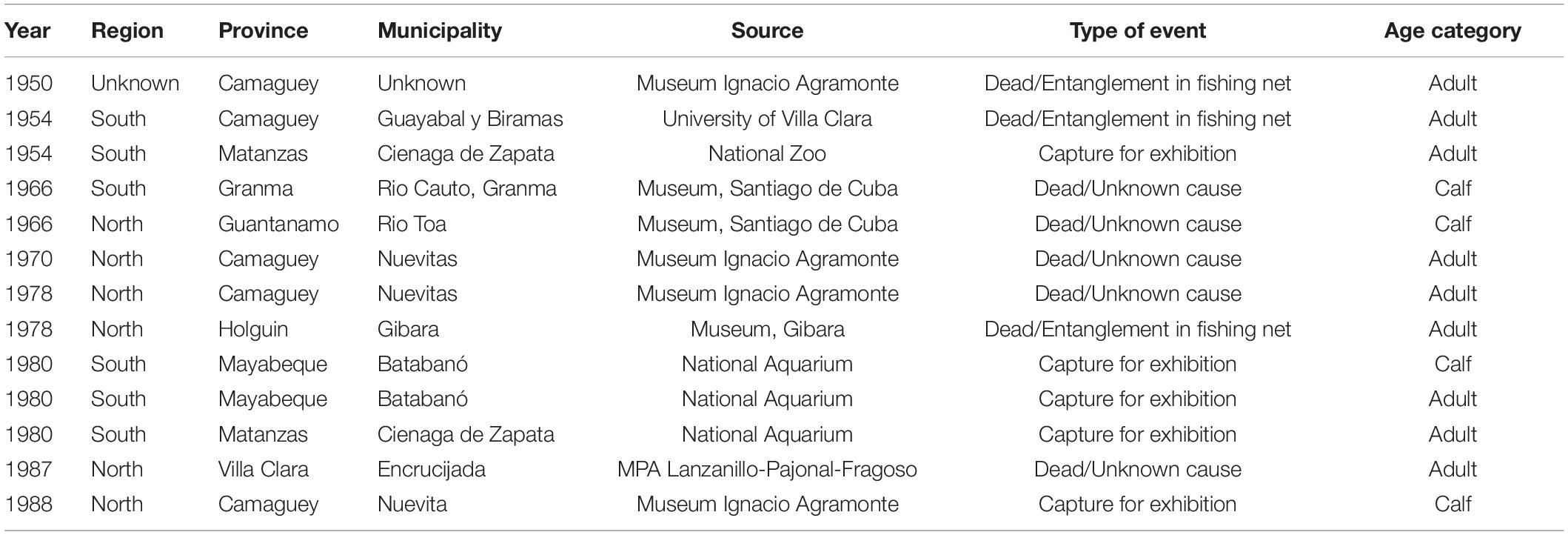
Table 2. Historical reports of events involving manatees between 1950 and 1988.
Events involving 85 manatees were recorded between 2001 and 2018. These cases were separated into three categories: dead (80 manatees), alive and released (3 manatees), and alive and subsequently died in captivity (2 manatees). The latter were calves that were only a few days old. The manatees that were released alive were found within MPAs, and they were entangled in fishing nets or exhibited signs of entanglement. Two manatees were observed in the Laguna del Tesoro, Zapata Swamp and the third in Villa Clara. The average number of deaths per year was 4.6, with a range of 1 to 14 individuals.
For the dead animals, 38% represented dried carcasses (usually loose bones with dried tissues attached). A further 42% of the dead animals were moderately to badly decomposed carcasses, and 20% of the dead manatees were fresh. In 20% of the cases, a physical examination of the carcass was not possible.
Across all mortalities, the causes of death recorded between 2001 and 2018 were poaching (38%), entanglement in fishing nets (10%), dependent calves (2.5%), and high probability of a collision with a watercraft (2.5%). One of these two cases involved a tourist boat operating in the Laguna del Tesoro section of the Zapata Swamp (Hurtado, pers. comm.). All deaths except those of dependent calves attributed directly to human activities (i.e., 50.5%). For the remaining 47% of carcasses, the cause of death was unknown primarily because the record was based on dried bones or highly decomposed carcasses. These records added uncertainty to the evaluation of human activities as a cause of mortality.
Historical and contemporary records for strandings and mortalities yielded information on the sex, size class, and, when it could be measured, the length of manatees found stranded or dead. Out of the 32 historical and contemporary reports with direct observations (n = 25) or genetic tests (n = 7) to determine sex, 22 manatees were females (18 detected visually and 4 with genetic tests) and only 10 were males (7 detected visually and 3 with genetic test). According to visual classifications of carcasses or body parts by observers, 63 individuals were identified as adults and 18 as calves. In addition, lengths were recorded for 26 manatees. The maximum length was 360 cm, and the minimum length was 129 cm. The animals were classified as calves (N = 8, mean length = 142 cm), subadults (N = 5, mean length = 201 cm), and adults (N = 13, mean length = 305 cm) according to categories in Mignucci-Giannoni et al. (2000), and as calves (N = 14, mean length = 169 cm), a subadult (N = 1, length = 250 cm), and adults (N = 11, mean length = 317 cm) according to Bonde et al. (2012a).
Historical or contemporary events occurred in 15 of the 16 provinces in Cuba, with the 85 contemporary events involving 57 manatees found in the north and 28 in the south (Figure 1). Events were sparsely distributed around the Cuban archipelago, with the exception of northern Villa Clara where a higher number of manatees has been found over the years (Figure 1).
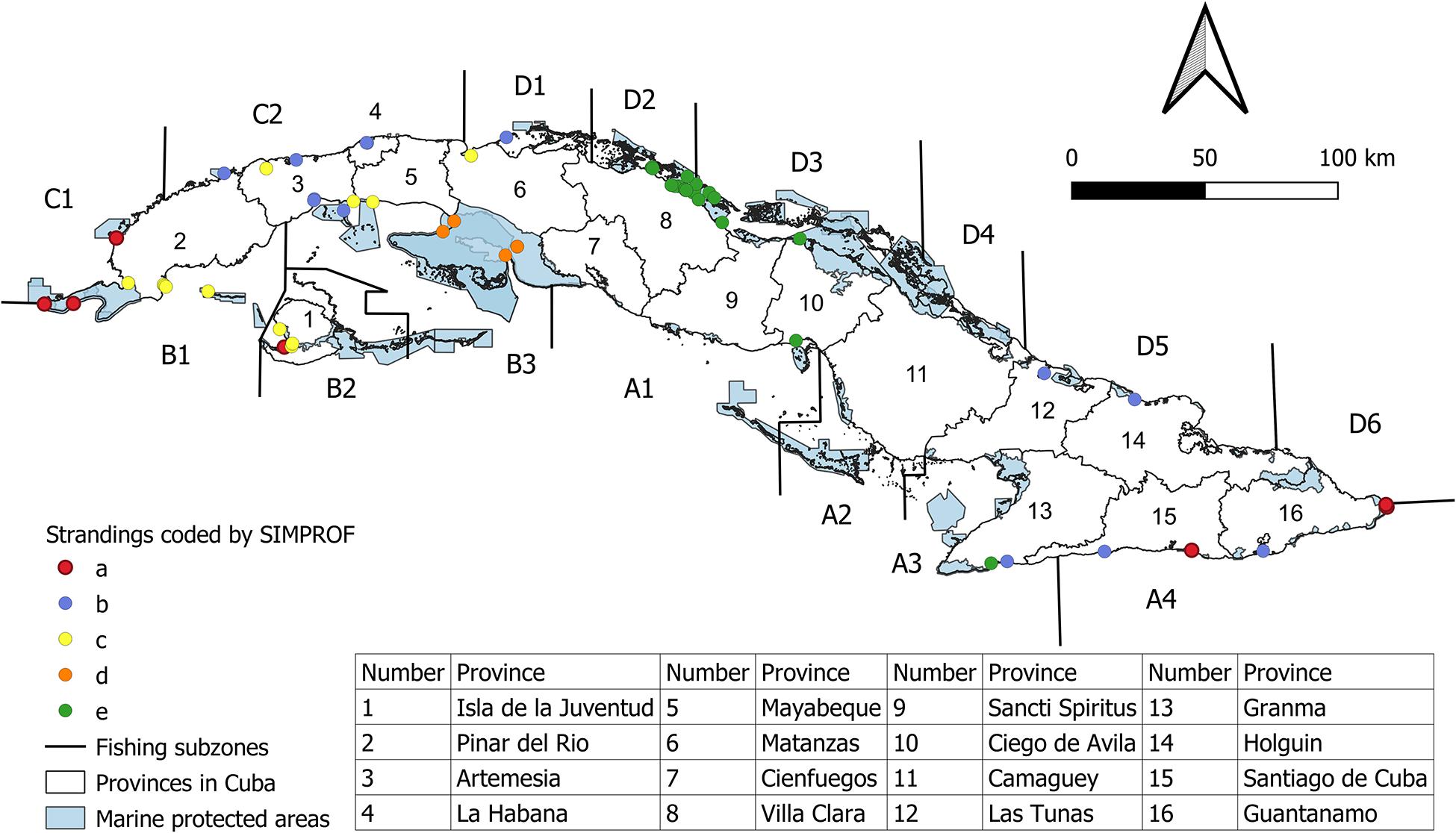
Figure 1. A map showing mortalities of manatees. Colored dots = events categorized in five groups (a–e) generated by a similarities profile analysis (SIMPROF), letters followed by numbers = fishing subzones, blue-shaded areas = approved marine protected areas, and numbers = provinces as listed in the table. Some colored dots overlap because mortalities occurred at the same place at different times.
Seventy-two percent of the 85 events were reported from inside 11 MPAs (Table 3 and Figure 1), and 52% of these cases involved mortality related to human activities. The types of protection associated with these areas varied, with some having a formal declaration, legal status, and administration at the national level and others being local efforts with less formal recognition (Table 3). Caches of bones from manatees were found buried in three nationally protected areas and one locally protected area, which was evidence of past mortality due to human activities (Table 3).
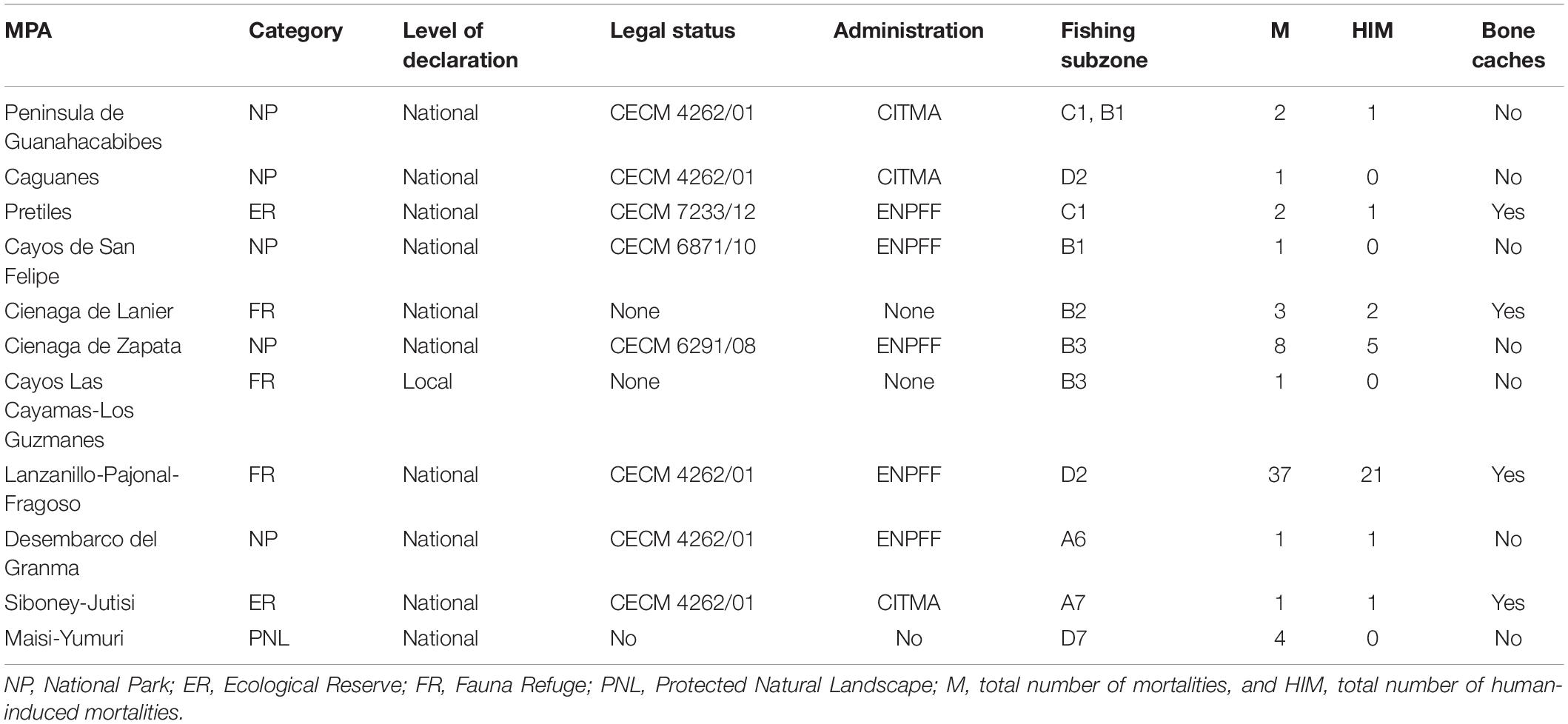
Table 3. Information about marine protected areas (MPAs) in Cuba and events involving manatees until 2018.
Manatee strandings and mortalities occurred in all four fishing zones (A, B, C, and D), and specifically, in 12 out of the 15 fishing subzones (Tables 3, 4 and Figure 1; Resolution 226/20005). The highest number of manatees was reported for zone D (46), followed by zone B (22), zone C (11), and zone A (6). Zone D also had a higher number of fishing boats (state-owned and private; Table 4).
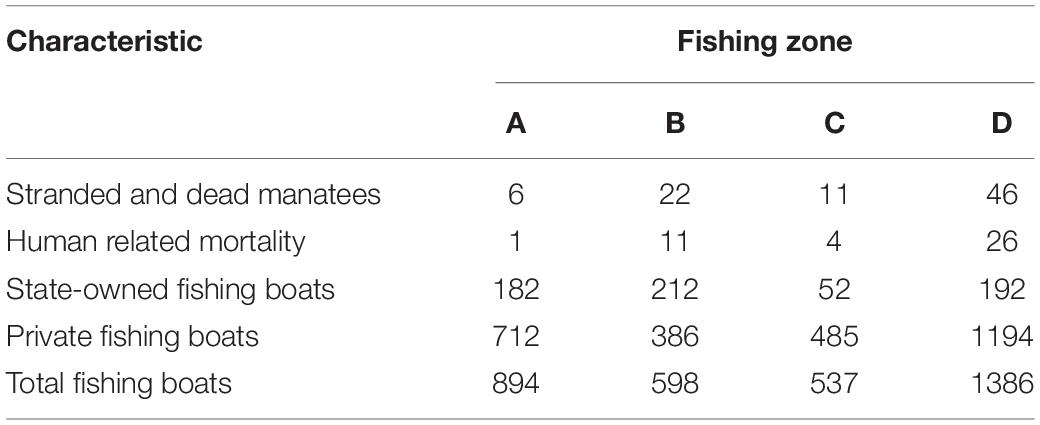
Table 4. Data related to contemporary events involving manatees, human-related mortality, and registered fishing boats per fishing zone between 2001 and 2018.
The SIMPROF analysis identified five groups of events (a–e; Figure 1). A group of 10 events (a) and a group of 13 events (b) were sparsely distributed across Cuba, while groups of 14 (c), 8 (d), and 40 (e) reports were concentrated in relatively small areas that were found mostly in the west of the country (Figure 1). Overall, group e accounted for 47% of the mortalities. Furthermore, results from the SIMPER analysis (Table 5) indicated that:
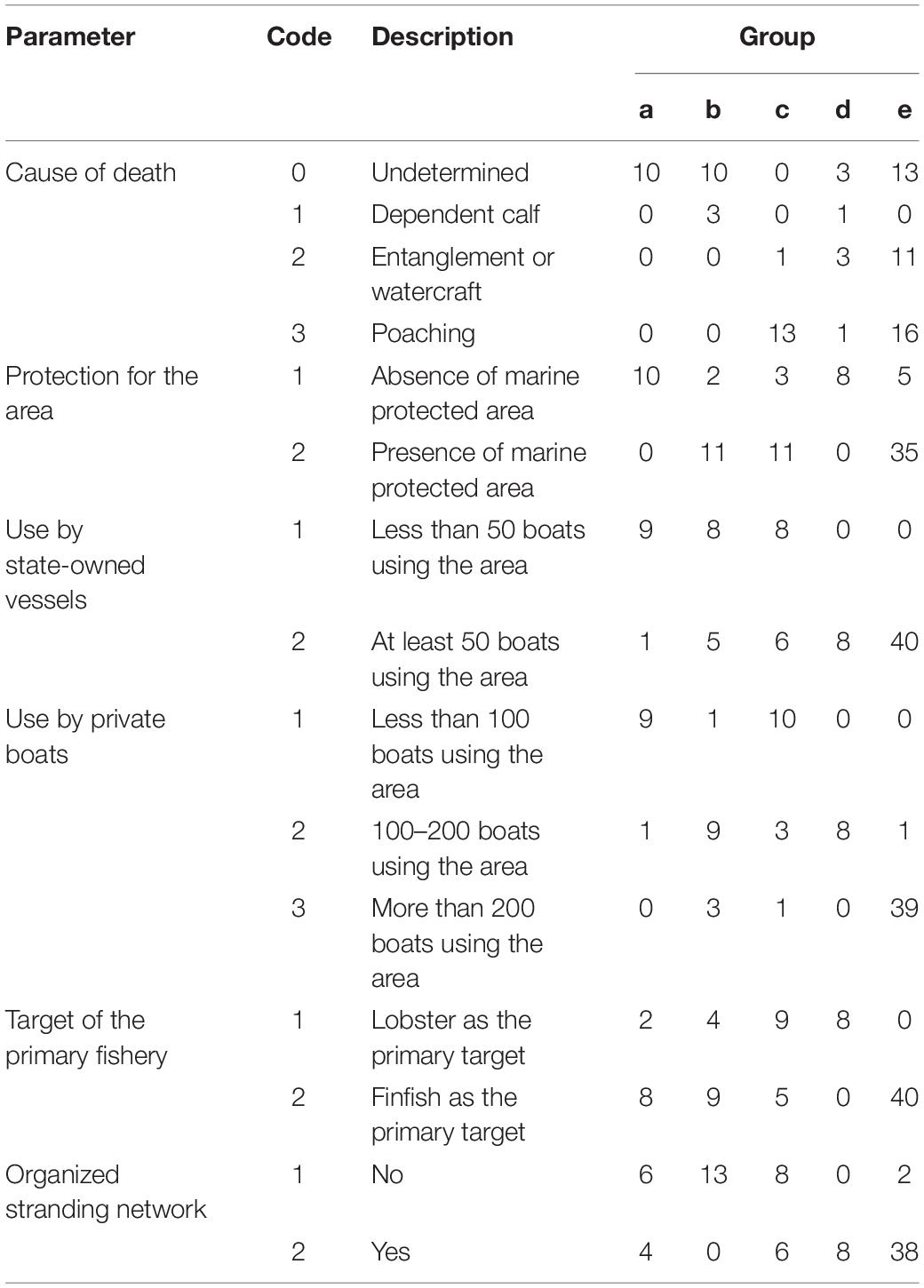
Table 5. Numbers of mortalities for each of the codes for the six parameters that characterize contemporary events affecting manatees in Cuba across the five groups of events generated in the SIMPROF analysis.
• Group a comprised mortalities with undetermined causes that occurred in areas without protection, few state-owned or private boats, some fishing for finfish, and possibly an organized stranding network;
• Group b comprised mortalities with undetermined causes or deaths of dependent calves that occurred primarily in areas with protection, a moderate to high number of boats, fishing mainly for finfish, and the absence of an organized stranding network;
• Group c comprised mortalities due to poaching that occurred primarily in areas with protection, a moderate to high number of boats, fishing for lobster and finfish, and possibly an organized stranding network;
• Group d comprised mortalities split among the causes that occurred in areas without protection, a moderate number of boats, fishing mainly for lobster, and the presence of an organized stranding network; and
• Group e comprised mortalities with undetermined causes or deaths caused by entanglement or poaching that occurred primarily in areas with protection, a high number of boats, fishing mainly for finfish, and the presence of an organized stranding network.
The high incidence of cases in group e found along the north coast of Cuba (Villa Clara; Figure 1) could have been due to the existence of monitoring programs and higher conservation effort. Nonetheless, the variable that denoted the presence or absence of organized surveys to detect strandings and mortalities, was responsible for an average of only 13% of the dissimilarity between group e and the other groups.
The estimated average age at death for 43 specimens (33 specimens from contemporary events and 9 from historical events) was 9 years old (individuals ranged from less than 1 to 40 years old). Thirty five percent of the manatees were 1 year old or less. Florida manatees (T. m. latirostris) reach sexual maturity between the ages of 2 and 6 years old (Marmontel, 1995), so using these values as references, 35–48% of the stranded or dead manatees were sexually immature. Sixty-seven percent of the manatees evaluated were less than 10 years old, and only seven manatees were more than 15 years old. The estimated average age for museum specimens was 8.8 years while for historical and contemporary carcasses it was 8.5 years. Based on analyses of periotic bones for contemporary cases and previous reports (Marmontel et al., 1992; Mignucci-Giannoni et al., 2000), manatees were classified as calves (<2 years, N = 13), subadults (2–7 years, N = 5), and adults (>7 years, N = 15).
Manatee strandings in Cuba primarily involved dead animals. Live strandings requiring assistance and rehabilitation were rare. The numbers of events, and presumably those specifically related to human activity, could be underestimated for several reasons. First, some areas are inaccessible to volunteers and biologists; therefore, carcasses are not detected or recovered. Furthermore, manatee hunting is illegal in Cuba (Decreto de Ley 164/1997; Ley No 129/2019), and it is likely that poachers tend to hide the carcasses to avoid punishment. Hidden piles of bones demonstrate that this attitude could exist among poachers, with loose bones or bones in boxes or bags found in some offshore localities (fishing subzones C1, B2, D2, and A7; Table 3). Finally, fishers or other members of a coastal community may not report a manatee carcass because they are afraid of being wrongly accused as responsible for its death. Despite this potential bias, human-related mortality was recorded frequently, with primary causes being poaching and entanglement in fishing nets.
This work documents poaching as a relatively frequent cause of death for West Indian manatees in Cuba (38%). Even higher rates were documented in Colombia, where 39–97% of the mortalities were attributed to poaching (Montoya-Ospina et al., 2001; Castelblanco-Martínez et al., 2009; Table 6). Poaching has been reported for other populations of Antillean manatees, but with a lower frequency of occurrence (Table 6). Puerto Rico and Brazil recorded poaching as the cause of 31 and 4% of the human-related mortalities, respectively (Mignucci-Giannoni et al., 2000; Meirelles, 2008). In Belize, very little recent mortality from poaching has been reported, and such events were not reported in Mexico (Castelblanco-Martínez et al., 2018).
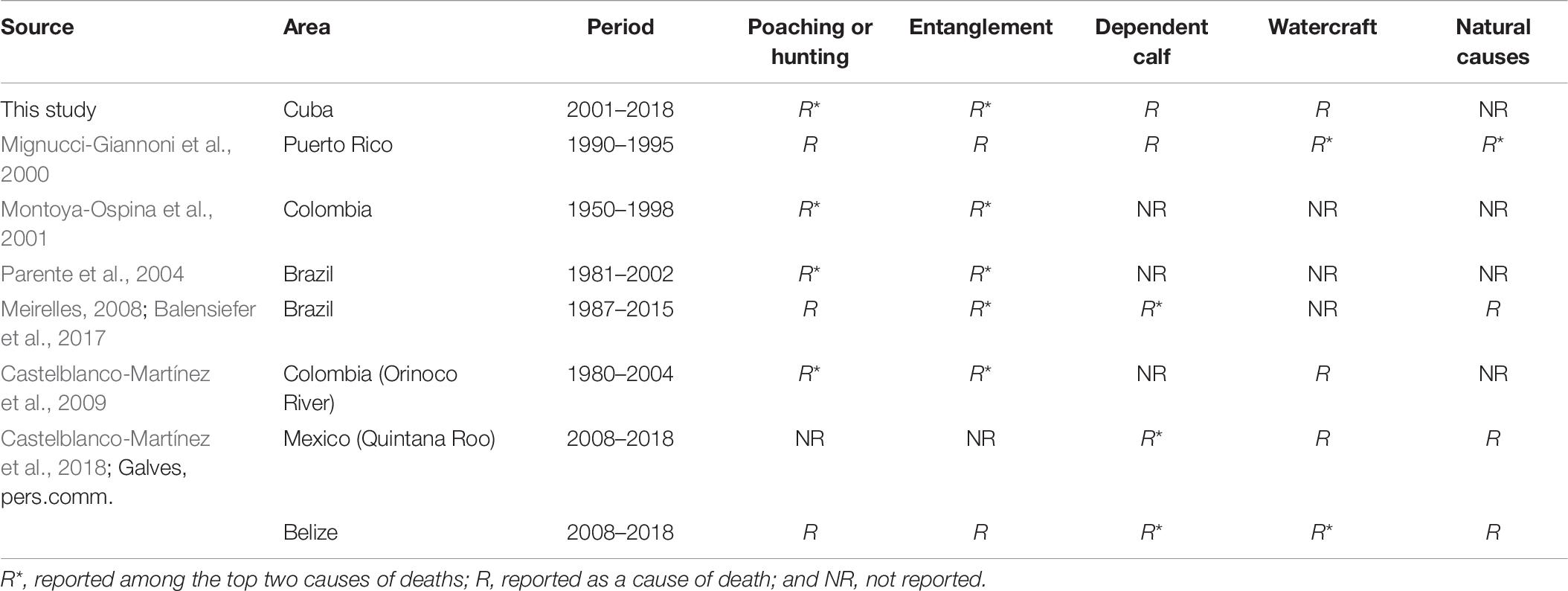
Table 6. Information about mortalities of Antillean manatees in six populations.
Entanglement in fishing nets has been reported as a cause of death for many populations of Antillean manatees (Mignucci-Giannoni et al., 2000; Meirelles, 2008; Castelblanco-Martínez et al., 2018). In Colombia, this was the most common cause of mortality, with 43% of the reported cases (Castelblanco-Martínez et al., 2009). Our study recorded the presence of this threat, but more detailed records are needed to determine if habitats that attract manatees also support more abundant stocks of valuable species that, in turn, attract fishers; if manatees represent a “targeted catch,” with an illegal market that generates demand as seen for marine turtles; or if the interactions typically are accidental (Forneiro et al., 2015).
Watercraft-related trauma was not observed frequently in Cuba, although it has been reported as a significant contributor to manatee mortality in Florida, Puerto Rico, and Belize (Mignucci-Giannoni et al., 2000; Lightsey et al., 2006; Bonde et al., 2012b; Castelblanco-Martínez et al., 2018, Galves, pers. comm.). Such a result is not unexpected given that high-powered watercraft are rare in Cuba, and manatees do not form large aggregations in busy waterways. However, a similar situation changed in Belize, so the future may yield different results (Castelblanco-Martínez et al., 2018, Galves, pers. comm.).
Females represented more carcasses than males. The same result occurred in Belize, where the sex of 145 carcasses was known, and 83 were females (Castelblanco-Martínez et al., 2018). This situation was not the case for Puerto Rico where males and females constituted 51.2 and 48.8% of the cases, respectively (Mignucci-Giannoni et al., 2000). In Quintana Roo Mexico, out of 32 manatees with known sex, 18 were males (Castelblanco-Martínez et al., 2018). Also, a study in Brazil recorded mortalities at a female:male ratio of 1:1 (Balensiefer et al., 2017). A previous study on the endangered giant panda (Ailuropoda melanoleuca), demonstrated that the severity of effects from poaching depend on the sex and age of the poached animals (Li et al., 2003). Poaching more females than males can reduce the size of the present generation and the number of new-born individuals contributing to the following generation (Li et al., 2003). Monitoring the incidence of stranded or dead females and finding a suitable method of assessing sex in decomposing carcasses remain critical needs.
Additional concerns about reduced contributions to future generations arise from strandings or mortalities involving calves. In many cases, these situations cannot be linked to an anthropogenic activity because the event that caused the calf to be separated from its mother is unknown. In this study, calves were involved in 22–54% of the incidents depending on the method used to classify individuals as calves. In most other studies, calves were involved in 12–83% of the recorded events (Mignucci-Giannoni et al., 2000; Montoya-Ospina et al., 2001; Meirelles, 2008; Castelblanco-Martínez et al., 2009, 2018; Balensiefer et al., 2017), with Parente et al. (2004) reporting that all 74 manatees that were stranded in northeast Brazil were calves. Overall, these data raise concerns regarding continued loss of reproductive potential.
The higher concentration of strandings and mortalities in north-central Cuba may be in part a result of a well-established network to report events in this region. Residents, fishers, coast guard personnel, and forest guards have a strong communication network that is activated when a dead or live manatee is found. This situation is especially the case in Villa Clara, and it has produced valuable information related to threats to manatees in this area. We recommend that a similar network be implemented across the rest of Cuba.
Mortality was relatively common inside MPAs. This result does not necessary imply that more manatees are dying or being killed inside protected areas, but one of the goals of MPAs should be to reduce the human-related mortalities of endangered species (Marsh et al., 2011). Therefore, the mortalities, suggest a need for improved management and conservation of such species.
Management of MPAs in Cuba is not consistent, and some of these areas could be considered “paper parks,” especially with regard to conservation (Perera-Valderrama et al., 2018). The primary barrier to reducing human-induced mortalities is the lack of resources and infrastructure to support enforcement (e.g., boats, vehicles, etc.; CNAP, 2013). Another potentially important issue is the overlap between fishing and MPAs (Perera-Valderrama et al., 2018). Restrictions on state and private fishing in the Sistema Nacional de Áreas Protegidas de Cuba are limited to terms found in an agreement with the Ministry of Food (Ministerio de la Industria Alimentaria; Perera-Valderrama et al., 2018). Overall, some protected areas in Cuba lack strictly enforced no-take areas, speed zones, or other regulations that reduce threats, and as a consequence, manatees are at risk of entanglement, poaching, collisions with watercraft, and potentially other detrimental interactions.
Two potentially productive steps toward elucidating interactions with fishing would be to explore relationships between habitats and the numbers of manatees and fishery species they attract and to facilitate networks of people to track manatees and their interactions with fishing, including consistent reporting of carcasses. Improved data collection could highlight between management objectives related to fisheries and those related to protecting endangered manatees. Resolving such conflicts could benefit all stakeholders.
The average age reported for Antillean manatees by Marmontel (1993) was 25 years old, which was substantially higher than the average age at death reported for manatees in Cuba. However, Marmontel (1995) estimated the age of 143 wild female Florida manatees and the average age was 12.6 years. Maximum age in manatees have been reported to be up to 60 years old in manatees and up to 73 in dugongs (Marsh et al., 2011). Uncertainty surrounds estimated ages of animals. For example, previous research indicated that counts of annual GLGs in the periotic domes of the tympanoperiotic complexes of manatees became less accurate for older animals, especially those over 15 years old (Lonati et al., 2019). However, the same study documented little to no resorption of annual GLGs for specimens ≤10 years old; therefore, we are confident in identifications of younger animals.
Negative repercussions could arise if individuals are removed before they are able to reproduce and leave offspring that contribute to the future gene pool (35–48% of contemporary mortalities were juveniles). Increased juvenile mortality means that adult survival rates need to increase if a population is to grow (Langtimm et al., 1998). Rigorous and consistent data on mortalities will be needed to evaluate these threats.
It is possible that poachers target certain sexes or size classes, as reported for indigenous hunters of dugong (Dugong dugon) in Australia that allegedly prefer breeding females to resting females or males because the former are fatter (Marsh et al., 2011). In our case, a well-designed, long-term monitoring program would provide valuable information.
Human related mortality constitutes an important group of threats that probably are underestimated for Cuba. Mortalities have persisted in fishing subzones where the use of nets is widespread, despite the existence of MPAs, and this result is unlikely to be due solely to disproportionate sampling effort. Overall, the persistence of mortality inside the system of MPAs suggests a need for examination of governing policies related to fishing and protection in those areas. Poaching also represents a threat, and the possibility that females and immature individuals are targeted needs to be evaluated and addressed through focused enforcement and increased public awareness of their protected status as these threats can have critical implications for recruitment. Lastly, like Florida and Belize, wild manatee viewing in certain areas could have an important economic return for the tourism industry and help incentivize government to improve enforcement of existing protections, re-evaluate the effects of fishing, and allocate resources to better protect Cuba’s endangered population of manatees.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
AA-A conceived the study and carried out the data collection. AA-A and CJ analyzed the data and prepared the tables and figures. All authors contributed to interpretation of the results and preparation and revision of the manuscript. All authors approved the submission.
This work was made possible by funding and support from the Sea to Shore Alliance, the John T. and Catherine D. MacArthur Foundation, the Eppley Foundation, the Waitt Foundation, and the Marine Mammal Commission (Grant MMC17-222). In addition, data collection was supported by the Tropical Conservation and Development Program at the University of Florida, and the GEF-UNDP-CNAP project “Regional approach to the management of the South Archipelagos of Cuba, 2009–2014.”
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We want to thank Empresa Nacional Para la Protección de Flora y Fauna, Centro Nacional de Áreas Protegidas and Centro de Investigaciones Marinas, Universidad de la Habana, Cuerpo de Guardabosques, Tropas Guardafronteras, and CITMA for their contributions to the Manatee Stranding Network in Cuba. This work resulted from a partnership among colleagues and volunteers across Cuba, and we thank D. Cobian, Y. Forneiro, Z. Hernandez, L. Rodriguez, Z. Navarro, A. Hurtado, J. A. Tamayo, F. Pina, L. Garcia, J. Angulo, D. Cruz, N. Hernandez, J. Izquierdo, A. Ruiz, Y. Lezcano, and R. Volta. Thanks to J. Hughes for the grammatical revision of the manuscript. Thanks also to K. Brill for processing and analyzing ear bones. Valuable advice and support were provided by J. Angulo, A. Read, P. Gerhartz, and R. Bonde. Finally, we thank the Cuban fishers for the information and valuable help they have provided during the collection of data and samples. The content of this manuscript was part of the first author’s dissertation at the University of Florida.
Alcolado, P. M. (2006). “Diversidad ecológica. Diversidad, utilidad y estado de conservación de los biotopos marinos,” in La Biodiversidad Marina de Cuba, ed. R. Claro (La Habana: Instituto de Oceanología Ministerio de Ciencia), 39.
Allen, A., Beck, C., Bonde, R., Powell, J., and Gomez, N. (2018). Diet of the Antillean manatee (Trichechus manatus manatus) in Belize, Central America. J. Mar. Biol. Associat. U K. 98, 1831–1840. doi: 10.1017/S0025315417000182
Alvarez-Alemán, A., Angulo-Valdés, J., Forneiro Martin-Viana, Y., and Powell, J. (2014). “Programa de monitoreo de manatí (Trichechus manatus),” in Estado actual de la biodiversidad marino-costera, en la región de los Archipiélagos del Sur de Cuba, ed. A. Hernádez- Ávila (La Habana: Centro Nacional de Áreas Protegidas), 200.
Alvarez-Alemán, A., García, E., Forneiro Martin-Viana, Y., Hernández-González, Z., Escalona-Domenech, R., Hurtado, A., et al. (2018). Status and conservation of manatees in Cuba: historical observations and recent insights. Bull. Mar. Sci. 94, 313–327. doi: 10.5343/bms.2016.1132
Anderson, M., Gorley, R. N., and Clarke, R. K. (2008). PERMANOVA+ for primer: Guide to software and Primer-E Limited. Devon: PRIMER-E Ltd.
Balensiefer, D. C., Attademo, F. L. N., Sousa, G., Freire, A. C., da Cunha, F. A. G. C., Barboza-Alencar, A. E., et al. (2017). Three decades of Antillean manatee (Trichechus manatus manatus) stranding along the Brazilian coast. Trop. Conserv. Sci. 10, 1–9. doi: 10.1177/1940082917728375
Bonde, R. K., Garret, A., Belanger, M., Askin, N., Tan, L., and Wittnich, C. (2012a). Biomedical health assessments of the Florida manatee in Crystal River - providing opportunities for training during the capture, handling, and processing of this endangered aquatic mammal. J. Mar. Anim. Ecol. 5, 17–28.
Bonde, R. K., Mignucci-Giannoni, A. A., and Bossart, G. D. (2012b). “Sirenian pathology and mortality assessment,” in Sirenian Conservation: Issues and Strategies in Developing countries, eds E. Hines, J. Reynolds, L. Aragones, G. A. A. Mignucci-Giannoni, and M. Marmontel (Gainesville, FL: University Press of Florida), 148–156. doi: 10.2307/j.ctvx079z0.23
Bonde, R. K., O’Shea, T. J., and Beck, C. A. A. (1983). A manual of procedures for the salvage and necropsy of carcasses of the West Indian manatee (Trichechus manatus latirostris). Virginia: National Technical Information Service.
Borges, J., Jung, L., Carvalho, V., De Oliveira Moreira, A., Attademo, F., Ramos, R., et al. (2017). Pulmonicola cochleotrema (Digenea: Opisthotrematidae) in Antillean manatees (Trichechus manatus manatus) from the North-eastern region of Brazil. J. Mar. Biol. Assoc. 97, 1581–1584. doi: 10.1017/s0025315416000941
Bossart, G. D., Baden, D. G., Ewing, R. Y., Roberts, B., and Wright, S. D. (1998). Brevetoxicosis in manatees (Trichechus manatus latirostris) from the 1996 epizootic: gross, histologic, and immunohistochemical features. Toxicol. Pathol. 26, 276–282. doi: 10.1177/019262339802600214
Bossart, G. D., Ewing, R. Y., Lowe, M., Sweat, M., Decker, S. J., Walsh, C. J., et al. (2002a). Viral papillomatosis in Florida manatees (Trichechus manatus latirostris). Exp. Mol. Pathol. 72, 37–48. doi: 10.1006/exmp.2001.2407
Bossart, G. D., Meisner, R. A., Rommel, S. A., Ghim, S. J., and Jenson, A. B. (2002b). Pathological features of the Florida manatee cold stress syndrome. Aquat. Mamm. 29, 9–17. doi: 10.1578/016754203101024031
Brill, K., Marmontel, M., Bolen-Richardson, M., and Stewart, R. E. (2016). Inter-lab comparison of precision and recommended methods for age estimation of Florida manatee (Trichechus manatus latirostris) using growth layer groups in earbones. NAMMCO Sci. Publ. 10:3786. doi: 10.7557/3.3786
Castelblanco-Martínez, D. N., Bermúdez-Romero, A. L., Gómez-Camelo, I. V., Rosas, F. C. W., Trujillo, F., and Zerda-Ordoñez, E. (2009). Seasonality of habitat use, mortality and reproduction of the vulnerable Antillean manatee Trichechus manatus in the Orinoco River, Colombia: implications for conservation. Oryx 43, 235–242. doi: 10.1017/S0030605307000944
Castelblanco-Martínez, D. N., Galves, J., Ramos, E. A., Searle, L., Niño-Torres, C. A., Padilla-Saldívar, J. A., et al. (2018). “High levels of mortality threaten the Antillean manatee along the Caribbean coast of Belize and Mexico,” in Presentation at 18 Reunión de Trabajo y XII Congreso de la Sociedad Latinoamericana de Especialistas en Mamíferos Acuáticos (SOLAMAC), (Lima: SOLAMAC).
Castelblanco-Martínez, D. N., Nourisson, C., Quintana-Rizzo, E., Padilla-Saldivar, J., and Schmitter-Soto, J. J. (2012). Potential effects of human pressure and habitat fragmentation on population viability of the Antillean manatee Trichechus manatus: a predictive model. Endang. Species Res. 18, 129–145. doi: 10.3354/esr00439
CNAP (2013). Plan del Sistema Nacional de Áreas Protegidas 2014–2020. Havana: Ministerio de Ciencias Tecnología y Medio Ambiente.
Dawson, T. P., Jackson, S. T., House, J. I., Prentice, I. C., and Mace, G. M. (2011). Beyond predictions: biodiversity conservation in a changing climate. Science 332, 53–58. doi: 10.1126/science.1200303
Deutsch, C. J. (2008). “Trichechus manatus ssp. latirostris,” in The IUCN Red List of Threatened Species, (United Kingdom: IUCN).
Edwards, H. (2013). Potential impacts of climate change on warm water megafauna: the Florida manatee example (Trichechus manatus latirostris). Clim. Change 121, 727–738. doi: 10.1007/s10584-013-0921-2
Forneiro, Y., Azanza, J., Moncada, F., and Gerhartz-Muro, J. L. (2015). Programa de monitoreo de tortugas marinas. Guatemala City: Proyecto Archipiélagos del Sur CNAP, 99–107.
FWC (2021). Research/manatee/rescue-mortality-response/statistics/mortality/yearly/. Tallahassee: FWC.
Hostetler, J. A., Edwards, H., Martin, J., and Schueller, P. (2018). Updated Statewide Abundance Estimates for the Florida Manatee. Florida Fish and Wildlife Conservation Commission, Fish and Wildlife Research Institute Technical Report No. 23. Florida, FL: Florida Fish and Wildlife Conservation Commission.
IUCN (2012). IUCN Red List categories and criteria, version 3.1, 2nd Edn. Gland: IUCN Species Survival Commission.
Langtimm, C. A., Beck, C. A., Edwards, H. H., Fick-Child, K. J., Ackerman, B. B., Barton, S. L., et al. (2004). Survival estimates for Florida manatees from the photo-identification of individuals. Mar. Mamm. Sci. 20, 438–463. doi: 10.1111/j.1748-7692.2004.tb01171.x
Langtimm, C. A., O’Shea, T. J., Pradel, R., and Beck, C. A. (1998). Estimates of annual survival probabilities for adult Florida manatees (Trichechus manatus latirostris). Ecology 79, 981–997. doi: 10.2307/176594
Lefebvre, L. W., Marmontel, M., Reid, J. P., Rathbun, G. B., and Domning, D. P. (2001). “Status and biogeography of the West Indian manatee,” in Biogeography of the West Indies, 2nd Edn, eds C. A. Woods and F. E. Sergile (Boca Raton: CRC Press), 425–474. doi: 10.1201/9781420039481.ch22
Li, Y., Zhongweia, G., Qisena, Y., Yushana, W., and Niemeläb, J. (2003). The implications of poaching for giant panda conservation. Biol. Conserv. 111, 125–136. doi: 10.1016/S0006-3207(02)00255-0
Lightsey, J. D., Rommel, S. A., Costidis, A. M., and Pitchford, T. D. (2006). Gross necropsy diagnosis of watercraft−related mortality in the Florida manatee (Trichechus manatus latirostris). J. Zoo. Wildlife Med. 37, 262–275. doi: 10.1638/04-095.1
Lonati, G. L., Howell, A. R., Hostetler, J. A., Schueller, P., de Wit, M., Bassett, B. L., et al. (2019). Accuracy, precision, and error in age estimation of Florida manatees using growth layer groups in earbones. J. Mammal. 100, 1350–1363. doi: 10.1093/jmammal/gyz079
Marmontel, M. (1995). “Age and reproduction in female Florida manatees,” in Population biology of the Florida manatee, eds T. J. O’Shea, B. B. Ackerman, and H. F. Percival (Washington, DC: National biological service), 98–119.
Marmontel, M. (1993). “Age estimation and population biology of the Florida manatee Trichechus manatus latirostris,” PhD Dissertation. Gainesville (Fl): University of Florida.
Marmontel, M., Odell, D. K., and Reynolds, J. E. (1992). “Reproductive biology of South American manatees,” in Reproductive Biology of South American Vertebrates, ed. W. C. Hamlett (New York, NY: Springer), 295–312. doi: 10.1007/978-1-4612-2866-0_20
Marsh, H., O’Shea, T. J., and Reynolds, I. I. I. J. E. (2011). Ecology and conservation of Sirenia, dugongs and manatees. New York, NY: Cambridge University Press.
Martin, J., Edwards, H. H., Fonnesbeck, C. J., Koslovsky, S. M., Harmak, C. W., Dane, T. M., et al. (2015). Combining information for monitoring at large spatial scales: first statewide abundance estimates of the Florida manatee. Biol. Conserv. 186, 44–51. doi: 10.1016/j.biocon.2015.02.029
Meirelles, A. C. O. (2008). Mortality of the Antillean manatee, Trichechus manatus, in Ceará State, northeastern Brazil. J. Mar. Biol. Assoc. U K. 88, 1133–1137. doi: 10.1017/S0025315408000817
Mignucci-Giannoni, A. A., Montoya-Ospina, R. A., Jiménez-Marrero, N. M., Rodriguez-Lopez, M. A., Williams, E. H., and Bonde, R. K. (2000). Manatee mortality in Puerto Rico. Environ. Manage. 25, 189–198. doi: 10.1007/s002679910015
Montoya-Ospina, R. A., Caicedo-Herrera, D., Millán-Sanchez, S. L., Mignucci-Giannoni, A. A., and Lefebvre, L. W. (2001). Status and distribution of the West Indian manatee, Trichechus manatus, in Colombia. Biol. Conserv. 102, 117–129. doi: 10.1016/S0006-3207(00)00062-8
Parente, C. I., Vergara-Parente, J. E., and Lima, R. P. (2004). Strandings of Antillean manatees, Trichechus manatus, in northeastern Brazil. Lat. Am. J. Aquat. Mamm. 3, 69–75. doi: 10.5597/lajam00050
Perera-Valderrama, S., Hernández-Ávila, A., González-Méndez, J., Moreno-Martínez, O., Cobián-Rojas, D., Ferro-Azcona, H., et al. (2018). Marine protected areas in Cuba. Bull. Mar. Sci. 94, 423–442. doi: 10.5343/bms.2016.1129
Pimiento, C., Leprieur, F., Silvestro, D., Lefcheck, J. S., Albouy, C., Rasher, D. B., et al. (2020). Functional diversity of marine megafauna in the Anthropocene. Sci. Adv. 6:eaay7650. doi: 10.1126/sciadv.aay7650
Quintana-Rizzo, E., and Reynolds, J. E. I. (2010). “Regional management plan for the West Indian manatee(Trichechus manatus),” in Caribbean Environment Programme Technical Report No. 48, (Kingston: United Nations Environment Programme).
Ripple, W. J., Wolf, C., Newsome, T. M., Betts, M. G., Ceballos, G., Courchamp, F., et al. (2019). Are we eating the world’s megafauna to extinction? Conserv. Lett. 2019:e12627. doi: 10.1111/conl.12627
Self-Sullivan, C., and Mignucci-Giannoni, A. (2008). “Trichechus manatus ssp. manatus,” in The IUCN Red List of Threatened Species 2008, (United Kingdom: IUCN).
Self-Sullivan, C., and Mignucci-Giannoni, A. (2012). “West Indian Manatees (Trichechus manatus) in the Wider Caribbean Region,” in Sirenian Conservation: Issues and strategies in developing countries, eds E. Hines, J. Reynolds, L. Aragones, G. A. A. Mignucci-Giannoni, and M. Marmontel (Florida: University Press of Florida), 36–46. doi: 10.2307/j.ctvx079z0.10
Tringali, M. D., Davis, M. C., Bolen, E. E., Sullivan, J. G., and Haubold, E. M. (2008). Simultaneous use of the X and Y-chromosome genes SMCX, SMCY, and DBY for sex determination in the Florida manatee (Trichechus manatus latirostris). Mar. Mamm. Sci. 24, 218–224. doi: 10.1111/j.1748-7692.2007.00147.x
Keywords: threats, poaching, fishing, observation networks, conservation
Citation: Alvarez-Alemán A, García Alfonso E, Powell JA, Jacoby CA, Austin JD and Frazer TK (2021) Causes of Mortality for Endangered Antillean Manatees in Cuba. Front. Mar. Sci. 8:646021. doi: 10.3389/fmars.2021.646021
Received: 24 December 2020; Accepted: 12 May 2021;
Published: 08 June 2021.
Edited by:
Nuno Queiroz, Centro de Investigacao em Biodiversidade e Recursos Geneticos (CIBIO-InBIO), PortugalReviewed by:
Antonio Mignucci, InterAmerican University of Puerto Rico, Puerto RicoCopyright © 2021 Alvarez-Alemán, García Alfonso, Powell, Jacoby, Austin and Frazer. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Anmari Alvarez-Alemán, YWFsdmFyZXpAY21hcXVhcml1bS5vcmc=
†ORCID: Anmari Alvarez-Alemán, orcid.org/0000-0002-9240-6141; James D. Austin, orcid.org/0000-0003-0643-8620
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.