 Haakon Hop1*
Haakon Hop1* Anette Wold1
Anette Wold1 Amelie Meyer1,2,3
Amelie Meyer1,2,3 Allison Bailey1
Allison Bailey1 Maja Hatlebakk4Slawomir Kwasniewski5
Maja Hatlebakk4Slawomir Kwasniewski5 Peter Leopold1
Peter Leopold1 Piotr Kuklinski5
Piotr Kuklinski5 Janne E. Søreide4
Janne E. Søreide4- 1Norwegian Polar Institute, Fram Centre, Tromsø, Norway
- 2Institute for Marine and Antarctic Studies, University of Tasmania, Hobart, TAS, Australia
- 3Australian Research Council Centre of Excellence for Climate Extremes, University of Tasmania, Hobart, TAS, Australia
- 4University Centre in Svalbard, Longyearbyen, Norway
- 5Institute of Oceanology, Polish Academy of Sciences, Sopot, Poland
The impact of the rapidly changing Arctic on zooplankton community structure and seasonal behaviour is not yet understood. Here we examine 6 months of under-ice zooplankton observations from the N-ICE2015 expedition (January to June 2015) in the Nansen Basin and on the Yermak Plateau north of Svalbard. Stratified sampling in the water column was done with MultiNet during the entire expedition, and sampling in the upper 5 m below sea ice was performed during April-May by divers using a hand-held net. Hydrographic conditions were dominated by northward-flowing warm and saline Atlantic Water at intermediate depth, and southward-flowing cold Polar Surface Water in the upper 100 m. The mesozooplankton was dominated by copepods. Most numerous were the small ubiquitous Oithona similis in the upper 200 m, with Microcalanus spp. and Triconia borealis further down the water column. Calanus finmarchicus dominated among the Calanus species while Metridia longa was also numerous. The most abundant deep-water copepods were Paraeuchaeta spp. and Spinocalanus spp. Arrow worms (Chaetognatha) and comb jellies (Ctenophora) were the most numerous non-copepods. The mesozooplankton community was more dependent on surrounding water mass characteristics, such as salinity and depth, than geographical location. Algal food availability, which was closely linked to seasonality, explained the community changes seen in surface waters in May and June due to seasonal ascent and recruitment. Seasonal changes from winter to spring mostly involved an increase in the herbivorous C. finmarchicus and its nauplii in the upper 200 m of the water column coinciding with the peak of the phytoplankton bloom in late May. The Yermak Plateau and adjacent Nansen Basin were characterised by oceanic North Atlantic and Arctic species, many of which are deep water specialists. Despite the late onset of the spring bloom due to consolidated sea ice, both North Atlantic and Arctic species successfully reproduced in the study area. This explains the species-rich mesozooplankton community in this region as opposed to the less productive central Arctic Ocean. Future prospects of less sea ice and earlier onset of the bloom will likely be positive for the overall secondary production by both Arctic and boreal zooplankton in this region.
Introduction
One of the main factors influencing the zooplankton community at high latitudes is strong seasonality. At 80°–82°N, the period of polar night without sunlight lasts for up to 4 months, although marine organisms may experience darkness for longer time depending on sea ice and snow cover (Cohen et al., 2020). The polar night is characterized by primary production close to zero, which is followed by a short and intense primary production season when the sun returns and the ice melts in the spring. Organisms living at high latitudes have adapted to the extreme seasonality and shortage of food (Conover and Huntley, 1991; Hagen and Auel, 2001). Diapause or reduced activity and metabolism during winter, combined with storing large lipid reserves are important adaptations of Arctic zooplankton (Varpe et al., 2009). The strong seasonality in primary production results in a short and pulsed appearance of the large, predominately herbivorous calanoid copepods in surface waters during the spring (Falk-Petersen et al., 2009). However, the annual timing of the copepod ascent from diapause differs from year to year and among regions (Daase et al., 2013). Further, depending on the bottom depth and closeness to land, massive outbursts of meroplankton may occur at the onset of the spring bloom or shortly after (Stübner et al., 2016). The Arctic Ocean is poorly explored since winter darkness, weather and sea ice conditions make it logistically challenging to access without expensive infrastructure. Most zooplankton studies in the Arctic Ocean are therefore performed in late summer-autumn when sea ice is at its minimum (Conover and Huntley, 1991; Conover and Siferd, 1993; Auel and Hagen, 2002; Kosobokova and Pertsova, 2005; Daase et al., 2008; Søreide et al., 2010; Hirche and Kosobokova, 2011; Freese et al., 2016; Ershova et al., 2021). Knowledge about the mesozooplankton community structure and development during the winter-spring period at the main entrance to the Arctic Ocean proper is particularly needed to investigate the expansion of boreal zooplankton into the Arctic Ocean (Wassmann et al., 2015).
Mesozooplankton studies from the Yermak Plateau and adjacent Nansen basin show that this area is rich in mesozooplankton compared to the other Arctic Ocean basins and is characterised by a mix of boreal/North Atlantic and Arctic species as well as deep-water specialists (Kosobokova et al., 1998; Mumm et al., 1998; Auel and Hagen, 2002; Hirche and Kosobokova, 2007). Calanus copepods, the dominant contributor to mesozooplankton biomass in this region and elsewhere in the Arctic Ocean (Ashjian et al., 2003; Ershova et al., 2021), descend to depth during the autumn after accumulating sufficient energy for overwintering (Falk-Petersen et al., 2009; Daase et al., 2013). They remain at depth (>300 m) during the dark winter and ascend in the spring (Darnis and Fortier, 2014), probably in response to increased light levels below the sea ice (Kowalczuk et al., 2017; Pavlov et al., 2017) or depletion of lipid reserves (Irigoien, 2004). Internal biological clocks, such as circadian clocks, can also play a major role in termination of diapause (Häfker et al., 2018). A mismatch between the algae bloom and presence of Calanus spp. in surface waters might reduce their reproductive success and subsequent survival of offspring (Leu et al., 2011; Daase et al., 2013). The seasonal migratory behaviour and physiological state of the zooplankton in the spring are important biological factors in this ecosystem because much of the energy flow in the pelagic system relies on the reproductive success of Calanus spp. and their subsequent population growth (Falk-Petersen et al., 2007). Plankton (autotrophs and heterotrophs) are the producers and the first-order consumers of the marine food webs and they respond rapidly to environmental changes with potential cascading trophic implications for the marine ecosystem (Falk-Petersen et al., 2007; Blachowiak-Samolyk et al., 2008; Wassmann and Reigstad, 2011; Darnis et al., 2012; Hop et al., 2019b). The Arctic is rapidly changing; the thicker multiyear ice cover is turning into a thinner and largely seasonal first-year ice cover and there is an increase in temperature of inflowing Atlantic Water (Beszczynska-Möller et al., 2012). These changes will likely impact the Arctic primary production and, hence, also the secondary production and the energy flow through the system (Polyakov et al., 2020).
Data collected during the N-ICE2015 expedition (The Norwegian young sea ICE expedition), a 6-month drift with the sea ice in the Arctic Ocean with a research vessel in 2015 (Granskog et al., 2018), provided a unique opportunity to study the distribution and development of zooplankton during the winter-spring period below sea ice. The N-ICE2015 time series of zooplankton community composition is the first of its kind from the dynamic Atlantic sector of the Arctic Ocean. Even though we understand many physiological processes of the Arctic zooplankton community, especially those related to migration, we still have insufficient knowledge of the links to environmental variables. Our study follows the natural course of events over seasons in the same year, while gaining spatial information from the drift tracks. This approach enables us to unravel some of the factors driving zooplankton behaviour and strategies in a rapidly changing region of the Arctic Ocean. Here we describe the zooplankton community along with changing environmental conditions in the Atlantic sector of the Arctic. We determined the vertical and horizontal distribution of zooplankton in the Nansen Basin and the Yermak Plateau area in order to assess the role of seasonal and oceanographic variability on the mesozooplankton community development during the winter-spring period. Winter-spring transition is the most critical time in zooplankton life history since this is, for many species, the main mating and reproduction season which is energy demanding and sensitive to individual’s energy status, and therefore drives fitness. Of particular interest is the temporal match of zooplankton’s presence in the surface waters and blooms of ice algae and phytoplankton, after the long and food-poor winter, as well as assessing the accumulated lipid stores and timing of reproduction.
Materials and Methods
N-ICE2015 Expedition and Area Description
The study was part of the N-ICE2015 expedition onboard R/V Lance between January and June 2015. The expedition completed four drifts in the Arctic Ocean north of Svalbard, during which the ship was moored each time to a sea ice floe (Figure 1). On each floe, an ice camp was set up and biogeochemical, oceanographic, atmospheric, sea ice, and snow data were collected (Granskog et al., 2016). For the purpose of marine ecological studies, samples of zooplankton, phytoplankton and ice algae (Assmy et al., 2017; Olsen et al., 2017; Fernández-Méndez et al., 2018) were collected throughout the period.
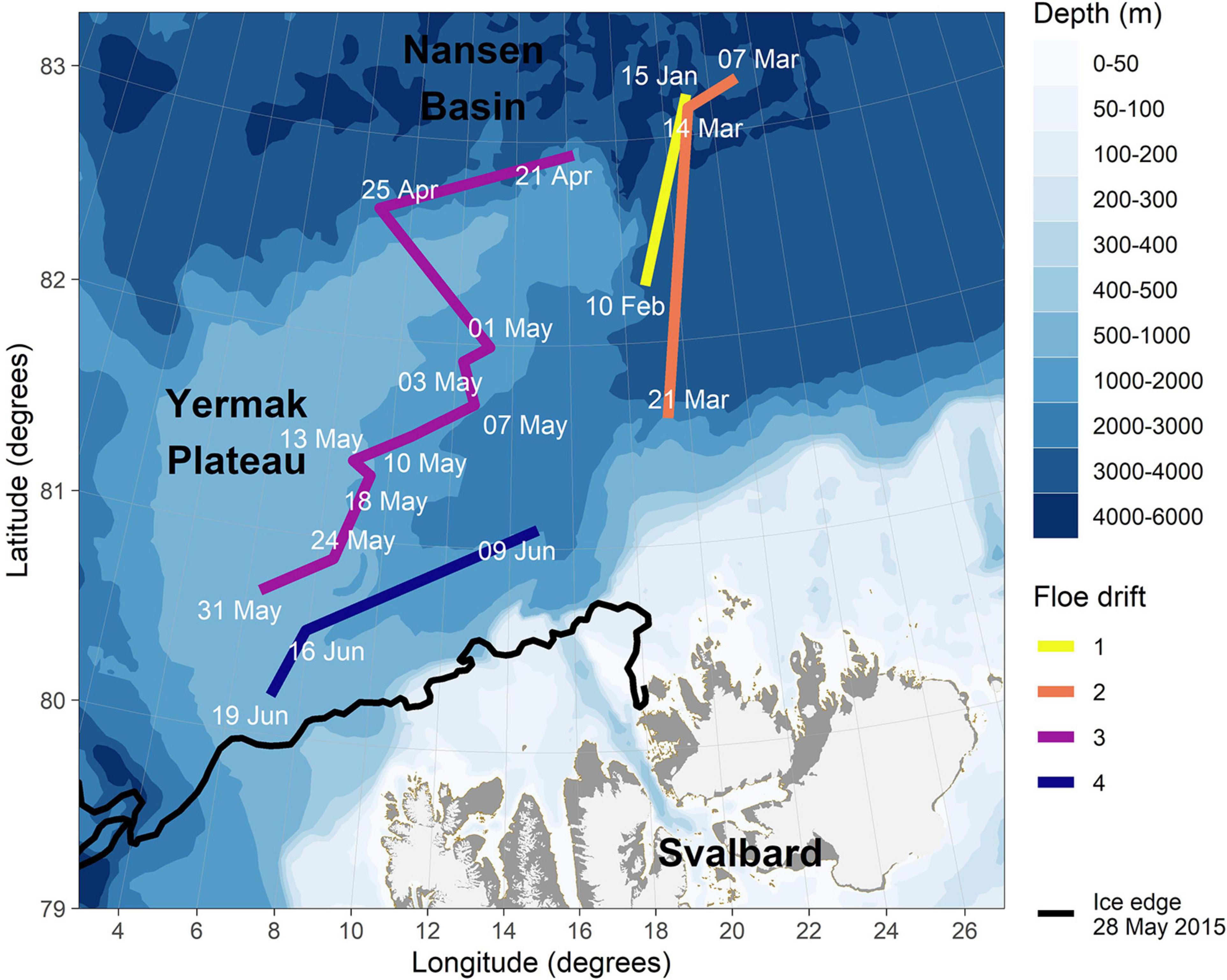
Figure 1. Zooplankton sampling stations during the four ice drifts during N-ICE2015. Floe 1 drift (10 February), Floe 2 drift (7, 14, and 21 March), Floe 3 drift (25 April, 3, 10, 18, 24, and 31 May) and Floe 4 drift (9, 16, and 19 June). The black line indicates the 10%+ ice edge on 28 May.
The N-ICE2015 expedition took place both in the Nansen Basin and over the Yermak Plateau, north-east of Fram Strait (Figure 1). Fram Strait is the only deep passage for water exchange between the North Atlantic Ocean and the Arctic Ocean, and its oceanography is dominated by northward flowing warm and saline Atlantic Water (AW) at intermediate depth on the east side and a southward flowing cold Polar Surface Water (PSW) in the surface on the west side (Rudels et al., 2000). The inflow of AW splits into two main paths as it hits the Yermak Plateau, with one recirculating in Fram Strait (von Appen et al., 2016) and the second directing eastward north of Svalbard. This flow of Atlantic Water north of Svalbard subdivides into the Svalbard Branch (or Atlantic Water boundary current) which follows the south-east side of the plateau (Renner et al., 2018), the Yermak Branch which circles anticyclonically around the Yermak Plateau, and the Yermak Pass Branch across the Yermak Plateau (Koenig et al., 2017). The Yermak Plateau is a local hotspot for vertical mixing affecting the primary production and hence the zooplankton production in the area (Padman and Dillon, 1991; Rudels et al., 2000; Daase and Eiane, 2007). The Floe 1 drift of the N-ICE2015 expedition lasted 38 days in January-February, passing over the Nansen Basin and the northern edge of the Yermak Plateau in deep waters (>3,000 m depth; Table 1). The Floe 2 drift lasted 24 days and passed over deep water in the Nansen Basin during February-March. The Floe 3 drift lasted the longest, for a period of 49 days from April to the end of May, from the northern slope of the Yermak Plateau to the southern edge of the Plateau. The majority of the zooplankton samples were collected during this third drift, which occurred in waters of 1,000–2,000 m depth with a shallower mixed layer and higher water temperatures than further north-east (Meyer et al., 2017b). Finally, the Floe 4 drift lasted 16 days and covered a similar track southeast of the Floe 3 drift over the Yermak Plateau. More details for each floe drift can be found in Meyer et al. (2017b).
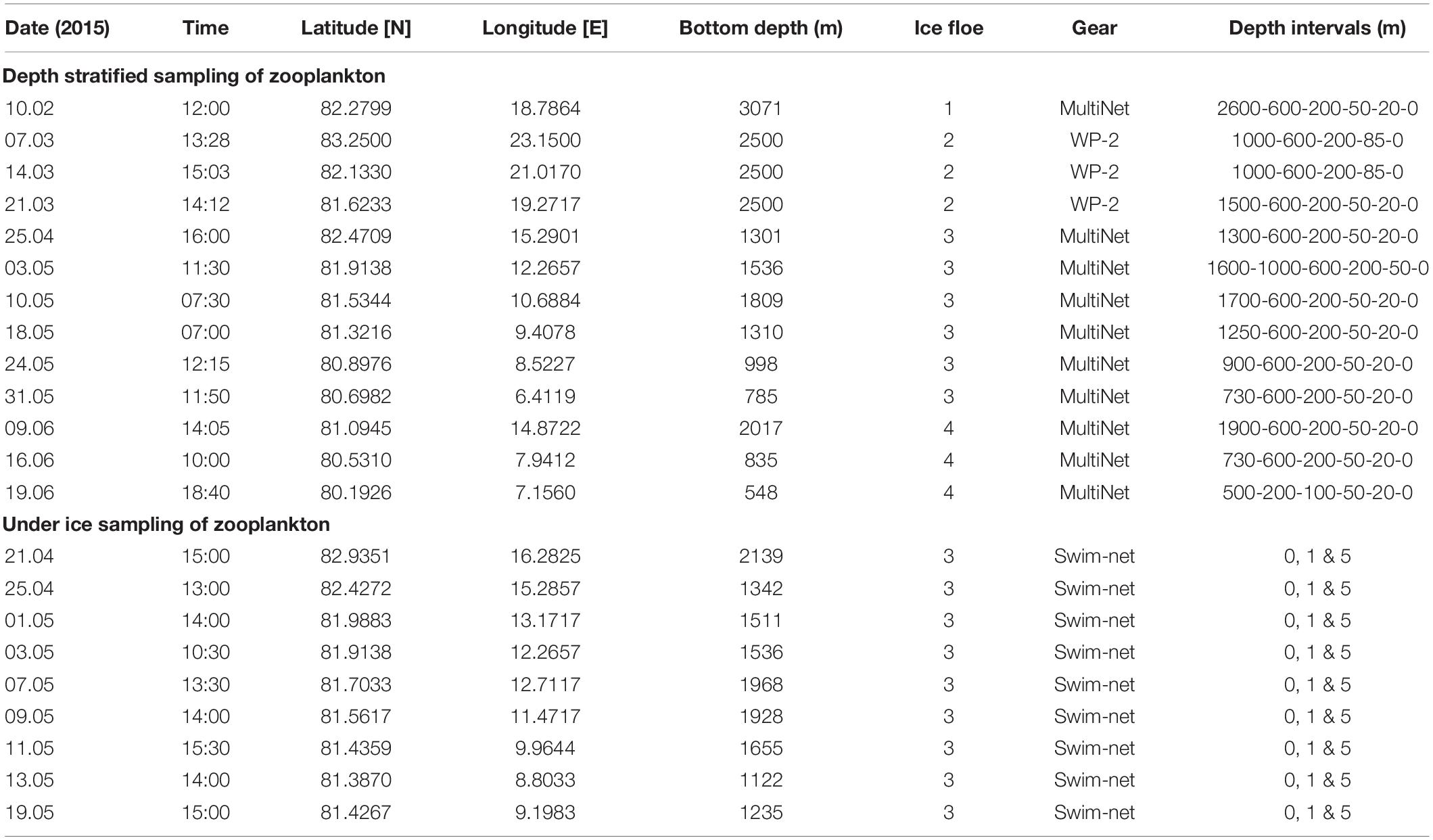
Table 1. Sampling stations for zooplankton with nets of mesh size 200 μm during N-ICE2015.
Water Masses, Chlorophyll a in Water Column and in Ice Cores
Six different water masses were identified using the classification by Rudels et al. (2000): Polar Surface Water, warm Polar Surface Water, Atlantic Water, Modified Atlantic Water, Intermediate Water, and Nordic Deep Water. For details, see Hydrography and Water Masses in Meyer et al. (2017b). The water-mass analyses were based on data from the ship CTD (Sea-Bird Electronics SBE911) and IAOOS buoys profiler (MSS-90 microstructure profilers). Mixed-layer depth was defined based on seasonal density criteria: in winter, it was defined as the depth in each profile where the potential density first exceeded the density at 20 m depth by 0.01 kg m–3. In spring, it was defined as the depth in each profile where the potential density first exceeded the near-surface value by 0.003 kg m–3 (see Meyer et al. (2017a) for details). Seawater samples for chlorophyll a (Chl a) were collected with 8 L Niskin bottles attached to a Sea-Bird rosette water sampler. A known volume of seawater from distinct depths was filtered on 25 mm GF/F filters (Whatman). Chlorophyll a was extracted in 100% methanol for 12 h at 5°C and measured fluorometrically using a Turner Fluorometer 10-AU (Turner Design, Inc.). Phaeopigments were also measured by fluorescence after acidification with 5% HCl (Holm-Hansen and Riemann, 1978). Chlorophyll a was measured, in the same manner as for seawater, from the bottom 10-cm layer of ice cores collected during 10 February-19 June. Sectioned cores were put into cleaned opaque plastic containers with lids and melted overnight at room temperature without addition of filtered seawater (Rintala et al., 2014).
Zooplankton Sampling and Taxonomical Analyses
Mesozooplankton was sampled from 10 February to 19 June (Table 1). Samples from the water column were collected during the entire study period with a Multiple Plankton Sampler (MultiNet type Midi, Hydro-Bios Kiel), consisting of five closing nets with 0.25 m2 square opening and 200 μm mesh size, pulled at a speed of about 1 m s–1. During 7–21 March, ice conditions were so severe that it was impossible to keep an opening in the ice next to the ship for deployment of the MultiNet. During this period, a WP-2 ring net with opening of 0.25 m2 and 200 μm mesh size was used instead, and samples were taken from four layers down to 1,000 m depth. Samples from underneath the sea ice were collected by scuba divers from 21 April to 19 May by use of a hand-held square net (25 cm × 25 cm, 0.125 m2) with 200 μm mesh size net bags. Horizontal transects of 80 m, swimming at about 1 m s–1 from the dive hole with return, were conducted at 0, 1, and 5 m below the sea ice, with three replicates per depth. Samples for taxonomical analyses were preserved with a 4% hexamethylenetetramine-buffered seawater formaldehyde solution immediately after collection.
All nets had the same mesh size, but net selectivity for the swim net is likely different from the vertically-hauled nets (MultiNet and WP-2). The swim net will sample more efficiently the water layer below sea ice, and results are therefore presented separately in tables and figures. The results from vertically-hauled nets are presented together, since their selectivity is similar, although the vertical resolution is more limited with the use of WP-2 net.
The organisms were identified and counted under a stereomicroscope equipped with an ocular micrometre, according to standard procedures (Harris et al., 2000; Kwasniewski et al., 2003). Typical mesozooplankters (organisms with total body length <5 mm, including most of Copepoda, juvenile stages of Pteropoda, Euphausiacea, Ostracoda, and Amphipoda) were identified and counted in sub-samples obtained from the fixed sample volume by pipette (approximately 500 individuals). Large zooplankters (organisms with body length >5 mm, including the largest Copepoda, Pteropoda, Euphausiacea, Ostracoda, Amphipoda, Decapoda, Appendicularia and Chaetognatha) were sorted out and identified from the whole sample. The three different Calanus species were differentiated based on morphology and prosome lengths according to Kwasniewski et al. (2003). Correct determination of C. finmarchicus and C. glacialis has been improved with genetic testing (Gabrielsen et al., 2012; Choquet et al., 2018), with some discrepancies relative to prosome length-based identification of the two species (Kwasniewski et al., 2003). However, it is assumed that the number of mis-determinations for this study is low (<10%) because the most northern Calanus spp. has clearer separation of their prosome length ranges (Choquet et al., 2018).
Calanus nauplii are not possible to determine to species according to size (Daase et al., 2011). Additional molecular analyses of Calanus nauplii were therefore conducted to identify which species being the main recruitment contributor. DNA was extracted following the HotSHOT protocol and species identification was done using six nuclear markers type InDels (Insertion or Deletion motifs) scored on a 3500xL genetic analyser (Applied Biosystems); see Smolina et al. (2014) and Choquet et al. (2017) for further methodological details. Up to 100 Calanus nauplii were randomly picked and analysed from each depth strata in June (the peak nauplii season). From these nauplii, the proportions of nauplii belonging to each of the Calanus species were calculated and applied to the quantitative community samples to estimate the abundance. Nauplii species were also determined genetically in February to see which Calanus species were producing eggs in winter.
Abundances of zooplankton were converted to biomass based on established conversion factors for zooplankton taxa (Hop et al., 2019b). Cnidarian and Ctenophores are listed with abundances only, and were not included in the calculations of total biomass because they were not sampled representatively by the MultiNet and have no reliable biomass conversion factors.
Lipid Content
Lipid content was measured for specimens of all three Calanus species that were picked out from samples from depth layers varying between 0 and 600 m. Copepods collected from 10 March to 3 June were photographed from lateral view through a stereo microscope (Leica MSV266) equipped with a photo adapter and a Leica MC170 HD camera. Lipid sac area and prosome length were measured from the pictures using imageJ (Schneider et al., 2012). Total lipid content (TL, mg) was estimated from lipid sac area (A, in mm2) with equation from Vogedes et al. (2010):
Data Treatment and Statistics
A canonical correspondence analysis (CCA) using CANOCO v5 (ter Braak and Šmilauer, 2002; Greenacre, 2016) was performed to reveal relationships between distribution of mesozooplankton and environmental variables. CCA is a constrained ordination analysis of taxon data, where the imaging of the taxa distribution pattern is constrained by the relationship between the taxa’s occurrence and environmental factors established using a linear model. The mesozooplankton abundance matrix used as input in the CCA included 111 taxa in 63 depth-stratified samples. The original data were used without transformation. The environmental variables included in the analysis were: date (Julian day), distance to shelf break (nm), mean depth of sampled layer (m), salinity, mean water temperature (°C) in sampled layer, water column Chl a (mg m–2), and Chl a concentration in the bottom 10 cm of ice (mg m–3). The contribution of the environmental variables was evaluated using Monte Carlo permutation test with 999 permutations and only variables that significantly (p < 0.05) explained the species composition were included in the CCA ordination model. The length of the environmental arrows and their orientation on the biplot indicate the relative importance of the variable to each axis, and the angle between arrows indicates the correlation between individual environmental variables. The distance between the samples (circles) approximates the dissimilarity of their species composition measured by the Euclidian distance. The closer the samples are clustered together, the more similar their species composition. For simplicity, only the 25 species with the largest fit value to the axis are shown.
Results
Seasonal Changes in Water Masses, Chlorophyll a and Zooplankton Abundance
During winter, the N-ICE drift was in an area showing classic Arctic Ocean properties: a cold (<0°C) relatively fresh and deep mixed Polar Surface Water down to 100 m, warmer and more saline Atlantic Water and Modified Atlantic Water between 100 and 500 m, and Intermediate Water and Nordic Deep Water down to 1,000 m depth and below (Figure 2).
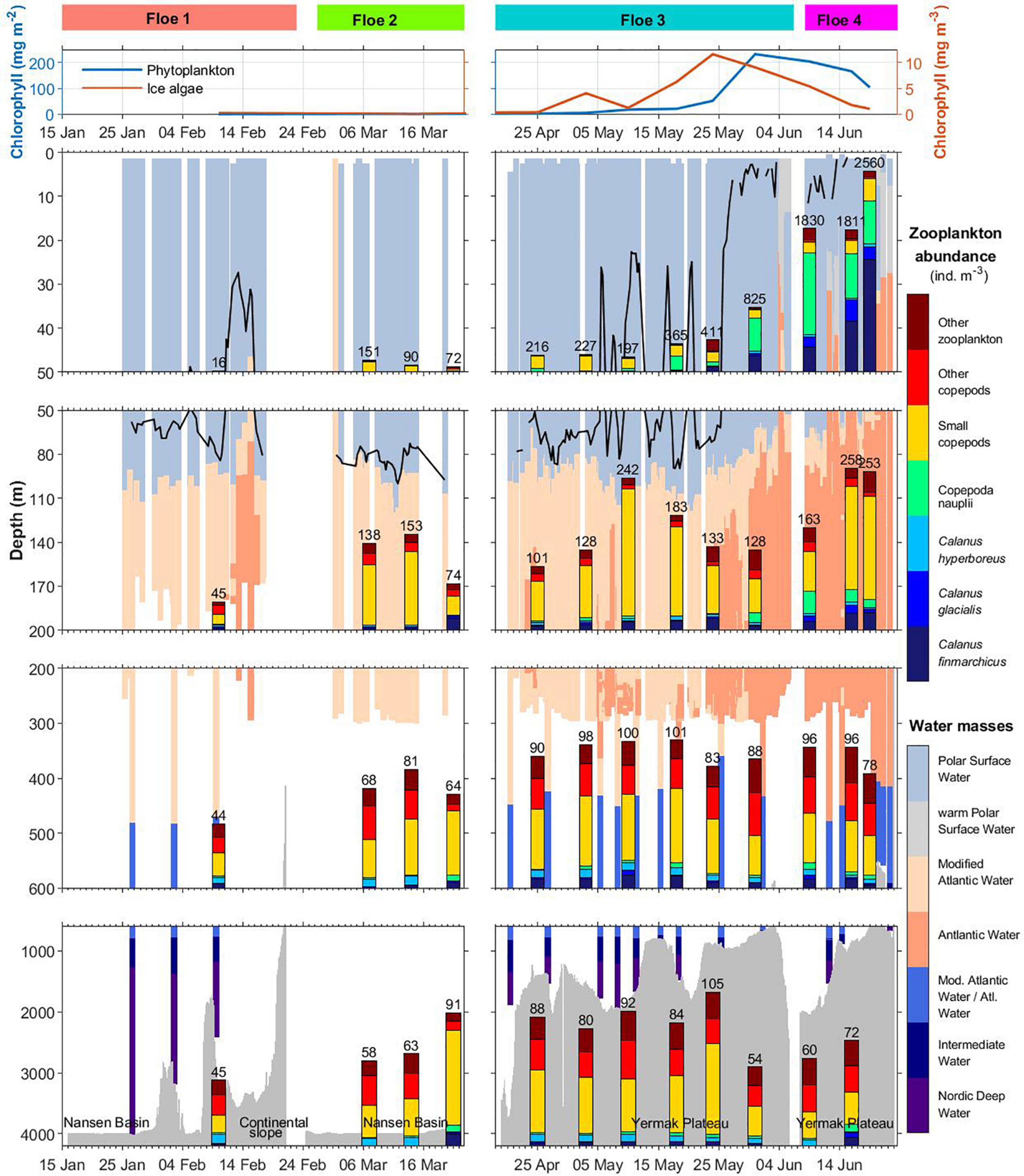
Figure 2. Chlorophyll a standing stock of phytoplankton (mg m–2) and ice algae in the bottom 10 cm of sea ice (mg m–3) during the N-ICE2015 drift (upper panels). Vertical distribution of water masses and abundance of key zooplankton species at different depth ranges: 0–50 m, 50–200 m, 200–600 m, and 600 m to deepest sample. Water-masses are labelled by colour: Atlantic Water (AW), Modified Atlantic Water (MAW), Polar Surface Water (PSW), warm Polar Surface Water (PSWw), Intermediate Water (IW) and Nordic Deep Water (NDW) following definitions by Rudels et al. (2000). Lack of colour indicates a lack of hydrography data, most notably between drifts and at depth. Overlain the water masses colour scale is a contour of the mixed-layer depth (black line). Zooplankton abundance axes are scaled to fit each figure, for better discrimination between groups, with the sum of abundance (ind. m–3) indicated above bars. Seafloor depth indicating topographic features is shaded in grey.
Hydrographic conditions changed significantly after 25 May, when the ship drifted over the Yermak Plateau (800–1,000 m depth), where the Atlantic Water was thicker and closer to the surface, while the mixed surface layer was thinner, fresher and warmer. The standing stock of Chl a in the surface water started to increase from early May, with the steepest increase happening after 25 May when the mixed layer was thinner, reaching maximum concentration on 31 May with 200 mg Chl a m–2. The highest surface values of Chl a reached 7.5 μg L–1 on 2 June (Assmy et al., 2017). The Chl a in the bottom layer of the sea ice started to increase from early May, peaking on 24 May with 11.6 mg m–3 in the bottom 10-cm sea ice.
The most pronounced change in zooplankton community was the appearance of the large calanoid copepods Calanus finmarchicus, Calanus glacialis as well as copepod nauplii in surface waters from early May (Tables 2A, 3). The CI and CII stages of copepods that showed the earliest increase in surface waters were those of Calanus hyperboreus, whereas those of C. glacialis appeared a week later. The largest increase in copepod nauplii was observed just after the peak in Chl a standing stock, on 9 June (Figures 2, 3A). However, from samples collected by divers from right below the sea ice, the increase in Calanus nauplii had already started in early May (Figure 4). The total abundance of copepods (excluding nauplii) in MultiNet samples from the upper 50 m varied from 12 to 380 ind. m–3 from February until the end of May and then increased rapidly to 1950 ind. m–3 on 19 June, mainly due to an increase in abundance of C. finmarchicus and C. glacialis (Table 2A and Figure 3A), which reached 1440 and 160 ind. m–3, respectively. Calanus hyperboreus contributed less to abundance (about 30 ind. m–3 from 31 May), but more to biomass because of its size (Figure 3B). Part of the zooplankton proliferation in the surface 50 m in June was due to presence of small species, such as Oithona similis (230 ind. m–3), Microcalanus spp. (45 ind. m–3) and Pseudocalanus spp. (36 ind. m–3), which were also collectively abundant (140 ind. m–3) in the intermediate layer from 50 to 200 m (Table 2B and Figure 3A).
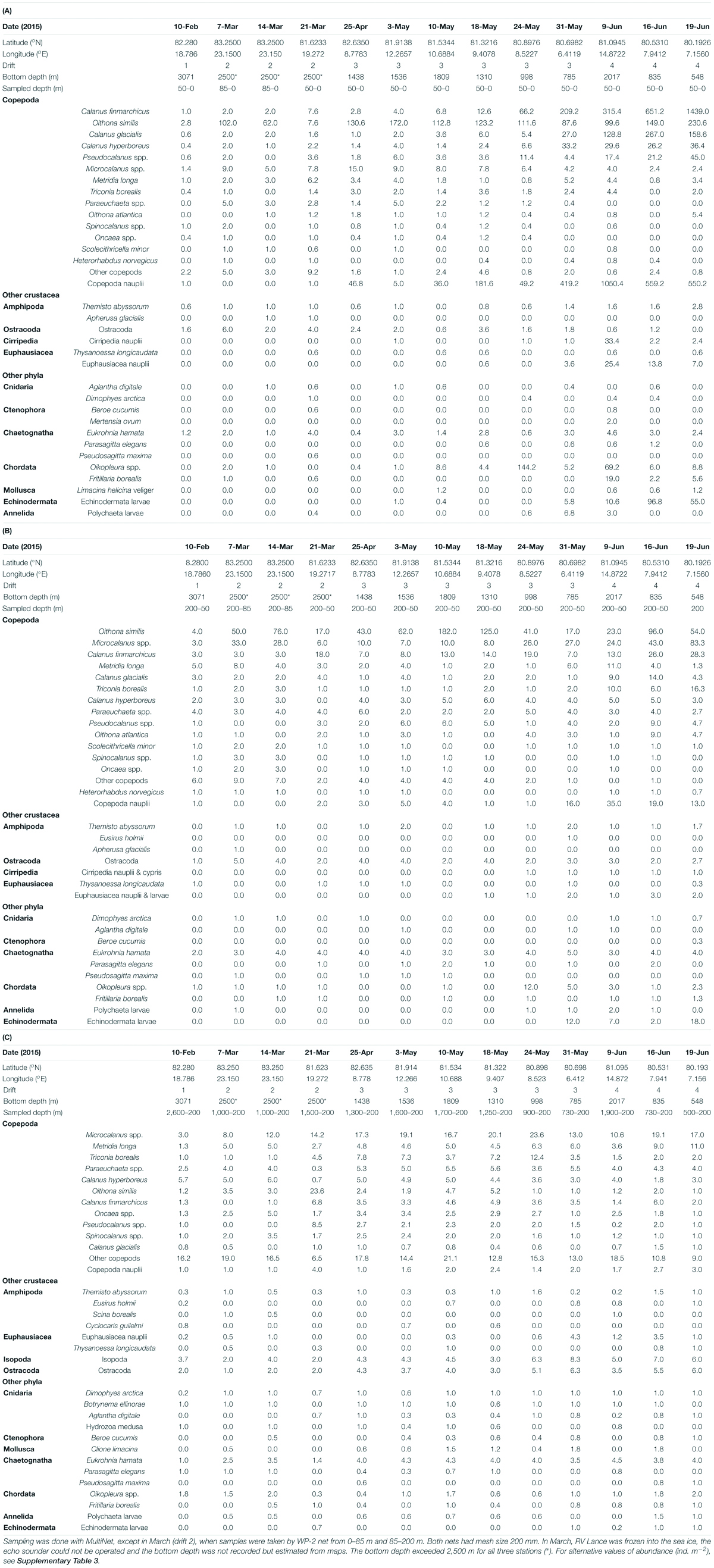
Table 2. Abundance of main zooplankton taxa (ind. m–3) in the Arctic Ocean at 0–50 m depth (A), 50–200 m depth (B), and 200 m to near bottom (C).
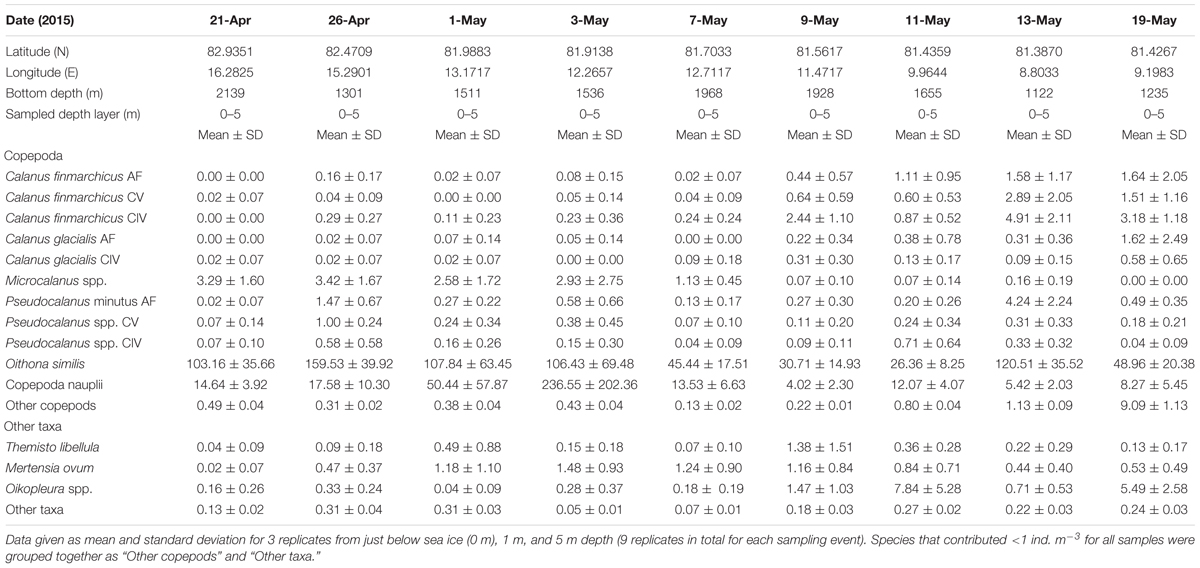
Table 3. Zooplankton taxa abundance (ind. m–3) for swim-net samples collected by divers from 21 April to 19 May 2015.
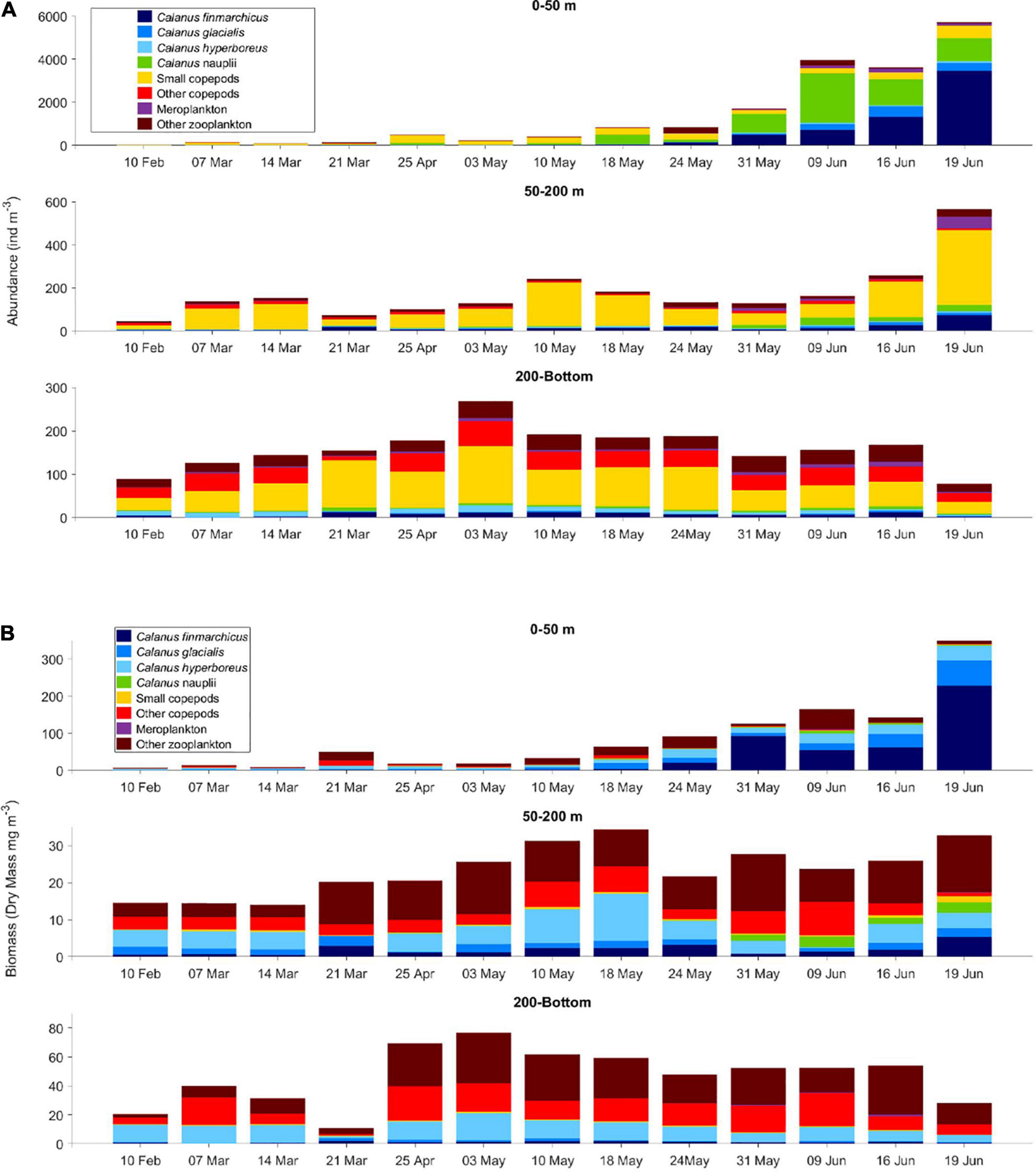
Figure 3. (A) Abundance (ind. m–3) and (B) biomass (dry mass, mg m–3) of main zooplankton taxa sampled with plankton nets (MultiNet and WP-2) in different water layers (depth ranges) from 10 February to 19 June, 2015. Note variations in scale for the different depth layers. The bottom depth on 19 June was 548 m.
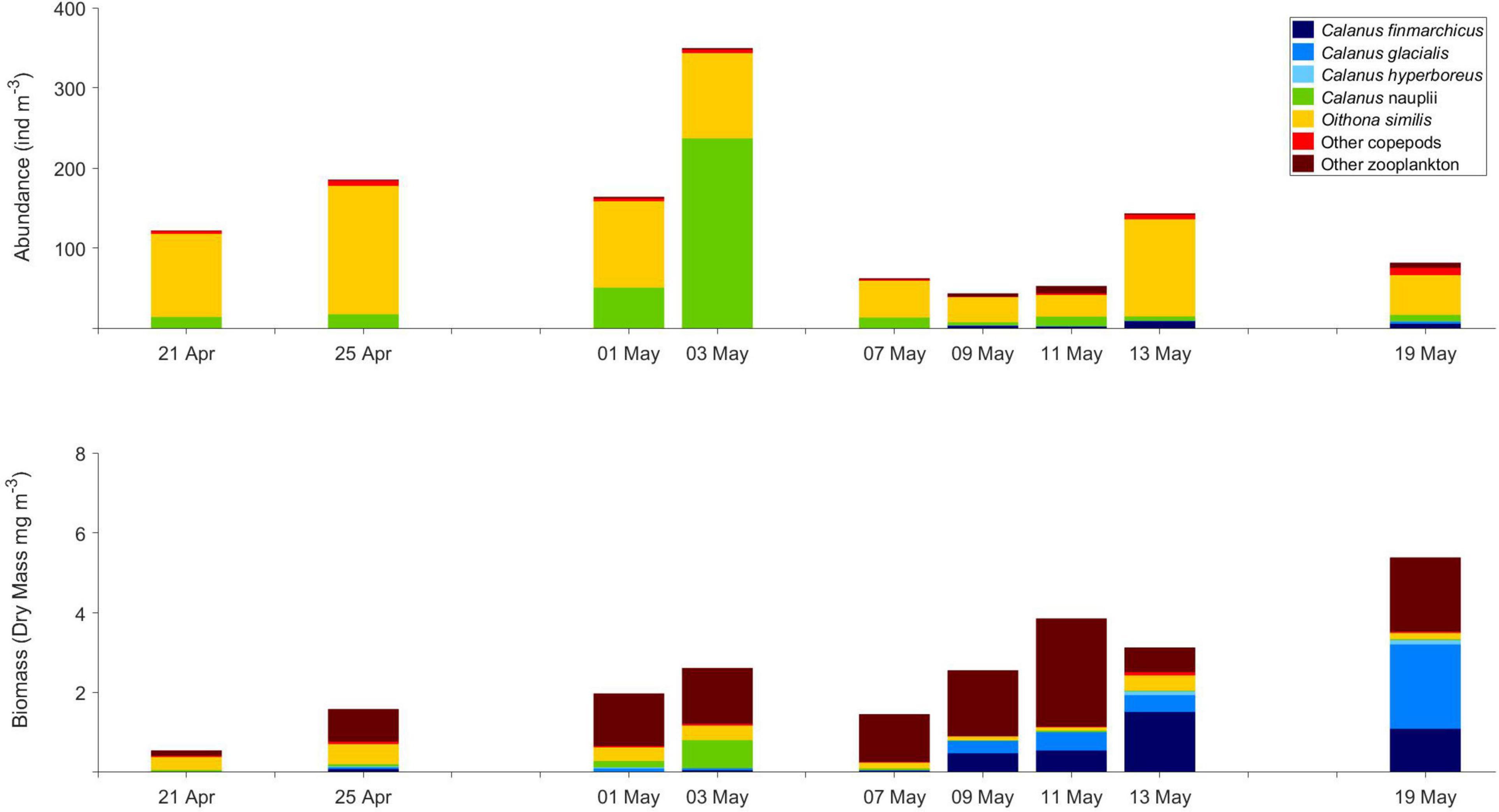
Figure 4. Abundance (ind. m–3) and biomass (dry mass, mg m–3) of main zooplankton taxa in the upper 5 m below the ice, collected with swim nets by scuba divers, from 21 April to 19 May 2015.
Other zooplankton species in the upper 50 m included the hyperiid amphipod Themisto abyssorum and appendicularians Oikopleura spp. In June, meroplankton increased in abundance when the ship drifted closer to the continental shelf (Figure 3A). Cirripedia dominated the meroplankton, followed by larvae of Polychaeta, and Echinodermata. Juvenile forms of Euphausiacea and Pteropoda (Limacina helicina), and Ostracoda were present in moderate numbers in most of the water column during the entire sampling period (Tables 2A–C).
The species composition and abundance of zooplankton in the two deepest layers (200–600 m and 600-bottom) changed little throughout the seasons (Figures 2B, 3A). The total abundance of copepods below 200 m varied from 36 to 76 ind. m–3 (Table 2C). The deep samples were dominated by smaller species (body length <1 mm) such as Oithona similis, Triconia borealis and Microcalanus spp. as well as by typical deep-water and large-bodied species, such as Heterorhabdus norvegicus, Gaetanus brevispinus, Paraeuchaeta norvegica and several Spinocalanus species. Other zooplankton was represented mainly by chaetognaths (Eukrohnia hamata), Oikopleura spp., Euphausiacea nauplii and larvae, and, to a lesser extent, by adult Thysanoessa inermis (Table 2C).
The biomass contributions of smaller copepods were minor despite their high abundance, while that of larger copepods dominated. In the surface waters (0–50 m), the increase in biomass in late May was mainly attributed to increased abundance of C. finmarchicus and partly C. glacialis (Figure 3B). This increase could be seen in mid-May in the intermediate layer (50–200 m) and in early May in the deepest layers (200 m-bottom). The increase in C. finmarchicus was especially large at the last sampling station on 19 June (Tables 2A,B and Figure 2), when the drift had approached the shelf break and the core of the Atlantic Water inflow. In the intermediate layer between 200 and 600 m, other zooplankton such as Clione limacina and Thysanoessa spp. together with larger copepods Paraeuchaeta spp. and C. hyperboreus contributed most to the overall mesozooplankton biomass. The biomass in the intermediate layer ranged from 15 to 51 mg m–3, except for low values on 21 March, when the ship was frozen completely into the sea ice and net sampling became difficult (Table 2). Biomass in the deepest layer ranged from 5 to 20 mg m–3 during winter, with an increase in late April to 54 mg m–3, which also corresponded to a change in the location of the ship from deeper to shallower waters (Figure 2). Metridia longa, Paraeuchaeta spp. and E. hamata, comprised a large fraction of the total biomass, and typical deep-water species H. norvegicus, G. brevispinus, and P. norvegica were also present (Supplementary Table 1).
Zooplankton Distribution in Relation to the Environmental Variables
The CCA showed that six of the tested environmental variables significantly explained the species distribution (Table 4). The total variation (inertia) was 2.318 and together these variables account for 48.7 % of the variance in the species distribution. Salinity explained the largest part (19.6%) followed by mean depth (8.5%), standing stock Chl a in the upper 50 m (7.2%), Chl a in the bottom 10 cm sea ice (6.3%), temperature (4.1%) and date (3.0 %). The first and second canonical axes explained 48.8 and 19.9% of the variance in the species data restricted by the environmental variables and 23.8 and 9.7 % of the total variance, respectively. Mesozooplankton in the surface waters (Figure 5A, blue and green sample marks) showed two distribution patterns, illustrated by the presence of two clusters, where the late May and June samples differed from the samples earlier in the season, and their arrangement was strongly related to the concentration of Chl a in the upper 50 m. In contrast, mesozooplankton in waters deeper than 200 m, and especially below 600 m (Figure 5A, purple and black sample marks), did not change much from winter to spring with the environmental factors studied, which is documented by the grouping of all samples collected in deep waters into one cluster.
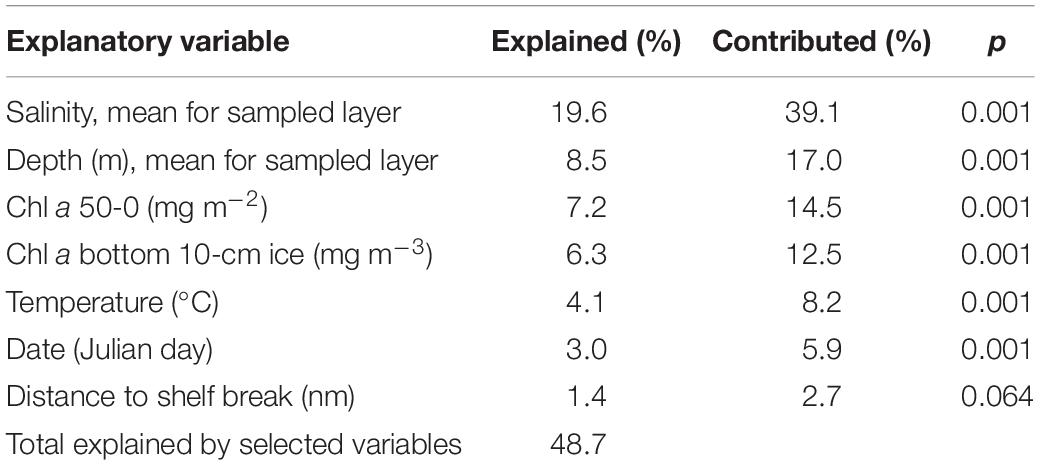
Table 4. Results of forward selection of environmental variables in CCA analysis.
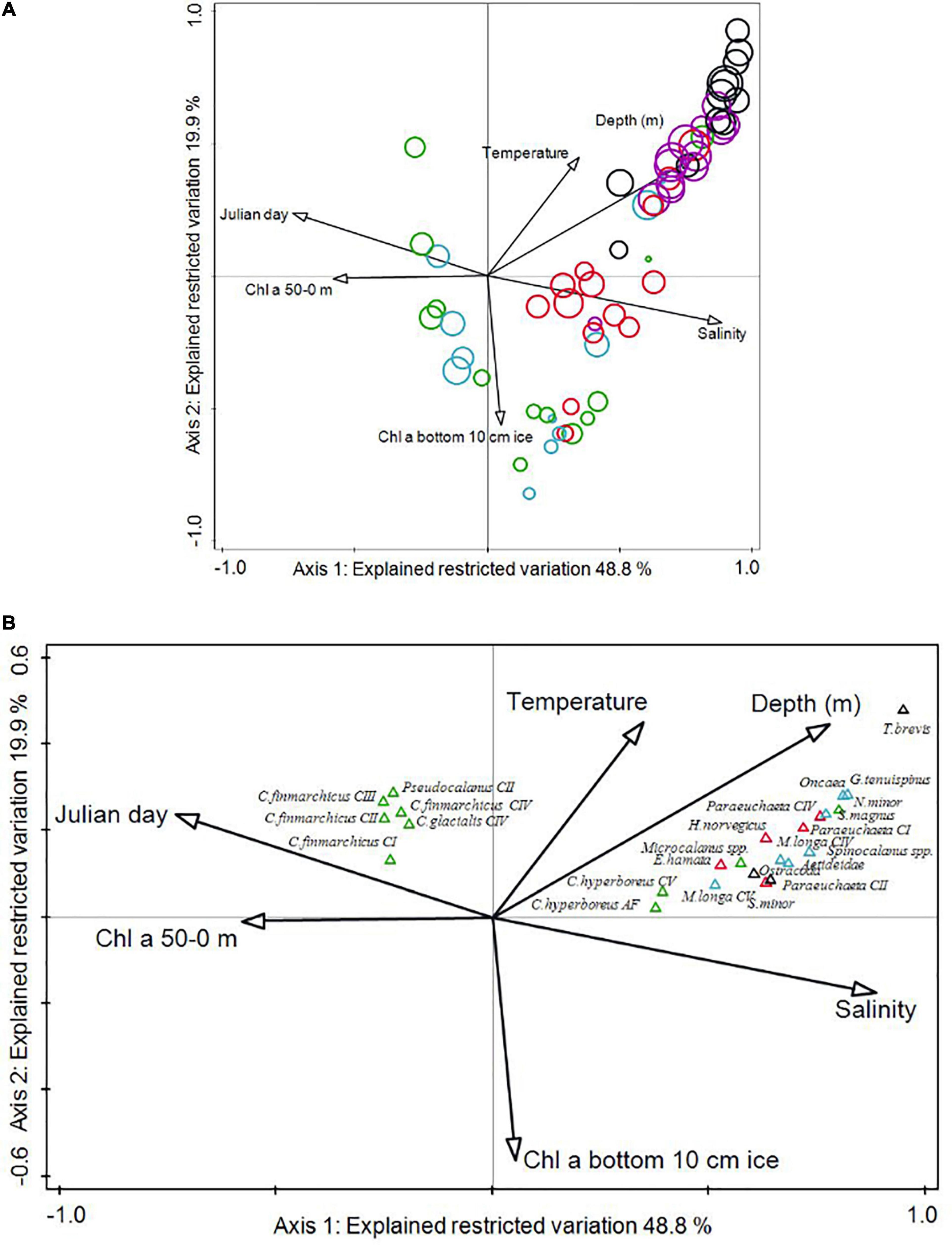
Figure 5. CCA biplots of (A) samples of mesozooplankton abundance (coloured circles) and environmental variables (arrows) and (B) species and environmental variables (arrows). Environmental variables are listed in Table 4. The first canonical axis explains 49% of the variation explained by the selected environmental variables. The second axis explains 20% of the variation explained by the selected environmental variables. In panel (A), the size of the sample circles reflects the count of species within the sample, the greater the size the higher species diversity within that sample. Samples from different depth layers are in different colours. The different colours of the species in panel (B) indicate the predominant trophic level of that species; herbivores are marked in green, carnivores in red and omnivores in blue and undefined in black. For simplicity reasons, only the 25 species with the largest fit value to the axis are shown. Some of the species’ names are given as abbreviations, the full names are given in Supplementary Table 2.
Temporal and spatial distribution of zooplankton species was also assessed with regards to their trophic levels, determined from literature (i.e., main trophic level of a species during spring-summer, even though it may change seasonally), with three groups: herbivores, carnivores and omnivores identified (marked green, red and blue, respectively, in Figure 5B). Surface samples in spring were typically dominated by herbivores such as Calanus spp., and larval stages of euphausiids (calyptopis and furcilia). Cirripedia nauplii were present in these spring samples as well, but absent from all deep samples and surface samples earlier in the season. The deep-water samples showed larger species diversity and were characterised by omnivorous species such as Gaetanus tenuispinus, Scaphocalanus magnus, Neomormonilla minor, Spinocalanus spp., Oncaea spp., Pseudocalanus spp. and Metridia longa, as well as carnivorous species, such as Paraeuchaeta spp. (including females of three species, P. norvegica, P. glacialis and P. barbata).
Lipid Stores and Reproduction in Calanus spp.
Calanus finmarchicus was present in low numbers in February and March, dominated by copepodids of stage IV and V (CIV and CV), likely overwintering stages, and a few adult females were also present (Supplementary Figure 1). Calanus glacialis was similarly distributed with depth and represented by similar stages as C. finmarchicus throughout winter and spring, but was more concentrated in surface waters compared to C. finmarchicus from mid-May. Calanus finmarchicus females peaked in abundance later than C. glacialis, and female C. glacialis were largely gone by the end of May when the phytoplankton biomass peaked. Few lipid data on C. finmarchicus CV and females were collected before June, but a distinct increase in lipid content from low winter values (March) was observed when algal food arrived in June (Figure 6). For C. glacialis, no trend in lipid content could be identified during the winter-spring transition, and samples varied considerably. Calanus hyperboreus was generally distributed deeper than the other two Calanus species and at lower abundance, but aggregated at the surface after mid-May, after which both CVs and females increased their lipid reserves (Figure 6). The amount of lipid reserves was most related to size of the specimens as indicated by prosome length, with C. finmarchicus being the smallest, C. glacialis intermediate, and C. hyperboreus the largest and most lipid-rich of the species (Supplementary Figure 2).
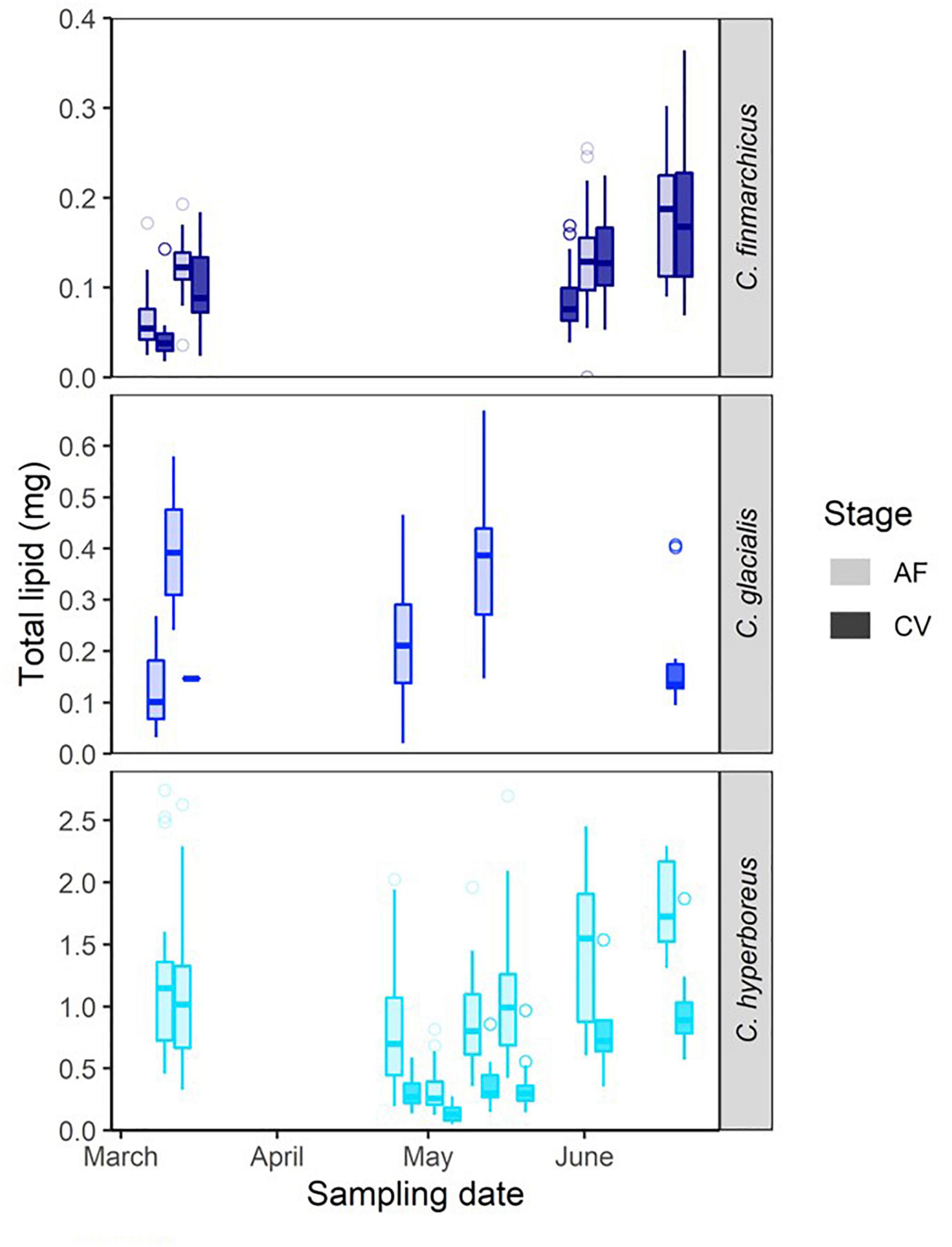
Figure 6. Total lipid in stage CV and AF (adult females) of three Calanus species over time, collected from the upper water column along the drifts (bottom depth of collection tow varies between 600 and 50 m). Boxplots present the following data: rectangle extent indicates the interquartile range (IQR), the 25 and 75% percentile, the middle line indicates the 50% quantile (the median), the whisker length extends to the last value within 1.5*IQR, and outlier points beyond this are indicated as points.
Young stages of C. hyperboreus (CII-CIII) were present in deep water (>200 m) from February, whereas CI appeared in the upper part of the water column at the end of April (Supplementary Figure 1). This coincided with the arrival of copepod nauplii in the upper 50 m beneath the sea ice from late April (Table 2A). Their abundance increased to 236 ind. m–3 in the 5-m layer below the ice at the beginning of May (Table 3 and Figure 4). Copepod nauplii reached approx. 1,000 ind. m–3 in the upper 50 m in early June (Table 2A). Thus, most of the seasonal signal was associated with the upper water layers, with the largest increase toward the end of the expedition (after 31 May), when the abundance of zooplankton (nauplii and larval forms excluded) increased rapidly from 390 to 1940 ind. m–3 in the upper 0–50 m layer and from 170 to 650 ind. m–3 in the 50–200 m layer below.
Molecular identification of Calanus nauplii in June showed that C. finmarchicus nauplii totally dominated in the upper 600 m with only a few C. hyperboreus and C. glacialis nauplii present in the upper 100 m (Figure 7). Calanus nauplii below 600 m depth were all identified as C. hyperboreus, and those present in April were mostly of the same species. Copepodid stages CI and CII peaked in surface waters by 31 May for C. hyperboreus and then a week later for C. glacialis. All young stages of C. finmarchicus increased in surface waters from 9 June at the onset of Floe 4 drift, which was still over 2,000 m depth but closer to the shelf and with the influence of Atlantic Water higher in the water column (Figure 2).
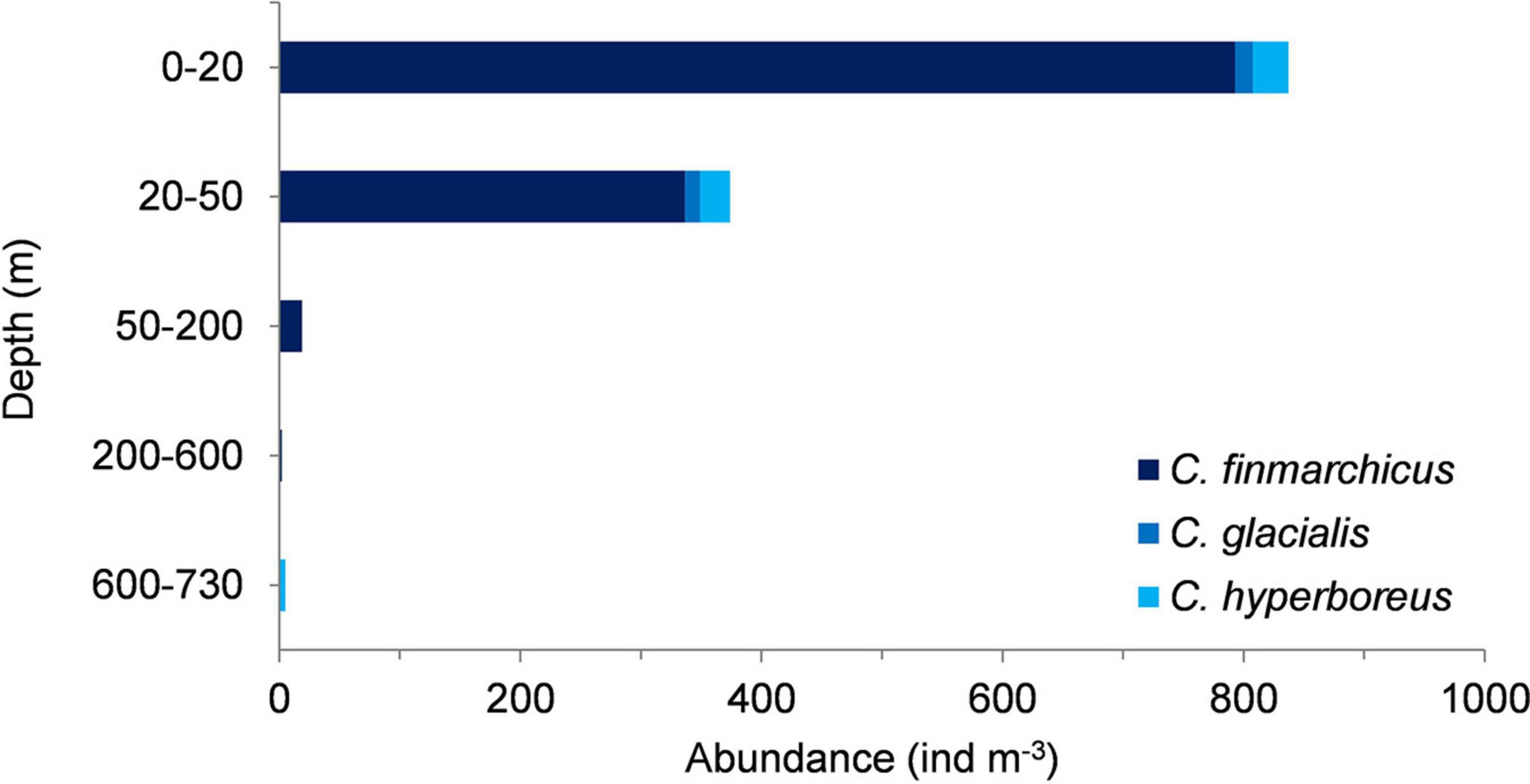
Figure 7. Abundance (ind. m–3) of Calanus nauplii sampled with MPS from bottom to surface in five depth layers on 16 June, 2015. Species composition estimated from genetically identified samples collected with MultiNet from the same depth layers immediately after the community samples (Figure 3).
Discussion
Changes in Zooplankton Community From Winter to Spring
During the N-ICE2015 sea ice drift, we moved from thicker pack-ice conditions above deep waters in winter to thinner ice near the outer ice edge and shelf during late May. Mesozooplankton in the upper and midwater layers consisted of advected Atlantic expatriates, such as C. finmarchicus, Scolecithricella minor, Heterorhabdus norvegicus and Oithona atlantica. The Atlantic Water core follows the continental slope north of Svalbard at depths from below the mixed layer down to 800 m depth (Meyer et al., 2017b), and we likely drifted into an area of more C. finmarchicus with a mixed assemblage of young stages, in late May-early June. The increase in C. finmarchicus and other zooplankton in late May and early June most likely reflected the vessel’s drift over an area with more Atlantic Water inflow combined with zooplanktons vertical seasonal migration to the surface in time for the rising primary production in surface waters.
Calanus finmarchicus is transported with Atlantic Water into the Arctic, and about 30% of its abundance likely continues eastwards in the Atlantic Water boundary current (Basedow et al., 2018; Hop et al., 2019a). The zooplankton is eventually carried into the Nansen Basin and the central Arctic Ocean (Kosobokova et al., 2011), with diminishing contribution by C. finmarchicus being subsequently replaced by C. glacialis and C. hyperboreus toward the central Arctic Ocean (Hirche and Mumm, 1992; Mumm et al., 1998; Kosobokova and Hirche, 2009; Ershova et al., 2021).
The variation in copepods and other zooplankton in the deep layer observed in this study likely reflected spatial changes including ocean depth due to our drift rather than seasonal and temporal patterns. However, some reduction in abundances of C. glacialis and C. finmarchicus were observed in the deeper layers when these species increased in the upper layers after mid-May (Supplementary Figures 1A,B).
Holoplanktonic Ostracoda, which are typically mesopelagic, were more abundant in deeper water and toward the end of the drift. Presence of meroplankton, the larval stages of benthic organisms, from Echinodermata, Gastropoda and Cirripedia, likely coincided with the drift over the Yermak Plateau (<1,000 m), and toward the continental shelf with the resultant proximity to benthic communities.
The sub-ice layer of ice algae started to develop in the area in late April (Kowalczuk et al., 2017), with rapid increase in algal biomass from mid-May below both first-year and second-year sea ice, and with maximum biomass around 21–23 May (Olsen et al., 2017). Ridges of first-year ice and infiltration layers between ice and snow are important habitats for ice algal growth, and the peak in Chl a biomass was found in ridges in the middle of May (Fernández-Méndez et al., 2018). The pelagic bloom peaked in early June, about 2 weeks after the peak of the ice-algal bloom (Assmy et al., 2017). The sudden increase in Chl a at the end of May coincided with a distinct change in the hydrographic conditions as the research vessel drifted south over the Yermak Plateau, where the Atlantic Water layer was thicker, reaching closer to the sea ice, with a very thin layer of fresh and cold water from ice melt on top. In June, the high Chl a in sea ice reflected infiltration by phytoplankton as well as accumulation of detritus (Kowalczuk et al., 2017).
The ice algae build-up in the ridges in mid-May and a week later below flat ice might have fuelled the reproduction of Calanus species and the following growth and development of nauplii observed in May. The reproductive success of some herbivorous zooplankton, such as C. glacialis, depends on a match between the timing of ice algae and phytoplankton blooms and specific life-history events, with ice algae fuelling gonad maturation and egg production, and the phytoplankton bloom providing food for development and growth of the succeeding offspring (Søreide et al., 2010). Females of C. glacialis likely benefitted from the ice algae bloom, rather than the phytoplankton bloom. Very few C. glacialis nauplii were present in June, and females had disappeared by mid-May, one to two weeks prior to the peak phytoplankton bloom. Calanus glacialis can switch from capital to income breeding when food becomes available, and the maximum potential for income breeding was therefore most likely not achieved for C. glacialis in this region. The larger C. hyperboreus is known to reproduce in winter, decoupled from the spring bloom (Hirche, 1997; Halvorsen, 2015). This is supported by our finding of nauplii and younger copepodids of this species in February. The presence of C. hyperboreus nauplii as late as June is interesting since this suggests that this species can produce eggs from November to June in this region (Halvorsen, 2015; this study) or potentially this could be explained by two different C. hyperboreus populations that co-exist in this area, one local and one advected with different timing of reproduction.
Spawning in the upper water layer underneath the ice likely happened in C. glacialis and C. hyperboreus before the algal bloom. At low ambient temperatures, e.g., in Polar Surface Water (<0oC), it takes about a week from spawning to egg hatching and another 10 days for the nauplii to develop to the first feeding stage, N3 (Daase et al., 2011; Weydmann et al., 2015). Species identification of younger developmental stages of Calanus needs to be validated by molecular methods, since there is a potential for misidentification (Daase et al., 2011; Gabrielsen et al., 2012; Choquet et al., 2017). Samples collected by scuba-divers underneath the ice showed an increase in copepod nauplii already in the beginning of May, during the early part of the ice algal bloom. The increase in copepod nauplii in the upper water column was also seen in samples from vertical net hauls from mid-May onward. The nauplii subsequently increased in surface waters until the beginning of June, in the wake of the peak in Chl a standing stock, providing a continuous supply of Calanus nauplii due to differences in timing of egg production among the three species.
The copepod nauplii observed in May most likely were C. glacialis, while those detected in June were genetically identified as C. finmarchicus and were three to five times more abundant in the water column. The North Atlantic C. finmarchicus is known to reproduce later than the Arctic shelf species C. glacialis, since it relies on freshly ingested food for egg production (Hirche, 1990; Swalethorp et al., 2011). In our study, C. glacialis females peaked in abundance earlier than C. finmarchicus, which is in line with this sequence in reproduction. The dominance of C. finmarchicus in this study, however, was likely due to the influence of Atlantic Water in this region and the continuous supply of C. finmarchicus from northward flowing Atlantic Water (Basedow et al., 2018).
Timing of Seasonal Vertical Migration
The large herbivorous Calanus spp. copepods perform seasonal vertical migration, overwintering at depth (Hirche, 1997; Falk-Petersen et al., 2009; Darnis and Fortier, 2014), while the omnivorous copepods, such as the large-bodied Metridia longa and small-bodied Oithona similis, Triconia borealis and Microcalanus spp. are believed to remain active year-round, not carrying out extensive seasonal migration (Lischka and Hagen, 2005). During the spring, Calanus spp. ascend to the upper water layers, but the timing varies with latitude and environmental conditions (Ashjian et al., 2003; Daase et al., 2013; Darnis and Fortier, 2014). Their ascent is most likely triggered by changes in light levels (Cohen et al., 2015), but could also be influenced by intrinsic factors, such as the circadian clock (Häfker et al., 2017, 2018), and the depletion of lipid reserves (Falk-Petersen et al., 2009). Zooplankton, and Calanus spp. in particular, are also known to perform diel vertical migrations (DVM), which are coordinated during winter-spring, but uncoordinated, limited or non-existent during the Arctic summer (Blachowiak-Samolyk et al., 2006; Cottier et al., 2006; Berge et al., 2009). Our sampling was generally during the day and not set up to account for DVM; this could have contributed to larger variability in our stratified samples, but not influenced the overall seasonal patterns. The three Calanus species overwinter at different depths in the study area, with C. hyperboreus descending to the deepest horizons (below 1,000 m in some areas, less in others), C. finmarchicus going less deep (600–1,000 m) and the shelf-species C. glacialis staying within the upper 300 m (Hirche, 1997; Vinogradov, 1997; Ashjian et al., 2003; Falk-Petersen et al., 2009; Darnis and Fortier, 2014). Calanus hyperboreus and C. glacialis can overwinter in the Arctic Ocean. The situation for C. finmarchicus is uncertain (Kosobokova et al., 2011), although we found overwintering stages in our samples from the Nansen Basin. The currently known overwintering region for the populations found in this study likely extends along the continental slope from northern Norway to Spitsbergen, in the eastern regions of the Norwegian and Greenland Seas (Halvorsen et al., 2003; Edvardsen et al., 2006).
The different centres of distributions for zooplankton species, their respective depths of overwintering and subsequent advection during early spring may affect their seasonal appearances in the Arctic Ocean, particularly in the upper water masses. Calanus spp. advected from the south are likely more advanced in their development than Arctic-hatched Calanus, which may influence the stage composition (Supplementary Figure 1). Our data indicate seasonal changes in abundance down to 200 m, whereas below this depth the variability is more likely related to changes in geographical locations. The biomass of C. hyperboreus and C. finmarchicus started to increase at 50–200 m depth in early May and in the upper 50 m a week later, with rapid increase in abundance toward late May. The zooplankton “wake-up call” after overwintering at depth, thus, seems to be in early May in the European sector of the Arctic Ocean, followed by an increase in biomass in the upper part of the water column toward the end of May. The increases in Calanus nauplii and meroplankton were also attributable to the shelf and shallow water settings in spring, such as those at the Yermak Plateau. These coastal domains are generally much more productive than the deep oligotrophic Arctic Ocean domain (Wassmann et al., 2015).
The observed increase in copepod nauplii underneath the sea ice just prior to the ice algal bloom, and the high numbers of nauplii and young copepodid stages during the peak in pelagic Chl a, indicate a match between reproduction of the main herbivorous species and the ice algal and phytoplankton blooms in the study year. However, in addition to the timing of reproduction relative to the timing of blooms, the reproductive success also depends on the bloom magnitude, duration and quality (Daase et al., 2011; Leu et al., 2015). During the N-ICE2015 drift, the phytoplankton bloom beneath the pack ice was dominated by the haptophyte Phaeocystis pouchetii until early June when diatoms had increased to half of the standing stock (Assmy et al., 2017). Sediment trap samples revealed that the bulk of the collected material was P. pouchetii aggregates, and that the species export rates increased during the bloom. Few faecal pellets in the sediment trap samples indicated low grazing rates or potentially rapid degradation due to microbial activity (Svensen et al., 2012). Phaeocystis pouchetii is generally not considered palatable by zooplankton because of the formation of large aggregations and its chemical composition (including e.g., exopolymers), but it can be ingested by large copepods such as Calanus spp. (Njestgaard et al., 2007). The contribution by diatoms at the bloom peak was likely more important for the young stages of Calanus spp. Thus, the timing of seasonal vertical migration happens in anticipation of the presence of algal food, but what is available seems rather unpredictable. It was apparent in our study, particularly from the dive samples, that the Calanus copepods arrived below the sea ice before the ice-algal and phytoplankton blooms started.
Lipid Content and Match for Reproduction
The lipid content of Calanus spp. is related to individual body size, with the largest specimens having the highest lipid content (Falk-Petersen et al., 2009; Renaud et al., 2018; Supplementary Figure 2). The energy content of zooplankton is proportional to their lipid content, and together with species abundances indicate their share of the energy pool at the secondary trophic level. The most important storage lipid for these species is wax esters, which represent a high-energy lipid (Lee, 1975; Sargent et al., 1981; Sargent and Falk-Petersen, 1988; Vogedes et al., 2010). The biosynthesis of specific long-chained fatty acids and wax esters is a special adaptation of Calanus spp. and some other zooplankters to cope with the strong seasonality in food availability (Sargent and Falk-Petersen, 1988; Hagen and Auel, 2001; Graeve and Greenacre, 2020). Capital breeders, such as C. hyperboreus and C. glacialis, contain enough lipids for gonad maturation and egg production, and can spawn without feeding (Falk-Petersen et al., 1990; Varpe et al., 2009; Schultz et al., 2020). However, C. glacialis can switch to income breeding when food becomes available and typically use ice algae as a high-energy nourishment source during spring to ensure early gonad maturation and reproduction (Søreide et al., 2010; Leu et al., 2011). As an income breeder, C. finmarchicus is dependent on ingesting food, most preferably from phytoplankton (autotrophic protists) at the end of May-beginning of June, to fuel reproduction (Varpe et al., 2009). The increase in total lipid in Calanus from May to June may indicate that they fed on the abundant Phaeocystis pouchetii, even if this species is not a preferred diet item (Njestgaard et al., 2007). They could also have been feeding selectively on the less abundant diatoms. During the peak of the bloom, when diatoms contributed about half of the standing stock, they likely represented an important food source for the developing copepodids.
Conclusion and Outlook
Sea ice is retreating in the Arctic Ocean and the duration and thickness of ice cover are decreasing regionally, with longer periods of open water and more leads in the pack ice (Barber et al., 2015; Polyakov et al., 2017). Thinner sea ice facilitates more ice algal growth (Matrai and Apollonio, 2013), while more open water early in the season may result in less ice algae (Barber et al., 2015). Earlier sea-ice melting causes more light to penetrate in the water column and can result in early and possibly longer phytoplankton bloom below sea ice (Assmy et al., 2017; Pavlov et al., 2017). The question is whether this causes a mismatch for seasonal zooplankton ascent and reproduction (Søreide et al., 2010). Thinner ice, more melt ponds and leads imply that more light is transmitted into the water column. If this happens in early spring, it is likely to trigger an earlier “wake-up call” for zooplankton at depth. Berge et al. (2020) have shown that artificial light affects zooplankton behaviour down to 200 m depth, and changes in natural light conditions can likely be sensed by zooplankton even deeper. The winter-spring “wake-up call” for zooplankton, likely mediated by seasonal changes in light, caused zooplankton to increase rapidly in the upper water layers in late May, with reproduction timed for the onset of the plankton bloom. Thus, with less sea ice in the Arctic Ocean, we anticipate zooplankton to arrive earlier in the upper part of the water column to match earlier algal blooms. We also expect that the contribution of ice algae to zooplankton diet in the Arctic Ocean will become less important because of shortened duration of their presence and reduced availability of growth supporting substrate (the sea ice). This will likely favour, to various degrees, all three Calanus species since nutritious algal food will arrive earlier and the productive season will be longer enabling C. hyperboreus to shorten it’s life cycle. C. glacialis females to be present when food is plentiful to maximize egg production and C. finmarchicus to increase its likelihood to reproduce and grow large enough before the productive season ends. With continued ocean warming the life cycle of C. finmarchicus may be sustained in the Arctic Ocean. The deeper zooplankton fauna, below 200 m depth, is expected to persist into the future with little change in communities.
Data Availability Statement
The data are available in the Norwegian Polar Institute’s Data Centre: N-ICE2015 bottle data from ship water sampler [v2.3; includes average Pressure, Temperature & Salinity measured by the CTD from 1 sec before the bottle was closed to 1 sec after the bottle was closed): https://data.npolar.no/dataset/516bc529-3019-497d-9b27-98d13e1adc47, N-ICE2015 water column biogeochemistry (v2.0; includes Chl a in the water column): https://data.npolar.no/dataset/3ebb7f64-a045-4627-9b71-694df089a903, N-ICE2015 sea ice biogeochemistry (Includes Chl a in sea ice): https://data.npolar.no/dataset/d3e93b31-0da8-460a-827f-5df2c5525456, and N-ICE2015 Zooplankton taxonomy and abundance (Includes zooplankton taxonomy as ind. m-2 for pelagic [Multinet & WP2] and under ice zooplankton [swim net)]: https://data.npolar.no/dataset/1beee2ec-873e-44b7-881f-9f227fcee8ad.
Author Contributions
AM contributed the oceanography. AW, AB, and MH did the Zooplankton sampling with MultiNet. PL, PK, and HH were part of the scientific dive team collecting samples. SK organized the zooplankton analyses. HH, AW, and AM assembled the first draft of this manuscript, with subsequent input from co-authors. All authors contributed to the article and approved the submitted version.
Funding
This study was supported by the Centre of Ice, Climate and Ecosystems at the Norwegian Polar Institute through the N-ICE2015 project. Additional support was provided by the Ministry of Foreign Affairs, Norway, through the ID Arctic project, and the FADE project of the Arctic Ocean Flagship of the Fram Centre. AM acknowledges support from the ARC Centre of Excellence for Climate Extremes (CE170100023). Financing support was also provided from the Polish Scientific Council project KongHau_ZOOM (2375/Svalbard/2012/2).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank the captain and crew of the RV Lance, N-ICE leader Harald Steen, scientific divers Pedro Duarte (NPI), and Pernilla Carlsson (NIVA) as well as scientists and technicians that contributed to the sampling campaign in the Arctic Ocean. Mikail Itkin (NPI) is thanked for help with the map, Marvin Choquet (Nord University, Bodø) for genetic sequencing of nauplii, and Michael Greenacre (Professor of statistics, Barcelona and Tromsø) is thanked for solving dilemmas with multivariate statistics. Marta Gluchowska (Institute of Oceanology, Sopot, Poland) is thanked for sample taxonomic analyses.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2021.609480/full#supplementary-material
Supplementary Figure 1 | Depth distribution of developmental stages of (A) Calanus finmarchicus, (B) C. glacialis, and (C) C. hyperboreus. Note that the scale of the abundance (ind. m–3) changes where there are red numbers.
Supplementary Figure 2 | Total lipid (mg) in relation to prosome length (mm) for Calanus finmarchicus, C. glacialis and C. hyperboreus, with their respective stages CIII-CV and adult females (AF).
Supplementary Table 1 | Zooplankton taxa abundance (ind. m–3) for each seasonal time period (drifts).
Supplementary Table 2 | Full taxon names of the abbreviated names used in Figure 5B.
Supplementary Table 3 | Abundance of main zooplankton taxa (ind. m–2) in the Arctic Ocean at 0–50 m depth (A), 50–200 m depth (B) and 200 m to near bottom (C). Sampling was done with MultiNet, except in March (drift 2), when samples were taken by WP-2 net from 0–85 m and 85–200 m. Both nets had mesh size 200 μm. In March, RV Lance was frozen into the sea ice, the echo sounder could not be operated and the bottom depth was not recorded but estimated from maps. The bottom depth exceeded 2,500 m for all three stations (∗).
References
Ashjian, C. J., Campbell, R. G., Welch, H. E., Butler, M., and Van Keuren, D. (2003). Annual cycle in abundance, distribution, and size in relation to hydrography of important copepod species in the western Arctic Ocean. Deep Sea Res. I 50, 1235–1261. doi: 10.1016/s0967-0637(03)00129-8
Assmy, P., Fernández-Méndez, M., Duarte, P., Meyer, A., Randelhoff, A., Mundy, C. J., et al. (2017). Leads in Arctic pack ice enable early phytoplankton blooms below snow-covered sea ice. Sci. Rep. 7:40850. doi: 10.1038/srep40850
Auel, H., and Hagen, W. (2002). Mesozooplankton community structure, abundance and biomass in the central Arctic Ocean. Mar. Biol. 140, 1013–1021. doi: 10.1007/s00227-001-0775-4
Barber, D. G., Hop, H., Mundy, C. J., Else, B., Dmitrenko, I. A., and Tremblay, J. -É, et al. (2015). Selected physical, biological and biogeochemical implications of a rapidly changing Arctic Marginal Ice Zone. Prog. Oceanogr. 139, 122–150. doi: 10.1016/j.pocean.2015.09.003
Basedow, S. L., Sundfjord, A., von Appen, W.-J., Halvorsen, E., Kwasniewski, S., and Reigstad, M. (2018). Seasonal variation in transport of zooplankton into the Arctic Basin through the Atlantic Gateway, Fram Strait. Front. Mar. Sci. 5:194. doi: 10.3389/fmars.2018.00194
Berge, J., Cottier, F., Last, K. S., Varpe, Ø, Leu, E., Søreide, J., et al. (2009). Diel vertical migration of Arctic zooplankton during the polar night. Biol. Lett. 5, 69–72. doi: 10.1098/rsbl.2008.0484
Berge, J., Geoffroy, M., Daase, M., Cottier, F., Priou, P., Cohen, J. H., et al. (2020). Artificial light during the polar night disrupts Arctic fish and zooplankton behaviour down to 200 m depth. Comm. Biol. 3:102. doi: 10.1038/s42003-020-0807-6
Beszczynska-Möller, A., Fahrbach, E., Schauer, U., and Hansen, E. (2012). Variability in Atlantic water temperature and transport at the entrance to the Arctic Ocean, 1997–2010. ICES J. Mar. Sci. 69, 852–863. doi: 10.1093/icesjms/fss056
Blachowiak-Samolyk, K., Kwasniewski, S., Richardson, K., Dmoch, K., Hansen, E., Hop, H., et al. (2006). Arctic zooplankton do not perform diel vertical migration (DVM) during periods of midnight sun. Mar. Ecol. Prog. Ser. 308, 101–116. doi: 10.3354/meps308101
Blachowiak-Samolyk, K., Søreide, J. E., Kwasniewski, S., Sundfjord, A., Hop, H., Falk-Petersen, S., et al. (2008). Hydrodynamic control of mesozooplankton abundance and biomass in northern Svalbard waters (79–81 degrees N). Deep Sea Res. II 55, 2210–2224. doi: 10.1016/j.dsr2.2008.05.018
Choquet, M., Hatlebakk, M., Dhanasiri, A. K. S., Kosobokova, K., Smolina, I., Søreide, J. S., et al. (2017). Genetics redraws pelagic biogeography of Calanus. Biol. Lett. 13:588. doi: 10.1016/b978-0-12-409548-9.11633-1
Choquet, M., Kosobokva, K., Kwasniewski, S., Hatlebakk, M., Dhanasiri, A. K. S., Melle, W., et al. (2018). Can morphology reliably distinguish between the copepods Calanus finmarchicus and C. glacialis, or is DNA the only way? Limnol. Oceanogr. Methods 16, 237–252. doi: 10.1002/lom3.10240
Cohen, J. H., Berge, J., Moline, M. A., Johnsen, J., and Zolich, A. P. (2020). “Light in the Polar Night,” in Polar Night Marine Ecology, eds J. Berge, G. Johnsen, and J. H. Cohen (Cham: Springer), 37–66. doi: 10.1007/978-3-030-33208-2_3
Cohen, J. H., Berge, J., Moline, M. A., Sørensen, A. J., Last, K., Falk-Petersen, S., et al. (2015). Is ambient light during the high Arctic polar night sufficient to act as a visual cue for zooplankton? PLoS One 10:e0126247. doi: 10.1371/journal.pone.0126247
Conover, R. J., and Huntley, M. E. (1991). Copepods in ice-covered seas: distribution, adaptation to seasonally limited food, metabolism, growth patterns and life cycle strategies in polar seas. J. Mar. Syst. 2, 1–40. doi: 10.1016/0924-7963(91)90011-i
Conover, R. J., and Siferd, T. D. (1993). Dark-season survival strategies of coastal zone zooplankton in the Canadian Arctic. Arctic 46, 303–311.
Cottier, F. R., Tarling, G. A., Wold, A., and Falk-Petersen, S. (2006). Unsynchronised and synchronised vertical migration of zooplankton in a high Arctic fjord. Limnol. Oceanogr. 51, 2586–2599. doi: 10.4319/lo.2006.51.6.2586
Daase, M., and Eiane, K. (2007). Mesozooplankton distribution in northern Svalbard waters in relation to hydrography. Polar Biol. 30, 969–981. doi: 10.1007/s00300-007-0255-5
Daase, M., Eiane, K., Aksnes, D. L., and Vogedes, D. (2008). Vertical distribution of Calanus spp. and Metridia longa at four Arctic locations. Mar. Biol. Res. 4, 193–207. doi: 10.1080/17451000801907948
Daase, M., Falk-Petersen, S., Varpe, Ø, Darnis, G., Søreide, J. E., Wold, A., et al. (2013). Timing of reproductive events in the marine copepod Calanus glacialis: a pan-Arctic perspective. Can. J. Fish. Aquat. Sci. 70, 871–884. doi: 10.1139/cjfas-2012-0401
Daase, M., Søreide, J. E., and Martynova, D. (2011). Effects of food quality on naupliar development in Calanus glacialis at subzero temperatures. Mar. Ecol. Prog. Ser. 429, 111–124. doi: 10.3354/meps09075
Darnis, G., and Fortier, L. (2014). Temperature, food and the seasonal vertical migration of key arctic copepods in the thermally stratified Amundsen Gulf (Beaufort Sea, Arctic Ocean). J. Plankton Res. 36, 1092–1108. doi: 10.1093/plankt/fbu035
Darnis, G., Robert, D., Pomerleau, C., Link, H., Archambault, P., Nelson, R. J., et al. (2012). Current state and trends in Canadian Arctic marine ecosystems: II. Heterotrophic food web, pelagic-benthic coupling, and biodiversity. Clim. Change 115, 179–205. doi: 10.1007/s10584-012-0483-8
Edvardsen, A., Pedersen, J. M., Slagstad, D., Semenova, T., and Timonin, A. (2006). Distribution of overwintering Calanus in the North Norwegian Sea. Ocean Sci. 2, 87–96. doi: 10.5194/os-2-87-2006
Ershova, E. A., Kosobokova, K. N., Banas, N. S., Ellingsen, I., Niehoff, B., Hildebrandt, N., et al. (2021). Sea ice decline drives biogeographical shifts of key Calanus species in the central Arctic Ocean. Glob. Change Biol. 27, 1–16. doi: 10.1111/gcb.15562
Falk-Petersen, S., Mayzaud, P., Kattner, G., and Sargent, J. R. (2009). Lipids and life strategy of Arctic Calanus. Mar. Biol. Res. 5, 18–39. doi: 10.1080/17451000802512267
Falk-Petersen, S., Pavlov, V., Timofeev, S., and Sargent, J. R. (2007). “Climate variability and possible effects on arctic food chains: the role of Calanus,” in Arctic-Alpine Ecosystems and People in a Changing Environment, eds J. B. Ørbæk, R. Kallenborn, I. Tombre, E. N. Hegseth, S. Falk-Petersen, and A. H. Hoel (Berlin: Springer Verlag), 147–166. doi: 10.1007/978-3-540-48514-8_9
Falk-Petersen, S., Sargent, J. R., and Hopkins, C. C. E. (1990). “Trophic relationships in the pelagic arctic food web,” in Trophic Relationships in the Marine Environment, eds M. Barnes and R. N. Gibson (Aberdeen: Scotland University Press), 315–333.
Fernández-Méndez, M., Olsen, L. M., Kauko, H. M., Meyer, A., Rösel, A., Merkouriadi, I., et al. (2018). Algal hot spots in a changing Arctic Ocean: sea-ice ridges and the snow-ice interface. Front. Mar. Sci. 5:75. doi: 10.3389/fmars.2018.00075
Freese, D., Søreide, J. E., Graeve, M., and Niehoff, B. (2016). A year-round study on metabolic enzymes and body composition of the Arctic copepod Calanus glacialis: implications for the timing and intensity of diapause. Mar. Biol. 164:3.
Gabrielsen, T. M., Merkel, B., Søreide, J. E., Johansson-Karlsson, E., Bailey, A., Vogedes, D., et al. (2012). Potential misidentifications of two climate indicator species of the marine arctic ecosystem: Calanus glacialis and C. finmarchicus. Polar Biol. 35, 1621–1628. doi: 10.1007/s00300-012-1202-7
Graeve, M., and Greenacre, M. J. (2020). The selection and analysis of fatty acid ratios: a new approach for the univariate and multivariate analysis of fatty acid trophic markers in marine pelagic organisms. Limnol. Oceanogr. Methods 18, 196–210. doi: 10.1002/lom3.10360
Granskog, M. A., Assmy, P. A., Gerland, S., Spreen, G., Steen, H., and Smedsrud, L. H. (2016). Arctic research on thin ice: consequences of Arctic sea ice loss. EOS Trans. AGU 97, 22–26. doi: 10.1029/2016EO044097
Granskog, M. A., Fer, I., Rinke, A., and Steen, H. (2018). Atmosphere-ice-ocean-ecosystem processes in a thinner Arctic sea ice regime: the Norwegian Young Sea Ice (N-ICE2015) expedition. J. Geophys. Res. Oceans 123, 1586–1594. doi: 10.1002/2017JC013328
Greenacre, M. (2016). Correspondence Analysis in Practice, 3rd Edn. Boca Raton, FL: Chapman & Hall, CRC.
Häfker, N. S., Meyer, B., Last, K. S., Pond, D. W., Hueppe, L., and Teschke, M. (2017). Circadian clock involvement in zooplankton diel vertical migration. Curr. Biol. 27:2194. doi: 10.1016/j.cub.2017.06.025
Häfker, N. S., Teschke, M., Last, K. S., Pond, D. W., Huppe, L., and Mayer, B. (2018). Calanus finmarchicus seasonal cycle and diapause in relation to gene expression, physiology, and endogenous clocks. Limnol. Oceanogr. 63, 2815–2838. doi: 10.1002/lno.11011
Hagen, W., and Auel, H. (2001). Seasonal adaptations and the role of lipids in oceanic zooplankton. Zoology 104, 313–326. doi: 10.1078/0944-2006-00037
Halvorsen, E. (2015). Significance of lipid storage levels for reproductive output in the Arctic copepod Calanus hyperboreus. Mar. Ecol. Prog. Ser. 540, 259–265. doi: 10.3354/meps11528
Halvorsen, E., Tande, K., Edvardsen, A., Slagstad, D., and Pedersen, O. P. (2003). Habitat selection of overwintering Calanus finmarchicus population in the NE Norwegian Sea and shelf waters off Northern Norway in 2000-02. Fish. Oceanogr. 12, 339–351. doi: 10.1046/j.1365-2419.2003.00255.x
Harris, R. P., Irigoien, X., Head, R. N., Rey, C., Hygum, B. H., Hansen, B. W., et al. (2000). Feeding, growth, and reproduction in the genus Calanus. ICES J. Mar. Sci. 57, 1708–1726. doi: 10.1006/jmsc.2000.0959
Hirche, H.-J. (1990). Egg production of Calanus finmarchicus at low temperature. Mar. Biol. 106, 53–58. doi: 10.1007/bf02114674
Hirche, H.-J. (1997). Life cycle of the copepod Calanus hyperboreus in the Greenland Sea. Mar. Biol. 128, 607–618. doi: 10.1007/s002270050127
Hirche, H.-J., and Kosobokova, K. (2007). Distribution of Calanus finmarchicus in the northern North Atlantic and Arctic Ocean - expatriation and potential colonization. Deep Sea Res. II 54, 2729–2747. doi: 10.1016/j.dsr2.2007.08.006
Hirche, H.-J., and Kosobokova, K. N. (2011). Winter studies on zooplankton in Arctic seas: the Storfjord (Svalbard) and adjacent ice-covered Barents Sea. Mar. Biol. 158, 2359–2376. doi: 10.1007/s00227-011-1740-5
Hirche, H.-J., and Mumm, N. (1992). Distribution of dominant copepods in the Nansen Basin, Arctic Ocean, in summer. Deep Sea Res. 39, S485–S505.
Holm-Hansen, O., and Riemann, B. (1978). Chlorophyll a determination: improvements in methodology. Oikos 30, 438–447. doi: 10.2307/3543338
Hop, H., Assmy, P., Wold, A., Sundfjord, A., Daase, M., Duarte, P., et al. (2019a). Pelagic ecosystem characteristics across the Atlantic water boundary current from Rijpfjorden, Svalbard, to the Arctic Ocean during summer (2010-2014). Front. Mar. Sci. 6:181. doi: 10.3389/fmars.2019.00181
Hop, H., Wold, A., Vihtakari, M., Daase, M., Kwasniewski, S., Gluchowska, M., et al. (2019b). “Zooplankton in Kongsfjorden (1996-2016) in relation to climate change,” in The Ecosystem of Kongsfjorden, Svalbard: Advances in Polar Ecology, Vol. 2, eds H. Hop and C. Wiencke (Cham: Springer Verlag), 229–303. doi: 10.1007/978-3-319-46425-1_7
Irigoien, X. (2004). Some ideas about the role of lipids in the life cycle of Calanus finmarchicus. J. Plankton Res. 26, 259–263. doi: 10.1093/plankt/fbh030
Koenig, Z., Provost, C., Villacieros-Robineau, N., Sennéchael, N., Meyer, A., Lellouche, J.-M., et al. (2017). Atlantic waters inflow north of Svalbard: insights from IAOOS observations and Mercator Ocean global operational system during N-ICE2015. J. Geophys. Res. Oceans 122, 1254–1273. doi: 10.1002/2016JC012424
Kosobokova, K., and Hirche, H.-J. (2009). Biomass of zooplankton in the eastern Arctic Ocean–a base line study. Prog. Oceanogr. 82, 265–280. doi: 10.1016/j.pocean.2009.07.006
Kosobokova, K. N., Hanssen, H., Hirche, H.-J., and Knickmeier, K. (1998). Composition and distribution of zooplankton in the Laptev Sea and adjacent Nansen Basin during summer, 1993. Polar Biol. 19, 63–76. doi: 10.1007/s003000050216
Kosobokova, K. N., Hopcroft, R. R., and Hirche, H.-J. (2011). Patterns of zooplankton diversity through the depths of the Arctic’s central basins. Mar. Biodivers. 41, 29–50. doi: 10.1007/s12526-010-0057-9
Kosobokova, K. N., and Pertsova, N. M. (2005). Zooplankton of the deep-water part of the White Sea at the end of the hydrological winter. Oceanology 45, 819–831.
Kowalczuk, P., Meier, J., Kauko, H. M., Pavlov, A. K., Zablocka, M., Peeken, I., et al. (2017). Bio-optical properties of Arctic drift ice and surface waters north of Svalbard from winter to spring. J. Geophys Res. Oceans 122, 4634–4660. doi: 10.1002/2016jc012589
Kwasniewski, S., Hop, H., Falk-Petersen, S., and Pedersen, G. (2003). Distribution of Calanus species in Kongsfjorden, a glacial fjord in Svalbard. J. Plankton Res. 25, 1–20. doi: 10.1093/plankt/25.1.1
Lee, R. F. (1975). Lipids of Arctic zooplankton. Comp. Biochem. Phys. B. 51, 263–266. doi: 10.1016/0305-0491(75)90003-6
Leu, E., Mundy, C. J., Assmy, P., Campbell, K., Gabrielsen, T. M., Gosselin, M., et al. (2015). Arctic spring awakening–steering principles behind the phenology of vernal ice algal blooms. Prog. Oceanogr. 139, 151–170. doi: 10.1016/j.pocean.2015.07.012
Leu, E., Søreide, J. E., Hessen, D. O., Falk-Petersen, S., and Berge, J. (2011). Consequences of changing sea-ice cover for primary and secondary producers in the European Arctic shelf seas: timing, quantity, and quality. Prog. Oceanogr. 90, 18–32. doi: 10.1016/j.pocean.2011.02.004
Lischka, S., and Hagen, W. (2005). Life histories of the copepods Pseudocalanus minutus, P. acuspes (Calanoida) and Oithona similis (Cyclopoida) in the Arctic Kongsfjorden (Svalbard). Polar Biol. 28, 910–921. doi: 10.1007/s00300-005-0017-1
Matrai, P., and Apollonio, S. (2013). New estimates of microalgae production based upon nitrate reductions under sea ice in Canadian shelf seas and the Canada Basin of the Arctic Ocean. Mar. Biol. 160, 1297–1309. doi: 10.1007/s00227-013-2181-0
Meyer, A., Fer, I., Sundfjord, A., and Peterson, A. K. (2017a). Mixing rates and vertical heat fluxes north of Svalbard from Arctic winter to spring. J. Geophys. Res. Oceans 122, 4569–4586. doi: 10.1002/2016JC012441
Meyer, A., Sundfjord, A., Fer, I., Provost, C., Villacieros-Robineau, N., Koenig, Z., et al. (2017b). Winter to summer oceanographic observations in the Arctic Ocean north of Svalbard. J. Geophys. Res. Oceans 122, 6218–6237. doi: 10.1002/2016JC012391
Mumm, N., Auel, H., Hanssen, H., Hagen, W., Richter, C., and Hirche, H.-J. (1998). Breaking the ice: large-scale distribution of mesozooplankton after a decade of Arctic and transpolar cruises. Polar Biol. 20, 189–197. doi: 10.1007/s003000050295
Njestgaard, J. C., Tang, K. W., Steinke, M., Dutz, J., Koski, M., Antajan, E., et al. (2007). Zooplankton grazing on Phaeocystis: a quantitative review and future challenges. Biogeochemistry 83, 147–172. doi: 10.1007/s10533-007-9098-y
Olsen, L. M., Laney, S. R., Duarte, P., Kauko, H. M., Fernández-Méndez, M., Mundy, C. J., et al. (2017). The seeding of ice algal blooms in Arctic pack ice: the multiyear ice seed repository hypothesis. J. Geophys. Res. Biogeosci. 122, 1529–1548. doi: 10.1002/2016JG003668
Padman, L., and Dillon, T. M. (1991). Turbulent mixing near the Yermak Plateau during the coordinated Eastern Arctic Experiment. J. Geophys. Res. 96, 4769–4782. doi: 10.1029/90JC02260
Pavlov, A. K., Taskjelle, T., Kauko, H. K., Hamre, B., Hudson, S. R., Assmy, P., et al. (2017). Altered inherent optical properties and estimates of the underwater light field during an Arctic under-ice bloom of Phaeocystis pouchetii. J. Geophys. Res. Oceans 122, 4939–4961. doi: 10.1002/2016jc012471
Polyakov, I. V., Alkire, M. B., Bluhm, B. A., Brown, K. A., Carmack, E. C., Chierici, M., et al. (2020). Borealization of the Arctic Ocean in response to anomalous advection from sub-Arctic seas. Front. Mar. Sci. 7:491. doi: 10.3389/fmars.2020.00491
Polyakov, I. V., Pnyushkov, A. V., Alkire, M. B., Ashik, I. M., Baumann, T. M., Carmack, E. C., et al. (2017). Greater role for Atlantic inflows on sea-ice loss in the Eurasian Basin of the Arctic Ocean. Science 356, 285–291. doi: 10.1126/science.aai8204
Renaud, P. E., Daase, M., Banas, N. S., Gabrielsen, T. M., Søreide, J. E., and Varpe, Ø, et al. (2018). Pelagic food-webs in a changing Arctic: a trait-based perspective suggests a mode of resilience. ICES J. Mar. Sci. 75, 1871–1881. doi: 10.1093/icesjms/fsy063
Renner, A. H. H., Sundfjord, A., Janout, M. A., Ingvaldsen, R. B., Beszczynska-Möller, A., Pickart, R. S., et al. (2018). Variability and redistribution of heat in the Atlantic water boundary current north of Svalbard. J. Geophys. Res. Oceans 123, 6373–6391. doi: 10.1029/2018jc013814
Rintala, J. M., Piiparinen, J., Blomster, J., Majaneva, M., Müller, S., Uusikivi, J., et al. (2014). Fast direct melting of brackish sea-ice samples results in biologically more accurate results than slow buffered melting. Polar Biol. 37, 1811–1822. doi: 10.1007/s00300-014-1563-1
Rudels, B., Meyer, R., Fahrbach, E., Ivanov, V. V., Østerhus, S., Quadfasel, D., et al. (2000). Water mass distribution in Fram Strait and over the Yermak Plateau in summer 1997. Ann. Geophys. 18, 687–705. doi: 10.1007/s005850000216
Sargent, J., and Falk-Petersen, S. (1988). The lipid biochemistry of calanoid copepods. Hydrobiologia 167/168, 101–114. doi: 10.1007/978-94-009-3103-9_9
Schneider, C. A., Rasband, W. S., and Eliceiri, K. W. (2012). NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 9, 671–675. doi: 10.1038/nmeth.2089
Schultz, M., Nielsen, T. G., and Møller, E. F. (2020). The importance of temperature and lipid accumulation for initiation and duration of Calanus hyperboreus spawning. J. Plankton Res. 42, 159–171. doi: 10.1093/plankt/fbaa003
Smolina, I., Kollias, S., Poortvliet, M., Nielsen, T. G., Lindeque, P., Castellani, C., et al. (2014). Genome- and transcriptome assisted development of nuclear insertion/deletion markers for Calanus species (Copepoda: Calanoida) identification. Mol. Ecol. Res. 14, 1072–1079.
Søreide, J. E., Leu, E., Berge, J., Graeve, M., and Falk-Petersen, S. (2010). Timing of blooms, algal food quality and Calanus glacialis reproduction and growth in a changing Arctic. Glob. Change Biol. 16, 3154–3163. doi: 10.1111/j.1365-2486.2010.02175x
Stübner, E. I., Søreide, J. E., Reigstad, M., Marquardt, M., and Blachowiak-Samolyk, K. (2016). Year-round meroplankton dynamics in high-Arctic Svalbard. J. Plankton Res. 38, 522–536. doi: 10.1093/plankt/fbv124
Svensen, C., Wexels Riiser, C., Reigstad, M., and Seuthe, L. (2012). Degradation of copepod faecal pellets in the upper layer: role of microbial community and Calanus finmarchicus. Mar. Ecol. Prog. Ser. 462, 39–49. doi: 10.3354/meps09808
Swalethorp, R., Kjellerup, S., Dünweber, M., Nielsen, T. G., Møller, E. F., Rysgaard, S., et al. (2011). Grazing, egg production, and biochemical evidence of differences in the life strategies of Calanus finmarchicus, C. glacialis and C. hyperboreus in Disko Bay, western Greenland. Mar. Ecol. Prog. Ser. 429, 125–144. doi: 10.3354/meps09065
ter Braak, C. J. F., and Šmilauer, P. (2002). CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (version 4.5). Ithaca, NY: Microcomputer Power.
Varpe, Ø, Jørgensen, C., Tarling, G. A., and Fiksen, Ø (2009). The adaptive value of energy storage and capital breeding in seasonal environments. Oikos 118, 363–370. doi: 10.1111/j.1600-0706.2008.17036.x
Vinogradov, M. E. (1997). Some problems of vertical distribution of meso- macroplankton in the ocean. Adv. Mar. Biol. 32, 1–92. doi: 10.1016/s0065-2881(08)60015-2
Vogedes, D., Varpe, Ø, Søreide, J. E., Graeve, M., Berge, J., and Falk-Petersen, S. (2010). Lipid sac area as a proxy for individual lipid content of arctic calanoid copepods. J. Plankton Res. 32, 1471–1477. doi: 10.1093/plankt/fbq068
von Appen, W.-J., Schauer, U., Hattermann, T., and Beszczynska-Möller, A. (2016). Seasonal cycle of mesoscale instability of the West Spitsbergen current. J. Phys. Oceanogr. 46, 1231–1254. doi: 10.1175/JPO-D-15-0184.1
Wassmann, P., Kosobokova, K. N., Slagstad, D., Drinkwater, K. F., Hopcroft, R. R., Moore, S. E., et al. (2015). The contiguous domains of Arctic Ocean advection: trails of life and death. Prog. Oceanogr. 139, 42–65. doi: 10.1016/j.pocean.2015.06.011
Wassmann, P., and Reigstad, M. (2011). Future Arctic Ocean seasonal ice zones and implications for pelagic-benthic coupling. Oceanography 24, 220–231. doi: 10.5670/oceanog.2011.74
Keywords: plankton bloom, phytoplankton, ice algae, zooplankton, Calanus, seasonal migration, Arctic, sea ice
Citation: Hop H, Wold A, Meyer A, Bailey A, Hatlebakk M, Kwasniewski S, Leopold P, Kuklinski P and Søreide JE (2021) Winter-Spring Development of the Zooplankton Community Below Sea Ice in the Arctic Ocean. Front. Mar. Sci. 8:609480. doi: 10.3389/fmars.2021.609480
Received: 23 September 2020; Accepted: 21 April 2021;
Published: 04 June 2021.
Edited by:
Anne Helene Solberg Tandberg, University Museum of Bergen, NorwayReviewed by:
Hauke Flores, Alfred Wegener Institute Helmholtz Centre for Polar and Marine Research (AWI), GermanyGerald Francis Darnis, Laval University, Canada
Copyright © 2021 Hop, Wold, Meyer, Bailey, Hatlebakk, Kwasniewski, Leopold, Kuklinski and Søreide. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Haakon Hop, SGFha29uLkhvcEBucG9sYXIubm8=