 Vanessa K. M. de Oliveira1*
Vanessa K. M. de Oliveira1* Drienne M. Faria1
Drienne M. Faria1 Haydée A. Cunha2Teresa E. C. dos Santos2Adriana C. Colosio3Lupércio A. Barbosa4Mylla Carla C. Freire1Ana Paula C. Farro1
Haydée A. Cunha2Teresa E. C. dos Santos2Adriana C. Colosio3Lupércio A. Barbosa4Mylla Carla C. Freire1Ana Paula C. Farro1- 1Laboratório de Genética e Conservação Animal, Departamento de Ciências Agrárias e Biológicas, Universidade Federal do Espírito Santo, São Mateus, Brazil
- 2Laboratório de Mamíferos Aquáticos e Bioindicadores, Faculdade de Oceanografia, Universidade do Estado do Rio de Janeiro, Rio de Janeiro, Brazil
- 3Instituto Baleia Jubarte (IBJ), Caravelas, Brazil
- 4Organização Consciência Ambiental (Instituto ORCA), Vila Velha, Brazil
The franciscana, Pontoporia blainvillei, is the most endangered small cetacean in the Southwestern Atlantic Ocean, occurring from Itaúnas, Espírito Santo, Brazil to Chubut province, Argentina. This area is divided into four Franciscana Management Areas (FMA). The northern portion of this species distribution is not continuous and a previous genetic study using mitochondrial DNA (mtDNA) separated it into FMAIa (Espírito Santo state) and FMAIb (North of Rio de Janeiro state). In order to increase the information about this population we expanded the sample number and evaluated mitochondrial and nuclear DNA diversity. Samples of 68 franciscanas found stranded on beaches from 2005 to 2020 were analyzed. Analyses included 350 bp of the mtDNA control region (D-loop) and 12 microsatellite loci. We identified three control region haplotypes in FMAIa, two of them not previously observed in this population, one being a new haplotype. Haplotype and nucleotide diversities were 0.0408 and 0.00012 respectively, the lowest reported for all FMAs analyzed until now. The Neutrality tests were not significant and Mismatch Distribution analysis did not reject the hypothesis of population expansion. One of the microsatellite loci was monomorphic, and for the other loci, two to nine alleles were identified, with expected heterozygosities ranging from 0.306 to 0.801. No substructure was revealed and effective population size (Ne) was estimated in 117.9 individuals. Even with an increased sample size, the high mitochondrial genetic homogeneity suggested for the population in a previous study was confirmed. Among six loci previously analyzed in other franciscana populations, five showed the lowest observed heterozygosities for the Espírito Santo population. The novel microsatellite data also showed low genetic diversity and could not reject the hypothesis of a single, panmitic population along the coast of Espírito Santo. This species has been intensively impacted in the last years by incidental capture during fishing activities and habitat degradation, caused by pollution, coastal development and environmental disasters in FMAIa. Considering that this population is small, isolated, and with low levels of genetic diversity, we reinforce the necessity of different conservation actions, focusing mainly on the reduction of bycatch of this species in the region.
Introduction
Pontoporia blainvillei (Gervais and D’orbigny, 1844), known as toninha or franciscana, is a small cetacean and the only member of the Pontoporiidae family. It is considered one of the smallest cetaceans and has as main characteristic a long beak and a bulky head (Crespo, 2018). This species inhabits estuarine regions and most records are in turbid waters up to 30 m deep (Di Beneditto and Ramos, 2001; ICMBIO, 2010).
Franciscanas are endemic to the southwestern coast of the Atlantic Ocean and occur from Itaúnas, Espírito Santo, Brazil, to Chubut province, in Argentina (Siciliano, 1994; Crespo et al., 1998; Bastida et al., 2007). The northern part of the species distribution, in Brazil, is not continuous. There are two gaps, one going from Santa Cruz, Espírito Santo to São Francisco de Itabapoana, Rio de Janeiro, and another from Armação de Búzios, Rio de Janeiro to Piraquara de Dentro, Rio de Janeiro. These gaps can occur due to depth, temperature, and water transparency (Siciliano et al., 2002; ICMBIO, 2010; Do Amaral et al., 2018).
Considering data about abundance, parasitology, growth, genetics, and morphology, four management areas, denominated “Franciscana Management Areas” (FMA) were proposed: FMAI, coast of Espírito Santo and Rio de Janeiro; FMAII, coast of São Paulo to Santa Catarina; FMAIII, coast of Rio Grande do Sul to Uruguay; and FMAIV, coast of Argentina (Secchi et al., 2003). The number and limits between FMAs have been refined using genetic data. Analyses of the mitochondrial marker D-loop, showed genetic structure between the two areas of occurrence within FMAI, and a new subdivision was proposed: FMAIa, Espírito Santo and FMAIb, north of Rio de Janeiro state (Cunha et al., 2014). These two FMA should be considered an Evolutionarily Significant Unit (ESUs). Therefore, the Espírito Santo state harbors the northernmost franciscana population, which is geographically isolated and genetically different from populations further southward, of which FMAIb is the closest both geographically and genetically (Cunha et al., 2014).
The franciscana is considered the most endangered small cetacean species in the western south Atlantic and its status of conservation is “Vulnerable” according with the International Union for the Conservation of Nature’s Red List of Threatened species (Zerbini et al., 2017). In Brazil, Pontoporia blainvillei is classified as “Critically Endangered,” according the Red Book of the Brazilian Fauna Threatened of Extinction and the List of Endangered Fauna and Flora list of Espírito Santo (ICMBIO, 2018; MCT, 2019).
The biggest risk factors for franciscanas are accidental catches in fishing (ICMBIO, 2010). In addition, franciscana populations are more vulnerable to environmental degradation processes since it is a costal species, with limited distribution and narrow preferential habitats. The FMAIa population shows an aggravating factor, by being an isolated population with probably a small number of individuals (ICMBIO, 2010).
A population that has been reduced in size for a long period of time or has suffered a bottleneck effect, tends to have low genetic diversity, decreasing its evolutionary potential and increasing the possibility of extinction in the face of environmental changes (Frankham et al., 2006). Therefore, the study of genetic diversity in a population like this is extremely important once the loss of diversity is directly related with the reduction of the evolutionary and reproductive potential (Frankham et al., 2006).
Molecular markers of the mitochondrial DNA (matrilinearly inherited) such as the control region (D-loop) and of the nuclear DNA (biparentaly inherited) such as microsatellite loci largely used in cetacean population genetics studies (Cunha et al., 2014; Gariboldi et al., 2015; Donato et al., 2019; Faria et al., 2020). These molecular markers can be used to access the genetic diversity of a population, to study the differences between the populations of a species and also to determine taxonomic relationships (Frankham et al., 2006). It is important to use mitochondrial and nuclear molecular markers for a better understanding of the populational genetics of a species.
D-loop is a region of mitochondrial DNA that shows some advantageous traits for these types of studies, such as being highly variable, having a small population size by being haploid, and lacking recombination (Frankham et al., 2006). Microsatellites or SSRs (Simple Sequence Repeats) have been extensile used for assessing nuclear DNA polymorphism for different species (Costa-Urrutia et al., 2012; Lima et al., 2017; Faria et al., 2020). They show codominance and high levels of polymorphism with many alleles per locus (Frankham et al., 2006). They have bi-parental transmission so they can be used to examine the flow of genes mediated by males and females (Oremus et al., 2007).
The present study aimed to evaluate the genetic diversity of the franciscana population from FMAIa, using sequences of the mitochondrial DNA (mtDNA) and twelve microsatellite loci, to increase the sample number and to test the occurrence of a single panmitic population in that area.
Materials and Methods
Sampling and DNA Extraction
Of a total of 68 samples, 50 were used in mitochondrial analyses and 43 in microsatellite loci analyses (Supplementary Table 1). Skin, liver or muscle samples were obtained from dead stranded franciscanas on the beaches of Espírito Santo coast (Figure 1 and Supplementary Table 1). The majority of samples were collected by the teams of Organização Consciência Ambiental (ORCA) and Instituto Baleia Jubarte (IBJ), responsible for assisting stranding of cetaceans and other marine mammals on the Espírito Santo coast. A few samples were collected by CTA – Serviços em Meio Ambiente, which performs the daily monitoring of Espírito Santo beaches, financed by PETROBRAS, a Brazilian company operating in the oil, natural gas and energy industry.
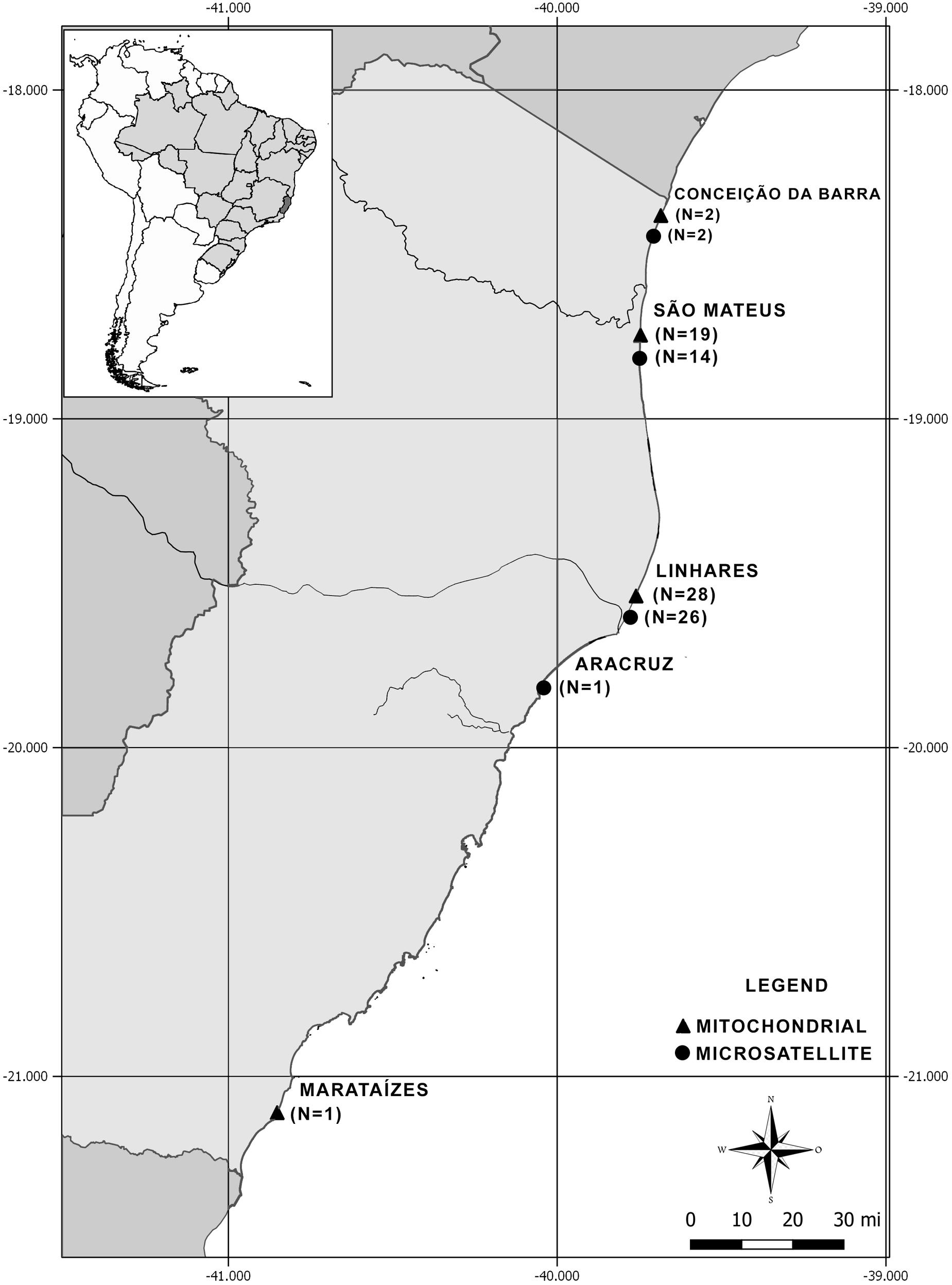
Figure 1. Sampling locations of stranded franciscanas (Pontoporia blainvillei) found dead on the beaches of Espírito Santo, Brazil, covering the area of Franciscana Management Area (FMAIa).
DNA extraction was performed using the protocols of phenol-chloroform (Sambrook et al., 1989) or saline extraction (Bruford et al., 1992). The mitochondrial DNA control region (D-loop) was amplified with the primers KRAdLp (Andrews et al., 2006) and DLp5 (Pichler et al., 2001).
mtDNA Amplification, Sequencing, and Data Analyses
Polymerase Chain Reactions were performed in 20 μL total volumes containing 20 ng DNA, Reaction Buffer 10X (Invitrogen), 200 μM of each dNTP, 2 mM of MgCl2, 0.5 units of Taq DNA polymerase (Invitrogen) and 0.2 μM of each primer. Cycle conditions were as follows: 95°C for 1 min, followed by 40 cycles of 94°C for 30 s, 54°C for 30 s and 72°C for 30 s, followed by a final extension of 72°C for 15 min.
Amplification products were sequenced in both directions (forward and reverse) by outsourced companies. The sequences were edited manually and aligned using the algorithm Muscle of MEGA 7.07 (Kumar et al., 2016). The generated sequences were compared with sequences deposited in GenBank1.
Arlequin v.3.11 (Excoffier and Lischer, 2010) was used to calculate haplotypic and nucleotide diversities, number of haplotypes, neutrality tests Fu’s Fs and Tajima’s D, and Mismatch distribution analysis. DnaSP v5 (Librado and Rozas, 2009) was used to identify polymorphic sites. A median joining haplotype network was built using Network (Bandelt et al., 1999).
For comparisons between the populations of Espírito Santo and Rio de Janeiro, sequences from north of Rio de Janeiro (N = 5) and south of Rio de Janeiro (N = 2) were added. Of these, four from north of Rio de Janeiro were generated in this study; one from north of Rio de Janeiro (Secchi et al., 1998) and two from south of Rio de Janeiro (Cunha et al., 2014) were downloaded from GenBank.
Population structuring among franciscanas from FMAIa was investigated using two putative populations: Central (N = 28, samples from Linhares) and North (N = 21, samples from Conceição da Barra and São Mateus). The only sample from south of Espírito Santo (Marataízes) was excluded from this analysis. AMOVA and pairwise FST were estimated with 10,000 random generations using Arlequin 3.5.2.2 (Excoffier and Lischer, 2010).
Microsatellite Amplification, Genotyping, and Analyses
Twelve microsatellite loci were amplified (Table 1). Polymerase Chain Reactions were performed in 10 μL total volumes containing around 30 ng DNA, Reaction Buffer 1X (Promega), 200 μM of each dNTP, 2.0–2.5 mM of MgCl2, 1U of Taq DNA polymerase (Promega) and 0.3 μM of each primer. Cycle conditions were as follows: 94°C for 4 min, followed by 30 cycles of 92°C for 45 s, T °C for 45 s and 72°C for 45 s, then 8 cycles of 92°C for 45 s, 53 °C for 45 s and 72°C for 45 s, followed by a final extension of 72°C for 30 min. Annealing temperatures for each of the primer pairs are listed in Table 1.
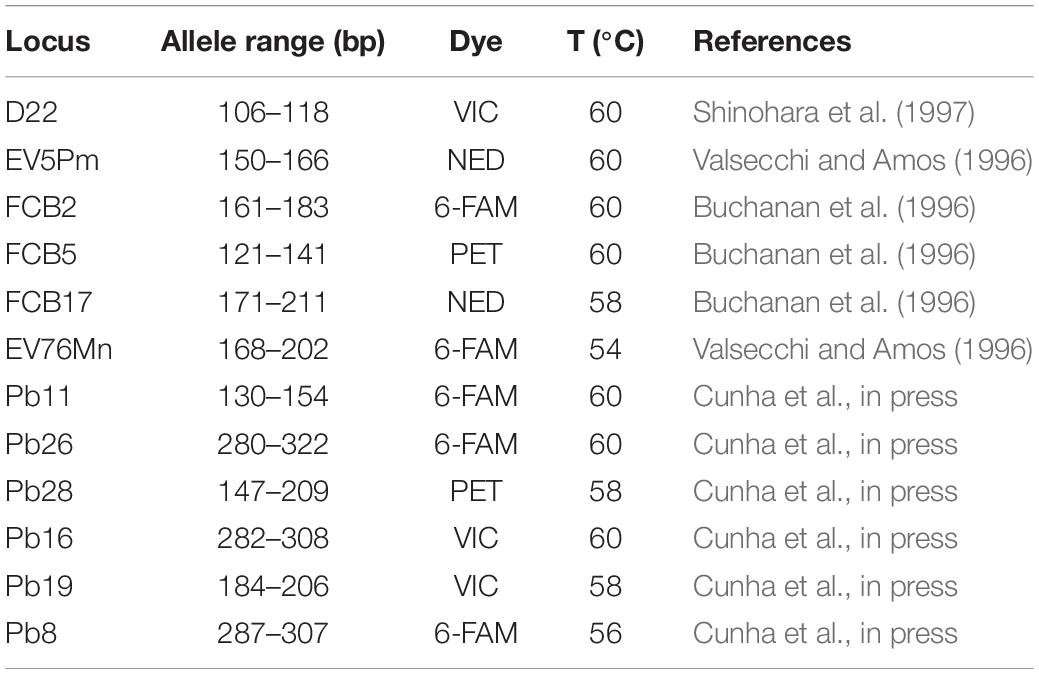
Table 1. Microsatellite loci used to access genetic diversity of franciscana (Pontoporia blainvillei). Locus name (Locus), allele range of amplified fragments size (base pairs, bp), fluorescent primer labeling (dye), annealing temperature (T), and original loci development (reference).
Capillary electrophoresis of PCR products was performed on ABI 3500 sequencer. Geneious version 2020.1.22 (Kearse et al., 2012) software was used to identify fragment sizes. Linkage disequilibrium was evaluated using GENEPOP on the Web (Raymond and Rousset, 1995) and testes of microsatellite loci for the presence of null alleles and/or genotyping errors were performed using the Microchecker 2.2.0.3 (Van Oosterhout et al., 2004).
Total number of alleles (Na), observed (Ho) and expected (He) heterozygosity, polymorphic information content (PIC), allelic richness (Ra) and inbreeding coefficient (FIS) were calculated using Arlequin 3.5.2.2 (Excoffier and Lischer, 2010) and Fstat v.2.9.3.2 (Goudet, 2001). Effective population size (Ne) was estimated using a method based on linkage disequilibrium (Waples, 1989; Waples and Do, 2010) as implemented in NeEstimator 2.0 (Do et al., 2014), we used a Pcrit value of 0.05 and 95% confidence intervals to reduce the potential bias for low frequency alleles (Do et al., 2014). A test for past events of population bottleneck based on allele frequencies using the software Bottleneck 1.2.02 was performed (Cornuet and Luikart, 1996); the one-tailed Wilcoxon sign-rank test for heterozygote excess was used under the stepwise mutational model (SMM) (Ohta and Kimura, 1973) and the two-phase model (TPM; 30 variations, mutational model 70 gradual, 10,000 interactions) (DiRenzo et al., 1994). A probability level of 0.05 was considered significant (Piry et al., 1999).
Tests for the existence of population structuring using microsatellite markers in franciscanas from FMAIa were also performed by using two putative populations: Central (N = 27), samples from Linhares e Aracruz, North (N = 16), samples from Conceição da Barra e São Mateus. The only sample from south of Espírito Santo (Marataízes) was excluded from this analysis. AMOVA and pairwise FST, based on number of different alleles, were estimated with 10,000 random generations using Arlequin 3.5.2.2 (Excoffier and Lischer, 2010).
A Bayesian clustering analysis was performed to estimate the most probable number of populations in Structure 2.3.2 (Pritchard et al., 2000), using an admixture model without prior population information and a correlated allele frequencies model. The burn-in length was set at 10,0000 steps, followed by 500,000 repetitions of Markov Chain Simulation and Monte Carlo (MCMC). Ten independent executions were performed for each value of K varying between one and ten to test the consistency of estimates of P (XK). The number of clusters that best fit in the data was assessed and visualized using Structure Harvester (Earl and VonHoldt, 2012) (available at3), a web-based program for collating results generated by the program Structure 2.3.2 (Earl and VonHoldt, 2012).
Analyses of genetic structuring were performed using three different combinations of loci: 10 loci (EV5Pm, FCB17, Pb11, Pb26, Pb16, Pb19, D22, FCB5, EV76Mn, and PB28); 8 loci (EV5Pm, FCB17, Pb11, Pb26, Pb16 e Pb19, D22, and Pb28); and 6 loci (EV5Pm, FCB17, Pb11, Pb26, Pb16, and Pb19).
Results
mtDNA
Sequences 350 bp-long were obtained for all 50 franciscanas and their identity (as Pontoporia blainvillei) were confirmed with 99–100% homology. A total of seven variable sites were identified (Table 2), representing three haplotypes. BLAST searches revealed that one of them was new MW248737. Haplotype H1 was the most frequent and was found in 48 of the 50 individuals. H2 was found in only one individual from Guriri, São Mateus. H3 was found in only one individual from Marataízes, an area within the species distribution gap. This latter haplotype is six mutational steps different from H2 and seven mutational steps different from H1.

Table 2. Variable sites of 350 bp of the mitochondrial DNA control region (D-loop) of franciscana (Pontoporia blainvillei) determining three haplotypes and their respective frequencies.
The Median-Joining network identified four haplotypes: H1, the most representative haplotype, shared by 48 individuals from Espírito Santo and one individual from north of Rio de Janeiro; H2, shared among four individuals from north of Rio de Janeiro and one individual from Espírito Santo; H3, an exclusive haplotype, represented by the individual from Marataízes and never reported for this species; H4, represented by two individuals from south of Rio de Janeiro. H3, found in Marataízes, is closer to the haplotypes of south of Rio de Janeiro than to the haplotypes of Espírito Santo. Given that, this individual that was found stranded in the beach of Marataízes (Espírito Santo) probably does not belong to Espírito Santo and was excluded from later population analyses in that region (Figure 2).
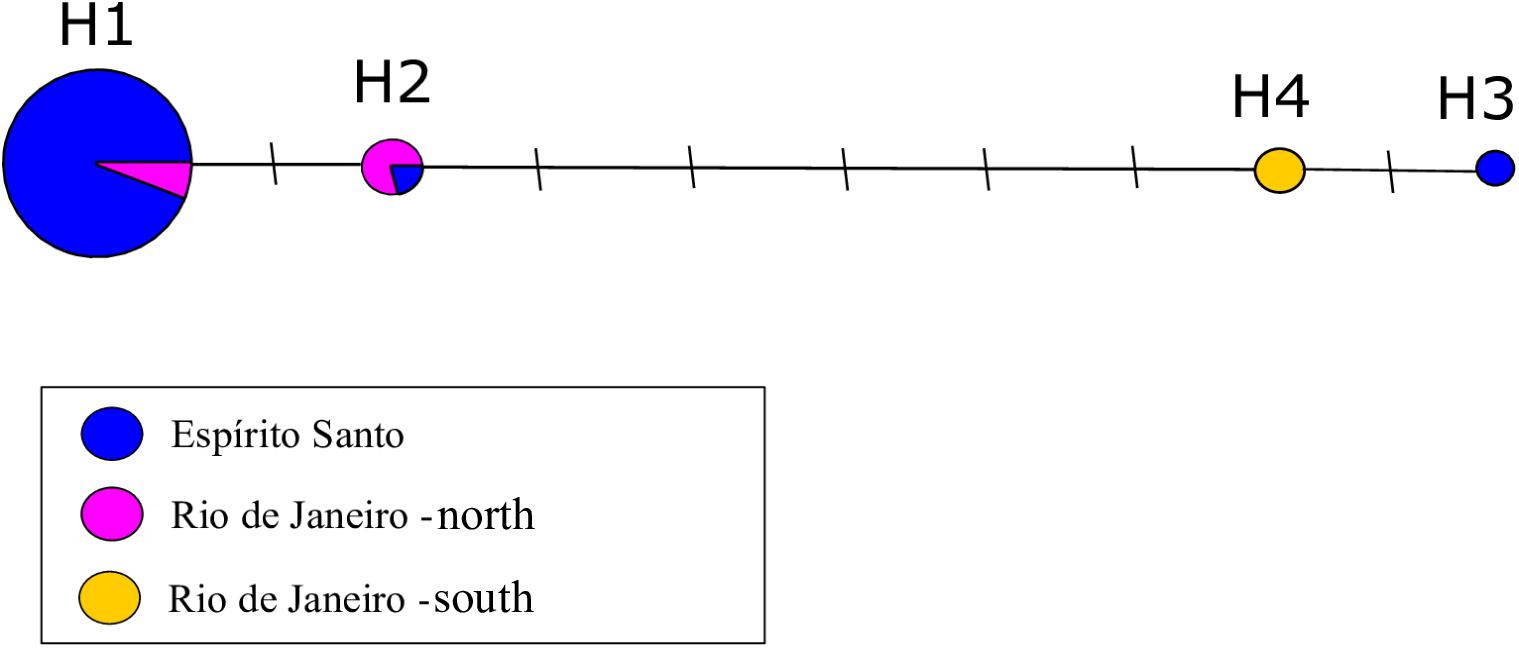
Figure 2. Haplotype network of the mitochondrial DNA control region (D-loop) for Pontoporia blainvillei (N = 57, 350 bp). Each circle corresponds to a haplotype and its size is proportional to its frequency in the population. Different colors represent the sampling locations. Each line between the haplotypes corresponds to the number of nucleotide differences between the haplotypes.
For the Espírito Santo population, the haplotype and nucleotide diversities were low, 0.0408 and 0.00012, respectively. The neutrality tests were not significant (Fu’s Fs = −1.619, P = 0.045; Tajima’s D = −1.104, P = 0.126). The Mismatch Distribution analysis showed that the distribution of the differences between the pairs of sequences is not statistically different from that expected in a population expansion scenario (PSsd = 0.100; PRaggedness = 0.920; Figure 3).
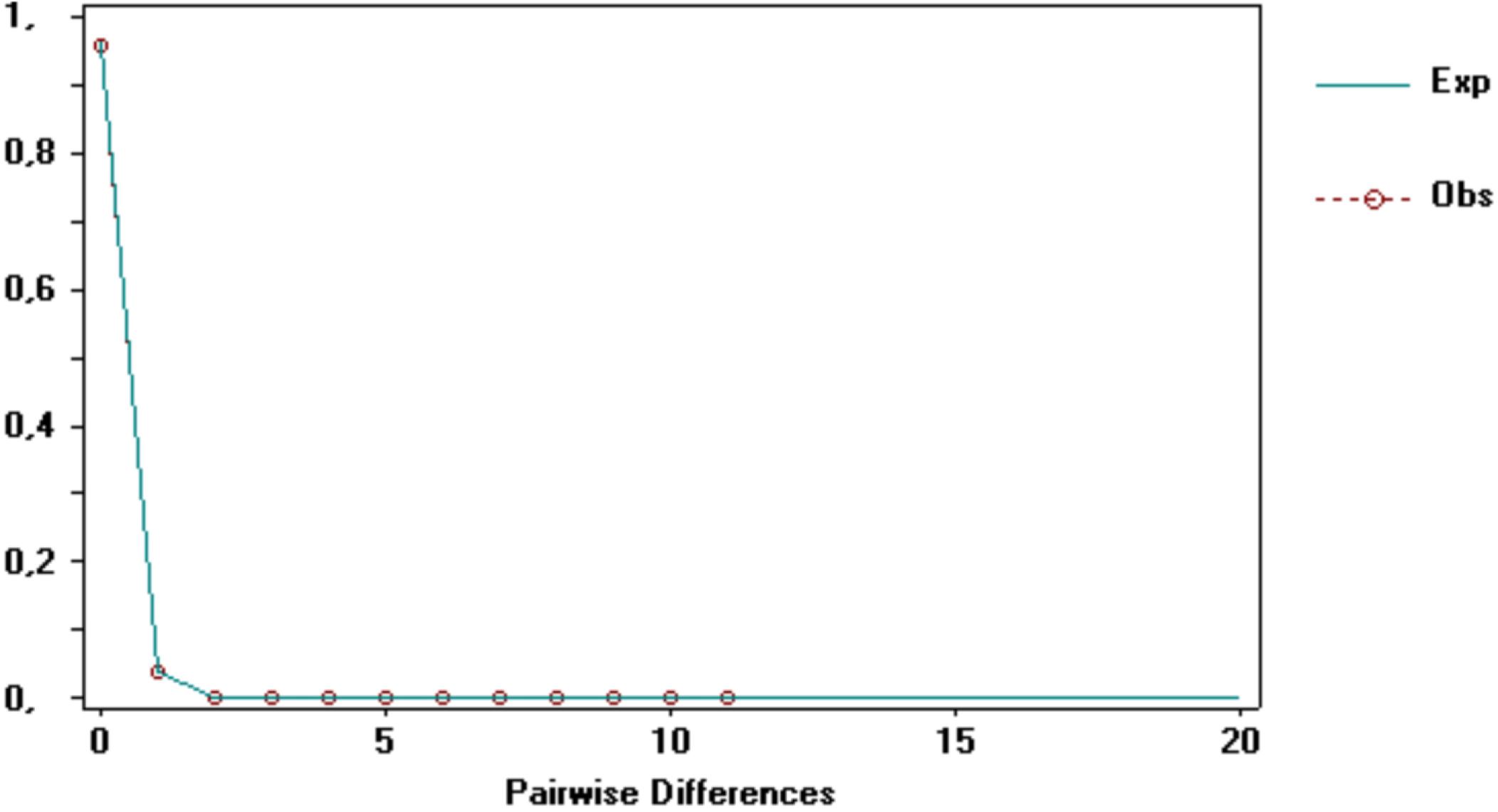
Figure 3. Mismatch Distribution for the mitochondrial DNA control region (D-loop) for Pontoporia blainvillei in the coast of Espírito Santo.
Genetic structuring tests using mtDNA showed no statistically significant FST values: FST = 0.01408, P = 0.435. AMOVA results showed that the highest levels of differentiation were within populations (locations) and no detectable structure was identified (Table 3).
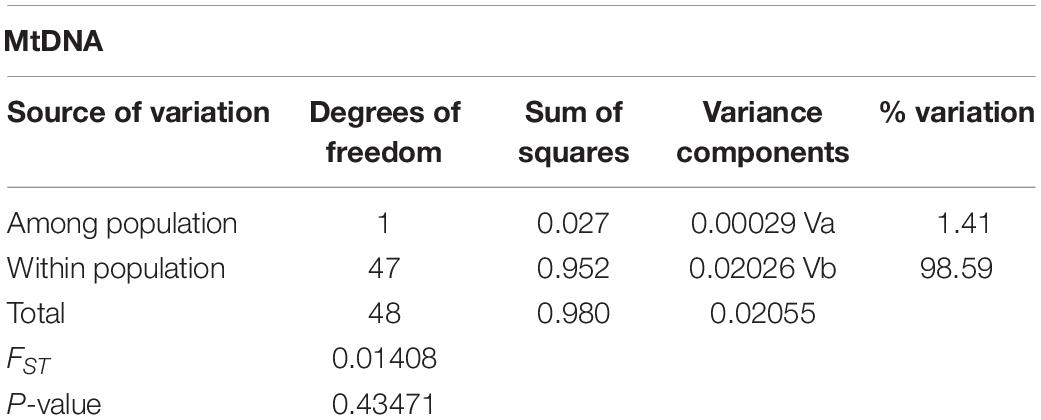
Table 3. AMOVA statistics for franciscana (Pontoporia blainvillei) of FMAIa for 350 bp of mtDNA control region (D-loop).
Microsatellite Loci
Forty-three franciscanas were genotyped. The diversity at 12 microsatellite loci was moderate, with from two to nine alleles, and expected heterozygosities ranging from 0.306 to 0.801 (Table 4). One locus was monomorphic (Pb8), four exhibited possible presence of null alleles with significant deviation from Hardy-Weinberg equilibrium (D22, FCB5, EV76Mn, and Pb28). None of the loci pairs was in linkage disequilibrium (Table 4).
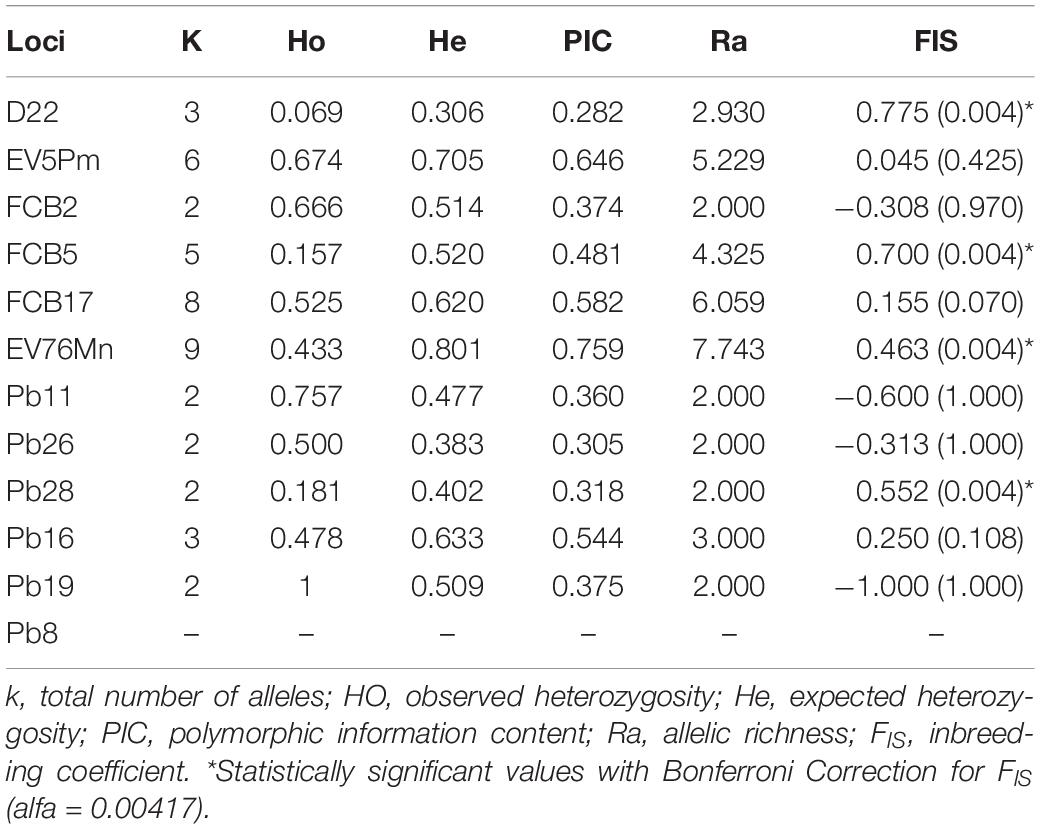
Table 4. Genetic diversity of franscicana for twelve microsatellite loci.
Effective population size (Ne) for franciscanas of FMAIa was estimated to be 117.9 (95% CI: 27.8–∞; harmonic mean sample size: 25.6). The Wilcoxon one-tailed test revealed that the franciscanas did not go through a bottleneck event based on the TPM mutational model (P = 0.12012), and SMM model (P = 0.51709).
The genetic structuring into two putative populations (Central and North) tested using three different combinations of loci showed no statistically significant FST values: 10 loci, pairwise FST = 0.006 (P = 0.415); 8 loci, pairwise FST = 0.001 (P = 0.411); 6 loci, pairwise FST = 0.001 (P = 0.406). AMOVA results showed that the highest levels of differentiation were within putative populations and no detectable structure was identified (Table 5).
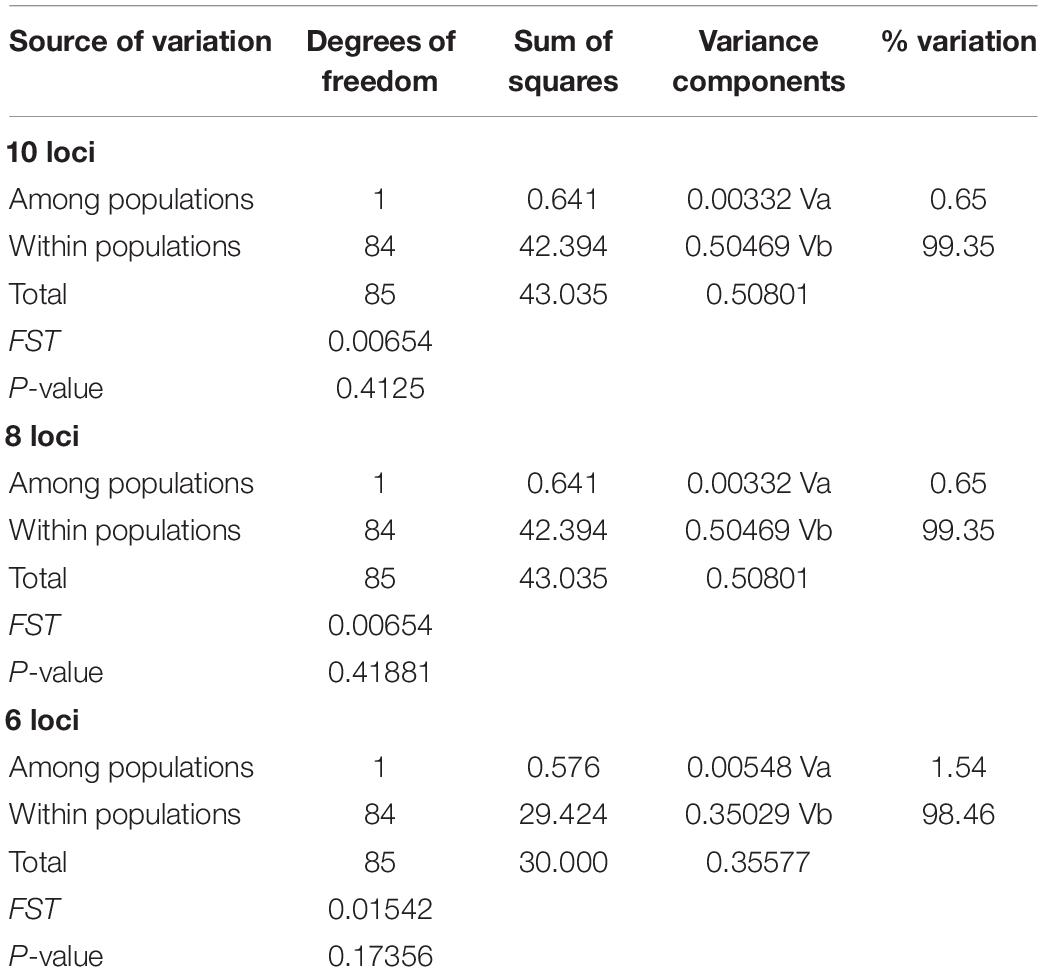
Table 5. AMOVA statistics of franciscana for three different combination of microsatellite loci.
The Bayesian clustering analysis did not detect structuring using the same combination of loci. The combination of 10 loci showed the clearest results. Delta K indicated K = 2 as the most likely scenario, but the greatest posterior probability was for K = 1. The analysis of Q for K = 2 showed all individuals to have similarly mixed ancestries (Figure 4 and Supplementary Figures 1, 2). These results thus reinforce the presence of just one population of franciscanas throughout FMAIa.
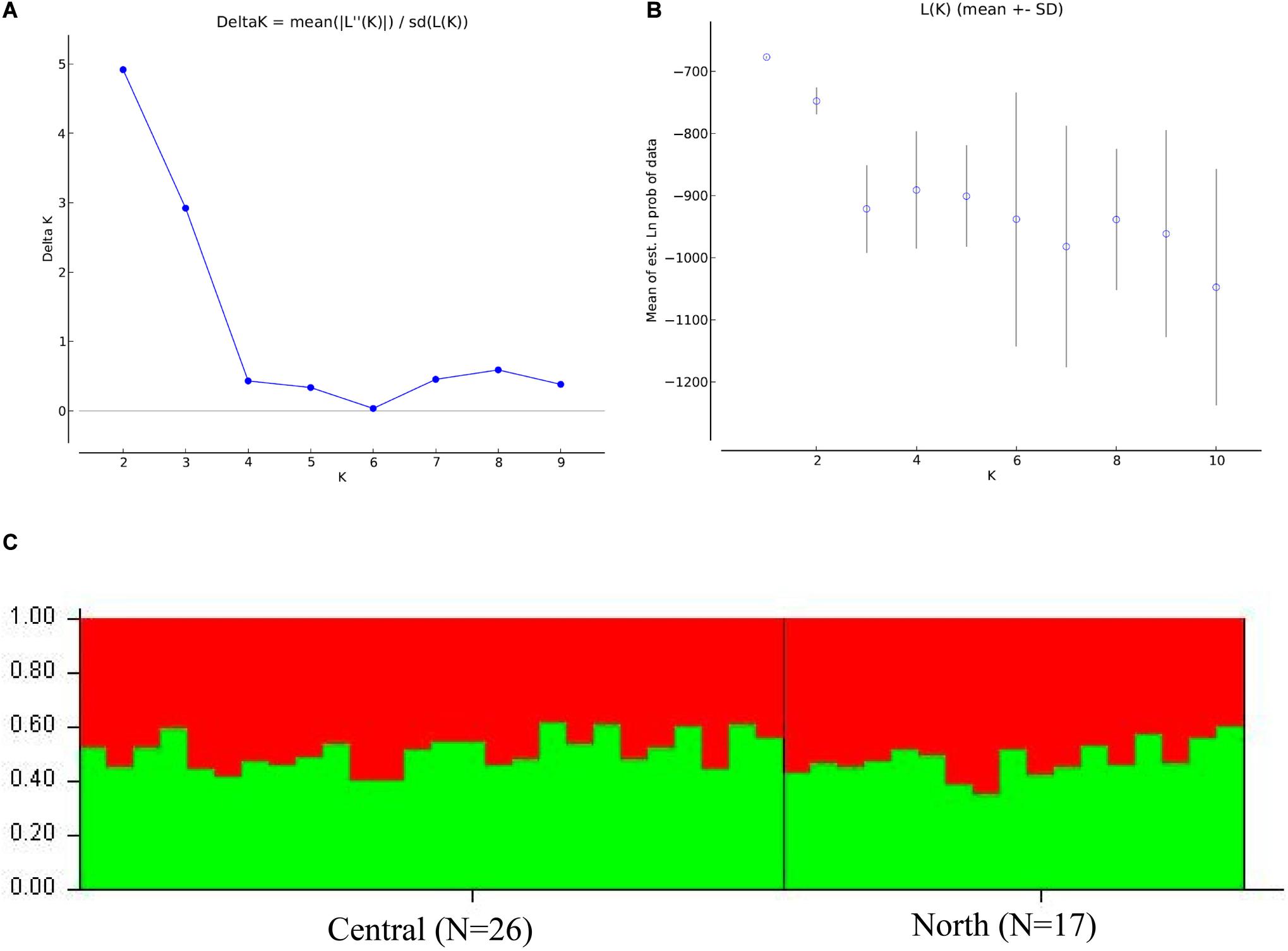
Figure 4. Bayesian clustering analysis inferred with the program Structure 2.3.2 for ten microsatellite loci of franciscana (Pontoporia blainvillei) of FMAIa: (A) ΔK from Evanno’s method is shown between successive K values, (B) mean log probability (Lk) is given for each K tested, and the (C) assignment probabilities of individuals to putative population clusters at K = 2 according to sampling putative populations.
Discussion
This is the first microscale study about Pontoporia blainvillei from northern Espírito Santo (FMAIa). In this study, we increased the number of samples evaluated using the mitochondrial marker (N = 50) in relation to the macroscale study developed by Cunha et al. (2014), and presented novel nuclear DNA data, providing important information that can assist in conservation strategies for this isolated population.
With increased sampling it was possible to detect a new haplotype for the population of Espírito Santo (FMAIa), not described by Cunha et al. (2014). Three haplotypes were found, including the new one, present in an individual who was found stranded in Marataízes, ES, an area known to be a gap in the species distribution. This haplotype showed many mutational steps from the most frequent haplotype in FMAIa, so this animal may have been caught by a fishing boat elsewhere and discarded in Espírito Santo. This would be unsurprising since incidental catch is one of the main threat factors for the species throughout its distribution.
The genetic diversity both at mitochondrial DNA and microsatellite loci was lower than already reported for P. blainvillei in other FMAs. Studies carried out with franciscanas from other FMAs found values ranging from 0.476 to 1.00 for haplotypic diversity (Table 6) and 0.0010 to 0.014 for nucleotide diversity. For microsatellite analyses, five out of six loci showed lower diversity in FMAIa than in previous studies (Mendez et al., 2007, 2010; Costa-Urrutia et al., 2012; Cunha et al., 2014; Gariboldi et al., 2015).
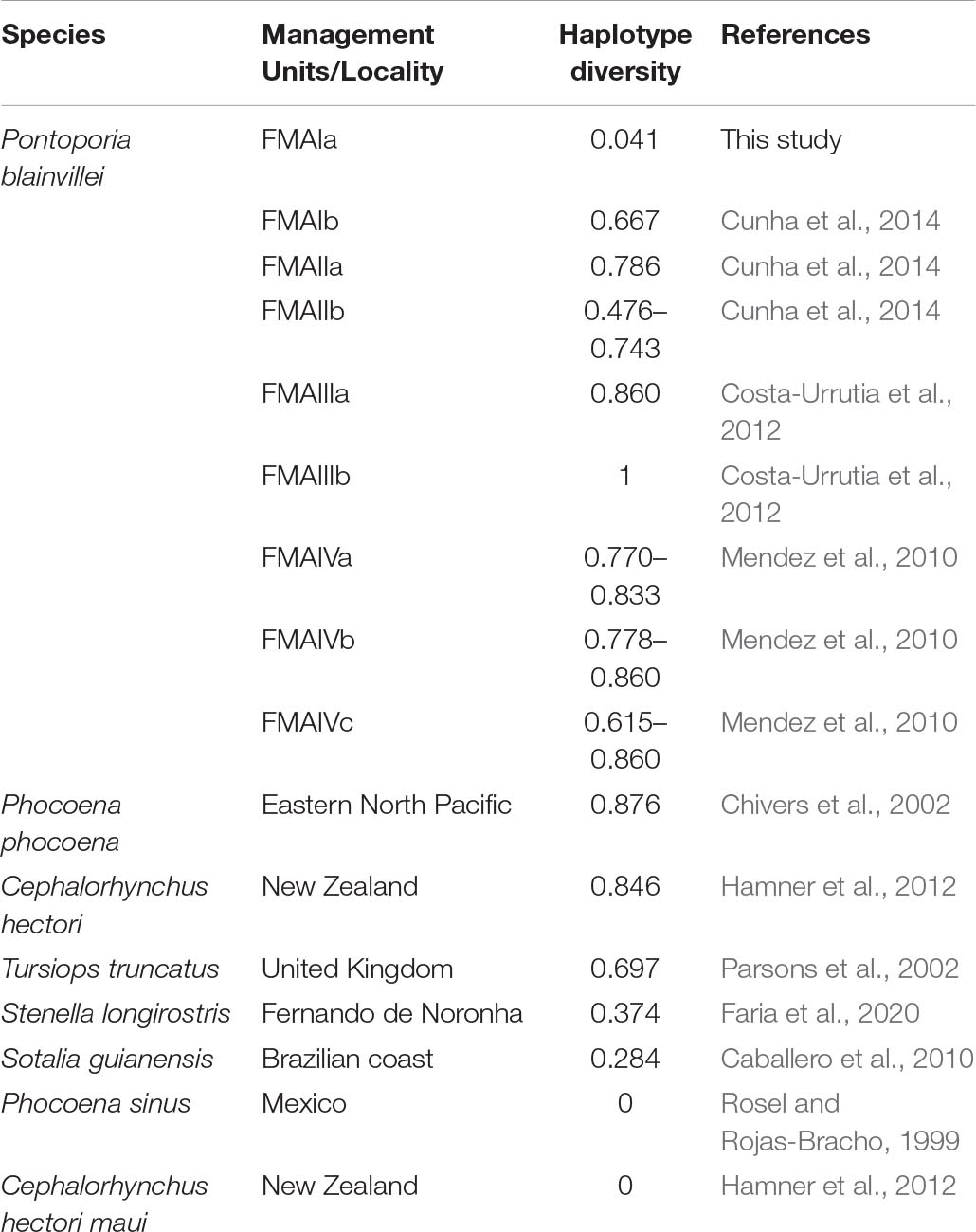
Table 6. Comparison of Pontoporia blainvillei haplotypic diversity between management units along species distribution and comparison with other small cetacean species.
The results presented here follow the same pattern reported in a macroscale study, which reported an increasing gradient of haplotypic diversity from the south to the north of the species distribution (Cunha et al., 2014).
When compared with other species of dolphins, threatened or not, franciscanas from FMAIa still have one of the lowest haplotype genetic diversity found (Rosel and Rojas-Bracho, 1999; Chivers et al., 2002; Parsons et al., 2002; Caballero et al., 2010; Hamner et al., 2012; Faria et al., 2020; Table 6).
The paucity of genetic variation may suggest that the situation of this endangered species in Espírito Santo is worrying. According to Frankham et al. (2006) species with small populations or that have suffered bottleneck events have low genetic diversity and suffer from accelerated endogamy, which leads to less reproductive and evolutionary potential. However, despite the low genetic variability seem in franciscanas from FMAIa, Fu (Fs), and Tajima (D) analyses were not significant, and mismatch distribution analysis did not discard the hypothesis of population expansion, while specific analysis with microsatellite data also did not detect signs of bottleneck events.
Implications for Management and Conservation
The conservation status and viability of the franciscana population in Espírito Santo is a known concern (Danilewicz et al., 2012). Franciscans were not seen more than 13 km from the coast (19 m depth), which suggests that in FMAIa they have more coastal habits than in other FMAs, and the size of this population is believed to be small (Danilewicz et al., 2012). In this study effective population size was estimated to be 117.9 for the franciscanas in FMAIa. Reduction in population size directly results in the loss of genetic diversity, which is already observed for this population. The loss of genetic diversity and the reduction of population size are factors considered to be vortices of extinction (Frankham et al., 2006).
Habitat degradation has been considered a critical factor for the conservation of coastal cetaceans. According to Reeves et al. (2003), coastal cetaceans are much more threatened than ocean species due to the intense use of these environments. Anthropic actions are the main threats for these species (ICMBIO, 2010). In the case of Pontoporia blainvillei, as with other coastal small cetaceans, interaction with fisheries plays a major role (Crespo et al., 1994; Capozzo et al., 2007; Negri et al., 2012).
Besides the mortality caused by accidental captures, franciscanas from Espírito Santo also suffered with extreme habitat degradation caused by the Fundão mining dam rupture, which occurred in November 2015 in the city of Mariana, MG, Brazil, was considered the biggest environmental disaster in South America (Marta-Almeida et al., 2016). After 14 months, 74 million tons of mining sediment were launched into the Doce River until reaching the Atlantic Ocean (Magris et al., 2019). The coastal area that immediately received the largest amount of trace-elements after the dam burst was the south side of the Doce river mouth (Magris et al., 2019), a region in FMAIa known to be important for franciscanas.
The presence of ore tailings in the franciscana habitat can affect their survival over time in several ways. High levels of trace-element accumulation in plankton, marine shrimp, and fishes were found after the disaster (RRDM, 2019). Cetaceans are top-of-the-chain animals, being highly affected by the bioaccumulation of chemicals in the food chain (Torre et al., 2012). A study comparing the concentration of trace-elements in the organs of franciscanas from the Espírito Santo coast before and after the disaster are being concluded (Manhães et al., in prep).
The franciscana population in Espírito Santo (FMAIa) is the smallest. It is geographically and genetically isolated and shows low genetic diversity. All of these characteristics enhance the vulnerability of this population, making even more important and urgent the development of conservation strategies in FMAIa, such as the creation of MPA (Marine Protected Areas) or maintenance/reinforcement of a fisheries exclusion zone in the impacted area. To avoid human intoxication, fishing has been prohibited in the Rio Doce mouth since the disaster. That measure could impact franciscanas positively, but it is uncertain to what extent this prohibition has been strictly followed. Thus, it is also worth mentioning the importance of monitoring accidental captures in FMAIa in the short, medium and long term.
Data Availability Statement
The new sequence found in this study, has already been deposited in a public database (https://www.ncbi.nlm.nih.gov/genbank/). The sequence accession number is MW248737. The sequence under that access number will only be available after the article is published with that number.
Ethics Statement
Ethical review and approval was not required for the animal study because the animals come from carcasses found stranded on beaches.
Author Contributions
VO: formal analysis of mtDNA, writing-original draft, writing-review and editing, conceptualization, discussion, and investigation and validation. DF: formal analysis of microsatellites, writing the microsatellites part, review and editing, conceptualization, and discussion. HC: writing-review and editing, conceptualization, discussion, and funding acquisition. TS: microsatellite laboratory part and writing-review. AC and LB: resources and writing-review and editing. MF: extraction of some samples and writing-review and editing. AF: conceptualization, discussion and investigation, funding acquisition, project administration, resources, supervision, and writing-review and editing. All authors contributed to the article and approved the submitted version.
Funding
This study was part of Projeto Conservação da Toninha na Área de Manejo I (Franciscana Management Area I – FMA I) and was funded with resources from a Term of Adjustment of Conduct signed by Chevron Brasil and the Ministério Público Federal, and managed by Fundo Brasileiro para a Biodiversidade – Funbio (Contract 81/2016).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank CTA – Serviços em Meio Ambiente for the samples provided. We thank Fábio Ribeiro Mendes for drawing the map. This study was part of Projeto Conservação da Toninha na Área de Manejo I (Franciscana Management Area I – FMA I) and was funded with resources from a Term of Adjustment of Conduct signed by Chevron Brasil and the Ministério Público Federal, and managed by Fundo Brasileiro para a Biodiversidade – Funbio (Contract 81/2016).
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2020.608276/full#supplementary-material
Footnotes
- ^ www.ncbi.nlm.nhi.gov/Genbank
- ^ http://www.geneious.com
- ^ http://taylor0.biology.ucla.edu/structureHarvester/
References
Andrews, K. R., Karczmarski, L., Au, W. W. L., Rickards, S. H., Vanderlip, C. A., and Robert, J. T. (2006). Patterns of genetic diversity of the Hawaiian spinner dolphin (Stenella longirostris). Atoll Res. Bull. 543, 65–73.
Bandelt, H. J., Forster, P., and Röhl, A. (1999). Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 16, 37–48. doi: 10.1093/oxfordjournals.molbev.a026036
Bastida, R., Rodríguez, D., Secchi, E., and Da Silva, V. (2007). Mamíferos Acuáticos de Sudamérica y Antártida. Buenos Aires:).
Bruford, M. W., Hanotte, O., Broohfield, J. F. Y., and Burke, T. (1992). “Single-locus and multilocus DNA fingerprinting,” in Molecular genetic analyses of populations: A Pratical Approach, ed. A. R. Hoelzel (Oxford: Oxford University Press), 225–269.
Buchanan, F. C., Friesen, M. K., Littlejohn, R. P., and Clayton, J. W. (1996). Microsatellites from the beluga whale Delphinapterus leucas. Mol. Ecol. 5, 571–575. doi: 10.1046/j.1365-294x.1996.00109.x
Caballero, S., Trujillo, F., Vianna, J. A., Barrios-Garrido, H., Montiel, M. G., Beltrán-Pedreros, S., et al. (2010). Mitochondrial DNA diversity, differentiation and phylogeography of the south american riverine and coastal dolphins Sotalia fluviatilis and Sotalia guianensis. Lajam 8, 69–79. doi: 10.5597/lajam00155
Capozzo, H. L., Negri, M. F., Pérez, F. H., Albareda, D., Monzón, F., and Corcuera, J. F. (2007). Incidental mortality of franciscana dolphin (pontoporia blainvillei) in argentina. Lajam 6, 127–137. doi: 10.5597/lajam00118
Chivers, S. J., Dizon, A. E., Gearin, P. J., and Robertson, J. M. (2002). Small-scale population structure of eastern North Pacific harbour porpoises (Phocoena phocoena) indicated by molecular genetic analyses. J. Cetacean Res. Manag. 4, 111–122.
Cornuet, J. M., and Luikart, G. (1996). Description and power analysis of two tests for detecting recent population bottlenecks from allele frequency data. Genetics 144, 01–14.
Costa-Urrutia, P., Abud, C., Secchi, E. R., and Lessa, E. P. (2012). Population Genetic Structure and Social Kin Associations of Franciscana Dolphin, Pontoporia blainvillei. J. Heredity 103, 92–102. doi: 10.1093/jhered/esr103
Crespo, E. A. (2018). “Franciscana Dolphin,” in Encyclopedia of Marine Mammals, eds B. Würsig, J. G. M. Thewissen, and K. M. Kovacs (Amsterdam: Elsevier), 388–392. doi: 10.1016/b978-0-12-804327-1.00133-3
Crespo, E. A., Corcuera, J., and Cazorla, A. L. (1994). Interactions between marine mammals and fisheries in some fishing areas of the coast of Argentina. Rep. Int. Whal. Comm. Special Iss. 15, 269–282.
Crespo, E. A., Harris, G., and González, R. (1998). Group size and distributional range of the franciscana, Pontoporia blainvillei. Mar. Mamm. Sci. 14, 845–849. doi: 10.1111/j.1748-7692.1998.tb00768.x
Cunha, H. A., Medeiros, B. V., Barbosa, L. A., Cremer, M. J., Marigo, J., Brito, J. L., et al. (2014). Population structure of the endangered franciscana dolphin (Pontoporia blainvillei): reassessing management units. PLoS One 9:e85633. doi: 10.1371/journal.pone.0085633
Cunha, H. A., dos Santos, T. E. C., Alvarenga, L. C., Cremer, M. J., Cavaleiro, N. P., Colosio, A., et al. (in press). Microsatellite markers for the endangered franciscana dolphin (Pontoporia blainvillei). Mol. Biol. Rep.
Danilewicz, D., Zerbini, A. N., Andriolo, A., Secchi, E. R., Sucunza, F., Ferreira, E., et al. (2012). “Abundance and distribution of an isolated population of franciscana dolphins (Pontoporia blainvillei) in southeastern Brazil: red alert for FMA I,” in International Whaling Commission, Scientific Committee Paper SC/64/SM17, Panama (Cambridge: International Whaling Commission).
Di Beneditto, A. P. M., and Ramos, R. M. A. (2001). Biology and conservation of the franciscana (Pontoporia blainvillei) in the north of Rio de Janeiro State. Brazil. J. Cetacean Res. Manag. 3, 185–192.
DiRenzo, A., Peterson, A. C., Garza, J. C., Valdes, A. M., and Slatkin, M. (1994). Mutational processes of simple-sequence repeat loci in human populations. Proc. Natl. Acad. Sci. 91, 3166–3170. doi: 10.1073/pnas.91.8.3166
Do Amaral, K. B., Danilewicz, D., Zerbini, A., Di Beneditto, A. P., Andriolo, A., Alvares, D. J., et al. (2018). Reassessment of the franciscana Pontoporia blainvillei (Gervais &d’Orbigny, 1844) distribution and niche characteristics in Brazil. J. Exp. Mar. Biol. Ecol. 508, 1–12. doi: 10.1016/j.jembe.2018.07.010
Do, C., Waples, R. S., Peel, D., Macbeth, G. M., Tillett, B. J., and Ovenden, J. R. (2014). NeEstimator v2: re-implementation of software for the estimation of contemporary effective population size (Ne) from genetic data. Mol. Ecol. Res. 14, 209–214. doi: 10.1111/1755-0998.12157
Donato, A., Siciliano, S., Weksler, M., Silva, D. A., Carvalho, E. F., Loiola, S., et al. (2019). Population structure and the conservation status of the rough-toothed dolphins based on the analysis of the mitochondrial control region. Forensic Sci. Int. Gen. Suppl. Ser. 7, 294–295. doi: 10.1016/j.fsigss.2019.09.103
Earl, D. A., and VonHoldt, B. M. (2012). Structure Harvester: a website and program for visualizing STRUCTURE output and implementing the Evanno method. Conservat. Gen. Res. 4, 359–361. doi: 10.1007/s12686-011-9548-7
Excoffier, L., and Lischer, H. E. L. (2010). Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Res. 10, 564–567. doi: 10.1111/j.1755-0998.2010.02847.x
Faria, D. M., da Silva, J. M. Jr., Pires Costa, L., Rezende Paiva, S., Marino, C. L., Rollo, M. M. Jr., et al. (2020). Low mtDNA diversity in a highly differentiated population of spinner dolphins (Stenella longirostris) from the Fernando de Noronha Archipelago, Brazil. PLoS One 15:e0230660. doi: 10.1371/journal.pone.0230660
Frankham, R., Ballou, J. D., and Briscoe, D. A. (2006). Fundamentos de Genética da Conservação. Ribeirão Preto: SBG, 262.
Gariboldi, M. C., Túnez, J. I., Dejean, C. B., Failla, M., Vitullo, A. D., Negri, M. F., et al. (2015). Population Genetics of Franciscana Dolphins (Pontoporia blainvillei): Introducing a New Population from the Southern Edge of Their Distribution. PLoS One 10:e0132854. doi: 10.1371/journal.pone.0132854
Goudet, J. (2001). FSTAT, a program to estimate and test gene diversities and fixation indices (version 2.9.3). Available online at http://www.unil.ch/izea/softwares/fstat.html
Hamner, E. M., Pichler, F. B., Heimeier, D., Constantine, R., and Baker, C. S. (2012). Genetic Differentiation and limited gene flow among fragmented populations of New Zealand endemic Hector’s and Maui’s dolphins. Conserv. Gen. 13, 987–1002. doi: 10.1007/s10592-012-0347-9
ICMBIO (2010). Plano de ação nacional para a conservação do pequeno cetáceo Toninha: Pontoporia blainvillei. Brasília: Instituto Chico Mendes de Conservação da Biodiversidade.
ICMBIO (2018). “Livro Vermelho da Fauna Brasileira Ameaçada de Extinção: Volume II - Mamíferos,” in Livro Vermelho da Fauna Brasileira Ameaçada de Extinção (Brasília: ICMBio), 622.
Kearse, M., Moir, R., Wilson, A., Stones-Havas, S., Cheung, M., Sturrock, S., et al. (2012). Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics. 28, 1647–1649. doi: 10.1093/bioinformatics/bts199
Kumar, S., Stecher, G., and Tamura, K. (2016). MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 35, 1547–1549. doi: 10.1093/molbev/msw054
Librado, P., and Rozas, J. (2009). DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 25, 1451–1452. doi: 10.1093/bioinformatics/btp187
Lima, J. Y., Machado, F. B., Farro, A. P. C., Barbosa, L. dA, da Silveira, L. S., and Medina-Acosta, E. (2017). Population genetic structure of Guiana dolphin (Sotalia guianensis) from the southwestern Atlantic coast of Brazil. PLoS One 12:e0183645. doi: 10.1371/journal.pone.0183645
Magris, R. A., Marta-Almeida, M., Monteiro, J. A. F., and Ban, N. C. (2019). A modelling approach to assess the impact of land mining on marine biodiversity: Assessment in coastal catchments experiencing catastrophic events (SW Brazil). Sci. Total Environ. 659, 828–840. doi: 10.1016/j.scitotenv.2018.12.238
Marta-Almeida, M., Mendes, R., Amorim, F. N., Cirano, M., and Dias, J. M. (2016). Fundão Dam collapse: Oceanic dispersion of River Doce after the greatest Brazilian environmental accident. Mar. Pollut. Bull. 112, 359–364. doi: 10.1016/j.marpolbul.2016.07.039
MCT. (2019). Fauna e flora ameaçadas de extinção no estado do Espírito Santo. Brazil: Ministério de Ciência e Tecnologia.
Mendez, M., Rosenbaum, H. C., and Bordino, P. (2007). Conservation genetics of the franciscana dolphin in Northern Argentina: population structure, by-catch impacts, and management implications. Conservat. Gen. 9, 419–435. doi: 10.1007/s10592-007-9354-7
Mendez, M., Rosenbaum, H. C., Subramaniam, A., Yackulic, C., and Bordino, P. (2010). Isolation by environmental distance in mobile marine species: molecular ecology of franciscana dolphins at their southern range. Mol. Ecol. 19, 2212–2228. doi: 10.1111/j.1365-294x.2010.04647.x
Negri, M. F., Denuncio, P., Panebianco, M. V., and Capozzo, H. L. (2012). Bycatch of franciscana dolphins Pontoporia blainvillei and the dynamic of artisanal fisheries in the species’ southernmost area of distribution. Brazil. J. Oceanogr. 60, 149–168. doi: 10.1590/s1679-87592012000200005
Ohta, T., and Kimura, M. (1973). A model of mutation appropriate to estimate the number of electrophoretically detectable alleles in a finite population. Gen. Res. 22, 201–204. doi: 10.1017/s0016672300012994
Oremus, M., Poole, M. M., Steel, D., and Baker, C. S. (2007). Isolation and interchange among insular spinner dolphin communities in the South Pacific revealed by individual identification and genetic diversity. Mar. Ecol. Prog. Ser. 336, 275–289. doi: 10.3354/meps336275
Parsons, K. M., Noblea, L. R., Reidb, R. J., and Thompsonc, P. M. (2002). Mitochondrial genetic diversity and population structuring of UK bottlenose dolphins (Tursiops truncatus): is the NE Scotland population demographically and geographically isolated? Biol. Conserv. 108, 175–182. doi: 10.1016/s0006-3207(02)00103-9
Pichler, F. B., Robineau, D., Goodall, R. N. P., Meyer, M. A., Olivarría, C., and Baker, C. S. (2001). Origin and radiation of Southern Hemisphere coastal dolphins (genus Cephalorhynchus). Mol. Ecol. 10, 2215–2223. doi: 10.1046/j.0962-1083.2001.01360.x
Piry, S., Luikart, G., and Cornuet, J. M. (1999). BOTTLENECK: a computer program for detecting recent reductions in the effective population size using allele frequency data. J. Heredity 90, 502–503. doi: 10.1093/jhered/90.4.502
Pritchard, J. K., Stephens, M., and Donnelly, P. (2000). Inference of population structure using multilocus genotype data. Genetics 155, 945–959.
Raymond, M., and Rousset, F. (1995). GENEPOP (version 1.2): population genetics software for exact tests and ecumenicism. J. Heredity 86, 248–249. doi: 10.1093/oxfordjournals.jhered.a111573
Reeves, R. R., Smith, B. D., Crespo, E., Notarbartolo, and Di Sciara, G. (2003). Dolphins, whales and porpoises. 2002 – 2010 Conservation Action Plan for the world’s cetaceans. Gland: International Union for the Conservation of Nature.
Rosel, P. E., and Rojas-Bracho, L. (1999). Mitochondrial Dna Variation in The Critically Endangered Vaquita Phocoena Sinus Norris And Macfarland, 1958. Mar. Mamm. Sci. 15, 990–1003. doi: 10.1111/j.1748-7692.1999.tb00874.x
RRDM (2019). Relatório Anual – Integração a partir dos índices ecotoxicológicos. RT-28, Programa de Monitoramento da Biodiversidade Aquática. Brazil: Fundação Espírito-Santense de Tecnologia.
Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989). Molecular Cloning: a Laboratory Manual. New York, NY: Cold Spring Harbor Laboratory Press.
Secchi, E. R., Danilewicz, D., and Ott, P. H. (2003). Applying the phylogeographic concept to identify franciscana dolphin stocks: implications to meet management objectives. J. Cetacean Res. Manag. 5, 61–68.
Secchi, E. R., Wang, J. Y., Murray, B., Rocha-Campos, C. C., and White, B. N. (1998). Population differentiation in the franciscana (Pontoporia blainvillei) from two geographic locations in Brazil as determined from mitochondrial DNA control region sequences. Canadian J. Zool. 76, 1622–1627. doi: 10.1139/cjz-76-9-1622
Shinohara, M., Domingo-Roura, X., and Takenaka, O. (1997). Microsatellites in the bottlenose dolphin Tursiops truncatus. Mol. Ecol. 6, 695–696. doi: 10.1046/j.1365-294x.1997.00231.x
Siciliano, S. (1994). “Review of small cetaceans and fishery interactions in coastal waters of Brazil,” in Gillnets and cetaceans. Reports of the Internation Whaling Commission, eds W. F. Perrin, G. P. Donovan, and J. Barlow (Cambridge: International Whaling Commission).
Siciliano, S., Di Beneditto, A. P. M., and Ramos, R. M. A. (2002). A toninha, Pontoporia blainvillei (Gervais & d’Orbigny, 1844) (Mammalia, Cetacea. Pontoporiidae), nos Estados do Rio de Janeiro e Espírito Santo, costa sudeste do Brasil: caracterização dos habitas e fatores de isolamento das populações. Boletim do Museu Nacional Zool. 476, 1–15.
Torre, A. L., Alonso, M., Martinez, M. A., Sanz, P., Shen, L., Reiner, E. J., et al. (2012). Dechlorane related compounds in franciscana dolphin (Pontoporia blainvillei) from Southeastern and Southern coast of Brazil. Environ. Sci. Technol. 302:934.
Valsecchi, E., and Amos, W. (1996). Microsatellite markers for the study of cetacean populations. Mol. Ecol. 5, 151–156. doi: 10.1111/j.1365-294x.1996.tb00301.x
Van Oosterhout, C., Hutchinson, W. F., Wills, D. P. M., and Shipley, P. (2004). MICRO-CHECKER: software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 4, 535–538. doi: 10.1111/j.1471-8286.2004.00684.x
Waples, R. S. (1989). A generalized approach for estimating effective population size from temporal changes in allele frequency. Genetics 121, 379–391.
Waples, R. S., and Do, C. (2010). Linkage disequilibrium estimates of contemporary Ne using highly variable genetic markers: a largely untapped resource for applied conservation and evolution. Evol. Appl. 3, 244–262. doi: 10.1111/j.1752-4571.2009.00104.x
Zerbini, A. N., Secchi, E., Crespo, E., Danilewicz, D., and Reeves, R. (2017). Pontoporia blainvillei (errata version published in 2018). The IUCN Red List of Threatened Species 2017: e.T17978A123792204. Available online at: http://dx.doi.org/10.2305/IUCN.UK.2017-3.RLTS.T17978A50371075.en. (accessed on April 07, 2020).
Keywords: cetacean, conservation, threatened, Franciscana Management Area, microsatellites, Pontoporiidae, D-loop
Citation: de Oliveira VKM, Faria DM, Cunha HA, dos Santos TEC, Colosio AC, Barbosa LA, Freire MCC and Farro APC (2020) Low Genetic Diversity of the Endangered Franciscana (Pontoporia blainvillei) in Its Northernmost, Isolated Population (FMAIa, Espírito Santo, Brazil). Front. Mar. Sci. 7:608276. doi: 10.3389/fmars.2020.608276
Received: 19 September 2020; Accepted: 23 November 2020;
Published: 18 December 2020.
Edited by:
Salvatore Siciliano, Oswaldo Cruz Institute, Oswaldo Cruz Foundation (Fiocruz), BrazilReviewed by:
Susana Caballero, University of Los Andes, ColombiaAna Paula Di Beneditto, State University of the North Fluminense Darcy Ribeiro, Brazil
Copyright © 2020 de Oliveira, Faria, Cunha, dos Santos, Colosio, Barbosa, Freire and Farro. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Vanessa K. M. de Oliveira, dmFuZXNzYS5rdWJveWFtYUBob3RtYWlsLmNvbQ==