
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Mar. Sci. , 10 December 2020
Sec. Marine Megafauna
Volume 7 - 2020 | https://doi.org/10.3389/fmars.2020.602565
This article is part of the Research Topic Pathologic Findings in Stranded Marine Mammals: A Global Perspective View all 29 articles
 Frances M. D. Gulland1,2*
Frances M. D. Gulland1,2* Ailsa J. Hall3
Ailsa J. Hall3 Gina M. Ylitalo4
Gina M. Ylitalo4 Kathleen M. Colegrove5
Kathleen M. Colegrove5 Tenaya Norris1
Tenaya Norris1 Pádraig J. Duignan1
Pádraig J. Duignan1 Barbie Halaska1
Barbie Halaska1 Karina Acevedo Whitehouse1,6Linda J. Lowenstine2Alissa C. Deming1,7Teresa K. Rowles8
Karina Acevedo Whitehouse1,6Linda J. Lowenstine2Alissa C. Deming1,7Teresa K. Rowles8The prevalence of cancer in wild California sea lions (Zalophus californianus) is one of the highest amongst mammals, with 18–23% of adult animals examined post-mortem over the past 40 years having urogenital carcinoma. To date, organochlorines, genotype and infection with Otarine herpesvirus-1 (OtHV-1) have been identified in separate studies using distinct animals as associated with this carcinoma. Multi-year studies using large sample sizes to investigate the relative importance of multiple factors on marine mammal health are rare due to logistical and ethical challenges. The objective of this study was to use a case control approach with samples from 394 animals collected over 20 years in a multifactorial analysis to explore the relative importance of distinct factors identified to date as associated with sea lion cancer in the likelihood of sea lion carcinoma. Stepwise regression indicated that the best model to explain carcinoma occurrence included herpesvirus status, contaminant exposure, and blubber depth, but not genotype at a single microsatellite locus, PV11. The odds of carcinoma was 43.57 times higher in sea lions infected with OtHV-1 (95% CI 14.61, 129.96, p < 0.001), and 1.48 times higher for every unit increase in the loge[contaminant concentrations], ng g–1 (an approximate tripling of concentration), in their blubber (95% CI 1.11, 1.97, p < 0.007), after controlling for the effect of blubber depth. These findings demonstrate the importance of contaminant exposure combined with OtHV1 infection, in the potential for cancer occurrence in wild sea lions.
California sea lions (Zalophus californianus) are long lived marine mammals that are resident along the west coast of North America, with most of the population of approximately 250,000 individuals breeding on islands off the central California coast (Laake et al., 2018). This population has among the highest prevalence of cancer in mammals, with 18–23% of adult animals examined post-mortem over the past 40 years having neoplasia (Gulland et al., 1996; Deming et al., 2018). The most common type of cancer is urogenital carcinoma (UGC) that is fatal due to widespread metastasis (Gulland et al., 1996). As cancer is rare in wild mammals, with the notable exception of transmissible tumors in Tasmanian devils Sarcophilus harrisii (Pearse and Swift, 2006), it is important to understand sea lion cancer pathogenesis and the potential implications for the health of humans who share the coastal environment and some seafood types with sea lions (Browning et al., 2015). Sea lion cancer is not clonal, unlike that of Tasmanian devils, thus exposure to exogenous factors such as environmental contaminants and infections as well as genetic susceptibility must be considered as causal factors (Leathlobhair et al., 2017).
The central California coast and its food web are exposed to high levels of potentially carcinogenic persistent organochlorines following dumping of industrial waste DDTs (dichlorodiphenyltrichloroethanes) in the 1960s and urbanization and industrialization of the coast increasing run-off of newer chemical contaminants (Young et al., 1976; McCarthy et al., 2008). Organochlorines have been associated with cancers in humans and are carcinogenic in laboratory animal models (Wolff et al., 1993; Longnecker et al., 1997; Lauby-Secretan et al., 2013). Exposure to pollutants has been associated with cancer in another marine mammal, the beluga (Delphinapterus leucas) of the St Lawrence River (De Guise et al., 1995; Martineau et al., 2002). Earlier studies on California sea lion cancer found samples of blubber from sea lions with UGC had significantly higher levels of polychlorinated biphenyls (PCBs) and DDTs than blubber from sea lions without carcinoma (Ylitalo et al., 2005b; Randhawa et al., 2015). Other factors, however, also have been associated with sea lion UGC. A virus, Otarine herpesvirus-1 (OtHV-1), detected in sea lion tumors is phylogenetically similar to the human oncogenic gammaherpesviruses Kaposi sarcoma virus and Epstein Barr virus, suggesting it could be important in oncogenesis (King et al., 2002; Lipscomb et al., 2005). Although observed in sea lion tumors, it also has been detected in secretions and tissues from healthy sea lions, raising questions about its role in cancer development (Buckles et al., 2006, 2007).
Genetic predisposition to cancer also is possible in some sea lions, as carcinoma is more common in relatively inbred California sea lions (Acevedo-Whitehouse et al., 2003). This effect is apparently due to a single microsatellite genetic marker (PV11) (Browning et al., 2014). PV11 maps to intron 9 of the heparanase 2 gene (HPSE2) locus, a large gene which in humans is associated with multiple carcinomas. Expression of heparanase 2 (HPA2) was only found in the genital tract of sea lions with UGC with a homozygous PV11 allele 1 genotype (Browning et al., 2014). Furthermore, a specific MHC class II locus, Zaca-DRB-A, was associated with an increased occurrence of cancer in a small sample of stranded sea lions (Bowen et al., 2005).
Carcinogenesis is a multistep process requiring multiple mutagenic events, often with a long latency period between the initial DNA damage and the onset of neoplasia (Nik-Zainal and Hall, 2019). Here we hypothesize that genetic predisposition in California sea lions interacts with herpesvirus infection and environmental factors including contaminant exposure as the multi-step process resulting in neoplasia. Research to date on the pathogenesis of California sea lion UGC has evaluated the association of single factors with this carcinoma. Each report has used small sample sizes and investigated the association of a single factor with sea lion carcinoma. The aim of this study is to determine the relative importance of the factors identified to date (persistent organic contaminant exposure, herpesvirus infection, and PV11 microsatellite genotype), as well as the confounding variables of sea lion age and body condition, in increasing the likelihood of occurrence of sea lion UGC. The objective is to use a case control study using samples from hundreds of wild sea lions collected over 20 years, an approach which is rare in marine mammals due to the logistical and ethical challenges in sampling significant numbers of animals and obtaining samples from non-diseased animals. This study builds on earlier data from stranded California sea lions. Samples from some cases have been used in previous studies examining the individual factors of interest in the pathogenesis of the disease (contaminants, herpesviruses, and PV11 genotype) and results have been published elsewhere (Ylitalo et al., 2005b; Buckles et al., 2006; Randhawa et al., 2015). Here we include additional samples not previously reported on to the dataset and combine all the results into an integrated analysis to determine the contribution of the various factors associated with sea lion cancer.
Adult female California sea lions that died from various causes (trauma, domoic acid toxicosis, cancer) at The Marine Mammal Center (TMMC) (Sausalito, CA, United States) following stranding along the California coast from San Luis Obispo to Humboldt Counties between 1st January 1998 and 30th December 2017 contributed cases or controls to this study (n = 394). California sea lions are a coastal species: after the breeding season when animals are concentrated on the Channel Islands, individuals migrate north along the coast as far as southern Canada to forage, with adult males migrating further north than females that return to the islands between foraging trips to nurse pups (DeLong et al., 2017). Sea lions that are observed stranded and appear sick or injured at various sites along the coast during the foraging season are taken to TMMC for treatment, and all come from the Channel Islands population (Carretta et al., 2018). Only female animals were included as few male sea lions without cancer are found stranded along the central California coast (Greig et al., 2005). Controls were mostly sea lions stranded with domoic acid toxicosis or trauma. All animal care procedures and sampling were authorized by the National Marine Fisheries Service (MMPA permit no 18786) and approved by The Marine Mammal Center’s Internal Animal Care and Use Committee. Within 24 h of death, each animal was examined at post-mortem, and straight length and blubber depth over the sternum measured. Representative samples of each organ including any grossly apparent masses, and the entire urogenital tract, were fixed in 10% neutral buffered formalin, then embedded in paraffin, processed routinely, sectioned at 5 μm and stained with hematoxylin and eosin for light microscopy. To determine the presence of UGC multiple tissues for histologic evaluation were sampled from the reproductive tract including uterus, ovaries, cervix, and vagina, and all histology was reviewed by one pathologist (KMC). A 5 mm3 skin sample from each animal was fixed in 70% ethanol for genetic testing, and 1 cm3 samples of cervix and/or vagina were frozen at −70°C for viral testing. Samples of liver, kidney, and skeletal muscle were archived from each animal at −20°C and used for genetic testing in cases for which skin samples were missing.
An approximately 100 g sample of blubber was collected from over the sternum, wrapped in Teflon and stored at -20°C until analysis for contaminants.
Each sea lion was assigned as a case or control based on histology: “controls,” no UGC observed on histology (n = 215); “cases,” UGC in reproductive tract and metastatic carcinoma in at least one lymph node or other organ (n = 179). In all cases, histologic features of UGC were as previously described (Gulland et al., 1996). Thus, controls came from the same population that gave rise to the cases and if they had cancer they would have been assigned as a case. The controls therefore represent those animals in the population that may have become cancer cases. All animals were selected independently of exposure.
Persistent organic contaminant concentrations in the blubber of sea lions represent past chemical exposure as these compounds are highly lipophilic and accumulate in the blubber tissue throughout the animals’ lifetime (Randhawa et al., 2015). However, the greatest exposure occurs in utero and during lactation when the legacy of contaminants is passed from mother to pup (Greig et al., 2007). Thus, there is a positive correlation between contaminants in the blubber of the pup and the milk of the mother; pups from females with high concentrations in their blubber and milk will start life with a higher exposure before they are even weaned and feeding independently.
Blubber samples were analyzed for PCBs, DDTs, and other organochlorine (OC) pesticides using either a gas chromatography/mass spectrometry (GC/MS) method coupled with gravimetric lipid determinations or a high-performance liquid chromatography/photodiode array (HPLC/PDA) method coupled with a thin layer chromatography/flame ionization detection (TLC/FID) method for percent lipid measures (Ylitalo et al., 2005a; Sloan et al., 2014). In general, the OC results obtained using these two methods have been shown to be comparable for a number of matrices, including marine mammal blubber (Ylitalo et al., 2005a). The blubber lipid content values determined gravimetrically or via TLC/FID also have been shown to be comparable, although the TLC/FID values tend to be approximately 10–15% lower (Ylitalo et al., 2005c).
The majority of the blubber samples (267 of 296) were analyzed for OCs and polybrominated diphenyl ether (PBDE) flame retardants using the GC/MS method (Sloan et al., 2014). Briefly, blubber samples were weighed (0.5–1.0 g), mixed with drying agents (sodium sulfate and magnesium sulfate), and extracted using dichloromethane in an accelerated solvent extraction procedure, followed by removal of approximately 1 mL of sample extract for gravimetric percent lipid determinations. Next, the sample extracts went through a two-step clean-up regime to remove polar compounds via a single stacked silica gel/alumina column and to separate the compounds of interest from bulk lipids and other biogenic compounds by high-performance size exclusion liquid chromatography. The cleaned-up extracts were then analyzed on a low-resolution quadrupole GC/MS system equipped with a 60-meter DB-5 GC capillary column. In this method, sum PCBs is the summed concentrations of congener PCBs 17, 18, 28, 31, 33, 44, 49, 52, 66, 70, 74, 82, 87, 95, 99, 101/90, 105, 110, 118, 128, 138/163/164, 149, 151, 153/132, 156, 158, 170, 171, 177, 180, 183, 187/159/182, 191, 194, 195, 199, 205, 206, 208, 209. Sum DDTs is the sum levels of o,p’-DDD, p,p’-DDD, o,p’-DDE, p,p’-DDE, o,p’-DDT and p,p’-DDT. Sum chlordanes is the summed concentrations of oxychlordane, gamma-chlordane, nona-III-chlordane, alpha-chlordane, trans-non-achlor, and cis-non-achlor. Sum hexachlorocyclohexanes (HCHs) includes the sum levels of alpha-, beta-, and gamma-HCH isomers and sum PBDEs is the sum concentrations of congeners 28, 47, 49, 66, 85, 99, 100, 153, 154, 183.
A further 29 blubber samples were analyzed using HPLC/PDA (Ylitalo et al., 2005b), and were weighed (0.2–0.3 g), mixed with hexane/pentane (1:1 v/v) and sodium sulfate (5 g), and then homogenized using a Tekmar Tissuemizers. The analytes of interest were separated from lipids and other interfering compounds on a gravity flow cleanup column that contained neutral, basic and acidic silica gels by eluting with hexane/dichloromethane (1:1 v/v). Prior to the cleanup step, approximately 1 mL of sample extract was removed for lipid quantitation by thin layer chromatography with flame ionization detection (TLC/FID) (Ylitalo et al., 2005a). Eight dioxin-like congeners (PCBs 77, 105, 118, 126, 156, 157, 169, 189) were resolved from other selected PCBs (PCBs 99/101/149/196, 110, 128/123, 138, 153/87, 170/194, 180, 190, 200) and six additional OCs [o,p’-DDD, p,p’-DDD, p,p’-DDE, o,p’-DDT, p,p’-DDT, hexachlorobenzene (HCB)] by HPLC on two Cosmosil PYE analytical columns, connected in series and cooled to 16°C. The OCs were measured by an ultraviolet (UV) photodiode array detector and were identified by comparing their UV spectra (200–310 nm) and retention times to those of reference standards. The analyte purity was confirmed by comparing spectra within a peak to the apex spectrum. In the HPLC/PDA method, concentrations of summed PCBs were calculated by summing the concentrations of the PCBs listed above (based on individual response factor) and summing the concentrations of other PCBs (calculated by summing areas of peaks identified as PCBs and using an average PCB response factor). Using an average PCB response factor was justified for the “other PCBs” measured in the samples due to similarities in the PCB congener response factors (ranging from 0.65 to 0.75) measured on the HPLC/PDA system. Based on the retention time and UV spectral data, the “other PCBs” were identified as PCB congeners 28, 52, 47, 66, 70, 182, 200, and 209. Summed DDT concentrations were calculated by adding the concentrations of the five DDT isomers.
Because a small number of blubber samples were analyzed for percent lipid by TLC/FID, we converted the TLC/FID values to approximate the gravimetric percent lipid measures using the following equation previously reported for California sea lion blubber samples (Randhawa et al., 2015):
For both OC analytical methods, a solvent method blank and a National Institute of Standards and Technology whale blubber Standard Reference Material (SRM, 1945) were analyzed with each sample batch and the results for the blubber SRM and other quality control samples met established laboratory criteria (Sloan et al., 2019).
Frozen cervical or vaginal samples from 162 control and 121 cancer cases were tested at the University of Florida College of Veterinary Medicine Zoological Medicine and Wildlife Disease Laboratory for the presence of herpesvirus using an OtHV1-specific qPCR (polymerase chain reaction) targeting the unique areas of the DNA polymerase gene (Dpol; forward primer: OtHV1qPCRF 5′-TCCCACGCTGTTTCGAATG-3′, reverse primer: OtHV1qPCRR 5′-AGCTCCGAGTCGTGTACACAGTAT-3′, probe: OtHV-1 Probe 5′-{FAM}-TCGCGCTCGCATCGGCA-{BNQ]-3′) with Black Hole Quencher probe and FAM reporter dye (Deming, 2018). Total DNA was extracted using a Qiagen kit according to manufacturer’s instructions (DNeasy Blood and Tissue, Qiagen Inc., Valencia, California, United States). DNA concentration of extracted samples were quantified using a NanoDrop 8000 spectrophotometer (Therma Fisher Scientific, Wilmington, Delaware, United States). qPCR reactions were run on 96-well polypropylene plates (Olympus Plastics, Genesee Scientific, San Diego, CA, United States). All plates had three no template (molecular-grade water) negative controls and standard curves (10–107 in triplicate). Each 20 μL reaction was composed of 4 μL DNA extract, 10 μL qPCR Master mix (TaqMan Fast Universal PCR Master Mix 2X, Applied Biosystems), 3 μL of molecular grade water, and 1 μL of each primer at a dilution of 18 μM. Samples were run in triplicate with an internal positive control of 18S ribosomal universal eukaryote DNA primer/probe (VIC Probe, Applied Biosystems) on a 7500 Fast Real-Time PCR System (Applied Biosystems) using the standard Fast protocol with thermocycling conditions as described in Deming (2018) (94°C for 20 s once, followed by 45 cycles at 94°C for 3 s and 60°C for 30 s). Data were analyzed using 7500 Fast Real-Time PCR System software. Samples with viral concentrations that were below the lower detection limit (1,000 viral copies per ng of DNA) were assigned an OtHV1 negative status and samples above the lower detection limit were assigned an OtHV1 positive status.
Vaginal samples from a further 15 control and three cancer animals were tested for OtHV-1 at the University of St. Andrews following PCR protocols described in Browning (2014) using OtHV-1 specific PCR primers that target a unique fragment of the OtHV-1 Dpol gene (forward primer: PolFor 5′-TTA CAC TTC TAC GTG ATG-3′, reverse primer: PolRev 5′-TCT TCG TCC AGT ATC ATT G G-3′) (Buckles et al., 2006; Browning, 2014). To confirm OtHV-1 positive PCR results, all samples that produced a positive band of expected size (approximately 740 bp) were gel extracted, sequenced (MRCPPU, College of Life Sciences, University of Dundee, Scotland)1 and aligned to the Dpol gene from OtHV-1 (GenBank: AF236050). All negative samples were re-run to confirm negative result.
Genotyping was carried out as described in Browning et al. (2014) (n = 317, Table 1). Briefly, genomic DNA (gDNA) was extracted following either a proteinase K-chelex DNA isolation method followed by phenol chloroform purification or using the PUREGENE DNA isolation method according to the manufacturer’s instructions. The microsatellite marker PV11 was amplified by PCR using Qiagen Multiplex Master Mix (Qiagen Inc.) and fluorescently tagged primers (Applied Biosystems). This enabled fragment analysis via automated capillary electrophoresis (ABI3700, Applied Biosystems or Beckman Coulter CEQ 8000) and subsequent allele identification. To check for errors in the amplification, a minimum of 10% of the samples were run in triplicate and two negative controls were included in each plate. Additional samples to those reported in Browning et al. (2014) were analyzed by Xelect Ltd., St Andrews, United Kingdom, using a Sanger sequencer to determine the fragment length following PCR including 10 repeats to ensure comparability. Five alleles were identified from fragment lengths (Browning, 2014), and homo- or heterozygosity assigned to each individual.
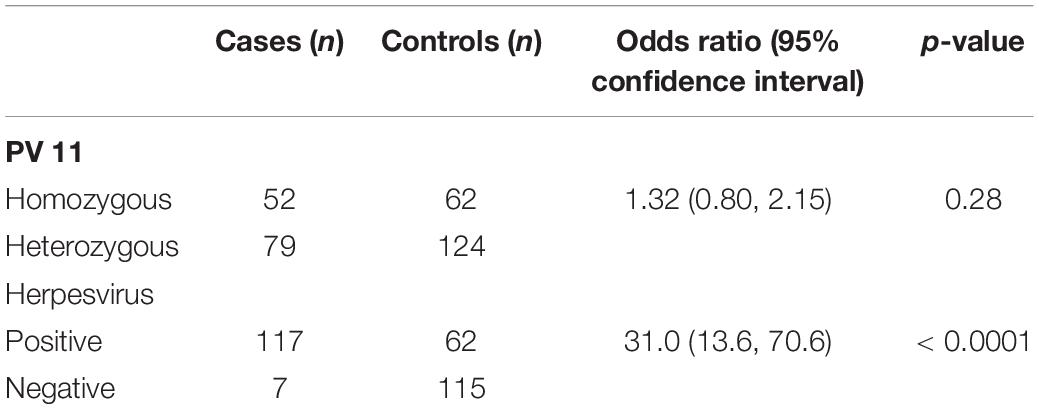
Table 1. Odds ratios for PV11 microsatellite genotype and herpesvirus status in sea lion cancer cases and controls.
Whilst every attempt was made to obtain results for all factors for all individuals, there were inevitably some missing results and data due to the loss of samples during storage and transport and due to analytical error. However, these losses occurred randomly and were not decisions made by the investigators.
We aimed to determine the herpesvirus status (positive or negative), genotype at PV11 (heterozygote or homozygote), and contaminant concentrations (sum PCB, DDTs, CHLDs, PBDEs, and HCHs on lipid weight basis) for each animal, but not all factors were available for each individuals in the dataset so sample sizes vary accordingly. Contaminant concentrations (on a lipid weight basis) were log-normally distributed and were therefore loge transformed after summation. The distribution of contaminants for the cases and controls are shown in Figure 1. The analysis was carried out using a staged approach with the relationship between the exposures (contaminants, genotype, and herpesvirus) and the probability of cancer first being considered singly, and with potential confounding factors, and then together using a multivariable approach. Fisher’s exact tests were carried out on the single exposure variable contingency tables to assess for independence between the variables and whether to reject the null hypothesis of the odds ratio being 1.0. Associations between the exposures and carcinoma were examined using logistic regression (generalized linear models with a binomial family and logit link function) to estimate odds ratios and their 95% confidence intervals using the package epitools in R (Aragon, 2018; R Core Team, 2020). Multivariable logistic regression analyses were then used to investigate the combined effects of the factors and odds ratios were calculated from the logistic regression coefficients for each factor classifying sea lions as controls and cases. The backward step function in R was used to determine the best fitting model which minimized Akaike’s Information Criterion (AIC), adjusting for covariates (Burnham and Anderson, 2004). The effect of blubber depth, as a potential confounding factor was investigated, together with any effect of year and season. A number of models were fitted to the data to determine the most parsimonious set of biologically plausible linear and non-linear predictors. Models were assessed for goodness of fit and those for which there was sufficient data which were therefore useful for making further inferences were included (see Supplementary Information).
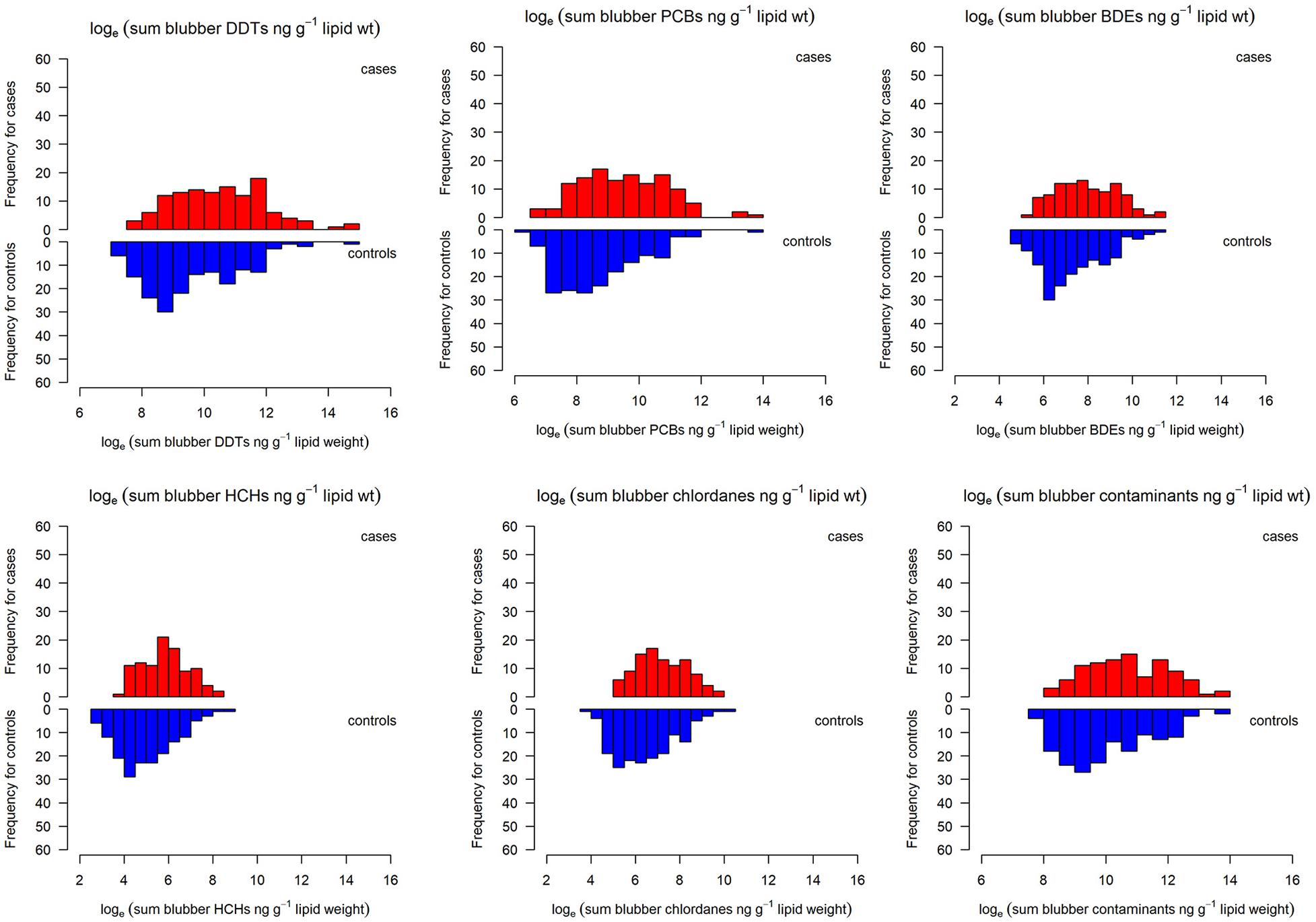
Figure 1. Blubber contaminant concentrations (loge transformed, ng g–1 on a lipid weight basis) in California sea lion cancer cases (red) and controls (blue) by contaminant classes.
A total of 179 adult female California sea lions with UGC and 215 adult female controls were included in the study. The potential role of each of the factors was investigated sequentially.
Carcinoma was not significantly associated with homozygosity (i.e., any combination of identical alleles compared to any combination of heterozygosity) at the PV11 microsatellite locus (n = 317, odds ratio = 1.32 (95% CI 0.80, 2.15), Fisher’s exact test p = 0.28, Table 1).
The probability of being a cancer case also was not significantly different among the four homozygous allele combinations (i.e., Group 1 alleles 1,1; Group 2 alleles 2,2; Group 3 alleles 3,3; Group 4 alleles 4,4—logistic regression intercept only model, null deviance = 157.16, homozygote groups model null deviance = 152.73, overall χ2 p = 0.218, Nagelkerke pseudo-R2 = 0.051, see Supplementary Information for further details).
The animals were then categorized according to their allele combination (i.e., 1,1; 1,2; 1,3; 1,4; 2,2; 2,3 etc.), with the aim of determining if the probability of cancer was related to a particular allele occurring in any combination, homozygote or heterozygote. None of the combinations were significantly associated with cancer (intercept only model, null deviance = 429.86; allele group model null deviance = 413.21; overall χ2 p = 0.163, Nagelkerke R2 = 0.069, see Supplementary Information for further details).
There was a significant association between herpesvirus status and carcinoma. The odds of being virus positive were significantly greater in the cases than the controls (n = 301, odds ratio = 31.0, Fisher’s exact p < 0.0001, Table 1).
Finally, the association between blubber contaminant concentrations and carcinoma was investigated. There was no relationship between contaminants and sea lion length (a proxy for age), and no difference in sea lion length between cases and controls (p = 0.812). The relationship between contaminant exposure and carcinoma was first investigated by combining the blubber concentrations of the five class (DDTs, PCBs, CHLDs, PBDEs, and HCHs) into one contaminant variable because there was a highly significant positive linear relationship among all five contaminant classes (Figure 2), as a combined variable as the sum of all contaminants measured. There was a significant negative relationship between blubber contaminant concentrations and blubber depth (p < 0.0001, adjusted R2 = 0.270, see Supplementary Information, and Figure S1). However, their variance inflation factors indicated a low degree of multicollinearity (blubber contaminant concentrations VIF = 1.27; blubber depth VIF = 1.23).

Figure 2. Relationship between blubber concentrations in different contaminant classes (loge transformed, lipid weight, ng g–1). DDTs, Dichlorodiphenyltrichloroethanes; PCBs, polychlorinated biphenyls; CHLDs, chlordanes; BDEs, polybrominated diphenyl ethers; HCHs, hexachlorocyclohexanes.
The odds ratio indicated that, after controlling for differences in blubber depth (cm) (four animals that were missing data on blubber depths were excluded from this analysis), the odds of cancer increased with increasing blubber contaminant concentrations. For the sum of all the contaminants measured, the odds ratio was 1.28 (n = 263, odds ratio 95% CI 1.02, 1.6, p = 0.03) indicating that holding blubber depth constant, the odds of cancer are 28% higher for each unit increase in loge[blubber contaminant concentrations, ng g–1] Table 2). As the contaminants have been loge transformed this equates to approximately a tripling (× 2.72) of concentration for each unit increase. Comparing the results and fit of a model which included loge[blubber contaminant concentrations] to one with both loge[blubber contaminant concentrations] and blubber depth indicated that the latter was more appropriate (analysis of deviance for the generalized linear model with a logit link function and binomial family with blubber contaminant concentrations as an independent variable without blubber depth vs. model with blubber depth; χ2 Pr = 0.0133, crude odds ratio = 1.48, adjusted odds ratio = 1.28, see Supplementary Information). There was thus a 13.5% change in the odds ratio between the two models, indicating blubber depth is a confounding variable. However, as animals will lose blubber mass during the disease process, resulting in an increase in their blubber contaminant concentrations (Hall et al., 2008), a further analysis was carried out including only those animals with a blubber depth > 1 cm (n = 139). For this subset, using the same model structure (Case or Control ∼ blubber depth + loge [blubber contaminant concentrations]), controlling for the effect of blubber depth the odds ratio was higher [odds ratio = 1.97 (95% CI 1.35, 2.89) p < 0.001, Nagelkerke R2 = 0.221]. This suggests that the observed relationship with contaminant exposure is not only because animals have lost blubber due to disease.
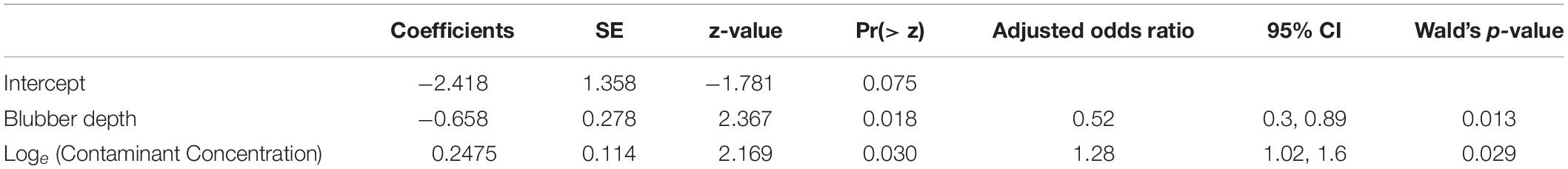
Table 2. Logistic regression model investigating probability of cancer in relation to blubber contaminant concentrations (loge[sum contaminant concentrations, ng g–1 lipid weight]) after controlling for blubber depth (glm[CaseControl ∼ BlubberDepth] + Loge[ContaminantConcentration], n = 263).
Stepwise multivariable logistic regression to investigate the relationship between carcinoma and the three exposures, genotype, herpesvirus, and blubber contaminants whilst accounting for the potential effect of confounding variables. There was no evidence for year or season being confounding variables (further details are given in Supplementary Information). The most parsimonious model included herpesvirus status, contaminant exposure, and blubber depth (Table 3, see also Supplementary Information). Genotype at microsatellite PV11 and body length were also not retained in the model. Models with interaction terms for the non-correlated variables (body length, blubber contaminants, year, season, and genotype) were tested but were not good fits to the data and were not retained in the final model.
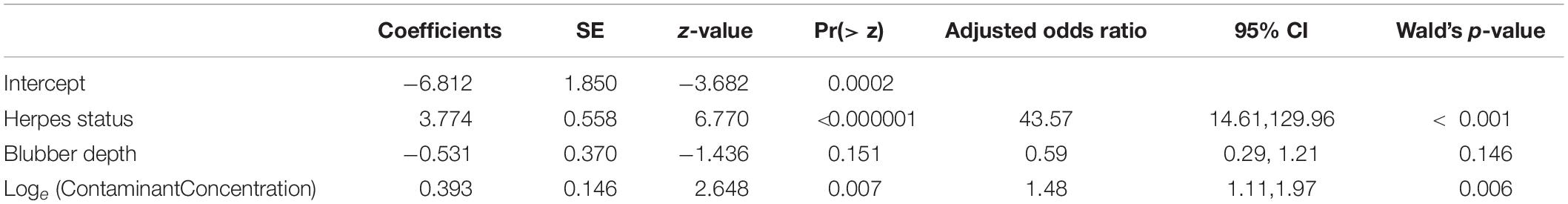
Table 3. Logistic regression model investigating probability of cancer in relation to herpesvirus status and blubber contaminant concentrations (loge[sum contaminant concentrations, ng g–1 lipid weight]) after controlling for blubber depth (glm[CaseControl ∼ HerpesStatus] + BlubberDepth + Loge[ContaminantConcentration], n = 252).
The adjusted odds ratio from the logistic regression model for herpesvirus was 43.57 (95% CI 14.61, 129.96, p < 0.001) and that for the contaminant concentration was 1.48 (1.11, 1.97, p < 0.007) again accounting for any effect of blubber depth. However, it should be noted that there were only seven sea lions with cancer that were negative for herpesvirus, and contaminant data were only available for four of these individuals (Figure 3). Nonetheless, the individual factor analysis indicates an effect of contaminants which is retained when herpesvirus status is accounted for. Probabilities estimated by the model (from the model logits) for animals with and without herpesvirus, for three different blubber depths, with increasing blubber contaminant concentrations, are shown in Figure 4. Animals with the highest blubber concentrations and without herpesvirus have a less than 20% probability of being a cancer case whereas for an animal with herpesvirus this increases from about a 30% probability in the least exposed to over 80% in the most exposed.
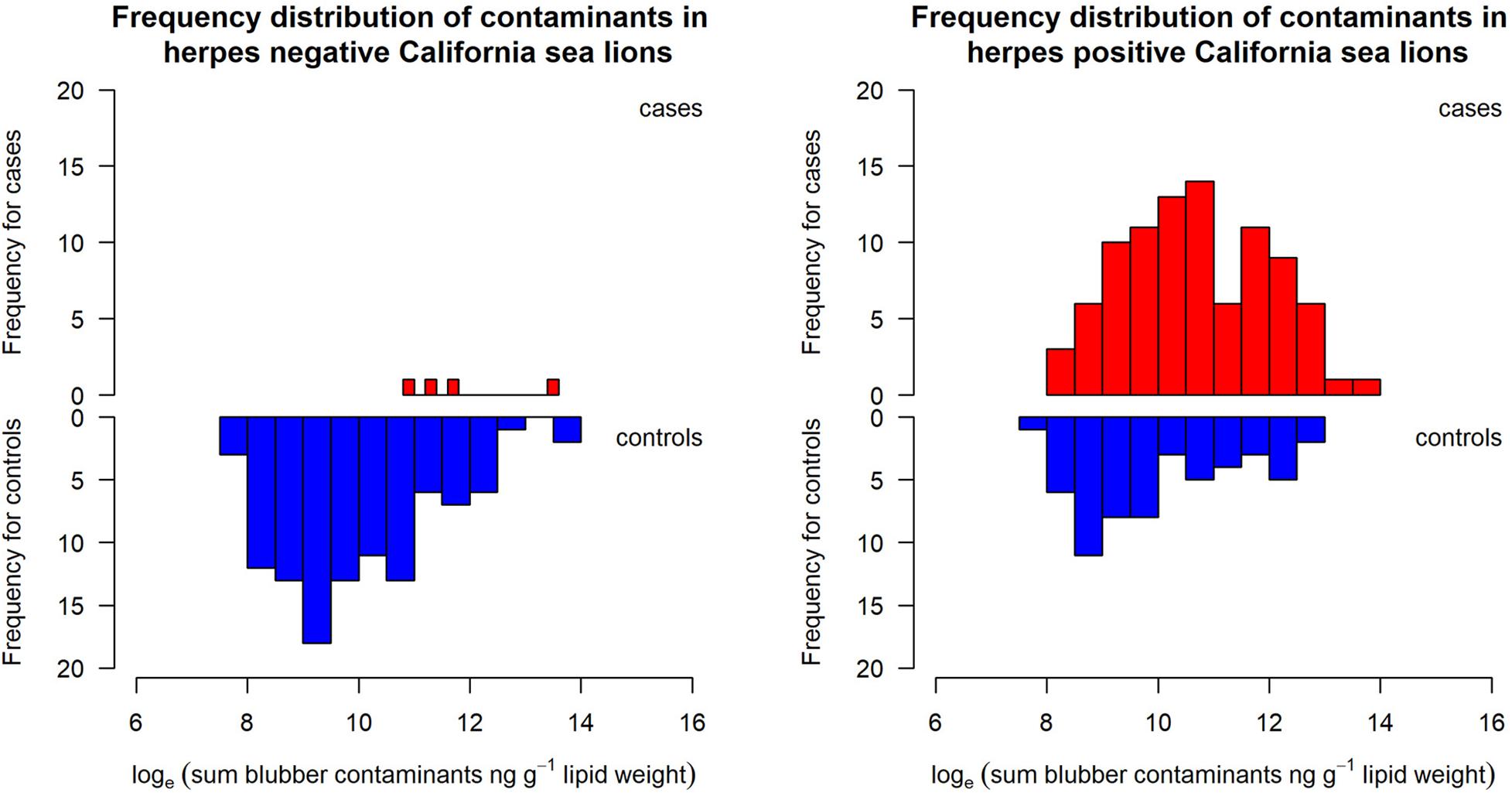
Figure 3. Distribution of blubber contaminant concentrations in herpesvirus negative (left panel) and herpesvirus positive (right panel) sea lions with (cases) (red) and without (controls) (blue) cancer. Note only four animals with cancer that also had contaminant data were negative for herpesvirus.
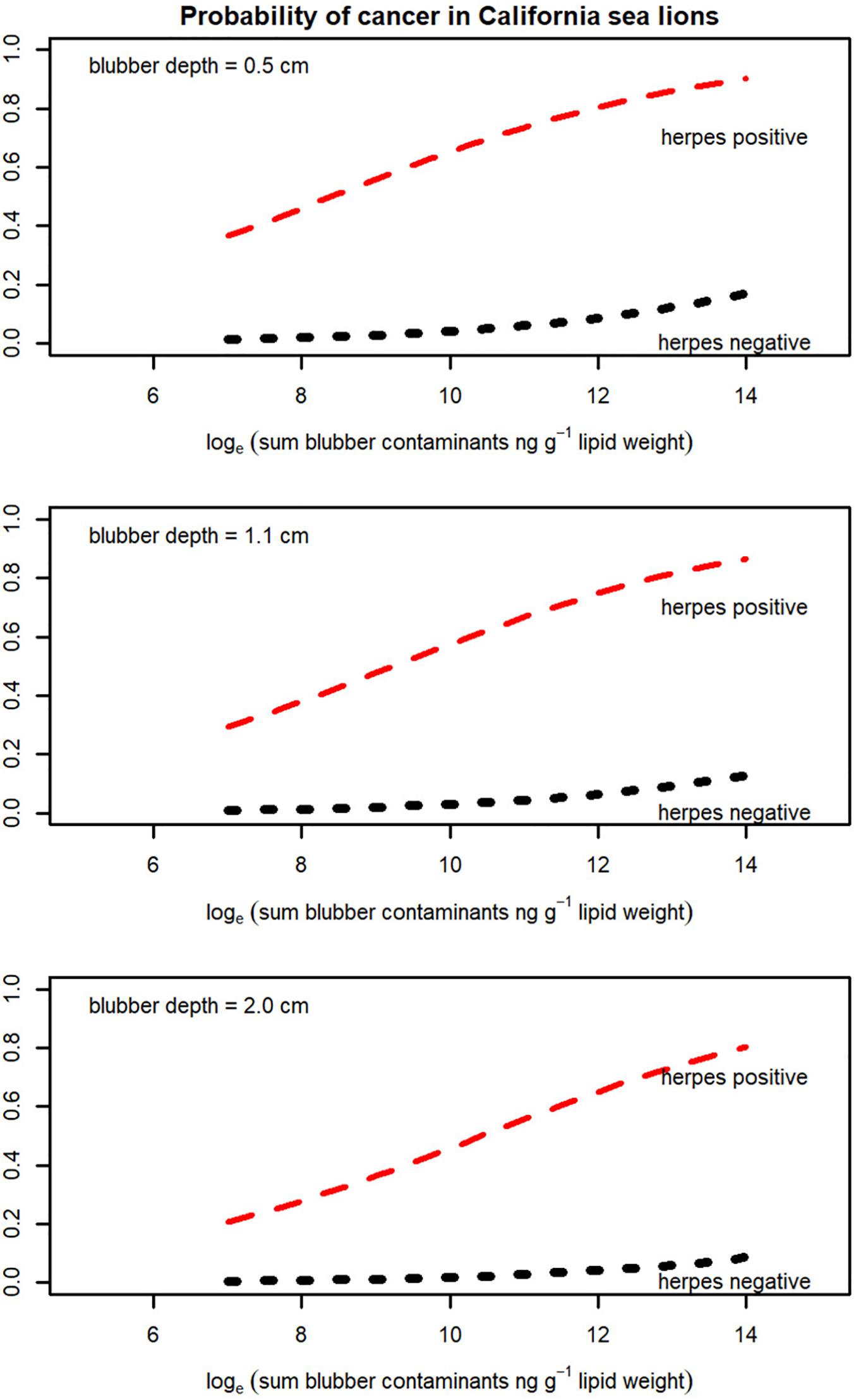
Figure 4. Probability of cancer in California sea lions with increasing blubber contaminant concentrations for animals with thin blubber (0.5 cm), the mean blubber thickness for the study group (1.1 cm) and with thick blubber (2.0 cm). Red = animals with herpesvirus; Black = animals without herpesvirus.
This California sea lion case control study provides evidence that herpesvirus infection (OtHV1) is critical to the likelihood of carcinoma occurrence, but in addition, the higher the animals’ blubber contaminant concentrations, the higher the odds of cancer. Such synergism between pollutants and virus in causing cancer has been suggested previously in humans but not wildlife: the high prevalence of esophageal carcinoma in people in the Huaihe River Basin in China is associated with human papillomavirus and exposure to 3-methylcholanthrene (Ghaffar et al., 2018). As carcinogenesis is a multistep process requiring multiple mutagenic events and may necessitate replication and fixing of the DNA damage caused by promoters, this study supports the hypothesis that carcinoma in California sea lions is a multifactorial disease. The gammaherpesvirus of sea lions is likely sexually transmitted, as is more common in sexually mature animals and is localized to the reproductive tract (Buckles et al., 2006, 2007). Exposure to contaminants, in contrast, occurs early in life, with California sea lions accruing PCBs and DDTs both in utero, across the placenta, and in milk (Greig et al., 2007). Thus, as there is often a long latency period between the initial DNA damage and the onset of neoplasia (Robertson and Hansen, 2020), this study suggests early exposure to persistent organic pollutants, followed by infection with OtHV1, are important in the pathogenesis of UGC in wild sea lions. Despite earlier work showing statistically significant associations between a single microsatellite genetic marker, PV11, and California sea lion carcinoma, this study with larger sample sizes and inclusion of other factors, did not find association between PV11 genotype and cancer, suggesting confounding factors may have been important in earlier studies.
Chemicals may induce neoplasia directly through DNA damage, or indirectly by acting as a promotor, or through immune suppression allowing infection with oncogenic viruses (Robertson and Hansen, 2020). PCBs are recognized as complete carcinogens (Lauby-Secretan et al., 2013), although this study suggests coinfection with herpesvirus is needed for urogenital cancer to occur in sea lions. Some contaminants have been linked to immune suppression in marine mammals (Ross et al., 1995; Levin et al., 2005; Williams et al., 2020), although others can increase lymphocyte proliferation and cytotoxicity (Peñín et al., 2018), suggesting that they can modulate anti-viral and tumor-surveillance activities of Natural Killer and cytotoxic T-cells. Such an effect would not necessarily lead to generalized states of immune suppression, which is improbable given that the sea lion population has been increasing over recent decades (Laake et al., 2018). Generalized immunosuppression is thus unlikely to be the mechanism through which persistent organic pollutants increase the likelihood of cancer in this species. Rather, it is possible that modulation of the immune system by organic contaminants can lead to variations in effectiveness of immune responses to OtHV-1 and cellular transformation. Another possibility is that persistent organic pollutants may increase the likelihood of cancer via their hormone mimicking properties, as sea lion reproductive tracts which are prone to cancer have estrogen and progesterone receptors and alteration of receptor expression is noted in UGC (Colegrove et al., 2009). The endocrine-disrupting properties of organochlorine pesticides are well recognized, as are the endocrine associations with cancer. For example, in humans, girls born to mothers receiving diethylstilbestrol are at higher risk of developing reproductive tract cancer (Colborn et al., 1993; Verloop et al., 2010). Also, human reproductive tract cancer is associated with higher blood levels of pesticides, and in utero exposure to DDTs increases the risk of breast cancer later in life (Mathur et al., 2008; Cohn et al., 2015). Experimental studies in laboratory animals have also shown interactions between organochlorines and hormone receptors in the genital tract. Mink exposed to organochlorines exhibit upregulation of hormone receptors (Patnode and Curtis, 1994). Neonatal mice exposed to estrogenic PCB metabolites developed cervicovaginal carcinomas (Martinez et al., 2005).
The blubber levels of persistent organic pollutants in these California sea lions are amongst the highest recorded in any marine mammal, presumably because of the persistence of compounds dumped along the California coast in the 1970s prior to the ban on their use and production in the United States, and the coastal foraging range of sea lions. Such high persistent organic pollutant levels were associated with carcinoma in two earlier studies (Ylitalo et al., 2005b; Randhawa et al., 2015). However, in neither of these studies was the effect of weight loss in animals with cancer considered. As persistent organic pollutants are lipid soluble and stored in adipose tissue of the blubber, when pinnipeds lose weight due to disease such as cancer, the blubber lipid content decreases and persistent organic pollutants are mobilized (Peterson et al., 2014). Serial sampling of sea lions that stranded suffering from domoic acid intoxication that lost weight then regained it after therapy demonstrated increases, followed by decreases, in blubber sum PCB and DDT concentrations, with different congeners having differing dynamics (Hall et al., 2008). To allow for these dynamics, in this study the effect of blubber thickness was controlled for, demonstrating persistent organic pollutant levels remain a significant factor in cancer development, even when considering the effects of weight loss. Age of female sea lions also likely affects blubber persistent organic pollutant burden, although in this study, there was no relationship between sea lion length, a proxy for age, and contaminants, and length was not retained in the model. In bottlenose dolphins (Tursiops truncatus), it has been shown that transfer of PCBs to offspring via the placenta and milk will decrease organochlorine burden with age in reproductively active female dolphins (Schwacke et al., 2002). Length at age for California sea lions has been established (Laake et al., 2016), and in this study most (80%) of these animals were over 5 years old and fully grown (> 153 cm long). Data on reproductive history for these wild animals are unavailable, but given the high fecundity of this species, and the high number of fully grown animals, it is unlikely that animals in the control group have different life histories from those in the case group. However, this remains a source of uncertainty in the results.
The clear association of OtHV-1 with carcinoma in this study supports a significant role for OtHV-1 in the pathogenesis of sea lion cancer. Although this virus has been previously demonstrated in California sea lion cancers, its presence in secretions from apparently healthy animals raised the question of its role in cancer development, and the possibility of it simply replicating in rapidly divided neoplastic tissue rather than playing a role in tumor development. Herpesviruses are associated with other mammalian tumors, including in primates and humans with the effects of infection typically depending upon multiple factors (Hunt et al., 1972; Touitou et al., 2001; Estep and Wong, 2013). For example, Epstein Barr virus (EBV) in humans is usually sub-clinical, or causes a self-limiting lymphoproliferative disease; however, when combined with co-factors, such as malaria or HIV infection, EBV infection can result in the development of lymphomas or nasal pharyngeal carcinoma (Touitou et al., 2001). Results of this larger study provides a clear example of a herpesvirus in a wild animal being strongly associated with cancer as indicated by the very high odds ratio, but a cofactor, persistent organic pollutants exposure, also may increase the likelihood of cancer occurrence. Further work on the mechanisms behind the association of OtHV1 infection with sea lion carcinoma is indicated by this study.
In conclusion, this case control study, designed around rigorous opportunistic sampling over two decades, demonstrates the value of long-term epidemiological studies that control for multiple factors and use large sample sizes. There is increasing concern over the exposure of wild marine mammals to multiple factors that could have cumulative impacts on health, yet studies to investigate these impacts are logistically and ethically challenging in ocean environments (National Academies of Sciences Engineering, and Medicine, 2016). This study demonstrates that two factors, herpesvirus infection and contaminant exposure, are positively associated with urogenital cancer, and future studies should focus on mechanistic experiments to explore tumorogenic pathways. This study has implications for human health, as virally associated cancer occurs in humans, and likelihood of cancer development could similarly be increased by exposure to environmental contaminants. Efforts to prevent ecosystem contamination with persistent organic pollutants must be improved to protect both wildlife and human health.
The research data underpinning this publication can be accessed at https://doi.org/10.17630/5e96ccde-3814-47ed-960b-6e8991da68df at the University of St. Andrews, Scotland, United Kingdom. The dataset is identified as Pure ID: 267128461; Title: Cancer in California Sea Lions (dataset).
The animal study was reviewed and approved by the Marine Mammal Center IACUC.
FG, TR, LL, KC, KA, GY, and AH conceived and designed the study. FG, PD, and BH conducted post-mortem examinations of animals and collected samples and archived samples. TN managed the data. KC performed the histological review. AD, GY, and KA conducted the laboratory analyses. AH carried out statistical analyses. FG, AH, KC, and AD drafted the manuscript. All authors contributed to the final manuscript.
This work was funded by the Geoffrey Hughes Fellowship, the National Institutes of Health (Fogarty International Center) and National Science Foundation joint program for the Ecology of Infectious Disease, the National Marine Fisheries Service Marine Mammal Heath and Stranding Program, and the Natural Environment Research Council grant number NE/R015007/. These funders did not design, analyze and report results of the study.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank Lauren Rust, Liz Wheeler, Denise Greig, Jennifer Soper, Carlos Rios and Christine Fontaine for collection and archiving of samples and data, all the staff and volunteers of the Marine Mammal Center for care of the animals and gross post mortem examinations, Helen Browning for herpesvirus testing and Debbie Russell for statistical advice. We appreciated the chemical contaminant analyses provided by our colleagues in the Environmental Chemistry Program at the Northwest Fisheries Science Center. We also thank the reviewers for helpful comments that improved the manuscript.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2020.602565/full#supplementary-material
Acevedo-Whitehouse, K., Gulland, F., Greig, D., and Amos, W. (2003). Inbreeding: disease susceptibility in California sea lions. Nature 422:35. doi: 10.1038/422035a
Aragon, T. J. (2018). epitools: Epidemiology Tools. R Package Version 0.5-10. Available online at: https://CRAN.R-project.org/package=epitools (accessed May 12, 2020).
Bowen, E., Aldridge, B. M., Delong, R., Melin, S., Buckles, E. L., Gulland, F. M. D., et al. (2005). An immunogenetic basis for the high prevalence of urogenital cancer in a free-ranging population of California sea lions (Zalophus californianus). Immunogenetics 56, 846–848. doi: 10.1007/s00251-004-0757-z
Browning, H. (2014). The Multi-Factorial Aetiology of Urogenital Carcinoma in the California Sea Lion (Zalophus californianus) – A Case-Control Study. Ph.D thesis, University of St Andrews, St Andrews.
Browning, H. M., Acevedo-Whitehouse, K., Gulland, F. M. D., Hall, A. J., Finlayson, J., Dagleish, M. P., et al. (2014). Evidence for a genetic basis of urogenital carcinoma in the wild California sea lion. Proc. R. Soc. Ser. B 281:20140240. doi: 10.1098/rspb.2014.0240
Browning, H. M., Gulland, F. M. D., Hammond, J. A., Colegrove, K. M., and Hall, A. J. (2015). Common cancer in a wild animal: the California sea lion (Zalophus californianus) as an emerging model for carcinogenesis. Philos. Trans. R. Soc. Lond. B 370:0140228.
Buckles, E. L., Lowenstine, L. J., DeLong, R. L., Melin, S. R., Vittore, R. K., Wong, H.-N., et al. (2007). Age-prevalence of otarine herpesvirus-1, a tumor-associated virus, and possibility of its sexual transmission in California sea lions. Vet. Microbiol. 120, 1–8. doi: 10.1016/j.vetmic.2006.10.002
Buckles, E. L., Lowenstine, L. J., Funke, C., Vittore, R. K., Wong, H.-N., St Leger, J. A., et al. (2006). Otarine herpesvirus-1, not papillomavirus, is associated with endemic tumours in California sea lions (Zalophus californianus). J. Comp. Pathol. 135, 183–189. doi: 10.1016/j.jcpa.2006.06.007
Burnham, K. P., and Anderson, D. R. (2004). Multimodel inference: understanding AIC and BIC in model selection. Soc. Method Res. 33, 261–304. doi: 10.1177/0049124104268644
Carretta, J. V., Forney, K. A., Oleson, E. M., Weller, D. W., Lang, A. R., Baker, J. D., et al. (2018). U.S. Pacific Marine Mammal Stock Assessments: NOAA Technical Memorandum NMFS-SWFSC-617. Seattle, WA: United States Department of Commerce.
Cohn, B. A., La Merrill, M., Krigbaum, N. Y., Yeh, G., Park, J.-S., Zimmermann, L., et al. (2015). DDT exposure in utero and breast cancer. J. Clin. Endocr. Metab. 100, 2865–2872.
Colborn, T., Vom Saal, F. S., and Sole, A. M. (1993). Developmental effects of endocrine-disrupting chemicals in wildlife and humans. Environ. Health Perspect. 101, 378–384. doi: 10.1289/ehp.93101378
Colegrove, K. M., Gulland, F. M. D., Naydan, D. K., and Lowenstine, L. J. (2009). Tumor morphology and immunohistochemical expression of estrogen receptor, progesterone receptor, p53, and Ki67 in urogenital carcinomas of California sea lions (Zalophus californianus). Vet. Pathol. 46, 642–655. doi: 10.1354/vp.08-vp-0214-c-fl
De Guise, S., Martineau, D., Béland, P., and Fournier, M. (1995). Possible mechanisms of action of environmental contaminants on St. Lawrence beluga whales (Delphinapterus leucas). Environ. Health Perspect. 103(Suppl. 4) 73–77. doi: 10.1289/ehp.95103s473
DeLong, R. L., Melin, S. R., Laake, J. L., Morris, P., Orr, A., and Harris, J. D. (2017). Age- and sex-specific survival of California sea lions (Zalophus californianus) at San Miguel Island, California. Mar. Mamm. Sci. 33, 1097–1125. doi: 10.1111/mms.12427
Deming, A. (2018). The Role of Otarine Herpesvirus 1 in California Sea Lion (Zalophus californianus) Urogenital Carcinoma. 45–48. Ph.D thesis, University of Florida, Gainesville, FL.
Deming, A. C., Colegrove, K., Duignan, P. J., Hall, A. J., Wellehan, F. X., and Gulland, F. M. D. (2018). Prevalence of urogenital carcinoma in stranded California sea lions (Zalophus californianus) from 2005-2015. J. Wildl. Dis. 54, 581–586. doi: 10.7589/2017-08-208
Estep, R. D., and Wong, S. W. (2013). Rhesus macaque rhadinovirus-associated disease. Curr. Opin. Virol. 3, 245–250. doi: 10.1016/j.coviro.2013.05.016
Ghaffar, M., Li, J., Zhang, L., Khodahemmati, S., Wang, M., Wang, J., et al. (2018). Water carcinogenicity and prevalence of HPV infection in esophageal cancer patients in Huaihe River Basin, China. Gastroenterol. Res. Pract. 2018:2028986. doi: 10.1155/2018/2028986
Greig, D. J., Gulland, F. M. D., and Kreuder, C. (2005). A decade of live California sea lion (Zalophus californianus) strandings along the Central California coast: causes and trends, 1991-2000. Aquat. Mamm. 31, 11–22. doi: 10.1578/am.31.1.2005.11
Greig, D. J., Ylitalo, G. M., Hall, A. J., Fauquier, D. A., and Gulland, F. M. D. (2007). Transplacental transfer of organochlorines in California sea lions (Zalophus californianus). Environ. Toxicol. Chem. 26, 37–44. doi: 10.1897/05-609r.1
Gulland, F. M., Trupkiewicz, J. G., Spraker, T. R., and Lowenstine, L. J. (1996). Metastatic carcinoma of probable transitional cell origin in 66 free-living California sea lions (Zalophus californianus), 1979 to 1994. J. Wildl. Dis. 32, 250–258. doi: 10.7589/0090-3558-32.2.250
Hall, A. J., Gulland, F. M. D., Ylitalo, G. M., Greig, D., and Lowenstine, L. J. (2008). Changes in blubber contaminant concentrations in California sea lions (Zalophus californianus) associated with weight loss and gain during rehabilitation. Environ. Sci. Technol. 42, 4181–4187. doi: 10.1021/es702685p
Hunt, R. D., Meléndez, L. V., García, F. G., and Trum, B. F. (1972). Pathologic features of Herpesvirus ateles lymphoma in cotton-topped marmosets (Saguinus oedipus). J. Natl. Cancer Inst. 49, 1631–1639. doi: 10.1093/jnci/49.6.1631
King, D. P., Hure, M. C., Goldstein, T., Aldridge, B. M., Gulland, F. M. D., Saliki, J. T., et al. (2002). Otarine herpesvirus-1: a novel gammaherpesvirus associated with urogenital carcinoma in California sea lions (Zalophus californianus). Vet. Microbiol. 86, 131–137. doi: 10.1016/s0378-1135(01)00497-7
Laake, J. L., Lowry, M. S., DeLong, R. L., Melin, S. R., and Carretta, J. V. (2018). Population growth and status of California sea lions. J. Wildl. Manage. 82, 583–595. doi: 10.1002/jwmg.21405
Laake, J. L., Melin, S., Greig, D., Prager, K., Delong, R., et al. (2016). California Sea Lion Sex-and Age-Specific Morphometry. NOAA Technical Memorandum NMFS-AFSC-312. Seattle, WA: United States Department of Commerce. doi: 10.7289/V5/TM-AFSC-312
Lauby-Secretan, B., Loomis, D., Grosse, Y., Ghissassi, F. E., Bouvard, V., Benbrahim-Tallaa, L., et al. (2013). Carcinogenicity of polychlorinated biphenyls and polybrominated biphenyls. Lancet Oncol. 14, 287–288.
Leathlobhair, M. N., Gulland, F. M. D., and Murchison, E. P. (2017). No evidence for clonal transmission of urogenital carcinoma in California sea lions (Zalophus californianus). Res. One 2:46. doi: 10.12688/wellcomeopenres.11483.1
Levin, M., De Guise, S., and Ross, P. S. (2005). Association between lymphocyte proliferation and polychlorinated biphenyls in free-ranging harbor seal (Phoca vitulina) pups from British Columbia, Canada. Environ. Toxicol. Chem. 24, 1247–1252. doi: 10.1897/04-206r.1
Lipscomb, T. P., Scott, D. P., Garber, R. L., Krafft, A. E., Tsai, M. M., Lichy, J. H., et al. (2005). Common metastatic carcinoma of California sea lions (Zalophus californianus): evidence of genital origin and association with novel gammaherpesvirus. Vet. Pathol. 37, 609–617. doi: 10.1354/vp.37-6-609
Longnecker, M. P., Rogan, W. J., and Lucier, G. (1997). The human health effects of DDT (dichlorodiphenyl−trichloroethane) and PCBs (polychlorinated biphenyls) and an overview of organochlorines in public health. Annu. Rev. Publ. Health 18, 211–244. doi: 10.1146/annurev.publhealth.18.1.211
Martineau, D., Lemberger, K., Dallaire, A., Labelle, P., Lipscomb, T. P., Michel, P., et al. (2002). Cancer in wildlife, a case study: beluga from the St. Lawrence Estuary, Quebec, Canada. Environ. Health Perspect. 110, 285–292. doi: 10.1289/ehp.02110285
Martinez, J. M., Stephens, L. C., and Jones, L. A. (2005). Long-term effects of neonatal exposure to hydroxylated polychlorinated biphenyls in the BALB/cCrgl mouse. Environ. Health Perspect. 113, 1022–1026.
Mathur, V., John, P. J., Soni, I., and Bhatnagar, P. (2008). “Blood levels of organochlorine pesticide residues and risk of reproductive tract cancer among women from Jaipur, India,” in Hormonal Carcinogenesis V. Advances in Experimental Medicine and Biology, Vol. 617, eds J. J. Li, S. A. Li, S. Mohla, H. Rochefort, and T. Maudelonde (New York, NY: Springer). doi: 10.1007/978-0-387-69080-3_37
McCarthy, S. G., Incardona, J. P., and Scholz, N. L. (2008). Coastal storms, toxic runoff and the sustainable conservation of fish and fisheries. Am. Fish. Soc. Symp. 64, 7–27.
National Academies of Sciences Engineering, and Medicine (2016). Approaches to Understanding the Cumulative Effects of Stressors on Marine Mammals. Washington, DC: The National Academies Press. doi: 10.17226/23479
Nik-Zainal, S., and Hall, B. A. (2019). Cellular survival over genetic perfection. Science 366, 802–803.
Patnode, K. A., and Curtis, L. R. (1994). 2,29,4,49,5,59- and 3,39,4,49,5,59-hexachlorobiphenyl alteration of uterine progesterone and estrogen receptors coincides with embryotoxicity in mink (Mustela vision). Toxicol. Appl. Pharmacol. 127, 9–18.
Pearse, A., and Swift, K. (2006). Transmission of devil facial-tumour disease. Nature 439:549. doi: 10.1038/439549a
Peñín, I., Levin, M., Acevedo-Whitehouse, K., Jasperse, L., Gebhard, E., Gulland, F. M. D., et al. (2018). Effects of polychlorinated biphenyls (PCB) on California sea lion (Zalophus californianus) lymphocyte functions upon in vitro exposure. Environ. Res. 167, 708–717. doi: 10.1016/j.envres.2018.08.028
Peterson, S. H., Hassrick, J. L., Lafontaine, A., Thome’, J.-P., Crocker, D. E., Debier, C., et al. (2014). Effects of age, adipose percent, and reproduction on PCB concentrations and profiles in an extreme fasting North Pacific marine mammal. PLoS One 9:e96191. doi: 10.1371/journal.pone.0096191
R Core Team (2020). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Randhawa, N., Gulland, F. M. D., Ylitalo, G. M., DeLong, R., and Mazet, J. A. K. (2015). Sentinel California sea lions provide insight into legacy organochlorine exposure trends and their association with cancer and infectious disease. One Health 1, 37–43.
Robertson, L. W., and Hansen, L. (eds) (2020). PCBs: Recent Advances in Environmental Toxicology and Health Effects: Medicine and Health Sciences, 2 Edn. Lexington, KY: University of Kentucky Press, 8.
Ross, P. S., De Swart, R. L., Reijnders, P. J., Van Loveren, H., Vos, J. G., and Osterhaus, A. D. M. E. (1995). Contaminant-related suppression of delayed-type hypersensitivity and antibody responses in harbor seals fed herring from the Baltic Sea. Environ. Health Perspect. 103, 162–167.
Schwacke, L., Viot, E. O., Hansen, L., Wells, R., Mitchum, G. B., Hohn, A., et al. (2002). Probabilistic risk assessment of reproductive effects of polychlorinated biphenyls on bottlenose dolphins (Tursiops truncatus) from the southeast United States coast. Environ. Toxicol. Chem. 21, 2752–2764.
Sloan, C. A., Anulacion, B. F., Baugh, K. A., Bolton, J. L., Boyd, D., Boyer, R. H., et al. (2014). Northwest Fisheries Science Center’s Analyses of Tissue, Sediment, and Water Samples for Organic Contaminants by Gas Chromatography/Mass Spectrometry and Analyses of Tissue for Lipid Classes by Thin Layer Chromatography/Flame Ionization Detection. NOAA Technical Memorandum NMFS-NWFSC-125. Seattle, WA: United States Department of Commerce.
Sloan, C. A., Anulacion, B. F., Baugh, K. A., Bolton, J. L., Boyd, D., Chittaro, P. M., et al. (2019). Quality Assurance Plan for Analyses of Environmental Samples for Polycyclic Aromatic Hydrocarbons, Persistent Organic Pollutants, Dioctyl Sulfosuccinate, Estrogenic Compounds, Steroids, Hydroxylated Polycyclic Aromatic Hydrocarbons, Stable Isotope Ratios, and Lipid Classes. NOAA Technical Memorandum NMFS-NWFSC-147. Seattle, WA: United States Department of Commerce. doi: 10.25923/kf28-n618
Touitou, R., Hickabottom, M., Parker, G., Crook, T., and Allday, M. J. (2001). Physical and functional interactions between the corepressor CtBP and the Epstein-Barr virus nuclear antigen EBNA3C. J. Virol. 75, 7749–7755.
Verloop, J., van Leeuwen, F. E., Helmerhorst, T. J., van Boven, H. H., and Rookus, M. A. (2010). Cancer risk in DES daughters. Cancer Causes Control 21, 999–1007.
Williams, R., ten Doeschate, M., Curnick, D. J., Brownlow, A., Barber, J. L., Davison, N. J., et al. (2020). Levels of polychlorinated biphenyls are still associated with toxic effects in harbor porpoises (Phocoena phocoena) despite having fallen below proposed toxicity thresholds. Environ. Sci. Technol. 54, 2277–2286.
Wolff, M. S., Toniolo, P. G., Lee, E. W., Rivera, M., and Dubin, N. (1993). Blood levels of organochlorine residues and risk of breast cancer. J. Natl. Cancer Inst. 85, 648–652.
Ylitalo, G. M., Buzitis, J., Boyd, D. T., Herman, D. P., Tilbury, K. L., and Krahn, M. M. (2005a). “Improvements to high-performance liquid chromatography/photodiode array detection (HPLC/PDA) method that measures dioxin-like polychlorinated biphenyls and other selected organochlorines in marine biota,” in Techniques in Aquatic Toxicology, Vol. 2, ed. G. K. Ostrander (Boca Raton, FL: CRC Press), 449–464.
Ylitalo, G. M., Stein, J. E., Hom, T. E., Johnson, L. J., Tilbury, K. L., Hall, A. J., et al. (2005b). The role of organochlorines in cancer-associated mortality in California sea lions (Zalophus californianus). Mar. Pollut. Bull. 50, 30–39.
Ylitalo, G. M., Yanagida, G. K., Hufnagle, L. Jr., and Krahn, M. M. (2005c). “Determination of lipid classes and lipid content in tissues of aquatic organisms using a thin layer chromatography/flame ionization detection (TLC/FID) microlipid method,” in Techniques in Aquatic Toxicology, Vol. 2, ed. G. K. Ostrander (Boca Raton, FL: CRC Press), 227–238.
Keywords: cancer, organochlorines, sea lions, herpesvirus, microsatellite, case control study
Citation: Gulland FMD, Hall AJ, Ylitalo GM, Colegrove KM, Norris T, Duignan PJ, Halaska B, Acevedo Whitehouse K, Lowenstine LJ, Deming AC and Rowles TK (2020) Persistent Contaminants and Herpesvirus OtHV1 Are Positively Associated With Cancer in Wild California Sea Lions (Zalophus californianus). Front. Mar. Sci. 7:602565. doi: 10.3389/fmars.2020.602565
Received: 03 September 2020; Accepted: 20 November 2020;
Published: 10 December 2020.
Edited by:
Alvin Cyril Camus, University of Georgia, United StatesReviewed by:
Gail Schofield, Queen Mary University of London, United KingdomCopyright © 2020 Gulland, Hall, Ylitalo, Colegrove, Norris, Duignan, Halaska, Acevedo Whitehouse, Lowenstine, Deming and Rowles. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Frances M. D. Gulland, ZnJhbmNlc2d1bGxhbmRAZ21haWwuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.