 Les Watling
Les Watling- 1School of Life Sciences, University of Hawai’i at Mānoa, Honolulu, HI, United States
- 2Mystic Aquarium—Sea Research Foundation, Mystic, CT, United States
- 3Department of Marine Sciences, University of Connecticut, Groton, CT, United States
The concept of “vulnerable marine ecosystem” (VME) was included in United Nations (UN) General Assembly Resolution 61/105 as part of an international effort to minimize the effects of seafloor fisheries on the high seas. However, defining a VME was left to be developed by technical guidance to the UN Food and Agricultural Organization. In that venue certain categories of organisms were deemed to be indicator species, suggesting that areas with those species would be considered VMEs with subsequent management measures implemented to conserve those ecosystem attributes. We note that on seamounts VME indicator species can be distributed widely, in dense clusters or sparsely. A dense cluster, for example, of scleractinian corals or sponges, is most often referred to as a VME, but we argue that any such dense cluster is not an ecosystem, rather it is a community, likely one of many that make up the ecosystem. Other communities on the seamount that are not part of that dense cluster could include many small species (some yet to be discovered) who are also part of the ecosystem because they are part of the web of interactions and flow of materials/energy on the seamount. We also suggest that a seamount ecosystem might extend over several or many seamounts in a biogeographic area. We conclude that the term “ecosystem” in the VME concept outlined by the United Nations needs to be re-evaluated from a classical ecological perspective leading to spatial management approaches that better address ecologically relevant space and time scales.
Introduction
Seamount habitats harbor diverse assemblages of suspension-feeding organisms, usually attached to the hard substrate, and often extending a meter or more into the overlying water. Many, if not most, of these organisms have life history characteristics that identify them as indicators of vulnerable marine ecosystems (VMEs, but we will use VMEcosystems, to distinguish between the ecosystem and the indicator species usage of the acronym VME). In general, the VME indicator species are long-lived, with ages extending from many decades to several centuries and longer; they are fragile, having become adapted to places where water flow, even when enhanced, generally does not exceed a few centimeters per second; their reproductive habits, when known, suggest infrequent or aperiodic spawning, often on scales of years between spawning events, the exception perhaps being in areas with seasonal and dense phytoplankton blooms with subsequent transport to the seafloor; and their larvae are mostly unknown, as is the distance over which they are capable of dispersing [Food and Agriculture Organization of the United Nations (FAO), 2009].
The problem with the identification of VMEcosystems and deciding how to manage them is that there is no agreement on delimitation of the ecosystem. Noteworthy is that the original UNGA resolution 61/105 included the phrase “vulnerable marine ecosystem” as critical terminology (paragraphs 76–95) so the Expert Consultation tasked to develop the initial draft of the guidelines needed to start with this as the foundation [Food and Agriculture Organization of the United Nations (FAO), 2009]. The panel considered how to address the ecosystem-scale scope of the term but reduced the level of complexity for implementation to the scales of populations, communities, and habitats (paragraphs 14–16 in the guidelines) which are the units that would experience any alterations. Indeed, the guidelines that describe and define VMEcosystems acknowledge that Regional Fisheries Management Organizations (RFMOs) and their member states would want some flexibility in implementation.
Determining whether a vulnerable marine ecosystem is present is predicated on finding pre-determined indicator species (UNGA Resolution 64/72). The characteristics of indicator species were determined to have a particular set of attributes that make them especially vulnerable to the impacts of bottom contact fishing gear [Food and Agriculture Organization of the United Nations (FAO), 2009]: (1) uniqueness or rarity of the species; (2) functionally significant to the habitat; (3) body fragility; (4) life-history characteristics that make probability of recolonization after impact low or unpredictable; and (5) species that serve as habitat for other species through their structural complexity. On seamounts much attention has been given to species of corals and sponges since they generally fit at least one, and often several, of the criteria to be considered as VME indicator species (Ardron et al., 2014).
The concept of vulnerability is a critical element for implementing the guidelines. Paragraph 14 of UNGA Resolution 62/72 states (italics ours for emphasis): “[v]ulnerability is related to the likelihood that a population, community, or habitat will experience substantial alteration from short-term or chronic disturbance, and the likelihood that it would recover and in what time frame. These are, in turn, related to the characteristics of the ecosystems themselves, especially biological and structural aspects. VME features may be physically or functionally fragile. The most vulnerable ecosystems are those that are both easily disturbed and very slow to recover, or may never recover.” Therefore, the concept of a VME is linked to the ecosystem in which populations, communities and habitats are nested and interact at a functional level.
In much of the subsequent literature and discussion on the issue, as well as the application of the guidelines, the concept of indicator species has been conflated with the ecosystem itself so that when some authors speak of VMEs they are simultaneously speaking of the presence of indicator species but also, by inference, evaluating the presence of a vulnerable ecosystem. The problem, of course, is that we see many examples where the occasional presence (i.e., sparse distribution) of indicator species is interpreted to mean that we are not in a VMEcosystem. However, it is important to remember that these are indicator species and are meant to represent all the other species not considered or sampled in the ecosystem.
Distribution of Indicator Species, Communities, and Ecosystems on Seamounts
Some seamounts, especially at high latitudes (but see Baco et al., 2017 for an example of a low latitude example) harbor extensive thickets or reefs created by colonies of hard corals. In the Southwest Pacific, the coral Solenosmilia variabilis, is the primary species of the coral thickets (Thresher et al., 2011). Elsewhere, Madrepora or Desmophyllum can fill the same role. The importance of these thickets is that they form habitat for an unknown number of other species, most of which have not been documented outside of the NE Atlantic. Images from the Tasmanian seamounts show most octocorals growing on the dead skeletons of the Solenosmilia colonies. In addition, large sea urchins and other invertebrates, including bryozoans and hydroids were utilizing this habitat (Thresher et al., 2011). When most people think of VMEcosystems, reef and thicket habitats are what come to mind. But seamounts, especially along ridges created by the erosion and slumping of the seamount flanks, can have high densities of bush-like octocorals with an understory of smaller species, including sponges, most if not all fragile and functionally significant as habitat (e.g., for commensal species). Diversity in these “coral gardens” or “forests” is probably quite high, but they have not yet been adequately sampled to better quantify this relationship.
In our opinion and extensive observation, most people, when referring to VMEcosystems are in fact really talking about VMCommunities (VMCs). Indeed it has been fundamental in ecological science that ecosystems are composed of many habitat types that support communities of several kinds (see Allee et al., 1949, for example). In the South Pacific, Rowden et al. (2017) mapped in detail the distribution of the hard coral, S. variabilis, over several seamounts. They found the distributions of brisingid starfish and crinoids to closely parallel that of the hard coral. They refer to this as the Solenosmilia VME, but in fact, it is the Solenosmilia community, one of many communities on those seamounts, the others generally not discussed in detail (but see Clark et al., 2015), that are made up of gorgonians, black corals, sponges, and small organisms living in the sand. With the possible exception of the sand, one could think of the coral groups and sponges as characterizing a suspension-feeder community, partly separate from, but perhaps also overlapping with the Solenosmilia community. The sand community comprises worms, nematodes, harpacticoid copepods, and others., but are poorly studied. The sand communities are connected through water flow, larval distribution, and the flux of materials from proximate coral and suspension-feeding communities (e.g., Bourque and Demopoulos, 2018; Pierrejean et al., 2020), thus justifying the concept of the seamount as a series of functionally interrelated and spatially overlapping communities making up part of an ecosystem which potentially comprises multiple seamounts.
Many studies, from Remotely Operated Vehicle (ROV) dives to towed camera transects, have demonstrated that VME indicator species are not homogeneously distributed across seamount substrates (e.g., Victorero et al., 2018). Seamounts are complex geomorphological structures composed largely of basalt rock in the form of ridges and flat plains, but also as vertical cliffs with all angles of slopes in between. Depending on the geological history of the seamount, there may also be areas of compressed ash, or breccias consisting of stones and boulders of various sizes, and biogenic sand. All these factors contribute to seamounts possessing a range of potential habitats (Figure 1) in which vulnerable species can live (Auster et al., 2005). In addition, seamount sandy habitats harbor unique small species, some of which have so far been found nowhere else. A good example are the 54 new species of the sediment-dwelling harpacticoid copepods found on the flat summit of Great Meteor Seamount in the Northeast Atlantic (George and Schminke, 2002). Dredge samples from the summits of the Meteor and Lusitanian seamount groups, which are separated by about 1,000 km, also produced 30 new species of small gastropods in the family Rissoidae (Gofas, 2007).

Figure 1. Typical communities commonly seen on Pacific seamounts that exhibit a diversity of species in different settings with no single indicator species that would characterize these places. (A) A mixed coral, coral rubble, and sand habitat at 803 m off Jarvis Island with roaming Bathycongrus. (B) Sparse corals and Neocyttus cf. acanthorhynchus along the margin of basaltic ridge and sand habitats, 1,140 m, Titov Seamount. (C) A mixed coral community on the flank of Swordfish Seamount, 970 m. (D) Diplacanthopoma sp. among octocoral branches in a mixed basalt and sand habitat. All images courtesy of NOAA Okeanos Program and CAPSTONE Science Team, fish identifications from annotations during the dives by B. Mundy, NOAA Fisheries.
The ICES Working Group on Deep-Sea Ecology (WGDEC) has been asked, over multiple years, to refine the operational definition of VMEcosystems and the associated use of indicator species in the decision process for spatial management. This effort has led to a series of reports and articles that address indicator species distribution, abundance, vulnerability, and uncertainties regarding indication of a VMEcosystem, in an effort to clarify recommended areas for protection in the North Atlantic where bottom trawl fishing is occurring (Morato et al., 2018, 2021). But, while numerical approaches are being used to identify vulnerable components of VMEcosystems, they do not provide insights into the functioning of vulnerable marine ecosystems. That is, the understanding and delimitation of the ecosystem also requires a “process-functional” approach which accounts for energy flow. Thus, “… we see the ecosystem as a dual entity. In one dimension it is structured according to constraints involving organism interaction and natural selection. In another dimension it is structured according to constraints that involve mass balance and thermodynamics” (O’Neill et al., 1986: 209). In the sea, which is a relatively open system, ecosystems are typically large, boundaries are created by water masses and fronts, thus encompassing several hundred thousand square kilometers measured at the ocean surface (Sherman and Duda, 1999). At depth, we do not yet know the extent of the ecosystem, although there are proposals for province boundaries (Watling et al., 2013).
Does the VMEcosystem cover only a single seamount or are there multiple seamounts in a VMEcosystem? The communities that make up an ecosystem can range over very large distances. Common examples are the communities of sandy beaches or rocky shores along continental coastlines, both of which are commonly thought of as ecosystems. Each contains their own common set of species, and each might not be contiguous throughout the range, but each also might have in common the dispersal of individuals from one part of the ecosystem to another, through the movements of either the larval or adult stages of the populations. The same can be said for seamounts within a biological province. A community dominated by several species of bamboo corals or the scleractinian Solenosmilia might extend along the whole or part of a seamount chain, or range over seamounts located in large sections of the ocean basin. Those distributions and estimates of the genetic connectivity or how much genetic variation exists over the whole of the range of a species is just becoming known (e.g., Thoma et al., 2009; Radice et al., 2016). Some information is available for species, such as brittle stars and chirostylid galatheid crabs associated with the corals, rather than the corals themselves (Samadi et al., 2006; Cho and Shank, 2010).
Determining the extent of VMEcosystems begins with mapping the distribution of biogeographic provinces in the ocean, which in turn uses the distributions of species that make up the communities that have so far been investigated. While deep-sea biogeography has been an area of study since the time of Ekman (Watling et al., 2013), it was only the recent funding for deep-diving ROVs and funding for ecological exploration of continental margins and deep sea regions that has allowed scientists to begin to map the bathyal benthos of the ocean away from the continental shelves (e.g., Kennedy et al., 2019; Lapointe et al., 2020). Much of the data thus procured has been archived in Ocean Biogeographic Information System (OBIS) and is available to anyone who wants to use it (e.g., Costello et al., 2017). One method of differentiating seamounts or seamount groups as VMEcosystems, is to determine which of the many seamounts in the ocean are similar to each other in some characteristic way. Clark et al. (2011) developed a classification system using several physical and biological features including summit depth, organic matter flux, distance to nearest seamount, and dissolved oxygen in the surrounding water. This method divided the estimated global 10,604 large seamounts (estimate from Kitchingman and Lai, 2004) with summit depths in the bathyal (<3,500 m) into 194 groups distributed over the 14 bathyal provinces of Watling et al. (2013). In the biogeographic province in which the Louisville seamounts are located (BY6), for example, there were 517 seamounts divided into 17 groups, of which four occurred at various points along the Louisville chain.
It is very rare that more than two or three ROV dives are conducted on any one seamount, so variation in community composition based on depth (water mass characteristics), orientation to principle current regime, and effects of species interactions are generally not well-known. Seamounts with multiple dives on the summit and flanks include Kelvin (Lapointe et al., 2020) and Anton Dohrn (Davies et al., 2015) in the North Atlantic, Davidson (McClain et al., 2010) and Cobb (Du Preez et al., 2016) in the Northeast Pacific, Necker Ridge (Morgan et al., 2015) in the Central Pacific, and the Graveyard seamounts NE of New Zealand (Clark and O’Driscoll, 2003). Other ROV programs, such as those of the Okeanos Explorer and Nautilus, are exploratory and rarely do more than two dives on any seamount (Kennedy et al., 2019). As a result we have a more or less detailed look at the distribution of organisms on only a few seamounts and so cannot say much about the distribution of communities on any one seamount. In the SW Pacific, a TowCam system was used to cover at least the summit area of a number of seamounts. In most cases those seamounts had relatively small summit areas so the TowCam transects were arranged to produce coverage of a significant proportion of the summit and approaching flanks. In other cases, such as the Louisville Seamounts, most of the transect coverage was on the summits (Clark et al., 2015).
In order to determine the boundary of an ecosystem it is important to know the limits of the component communities, both horizontally and vertically. Efforts to plot or model the distribution of the most abundant indicator species may not account for the whole ecosystem or spatially distinct but nonetheless vulnerable communities within the seamount landscape (e.g., Rowden et al., 2017). As well, we know that fishing at a selective depth can have impacts far deeper than the target fishing depth due to the vertical movements and other density dependent responses of fish species (Bailey et al., 2009; Priede et al., 2011). For several seamounts in the Louisville Seamount chain, various models were applied to towed camera image data to determine that three VME indicator species, S. variabilis coral heads and matrix, brisingid starfish, and crinoids, were located near the summit edges, and that other parts therefore could be trawled due to the absence or low abundance of those species. Rowden et al. (2017) refer to the S. variabilis matrix as if it is a VMEcosystem on its own, and so, by presumption, the remainder of the summit of the seamount does not belong to that ecosystem and is therefore not vulnerable. We have already suggested that Rowden et al. (2017) were actually determining the extent of the Solenosmilia community, which is only a part of the larger ecosystem. Whole seamounts need to be treated as VMEcosystems (Watling and Auster, 2017), or more likely, as part of a larger ecosystem.
Conclusion and Recommendations
We find the term “ecosystem” as currently applied to VMEs to be confused in use and application since its roots in the ecosystem concept are broader and more holistic. This confusion has led to use of VME indicator species being conflated to indicate whether a VMEcosystem is present or not, often ignoring the connections of such species to the communities of organisms in which they reside. Most importantly, this interpretation leads to gaps in meeting overall conservation goals. Seamount communities are not well known due to minimal or a wholesale lack of survey effort on individual seamounts. Focusing on single easily detected species (i.e., captured in fishing gear as bycatch or in image surveys) does not equate to the extent of the interactions defining the ecosystem or the connectivity of the vulnerable seafloor community. Protecting only part of a seamount does not necessarily protect the integrity of the seamount communities, making the whole community vulnerable. Furthermore, the seamount ecosystem most likely will extend over multiple seamounts, the extent to be determined by geography and hydrography. “An “ecosystem plan” that fails to protect all of the species associated with a particular natural community can only be characterized as deficient” (Wilcove, 1994: 327). This statement was written as part of the legal argument for saving the whole of the old growth forest of the northwestern United States as habitat for the northern spotted owl. Wilcove showed that a management plan targeted at a single species was unlikely to protect the full diversity of interacting and vulnerable species in the ecosystem and thus the spotted owl, itself.
Identifying the presence, distribution, and abundance of an indicator species defines the state of that species at a moment (or period) in time. It does not define the composition of an associated community, the suite of species interactions that define and sustain the community, or the flows of materials and energy that define the bounds of the ecosystem. Most important, it is those details about species interactions (including population connectivity, energy flow that mediates growth and reproduction, and interactions mediated by the local oceanographic regime) that will be needed to understand and predict the extent to which fishing and other human activities produce significant adverse impacts. Significant adverse impacts to indicator species alone simply defines a set of minimum bounds on the effects of human actions on VMEcosystems.
From these considerations we make the following recommendations: (1) use indicator species to identify individual seamount VMCommunities recognizing that protecting part of a seamount identified only by the presence and distribution of an indicator species is not enough (Watling and Auster, 2017); (2) use the seamount classification system of Clark et al. (2011) or it’s equivalent to delimit groups of similar seamounts to focus conservation management efforts and to distinguish between rare and abundant seamount types; (3) examine the similarities among adjacent groups of seamounts to see whether they should be considered to be part of a larger ecosystem group; and (4) evaluate the spatial extent of these larger units so that “significant adverse impacts” measures can be used to determine whether to allow some bottom fishing within a seamount ecosystem group (e.g., allowing one or two seamounts in each ecosystem group to be fished). We show in Figure 2 the classification system as applied to the New England and Corner Rise Seamounts in the NW Atlantic along with the distribution of megafaunal assemblages on some of those seamounts as determined by Lapointe et al. (2020) to illustrate the complexity of the ecosystem problem. Only by defining the boundaries in which fishing impacts occur can we evaluate the impacts to VMEcosystems and those interacting communities within.
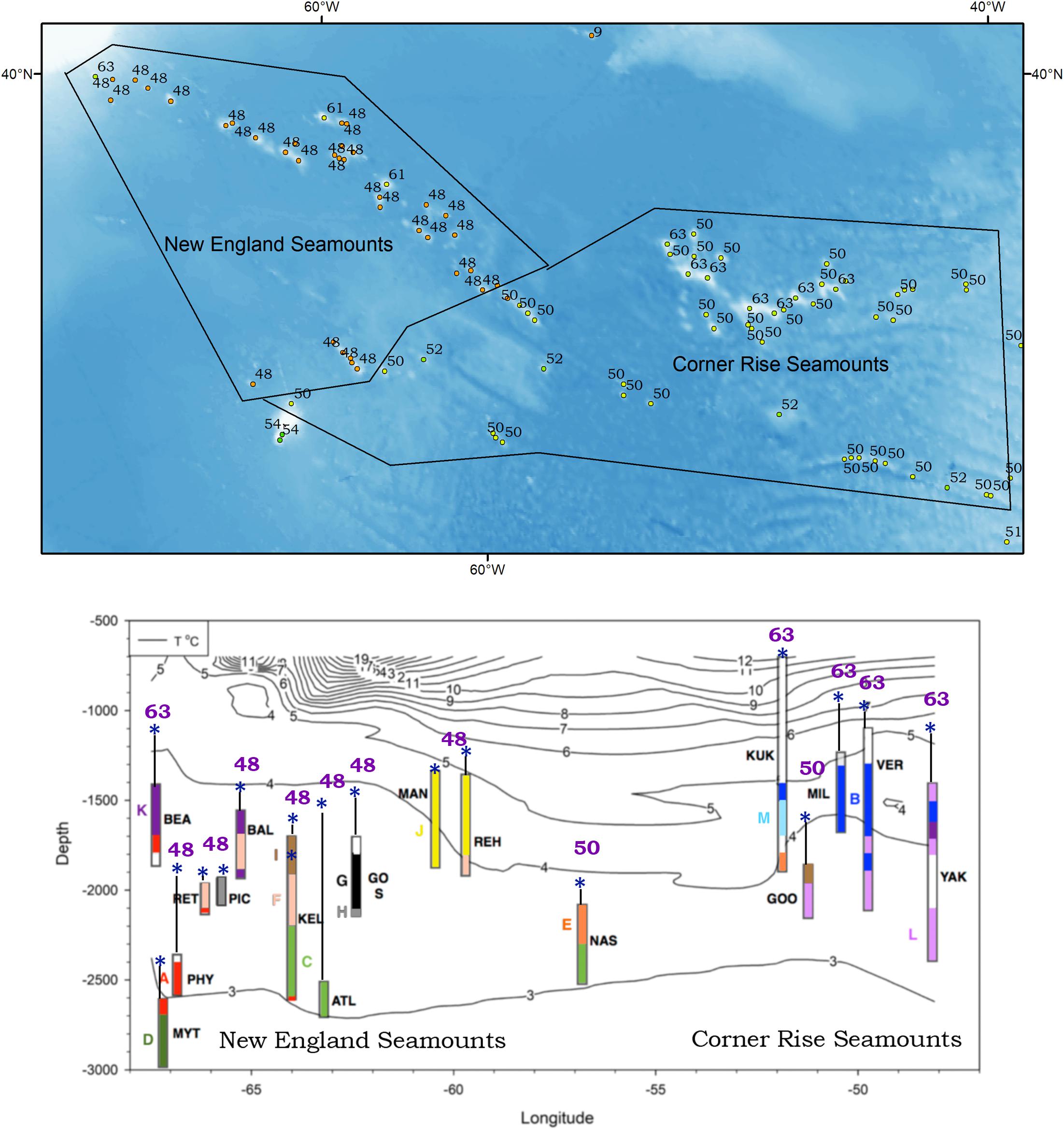
Figure 2. Application of the Clark et al. (2011) seamount classification system to the seamounts of the New England and Corner Rise seamount groups, NW Atlantic whose summits were shallower than 3,500 m. Top panel: Numbers indicate in which group the seamount was included. Seamount groups were distinguished globally using factors such as summit depth, organic matter flux to the seamount summit, temperature at the summit, etc. Lower panel: A chart showing the arrangement of megafaunal assembles on the flanks of the seamounts from the upper panel that were surveyed by submersible or ROV (modified from Lapointe et al., 2020). Colors represent distinct megafaunal assemblages (white indicates areas not surveyed), asterisks indicate summit depths, numbers above the asterisks are the classification group from Clark et al. (2011), and the three letter acronyms represent seamount names (see Lapointe et al., 2020 for details).
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author/s.
Author Contributions
LW and PA drafted and reviewed the manuscript, and prepared it for publication. Both authors contributed to the article and approved the submitted version
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This perspective is modified from a position manuscript by the authors submitted as background for the 8th Scientific Committee of the South Pacific Regional Fisheries Management Organization. The comments of D. Currie, A. Grehan, and the reviewers are much appreciated. The views expressed herein are those of the authors and do not necessarily reflect the views of the authors institutions, collaborators, or funding agencies. This is publication number 108 from the School of Life Sciences, University of Hawai’i at Mānoa.
References
Allee, W. C., Emerson, A. E., Park, O., Park, T., and Schmidt, K. P. (1949). Principles Of Animal Ecology. Philadelphia: W.B. Saunders Co.
Ardron, J. A., Clark, M. R., Penney, A. J., Hourigan, T. F., Rowden, A. A., Dunstan, P. K., et al. (2014). A systematic approach towards the identi fi cation and protection of vulnerable marine ecosystems. Mar. Policy 49, 146–154. doi: 10.1016/j.marpol.2013.11.017
Auster, P. J., Moore, J., Heinonen, K. B., and Watling, L. (2005). “A habitat classification scheme for seamount landscapes: assessing the functional role of deep-water corals as fish habitat,” in Cold-water Corals and Ecosystems, eds A. Freiwald and J. M. Robeerts (Berlin: Springer-Verlag), 761–769. doi: 10.1007/3-540-27673-4_40
Baco, A. R., Morgan, N., Roark, E. B., Silva, M., Shamberger, K. E. F., and Miller, K. (2017). Defying dissolution: discovery of deep-sea scleractinian coral reefs in the North Pacific. Sci. Rep. 7, 1–11. doi: 10.1038/s41598-017-05492-w
Bailey, D. M., Collins, M. A., Gordon, J. D. M., Zuur, A., and Priede, I. G. (2009). Long-term changes in deep-water fish populations in the northeast Atlantic: a deeper reaching effect of fisheries? Proc. R. Soc. B Biol. Sci. 276, 1965–1969. doi: 10.1098/rspb.2009.0098
Bourque, J. R., and Demopoulos, A. W. J. (2018). The influence of different deep-sea coral habitats on sediment macrofaunal community structure and function. PeerJ 6:e5276. doi: 10.7717/peerj.5276
Cho, W., and Shank, T. M. (2010). Incongruent patterns of genetic connectivity among four ophiuroid species with differing coral host specificity on North Atlantic seamounts. Mar. Ecol. 31, 121–143. doi: 10.1111/j.1439-0485.2010.00395.x
Clark, M., and O’Driscoll, R. (2003). Deepwater fisheries and aspects of their impact on seamount habitat in New Zealand. J. Northw. Atl. Fish. Sci. 31, 441–458. doi: 10.2960/j.v31.a34
Clark, M. R., Anderson, O. F., Bowden, D. A., Chin, C., George, S. G., Glasgow, D. A., et al. (2015). Vulnerable marine ecosystems of the Louisville Seamount chain: voyage report of a survey to evaluate the efficacy of preliminary habitat suitability models. New Zealand Aquatic Environment and Biodiversity Report (Wellington). Wellington: New Zealand Aquatic Environment, 1–87.
Clark, M. R., Watling, L., Rowden, A. A., Guinotte, J. M., and Smith, C. R. (2011). A global seamount classification to aid the scientific design of marine protected area networks. Ocean Coast. Manag. 54, 19–36. doi: 10.1016/j.ocecoaman.2010.10.006
Costello, M. J., Tsai, P., Wong, P. S., Cheung, A. K. L., Basher, Z., and Chaudhary, C. (2017). Marine biogeographic realms and species endemicity. Nat. Commun. 8, 1–9. doi: 10.1038/s41467-017-01121-2
Davies, J. S., Stewart, H. A., Narayanaswamy, B. E., and Jacobs, C. (2015). Benthic Assemblages of the Anton Dohrn Seamount (NE Atlantic): Defining Deep-Sea Biotopes to Support Habitat Mapping and Management Efforts with a Focus on Vulnerable Marine Ecosystems. PLoS One 10:e0124815. doi: 10.1371/journal.pone.0124815
Food and Agriculture Organization of the United Nations [FAO] (2009). International Guidelines for the Management of Deep-sea Fisheries in the High Seas. Rome: Food and Agriculture Organization of the United Nations.
George, K. H., and Schminke, H. K. (2002). Harpacticoida (Crustacea, Copepoda) of the Great Meteor Seamount, with first conclusions as to the origin of the plateau fauna. Mar. Biol. 141, 887–895. doi: 10.1007/s00227-002-0878-6
Gofas, S. (2007). Rissoidae (Mollusca: Gastropoda) from northeast Atlantic seamounts. J. Nat. Hist. 41, 779–885. doi: 10.1080/00222930701298085
Kennedy, B. R., Cantwell, K., Malik, M., Kelley, C., Potter, J., Elliott, K., et al. (2019). The unknown and the unexplored: insights into the Pacific deep-sea following NOAA CAPSTONE expeditions. Front. Mar. Sci. 6, 1–21. doi: 10.3389/fmars.2019.00480
Kitchingman, A., and Lai, S. (2004). Inferences on potential seamount locations from mid-resolution bathymetric data. Univ. Br. Columbia Fish. Cent. Res. Rep. 12, 7–12.
Lapointe, A. E., Watling, L., France, S. C., and Auster, P. J. (2020). Megabenthic assemblages in the lower bathyal (700–3000 m) on the New England and Corner Rise Seamounts, Northwest Atlantic. Deep. Res. Part I Oceanogr. Res. Pap. 165:103366. doi: 10.1016/j.dsr.2020.103366
McClain, C. R., Lundsten, L., Barry, J., and DeVogelaere, A. (2010). Assemblage structure, but not diversity or density, change with depth on a northeast Pacific seamount. Mar. Ecol. 31, 14–25. doi: 10.1111/j.1439-0485.2010.00367.x
Morato, T., Pham, C. K., Fauconnet, L., Taranto, G. H., Chimienti, G., Cordes, E. E., et al. (2021). North Atlantic basin-scale multi-criteria assessment database to inform effective management and protection of Vulnerable Marine Ecosystems. Front. Mar. Sci. 8, 1–7. doi: 10.3389/fmars.2021.637078
Morato, T., Pham, C. K., Pinto, C., Golding, N., Ardron, J. A., Muñoz, P. D., et al. (2018). A multi criteria assessment method for identifying vulnerable marine ecosystems in the North-East Atlantic. Front. Mar. Sci. 5:460. doi: 10.3389/fmars.2018.00460
Morgan, N. B., Cairns, S., Reiswig, H., and Baco, A. R. (2015). Deep-Sea Research I Benthic megafaunal community structure of cobalt-rich manganese crusts on Necker Ridge. Deep. Res. 1 Oceanogr. Res. Pap. 104, 92–105. doi: 10.1016/j.dsr.2015.07.003
O’Neill, R. V., DeAngelis, D. L., Waide, J. B., and Allen, T. F. H. (1986). A hierarchical concept of ecosystems. Princeton: Princeton University Press.
Pierrejean, M., Grant, C., de Moura Neves, B., Chaillou, G., Edinger, E., Blanchet, F. G., et al. (2020). Influence of Deep-Water Corals and Sponge Gardens on Infaunal Community Composition and Ecosystem Functioning in the Eastern Canadian Arctic. Front. Mar. Sci. 7:495. doi: 10.3389/fmars.2020.00495
Du Preez, C., Curtis, J. M. R., and Clarke, M. E. (2016). The structure and distribution of benthic communities on a shallow seamount (Cobb Seamount, Northeast Pacific Ocean). PLoS One 11:e0165513. doi: 10.1371/journal.pone.0165513
Priede, I. G., Godbold, J. A., Niedzielski, T., Collins, M. A., Bailey, D. M., Gordon, J. D. M., et al. (2011). A review of the spatial extent of fishery effects and species vulnerability of the deep-sea demersal fish assemblage of the Porcupine Seabight, Northeast Atlantic Ocean (ICES Subarea VII). ICES J. Mar. Sci. 68, 281–289. doi: 10.1093/icesjms/fsq045
Radice, V. Z., Quattrini, A. M., Wareham, V. E., Edinger, E. N., and Cordes, E. E. (2016). Vertical water mass structure in the North Atlantic influences the bathymetric distribution of species in the deep-sea coral genus Paramuricea. Deep. Res. 1 Oceanogr. Res. Pap. 116, 253–263. doi: 10.1016/j.dsr.2016.08.014
Rowden, A. A., Anderson, O. F., Georgian, S. E., Bowden, D. A., Clark, M. R., Pallentin, A., et al. (2017). High-resolution habitat suitability models for the conservation and management of vulnerable marine ecosystems on the Louisville Seamount Chain, South Pacific Ocean. Front. Mar. Sci. 4:335. doi: 10.3389/fmars.2017.00335
Samadi, S., Bottan, L., Macpherson, E., De Forges, B. R., and Boisselier, M. C. (2006). Seamount endemism questioned by the geographic distribution and population genetic structure of marine invertebrates. Mar. Biol. 149, 1463–1475. doi: 10.1007/s00227-006-0306-4
Sherman, K., and Duda, A. M. (1999). An ecosystem approach to global assessment and management of coastal waters. Mar. Ecol. Prog. Ser. 190, 271–287. doi: 10.3354/meps190271
Thoma, J. N., Pante, E., Brugler, M. R., and France, S. C. (2009). Deep-sea octocorals and antipatharians show no evidence of seamount-scale endemism in the NW Atlantic. Mar. Ecol. Prog. Ser. 397, 25–35. doi: 10.3354/meps08318
Thresher, R. E., Adkins, J., Fallon, S. J., Gowlett-Holmes, K., Althaus, F., and Williams, A. (2011). Extraordinarily high biomass benthic community on Southern Ocean seamounts. Sci. Rep. 1:119. doi: 10.1038/srep00119
Victorero, L., Robert, K., Robinson, L. F., Taylor, M. L., and Huvenne, V. A. I. (2018). Species replacement dominates megabenthos beta diversity in a remote seamount setting. Sci. Rep. 8:4152. doi: 10.1038/s41598-018-22296-8
Watling, L., and Auster, P. J. (2017). Seamounts on the high seas should be managed as vulnerable marine ecosystems. Front. Mar. Sci. 4:14. doi: 10.3389/fmars.2017.00014
Watling, L., Guinotte, J., Clark, M. R., and Smith, C. R. (2013). A proposed biogeography of the deep ocean floor. Prog. Oceanogr. 111, 91–112. doi: 10.1016/j.pocean.2012.11.003
Keywords: vulnerable marine ecosystem, seamount, marine communities, Solenosmilia, fisheries
Citation: Watling L and Auster PJ (2021) Vulnerable Marine Ecosystems, Communities, and Indicator Species: Confusing Concepts for Conservation of Seamounts. Front. Mar. Sci. 8:622586. doi: 10.3389/fmars.2021.622586
Received: 28 October 2020; Accepted: 26 April 2021;
Published: 25 May 2021.
Edited by:
Anthony Grehan, National University of Ireland Galway, IrelandReviewed by:
Chris Yesson, Zoological Society of London, United KingdomNadia Papadopoulou, Hellenic Centre for Marine Research (HCMR), Greece
Copyright © 2021 Watling and Auster. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Les Watling, watling@hawaii.edu