 Jhostin Ramos
Jhostin Ramos Marco Boto1
Marco Boto1 Juan Felipe Blanco-Libreros
Juan Felipe Blanco-Libreros José M. Riascos
José M. Riascos- 1Programa de Ecología de Zonas Costeras, Universidad de Antioquia – Sede Ciencias del Mar, Turbo, Colombia
- 2Instituto de Biología Universidad de Antioquia, Medellín, Colombia
- 3Corporación Académica Ambiental, Universidad de Antioquia – Sede Ciencias del Mar, Turbo, Colombia
- 4Corporación Centro de Excelencia en Ciencias Marinas – CEMarin, Bogotá, Colombia
Periwinkles (Littorinidae: genus Littoraria) are one of the very few molluscan clades showing an adaptive radiation closely associated to the mangrove habitat. However, pervasive land use changes associated to urbanization is prompting mangrove loss or degradation, with unknown consequences for mangrove-associated fauna. In the southern Colombian Caribbean, mangrove ecosystems have been encroached by human settlements and different populations of Littoraria angulifera (Lamarck, 1822) now inhabit anthropogenic intertidal substrates in urban areas, but the demographic traits of populations thriving in these novel environments are unknown. We studied the relative abundance and size structure of L. angulifera in remnant mangrove patches, woody debris and anthropogenic substrates (boulder seawalls and built structures) in 13 locations throughout the Urabá Gulf, a human-transformed tropical estuarine system. The abundance of L. angulifera was up to two orders of magnitude higher in anthropogenic than in quasi-natural or natural substrates. Snails also displayed a significant preference for wave protected positions in boulder seawalls and built structures exposed to heavy wave action, which was not previously reported in mangrove forests. Moreover, snail populations in anthropogenic substrate were consistently dominated by individuals of small sizes in comparison with mangroves or driftwood. We argue that the anthropogenic disturbances caused by the expansion of Turbo city during nearly one century in a coast formerly dominated by mangrove forests are providing novel and expanding habitats, whose quality might be good enough as to support high-density populations of L. angulifera. However, we hypothesize that shifted thermal regimes in hard and novel wave-exposed urban seascapes might also be prompting behavioral adjustments and the selection of smaller size ranges than those observed in mangrove forests.
Introduction
The establishment of modern cities and its subsequent expansion has long been identified as a major issue for the conservation of mangrove forests and their associated biodiversity (e.g., Hinrichsen, 1994; Ellison and Farnsworth, 1996; Lee et al., 2006; Branoff, 2017). As current trends predict that coastal tropical areas will experience some of the greatest urbanization rates over the next decades (Seto et al., 2012) this ubiquitous process is expected to have pervasive ecological consequences for mangrove ecosystems, similar to the consequences reported for urbanized estuaries and coasts worldwide (Alberti, 2015). In human-altered systems, and particularly in cities, most species are declining and being replaced by a small number of expanding species able to capitalize on the new conditions, which dictates the balance between “winners” and “losers” of human actions (McKinney and Lockwood, 1999; Alberti et al., 2020). From a functional point of view, specialists are more sensitive to ongoing human impacts and are being rapidly replaced by generalist species in diverse systems, prompting a global functional homogenization that could alter ecosystem functioning (Clavel et al., 2011; Dharmarajan et al., 2021; Filgueiras et al., 2021). This is critically important in ecosystems where functional redundancy is extremely low, as tropical mangrove forests (see Cannicci et al., 2021) because eliminating specialists may also imply loosing key ecosystem functions.
Periwinkles (Littorinidae: genus Littoraria) are one of the very few molluscan clades showing an adaptive radiation closely associated to the mangrove habitat; phylogenetic reconstructions of ancestral habitats suggests that periwinkles inhabited mangrove or wood substrates since its origin (Reid et al., 2010). The species Littoraria angulifera (Lamarck, 1822) is found in quiet brackish water inlets lined with thickets of Rhizophora mangle, crawling on trunks, prop roots, leaves and branches from the water line up to a few meters above the high tide mark (Gutierrez, 1988; Merkt and Ellison, 1998; Rodrigues et al., 2016). For closely associated mollusks, mangrove forests represent a refuge from strong wave action and extremes of temperature or evaporation, commonly observed in rocky and sandy environments (Sasekumar, 1974; Skilleter and Warren, 2000; Rodrigues et al., 2016). Accordingly, shell morphological variation of L. angulifera and other species in the genus has been related to local differences in forests conditions, including canopy height, food availability, average rainfall, and temperature (Merkt and Ellison, 1998; Tanaka and Maia, 2006; Riascos and Guzman, 2010; Silva et al., 2013). These ecological traits suggest that L. angulifera operates as a mangrove specialist, but an increasing number of studies show that it may actually occupy anthropogenic habitats (Gallagher and Reid, 1979; Costa et al., 2013; Zeidan et al., 2020) and display a high phenotypic plasticity (Janson, 1985; Reid and Mak, 1999); two traits typically observed in generalist species (Duarte et al., 1995).
The Urabá gulf (Colombian Caribbean) holds the southernmost mangrove forests of the Caribbean Sea (Figure 1). Currently, those forests are mainly located in the innermost southern region of the gulf, because cumulative alterations and land use changes have left mangrove forest destroyed or reduced to small patches in the outer-most part of the gulf. While studying the environmental drivers of the region-wide geographic distribution of L. angulifera in the southern Caribbean coast of Colombia, Ortiz and Blanco-Libreros (2012) found that despite the degradation of mangroves forests the species was mainly restricted to the external region of the gulf, reflecting its low tolerance to lower salinities prevailing in the internal, estuarine region. Interestingly, L. angulifera is also locally distributed in Turbo, a city port founded in 1840, now with∼125,000 inhabitants (Blanco-Libreros, 2016) located in the interior, estuarine zone of the gulf, which is characterized by relatively high salinity in surface waters and a severe environmental degradation. Therefore, in this heavily altered coast L. angulifera occupy small, patchily distributed mangrove areas, woody debris along the supra-littoral zone in sandy shores and hard anthropogenic structures.
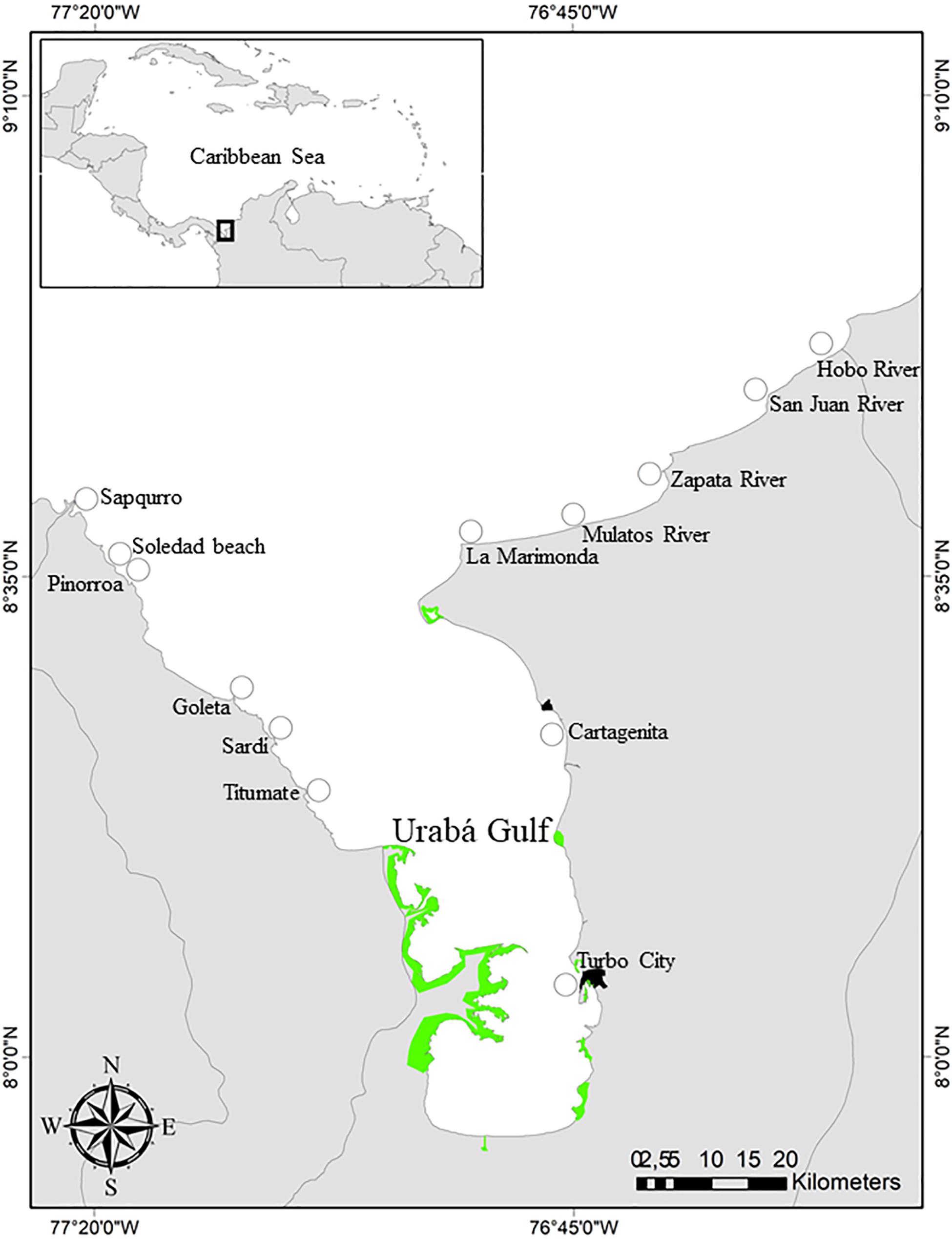
Figure 1. Map of the Urabá Gulf and the sampling sites in the Southern Caribbean of Colombia. The distribution of mangrove areas are shown in green.
In this context, we hypothesized that the severe transformations of the supra-littoral zone across the Urabá Gulf offer a spatial template where ecological pressures may alter behavior and population characteristics of L. angulifera. Specifically, we evaluated changes in abundance and size structure among populations thriving in distinct habitats. Moreover, we assessed behavioral patterns in the position of snails with respect to wave action in anthropogenic habitats.
Materials and Methods
Study Area
The study was performed in the Urabá Gulf (Figure 1). Located near the Colombia-Panama border, the gulf is a north-facing embayment that represents the southernmost region of the Caribbean Sea. The oceanographic setting and dynamics off the gulf are largely defined by the annual sediment and freshwater discharge of the Atrato river, considered the second largest river in the Southern Caribbean (Beier et al., 2017). As observed in major tropical river deltaic systems worldwide, mangrove forests are the most representative ecosystem in the gulf. These dense, monospecific stands are considered the most productive Rhizophora mangle-stands in the world (Riascos and Blanco-Libreros, 2019) and are seemingly different from other mangrove forests in the region in terms of species composition and dominance, forest structure, and ecosystem function (Urrego et al., 2014; Blanco-Libreros et al., 2015).
Field Work
Two samplings were performed for this study. In July 2009, 39 coastal localities (Figure 1) were visited to assess the distribution, relative abundance and length structure of L. angulifera in the Urabá Gulf (Ortiz and Blanco-Libreros, 2012). Where L. angulifera was present, the type of habitat was registered and the relative abundance and shell length were measured. The number of snails captured by two people during 20 min along a 50 m transect was taken as a measure of relative abundance in a given place and the shell length (i.e., the maximum distance between the apex and the anterior lip) for each snail was measured with a caliper. A second sampling was performed in November 2019 and February-March 2020, in 13 locations experiencing fast urban expansion and loss or reduction of mangrove cover (Figure 1). To assess demographic patterns, the relative abundance and the shell length were registered in natural (mangroves, woody debris) and anthropogenic substrates (boulder seawalls and concrete structures) in those places. Moreover, we evaluated behavioral preferences for exposed or protected positions with respect to wave action in the intertidal zone by comparing the number of snails in each position in woody debris and anthropogenic substrates. All the snails were released after measurements.
Data Analysis and Visualization
Differences in the relative abundance of L. angulifera among types of substrate were assessed with a Kruskal-Wallis ANOVA by ranks. Length–frequency distributions of L. angulifera collected in different habitats were plotted together to assess spatial patterns across the Urabá Gulf. To assess preferences for exposed/protected position to wave action, we used Mann-Whitney U tests to compare the number of snails in protected and exposed positions in woody debris, boulder seawalls and concrete structures.
Results
Distribution
Littoraria angulifera was spatially restricted to the north-western and north-eastern flanks and the outer margin of the Gulf (Figure 1). In those areas, L. angulifera was mainly observed in woody debris in beaches formerly dominated by mangroves and rarely in remaining mangrove patches near river mouths. In peri-urban areas of Turbo city, where mangrove forests have almost disappeared, L. angulifera was observed in two types of anthropogenic substrates: boulder seawalls protecting urban infrastructure and concrete structures.
Relative Abundance
Littoraria angulifera showed differences in relative abundances among substrates (H [4; N = 49] = 29,782; p < 0.001; Figure 2). Anthropogenic habitats showed higher (up to two orders of magnitude) and more variable relative abundances than those of natural habitats (Figure 2). Noteworthy, the lowest relative abundances of L. angulifera was observed in wild mangrove forests at Titumate.
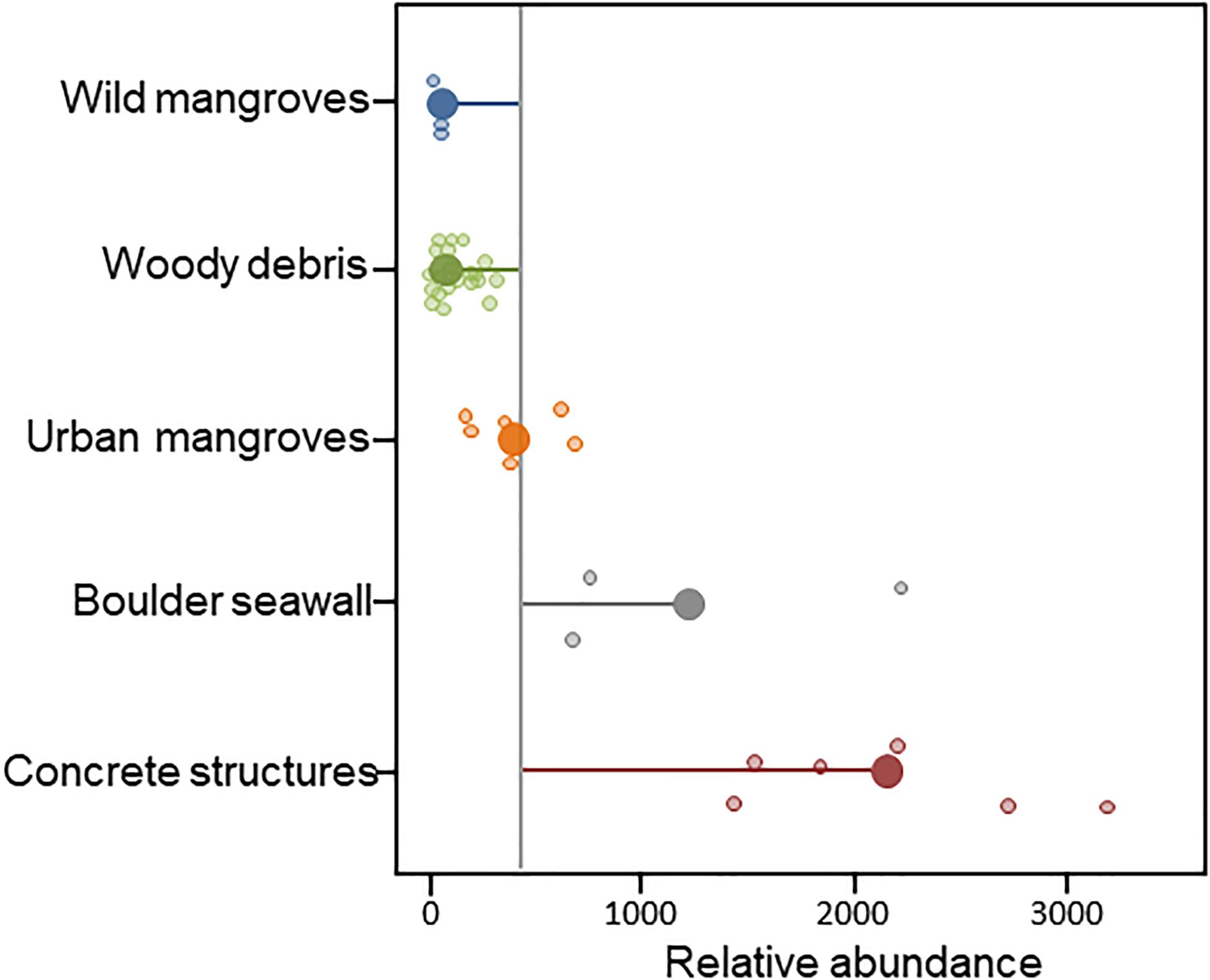
Figure 2. Jitter-pop plot of the relative abundance (number of snails per 20 m sampling) of Littoraria angulifera in different habitats at the Urabá Gulf estimated from data taken in 2009, 2019, and 2020. The plot was generated in R using the “ggplot2” library. Gray line: overall mean; lollypops: mean for each habitat and distance to the general mean.
Length-Frequency Distribution
The joy-plots of length-frequency distributions of L. angulifera populations revealed high spatial variability across both flanks of the Urabá Gulf (Figure 3). First, southern populations, particularly in the eastern flank, tend to be numerically dominated by small, presumably sexually immature snails, whereas northern populations tend to be dominated by larger, adult snails. Second, only populations located at the northernmost areas, at the entrance of the gulf, showed balanced numbers of adults and juveniles. There were no obvious distinctive patterns in size frequency distribution among habitats, perhaps reflecting the fact that mangrove trees and hard bottoms were observed in a few places, whereas woody debris were more commonly observed through the coast.
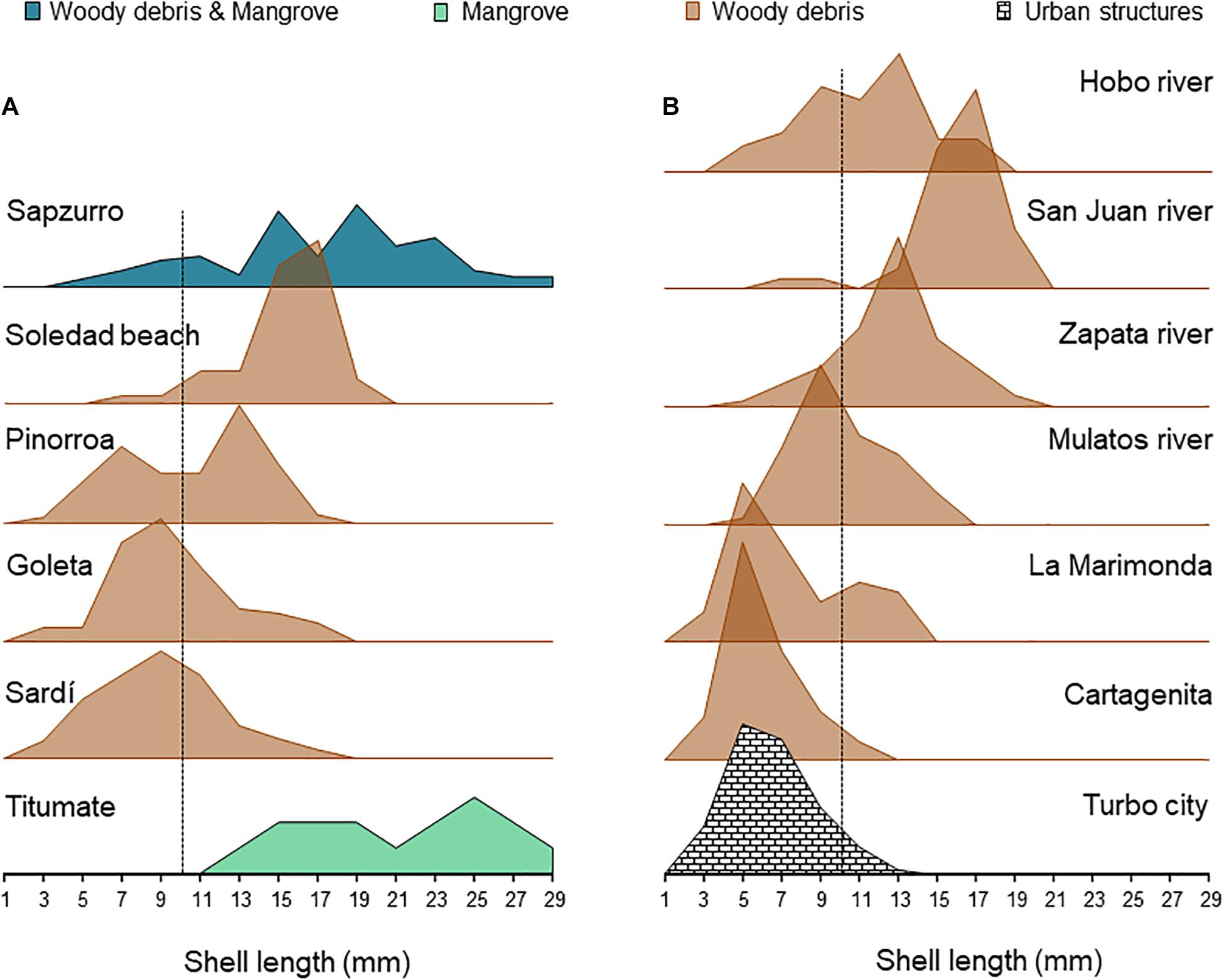
Figure 3. Joy-plot of the north-south changes in length-frequency distributions of Littoraria angulifera populations across the western (A) and eastern (B) flanks of the Urabá Gulf from data taken in 2009, 2019, and 2020. Y-axis represents the percentage of each shell length with respect to total abundance in each place; dotted lines represent the approximate mean size at sexual maturity of L. angulifera in Panamá (Gutierrez, 1988) and Brazil (Costa et al., 2013).
Position
The number of snails was significantly lower in exposed positions than in protected positions in woody debris, boulder seawalls, and concrete structures (Table 1 and Figure 4).
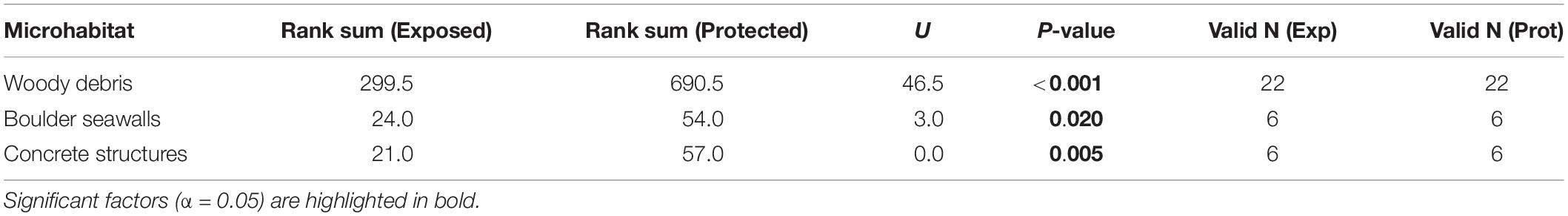
Table 1. Mann-Whitney U tests assessing differences in the number of Littoraria angulifera between exposed and protected positions in different microhabitats.
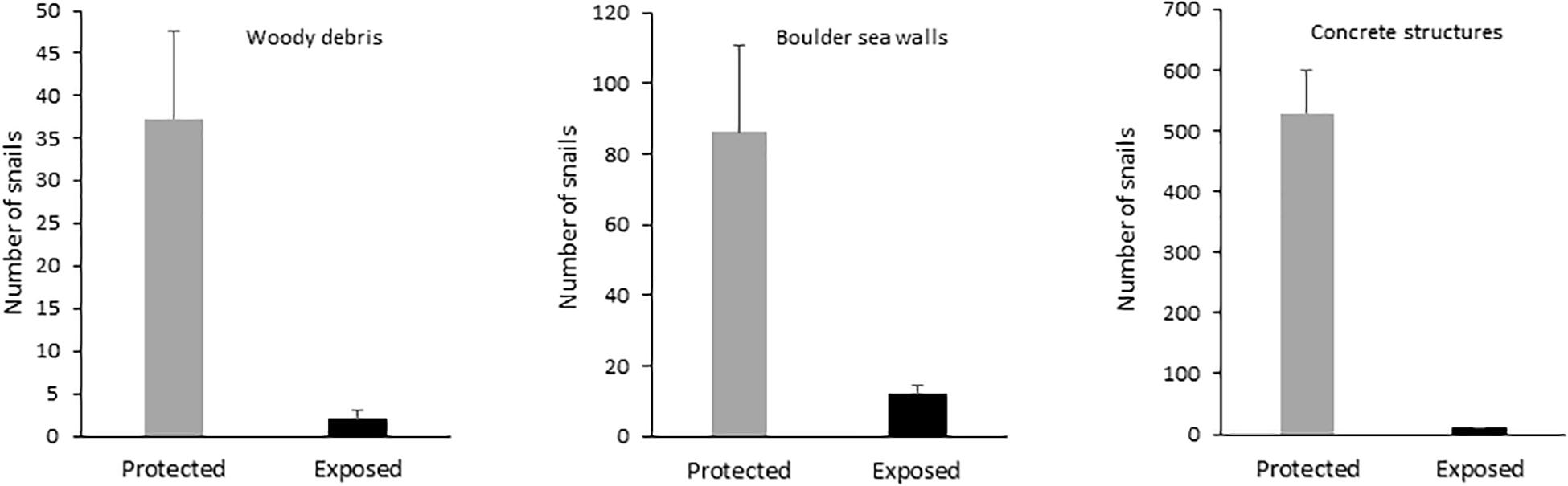
Figure 4. Comparison of the number of Littoraria angulifera in wave-protected and wave-exposed positions in woody debris, boulder seawalls and concrete structures in the Urabá Gulf, Antioquia, Colombia. Error bars are standard errors.
Discussion
The distribution of L. angulifera in the northern, external region of the Urabá Gulf and around Turbo city coincided with the most altered zone in the gulf. Owing to the strategic position of the gulf between Central and South America and the abundance of natural resources, this zone has seen growing agricultural land-use changes, urban expansion and a diverse array of legal and illicit activities for decades (Tenthoff, 2008; Lombana-Reyes, 2012) that resulted in a strong degradation or destruction of mangrove forests. The observed distribution of L. angulifera suggest that this species is able to thrive in alternative habitats which are not permanently influenced by the large discharge of the Atrato River and thus by less saline waters, which represents a geographic barrier for stenohaline marine species such as L. angulifera (Ortiz and Blanco-Libreros, 2012). Hence, L. angulifera in the Urabá Gulf can be considered a species thriving in harsh oceanographic and biotic conditions, with populations in peri-urban areas of Turbo city representing a marginal population.
Studies on the demography of marginal populations show that they commonly have low density relative to those thriving in core habitats: more individuals commonly disperse from the core to marginal habitats. Hence marginal populations often represent demographic sinks – a pattern observed at different spatial scales (reviewed by Kawecki, 2008). It is therefore a remarkable finding that the relative abundance of L. angulifera was one or two orders of magnitude higher in its southernmost, urbanized and osmotic-limiting boundary than in remnant mangrove patches or in woody debris at sandy beaches. Although the fossil evidence and a molecular phylogenetic reconstruction of ancestral habitats indicate that L. angulifera is an obligate mangrove dweller (Reid et al., 2010), several studies have reported this species thriving in hard natural or artificial substrates (e.g., Gallagher and Reid, 1979; Janson, 1985; Antwi and Ameyaw-Akumfi, 1987; Reid and Mak, 1999; Costa et al., 2013; Zeidan et al., 2020). Thus, this may be thought of as a sign of contemporary habitat diversification – likely reflecting on-going coastal urban sprawl – and the outstandingly high abundance of L. angulifera suggest that hard artificial substrates are not necessarily low-quality habitats when compared with core mangrove habitats outside the Urabá Gulf.
A striking demographic feature of the L. angulifera population inhabiting seawalls and concrete structures in Turbo was the rather small shell size range (1–15 mm) relative to the size range observed in mangroves or woody debris in other locations. As this species reach sexual maturity at ∼10 mm (Gutierrez, 1988; Costa et al., 2013), the population recorded in Turbo city seems almost exclusively composed of recruits and juveniles. Hence, one could reasonably argue that the lack of adults and the high abundance in this urban population may simply reflect a recent pulse of immigrants – allegedly a characteristic of marginal, transient populations that operate as demographic sinks (e.g., Kawecki, 2008). We could not rule out this possibility from our snapshot samplings, but L. angulifera – as most tropical mollusks do – display a continuous reproduction and hence populations are not commonly dominated by clearly defined cohorts (Urban and Riascos, 2001; Boehs and Freitas, 2021). Indeed, it was precisely the observed persistence of a population thriving in artificial substrates in Turbo and elsewhere through the distribution range (Gallagher and Reid, 1979; Costa et al., 2013; Zeidan et al., 2020) was what motivated this study. Hence, an alternative hypothesis to explain the smaller size range of L. angulifera in urban anthropogenic substrates would be that it represents an urban signature of phenotypic change, as reported for many various marine species elsewhere (Alberti, 2015; Alberti et al., 2020). First, a significantly smaller shell length has been here reported for L. angulifera populations inhabiting urban anthropogenic substrates as compared with populations in mangrove forests studied in the Brazilian states of Espírito Santo (Costa et al., 2013) and Bahía (Zeidan et al., 2020), suggesting that shifts to smaller body sizes are related to urbanization. Second, mangrove clearing has been empirically related to increased temperature in both water and sediment matrices (Granek and Ruttenberg, 2008; Kon et al., 2010) and temperature is higher in intertidal artificial structures than in natural adjacent habitats (Aguilera et al., 2019). Thus, L. angulifera may respond to warmer conditions associated urban expansion on mangrove forests with shrinking body size, which has been heralded as a “universal ecological response to global warming in aquatic systems” and the “temperature-size rule,” to which the great majority of ectotherms accommodates (Daufresne et al., 2009; Verberk et al., 2020). If a smaller urban phenotype has been selected as an outcome of this thermal-reaction norm, we should expect that the L. angulifera population in Turbo also reaches maturity at a smaller size and therefore the proportion of adults is actually higher than what the distribution of size frequencies suggests. In turn, the less dense populations inhabiting woody debris may reflect the unpredictable and transient nature of fallen trees as habitats in estuarine systems (Gonor et al., 1988) and therefore we hypothesize that these are low-quality habitats. Owing to its southern position and the patterns of water circulation within the Gulf, Ortiz and Blanco-Libreros (2012) suggested that the population of L. angulifera from Turbo may be reproductively isolated and not dependent on gene flow from populations in the Caribbean Sea. If this is correct, the urban population in Turbo may export larvae to nearby locations, thus explaining the observed prevalence of small snails in woody debris in neighbor areas (e.g., Cartagenita; Figure 1). In addition, during July 2016, consistent with high air temperature and high surface water salinity, many individuals of L. angulifera were observed in drift wood, fallen Rhizophora mangle trees but not on fringing mangroves to the south of Turbo, in an area where they have not been previously recorded (JF Blanco, personal observations). This suggests that the transient nature of populations in the SW part of the Gulf (Punta Coquito-Rio Guadualito afar from urbanized areas) is modulated by salinity regimes. However, more data and continuous observations are needed to assess these hypotheses.
Past and current efforts to understand and predict biological responses to on-going climate change have tended to focus on physiological changes, overlooking the importance of behavioral adaptations as traits that confer advantages in facing altered conditions. In line with the observed regulatory behaviors displayed by other littorinid snails to buffer changes in temperature, relative humidity, salinity and physical stress (Bingham, 1972; Garrity, 1984; Ng et al., 2017), our results strongly suggest that the observed positioning of L. angulifera in hard artificial structure reflects the need to cope with increased thermal conditions and perhaps also increased desiccation and physical stress caused by increased wave action.
The replacement of specialist for generalist species in human-modified environments result primarily from wholesale changes in the spectrum of available abiotic and biotic resources rather than from relaxed intra-guild competition among species (Filgueiras et al., 2021). Thus, we should ask about which resources are significantly changed as a consequence of urbanization and how periwinkles may take advantage of them. Apart from providing new, yet highly modified intertidal habitats, urbanization in estuarine systems alters nutrient flows and regimes (Lee et al., 2006; Lee, 2016). Taking into account that L. angulifera feeds on green algae, fungi and bacterial films closely associated to Rhizophora mangle prop roots (Kohlmeyer and Bebout, 1986), it is reasonable to assume that shifted nutrient regimes modify the amount of quality of food in anthropogenic substrates. Therefore, understanding the dietary shifts of periwinkles in urban habitats will be crucial to understand its role in urban ecosystem functioning and the goods and services they will provide.
Concluding Remarks
Our study undercovered a spatial pattern suggesting some demographic processes that characterize the persistence of L. angulifera in heavily altered coastal habitats from a tropical estuarine system. The remarkably high relative abundance and the behavioral changes displayed by snails in hard, human-built substrates seemingly reflect the fact that these novel and expanding environments offer opportunities probably related to increased food availability, but also impose harsh conditions. Moreover, among the ecological changes involved in the transition from mangrove forest to boulder seawalls or concrete structures shifts in thermal regimes seemingly play a key role, and we hypothesize that the small size range that characterize the populations thriving there may represent an urban phenotype that may confer advantages to cope these altered thermal conditions.
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Author Contributions
JRa, MB, and JRi conceived the project and performed the samplings. JRi conducted the statistical analyses. JB-L provided additional data. JRa, JB-L, and JRi wrote the manuscript. All authors revised the final draft of the current manuscript.
Funding
This research was received financial support by the Universidad de Antioquia – Corporación Académica Ambiental and Gobernación de Antioquia – project “Estuarine Expedition Urabá Gulf Phase 1,” within the larger Project “Expedición Antioquia 2013.”
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
Thanks to Vladimir Toro for organizing the logistics for the field trip that resulted in this work.
References
Aguilera, M. A., Arias, R. M., and Manzur, T. (2019). Mapping microhabitat thermal patterns in artificial breakwaters: alteration of intertidal biodiversity by higher rock temperature. Ecolo. Evol. 9, 12915–12927. doi: 10.1002/ece3.5776
Alberti, M. (2015). Eco-evolutionary dynamics in an urbanizing planet. Trends Ecol. Evol. 30, 114–126. doi: 10.1016/j.tree.2014.11.007
Alberti, M., Palkovacs, E. P., Roches, S. D., Meester, L. D., Brans, K. I., Govaert, L., et al. (2020). The complexity of urban eco-evolutionary dynamics. Bioscience 70, 772–793.
Antwi, D. A., and Ameyaw-Akumfi, C. (1987). Migrational orientation in two species of littoral gastropods (Littorina angulifera and Nerita senegalensis). Mar. Biol. 94, 259–263. doi: 10.1007/bf00392938
Beier, E., Bernal, G., Ruiz-Ochoa, M., and Barton, E. D. (2017). Freshwater exchanges and surface salinity in the Colombian basin. Caribbean Sea. PLoS One 12:e0182116. doi: 10.1371/journal.pone.0182116
Bingham, F. O. (1972). The mucus holdfast of Littorina irrorata and its relationship to relative humidity and salinity. Veliger 15, 48–50.
Blanco-Libreros, J. F. (2016). Global changes in Gulf of Urabá mangroves (Colombia): between the changing coastline and the expanding agricultural frontier. Actual. Biol. 38, 53–70.
Blanco-Libreros, J. F., Ortiz-Acevedo, L. F., and Urrego, L. E. (2015). Above-ground biomass and carbon reservoirs in Urabá Gulf mangroves (Colombian Caribbean). Actual. Biol. 37, 131–141. doi: 10.17533/udea.acbi.v37n103a02
Boehs, G., and Freitas, L. A. (2021). Population attributes of Littoraria angulifera (Gastropoda: Littorinidae) in mangroves in Bahia State, north-eastern Brazil. Braz. J. Biol. 82:e243114.
Branoff, B. L. (2017). Quantifying the influence of urban land use on mangrove biology and ecology: a meta analysis. Global Ecol. Biogeogr. 26, 1339–1356. doi: 10.1111/geb.12638
Cannicci, S., Lee, S. Y., Bravo, H., Cantera-Kintz, J. R., Dahdouh-Guebas, F., Fratini, S., et al. (2021). A functional analysis reveals extremely low redundancy in global mangrove invertebrate fauna. Proc. Natl. Acad. Sci. 118:e2016913118. doi: 10.1073/pnas.2016913118
Clavel, J., Julliard, R., and Devictor, V. (2011). Worldwide decline of specialist species: toward a global functional homogenization? Front. Ecol. Environ. 9, 222–228.
Costa, M. B., Zamprogno, G. C., Pedruzzi, F. C., Dalbem, G. B., and Tognella, M. M. P. (2013). Assessing the continuous impact of tributyltin from antifouling paints in a Brazilian mangrove area using intersex in Littoraria angulifera (Lamarck, 1822) as biomarker. Int. J. Oceanogr. 2013:769415. doi: 10.1155/2013/769415
Daufresne, M., Lengfellner, K., and Sommer, U. (2009). Global warming benefits the small in aquatic ecosystems. Proc. Nat. Acad. Sci. U. S. A. 106, 12788–12793. doi: 10.1073/pnas.0902080106
Dharmarajan, G., Gupta, P., Vishnudas, C. K., and Robin, V. V. (2021). Anthropogenic disturbance favours generalist over specialist parasites in bird communities: implications for risk of disease emergence. Ecol. Lett. 24, 1859–1868. doi: 10.1111/ele.13818
Duarte, C. M., Nielsen, S. L., Enriquez, S., and Agusti, S. 1995. Comparative functional-plant ecology – rationale and potentials. Trends Ecol. Evol. 10, 418–421.
Ellison, A. M., and Farnsworth, E. J. (1996). Anthropogenic disturbance of Caribbean mangrove ecosystems: past impacts, present trends, and future predictions. Biotropica 28, 549–565. doi: 10.2307/2389096
Filgueiras, B. K., Peres, C. A., Melo, F. P., Leal, I. R., and Tabarelli, M. (2021). Winner–Loser Species Replacements in Human-Modified Landscapes. Trends Ecol. Evol. 36, 545–555. doi: 10.1016/j.tree.2021.02.006
Gallagher, S. B., and Reid, G. K. (1979). Population dynamics and zonation in the periwinkle snail, Littorina angulifera, of the Tampa Bay, Florida region. Nautilus 94, 162–178.
Garrity, S. D. (1984). Some adaptations of gastropods to physical stress on a tropical rocky shore. Ecology 65, 559–574. doi: 10.2307/1941418
Gonor, J. J., Sedell, J. R., and Benner, P. A. (1988). “What we know about large trees in estuaries, in the sea, and on coastal beaches” in From the Forest to the Sea, a story of Fallen Trees. eds C. Maser, R. F. Tarrant, J. M. Trappe, and J. F. Franklin (United States: USDA). 83–112USDA For. Serv. Gen. Tech. Rep. GTR-PNW-229, Pacific Northwest Res. Sta., Portland, OR.
Granek, E., and Ruttenberg, B. I. (2008). Changes in biotic and abiotic processes following mangrove clearing. Estuar. Coast. Shelf. Sci. 80, 555–562. doi: 10.1016/j.ecss.2008.09.012
Gutierrez, P. C. (1988). The ecology and behavior of the mangrove periwinkle. Littorina angulifera. Biotropica 20, 352–356. doi: 10.2307/2388333
Janson, K. (1985). Genetic and morphologic variation within and between populations of Littorina angulifera from Florida. Ophelia 24, 125–134. doi: 10.1080/00785236.1985.10426625
Kawecki, T. J. (2008). Adaptation to marginal habitats. Ann. Rev. Ecol. Evol. Syst. 39, 321–342. doi: 10.1146/annurev.ecolsys.38.091206.095622
Kohlmeyer, J., and Bebout, B. (1986). On the occurrence of marine fungi in the diet of Littorina angulifera and observations on the behavior of the periwinkle. Mar. Ecol. 7, 333–343. doi: 10.1111/j.1439-0485.1986.tb00168.x
Kon, K., Kurokura, H., and Tongnunui, P. (2010). Effects of the physical structure of mangrove vegetation on a benthic faunal community. J. Exp. Mar. Biol. Ecol. 383, 171–180. doi: 10.1016/j.jembe.2009.11.015
Lee, S. Y. (2016). From blue to black: anthropogenic forcing of carbon and nitrogen influx to mangrove-lined estuaries in the South China Sea. Mar. Poll. Bull. 109, 682–690. doi: 10.1016/j.marpolbul.2016.01.008
Lee, S. Y., Dunn, R. J. K., Young, R. A., Connolly, R. M., Dale, P. E. R., Dehayr, R., et al. (2006). Impact of urbanization on coastal wetland structure and function. Austral Ecol. 31, 149–163. doi: 10.1111/j.1442-9993.2006.01581.x
Lombana-Reyes, M. (2012). La configuración espacial de Urabá en cinco décadas. Cienc. Polít. 13, 40–79.
McKinney, M. L., and Lockwood, J. L. (1999). Biotic homogenization: a few winners replacing many losers in the next mass extinction. Trends Ecol. Evol. 14, 450–453. doi: 10.1016/s0169-5347(99)01679-1
Merkt, R. E., and Ellison, A. M. (1998). Geographic and habitatspecific morphological variation of Littoraria (Littorinopsis) angulifera (Lamarck, 1822). Malacologia 40, 279–295.
Ng, T. P., Lau, S. L., Seuront, L., Davies, M. S., Stafford, R., Marshall, D. J., et al. (2017). Linking behaviour and climate change in intertidal ectotherms: insights from littorinid snails. J. Exp. Mar. Biol. Ecol. 492, 121–131. doi: 10.1016/j.jembe.2017.01.023
Ortiz, L. F., and Blanco-Libreros, J. F. (2012). Distribución de los gasterópodos del manglar, Neritina virginea (Neritidae) y Littoraria angulifera (Littorinidae) en la Ecorregión Darién, Caribe colombiano. Rev. Biol. Trop. 60, 219–232.
Reid, D. G., Dyal, P., and Williams, S. T. (2010). Global diversification of mangrove fauna: a molecular phylogeny of Littoraria (Gastropoda: littorinidae). Mol. Phylogenetics Evol. 55, 185–201. doi: 10.1016/j.ympev.2009.09.036
Reid, D. G., and Mak, Y. M. (1999). Indirect evidence for ecophenotypic plasticity in radular dentition of Littoraria species (Gastropoda: littorinidae). J. Moll. Stud. 65, 355–370. doi: 10.1093/mollus/65.3.355
Riascos, J. M., and Blanco-Libreros, J. F. (2019). Pervasively high mangrove productivity in a major tropical delta throughout an ENSO cycle (Southern Caribbean. Colombia). Estuar. Coast. Shelf Sci. 227:106301. doi: 10.1016/j.ecss.2019.106301
Riascos, J. M., and Guzman, P. A. (2010). The ecological significance of growth rate, sexual dimorphism and size at maturity of Littoraria zebra and L. variegata (Gastropoda: littorinidae). J. Moll. Stud. 76, 289–295. doi: 10.1093/mollus/eyq011
Rodrigues, C. A., Ribeiro, R. P., Santos, N. B., and Almeida, Z. S. (2016). Patterns of mollusc distribution in mangroves from the São Marcos Bay, coast of Maranhão State, Brazil. Acta Amazon. 46, 391–400. doi: 10.1590/1809-4392201600493
Sasekumar, A. (1974). Distribution of macrofauna on a Malayan mangrove shore. J. Anim. Ecol. 43, 51–69. doi: 10.2307/3157
Seto, K. C., Güneralp, B., and Hutyra, L. R. (2012). Global forecasts of urban expansion to 2030 and direct impacts on biodiversity and carbon pools. Proc. Nat. Acad. Sci. U. S. A. 109, 16083–16088. doi: 10.1073/pnas.1211658109
Silva, S. E., Silva, I. C., Madeira, C., Sallema, R., Paulo, O. S., and Paula, J. (2013). Genetic and morphological variation in two littorinid gastropods: evidence for recent population expansions along the East African coast. Biol. J. Linn. Soc. 108, 494–508. doi: 10.1111/j.1095-8312.2012.02041.x
Skilleter, G. A., and Warren, S. (2000). Effects of habitat modification in mangroves on the structure of mollusc and crab assemblages. J. Exp. Mar. Biol. Ecol. 244, 107–129. doi: 10.1016/s0022-0981(99)00133-1
Tanaka, M. O., and Maia, R. C. (2006). Shell morphological variation of Littoraria angulifera among and within mangroves in NE Brazil. Hydrobiologia 559, 193–202. doi: 10.1007/s10750-005-1449-x
Tenthoff, M. (2008). El Urabá: donde el Desarrollo Alternativo se confunde con intereses económicos y la reinserción del paramilitarismo. Amsterdam: Transnational Institute.
Urban, H. J., and Riascos, J. M. (2001). Estimating gonado-somatic índices in bivalves with fused gonads. J. Shellfish Res. 21, 249–253.
Urrego, L. E., Molina, E. C., and Suárez, J. A. (2014). Environmental and anthropogenic influences on the distribution, structure, and floristic composition of mangrove forests of the Gulf of Urabá (Colombian Caribbean). Aquat. Bot. 114, 42–49. doi: 10.1016/j.aquabot.2013.12.006
Verberk, W. C., Atkinson, D., Hoefnagel, K. N., Hirst, A. G., Horne, C. R., and Siepel, H. (2020). Shrinking body sizes in response to warming: explanations for the temperature–size rule with special emphasis on the role of oxygen. Biol. Rev. 96, 247–268. doi: 10.1111/brv.12653
Keywords: marine urban sprawl, urban expansion, Eco-evo dynamics, mangrove ecology, human-dominated ecosystem, population structure
Citation: Ramos J, Boto M, Blanco-Libreros JF and Riascos JM (2021) The Mangrove Periwinkle Littoraria angulifera (Mollusca: Littorinidae) in the Urabá Gulf (Colombian Caribbean): Finding Ways in an Urbanizing Coast? Front. Mar. Sci. 8:641567. doi: 10.3389/fmars.2021.641567
Received: 14 December 2020; Accepted: 26 August 2021;
Published: 16 September 2021.
Edited by:
Nuria Marba, Consejo Superior de Investigaciones Científicas, SpainReviewed by:
Luis Ernesto Arruda Bezerra, Instituto de Ciências do Mar, BrazilChristopher Henderson, University of the Sunshine Coast, Australia
Copyright © 2021 Ramos, Boto, Blanco-Libreros and Riascos. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: José M. Riascos, am9zZS5yaWFzY29zQHVkZWEuZWR1LmNv