
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Mar. Sci. , 28 May 2021
Sec. Deep-Sea Environments and Ecology
Volume 8 - 2021 | https://doi.org/10.3389/fmars.2021.600087
This article is part of the Research Topic Deep-sea Sponge Ecosystems: Knowledge-based Approach Towards Sustainable Management and Conservation View all 29 articles
 Joana R. Xavier1,2*David J. Rees2Raquel Pereira3Ana Colaço4,5Christopher K. Pham4,5Francisca C. Carvalho2,6
Joana R. Xavier1,2*David J. Rees2Raquel Pereira3Ana Colaço4,5Christopher K. Pham4,5Francisca C. Carvalho2,6Lithistid sponges are globally distributed in temperate and sub-tropical areas, constituting an important component of deep-sea benthic communities where they form structurally complex and vulnerable marine ecosystems (VMEs). In this study, we assess the diversity and investigate the spatial and bathymetric distribution of the lithistid sponges of the Azores archipelago (North Atlantic) based on historical records and examination of samples accidentally collected during deep-sea longline fishing operations in the region. Eleven lithistid species are recognized to occur in the Azores, including Leiodermatium tuba, recently described from material collected in several Northeast Atlantic seamounts that is hereby reported for the first time to the archipelago. We provide molecular barcodes (mtDNA COI and rRNA 28S) for seven of these species, including Discodermia ramifera, Macandrewia azorica, and Exsuperantia archipelagus, for which the Azores constitutes the type locality. We further discuss the phylogenetic and biogeographic affinities of the Azorean lithistids in the context of the Porifera classification, and the wider Northeast Atlantic upper bathyal fauna. Our study also warrants the addition of some lithistid species to the list of VME indicators for the Northeast Atlantic in support of the sustainable management and conservation of these species and habitats, as well as the ecological functions they deliver.
Lithistids are a polyphyletic group of demosponges, distributed across three orders and 13 families, which share the presence of a hypersilicified skeleton formed by an intricately articulated spicule type called desmas (Pisera and Lévi, 2002). They are found distributed globally in temperate and sub-tropical areas largely in the upper bathyal zone, and seem to be particularly prevalent in oceanic (island and seamount) environments (e.g., Schlacher-Hoenlinger et al., 2005; Carvalho et al., 2015, 2020; Schuster et al., 2018). Knowledge of the diversity, ecology and phylogenetic affinities of these sponges has greatly increased in recent years, with new species and habitats described for understudied areas (Carvalho et al., 2015, 2020; Maldonado et al., 2015; Schuster et al., 2018; Ríos et al., 2020), (phylo)diversity and distribution patterns assessed (Pomponi et al., 2001; Kelly et al., 2007; Schuster et al., 2021), and considerable changes proposed regarding their placement within the poriferan classification system proposed (Cárdenas et al., 2011; Morrow and Cárdenas, 2015; Schuster et al., 2015).
Recently, lithistid sponges were found forming structural habitats, aggregations and reef-like formations, in the upper bathyal in the western Mediterranean (Maldonado et al., 2015) and the northeast Atlantic (Ríos et al., 2020), possibly analogous to reef habitats which developed and thrived during the Mesozoic in shallower waters (Kauffman et al., 2000). While the ecological role of lithistid aggregations is yet to be investigated, sponge habitats dominated by other groups (e.g., such as geodiids and hexactinellids) were shown to serve as habitat for numerous invertebrate and fish species (e.g., Hawkes et al., 2019), also performing numerous functions in bentho-pelagic coupling and nutrient recycling (Cathalot et al., 2015; Maldonado et al., 2020; Rooks et al., 2020).
Sponge habitats meet several of the vulnerability criteria, viz. uniqueness or rarity, functional significance, fragility, low recovery potential, and structural complexity, defined for vulnerable marine ecosystems (VMEs) by the Food and Agriculture Organization within the scope of the International Guidelines for the Management of Deep-Sea Fisheries in the High Seas (FAO, 2009, ICES, 2020). Therefore, uncovering their distribution patterns is critical to inform the development of appropriate conservation measures to ensure long-term sustainability and minimize the impacts of the various anthropogenic activities operating over the deep seafloor.
In the Northeast Atlantic, the Azores archipelago constitutes the region for which the lithistid fauna has been most comprehensively investigated. Since the description of the first lithistid for the area (Macandrewia azorica Gray, 1859), nine species were reported to the archipelago largely thanks to the studies by the French sponge taxonomist Émile Topsent, resulting from the historical expeditions of Prince Albert I of Monaco between 1885 and 1915 onboard the vessels Princesse Alice I and II and Hirondelle I and II (Topsent, 1892, 1898, 1904, 1928; see also Porteiro, 2009). However, it was not until recent that new data and samples became available as a result of an increase of the exploration efforts toward the deeper environments of this archipelago. These, alongside recent studies describing the lithistids of the Macaronesian archipelagos of Madeira and Selvagens (Carvalho et al., 2015) and seamounts off the Iberian margin and South of the Azores (Carvalho et al., 2020), allow for the first time a comparison of the lithistid assemblages of the Lusitanian and Mediterranean marine provinces.
Thus, the main aims of this study were to: (i) investigate the patterns of spatial and bathymetric distribution of the lithistid sponges throughout the Azores archipelago from the integration of historical records and newly obtained data and samples; (ii) assess their diversity and phylogenetic relationships; and (iii) explore the biogeographic affinities of the species occurring on the Azores in the context of the wider Northeast Atlantic and Mediterranean bathyal fauna.
The Azores is an oceanic volcanic archipelago that lies above the Mid-Atlantic Ridge in the North Atlantic Ocean. The seafloor is mostly deep, but a large number of seamounts, a fraction of the Mid-Atlantic Ridge, and the slopes of the islands compose the shallowest parts (Perán et al., 2016). Underwater it is characterized by the presence of numerous seamounts, isolated or associated with the Mid-Atlantic ridge (Morato et al., 2008), that host a wide variety of open-ocean and deep-sea organisms. The region is considered a biodiversity hotspot for several groups of organisms, from cetaceans and other megafauna (Silva et al., 2014; Afonso et al., 2020) to benthic corals (Braga-Henriques et al., 2013; Sampaio et al., 2019). A convergence of oceanographic processes manifests in the Azores Archipelago, designated as a confluence zone between the west and the east North Atlantic. Different dominant oceanographic processes affect the west and the east groups not only by the water mass composition but also in their nutrient (climatological) profiles (Caldeira and Reis, 2017). Various water masses, e.g., the Eastern North Atlantic Central Water (ENACW), the Mediterranean Outflow Water (MOW), and the North Atlantic Deep Water (NADW) are found in the region at different depths (Bashmachnikov et al., 2015 and references therein). The Azores region is located in oligotrophic waters where primary production and carbon flux to the seafloor are lower than other Atlantic areas (Abell et al., 2013). Seafloor environmental conditions show high spatial and temporal (seasonal/inter-annual) variability (Amorim et al., 2017).
An extensive review of the sponge taxonomic literature (e.g., Gray, 1859; Topsent, 1892, 1898, 1904, 1928; Lévi and Vacelet, 1958) as well as biodiversity data portals (e.g., OBIS, GBIF) was performed and all records of lithistid sponges contained within the Azores EEZ were compiled in an excel spreadsheet following Darwin Core (DwC) data standards (Wieczorek et al., 2012). Five specimens, collected outside the EEZ, on the Irving and Plato seamounts located southern of the archipelago were also included for completeness. For each record, information on expedition (vessel, year), location (station no., coordinates and depth), sampling gear (e.g., trawl, longline), and taxonomic identification (as reported and currently accepted) was assembled (see Supplementary Material 1). Collection data of the specimens examined here and occurrences, both historical and recent, are publicly available in the PANGAEA data repository1, and in the SponGIS data portal2 developed in scope of the H2020 SponGES project3.
New material and information were collected during several scientific expeditions (2006–2008; 2010–2011; 2017), experimental fishing surveys and bycatch monitoring through fisheries observers in the course of various projects and programs, both past and ongoing in the region, such as SponGES, MERCES, Discardless, CoralFish (see also section “Acknowledgments”). Sampling was performed according to national Portuguese laws and approved by the Azorean Directorate of Sea Affairs of the Azores Autonomous region through the CCPI 03/2017/DRCT for the samples collected during the SponGES project.
Specimens were identified from analyses of external and/or internal morphological characteristics such as habitus, texture, consistency, surface ornamentation and oscula distribution, skeletal arrangement and spicular composition, following standard methods (Boury-Esnault and Rützler, 1997). Detailed morphological descriptions are not provided in this study as taxonomic studies of neighboring areas (Madeira and Selvagens archipelagos, Northeast Atlantic seamounts), including many of the species herein reported, were recently presented (Carvalho et al., 2015, 2020; Carvalho and Pisera, 2019).
Genomic DNA was extracted from small pieces of choanosomal tissue using a DNeasy Blood and Tissue kit (QIAGEN), following the instructions of the manufacturer. The Folmer region of the mitochondrial cytochrome c oxidase subunit I (COI), and the C1–D2 region of the nuclear ribosomal 28S gene were amplified and sequenced. The COI and 28S fragments were amplified using the primer pairs LCO1490 and HCO2198 (Folmer et al., 1994) or dgLCO1490 and dgHCO2198 (Meyer et al., 2005), and C1’ASTR and D2 (Lê et al., 1993; Cárdenas et al., 2010), respectively. The polymerase chain reaction conditions used follow the protocols as in Schuster et al. (2015). Obtained amplicons were purified with a solution of Exonuclease 1 (EXO, 10 U μL-1) and shrimp alkaline phosphatase (SAP, 10 U μL-1 USB®), and Sanger sequenced in both directions with Big Dye® Terminator v3.1, on a capillary-based Applied Biosystem 3730XL Analyzer. The molecular work was carried out at Biodiversity Laboratories (BDL, DNA Section) at the Department of Biological Sciences (University of Bergen). Generated sequences were deposited in GenBank under accession nos. MW006490– MW006541 for rRNA 28S (C1–D2) and MW000672– MW000696 for mtDNA COI (see Supplementary Material 2).
Obtained sequences were verified using the Basic Local Alignment Search Tool (BLAST) available at http://www.ncbi.nlm.nih.gov/BLAST/. Sequence quality checks, assembly and editing were performed in Geneious v.10.0.94 (Kearse et al., 2012) and aligned using Muscle (Edgar, 2004). Several sequences of the orders Tetractinellida Marshall, 1876 and Sphaerocladina Schrammen, 1924 were downloaded from GenBank and used in these alignments (see Supplementary Material 2, also deposited in PANGAEA5) according to the new classification of Demospongiae Sollas, 1885 (Morrow and Cárdenas, 2015). Sequences belonging to the order Sphaerocladina were used as outgroups. The final datasets comprise 192 sequences for mtDNA COI and 212 sequences for rRNA 28S, of which 25 and 48, respectively, are new. Phylogenetic reconstructions were performed for the individual markers under Bayesian Inference, as implemented in MrBayes v3.2 (Ronquist et al., 2012) and Maximum Likelihood as implemented in RaxML v.8 (Stamatakis, 2014) via the CIPRES Science Gateway V.3.36. Reconstructions under Bayesian Inference were obtained under two runs and four Metropolis-coupled Markov chains (MCMC) over 100 million generations, sampled every hundredth generation and with a burnin of 0.25, and the best-fitting model for both genes (GTR + G + I) as estimated by jModelTest v.2.1.7 (Darriba et al., 2012) calculated under the Akaike Information Criterion (AIC). For the Maximum Likelihood analysis, the GTRGAMMAI model was used and 1000 rapid bootstrapping replicates performed. The obtained phylogenetic trees were visualized in FigTree v.1.4.47 and edited in Adobe® Illustrator® (2019).
To examine the biogeographic affinities of the lithistid fauna in the context of the wider Northeast Atlantic and Mediterranean we built a presence/absence data matrix based on literature records and our own data for five oceanic archipelagos (Azores, Madeira, Selvagens, Canaries, and Cape Verde), two seamount groups (the oceanic seamounts located south of the Azores, and the Lusitanian seamounts off the Southwestern European shelf), as well as for the continental slopes of the Iberian margin, the Gulf of Cadiz and the Mediterranean Sea. This species × area matrix was then used to build a pairwise similarity matrix using the Bray-Curtis coefficient as implemented in PRIMER 7 (Clarke and Gorley, 2015).
A total of 47 individual records, representing 10 species, seven genera, distributed across seven families and two orders (Tetractinellida and Bubarida Morrow and Cárdenas, 2015), were compiled from the literature for the region. Detailed information on such records, including nomenclatural changes as verified in the World Porifera Database (Van Soest et al., 2021), can be found in Supplementary Material 1. In addition, 82 specimens collected as bycatch from bottom longline fishing operations were examined and assigned to seven species. Combined, the two datasets, elevate to 11 the total number of lithistid species reported from the Azores archipelago (see systematic list below, and following sections) (Figures 1, 2). Of these, Neophrissospongia nolitangere (Schmidt, 1870) and Macandrewia azorica Gray, 1859 were the most commonly captured, with 22 and 20 individuals collected, followed by Leiodermatium spp. (N = 25, collectively) and Exsuperantia archipelagus Carvalho and Pisera (2019) (N = 12) (Supplementary Material 1). Specimens within the genus Leiodermatium Schmidt, 1870 could not be assigned with full confidence to the two species previously reported for the Azores archipelago, i.e., Leiodermatium lynceus Schmidt (1870) and Leiodermatium pfeifferae (Carter, 1873), either because we did not find consistency in the characters traditionally used to distinguish the two species, i.e., location of the inhalant and exhalant openings (ostia and oscula), the thickness of the plate and the hispidity of its margin; or, in some instances, because we only had access to small fragments which prevented such analyses. These were therefore retained as L. cf. lynceus and L. cf. pfeifferae when they seemed more similar to one of the species, and as Leiodermatium sp. indet., following the open nomenclature standards proposed by Sigovini et al. (2016) and Horton et al. (2021), when they appeared somewhat intermediate or only small fragments were available.
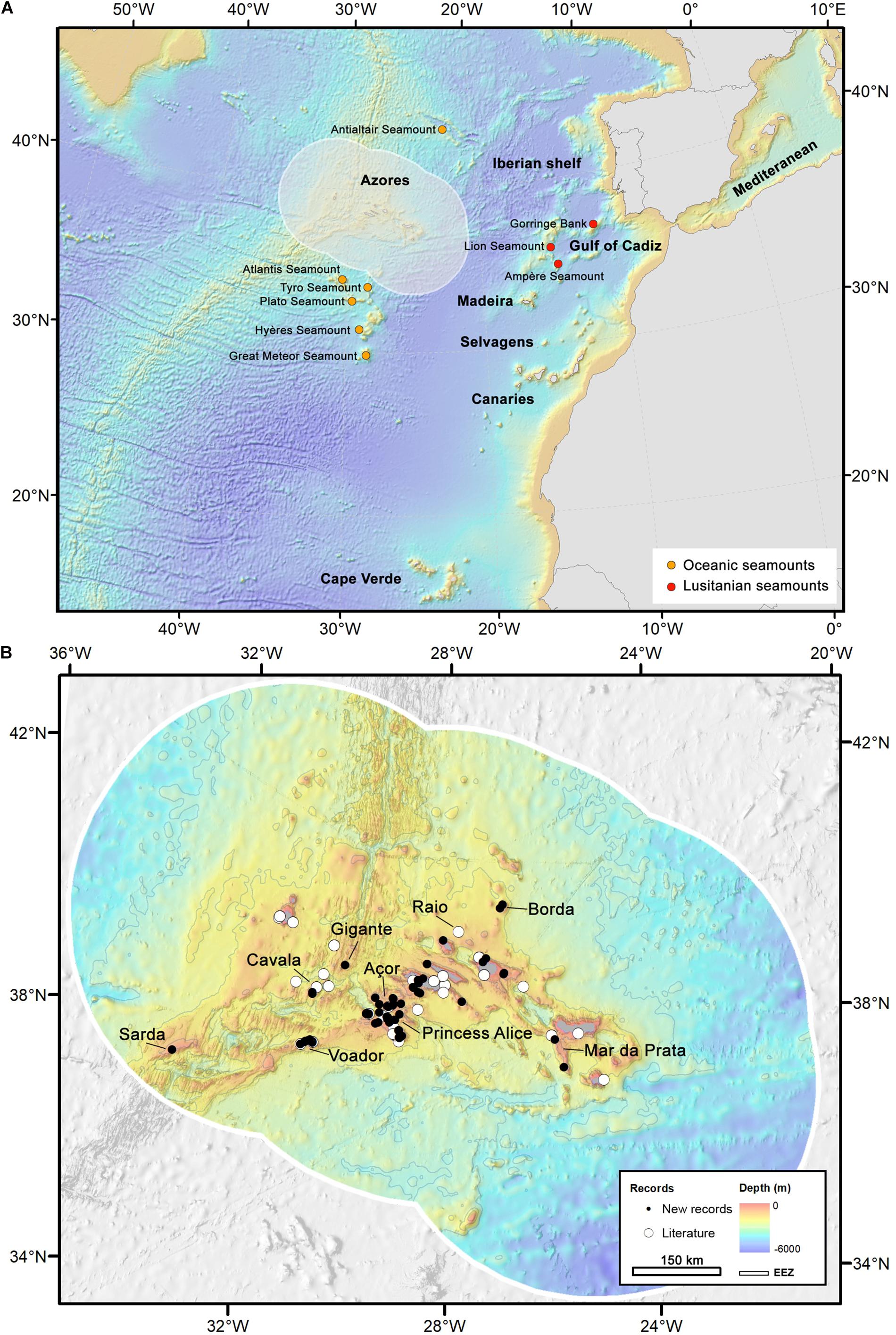
Figure 1. Map of the study area. (A) Overall geographical setting, Northeast Atlantic and Mediterranean Sea, with archipelagos and main seamounts indicated. (B) The Azores EEZ with some geomorphological features indicated, and distribution of occurrence records: white circles represent records from the literature whereas black dots represent specimens analyzed in this study.

Figure 2. Example of lithistid sponges of the Azores, collected and analyzed in the present study. (A) Neophrissospongia nolitangere (DOP 5323), (B) Discodermia ramifera (DOP 6477), (C) Exsuperantia archipelagus (DOP 6212), (D) Macandrewia azorica (DOP 6601), (E) Leiodermatium cf. lynceus (DOP 6429), (F) Leiodermatium cf. pfeifferae (DOP 11102), (G) Leiodermatium tuba (DOP 2975).
Phylum PORIFERA Grant, 1836
Class DEMOSPONGIAE Sollas, 1885
Subclass HETEROSCLEROMORPHA Cárdenas et al., 2012
Order TETRACTINELLIDA Marshall, 1876
Suborder ASTROPHORINA Sollas, 1877
Family CORALLISTIDAE Sollas, 1888
Genus Neophrissospongia Pisera and Lévi, 2002
Species Neophrissospongia nolitangere (Schmidt, 1870)
Family THEONELLIDAE Lendenfeld, 1903
Genus Discodermia du Bocage, 1869
Species Discodermia ramifera Topsent, 1892∗
Family MACANDREWIIDAE Schrammen, 1924
Genus Macandrewia Gray, 1859
Species Macandrewia azorica Gray, 1859∗
Species Macandrewia ramosa Topsent, 1904∗
Species Macandrewia robusta Topsent, 1904∗
Family PHYMARAPHINIIDAE Schrammen, 1924
Genus Exsuperantia Özdikmen, 2009
Species Exsuperantia archipelagus Carvalho and Pisera, 2019∗
Suborder SPIROPHORINA Bergquist and Hogg, 1969
Family AZORICIDAE Sollas, 1888
Genus Leiodermatium Schmidt, 1870
Species Leiodermatium (cf.) lynceus Schmidt, 1870
Species Leiodermatium (cf.) pfeifferae (Carter, 1873)
Species Leiodermatium tuba Carvalho and Xavier, 2020
Family SIPHONIDIIDAE Lendenfeld, 1903
Genus Siphonidium Schmidt, 1879
Species Siphonidium ramosum (Schmidt, 1870)
Order BUBARIDA Morrow and Cárdenas, 2015
Family DESMANTHIDAE Topsent, 1893
Genus Petromica Topsent, 1898
Subgenus Petromica (Petromica) Topsent, 1898
Species Petromica (Petromica) grimaldii Topsent, 1898∗
Highlighted in bold font are species found in the present study; ∗ denote species for which the type-locality is in the Azores archipelago.
Of the seven species identified from the material collected as bycatch from longline fisheries in the Azores, one (Leiodermatium tuba) constitutes a new record for the archipelago. Similarly, the specimens DOP 5850 and DOP 5853 collected at 461–501 m depth in the Plato seamount constitute the first record of Neophrissospongia nolitangere in this group of oceanic seamounts, thereby extending the known distribution of the species.
Overall, both historical and recent records are widely distributed across the archipelago, on island slopes and offshore seamounts, but are mostly concentrated in the central island group, south of the islands of Faial and Pico (Figures 1, 3). Except for the five specimens collected from the Plato and Irving seamounts, all specimens were collected on the Azores plateau and within the EEZ. The genus Macandrewia (family Macandrewiidae) is the most widely distributed, with the species Macandrewia azorica being found on all islands’ groups. In the Eastern island group, no records were found for N. nolitangere, Discodermia ramifera, Siphonidium ramosum, and Petromica (Petromica) grimaldii. On the other hand, in the Western island group only P. (P.) grimaldii, M. azorica, M. ramosa, and Exsuperantia archipelagus have been recorded. Several species of Leiodermatium spp., M. azorica, E. archipelagus, D. ramifera, and S. ramosum are also recorded in the topographic highs of the Mid-Atlantic Ridge between the Western and Central island groups (Figure 3).
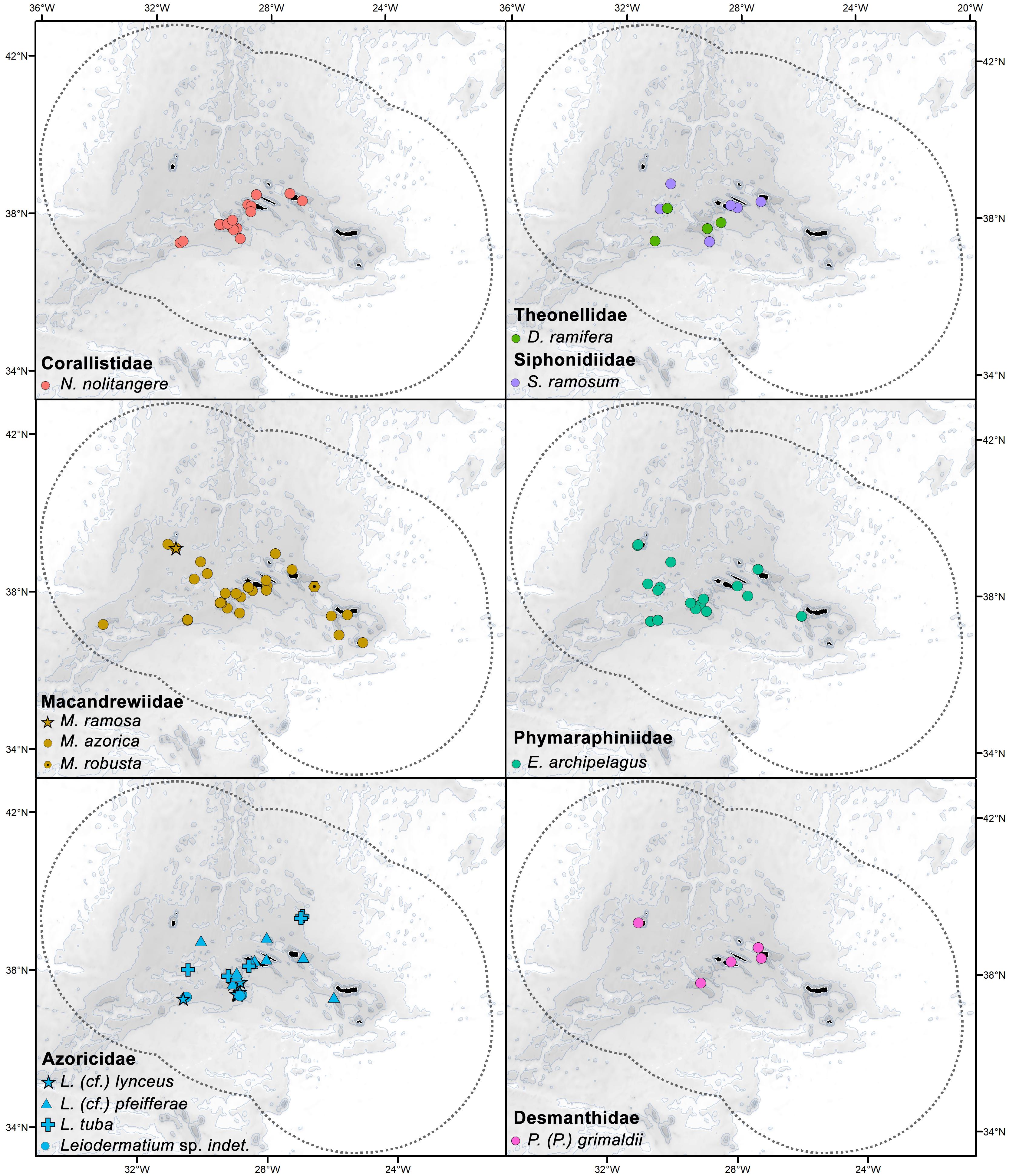
Figure 3. Distribution of lithistid species within the Azores EEZ based on literature records and analyzed specimens.
With regards to bathymetric distribution, the majority of the records (N = 82) and species (S = 9) were found in the upper bathyal between 200–800 m depth, peaking at 300–400 m depth. Comparatively, the number of records and species for the circalittoral zone above 200 m or for the lower bathyal zone below 800 m depth were much lower, with 14 records (S = 6) for the former, and 13 records (S = 5) for the latter. The shallowest record is that of D. ramifera recorded at 98 m depth east of the Açor bank, and the deepest of M. azorica found at 1495 m depth in the deep slope east of Santa Maria Island. The species N. nolitangere, D. ramifera, M. azorica, and L. (cf.) lynceus were found to be restricted to depths above 800 m, whereas E. archipelagus, L. (cf.) pfeifferae, and P. grimaldii further extend to deeper areas (down to 1250 m, depending on the species). Macandrewia ramosa and M. robusta (family Macandrewiidae) are the only two lithistid species exclusively found in the deeper zones (>1100 m depth), whereas L. tuba and S. ramosum were only found in the upper bathyal, between 622–723 m and 349–793 m depth, respectively (Figure 4 and Supplementary Material 1).
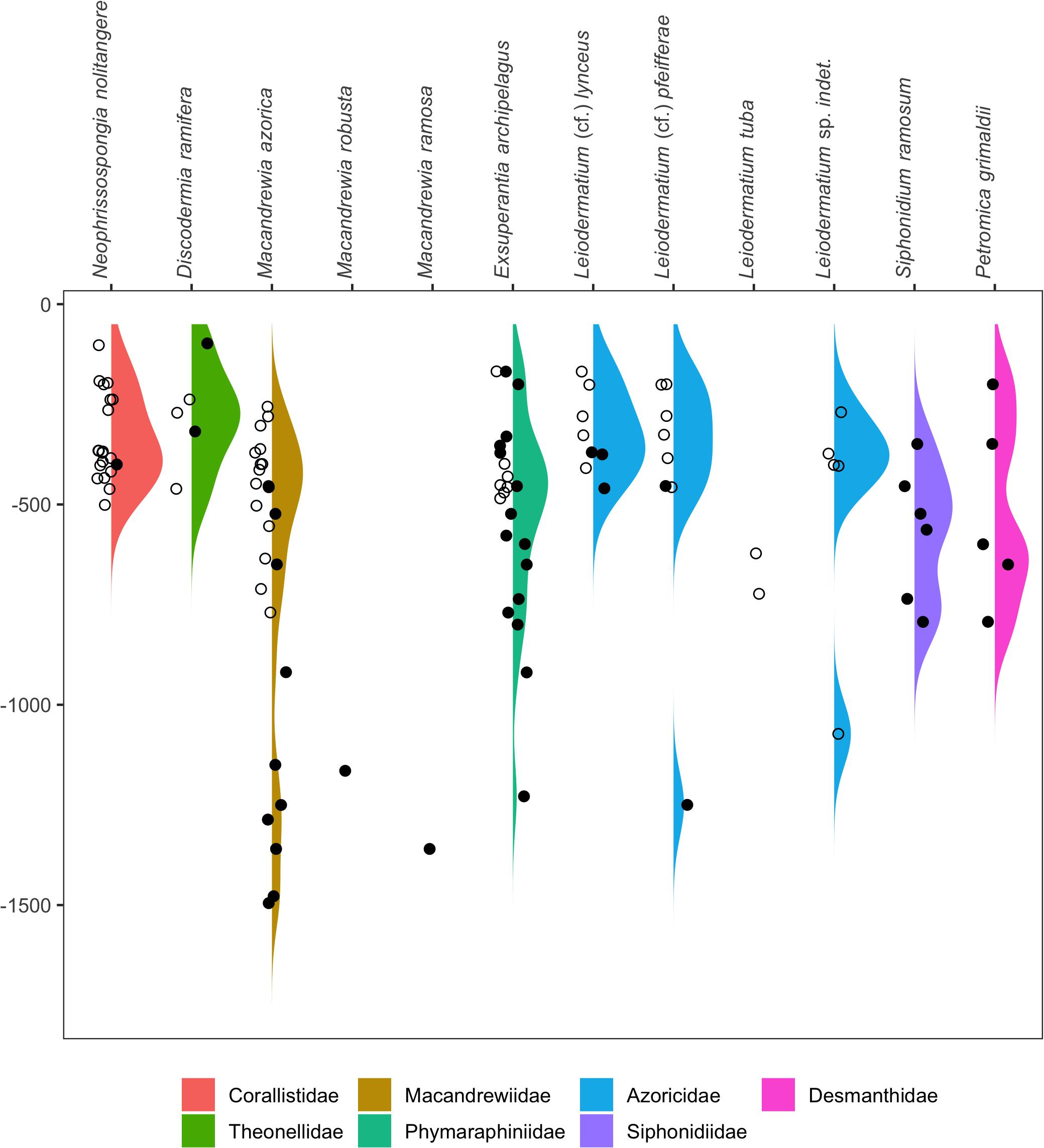
Figure 4. Ridge plots of bathymetric distribution records for each individual species based on georeferenced literature records (closed circles) and analyzed specimens (open circles). Colors correspond to each individual family.
In total, we obtained 25 mtDNA COI and 48 partial rRNA 28S sequences representing four and seven species, respectively (Supplementary Material 2). Phylogenetic trees obtained separately for the mtDNA COI and the 28S rRNA regions were largely congruent, both in overall topology as well as with high support at species and genus levels with a decrease in support toward deeper (family level) nodes (Figures 5, 6 and Supplementary Material 3).
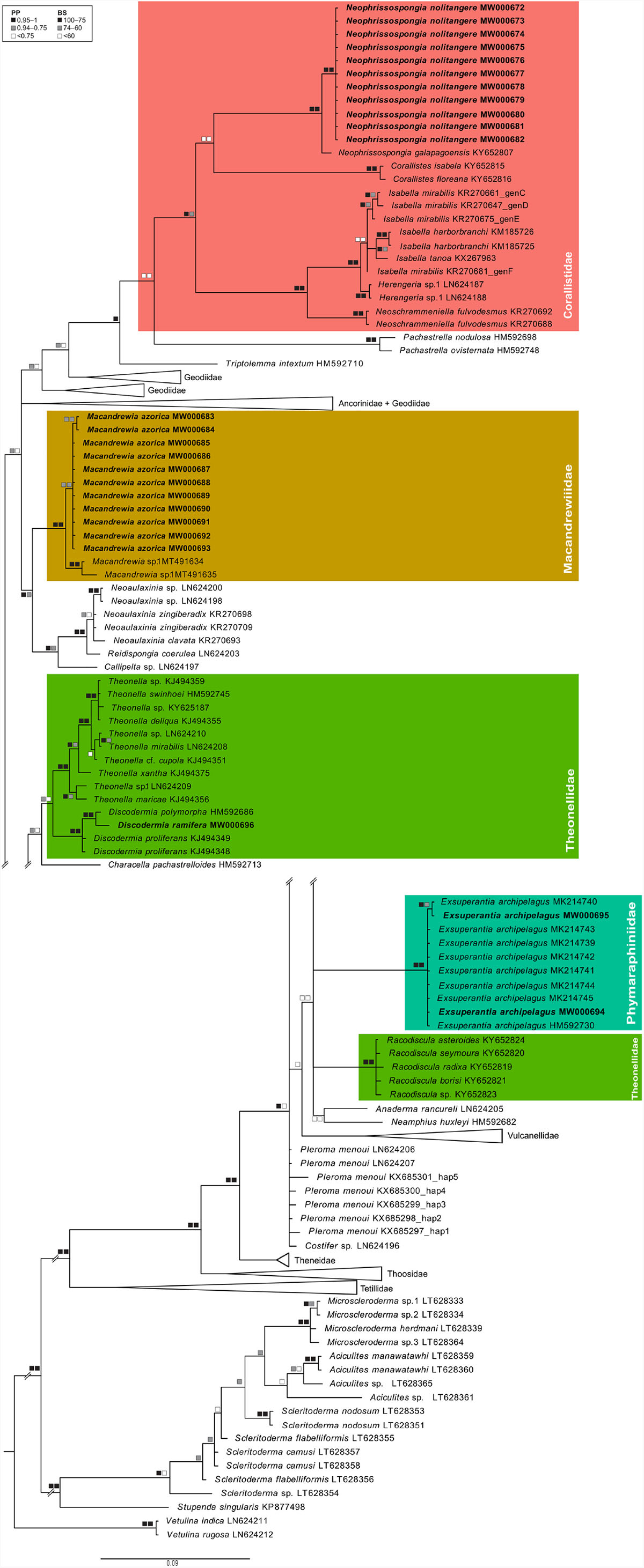
Figure 5. Phylogenetic reconstruction of the mtDNA COI gene for lithistids recorded in the Azores archipelago. Bayesian posterior probabilities (PP, left square) and maximum likelihood bootstrap support (BS, right square) values are provided on the branches. If only one square is shown, it means the clade was not retrieved in the Maximum Likelihood analysis. Sequences generated in this study are in bold. Families encompassing lithistid species found in the present study are highlighted in color.

Figure 6. Phylogenetic reconstruction of the 28S rRNA C1–D2 region for lithistids recorded in the Azores archipelago. Bayesian posterior probabilities (PP, left square) and maximum likelihood bootstrap support (BS, right square) values are given on the branches. If only one square is shown, it means the clade was not retrieved in the Maximum Likelihood analysis. Sequences generated in this study are in bold. Families encompassing lithistid species found in the present study are highlighted in color.
Within the family Macandrewiidae, all specimens assigned to Macandrewia azorica formed a clade with maximum support with a single rRNA 28S sequence of M. rigida Lévi and Lévi (1989), from the Solomon Islands, available on GenBank (PP28S = 1; BS28S = 100) (Figure 6). In the COI tree, the clade comprising all of the Azorean specimens was retrieved with only moderate support (PPCOI = 0.77; BSCOI = 62), sister to a clade formed by two sequences of unidentified Macandrewia spp. from South Africa, within a highly supported Macandrewia sp. clade (PPCOI = 1; BSCOI = 79). Within the Azorean Macandrewia clade, two sequences (DOP 2866 and DOP 6249) formed a separate but only moderately supported sub-clade (PPCOI = 0.80, BSCOI = 64) (Figure 5).
Sequences obtained for Discodermia ramifera were retrieved in a highly supported clade (PP28S = 1; BS28S = 93; PPCOI = 1, BSCOI = 98) with the Mediterranean species D. polymorpha Pisera and Vacelet (2011) in an equally monophyletic Discodermia spp. clade (PP28S = 1, BS28S = 93; PPCOI = 1; BSCOI = 98), sister to Theonella spp. Similarly, all sequences of Exsuperantia archipelagus were grouped into a clade with maximum support (PP28S = 1; BS28S = 100; PPCOI = 1; BSCOI = 100) together with sequences (available in GenBank) previously obtained for this archipelago and the seamounts located south of the Azores. In the COI tree, some sub-structuring was observed with two sequences, presenting 1 bp difference to the remaining ones, forming a sub-clade strongly supported in the Bayesian analysis (PPCOI = 0.97) and moderately supported by maximum likelihood (BSCOI = 64) (Figures 5, 6).
In the COI tree, all specimens of Neophrissospongia nolitangere collected in the Azores archipelago formed a highly supported clade (PPCOI = 0.98, BSCOI = 99) sister to N. galapagoensis Schuster, 2018 from the Galapagos islands (Figure 5). However, in the 28S tree and with the addition of two sequences of Neophrissospongia spp. from Western Australia, the Azorean clade is not supported (PP28S = 0.53, BS28S = 78) (Figure 6). This Neophrissospongia spp. clade is retrieved as sister to a larger clade comprising the genera Herengeria Lévi and Lévi (1988), Corallistes Schmidt (1870), Neoschrammeniella Pisera and Lévi (2002), and Isabella Schlacher-Hoenlinger et al. (2005) within family Corallistidae.
For the Spirophorina suborder we were only able to obtain 28S sequences and all attempts at amplifying the mtDNA COI marker failed. Within this suborder, our reconstructions show all our sequences of Leiodermatium spp. in a highly supported clade (PP28S = 1; BS28S = 97), within which two groups are found: one clade grouping specimens identified as L. cf. pfeifferae, L. cf. lynceus and Leiodermatium sp. (PP28S = 0.98, BS28S = 85) and one group with specimens identified as L. tuba (DOP 4862, DOP 11254, DOP 11445) only strongly supported in the maximum likelihood analysis (PP28S = 0.55, BS28S = 84). Leiodermatium spp. is found as sister to Aciculites spp., but only with moderate to low support (PP28S = 0.83, BS28S = 59). A monophyletic clade comprising representatives of the genera Leiodermatium (family Azoricidae) and Aciculites Schmidt, 1879 + Scleritoderma Sollas, 1888 + Microscleroderma Kirkpatrick (1903) (family Scleritodermidae Sollas, 1888) is retrieved with maximum support in our Bayesian analysis (PP28S = 1) but only moderately supported in the maximum likelihood analysis (BS28S = 69) (Figure 6). Unfortunately, no specimens, and therefore no sequences were obtained for Macandrewia robusta and M. ramosa (family Macandrewiidae), Siphonidium ramosum (family Siphonidiidae) or Petromica (Petromica) grimaldii (family Desmanthidae), all known to occur in the archipelago.
Examining the faunal similarities across the Northeast Atlantic and the Mediterranean Sea, we found the Azores lithistid assemblage to be most similar (51.9%) to the one occurring in the oceanic seamounts located to the south of the archipelago with seven species shared. This was followed by the Canaries and Madeira archipelagos, with 45.5 and 44.4% (five and four species shared), respectively. The lowest levels of similarity were found between the Azores and Iberian slope (no species shared) and the Cape Verde archipelago with which it only shares Leiodermatium pfeifferae (Figure 7, Table 1, and see also Supplementary Material 4). Of the 38 lithistid species occurring in the Northeast Atlantic and Mediterranean Sea, only one (Macandrewia ramosa Topsent, 1904) has been exclusively reported in the Azores (Table 1).
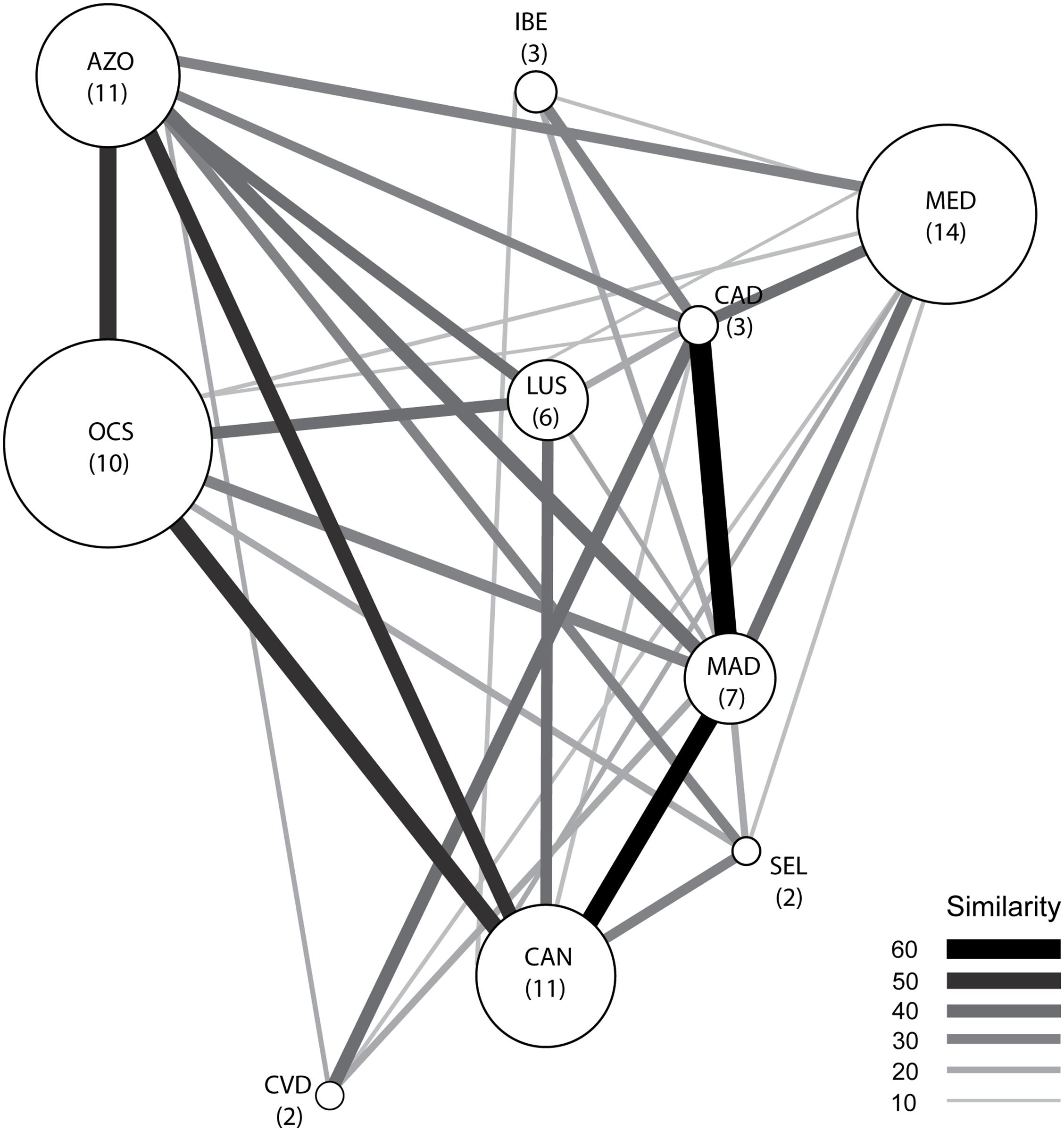
Figure 7. Network of the lithistid faunal similarities across the Northeast Atlantic and Mediterranean Sea. Size of the nodes (areas) is proportional to the number of species in that area (indicated between brackets), whereas thickness and color of the lines between nodes represents Bray–Curtis similarity (%). Areas codes: AZO, Azores; MAD, Madeira; SEL, Selvagens; CAN, Canaries; CVD, Cape Verde; OCS, oceanic seamounts; LUS, Lusitanian seamounts; IBE, Iberian Atlantic margin; CAD, Gulf of Cadiz; MED, Mediterranean Sea.
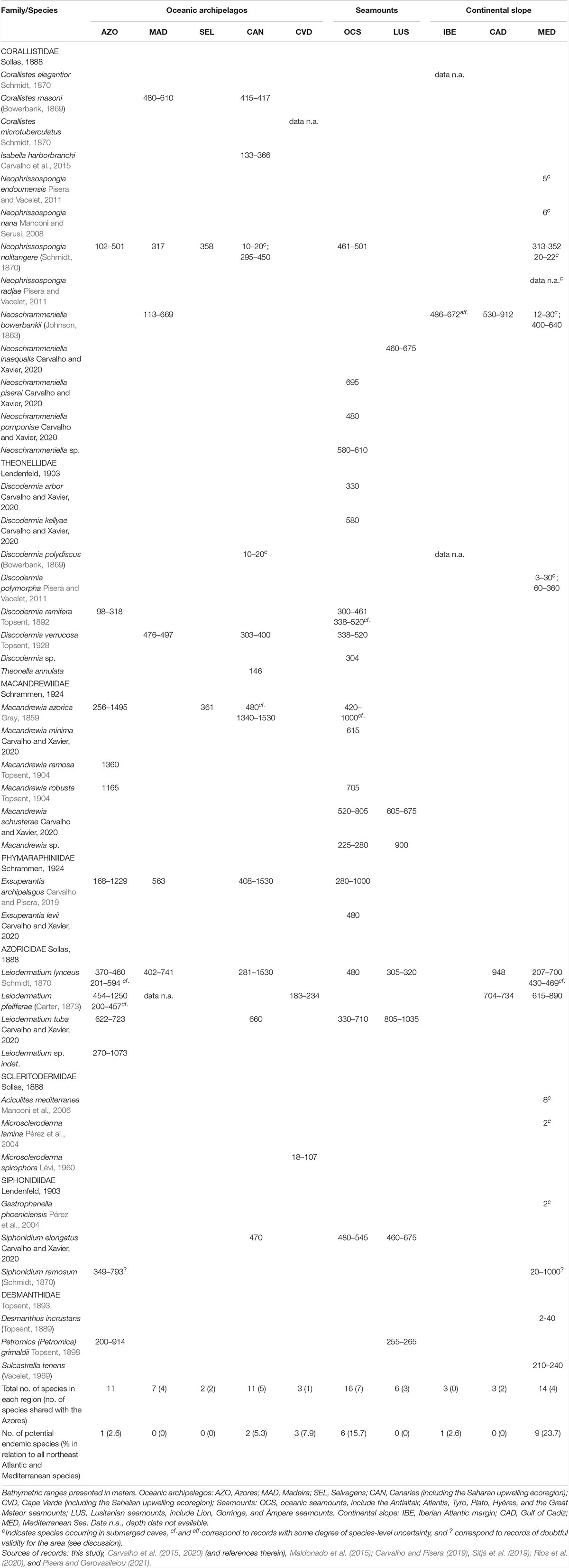
Table 1. Spatial and bathymetric distribution of lithistid sponges (Porifera, Demospongiae, Heteroscleromorpha) across the Northeast Atlantic and Mediterranean Sea.
From the integration of historical records and newly obtained data and samples we expanded considerably our knowledge of the diversity and spatial distribution of an important and vulnerable group of deep-sea sponges (lithistid demosponges) in the Azores archipelago. Leiodermatium tuba, recently described from material collected on various oceanic seamounts located South of the Azores as well as on the Gorringe Bank and the Canaries (Carvalho et al., 2020), is reported here for the first time raising to 11 the number of lithistids in the Azores archipelago. When compared with other areas of the Atlanto-Mediterranean region this diversity is the third highest, only lower than that found in the oceanic seamounts (16 species) and the Mediterranean Sea (14 species) and similar to the richness found in the Canaries. Together, the Azores and the oceanic seamounts harbor 20 species, i.e., 54% of all lithistid species of this region, and may therefore be considered a lithistid diversity hotspot of the central North Atlantic. However, such diversity is still modest when compared with the tropical Western Atlantic, a region where latest estimates suggest the occurrence of at least 88 lithistid species (Schuster et al., 2021) and that is also recognized by its high Poriferan richness overall (Van Soest et al., 2012). Considering that the Azores harbor an additional 270+ (non-lithistid) deep-sea sponge species (Topsent, 1904, Xavier et al. unpublished data), as well as a rich cold-water coral fauna (Braga-Henriques et al., 2013; Sampaio et al., 2019) further emphasizes the value of this region as a reservoir of deep-sea biodiversity.
In the Azores, the spatial patterns of distribution, with a considerably higher number of records southwest of Faial and Pico islands (central island group) reflect collection efforts. This area has a complex topography and comprises several seamounts (e.g., Açor and Princesse Alice banks) which were surveyed during historical expeditions and are traditionally used as fishing grounds of the small-scale longline fisheries fleet in the region.
The Azorean lithistid fauna is composed of a mix of steno- and eurybathic species, mostly overlapping in upper bathyal depths between 200 and 600 m. In general, distribution ranges observed for the various species in the Azores are similar to those reported for other northeast Atlantic archipelagos and seamounts (e.g., Carvalho et al., 2015, 2020, see also Table 1). However, it is important to note the uneven number of records for some species (e.g., only two records for Macandrewia ramosa and M. robusta below 1100 m in the literature), as well as some uncertainty regarding the taxonomic identification of some specimens both examined in this and previous studies (e.g., Leiodermatium spp. and M. azorica), which may be confounding observed spatial and bathymetric ranges.
The highest species diversity and number of records found in the upper bathyal (200–800 m) suggests that oceanographic conditions may be optimal at this depth range for lithistids. The pattern here observed for lithistids agrees well with previous findings of highest diversity of cold-water corals between 300 and 900 m in the archipelago (Braga-Henriques et al., 2013). However, this pattern may also partly result from considerably higher collection efforts in this depth range targeted by the fisheries from which the specimens of both studies have been obtained.
While we lack high-resolution data to investigate which environmental factors drive the distribution of the various species or the assemblage as a whole, it is accepted that availability and diversity of geomorphological features with steep slopes and limited sedimentation (island slopes, near shore banks and offshore seamounts) play an important role in providing the hard substrate needed for lithistid settlement and growth (Pomponi et al., 2001; Kelly et al., 2007). Investigating the role of other factors such as depth, temperature, salinity, and nutrients shown to be important predictors for the distribution of deep-sea sponge species and communities (e.g., Knudby et al., 2013; Howell et al., 2016; Alvarez et al., 2017; Murillo et al., 2018) will be critical to better understand the ecology of this important group.
Phylogenetic reconstructions based on two molecular markers traditionally used in the group, i.e., the mitochondrial COI and the ribosomal 28S, confirmed our morphology-based assignments, corroborated previous findings regarding the classification and phyletic status of some groups (e.g., Carvalho and Pisera, 2019; Schuster et al., 2021) and further shed light into some long-standing species-level taxonomic problems. In the majority of cases, specimens assigned to a given species formed species-level monophyletic clades or clustered with GenBank sequences of other congeneric species. This was the case for Neophrissospongia nolitangere, Macandrewia azorica, Discodermia ramifera, and Exsuperantia archipelagus.
In N. nolitangere, all specimens had identical (COI) or near-identical (28S) sequences and formed a robust clade with the Pacific N. galapagoensis. In the future it is important to compare the Azores specimens with the two new, as yet undescribed, species found in the Caribbean (Schuster et al., 2021) as these may prove much more closely related. Similarly, all our M. azorica specimens form a clade consistent with a species level relationship, although the phylogenetic support was only moderate in the COI tree. The 1 bp difference found in COI for two specimens (DOP 2866 and DOP 6249) is likely to reflect some intraspecific genetic variability associated with the depth at which the specimens were found (635 and 711 m), considerably deeper than most other specimens. The 1 bp difference found in COI for one specimen of E. archipelagus confirms the variability previously reported for this species by Carvalho and Pisera (2019). In this case, no distinctive patterns were found in relation to spatial or bathymetric distribution. Such small levels of intraspecific genetic variability, while not commonly reported, have been previously found in other tetractinellid sponges, such as Geodia barretti Bowerbank, 1858 (Cárdenas et al., 2013). Comparatively higher levels of variation in COI were also found in the genus Isabella from the Norfolk Ridge (off New Caledonia), warranting the description of a new species (Isabella tanoa Ekins et al., 2016) and the recognition of a morphologically cryptic species complex for Isabella mirabilis Schlacher-Hoenlinger et al. (2005; Ekins et al., 2016).
A close relationship of Discodermia ramifera with the Mediterranean D. polymorpha was strongly supported, in comparison to another congener from the southwest Pacific (D. proliferans Lévi and Lévi, 1983). Unfortunately, no sequences were available of other Discodermia spp. from the Northeast Atlantic [D. arbor Carvalho and Xavier, 2020, D. kellyae Carvalho and Xavier 2020, D. polydiscus (Bowerbank, 1869) and D. verrucosa Topsent, 1928] which would allow to better understand their phylogenetic and biogeographic relationships.
An important finding of our study was the fact that neither through morphological examination nor molecular data we were able to confidently assign our Leiodermatium specimens to the two species – Leiodermatium lynceus and Leiodermatium pfeifferae (see section “Results”) – long reported to occur in the region (Topsent, 1892, 1904; Lévi and Vacelet, 1958) and throughout the Northeast Atlantic and Mediterranean Sea. In fact, all specimens which morphologically seemed most similar to L. pfeifferae or L. lynceus, and those which we could not assign to either species (Leiodermatium sp. indet.), grouped in a strongly supported clade in our 28S tree. Similar to previous studies, we failed to obtain COI sequences for any of our Leiodermatium specimens, further corroborating the hypothesis that a type I intron, similar to those found in other genera of the family Scleritodermidae (Schuster et al., 2017) may also be present in this genus as also suggested by Schuster et al. (2021).
Our results suggest that the Azorean Leiodermatium sp. indet. specimens, and possibly other records previously assigned in the literature to L. pfeifferae and L. lynceus, may represent either (i) a single species of Leiodermatium with some morphological variability; (ii) a complex of species where reduced genetic differentiation and morphological plasticity hamper unequivocal species-level discrimination, or (iii) a case of hybridization or incomplete lineage sorting between closely related species. If the former hypothesis is confirmed, L. pfeifferae would become a junior synonym of L. lynceus, type-species of the genus, following the priority principle (Article 23) of the International Code of Zoological Nomenclature (ICZN, 1999). However, a detailed taxonomic revision including examination of additional specimens collected from both species’ type-localities (Portugal mainland slope for L. lynceus and Madeira island for L. pfeifferae), and other Northeast Atlantic localities, as well as an improved molecular dataset including other mitochondrial and nuclear markers, would be needed to support this or other nomenclatural act(s). A future comparison, both morphological and molecular, with the eight Leiodermatium species recently proposed by Schuster et al. (2021) for the tropical western Atlantic would further allow a complete review of the systematics and biogeography of this genus in the Atlantic.
The Azores lithistid fauna is largely composed of species with a broader northeast Atlantic or Atlanto-Mediterranean distribution, and very low endemism (one single species). This is in accordance with previous findings both for sponges (Xavier and Van Soest, 2012) and other taxonomic groups in shallow waters (Freitas et al., 2019), as well as for cold-water corals in which a putative endemism of 14–15% was observed in the region (Braga-Henriques et al., 2013; Sampaio et al., 2019). Comparably low endemism levels have also been reported for the New Zealand lithistid assemblages at similar spatial scales (Kelly et al., 2007).
The highest similarity found between the Azores and the oceanic seamounts lithistid assemblages was expected given the geographical proximity and similar environmental conditions (temperature, salinity, oxygen and major nutrients) observed at the seafloor (Amorim et al., 2017). The occurrence of Leiodermatium tuba in the Azores and of Neophrissospongia nolitangere on these oceanic seamounts, herein reported for the first time, further suggests that as surveys and taxonomic efforts increase, more species will be found to be shared between these two regions, and possibly other, as previously posited by Carvalho et al. (2020).
The upper bathyal (200–1000 m depth) of the area comprising the Azores archipelago and the oceanic seamounts, where the vast majority of the lithistid records are found, is dominated by the North Atlantic Central Water, which presents a salinity of 35.2 – 36.7 PSU, temperatures between 8.5°–19°C and high median concentration of oxygen (94%) (Palma et al., 2012; Bashmachnikov et al., 2015). Below this depth there is only a limited influence of Subarctic Intermediate Water and Mediterranean Outflow Water below this depth (Bashmachnikov et al., 2015).
The major North Atlantic water masses and current pathways have been shown to structure the diversity and distribution of deep-sea sponges and corals, as well as the habitats they form (Puerta et al., 2020; Roberts et al., 2021). The large-scale circulation of the temperate northeast Atlantic is dominated by the Azores Current (AzC) that flows eastwards toward the Iberian margin (Carracedo et al., 2014). Importantly, the AzC southward and eastward branches have been identified as an important transport pathway between the Azores archipelago and the oceanic seamounts of the Great Meteor complex and the Lusitanian seamounts, respectively (Lima et al., 2020). The AzC therefore provides the physical connectivity needed for the dispersal of sponge larvae between these regions. However, the considerably largest distances between the Azores, the Lusitanian seamounts and Iberian shelf, over abyssal areas is likely to limit successful dispersal of the generally short-lived and lecitotrophic larvae typical of sponges, resulting in a much lower similarity between these regional assemblages.
Our results suggest that the complex and dynamic oceanographic circulation patterns of the northeast Atlantic, as well as a rich topography with numerous seamounts provide the conditions for these typically bathyal species to disperse and maintain regional-scale distributions and low endemism. This has in fact been observed in other tetractinellid groups (Cárdenas et al., 2013; Cárdenas and Rapp, 2015) and cladorhizids (Hestetun et al., 2015, 2017) with similar depth distributions.
In our study, the vast majority (87%) of the lithistid records obtained as a result of bycatch of longline fisheries corresponded to large and potentially habitat-building species (Neophrissospongia nolitangere, Macandrewia azorica, and Leiodermatium spp.). This finding suggests that sponge species attaining large sizes, with a more complex morphology and with a dense hypersilicified skeleton may be more vulnerable to this activity than smaller species (Discodermia ramifera), or those with looser or softer skeletal structure [Petromica (Petromica) grimaldii]. These results concur with previous studies which showed that although bottom longline fishing has a reduced impact on deep-sea corals (another VME-indicator taxon) when compared to bottom trawling, the removal rates were significantly higher in corals with more complex morphology (Sampaio et al., 2012; Pham et al., 2014).
Notably, deep-sea sponges are presumed to exhibit slow growth rates and high longevity (Leys and Lauzon, 1998; Fallon et al., 2010; Kahn et al., 2016), which renders them particularly vulnerable to both environmental and anthropogenic disturbances. An age between 135 and 160 years was established for a specimen of Corallistes undulatus Lévi and Lévi (1983) (family Corallistidae), roughly 30 cm in diameter, collected on the Norfolk Ridge (Ellwood et al., 2007). Accidental longline capture of specimens of similar and larger sizes, thus possibly centennial in age, in the Azores is not uncommon. So, while the impact of bottom longline fisheries may be negligible in terms of the number of individuals removed from the seafloor, over time its selectivity for larger/older individuals may cause shifts in community composition and population structure and consequently alter or compromise the function(s) of the ecosystem at local scales.
Taking such considerations into account, our results further support the recent addition of lithistid tetractinellids, particularly those in the genera Leiodermatium, Neophrissospongia, Neoschrammeniella, and Macandrewia to the list of VME indicators for the Northeast Atlantic (ICES, 2020), and their proposed inclusion in Annex III of the EU deep-sea access regulations (EU Reg. 2016/2336) toward preventing significant adverse impacts on these VMEs (EU, 2016).
Whether these lithistid species form dense habitats as those found dominated by Leiodermatium pfeifferae on seamounts in the Western Mediterranean (Maldonado et al., 2015), and suspected for M. azorica on the Hyères seamount (Carvalho et al., 2020) requires verification using in situ observation systems such as remotely operated or autonomous underwater vehicles (ROVs/AUVs), and cameras.
Larger scale investigations, currently underway, of the patterns of diversity and distribution of lithistids across the entire North Atlantic, as well as their underpinning ecological and evolutionary drivers will allow to coordinate efforts toward conservation of these species, habitats and the ecological functions they deliver.
The datasets generated and analyzed in this study can be found in the PANGAEA data repository (https://doi.pangaea.de/10.1594/PANGAEA.921772 and https://doi.pangaea.de/10.1594/PANGAEA.929934), and in the SponGIS data portal (www.spongis.org). Sequences are deposited in GenBank under accession nos. MW006490–MW006541 for rRNA 28S (C1–D2) and MW000672–MW000696 for mtDNA COI gene fragments.
JX and FC designed the study. JX, RP, and FC identified the samples. DR and FC generated the sequence data. JX, RP, CP, and FC analyzed the data. AC and CP contributed the samples and data. JX wrote the draft of the manuscript. JX, RP, CP, and FC prepared the figures and tables. All the authors contributed the text and reviewed the manuscript.
This research has been performed in the scope of the SponGES project, which received funding from the European Union’s Horizon 2020 Research and Innovation Programme under grant agreement No. 679849. This document reflects only the authors’ views and the Executive Agency for Small and Medium-sized Enterprises (EASME) is not responsible for any use that may be made of the information it contains. JX research is further supported by national funds through FCT Foundation for Science and Technology within the scope of UIDB/04423/2020, UIDP/04423/2020, and CEECIND/00577/2018. RP research is supported by the Swedish Taxonomic Species Initiative (Svenska artprojektet) dha 159/09 1.4 grant. AC is supported by Foundation for Science and Technology (FCT) investigator contract IF/00029/2014/CP1230/CT0002. AC and CP are supported through the strategic project UIDB/05634/2020 and UIDP/05634/2020, and through the project FunAzores-ACORES 01-0145-FEDER-000123.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank all fishers, fisheries observers, students, and technicians involved in the collection and curation of the specimens, and particularly to Cristina Gutiérrez-Zárate and Íris Sampaio for their support with the specimens and data from the COLETA. Filipe Mora Porteiro (IMAR/Okeanos, University of the Azores) is also acknowledged for his support in the early stages of this study. We further thank our colleague Astrid Schuster (CIIMAR and University of Southern Denmark) for support with the translation from German of some original descriptions. We acknowledge the two reviewers for their thoughtful suggestions that improved the manuscript. This study is dedicated to our dear colleague and friend Hans Tore Rapp, sponge scientist and deep-sea explorer, whom we deeply miss.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2021.600087/full#supplementary-material
Abell, R. E., Brand, T., Dale, A. C., Tilstone, G. H., and Beveridge, C. (2013). Variability of particulate flux over the Mid-Atlantic ridge. Deep Sea Res. Part II Top. Stud. Oceanogr. 98, 257–268. doi: 10.1016/j.dsr2.2013.10.005
Afonso, P., Fontes, J., Giacomello, E., Magalhães, M. C., Martins, H. R., Morato, T., et al. (2020). The Azores: a mid-Atlantic hotspot for marine megafauna research and conservation. Front. Mari. Sci. 6:826. doi: 10.3389/fmars.2019.00826
Alvarez, B., Frings, P. J., Clymans, W., Fontorbe, G., and Conley, D. J. (2017). Assessing the Potential of Sponges (Porifera) as indicators of ocean dissolved si concentrations. Front. Mar. Sci. 4:373. doi: 10.3389/fmars.2017.00373
Amorim, P., Perán, A. D., Pham, C. K., Juliano, M., Cardigos, F., Tempera, F., et al. (2017). Overview of the ocean climatology and its variability in the Azores region of the North Atlantic including environmental characteristics at the seabed. Front. Mar. Sci. 4:56. doi: 10.3389/fmars.2017.00056
Bashmachnikov, I., Nascimento, Å., Neves, F., Menezes, T., and Koldunov, N. V. (2015). Distribution of intermediate water masses in the subtropical northeast Atlantic. Ocean Sci. 11, 803–827. doi: 10.5194/os-11-803-2015
Bergquist, P. R., and Hogg, J. J. (1969). Free amino acid patterns in Demospongiae: a biochemical approach to sponge classification. Cahiers de Biologie Marine 10, 205–220.
Boury-Esnault, N., and Rützler, K. (1997). Thesaurus of Sponge Morphology: Smithsonian Contributions to Zoology. Washington, DC: Smithsonian Institution Press.
Bowerbank, J. S. (1869). A Monograph of the Siliceo-fibrous Sponges. Part I. Proc. Zool. Soc. London 1, 66–100. doi: 10.1111/j.1469-7998.1869.tb07296.x
Braga-Henriques, A., Porteiro, F. M., Ribeiro, P. A., de Matos, V., Sampaio, Í, Ocana, O., et al. (2013). Diversity, distribution and spatial structure of the cold-water coral fauna of the Azores (NE Atlantic). Biogeosciences 10, 529–590. doi: 10.5194/bgd-10-529-2013
Caldeira, R. M. A., and Reis, J. C. (2017). The azores confluence zone. Front. Mar. Sci. 4:37. doi: 10.3389/fmars.2017.00037
Cárdenas, P., Pérez, T., and Boury-Esnault, N. (2012). “Sponge systematics facing new challenges,” in Advances in Sponge Science: Phylogeny, Systematics, Ecology, eds M. A. Becerro, M. J. Uriz, M. Maldonado, and X. Turon (Amsterdam: Elsevier), 79–209. doi: 10.1016/B978-0-12-387787-1.00010-6
Cárdenas, P., and Rapp, H. T. (2015). Demosponges from the northern Mid-Atlantic Ridge shed more light on the diversity and biogeography of North Atlantic deep-sea sponges. J. Mar. Biol. Assoc. U.K. 95, 1475–1516. doi: 10.1017/S0025315415000983P
Cárdenas, P., Rapp, H. T., Klitgaard, A. B., Best, M., Thollesson, M., and Tendal, O. S. (2013). Taxonomy, biogeography and DNA barcodes of Geodia species (Porifera, Demospongiae, Tetractinellida) in the Atlantic boreo-arctic region. Zool. J. Linnean Soc. 169, 251–311. doi: 10.1111/zoj.12056
Cárdenas, P., Rapp, H. T., Schander, C., and Tendal, O. S. (2010). Molecular taxonomy and phylogeny of the Geodiidae (Porifera, Demospongiae, Astrophorida) - combining phylogenetic and Linnaean classification. Zool. Scripta 39, 89–106. doi: 10.1111/j.1463-6409.2009.00402.x
Cárdenas, P., Xavier, J. R., Reveillaud, J., Schander, C., and Rapp, H. T. (2011). Molecular phylogeny of the Astrophorida (Porifera, Demospongiaep) reveals an unexpected high level of spicule homoplasy. PLoS One 6:e18318. doi: 10.1371/journal.pone.0018318
Carracedo, L. I., Gilcoto, M., Mercier, H., and Pérez, F. F. (2014). Seasonal dynamics in the Azores-Gibraltar Strait region: a climatologically-based study. Prog. Oceanogr. 122, 116–130. doi: 10.1016/j.pocean.2013.12.005
Carter, H. J. (1873). On the Hexactinellidae and Lithistidae generally, and particularly on the Aphrocallistidae, Aulodictyon, and Farreae, together with facts elicited from their deciduous structures and descriptions respectively of three new species. Ann. Magaz. Nat. Hist. 12, 349–373. doi: 10.1080/00222937308680782
Carvalho, F. C., Cárdenas, P., Ríos, P., Cristobo, J., Rapp, H. T., and Xavier, J. R. (2020). Rock sponges (lithistid demosponges) of the Northeast Atlantic seamounts, with the description of 10 new species. PeerJ 8:e8703. doi: 10.7717/peerj.8703
Carvalho, F. C., and Pisera, A. (2019). Revision of the genus Exsuperantia Özdikmen, 2009 (Tetractinellida: Phymaraphiniidae) with description of a new species from the Atlantic Ocean. Zootaxa 4613, 135–151. doi: 10.11646/zootaxa.4613.1.7
Carvalho, F. C., Pomponi, S. A., and Xavier, J. R. (2015). Lithistid sponges of the upper bathyal of Madeira, Selvagens and Canary Islands, with description of a new species of Isabella. J. Mar. Biol. Assoc. U.K. 95, 1287–1296. doi: 10.1017/s0025315414001179
Cathalot, C., Van Oevelen, D., Cox, T. J. S., Kutti, T., Lavaleye, M., Duineveld, G., et al. (2015). Cold-water coral reefs and adjacent sponge grounds: hotspots of benthic respiration and organic carbon cycling in the deep sea. Front. Mar. Sci. 2:37. doi: 10.3389/fmars.2015.00037
Darriba, D., Taboada, G. L., Doallo, R., and Posada, D. (2012). jModelTest 2: more models, new heuristics and parallel computing. Nat. Methods 9, 772–772. doi: 10.1038/nmeth.2109
Edgar, R. C. (2004). MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 32, 1792–1797. doi: 10.1093/nar/gkh340
Ekins, M., Erpenbeck, D., Wörheide, G., and Hooper, J. N. A. (2016). A new species of lithistid sponge hiding within the Isabella mirabilis species complex (Porifera: Demospongiae: Tetractinellida) from seamounts of the Norfolk Ridge. Zootaxa 4136, 433–460. doi: 10.11646/zootaxa.4136.3.2
Ellwood, M. J., Kelly, M., and Richer de Forges, B. (2007). Silica banding in the deep-sea lithistid sponge Corallistes undulatus: investigating the potential influence of diet and environment on growth. Limnol. Oceanogr. 52, 1865–1873. doi: 10.4319/lo.2007.52.5.1865
EU (2016). Regulation (EU) 2016/2336 of the European Parliament and of the council of 14 December 2016 Establishing Specific Conditions for Fishing for Deep-Sea Stocks in the North-East Atlantic and Provisions for Fishing in International Waters of the North-East Atlantic and Repealing Council Regulation (EC) No 2347/2002. Brussels: European Union.
Fallon, S. J., James, K., Norman, R., Kelly, M., and Ellwood, M. J. (2010). A simple radiocarbon dating method for determining the age and growth rate of deep-sea sponges. Nuclear Instrum. Methods Phys. Res. Sect. B 268, 1241–1243. doi: 10.1016/j.nimb.2009.10.143
FAO (2009). International Guidelines for the Management of Deep-sea Fisheries in the High Seas. Directives Internationales sur la Gestion de la pêche Profonde en haute mer. Directrices Internacionales para la Ordenación de las Pesquerías de Aguas Profundas en Alta Mar. 73 pp. Rome: Food and Agriculture Organization of the United Nations.
Folmer, O., Black, M., Hoeh, W., Lutz, R., and Vrijenhoek, R. (1994). DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 3, 294–299.
Freitas, R., Romeiras, M., Silva, L., Cordeiro, R., Madeira, P., González, J. A., et al. (2019). Restructuring of the ‘Macaronesia’ biogeographic unit: a marine multi-taxon approach. Sci. Rep. 9:15792. doi: 10.1038/s41598-019-51786-6
Gray, J. E. (1859). Description of MacAndrewia and Myliusia, two new forms of Sponges. Proc. Zool. Soc. Lond. 27, 437–440.
Hawkes, N., Korabik, M., Beazley, L., Rapp, H. T., Xavier, J. R., and Kenchington, E. (2019). Glass sponge grounds on the Scotian Shelf and their associated biodiversity. Mar. Ecol. Prog. Ser. 614, 91–109. doi: 10.3354/meps12903
Hestetun, J. T., Fourt, M., Vacelet, J., Boury-Esnault, N., and Rapp, H. T. (2015). Cladorhizidae (Porifera, Demospongiae, Poecilosclerida) of the deep Atlantic collected during Ifremer cruises, with a biogeographic overview of the Atlantic species. J. Mar. Biol. Assoc. U. K. 95, 1311–1342. doi: 10.1017/s0025315413001100
Hestetun, J. T., Tompkins-MacDonald, G., and Rapp, H. T. (2017). A review of carnivorous sponges (Porifera: Cladorhizidae) from the Boreal North Atlantic and Arctic. Zool. J. Linnean Soc. 181, 1–69. doi: 10.1093/zoolinnean/zlw022
Horton, T., Marsh, L., Bett, B. J., Gates, A. R., Jones, D. O. B., Benoist, N. M. A., et al. (2021). Recommendations for the standardisation of Open taxonomic nomenclature for image-based identifications. Front. Mar. Sci. 8:620702. doi: 10.3389/fmars.2021.620702
Howell, K. L., Piechaud, N., Downie, A. L., and Kenny, A. (2016). The distribution of deep-sea sponge aggregations in the North Atlantic and implications for their effective spatial management. Deep Sea Res. Part I Oceanogr. Res. Pap. 115, 309–320. doi: 10.1016/j.dsr.2016.07.005
ICES (2020). ICES/NAFO Joint Working Group on Deep-water Ecology (WGDEC). ICES Sc. Rep. 2:171. doi: 10.17895/ices.pub.6095
ICZN (1999). International Code of Zoological Nomenclature, 4th Edn, London: The International Trust for Zoological Nomenclature.
Johnson, J. Y. (1863). Description of a new siliceous sponge from the coast of Madeira. Proc. Zool. Soc. London, 257–259.
Kahn, A. S., Vehring, L. J., Brown, R. R., and Leys, S. P. (2016). Dynamic change, recruitment and resilience in reef-forming glass sponges. J. Mar. Biol. Assoc. U. K. 96, 429–436. doi: 10.1017/s0025315415000466
Kauffman, E. G., Herm, D., Johnson, C. C., Harries, P., and Höfling, R. (2000). The ecology of Cenomanian lithistid sponge frameworks, Regensburg area, Germany. Lethaia 33, 214–235. doi: 10.1080/00241160025100071
Kearse, M., Moir, R., Wilson, A., Stones-Havas, S., Cheung, M., Sturrock, S., et al. (2012). Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 28, 1647–1649. doi: 10.1093/bioinformatics/bts199
Kelly, M., Ellwood, M., Tubbs, L., and Buckeridge, J. (2007). “The lithistid Demospongiae in New Zealand waters: species composition and distribution,” in Porifera Research. Biodiversity, Innovation and Sustainability. Livros de Museu Nacional 28, eds M. R. Custódio, G. Lôbo-Hajdu, E. Hajdu, and G. Muricy (Rio de Janeiro: Routledge), 393–404.
Kirkpatrick, R. (1903). Descriptions of South African Sponges. Part II. Marine Investigations in South Africa 2, 171–180, pl. IV.
Knudby, A., Kenchington, E., and Murillo, F. J. (2013). Modeling the distribution of Geodia Sponges and Sponge grounds in the Northwest Atlantic. PLoS One 8:e82306. doi: 10.1371/journal.pone.0082306
Lê, H. L. V., Lecointre, G., and Perasso, R. (1993). A 28S rRNA-based phylogeny of the gnathostomes: first steps in the analysis of conflict and congruence with morphologically based cladograms. Mol. Phylogenet. Evol. 2, 31–51. doi: 10.1006/mpev.1993.1005
Lévi, C. (1960). Spongiaires des côtes occidentales africaines. Bulletin de l’Institut fran ais d’Afrique noire. A. Sciences naturelles 22, 743–769.
Lévi, C., and Lévi, P. (1983). Eponges Tétractinellides et Lithistides bathyales de Nouvelle-Calédonie. Bull. du Muséum national d’Histoire naturelle (4) 5, 101–168.
Lévi, C., and Lévi, P. (1988). Nouveaux Spongiaires Lithistides bathyaux à affinités crétacées de la Nouvelle-Calédonie. Bulletin du Muséum national d’Histoire naturelle 10, 241–263.
Lévi, C., and Lévi, P. (1989). “Spongiaires (MUSORSTOM 1 & 2),” in Résultats des Campagnes MUSORSTOM 4. Mémoires du Muséum national d’Histoire naturelle, Vol. 143, ed. J. Forest (Série A: Zoologie), 25–103.
Lévi, C., and Vacelet, J. (1958). Éponges récoltées dans l’Atlantique oriental par le ‘Président Théodore-Tissier’ (1955-1956). Revue Travaux l’Institut Pêches Marit. 22, 225–246.
Leys, S. P., and Lauzon, N. R. J. (1998). Hexactinellid sponge ecology: growth rates and seasonality in deep water sponges. J. Exper. Mar. Biol. Ecol. 230, 111–129. doi: 10.1016/s0022-0981(98)00088-4
Lima, M. J., Sala, I., and Caldeira, R. M. A. (2020). Physical connectivity between the NE Atlantic seamounts. Front. Mar. Sci. 7:238. doi: 10.3389/fmars.2020.00238
Maldonado, M., Aguilar, R., Blanco, J., García, S., Serrano, A., and Punzón, A. (2015). Aggregated clumps of lithistid sponges: a singular, reef-like bathyal habitat with relevant paleontological connections. PLoS One 10:e0125378. doi: 10.1371/journal.pone.0125378
Maldonado, M., López-Acosta, M., Sitjà, C., García-Puig, M., Galobart, C., Ercilla, G., et al. (2020). Sponge skeletons as an important sink of silicon in the global oceans. Nat. Geosci. 19, 815–822. doi: 10.1038/s41561-019-0430-7
Manconi, R., and Serusi, A. (2008). Rare sponges from marine caves: discovery of Neophrissospongia nana nov. sp. (Demospongiae, Corallistidae) from Sardinia with an annotated checklist of Mediterranean lithistids. ZooKeys 4, 71–87. doi: 10.3897/zookeys.4.39
Manconi, R., Serusi, A., and Pisera, A. (2006). A new Mediterranean ‘lithistid’ sponge, Aciculites mediterranea sp. nov. (Porifera: Demospongiae) from a dark marine cave in Sardinia. J. Mar. Biolog. Assoc. U.K. 86, 691–698. doi: 10.1017/S0025315406013580
Meyer, C. P., Geller, J. B., and Paulay, G. (2005). Fine scale endemism on coral reefs: archipelagic differentiation in turbinid gastropods. Evolution 59, 113–125. doi: 10.1554/04-194
Morato, T., Machete, M., Kitchingman, A., Tempera, F., Lai, S., Menezes, G., et al. (2008). Abundance and distribution of seamounts in the Azores. Mar. Ecol. Prog. Ser. 357, 17–21. doi: 10.3354/meps07268
Morrow, C., and Cárdenas, P. (2015). Proposal for a revised classification of the Demospongiae (Porifera). Front. Zool. 12:7. doi: 10.1186/s12983-015-0099-8
Murillo, F. J., Kenchington, E., Tompkins, G., Beazley, L., Baker, E., Knudby, A., et al. (2018). Sponge assemblages and predicted archetypes in the eastern Canadian Arctic. Mar. Ecol. Prog. Ser. 597, 115–135. doi: 10.3354/meps12589
Özdikmen, H. (2009). Substitute names for eight sponge genus group names (Porifera). Munis Entomology & Zoology 4, 212–218.
Palma, C., Lillebø, A. I., Borges, C., Souto, M., Pereira, E., Duarte, A. C., et al. (2012). Water column characterisation on the Azores platform and the sea mounts south of the archipelago. Mar. Pollut. Bull. 64, 1884–1894. doi: 10.1016/j.marpolbul.2012.06.015
Perán, A. D., Pham, C. K., Amorim, P., Cardigos, F., Tempera, F., and Morato, T. (2016). Seafloor characteristics in the Azores region (North Atlantic). Front. Mar. Sci. 3:204. doi: 10.3389/fmars.2016.00204
Pérez, T., Vacelet, J., Bitar, G., and Zibrowius, H. (2004). Two new lithistids (Porifera: Demospongiae) from a shallow eastern Mediterranean cave (Lebanon). J. Mar. Biolog. Assoc. U.K. 84, 15–24. doi: 10.1017/S0025315404008859h
Pham, C., Diogo, H., Menezes, G., Porteiro, F., Braga-Henriques, A., Vandeperre, F., et al. (2014). Deep-water longline fishing has reduced impact on vulnerable marine ecosystems. Sci. Rep. 4:4837. doi: 10.1038/srep04837
Pisera, A., and Gerovasileiou, V. (2021). Lithistid demosponges of deep-water origin in marine caves of the north-eastern Mediterranean Sea. Front. Mar. Sci. 8:630900. doi: 10.3389/fmars.2021.630900
Pisera, A., and Lévi, C. (2002). “’Lithistid’ Demospongiae,” in Systema Porifera. A Guide to the Classification of Sponges 1, eds J. N. A. Hooper and R. W. M. Van Soest (New York, NY: Kluwer Academic Plenum), 299–301. doi: 10.1007/978-1-4615-0747-5_33
Pisera, A., and Vacelet, J. (2011). Lithistid sponges from submarine caves in the Mediterranean: taxonomy and affinities. Scientia Marina 75, 17–40. doi: 10.3989/scsimar.2011.75n1017
Pomponi, S. A., Kelly, M., Reed, J. A., and Wright, E. A. (2001). Diversity and bathymetric distribution of lithistid sponges in the tropical western Atlantic region. Bull. Biol. Soc. Washington 10, 344–353.
Porteiro, F. M. (2009). The importance of Prince’s Albert I of monaco campaigns to the knowledge of the Azores Seas. Boletim Nuìcleo Cult. Horta 18, 189–219.
Puerta, P., Johnson, C., Carreiro-Silva, M., Henry, L.-A., Kenchington, E., Morato, T., et al. (2020). Influence of Water masses on the biodiversity and biogeography of deep-sea benthic ecosystems in the North Atlantic. Front. Mar. Sci. 7:239. doi: 10.3389/fmars.2020.00239
Ríos, P., Prado, E., Carvalho, F. C., Sánchez, F., Rodríguez-Basalo, A., Xavier, J. R., et al. (2020). Community composition and habitat characterization of a rock sponge aggregation (Porifera, Corallistidae) in the Cantabrian Sea. Front. Mar. Sci. 7:578. doi: 10.3389/fmars.2020.00578
Roberts, E. M., Bowers, D. G., Meyer, H. K., Samuelsen, A., Rapp, H. T., and Cárdenas, P. (2021). Water masses constrain the distribution of deep-sea sponges in the North Atlantic Ocean and Nordic Seas. Mar. Ecol. Prog. Ser. 659, 75–96. doi: 10.3354/meps13570
Ronquist, F., Teslenko, M., van der Mark, P., Ayres, D. L., Darling, A., Höhna, S., et al. (2012). MrBayes 3.2: efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 61, 539–542. doi: 10.1093/sysbio/sys029
Rooks, C., Fang, J. K.-H., Mørkved, P. T., Zhao, R., Rapp, H. T., Xavier, L., et al. (2020). Deep-sea sponge grounds as nutrient sinks: denitrification is common in boreo-Arctic sponges. Biogeosciences 17, 1231–1245. doi: 10.5194/bg-17-1231-2020
Sampaio, Í, Braga-Henriques, A., Pham, C., Ocaña, O., de Matos, V., Morato, T., et al. (2012). Cold-water corals landed by bottom longline fishery in the Azores. J. Mar. Biol. Assoc. U. K. 92, 1547–1555. doi: 10.1017/s0025315412000045
Sampaio, Í, Freiwald, A., Porteiro, F. M., Menezes, G., and Carreiro-Silva, M. (2019). Census of Octocorallia (Cnidaria: Anthozoa) of the Azores (NE Atlantic) with a nomenclature update. Zootaxa 4550, 451–498. doi: 10.11646/zootaxa.4550.4.1
Schlacher-Hoenlinger, M. A., Pisera, A., and Hooper, J. N. A. (2005). Deep-sea “lithistid” assemblages from the Norfolk Ridge (New Caledonia), with description of seven new species and a new genus (Porifera, Demospongiae). Zoosystema 27, 649–698.
Schmidt, O. (1870). Grundzüge einer Spongien-Fauna des Atlantischen Gebietes. Leipzig: Wilhelm Engelmann.
Schuster, A., Cárdenas, P., Pisera, A., Pomponi, S. A., Kelly, M., Wörheide, G., et al. (2018). Seven new deep-water Tetractinellida (Porifera: Demospongiae) from the Galápagos Islands - morphological descriptions and DNA barcodes. Zool. J. Linnean Soc. 184, 273–303. doi: 10.1093/zoolinnean/zlx110
Schuster, A., Erpenbeck, D., Pisera, A., Hooper, J. N. A., Bryce, M., Fromont, J., et al. (2015). Deceptive desmas: molecular phylogenetics suggests a new classification and uncovers convergent evolution of lithistid demosponges. PLoS One 10:e116038. doi: 10.1371/journal.pone.0116038
Schuster, A., Lopez, J. V., Becking, L. E., Kelly, M., Pomponi, S. A., Wörheide, G., et al. (2017). Evolution of group I introns in Porifera: new evidence for intron mobility and implications for DNA barcoding. BMC Evol. Biol. 17:82. doi: 10.1186/s12862-017-0928-9
Schuster, A., Pomponi, S., Pisera, A., Cárdernas, P., Kelly, M., Wörheide, G., et al. (2021). Systematics of ‘lithistid’ tetractinellid demosponges from the Tropical Western Atlantic - implications for phylodiversity and bathymetric distribution. PeerJ 9:e10775.
Sigovini, M., Keppel, E., and Tagliapietra, D. (2016). Open nomenclature in the biodiversity era. Methods Ecol. Evol. 7, 1217–1225.
Silva, M. A., Prieto, R., Cascão, I., Seabra, M. I., Machete, M., Baumgartner, M. F., et al. (2014). Spatial and temporal distribution of cetaceans in the mid-Atlantic waters around the Azores. Mar. Biol. Res. 10, 123–137. doi: 10.1080/17451000.2013.793814
Sitjà, C., Maldonado, M., Farias, C., and Rueda, J. L. (2019). Deep-water sponge fauna from the mud volcanoes of the Gulf of Cadiz (North Atlantic, Spain). J. Mar. Biol. Assoc. U. K. 99, 807–831. doi: 10.1017/S0025315418000589
Stamatakis, A. (2014). RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 30, 1312–1313. doi: 10.1093/bioinformatics/btu033
Topsent, E. (1889). Quelques spongiaires du Banc de Campêche et de la Pointe-à-Pître. Mémoires de la Société zoologique de France 2, 30–52.
Topsent, E. (1892). Contribution aì l’eìtude des Spongiaires de l’Atlantique Nord (Golfe de Gascogne, Terre-Neuve, Acçores). Reìsult. Campag. Sci. Accompl. Prince Albert I Monaco 2, 1–165.
Topsent, E. (1893). Nouvelle série de diagnoses d’éponges de Roscoff et de Banyuls. Archives de Zoologie expérimentale et générale. 3) 1 (Notes et Revue 10), xxxiii–xliii.
Topsent, E. (1904). Spongiaires des Acçores. Reìsult. Campag. Sci. Accompl. Prince Albert I Monaco 25, 1–280.
Topsent, E. (1928). Spongiaires de l’Atlantique et de la Meìditerraneìe provenant des croisieÌres du Prince Albert ler de Monaco. Reìsult. Campag. Sci. Accompl. Prince Albert I Monaco 74, 1–376.
Vacelet, J. (1969). Eponges de la Roche du Large et de l’étage bathyal de Méditerranée (Récoltes de la soucoupe plongeante Cousteau et dragages). Mémoires du Muséum national d’Histoire naturelle. A, Zoologie 59, 145–219.
Van Soest, R. W. M., Boury-Esnault, N., Hooper, J. N. A., Rützler, K., de Voogd, N. J., Alvarez, B., et al. (2021). World Porifera Database. Available online at: http://www.marinespecies.org/porifera (accessed January 03, 2021).
Van Soest, R. W. M., Boury-Esnault, N., Vacelet, J., Dohrmann, M., Erpenbeck, D., de Voogd, N. J., et al. (2012). Global diversity of Sponges (Porifera). PLoS One 7:e35105. doi: 10.1371/journal.pone.0035105
Wieczorek, J., Bloom, D., Guralnick, R., Blum, S., Döring, M., Giovanni, R., et al. (2012). Darwin Core: an evolving community-developed biodiversity data standard. PLoS One 7:e29715. doi: 10.1371/journal.pone.0029715
Keywords: bathyal, rock sponges, oceanic islands, seamounts, biodiversity, biogeography, vulnerable marine ecosystems, conservation
Citation: Xavier JR, Rees DJ, Pereira R, Colaço A, Pham CK and Carvalho FC (2021) Diversity, Distribution and Phylogenetic Relationships of Deep-Sea Lithistids (Porifera, Heteroscleromorpha) of the Azores Archipelago. Front. Mar. Sci. 8:600087. doi: 10.3389/fmars.2021.600087
Received: 28 August 2020; Accepted: 12 April 2021;
Published: 28 May 2021.
Edited by:
Jose Angel Alvarez Perez, Universidade do Vale do Itajaí, BrazilReviewed by:
Cristiana Castello-Branco, Smithsonian National Museum of Natural History (SI), United StatesCopyright © 2021 Xavier, Rees, Pereira, Colaço, Pham and Carvalho. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Joana R. Xavier, jxavier@ciimar.up.pt
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.