 Gregory A. Kolbasov
Gregory A. Kolbasov Alexandra S. Savchenko2
Alexandra S. Savchenko2 Benny K. K. Chan
Benny K. K. Chan- 1White Sea Biological Station, Biological Faculty, Moscow State University, Moscow, Russia
- 2Department of Invertebrate Zoology, Biological Faculty, Moscow State University, Moscow, Russia
- 3Marine Biological Research Division, Scripps Institution of Oceanography, La Jolla, CA, United States
- 4Biodiversity Research Center, Academia Sinica, Taipei, Taiwan
A new ectoparasitic ascothoracidan species has been discovered off northern Tasman Sea at a depth of 520 m. Single female, described herein as Waginella ebonita Kolbasov and Newman sp. nov. in the family Synagogidae Gruvel, 1905, was collected on the column of crinoid Metacrinus sp. This is the first study of a new form of Waginella to be based on the extensive use of both light and scanning electron microscopy to document the fine-scale external morphology. We studied the external morphology of other two species of Waginella, W. cf. axotremata and W. sandersi utilizing SEM and compared fine structures of all congeners. The genus Waginella represents a monophyletic taxon including at least 4 species having similar and unique morphology even on the ultrastructural level. Two species, W. cf. axotremata and W. cf. metacrinicola may represent at least two complexes of species. The morphology of an attachment apparatus in all species of Waginella suggests that they remain attached to the host for a considerable period of time. The lattice organs of Waginella have a unique structure distinguishing from all Thecostraca. Waginella ebonita sp. nov. is the second species of this genus known to harbor hyperparasitic cryptoniscid isopods representing parasitic castrators.
Introduction
The Ascothoracida are relatively little known, exclusively parasitic crustaceans that range from ecto- to endoparasites found on cnidarians (Alcyonacea, Antipatharia, Scleractinia, and Zoantharia) and echinoderms (Asteroidea, Crinoidea, Echinoidea, and Ophiuroidea) when compared to other crustacean parasitic taxa including parasitic copepods and isopods (Figure 1). Currently the Ascothoracida is comprised of 114 described species assigned to two orders (Grygier, 1987a, 1996): the Laurida, species of which are parasites of anthozoans except for those of Waginella Grygier, 1983a, which are ectoparasites of crinoids, and the Dendrogastrida, species of which are parasites of non-crinoid echinoderms. The monophyly of these two orders is yet to be tested by molecular methods (Chan et al., 2021). The ascothoracidans are largely dioecious, with larger females and smaller, sometimes dwarf cypridiform males (Grygier and Fratt, 1984; Grygier, 1985, 1987b, 1991a, 1991b; Kolbasov, 2007). The family Petrarcidae, and possibly some of Ctenosculidae, are endoparasites in scleractinian corals and sea stars and have secondarily evolved hermaphroditism (Okada, 1938; Grygier, 1983b,c).
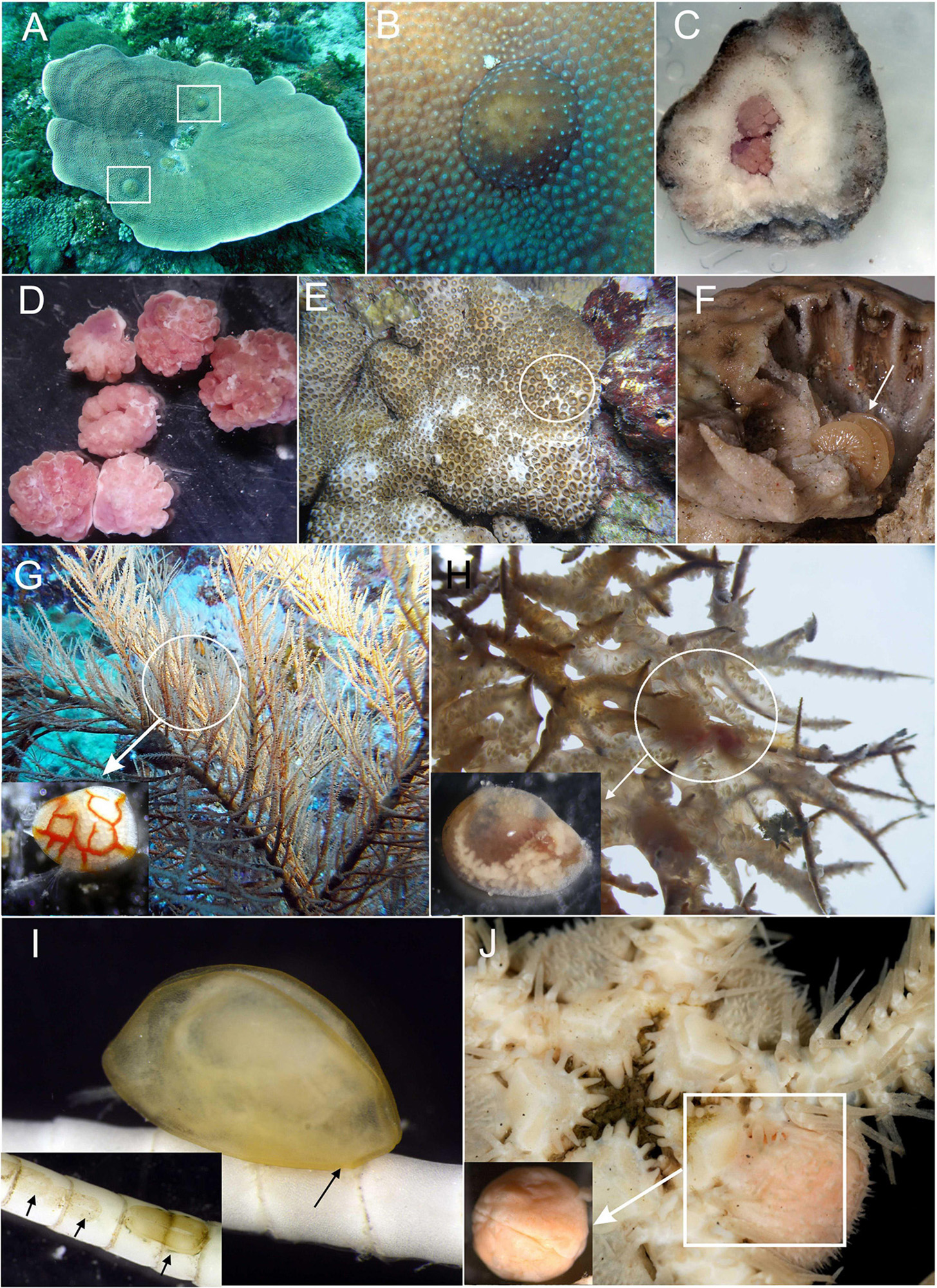
Figure 1. Diversity of parasitic Ascothoracida. (A) Petrarca—genus endoparasitic in Turbinaria corals, forming globular galls. (B) Gall formed by Petrarca on coral surface. (C) Underneath of gall chipped off from coral, showing two individuals of Petrarca. (D) Individuals of Petrarca isolated from their galls. (E) Zoanthid Palythoa sp.—host of endoparasitic genus Baccalaureus. (F) Dissected zoanthid colony showing female of Baccalaureus (indicated by arrow). (G) Black coral (antipatharian) colony—host of free-swimming, ectoparasitic genus Synagoga, (isolated female in lower left corner, modified after Kolbasov et al., 2019). (H) Anastomosing colony of black coral (antipatharian)—host of endoparasitic but vagile genus Sessilagoga (isolated female in lower left corner, modified after Kolbasov et al., 2020). (I) Waginella—genus ectoparasitic in stalked crinoids (inserts showing multiple attachment pads on host after parasites were detached). (J) Genus Ascothorax—mesoparasite of ophiuroids (isolated female in lower left corner, modified after Kolbasov and Petrunina, 2018).
The life cycle of ascothoracidans is depending on species, the larval stages may be free-swimming or brooded. It potentially includes up to six planktotrophic or lecithotrophic naupliar instars, albeit in a few species the naupliar phase is condensed or even omitted (Høeg et al., 2014), followed by one or two instars of a specialized non-feeding ascothoracid larva that infects the host. The family Synagogidae represents the basal group of ascothoracidans, the most generalized of which belong to the genera Synagoga Norman, 1888, Sessilogoga Grygier, 1990b, and Waginella Grygier, 1983a. Adults of these genera retain some degree of vagility throughout their lives and are characterized by a bivalve carapace enclosing the whole body, the head with a pair of W-shaped, six-segmented prehensile antennules and an oral cone enclosing piercing mouthparts. The trunk consists of 11 segments including six thoracomeres with biramous thoracopods, a genital somite (first abdominal segment) bearing a sexually dimorphic penis which is vestigial in females, three limbless abdominal somites, and a telson bearing a pair of furcal rami. Three species of Synagoga are known to be free-swimming ectoparasites or grazers of antipatharians and probably others feed likewise on black corals (Norman, 1888, 1913; Kolbasov and Newman, 2018; Kolbasov et al., 2019). Sessilogoga includes two species representing the vagile endoparasites of antipatharians (Grygier, 1990a; Kolbasov et al., 2020).
Genus Waginella was proposed by Grygier (1983a) for two former species of Synagoga: W. metacrinicola (Okada, 1926) and W. sandersi (Newman, 1974), as well as for his new species W. axotremata Grygier, 1983. Among these species, adults of W. sandersi were collected as free-swimming, their food source or host being unknown (Newman, 1974), while the other two species, W. metacrinicola and W. axotremata, are (temporarily?) cemented to their host crinoids of the subfamily Metacrininae (Grygier, 1983a, 1990c; Grygier and Itô, 1995). W. sandersi was collected from abyssal depths off Patagonia, south Atlantic (Newman, 1974) whereas both W. metacrinicola and W. axotremata range from the lower subtidal to upper bathyal zone of the Indo-Pacific and maybe sympatric (Grygier and Itô, 1995). The descriptions of species of Waginella were mainly done by light microscopy (Newman, 1974; Grygier, 1983a). Grygier and Itô (1995) subsequently employed scanning electron microscopy (SEM) to study the external morphology of a female W. metacrinicola.
The new species, Waginella ebonita sp. nov. was collected off northern Tasman Sea at a depth of 520 m, on the column of crinoid Metacrinus sp. (Figure 2). It differs morphologically from all congeners. In contrast to its congeners previously described, the new species is based on the extensive use of both light and scanning electron microscopy to document the fine-scale external morphology. Furthermore, we studied the external morphology of other two species of Waginella, W. cf. axotremata from Taiwan and W. sandersi from type locality utilizing SEM and thus were able to compare fine structures of all species of Waginella.
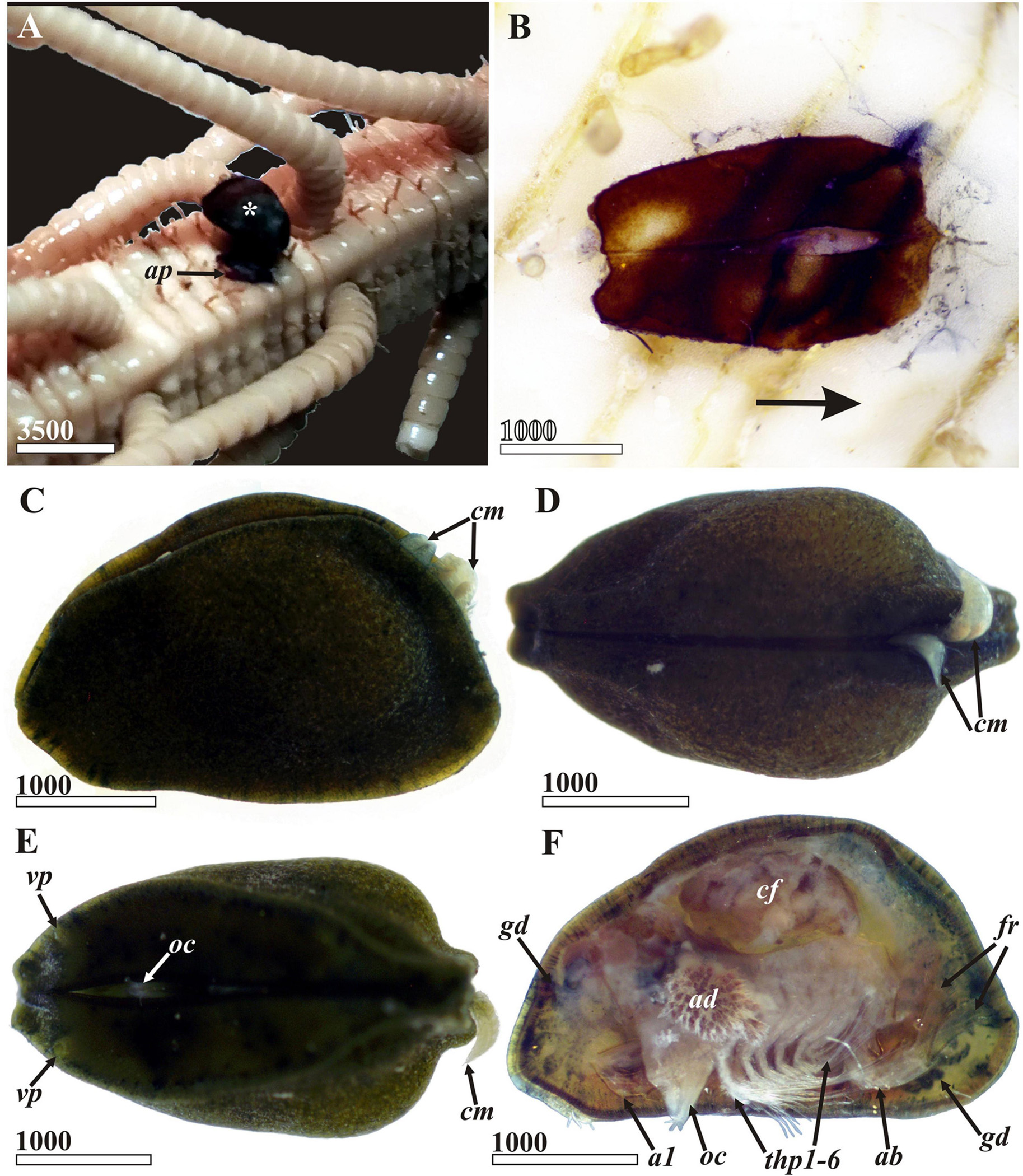
Figure 2. Waginella ebonita sp.nov. General appearance, light microscopy, female, holotype (ethanol fixation). (A) Holotype (indicated by asterisk) on the column of host, Metacrinus sp. partially detached during preparation. (B) Attachment pad of cement on host column (anterior direction of parasite indicated by arrow). (C) General view lateral, left side, anterior end left (two males of a hyperparasitic cryptoniscid isopod clinging to posteriodorsal side of entrance to mantle cavity seen also in (D,E). (D) Dorsal view, anterior end left. (E) Ventral side, anterior end left (tip of oral cone outstretched between valves in anterior third). (F) General view lateral, left valve removed (female of hyperparasitic cryptoniscid isopod occupies brooding chamber of ascothoracidan). a1, antennule; ap, attachment pad; ab, abdomen; ad, adductor muscle; cf, female of cryptoniscid isopod; cm, males of cryptiniscid isopods; fr, furcal rami; gd, gut diverticulum; oc, oral cone; thp1–6, thoracopods 1–6; vp, anterioventral carapace pore. Scale bars in μm.
Materials and Methods
The holotype female of the new species, Waginella ebonita sp. nov., discovered during the survey of the crinoid collections of the Zoological Institute RAS (St.-Petersburg). It was attached to the column of Metacrinus sp. (Figure 2A), and two other attachment pads were found on the columns of two other Metacrinus specimens. The crinoids had been collected in a Sigsbee trawl at a depth 520 m in northern part of Tasman Sea. A holotype was detached from host column and studied with light microscope and SEM. Its carapace was then dissected in order to observe morphology of body proper and its structures (Figure 2F). Carapace valves, dissected mouth parts, antennules, trunk with furcal rami and all thoracopods were mounted in glycerol on glass slides and examined using Olympus BX 43 light microscope. Line drawings were made using a drawing tube on the same microscope. The carapace valves, left antennule and trunk with furcal rami were washed in deionized water, postfixed in 2% OsO4 for 2 h, dehydrated in ethanol and acetone, and critical point dried by CO2, sputter-coated with platinum–palladium and examined on JEOL JSM-6380LA scanning electron microscope at operating voltages of 15–20 kV at the University of Moscow. Resulting photographs were touched up using the CorelDraw X3 Graphics Suite.
Three specimens (2 females and 1 male) of W. cf axotremata and two individuals (females) of W. sandersi were studied with SEM. Specimens of W. cf axotremata came from different locations off east Taiwan (collections of the National Museum of Natural History, Taichung, Taiwan). These were one female from a cirrus of Metacrinus sp., 24°34.60′N, 122°5.84′E, 223–260 m, R/V Ocean Researcher 1, 26.08.2003; another on a cirrus of Metacrinus interruptus Carpenter 1884, 22°12.376′N, 120°5.272′E, 699–912 m, R/V Ocean Researcher 3, 11.07.2008; and one male on cirrus of Saracrinus sp., 22°1.207′N, 118°53.827′E, 757–1311 m, R/V Ocean Researcher 1, 30.04.2016. Two females of W. sandersi were sent to GAK from collections of Scripps Institution of Oceanography (United States); locality: C 8422, cruise No. 60, sta. 274a, S.W. Atlantic, 43.0°33.0′S 48.0°58.0′W, 5200.0 m, large epibenthic sled, 17.03.1971, R/V Atlantis II, coll. H.L. Sanders, det. W. Newman.
Results
Systematics
Subclass Ascothoracida de Lacaze-Duthiers, 1880
Order Laurida Grygier, 1987a
Family Synagogidae Gruvel, 1905
Genus Waginella Grygier, 1983
Type species: Waginella metacrinicola (Okada, 1926)
Waginella ebonita Kolbasov and Newman sp. nov., herein

Figure 3. Waginella ebonita sp. nov., female, holotype. General morphology. (A) Left valve of carapace, lateral view, inner side, anterior end right. (B,C) Anterior and posterior gut diverticula, right valve of carapace, inner side. (D) Frontal filament complex, anterior margin right. (E) Posterior part of trunk (thoracic segments numbered (Roman numerals), abdominal segments (Arabic numerals). (F) Rudimentary penis. (G) Telson with furcal rami. (H) Posterior end of furcal ramus. 5(te), fifth abdominal segment (telson); ad, adductor muscle; eu, epaulet; ffc, frontal filament complex; fr, furcal rami; gd, gut diverticulum; per, rudimentary penis; ts, telsonic spines. Scale bars in μm.
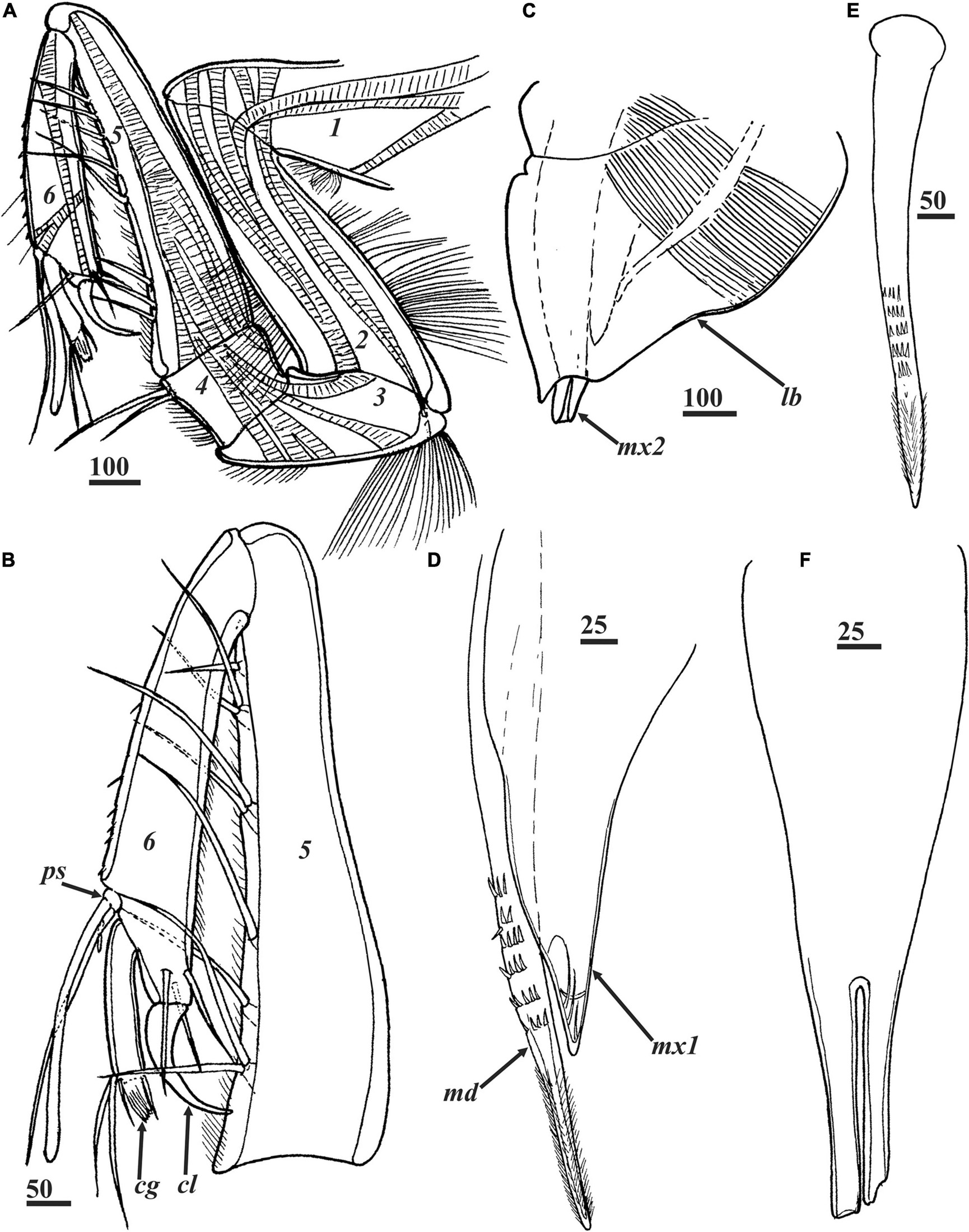
Figure 4. Waginella ebonita sp. nov., female, holotype. Head appendages. (A) Left antennule with musculature, segments numbered. (B) Right antennule, distal part, segments numbered. (C) Oral cone. (D) Mandible and maxillule. (E) Mandible. (F) Maxillae, proximal part, distal tips broken. cg, claw guard; cl, claw; lb, labrum; md, mandible; mx1, maxillule; mx2, maxillae; ps, proximal sensory process. Scale bars in μm.
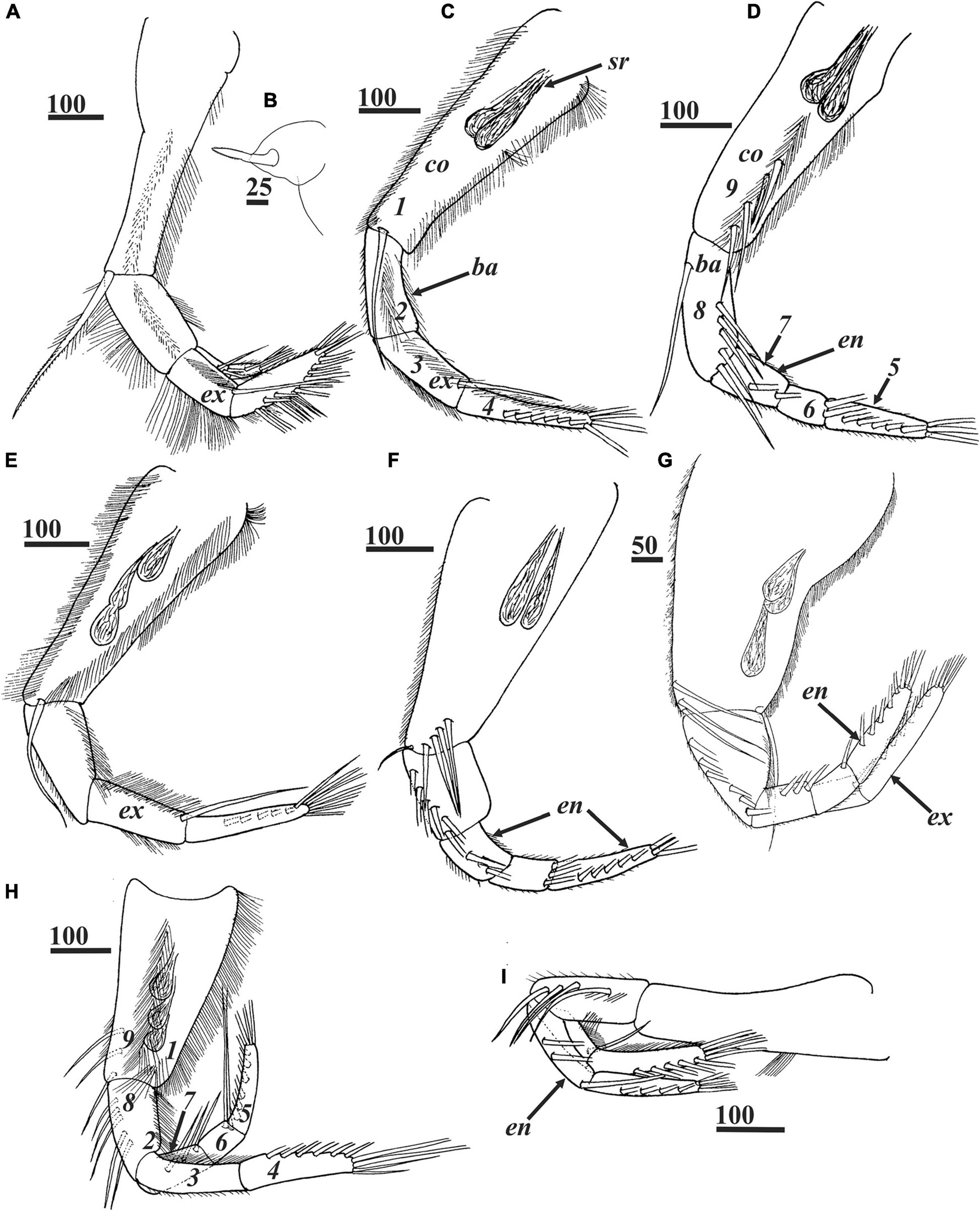
Figure 5. Waginella ebonita sp. nov., female, holotype. (A) Left thoracopod 1. (B) Filamentary appendage at base of thoracopod 1. (C,D) Left and right thoracopods 2. (E,F) Left and right thoracopods 3. (G) Right thoracopod 4. (H) Left thoracopod 5. (I) Right thoracopod 6. Ampuliform seminal receptacles are situated in upper outer parts of coxae of thoracopods 2–5 (C–H). Numbers indicating positions for setal counts in description (1–9) are shown for thoracopods 2 (C,D). ba, basis; co, coxa; en, endopod; ex, exopod; sr, seminal receptacles. Scale bars in μm.

Figure 6. Waginella ebonita sp. nov., female, holotype. Morphology of attachment pad (SEM). (A) Attachment pad on host column, anterior end left (location of feeding hole in rectangle area “B”). (B) Feeding hole burrowed by mouth parts of parasite (presumably mandibles). (C) Surface of cement of attachment pad. (D) Enlarged part of attachment cement. Scale bars in μm.
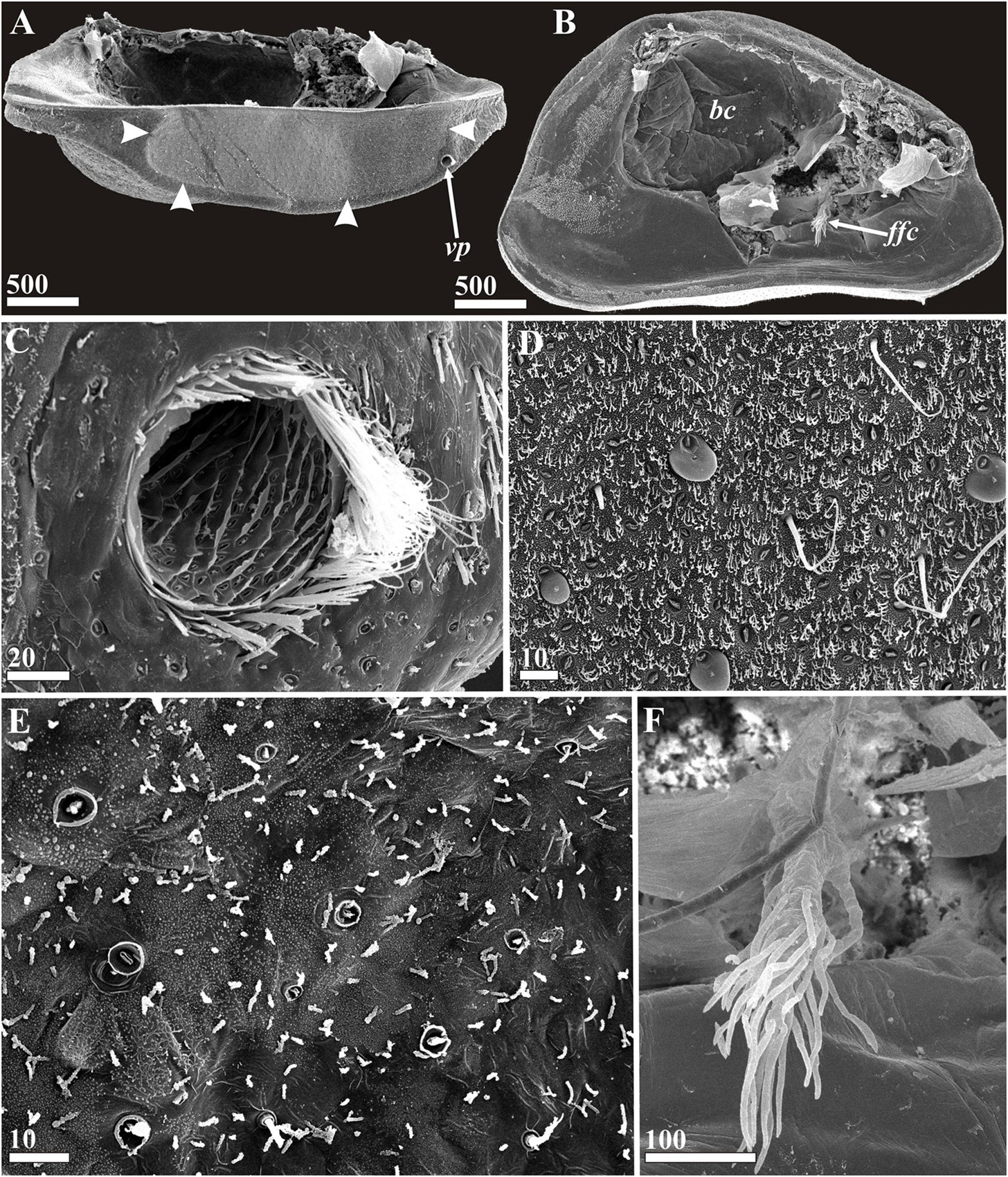
Figure 7. Waginella ebonita sp. nov., female, holotype. Carapace structures (SEM). (A) Left valve of carapace, ventral view, anterior end right (border of area producing attachment cement indicated by arrowheads). (B) Left valve of carapace, lateral view, inner side, anterior end right. (C) Anterioventral pore, anterior end right. (D) Surface of ventral part of carapace producing cement (note gland tubes, dense cuticular villi and setae, some gland tubes (4) with bulbs of cement). (E) Surface of ventral part of carapace not producing cement (note gland tubes without bulbs of cement, rare cuticular villi and setae). (F) Frontal filament complex. bc, brood chamber; ffc, frontal filament complex; vp, anterioventral carapace pore. Scale bars in μm.
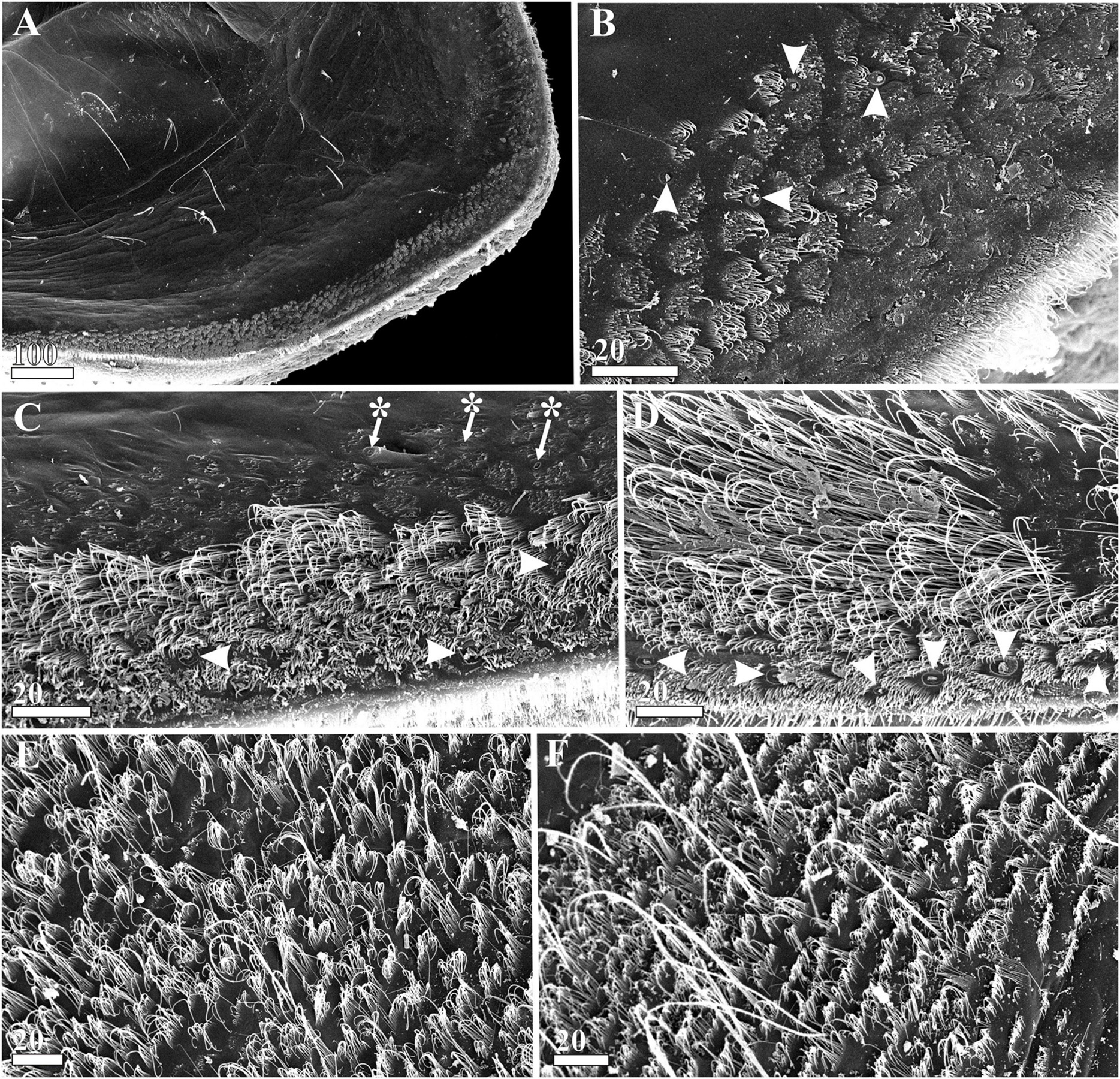
Figure 8. Waginella ebonita sp. nov., female, holotype. Carapace mantle structures (SEM). (A) Surface of mantle, anterioventral part. (B) Ornamentation of mantle surface at anterior margin (gland tubes indicated by arrowheads). (C) Ornamentation of mid-ventral part of mantle surface (gland tubes indicated by arrowheads, cuticular pores—by asterisks). (D) Ornamentation of posterioventral part of mantle surface (marginal gland tubes indicated by arrowheads). (E) Ornamentation of mantle surface at posterior margin. (F) Ornamentation of mantle surface at posteriodorsal margin. Scale bars in μm.

Figure 9. Waginella ebonita sp. nov., female, holotype. Morphology of trunk (SEM). (A) Posterior part of trunk, left furcal ramus removed (thoracic segments—Roman numerals, abdominal segments—Arabic numerals). (B) Rudimentary penis. (C,D) Epaulet and its enlarged surface respectively. (E) Posterior margin of telson with proximal part of furcal ramus. (F) Upper margin of furcal ramus, middle part (insertions of broken setae indicated by arrowheads). 5(te), fifth abdominal segment (telson); eu, epaulet; fr, furcal rami; per, rudimentary penis; ts, telsonic spines. Scale bars in μm.

Figure 10. Waginella ebonita sp. nov., female, holotype. Morphology of antennules (SEM). (A) Left antennule, segments numbered. (B) Distal part of left antennule, segments numbered. (C) Distal part of sixth antennular segment. (D) Proximal part of proximal sensory process (setae indicated by asterisks). (E) Tip of claw guard (setae indicated by asterisks). (F) Inner surface of claw, middle part. cg, claw guard; cl, claw; ps, proximal sensory process. Scale bars in μm.
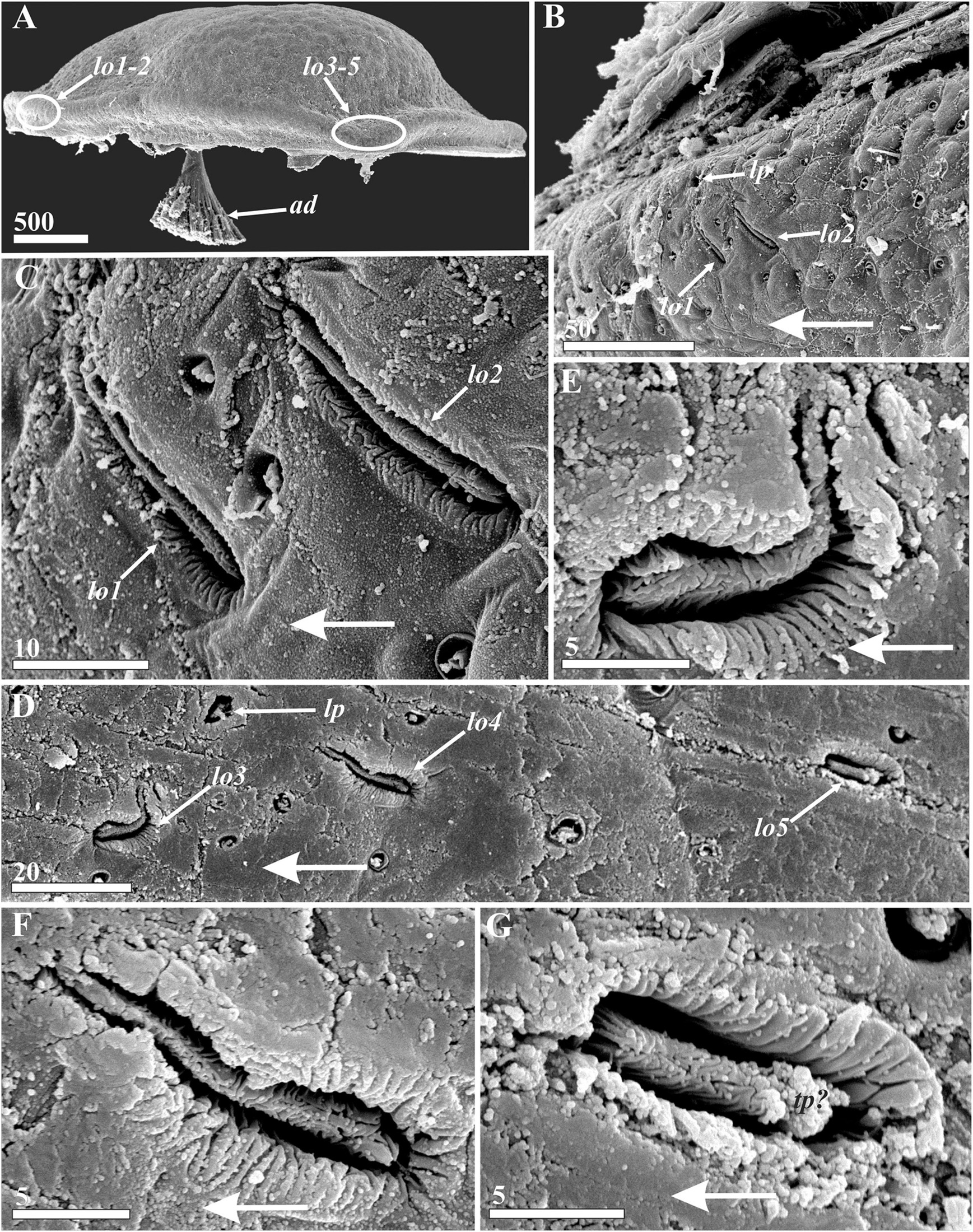
Figure 11. Waginella ebonita sp. nov., female, holotype. Lattice organs, with large arrows indicating anterior direction (SEM). (A) Locations of lattice organs (indicated by oval outlines) on right valve of carapace, anterior end left. (B) Anterior lattice organs (1, 2) on left carapace valve. (C) Enlarged left anterior lattice organs (1, 2). (D) Left posterior lattice organs (3–5). (E–G) Left posterior lattice organs of 3, 4, and 5 pair, respectively. ad, adductor muscle; lo1–5, lattice organs; lp, large pore associated with lattice organs; tp?, probable terminal pore of lattice organ. Scale bars in μm.
Type Locality
R/V “Akademik Oparin,” 7th cruise, st. 159, 11.09.1988, northern part of Tasman Sea, 32°27.0′S, 162°35.9′E, 520 m, Sigsbee trawl, coll. A.V. Smirnov, on Metacrinus sp.
Material Examined
One female specimen of the new species, Waginella ebonita sp. nov., was collected from the column of crinoid Metacrinus sp. Slides of the mouth parts, right antennule and thoracopods and SEM stub with carapace, trunk and left antennule of holotype Mg 1246 are deposited in the Zoological Museum of Moscow State University in Moscow, Russian Federation.
Diagnosis
Diagnosis is provided for the female, and a full list of interspecific differences is given in Table 1.

Table 1. Main diagnostic characters of species of the genus Waginella (modified from Newman, 1974; Grygier and Itô, 1995).
Female attached to crinoid column; carapace elliptical, dark, almost black colored, anterior and posterior ends angular in side view, 3.5 mm long, 1.9 mm high, and 2.0 mm wide. Fourth antennular segment with a pair of long, slender simple setae; fifth segment with 8 or 9 long, slender setae. Distal segment of exopod of thoracopod 1 with 16 setae; coxae of thoracopods 4 and 5 with 3 seminal receptacles. Telsonic spines about 1/3 of blade length of furcal ramus; furcal rami 3 times longer than high, inner surface of furcal ramus with 16 medial setae.
Etymology
From ancient Greek “ε’´βενoς” (ebenos)—ebony tree, referring to the dark carapace coloration.
Description
Female attached to column of crinoid, coloration of carapace in ethanol dark, almost black (Figures 2A,C–E). Attachment (cement) pad dark brown (Figure 2B), 2.1–2.3 mm long, consisting of two elongated halves (Figure 6A) diverging anterior and corresponding to flat ventral surfaces of carapace. A small rounded feeding hole about 45 μm in diameter and corresponding to tip of oral cone lies between halves of anterior third of the attachment pads (Figures 6A,B). Surface of attachment pad spongy, with rounded, 5–10 μm in diameter pits corresponding to bulbs of cement producing by cement glands and numerous tips of gland tubes imbedded in attachment cement (Figures 6C,D, 7D).
Carapace bivalved, ellipsoid, 3.5 mm long, 1.9 mm high, and 2.0 mm wide, anterior and posterior ends angular (Figures 2C,F, 3A). Dorsal margin of valves convex, valves joined by a hinge along dorsal margin and between them having an elongate posteriodorsal and a smaller rounded anteriodorsal aperture (Figures 2C,D,F) plus two dorsolateral spherical inflations corresponding to brood chamber (Figures 2D, 11A). Ventral side flattened, with large anterior pore on each valve and narrow gap in anterior half between them allowing the oral cone to protrude into the host (Figures 2E, 7A). Large anterioventral pore round, 77 μm in diameter, with tufts of fine setae arise from sockets along its rim especially dense at anterior end and ranging to absent in posterior part (Figure 7C). Surface of pore canal with lamellar ridges forming lattice pattern with each square having one small oval pore 3–4 μm long (Figure 7C). Right and left gut diverticula (Figures 1F, 3A–C) lying within their respective carapace valves, each with anterior branch shorter than posterior, numerous simple and bifid small branches extending from them in various directions; posterior branch circumflex brood chamber.
Exterior of carapace slightly uneven with more or less regular array of papilliform gland tubes in circular pits about 3–6.5 μm in diameter and rare small setae covering whole surface of valves (Figures 7C–E, 11A,D). The dense carpet of cuticular villi around each gland tube and seta denser in attachment zone of ventral side (Figures 7A,D). Several gland tubes in the attachment zone have bulbs of attachment cement exuding from them (Figure 7D). Gland tubes, setae and cuticular villi are not so frequent behind attachment zone and lack bulbs of cement (Figure 7E).
Inner surface of carapace valves with cuticular lining (Figure 7B). Cephalic cuticle confluent with anterior part in zone extending anteriodorsally from a point near adductor muscle and gut diverticulum on each side (Figures 3A, 11A). Remainder of the body hanging freely within mantle cavity (Figure 2F). An oval brood chamber occupies the mid-dorsal portion of each valve, with smooth cuticle and sparse long, thin, simple setae at the posterior opening (Figures 3A, 7B). Frontal filament complex (Figures 3A,D, 7B,F) originating on mantle beneath entrance of adductor muscle, tripartite, with long ramus (ca. 320 μm) and densely covered by long setiform protrusions and two ampuliform, short basal processes/rami (ca. 100 and 57 μm) with smooth cuticle.
Cuticular armament of mantle similar to that in W. cf. metacrinicola (see Grygier and Itô, 1995). Main cuticular structures of mantle arrayed along its margin (Figure 8): cuticular denticles 5–10 μm long form dense, brush-lake scales with rare gland tubes along anterior and anterioventral sides (Figures 8A,B). Ventral side with dense pelage consisting of 20–40 μm long setiform denticles underlying by almost linear row of gland tubes and dense short denticles, and upper short brush-like scales with cuticular pores (Figures 8C,D). Posterior margin of mantle with dense irregular scales of 20–40 μm long setiform denticles (Figure 8E); similar scales arranged in several transverse rows and thin setae with fine setules observed in posteriodorsal side (Figure 8F); gland tubes absent in posterior and posteriodorsal sides.
Body proper consisting of unsegmented head and segmented thorax and abdomen (Figures 2F, 3E, 9A). Head bearing W-shaped antennules followed by developed ventral oral cone (Figures 2F 4A,C, 10A). Thorax consisting of six segments (Figures 2F, 3E, 9A), each with pair of biramous natatory thoracopods. Dorsal and lateral sides of segments covered with rare thin, long setae; tufts of thin, long setae inserted mid-laterally between segments (Figures 3E, 9A). Posterioventral angles of sixth thoracic segment with conspicuous epaulets (Figures 3E, 9A) consisting of proximal part with smooth cuticle and inflated, rounded distal part covered by polygonal cuticular knobs with fine, spherical plaques (Figures 9C,D).
Extendable subchelate prehensile antennules folded into W-shape consist of six segments with complex of intrinsic and extrinsic flexor and extensor muscles (Figures 4A,B, 10A,B). First segment trapezoidal, narrowing somewhat distally, with thin setae on preaxial/dorsal margin (Figures 4A, 10A). Second segment oblong, rectangular, with dense, thin omniserrate setae along dorsal and postaxial/ventral margins (Figures 4A, 10A). Third segment trapezoid, narrowing toward proximal end; dorsal margin with proximal tuft of dense, long thin omniserrate setae and smaller distal tuft of short setae (Figures 4A, 10A). Fourth segment narrow, trapezoid, narrowing toward ventral margin, with two conspicuous setae and row of short thin setae on dorsal margin (Figures 4A, 10A,B). Fifth segment longest, conical, forming a palm against which sixth segment can fold, with 8–9 strong, simple setae and row of ctenoid scales and short thin setae along dorsal margin (Figures 4A,B, 10A,B). Sixth segment significantly shorter than fifth segment (about 2/3 of its length) and armed with sensory and grasping structures (Figures 4A,B, 10A,B). Subterminal rudimentary proximal sensory process at base of claw guard (Figures 4B, 10B–D), with 3 terminal setae on small bump, 1 thick, long, blunt seta or aesthetasc, 1 long simple seta and 1 small seta with denticles (Figures 4B, 10C,D). Curved claw on distal end of sixth segment apparently with muscles attached (Figures 4A,B, 10B,C); concave margin of claw serrate, with microscopic denticles in middle part (Figure 10F); three small setae at base of claw, two lateral on inner and outer surfaces and one on anterior dorsal margin (Figures 4B, 10C). Grooved claw guard (Figures 4B, 10B,C), approximately 190 μm long, with thin, membranous, apical ctenoid hood (Figure 10E) and 4 small terminal setae including 2 longer and 1 tiny subapical, and 1 tiny apical seta (Figure 10E).
Oral cone prominent, approximately 650 μm long; distal end often protruding outside carapace (Figures 2F, 4C); formed by cone-shaped labrum surrounding piercing mouth parts. Posterior margins of labrum free, unfused; suraface of labrum without setae; developed bulk of pharyngeal mussels, indicating on suctorial way of feeding, lies in posterior part of labrum around pharynx (Figure 4C). Styliform rasp-like mandibles, approximately 660 μm long (Figures 4E,D); distal part with basally directed, dense and delicate setiform denticles, proximally basally directed robust denticles associated in 5–6 transverse rows. Maxillules triangular, narrowing distally, without any armament, except feeble cutiular ridges at tip (Figure 4D). Maxillae thin, fused at bases, distal ends broken (probably stayed in host) (Figure 4F).
All thoracopods natatory and biramous, with long, plumose setae and lined with dense fine setae (Figure 5). Seminal receptacles found in lateral proximal parts of coxae of thoracopods 2–5 (Figures 5C–H), consisting of ampuliform sacs with proximal parts converging but external opening(s) not observed; at least thoracopods 4 and 5 with 3 seminal receptacles, thoracopods 2 and 3 with two. Thoracopodal setation summarized in Table 2. First thoracopod slightly separated from others, with elongate protopod comprised of coxa and basis and two-segmented exopod and endopod (Figure 5A); coxa with large, distal seta in position “1” (see Table 2 for further explanation); exopod twice longer than endopod, with 19 long, plumose setae on distal segment; both segments of endopod bearing 3 long, plumose setae. Filamentary appendage with pointed process at base of thoracopod 1 (Figure 5B). Thoracopods 2–5 with three-segmented endopods and two-segmented exopods (Figures 5C–H). Coxae of thoracopods 2, 3, 4 (Figures 5C,E,G) with distal seta in position “1”. Thoracopods 2–5 with 23, 27, 23 and 24 long pulmose setae totally on both rami (Figures 5C–H). Protopod of thoracopod 6 (Figure 5I) narrow; coxa without setae; both rami two-segmented with 9 and 8 long, plumose setae on distal segments of exopod and endopod respectively.
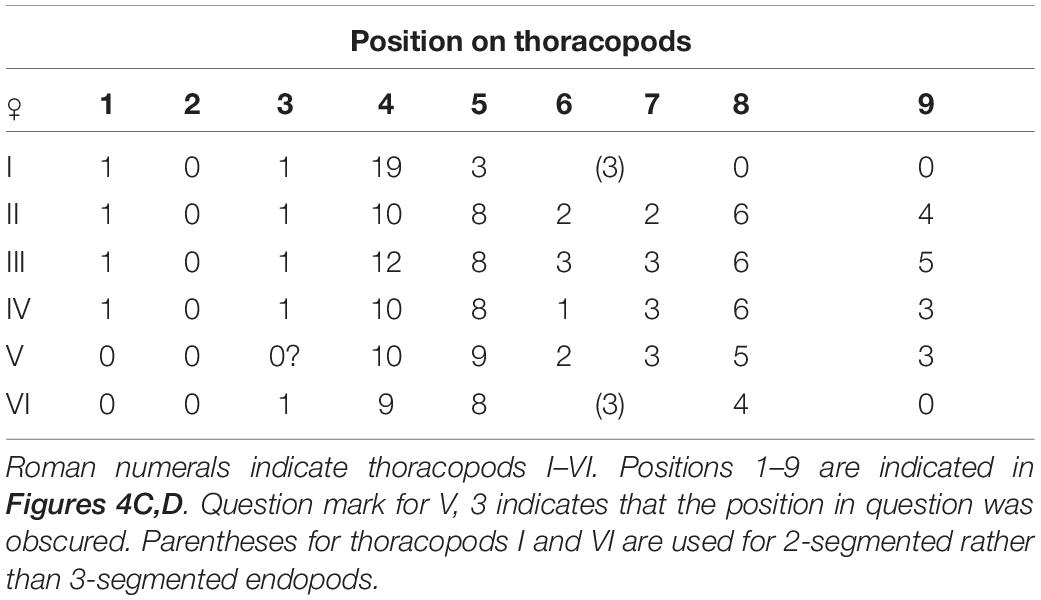
Table 2. Thoracopodal setal counts in Waginella ebonita sp. nov. (ignoring tiny setae).
Abdomen U-shaped, five-segmented, including telson (Figures 2F, 3E, 9A). First segment with vestigial penis on ventral side (Figures 3E,F, 9A,B), an unpaired process about 110 μm long, its tip and lateral sides bearing ctenoid scales (Figure 9B). Dorsal surface of first segment with ctenoid scales and row of dense scales with long setiform denticles along posterior margin (Figure 9A). Second segment trapezoid, about 330 μm long, bigger than either third or fourth, with ctenoid scales on ventral surface. Last body segment (telson) trapezoid, about 300 μm long, its ventral margin bearing rows of ctenoid scales and pair of conspicuous telsonic spines (Figures 3E, 9A,E) approximately 155 μm long with row of numerous, small sharp denticles along their dorsal margins (Figure 9E). Furcal rami blade-like (Figures 3G,H, 9A,E,F), about 500 μm long and 159 μm wide (l/h ratio 3.1); inner surface with rare ctenoid scales; ventral margin with 14–15 large sharp denticles (Figures 3G, 9A,E); row of 24 ctenoid scales with long denticles along dorsal margin covering insertions of 16 medial setae with long fine setules (Figures 3G, 9A,F). Distal margin uneven, with 4 long stout setae grouped into upper and lower pairs (Figures 3G,H); lower and upper setae with short denticles, middle setae with long, fine setules; small but stout plumose seta at base of upper distal seta (Figure 3H).
Lattice Organs
Five bilateral pairs of trough-like lattice organs (Figure 11) situated in 2 groups along hinge line on outer surface of carapace: anterior pairs lo1 and lo2 and posterior pairs lo3, lo4, and lo5 (Figures 11A,B,D). Lattice organs partially hidden by bacteria or other debris. Most of pairs of lattice organs straight, each trough containing crest; cuticle of trough folded, cuticle of crests of all lattice organs strongly wrinkled or plicate, not perforated by small pores (Figures 11C,E–G). Anterior lattice organs situated on small bump with apical large pore just posterior to point of divergence of carapace valves (Figures 11A,B), almost perpendicular to hinge line (70°, converge anteriorly) and parallel to each other (Figure 11B). Lo1 slit-like, 22 μm long and 2.8 μm wide, and located 22 μm from hinge line (Figure 11C). Lo2 16 μm behind lo1, 25 μm long and 3.3 μm wide, slit-like and located 24 μm from hinge line (Figure 11C). Terminal pores of lo1 and lo2 not observed, but morphology of organs may evidence on anterior position (closer to hinge line). Posterior lattice organs situated somewhat anterior to point of divergence of carapace valves 1900 μm behind anterior organs, have different orientations; small bump with apical large pore lies between lo 3 and lo4 closer to hinge line (Figures 11A,D). Lo3 undulating, S-shaped, almost perpendicular to hinge line, 17 μm long and 4 μm wide (Figure 11E), located 83 μm from hinge line, terminal pore not observed, but morphology of organ suggests its position is closer to hinge line. Lo4 slightly undulating and almost parallel to hinge line (converge anteriorly), 32 μm behind lo3, 20.5 μm long and 3 μm wide (Figure 11F), located 76 μm from hinge line, terminal pore not observed, but morphology of organ suggests anterior position. Lo5 68 μm behind lo4 short, straight, almost parallel to hinge line (converge anteriorly), 12.4 μm long and 4 μm wide (Figure 11G), located 78 μm from hinge line, terminal pore rather posterior but hidden by debris.
Associated Fauna
A new species, Waginella ebonita sp. nov., was infested with cryptoniscid isopods (Epicaridea) of an undescribed type (Figures 2C–F). A big brooding female occupied the whole brood chamber of Waginella (Figure 2F, “cf”) and may have consumed the host embryos for feeding (it was not attached by its mouthparts and no embryos of the host were found). Two smaller males were found in posteriodorsal opening of host carapace (Figures 2C–E). These males were apparently trying to enter its mantle cavity to copulate with the female isopod since it was not brooding eggs of its own. Similar parasitic isopods were found in the brood chamber of 4 specimens of W. sandersi (Newman, 1974; Grygier, 1983a). Hyperparasitic cryptoniscid isopods have also been found in association with many other ascothoracidans (Pyefinch, 1939; Newman, 1974; Grygier, 1981b, 1983b,c, 1984, 1985, 1990d; Moyse, 1983; Kolbasov and Petrunina, 2018), although only one species from among them has been described (Grygier, 1981a). Following to Grygier (1991b) we conclude that large female isopods feed on the host embryos and “completely prevent the host’s brood deposition and are therefore parasitic castrators.”
Williams and Bunkley-Williams (2019), when discussing hyperparsitism of ascothoracidans, note that some were considered “symbiotic egg predators” by Kuris et al. (2005). They go on to observe that since these not only damage and feed off their hosts but (rarely) leave them, they should be consider “true parasites,” and with this we agree. However, they also propose a common name for the Ascothoracida; namely, the “copebarnacles” as though they resembled copepods more than some other crustaceans. While having a common name for a group that most carcinologists much less the general public have never seen seem of little utility, but if one were needed we would propose “cypribarnacles” since they have a bivalved carapace like ostracods as well as the so-called “cyprid larvae” of barnacles.
Comparison With Other Congeners
The new species, Waginella ebonita sp. nov. and its Indo-West Pacific congeners, W. cf. axotremata and W. cf. metacrinicola are distinguished from W. sandersi, found in south Atlantic (Newman, 1974; Grygier, 1983a; current data; Table 1) in having (i) angular anterior and posterior ends of carapace (rounded in W. sandersi, Figure 12E), (ii) significantly shorter furcal rami (see Table 1 and Supplementary Figure 2C), (iii) inner surface of furcal rami with 16 medial setae (∼20 in W. sandersi), (iv) ventral 2 terminal setae of furcal rami sit closer (widely spaced in W. sandersi), (v) longer setae on 4th antennular segment (less than 100 μm long in W. sandersi, Supplementary Figure 3C), and (vi) row of dense scales with long setiform denticles on dorsal surface of first abdominal segment. Waginella ebonita sp. nov. differs from W. cf. metacrinicola and W. cf. axotremata (Grygier, 1983a, 1990c; Grygier and Itô, 1995) in having black colored carapace and by a large number of medial setae of furcal rami (Figures 9A,E,F, Supplementary Figures 2E,F, and Table 1). The new species is further from W. cf. metacrinicola by setation of thoracopods (Table 1). Finally, it can be distinguished from W. cf. axotremata by larger carapace, more elongated furcal rami, number of seminal receptacles of thracopods 4, 5, more setae on the second segment of exopod of thoracopod 1 and on third segment of endopod in thoracopods 2–5 (Table 1). A new species attaches to the column of crinoid host (Figures 2A,B), while W. cf. axotremata seems to inhabit cirri of crinoids (Figures 13A–D). And finally Waginella ebonita sp. nov. has been found deeper than W. cf. metacrinicola, and all three Indo-west Pacific species are allopatric (see Table 1).
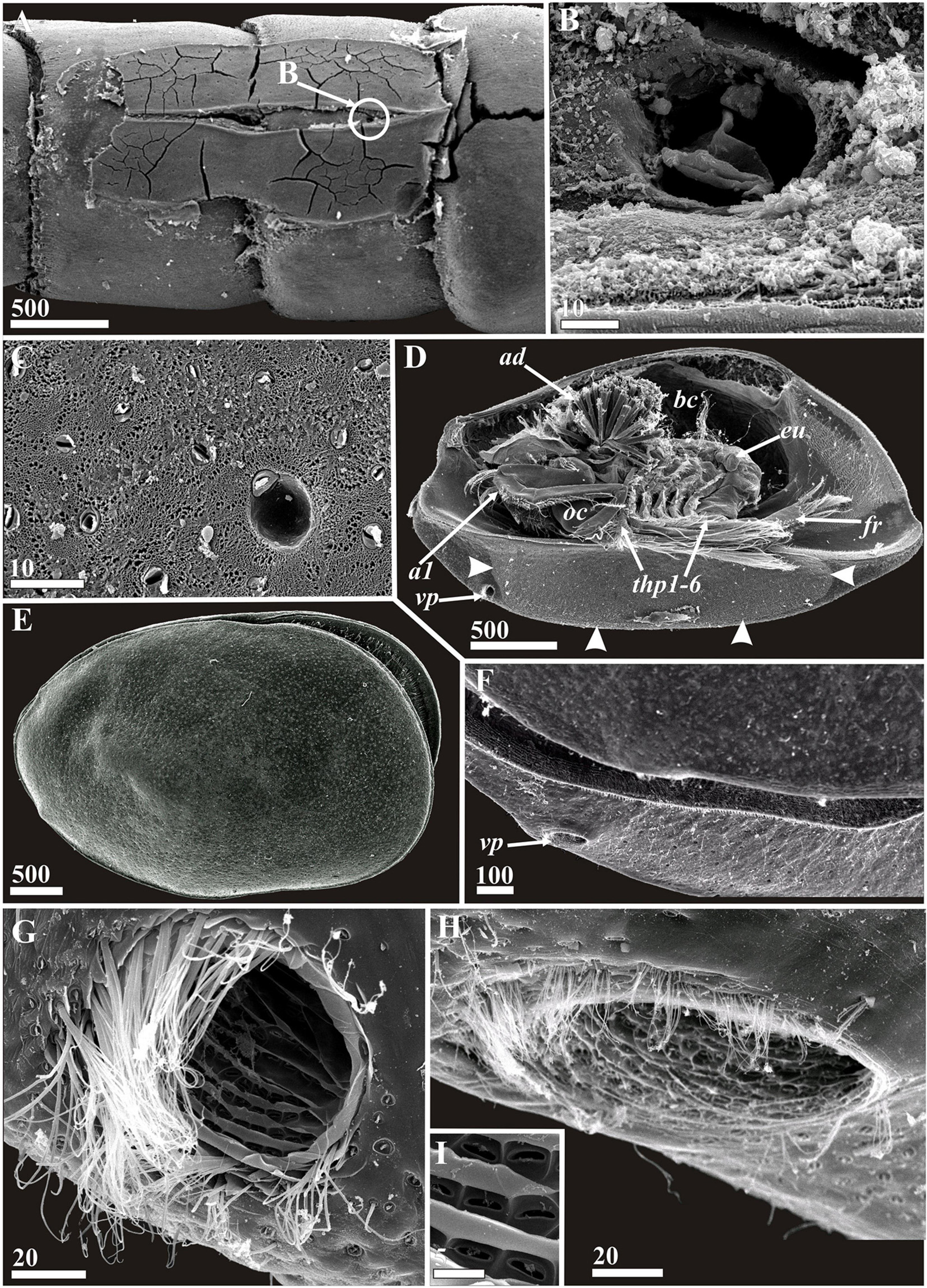
Figure 12. Waginella cf. axotremata (A–D,G,I) and Waginella sandersi (E,F,H), females. Attachment pad, general morphology, carapace structures (SEM). (A) Attachment pad on crinoid host cirrus, anterior end right (location of feeding hole indicated by round outline “B”). (B) Feeding hole burrowed by mouth parts of parasite (presumably mandibles). (C) Surface of cement of attachment pad. (D) General vetrolateral view, left valve of carapace removed, anterior end left (border of ventral area producing attachment cement indicated by arrowheads). (E) General lateral view, anterior end left. (F) Anterioventral part of carapace, anterior end left. (G,H) Anterioventral carapace pores, anterior end left. (I) Enlarged part of inner canal of anterioventral carapace pore. a1, antennule; ad, adductor muscle; bc, brood chamber; eu, epaulet; fr, furcal rami; oc, oral cone; thp1–6, thoracopods 1–6; vp, anterioventral carapace pore. Scale bars in μm.
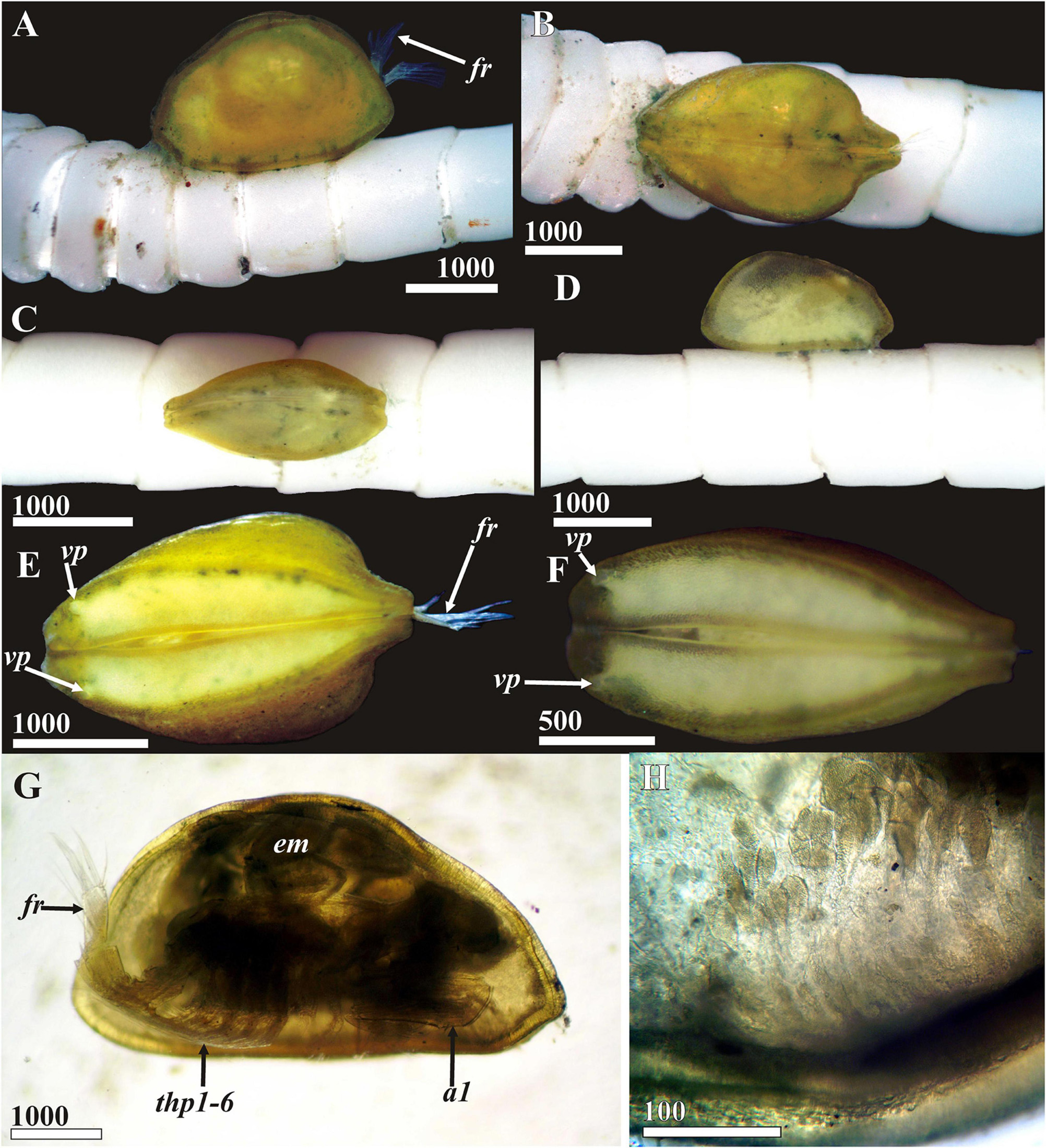
Figure 13. Waginella cf. axotremata. General appearance, light microscopy, female (A,B,E,G,H) and male (C,D,F) (ethanol fixation). (A–D) General view of attached parasite to crinoid cirrus (A,D, lateral side, B,C, dorsal side). (E,F) General view of detached parasite, ventral side. (G) General view lateral, right valve removed. (H) Carapace glands (cement?) of anterioventral side. a1, antennule; em, embryos; fr, furcal rami; thp1–6, thoracopods 1–6; vp, anterioventral carapace pore. Scale bars in μm.
Comparison With the Other Generalized Ascothoracida
Waginella differs from the other generalized ascothoracidan genera Synagoga and Sessilogoga parasitizing in black corals by the following characters, which are all probably synapomorphies: (i, ii) flattened ventral margins of carapace valves with a pair of large external anterior pores; (iii) carapace with numerous pits each with short sensillum inside (gland tube); (iv) fourth antennular segment with two thin dorsal setae not forming a fork; (v) the proximal sensory process of sixth antennular segment inserted close to the claw guard; and (vi) all anterior lattice organs refracted on 60–70° to the hinge line (see Grygier, 1983a; Kolbasov et al., 2020). The species W. sandersi on the other hand “occupies an isolated position with respect to W. metacrinicola and W. axotremata” (Grygier, 1983a). It is characterized by carapace with rounded anterior and posterior ends and less anteroiventral flattening compared to W. cf. axotremata and W. cf. metacrinicola. The shape of carapace of W. sandersi resembles that of the free-swimming Synagoga and may be considered as plesiomorphic (Grygier, 1983a). He also noted the absence in W. sandersi of both long setiform denticles on the dorsal side of first abdominal segment and two vestigial setae of claw guard adjacent to the claw. He concluded that W. sandersi was clearly the plesiomorphic sister group of the Pacific species but did not consider it as a separate genus or subgenus (Grygier, 1983a).
Discussion
Later the fine morphology of W. cf. metacrinicola was examined with SEM (Grygier and Itô, 1995). This study revealed a row of unique characters of carapace. They also studied the morphology of the anterior lattice organs, antennules, frontal filament complex, trunk somites and epaulets (Grygier and Itô, 1995).
During our SEM survey of Waginella we obtained additional information on the fine structure of other three congeners, W. cf. axotremata, W. sandersi, and Waginella ebonita sp. nov. Our data allowed us to compare the morphology of all species of Waginella and establish both relations and differences between them.
Attachment Pads and Cement
The attachment pads of Waginella were studied for the first time (Figures 6, 12A–C, 13H). They are of similar structure in both W. cf axotremata and Waginella ebonita sp. nov. The surfaces of these attachment pads have a spongy texture in which numerous tips of gland tubes are imbedded, and there are rare rounded pits corresponding to the bulbs excreting some substance via gland tubes (Figures 6C,D, 13C). The cement fibers form a clear pattern with numerous hollows corresponding to cuticular villi on the attachment side of carapace (Figure 12C). The presence of imbedded tips of gland tubes in attachment pad is evidence that the parasite is glued permanently to the crinoid host, at least until the next molt, or perhaps by dissolution of this cement pad.
A small rounded feeding hole bored by rasping mandibles and corresponding in diameter to the tip of oral cone always lies between the halves of attachment pad (Figures 6A,B, 12A,B). Its form and small size (∼30–45 μm in diameter) suggest feeding on the coelomic fluid of a host.
Groups of ampuliform structures with a granulated content resembling unicellular glands were observed in the ventral attachment zone of carapace (Figure 13H). These structures may be the cement glands, if not glands secreting a cement dissolving substance.
Carapace Ornamentation
Ornamentation of carapace is similar in all species of Waginella (Figures 7, 8, 11, 12D–I and Supplementary Figures 1A,B,E–I, 4, 5). The whole external surface of carapace is covered with numerous pitted glandular tubes and short rare setae, which are denser on ventral side (Figures 7D,E, 11C,D and Supplementary Figures 1A,B, 4A,D, 5A,D; see Grygier and Itô, 1995 for W. cf. metacrinicola). Similar glandular tubes are found in the interior of the carapace, along the ventral margin (Figures 8B–D and Supplementary Figures 1E,F). The ventral side has a carpet of cuticular villi especially denser in attachment site (Figures 7D,E and Supplementary Figures 1A,B), where sets of longer villi surround numerous tiny ones and gland tubes. These cuticular structures represent synapomorphies of the genus Waginella. Although seems in other Ascothoracida curiously they are absent in Synagoga and Sessilogoga which have smooth exterior carapaces with rare, minute pores (Kolbasov and Newman, 2018; Kolbasov et al., 2019, 2020).
The anatomy of gland tubes and cuticular villi remains unknown so we can only speculate on their function, such as secreting some substance on at least the ventral side of carapace (Figure 7D and Supplementary Figure 1B). It is tempting to assume these structures to be cement glands, but their position on the whole carapace makes that unlikely. Their secretions may dissolve cement and serve for disattachment of the parasite from host and several lacunae on the attachment pad situated below some of gland tubes may indicate of this function (Figures 6C,D, 12C).
Similar papilliform structures (opercular papillae) associated with burrowing apparatus were reported in many acrothoracican species that apparently secrete a substance softens and dissolves the carbonate substrate of their hosts (Kolbasov, 2009). On the other hand, the cuticular villi found in Waginella are similar to cuticular projections on the attachment disk of the acrothoracicans (Kolbasov, 2009) and we consider both as secretory structures for cement that holds them in place.
All species of Waginella possess big anterioventral pores having external and internal parts with an identical morphology (Figures 7C, 12G–I; see Grygier and Itô, 1995 for W. cf. metacrinicola). The surface of the pore canal has lamellar ridges forming a lattice pattern, where each square has one small oval pore. The external rim of pore is guarded by tufts of fine setae, shorter in W. sandersi in comparison with other congeners.
Inner surface of carapace has similar ornamentation in all species of Waginella (Figure 8 and Supplementary Figures 1E–I; see Grygier and Itô, 1995 for W. cf. metacrinicola). An array of different scales, denticles, gland tubes and pores are spread along the ventral margin of carapace (see Description herein). The pattern of distribution of main cuticular structures of the carapace is similar in all congeners. Its armament is better developed in Waginella than in Synagoga and Sessilogoga having setae, small pores and underlying ctenae of lesser density.
Both sexes of Waginella, as in many others ascothoracidans, possess frontal filament complex (Figure 7F and Supplementary Figures 1C,D; see Grygier and Itô, 1995 for W. cf. metacrinicola). It is tripartite and better developed in males with anterior ramus, longer posterior ramus and short, rounded basal process (Supplementary Figures 1C). Only the posterior ramus is developed in females, the other two parts being vestigial. This ramus in W. sandersi (Supplementary Figure 1D) has extremely densely packed numerous setiform projections in comparison with other three Indo-Pacific species. Such a unity of carapace ornamentation in all species of Waginella supports its monophyly.
Body Morphology
Besides identical carapace ornamentation, all species of the genus Waginella possess well developed epaulets of similar form and morphology on sixth thoracic somite (Figures 9C,D and Supplementary Figures 2A,B; see Grygier and Itô, 1995 for W. cf. metacrinicola). Each epaulet consists of stem-like proximal part with a smooth cuticle, which may be perforated by a few small pores in W. cf. metacrinicola and W. cf. axotremata (Supplementary Figure 5A) before swelling into a rounded distal part divided by fine cuticular furrows into polygonal knobs with fine, spherical plaques.
Grygier and Itô (1995) suggested the epaulets possibly functioned as respiratory organs or gills but there has been no substantion since. From 23 ascothoracidan genera they present in 7 genera of generalized Synagogidae (except Sessilogoga) and in several species of the genus Ascothorax – most generalized taxon of the order Dendrogastrida. These structures are absent in advanced endoparasitic forms and reduction of epaulets is an apomorphic condition for the Ascothoracida.
In spite of different ratios (l/h), furcal rami of all species of Waginella possess a similar morphology (Figures 9A,E,F and Supplementary Figures 2C–F), e.g., a transverse row of long medial setae on inner surface (8–20) as does Synagoga, 4 long setae on distal margin and the absence of ventral setae.
Antennules of all species of Waginella have similar form, morphology and ornamentation (Figure 10 and Supplementary Figure 3; see Grygier and Itô, 1995 for W. cf. metacrinicola). Sixth segment is significantly shorter than fifth and movable claw cannot reach fourth segment to form a lock with its setae that has apparently led to their reduction, since the movable claw reaches fourth antennular segment in both Synagoga and Sessilogoga and two setae of this segment, forming a lock with claw, are stout and spiniform. A rudimentary proximal sensory process is always present at the base of claw guard in all species of Waginella. The ultrastructure of antennules in W. sandersi differs slightly from that in other congeners: (i) first segment with dense ctenoid scales and lacking setae (Supplementary Figure 3A) and (ii) claw guard apparently having but 2 long setae, without 2 vestigial setae of ctenoid hood.
Lattice Organs
Lattice organs of all representatives of Waginella are characterized by some similarities in morphology, but their form generally differs in congeners (Figure 11 and Supplementary Figures 4, 5; see Grygier and Itô, 1995 for W. cf. metacrinicola). All their lattice organs have a distinct crest in a deep trough. Each trough has folded or wrinkled cuticle (Figures 11E–G and Supplementary Figures 4B,C,E–G), but the cuticle of trough in W. sandersi is without distinct folders (Supplementary Figures 5B,C,E–G). In all species of Waginella the cuticle of crests of the lattice organs is strongly wrinkled or plicated, and without distinct terminal pores. Such unique crests and troughs are found only in Waginella but not in other Thecostraca. Moreover the anterior lattice organs (lo1, lo2) in all species of Waginella are situated on noticeable or small hump with apical large pore, they are almost perpendicular to hinge line (60–70°) and converge anteriorly. Distal edges of crests of lo1 and lo2 are oriented anteriorly and closer to the hinge line in all species of the genus (Figures 11A,B and Supplementary Figures 4A,B, 5A,B). Posterior lattice organs in Waginella differ in orientation and form in congeners (Figure 11D and Supplementary Figures 4D, 5D) albeit Grygier and Itô (1995) did not provide information about orientation and structure of lo3–5 in W. cf. metacrinicola). Lo3 and lo4 are located near the large pore, often on the small hump.
All posterior lattice organs are straight in W. sandersi (Supplementary Figures 5D–G), distal end of crest of lo5 is posterior, but the orientation of distal/terminal end in other organs is unclear. In both W. cf. axotremata and Waginella ebonita sp. nov. crest of lo5 is also straight, with posterior distal end bearing putative terminal pore (Figure 11G and Supplementary Figure 4G); while lo3 and lo4 are S-shaped or undulating (Figures 11F,G and Supplementary Figures 4E,F). While lo3 in both species are S-shaped and have the distal end turned to the hinge line, it was not possible to determine anterior or posterior orientation for this organ due to its perpendicular orientation (Figure 11E and Supplementary Figure 4E). Lo4 is undulating, with anterior terminal end of the crest (Figure 11F and Supplementary Figure 4F).
The orientation, structure and their form of the lattice organs (especially anterior pairs) varies significantly within the adult Ascothoracida. While all co-linear in Synagoga (Grygier and Ohtsuka, 1995; Kolbasov and Newman, 2018; Kolbasov et al., 2019), the anterior lattice organs are almost perpendicular to the hinge line in both Sessilogoga (lo1 in female both lo1 and lo2 in male) and Waginella (Kolbasov et al., 2020, herein). This orientation of the anterior lattice organs is considered an apomorphy of these genera (Kolbasov et al., 2020).
The orientation and form of the lattice organs differ not only among thecostracan taxa but also in various pairs within the same species. Although a fully co-linear arrangement, as well as posterior terminal pores (= distal ends) of all lattice organs has been inferred to be plesiomorphic not only for ascothoracidans but also for all thecostracans (Jensen et al., 1994; Høeg and Kolbasov, 2002; Celis et al., 2008; Kolbasov and Newman, 2018), the present findings could strengthen Grygier and Ohtsuka’s (1995) and Kolbasov et al. (2019, 2020) contention that the configuration of lattice organs in ascothoracidans is not constant. Actually, such a plesiomorphic orientation (as well as a structure) of the lattice organs was inferred only for the ascothoracid larva of Ulophysema (Jensen et al., 1994) in comparison with the lattice organs of other cypridiform larvae of Thesostraca, e.g., the forms where these organs first appeared. A similar plesiomorphic structure of the lattice organs was discovered in the cypridiform larvae of Facetotecta (Høeg and Kolbasov, 2002; own data).
Lattice organs have been described for many adults of the Ascothoracida having a carapace (Grygier and Itô, 1995; Grygier and Ohtsuka, 1995; Kolbasov et al., 2008, 2019, 2020; Kolbasov and Petrunina, 2018; Kolbasov and Newman, 2018) but only three times for the ascothoracid larvae (Itô and Grygier, 1990; Jensen et al., 1994; Kolbasov et al., 2008). Kolbasov et al. (2008) studied modified lattice organs of the second ascothoracid larva of Dendrogaster spp., a highly modified ascothoracidan, having only 4 pairs of lattice organs. They showed that morphology and orientation of the lattice organs can change after molting into an adult male. On the other hand, the ascothoracid or Tessmann larva of Baccalaureus possesses all 5 pairs of lattice organs which are plesiomorphic in structure including posterior terminal pores (Itô and Grygier, 1990; own unpublished data). While its dwarf male retains all 5 pairs of lattice organs, their position changes from co-linear to perpendicular in all pairs (own unpublished data). This fact supports the hypothesis that in Thecostraca the morphology of lattice organs in homologous stages (e.g., cypridiform larvae for all taxa) can be used in phylogenetic reconstructions.
Monophyly of the Genus Waginella and Species Recognition
Fine morphology also supports monophyly of Waginella. To the six morphological characters mentioned earlier uniting Waginella the following can be added: (i) all species possess a unique attachment apparatus consisting of gland tubes and cuticular villi on the ventral side of carapace; (ii) large anterioventral pores having the same structure in all species; (iii) the distal part of the epaulets being covered by polygonal knobs with fine, spherical plaques; (iv) furcal rami without ventral setae; (v) sixth antennular segment significantly shorter than fifth; and (vi) cuticle of crests of all lattice organs strongly wrinkled or plicated.
While the Indo-Pacific species of Waginella appear have no distinct distinguishing characters (see Table 1), the South Atlantic W. sandersi forms a distinct morphospecies. The type locality for W. metacrinicola is Japan and Luzon Island (Philippines), whereas W. axotremata is from Indonesia (Okada, 1926; Grygier, 1983a). Subsequently many specimens of Waginella were found in different localities from Indo-West Pacific, but in spite of several differences in setation of thoracopods, antennules and furcal rami, and location on hosts that may indicate different species, most were assigned into W. axotremata albeit most tentatively (Grygier, 1990c; Grygier and Itô, 1995). Grygier and Itô (1995) studied W. metacrinicola and W. axotremata and concluded that only “… the size and shape of the carapace of brooding females remain the most useful taxonomic characters to distinguish these two nominal species.” Species of other generalized synagogids, Synagoga and Sessilogoga, also have very similar morphology of congeners that can only be distinguished by such taxonomic characters as setation of antennules, furcal rami and thoracopods (Kolbasov and Newman, 2018; Kolbasov et al., 2019, 2020). Thus, two species of Synagoga living in remote locations, S. arabesque from Taiwan can be distinguished from S. grygieri from Macaronesia only by fewer setae on the fifth antennular segment of males and more setae on the inner side of the furcal ramus of females. Thus, it appears that W. axotremata and probably W. metacrinicola represent species complexes that need to be revised morphologically, and if possible with genetic confirmation.
Summary and Outlook
We have shown that genus Waginella represents a monophyletic taxon including at least 4 species having similar and unique morphology even on the ultrastructural level (e.g., structure of the lattice organs and attachment apparatus) albeit the Atlantic morphotype is distinct form that of the Indo-Pacific. Presence of an attachment apparatus in all species of the genus suggests that they remain attached to the host for a considerable period of time, at least until the next molt or dissolution of attachment cement by a special secretory apparatus for which we present some evidence. Two species, W. cf. axotremata and W. cf. metacrinicola may represent at least two complexes of species. The lattice organs of Waginella have a unique structure (cuticle of crests of all lattice organs is strongly wrinkled) distinguishing from all Thecostraca. And finally, it appears that only the morphology of lattice organs in homologous stages (e.g., cypridiform larvae) can currently be used in the phylogenetic reconstructions of Thecostraca.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: urn:lsid:zoobank.org:pub:49C48116-770E-4073-BE53-02DF5AE66761 and urn:lsid:zoobank.org:act:8F743E91-F94B-4044-8A92-5DD81315888D.
Author Contributions
GK and WN identified and examined the specimens. All authors involved in writing of the manuscript, contributed to the article, and approved the submitted version.
Funding
GK and AS were financially supported by the Russian Foundation for Basic Research (grant 21-54-52003 MNT_a). BC was supported by the Taiwan Russia bilateral grants from Ministry of Science and Technology, Taiwan (MOST-110-2923-B-001-003-MY3).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
Two referees are thanked for very constructive comments and criticism. We thank the collaborators of the Laboratory of Electronic Microscopy of Moscow State University for assistance in SEM studies.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2021.616001/full#supplementary-material
References
Celis, A., Alvarez, F., Newman, W. A., and Sumida, P. Y. G. (2008). Descriptions and phylogenetic significance of the fronto-lateral gland pores and dorsal lattice organs of cyprid larvae of seven species of barnacles (Cirripedia: thoracica: pedunculata). J. Crust. Biol. 28, 203–215. doi: 10.1163/20021975-99990365
Chan, B. K. K., Gale, A. S., Glenner, H., Dreyer, N., Ewers Saucedo, C., Péres-Losada, M., et al. (2021). The Evolutionary Diversity of the Barnacles with an Updated Classification of fossil and living forms. Zool. J. Linn. Soc. zlaa160, 1–58.
de Lacaze-Duthiers, H. (1880). Histoire de la Laura gerardiæ, type nouveau de Crustacé parasite. Archs. Zool. Exp. Gen. 8, 537–581.
Grygier, M. J. (1981a). Redescription of Gorgonolaureus bikiniensis (Crustacea: ascothoracica), with a reevaluation of its familial affinities. Micronesica 17, 67–76.
Grygier, M. J. (1981b). Gorgonolaureus muzikae sp. nov. (Crustacea: ascothoracida) parasitic on a Hawaiian gorgonian, with special reference to its protandric hermaphroditism. J. Nat. Hist. 15, 1019–1045. doi: 10.1080/00222938100770771
Grygier, M. J. (1983a). Revision of Synagoga (Crustacea: maxillopoda: ascothoracida). J. Nat. Hist. 17, 213–239. doi: 10.1080/00222938300770181
Grygier, M. J. (1983b). Ctenosculum hawaiiense Heath: confirmation of its affinities (Crustacea: ascothoracida - ex Mollusca: gastropoda). J. Crust. Biol. 3, 257–265. doi: 10.2307/1548261
Grygier, M. J. (1983c). Introcornia conjugans n. gen. n. sp., parasitic in a Japanese ahermatypic coral (Crustacea: ascothoracida: petrarcidae). Senck. Biol. 63, 419–426.
Grygier, M. J. (1984). Ascothoracida (Crustacea: maxillopoda) parasitic on Chrysogorgia (Gorgonacea) in the Pacific and western Atlantic. Bull. Mar. Sci. 34, 141–169.
Grygier, M. J. (1985). Lauridae: taxonomy and morphology of ascothoracid crustacean parasites of zoanthids. Bull. Mar. Sci. 36, 278–303.
Grygier, M. J. (1987a). Nauplii, antennular ontogeny and the position of the Ascothoracida within the Maxillopoda. J. Crust. Biol. 7, 87–104. doi: 10.1163/193724087X00072
Grygier, M. J. (1987b). Reappraisal of sex determination in the Ascothoracida. Crustaceana 52, 149–162. doi: 10.1163/156854087X00196
Grygier, M. J. (1990a). A crustacean endoparasite (Ascothoracida: synagogidae) of an antipatharian from Guam. Micronesica 23, 15–25.
Grygier, M. J. (1990b). Early planktotrophic nauplii of Baccalaureus and Zibrowia (Crustacea: ascothoracida) from Okinawa, Japan. Galaxea 8, 321–337.
Grygier, M. J. (1990c). New records of Waginella (Crustacea: ascothoracida) ectoparasitic on stalked crinoids from Japan and Australasia. Galaxea 8, 339–350.
Grygier, M. J. (1990d). Five new species of bathyal Atlantic Ascothoracida (Crustacea: maxillopoda) from the equator to 50°N latitude. Bull. Mar. Sci. 46, 655–676.
Grygier, M. J. (1991a). Additions to the ascothoracidan fauna of Australia and South-east Asia (Crustacea, Maxillopoda): synagogidae (part), Lauridae and Petrarcidae. Rec. Austr. Mus. 43, 1–46. doi: 10.3853/j.0067-1975.43.1991.39
Grygier, M. J. (1991b). Redescription, ontogeny, and demography of Parascothorax synagogoides (Crustacea: ascothoracida), parasitic on Ophiophthalmus normani, a dominant ophiuroid in the bathyal basins off southern California. Proc. San Diego Soc. Nat. Hist. 6, 1–20.
Grygier, M. J. (1996). “Sous-classe des Ascothoracides (Ascothoracida Lacaze-Duthiers, 1880)”in Traité de Zoologie: Anatomie, Systématique, Biologie. Tome VII: Crustacés, Fascicule 2: Généralités (suite) et Systématique. ed. J. Forest (Paris: Masson) 433–452.
Grygier, M. J., and Fratt, D. B. (1984). The ascothoracid crustacean Ascothorax gigas: redescription, larval development, and notes on its infestation of the Antarctic ophiuroid Ophionotus victoriae. Antarct. Res. Ser. 41, 43–58.
Grygier, M. J., and Itô, T. (1995). “SEM-based morphology and new host and distribution records of Waginella (Ascothoracida)”in New Frontiers in Barnacle Evolution (Crustacean Issues 10), eds F. R. Schram and J. T. Høeg (Rotterdam :A.A. Balkema) 209–228.
Grygier, M. J., and Ohtsuka, S. (1995). New species of Synagoga (Crustacea: ascothoracida) from plankton off Okinawa, Japan, with an SEM study of the carapace. Publ. Seto Mar. Biol. Lab. 36, 293–311. doi: 10.5134/176250
Høeg, J. T., and Kolbasov, G. A. (2002). Lattice organs in y-cyprids of the Facetotecta and their significance in the phylogeny of the Crustacea Thecostraca. Acta Zool. 83, 67–79. doi: 10.1046/j.0001-7272.2001.00100.x
Høeg, J. T., Chan, B. K. K., Kolbasov, G. A., and Grygier, M. J. (2014). “Ascothoracida” in Atlas of Crustacean Larvae eds J. W. Martin, J. Olesen, and J. T. Høeg (Baltimore: Johns Hopkins University Press) 104–106.
Itô, T., and Grygier, M. J. (1990). Description and complete larval development of a new species of Baccalaureus (Crustacea: ascothoracida) parasitic in a zoanthid from Tanabe Bay, Honshu, Japan. Zool. Sci. 7, 485–515.
Jensen, P. G., Moyse, J., Høeg, J., and Al-Yahya, H. (1994). Comparative SEM studies of lattice organs: putative sensory structures on the carapace of larvae from Ascothoracida and Cirripedia (Crustacea Maxillopoda Thecostraca). Acta. Zool. 75, 125–142. doi: 10.1111/j.1463-6395.1994.tb01117.x
Kolbasov, G. A. (2007). The external morphology of parasitic crustaceans of the genus Dendrogaster (Thecostraca, Ascothoracida). Zool. Z. 86, 654–670.
Kolbasov, G. A. (2009). Acrothoracica, burrowing crustaceans. Moscow: KMK Scientific Press Ltd. 452.
Kolbasov, G. A., and Newman, W. A. (2018). A new species of Synagoga (Crustacea: thecostraca: ascothoracida) parasitic on an antipatharian from the Azores and Cape Verde Islands, with notes on its morphology, sexuality, host specificity, and biogeography. Mar. Biodivers. 49, 1101–1122. doi: 10.1007/s12526-018-0892-7
Kolbasov, G. A., and Petrunina, A. S. (2018). The family Ascothoracidae Grygier, 1987, a review with descriptions of new abyssal taxa parasitizing ophiuroids and remarks on the invalidity of the genus Parascothorax Wagin, 1964 (Crustacea: thecostraca: ascothoracida). Mar. Biodivers. 49, 1417–1447. doi: 10.1007/s12526-018-0921-6
Kolbasov, G. A., Grygier, M. J., Høeg, J. T., and Klepal, W. (2008). External morphology of ascothoracid-larvae of the genus Dendrogaster (Crustacea, Thecostraca, Ascothoracida), with remarks on the ontogeny of the lattice organs. Zool. Anz. 247159–183. doi: 10.1016/j.jcz.2007.07.007
Kolbasov, G. A., Petrunina, A. S., Ho, M. J., and Chan, B. K. K. (2019). A new species of Synagoga (Crustacea, Thecostraca, Ascothoracida) parasitic in an antipatharian from Green Island, Taiwan, with notes on its morphology. ZooKeys 87655–85. doi: 10.3897/zookeys.876.35443
Kolbasov, G. A., Petrunina, A. S., Olesen, J., Ho, M. J., Chan, B. K. K., and Grygier, M. J. (2020). A new species of Sessilogoga Grygier, 1990 parasitic in an antipatharian from Green Island, Taiwan, with notes on its nauplius larvae and the synapomorphies and apparent gonochorism of the genus (Crustacea: thecostraca: ascothoracida). Mar. Biodivers. 50, 1–29. doi: 10.1007/s12526-020-01062-y
Kuris, A. M., Lafferty, K. D., and Torchin, M. E. (2005). “Biological control of the European green crab, Carcinus maenas: natural enemy evaluation and analysis of host specificity” in Second international symposium on biological control of arthropods. (Washington, D.C: USDA Forest Service). 102–115.
Moyse, J. (1983). Isidascus bassindelei gen. nov., sp. nov. (Ascothoracida: crustacea) from north-east Atlantic with a note on the origin of barnacles. J. Mar. Biol. Assoc. U. K. 63, 161–180.
Newman, W. A. (1974). Two new deepdsea Cirripedia (Ascothoracica and Acrothoracica) from the Atlantic. J. Mar. Biol. Assoc. U. K. 54, 437–456. doi: 10.1017/S002531540005863X
Norman, A. M. (1888). “Report on the occupation of the Table” in Report of the Fifth-Seventh Meeting of the British Association for the Advancement of Science Held at Manchester in August and September 1887, London. 85–86.
Norman, A. M. (1913). Synagoga mira, a crustacean of the order Ascothoracica. Trans. Linn. Soc. Lond. 2nd Ser. Zool. 11, 161–166. doi: 10.1111/j.1096-3642.1913.tb00191.x
Okada, Y. K. (1926). Contribution à l’étude des Cirripèdes Ascothoraciques. II. Note sur l’organisation de Synagoga. Bull. Mus. Natl. Hist. Nat. 32, 69–73.
Pyefinch, K. A. (1939). Ascothoracica (Crustacea, Cirripedia). The John Murray Expedition, 1933-34. Sci. Rep. 5, 247–262.
Keywords: parasitic crustaceans, Ascothoracida, taxonomy, SEM, ultrastructure, lattice organs, distribution
Citation: Kolbasov GA, Savchenko AS, Newman WA and Chan BKK (2021) A New Species of Waginella (Crustacea: Thecostraca: Ascothoracida) Parasitic on a Stalked Crinoid From Tasman Sea, With Notes on Morphology of Related Genera. Front. Mar. Sci. 8:616001. doi: 10.3389/fmars.2021.616001
Received: 10 October 2020; Accepted: 06 May 2021;
Published: 09 June 2021.
Edited by:
Michel Roux, Muséum National d’Histoire Naturelle, FranceReviewed by:
Maria Pia Miglietta, Texas A&M University at Galveston, United StatesKaori Wakabayashi, Hiroshima University, Japan
Copyright © 2021 Kolbasov, Savchenko, Newman and Chan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Benny K. K. Chan, chankk@gate.sinica.edu.tw
†Deceased