
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Mar. Sci. , 08 September 2020
Sec. Global Change and the Future Ocean
Volume 7 - 2020 | https://doi.org/10.3389/fmars.2020.542563
This article is part of the Research Topic Biogeochemical Consequences of Climate-Driven Changes in the Arctic View all 11 articles
 Maeve McGovern1,2,3*
Maeve McGovern1,2,3* Alexey K. Pavlov4,5
Alexey K. Pavlov4,5 Anne Deininger1,6
Anne Deininger1,6 Mats A. Granskog7
Mats A. Granskog7 Eva Leu4
Eva Leu4 Janne E. Søreide3
Janne E. Søreide3 Amanda E. Poste1*
Amanda E. Poste1*Climate-change driven increases in temperature and precipitation are leading to increased discharge of freshwater and terrestrial material to Arctic coastal ecosystems. These inputs bring sediments, nutrients and organic matter (OM) across the land-ocean interface with a range of implications for coastal ecosystems and biogeochemical cycling. To investigate responses to terrestrial inputs, physicochemical conditions were characterized in a river- and glacier-influenced Arctic fjord system (Isfjorden, Svalbard) from May to August in 2018 and 2019. Our observations revealed a pervasive freshwater footprint in the inner fjord arms, the geochemical properties of which varied spatially and seasonally as the melt season progressed. In June, during the spring freshet, rivers were a source of dissolved organic carbon (DOC; with concentrations up to 1410 μmol L–1). In August, permafrost and glacial-fed meltwater was a source of inorganic nutrients including NO2 + NO3, with concentrations 12-fold higher in the rivers than in the fjord. While marine OM dominated in May following the spring phytoplankton bloom, terrestrial OM was present throughout Isfjorden in June and August. Results suggest that enhanced land-ocean connectivity could lead to profound changes in the biogeochemistry and ecology of Svalbard fjords. Given the anticipated warming and associated increases in precipitation, permafrost thaw and freshwater discharge, our results highlight the need for more detailed seasonal field sampling in small Arctic catchments and receiving aquatic systems.
Recent climate change driven increases in air temperature and precipitation are changing the timing, magnitude and geochemical nature of freshwater runoff with unknown implications for Arctic coastal waters. The observed changes in climate have been distinct in the high-Arctic Svalbard archipelago (e.g., Adakudlu et al., 2019; van Pelt et al., 2019) where marine and land-terminating glaciers are shrinking in size (van Pelt et al., 2019) and where the upper layer of permafrost, where large amounts of organic carbon are stored (Tarnocai et al., 2009) is warming (Grosse et al., 2016; Biskaborn et al., 2019), and active layer depth is increasing (Christiansen et al., 2005). Together with increased precipitation and freshwater discharge (Peterson et al., 2002; McClelland et al., 2006; Adakudlu et al., 2019), the thawing terrestrial cryosphere is expected to lead to the mobilization and transport of dissolved and particulate organic and inorganic matter from Arctic watersheds to coastal waters (Parmentier et al., 2017).
In central Svalbard, snowmelt typically occurs in June (van Pelt et al., 2016) alongside high river discharge (Hodson et al., 2016). The permafrost active layer is deepest in August (Christiansen et al., 2005), a typically low discharge period (Hodson et al., 2016) when glacial-meltwater has higher residence time in the catchment. Seasonal changes in catchment hydrology have implications for the transport and bioavailability of carbon and nutrients in glacial meltwater on Svalbard (Nowak and Hodson, 2015; Koziol et al., 2019) and elsewhere in the Arctic (Neff et al., 2006; Holmes et al., 2008; Spencer et al., 2008). For example, carbon delivered during spring freshet in Alaskan rivers is more labile compared to aged, microbially reworked carbon delivered later in the summer (Holmes et al., 2008). While seasonal changes in river physicochemistry have been well documented for the Great Arctic rivers (e.g., Holmes et al., 2011), seasonal data from small Arctic catchments are scarce, making it difficult to assess potential impacts on receiving near-shore and coastal waters.
Arctic fjord estuaries are biogeochemical hotspots for the cycling of organic matter (OM) (Bianchi et al., 2020) and burial of carbon (Smith et al., 2015; Bianchi et al., 2018). The fate of terrestrial materials in the marine system is linked to physical and biological processes in the water column. Flocculation and sedimentation at the land-ocean interface (Meslard et al., 2018), and photodegradation and mineralization can act to remove OM from the water column while uptake by coastal biota can integrate terrestrial OM into the marine food-web (Parsons et al., 1989; Harris et al., 2018). Turbid freshwater plumes can also stratify the water column and inhibit nutrient-rich deep water renewal (Torsvik et al., 2019), while also rapidly attenuating light critical for photosynthesis (Murray et al., 2015; Holinde and Zielinski, 2016; Pavlov et al., 2019), with implications for the autotrophic: heterotrophic balance in nearshore areas (Wikner and Andersson, 2012). Despite the rapid warming documented in the high Arctic (IPCC, 2014; Adakudlu et al., 2019), little is known regarding how these changes will affect the quantity and quality of materials transported to and through near-shore, fjord and coastal systems and thus their potential impacts on local and regional biogeochemical cycles (Parmentier et al., 2017).
To address these knowledge gaps, we studied the impacts of inputs from marine terminating glaciers and rivers on light, stratification, nutrient and OM dynamics in Isfjorden (Svalbard). To evaluate seasonal changes in runoff and associated impacts (snow melt vs. glacial melt/permafrost erosion), we targeted three stages of the melt season (1) pre-freshet in May, (2) spring freshet in June, and (3) late-summer runoff in August. Specifically, we aimed to identify the spatial and seasonal response in fjord physicochemical conditions and OM characteristics and evaluate how these might change with the future projected changes in freshwater runoff on Svalbard.
Fieldwork took place in 2018 and 2019 in Isfjorden, the largest fjord system on the West coast of Spitsbergen, Svalbard (Figures 1a,b). Isfjorden exchanges waters with the west Spitsbergen shelf, where the West Spitsbergen Current (WSC) and the Spitsbergen Polar Current (SPC) bring Atlantic and Arctic waters, which enter the fjord along the southern shore and exit the fjord along the northern coastline (Nilsen et al., 2016; Figure 1c). Isfjorden has several fjord arms (e.g., Fraser et al., 2018). Tempelfjorden and Billefjorden and the northern side of Isfjorden have marine terminating glaciers, which are absent from the southern side of Isfjorden, including Adventfjorden (Figure 1b). Of the sampled fjord arms, only Billefjorden has a shallow sill (50 m) at the entrance, which typically inhibits water mass exchange with adjacent (or central) parts of Isfjorden (Nilsen et al., 2008). The intrusion of warm and saline Atlantic water from the WSC (Fraser et al., 2018) facilitates the melting of Svalbard glaciers (Luckmann et al., 2015). In turn, runoff from glaciers and rivers contribute to estuarine circulation in the fjord (Torsvik et al., 2019). The rivers sampled in this study have catchments ranging from (55–725 km2) in size with varying degrees of glacial cover (10–51%; pers. com. Guerrero, 2019).
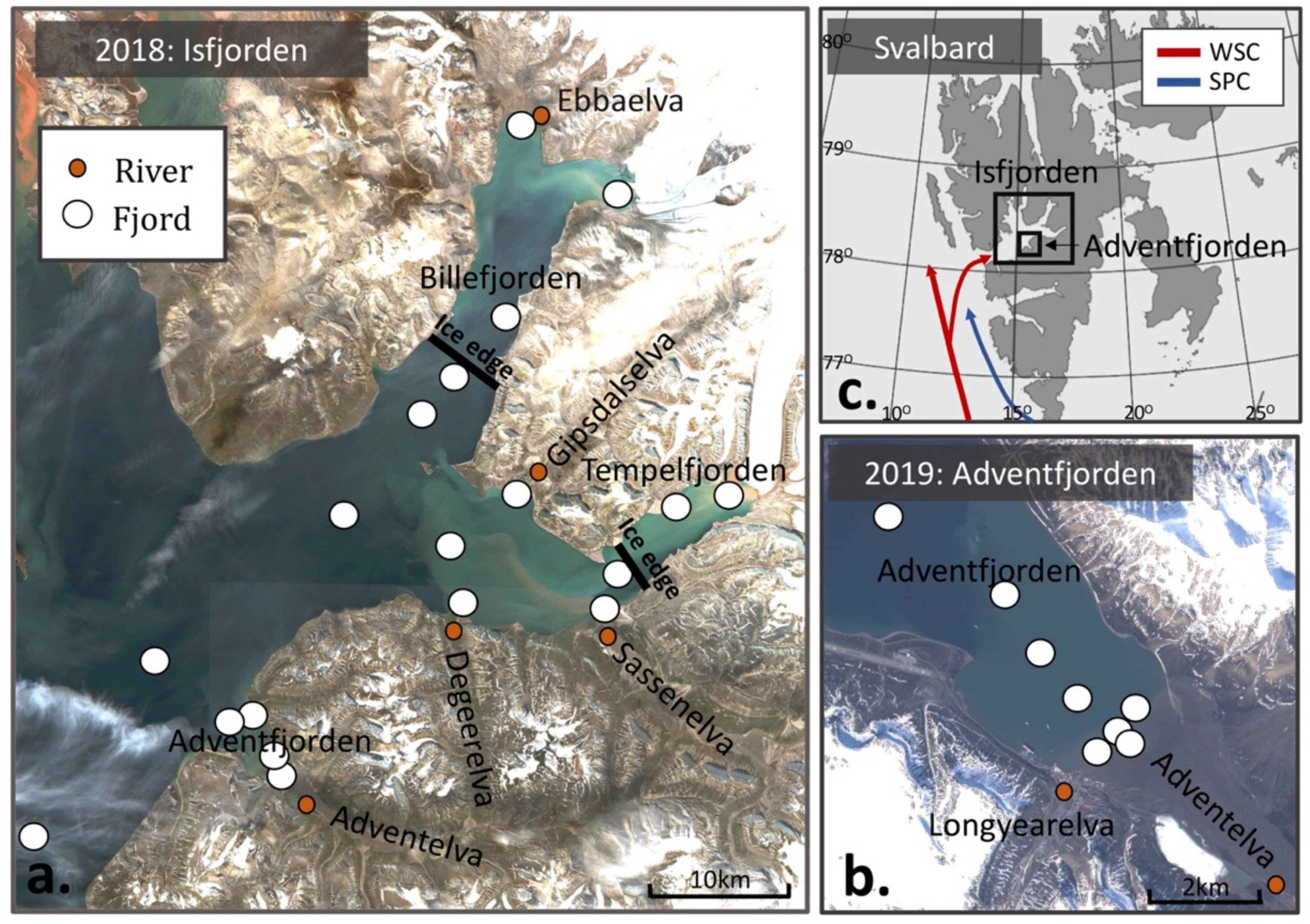
Figure 1. (a) Station map of Isfjorden (sampled in 2018), and (b) Adventfjorden (sampled in 2019), superimposed on satellite images taken from the same week as sampling [August 20, 2018 and June 14th, 2019; Sentinel-2 (https://scihub.copernicus.eu/)]. The location of the ice edge in May 2018, when land-fast ice covered the inner fjord arms, is indicated in black. (c) Map of Svalbard with the West Spitsbergen Current (WSC) and Spitsbergen Polar Current (SPC) depicted in red and blue respectively.
In 2018, samples were collected in May (10th–11th), June (18th–24th), and August (16th–24th), from a total of 17 different stations in Isfjorden along gradients from rivers and glaciers to the outer fjord (Figure 1a). The number of stations sampled each month varied due to presence of ice in May (Figure 1a), when additional fjord transect stations were sampled at the land-fast ice edge in the fjord arms, and where the innermost stations were not accessible. In 2019, the same sampling techniques were used in Adventfjorden with a higher spatial resolution, during June (15th–17th) and August (7th–9th; Figure 1b). Samples were collected from 8 stations in Adventfjorden as well as 2 rivers (Adventelva and Longyearelva). At each fjord station, water samples were collected from up to 5 depths (surface, 2 m, 5 m, 15 m, and 30 m) depending on station depth.
In both sampling years, a CTD profiler (SD204, SAIV A/S or Seabird SBE 911) was used to collect vertical profiles of salinity, temperature and chlorophyll fluorescence. Secchi depth was measured and light measurements were made using optical sensors. In 2018, a PAR cosine-corrected sensor was used to obtain vertical profiles of photosynthetically active radiation (PAR, 400–700 nm) while in 2019, TrioS Ramses ACC-VIS hyperspectral radiometers (one for profiling, one as a surface reference) were used to obtain downwelling planar irradiance profiles. At all stations, water was collected from the surface and 15 m using a Niskin bottle. At stations shallower than 17 m, water was collected from the surface and from 2 m above the bottom. A multiparameter sensor (Hanna instruments, HI 98195) and handheld turbidity meter (Thermo Scientific Eutech TN-100) were used in the field to record temperature, salinity, pH, conductivity and turbidity for each sample in a well-mixed bucket of sample water immediately after collection. Water was collected directly from the Niskin bottle into 20 liter jugs for further processing at the University Centre in Svalbard (UNIS).
Samples for analysis of dissolved organic carbon (DOC), dissolved nutrients [ammonium (NH4), phosphate (PO4), nitrite + nitrate (NO2 + NO3), and silica (SiO2)] were filtered through 0.2 μm polycarbonate membrane filters and preserved with 4M H2SO4 (final concentration of 1% by volume) in 100 mL pre-cleaned amber glass bottles (DOC) or 100 mL acid-washed HDPE bottles (dissolved nutrients). Samples were stored in the dark at 4°C until analysis. For characterization of chromophoric dissolved organic matter (cDOM), water was filtered through 0.2 um polycarbonate filters and stored in 100 mL amber glass bottles in the dark at 4°C. To determine the concentration of suspended particulate matter (SPM), water was filtered onto pre-combusted and pre-weighed glass fiber filters (Whatman GF/F, nominal pore size 0.7 μm). For particulate organic carbon (POC) and particulate nitrogen (PartN) and analysis of stable carbon and nitrogen isotopes (SIA), up to 1.5L of water was filtered onto pre-combusted 25 mm GF/F filters. Particulate phosphorus (PartP) and chlorophyll a (Chla) samples were filtered onto a non-combusted GF/F filters. All filters were stored frozen at −20°C until analysis.
Nutrient, DOC, and PartP analyses were carried out at the Norwegian Institute for Water Research (NIVA, Oslo, Norway) using standard and accredited methods (as described in Kaste et al., 2018). Filters for SPM were dried and reweighed to determine SPM concentrations. Chlorophyll a was determined fluorometrically on a Turner 10-AU fluorometer after methanol extraction (Parsons, 2013). Pheophytin was measured on the same samples following acidification with 3 drops of 1M HCl. Stable isotope analysis of particulate organic matter (POM) was carried out at the University of California, Davis (UC Davis Stable Isotope Facility, United States). For PartN, filters were dried and packed into tin capsules for analysis. For POC, filters were fumigated for 24–48 h in a desiccator with concentrated HCl to remove inorganic carbonates prior to encapsulation. δ13C, δ15N, as well as total C and N content were measured using an elemental analyzer interfaced to an isotope ratio mass spectrometer. Run-specific standard deviations at UC Davis were ± 0.09‰ for 13C and 0.05‰ for 15N in 2018 and ± 0.08‰ for 13C and 0.05‰ for 15N in 2019. Stable carbon and nitrogen isotope values are presented using delta notation, relative to international standards (Vienna PeeDee Belemnite for C, and atmospheric N for nitrogen) (Peterson and Fry, 1987). For analysis of cDOM properties (Table 1), absorbance was measured at 1 nm intervals across a wavelength range of 200–900 nm with a Perkin-Elmer Lambda 40P UV/VIS Spectrophotometer using a cuvette with a 5 cm path-length. Absorbance values were blank corrected (Milli-Q) and the average absorbance from 700–900 nm was subtracted from the spectra to correct for possible absorption offset (Helms et al., 2008). Values were converted to Naperian absorption coefficients by multiplying the raw absorbance values by 2.303 and dividing by the pathlength (m) (Hu et al., 2002). Spectral slopes (S) (Table 1), which serve as proxies for the composition and source of DOM, with steeper S275–295 and increasing slope ratio (SR; S275–295:S350–400) indicative of marine, low molecular weight OM (Helms et al., 2008), were calculated from the spectral absorption data. Meanwhile, specific UV absorbance at 254 nm (SUVA254), which is positively related to aromaticity of DOM (Weishaar et al., 2003), was calculated by dividing absorbance at 254 nm by the DOC concentration (Weishaar et al., 2003).

Table 1. Optical characteristics of cDOM based on absorption spectra.
Spectral irradiance obtained using TriOS Ramses ACC-VIS sensors in 2019 was integrated over the PAR range (400–700 nm). The diffuse attenuation coefficient Kd(PAR) (m-1) was calculated in the top 1 m using the following equation (Kirk, 2010), which assumes the exponential attenuation of light with depth (Beer’s Law):
where Ed(PAR, 0) and Ed(PAR, Z) represent the downwelling irradiance just below the surface and at depth Z, respectively.
The euphotic depth (Zeu) was calculated as 1% of surface values (just below the water surface) based on irradiance profiles. In cases when Zeu exceeded the station depth, light profiles were extrapolated using the best exponential fit to estimate Zeu.
Freshwater content (FWC) relative to a salinity of 34.7 in the top 10 m was calculated from CTD profiles at all stations using the following equation (Proshutinsky et al., 2009):
The reference salinity, Sref is taken as 34.7, which represents the boundary between surface waters and advected waters in Isfjorden (Nilsen et al., 2008). S is the water salinity at depth z. Change in FWC is a measure of how much liquid freshwater has accumulated or been lost from the ocean column bounded by the 34.7 isohaline. In this study, FWC in the surface layer is used as an indicator of degree of freshwater influence in Isfjorden. In addition, a difference in salinity (dS) between the surface and 10 m is used as a simple indicator of water column stratification at the time of sampling.
All statistical analyses were carried out using R (version 3.4.3, R Core Team, 2017). Temperature-Salinity (TS) diagrams were made using the PlotSvalbard package (Vihtakari, 2019). Water mass determinations were made based on Nilsen et al. (2008); Surface waters (SW) = Sal < 34, T > 1°C, intermediate waters (IW) = 34 < Sal < 34.7, T > 1°C, Atlantic waters (AW) = Sal > 34.9, T > 3°C), transformed Atlantic water (TAW) = Sal > 34.7, T > 1°C, Arctic water (ArW) = 34.4 < Sal < 34.8, −1.5 > T < 1°C, winter cooled water (WCW) = Sal > 34.74, T < −0.5°C) and local water (LW) = T < 1°C. For Table 2, discrete waters samples are grouped by fjord surface water (salinity < 34.7) and fjord advected water (salinity > 34.7). Spearman rank correlations were used to evaluate relationships between water chemistry parameters and salinity (Supplementary Figure S1).
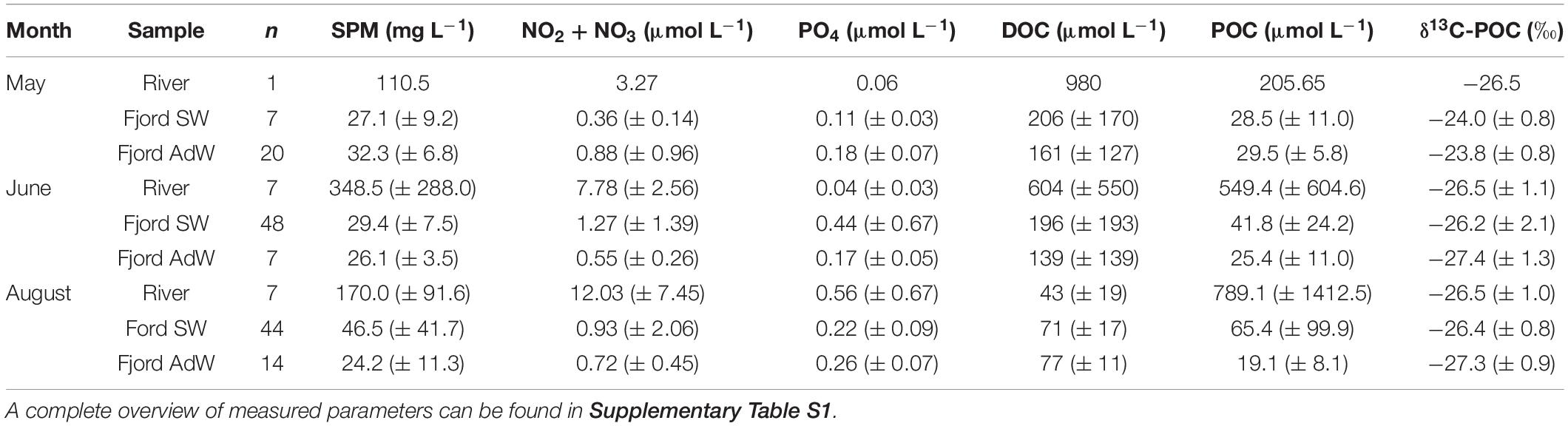
Table 2. Key water chemistry parameters (averages ± SD) of river water, fjord surface water (SW), and fjord advected water (AdW) samples from 2018 to 2019 for each month.
Redundancy analysis (RDA) was performed on scaled data using the vegan package (Oksanen et al., 2018) to test whether terrestrial inputs explain variation in water chemistry parameters as well as the source and quality of OM. Explanatory variables included salinity, turbidity, temperature and sampling month. To avoid overestimation of the explained variation, constraining variables were selected using forward model selection with a double-stopping criterion (Blanchet et al., 2008). For the water chemistry RDA, salinity, turbidity, temperature and sampling month were chosen via forward selection and all explained a significant amount of variation. For the organic matter RDA, turbidity was not significant, and instead salinity, temperature and sampling month were chosen for the RDA model.
In May, sampling took place when land-fast ice still covered much of inner Billefjorden and Tempelfjorden (Figure 1). Of the six rivers sampled in this study, only one (Adventelva) was running in May, and the water column at all sampling stations comprised of WCW and LW (Figure 2). In June and August, freshwater input from all of the rivers, as well as glacial melt and diffuse runoff along the coast, resulted in extensive freshening of surface waters in both years (Figure 2). This freshening was accompanied by increased stratification (based on dS) and fresh water content (FWC) between the surface and 10 m in June and August (Figures 3A,B). Riverine and glacial inputs delivered high concentrations of SPM to nearshore waters in Isfjorden (Figure 3D), resulting in turbid freshwater plumes associated with increased light attenuation, and thus a decreased depth of the euphotic zone (Zeu) in affected areas of the fjord (Figure 3C). Meanwhile, in the deeper waters, the intrusion of cold saline ArW and warmer saline TAW from the shelf was observed at the outer Isfjorden stations in June and August (Figure 2).
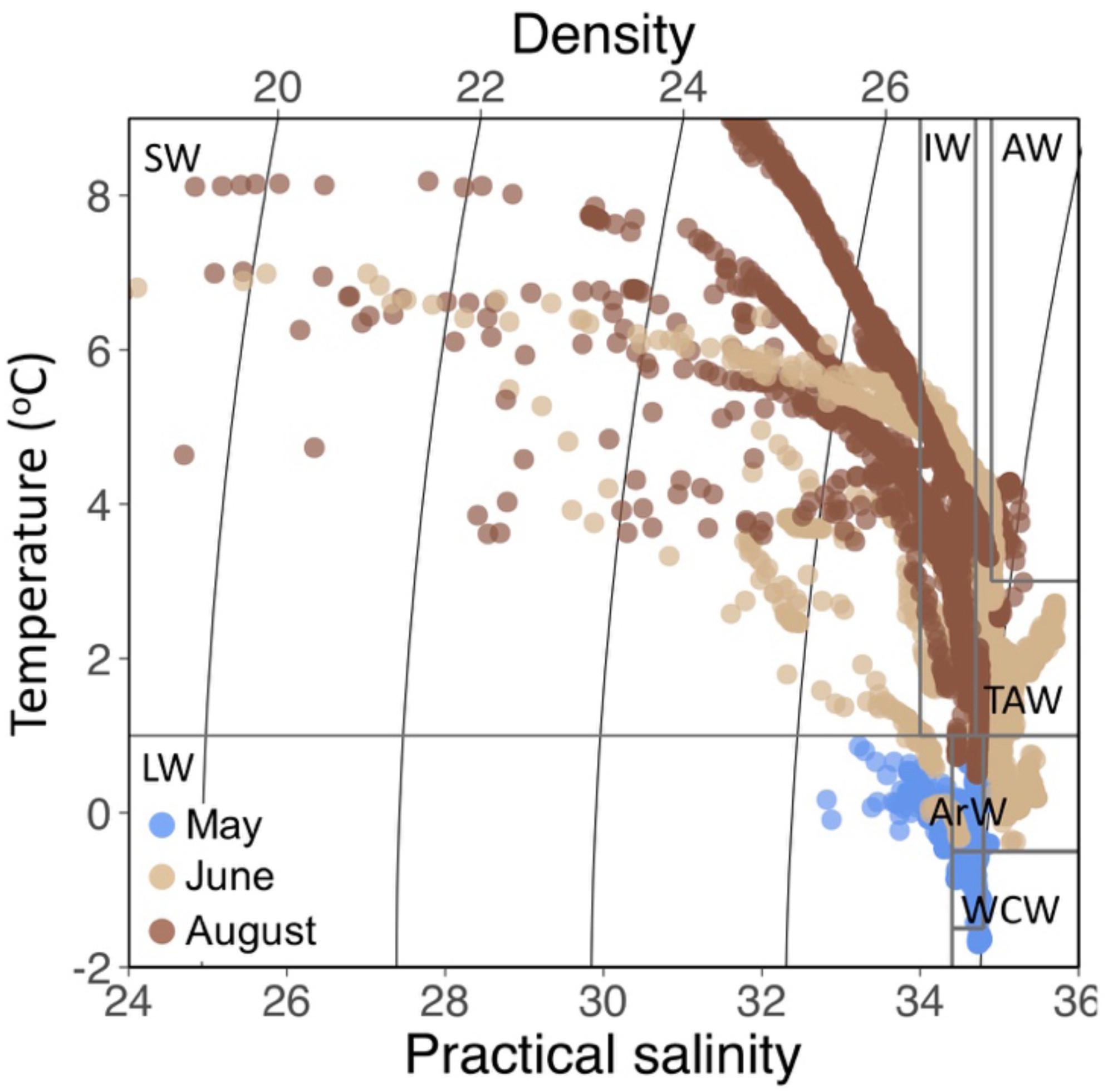
Figure 2. TS diagram based on all CTD profiles by sampling month. Water masses were determined using categories specific to Isfjorden (Nilsen et al., 2008).
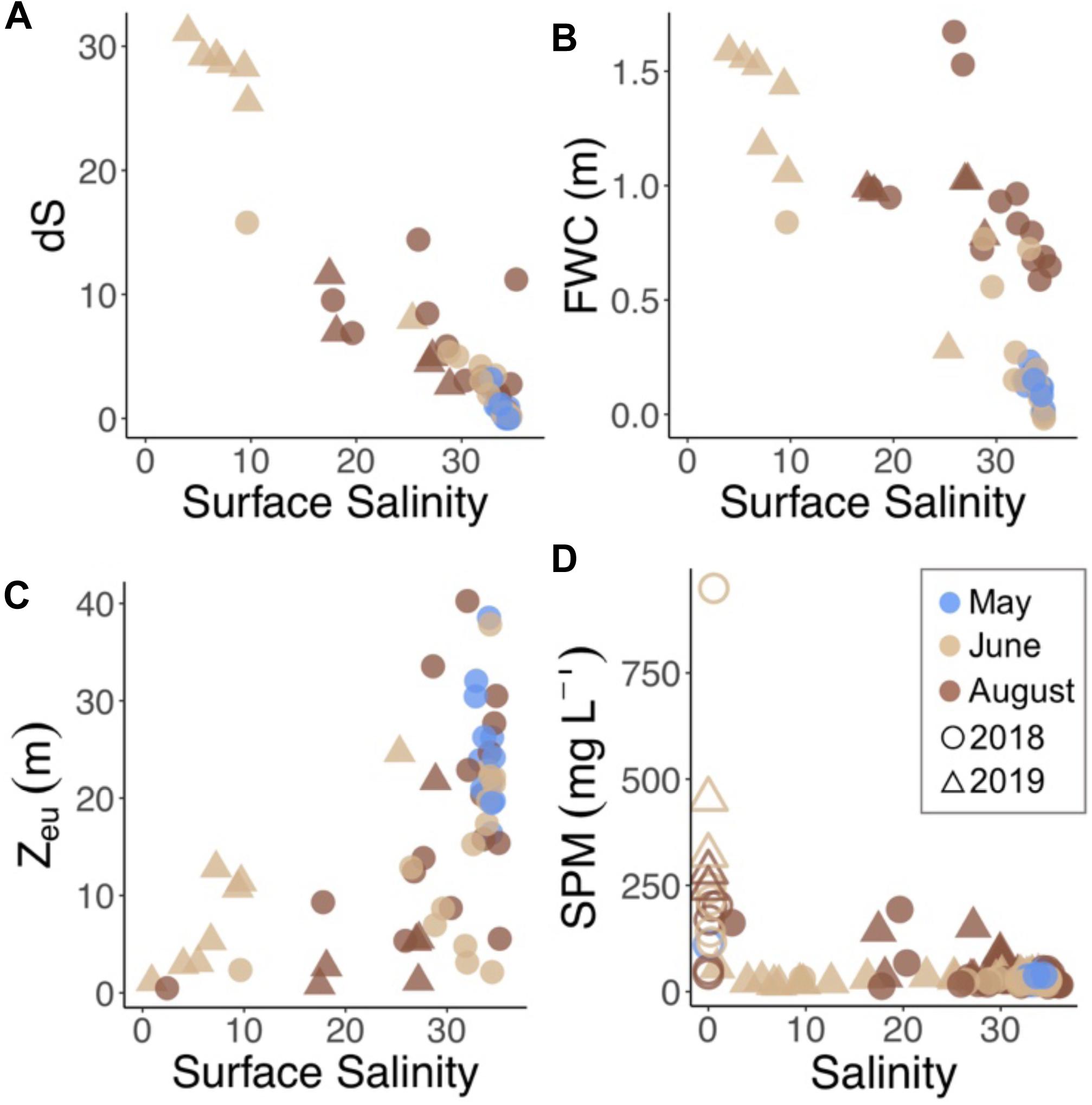
Figure 3. (A) Difference in salinity (dS) between surface and 10 m, (B) fresh water content (FWC) between surface and 10 m, (C) depth of euphotic zone (Zeu), and (D) suspended particulate matter (SPM) by year (symbol) and month (color). River samples are further distinguished by open symbols.
River samples had high concentrations of carbon early in the melt season (Figure 4A). In May, the DOC concentration in Adventelva was 980 μmol L–1. In June, DOC in Adventelva was much lower (40 μmol L–1) while the other rivers sampled had concentrations ranging from 670 to 1410 μmol L–1 (average 604 ± 550 μmol L–1; Table 2). All rivers had much lower concentrations of DOC in August, similar to those of Adventelva in June (range: 30–80 μmol L–1; average: 43 ± 19 μmol L–1). POC was also highly variable between rivers (Figure 4B) and was much higher than concentrations observed for advected water (Table 2). Results of δ13C-POC (Figure 4C) indicate that marine phytoplankton dominated the particulate matter pool in May during the spring phytoplankton bloom (δ13C: −23.9 ± 0.8‰). Meanwhile, terrestrial carbon dominated POC in June (δ13C: −26.4 ± 2.0‰) and August (δ13C: −26.6 ± 0.9‰) at all fjord sampling locations. CN ratios (Figure 4D) increased from May to June to August in both the rivers and the water column and decreased across the salinity gradient.
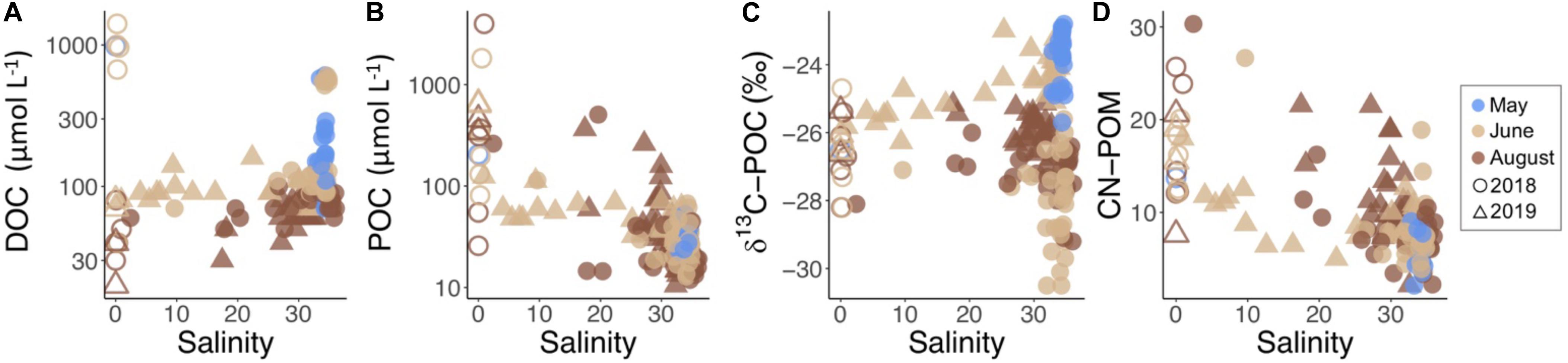
Figure 4. Concentrations of (A) DOC, (B) POC, and (C) δ13C-POC and (D) CN ratio of POM vs. salinity for each water sample by year (symbol) and month (color). River samples are further distinguished by open symbols.
Concentrations of NO2 + NO3 and SiO2 were highest in the river samples and decreased across the salinity gradient (Figure 5). These nutrients had high spatial and seasonal variability in Isfjorden with increasing concentrations from May to August at the near-shore stations (Figure 5). In May, 2018, sampling occurred during the end of the spring bloom. Concentrations of NO2 + NO3 in surface waters averaged 0.36 ± 0.14 μmol L–1 in SW, and 0.88 ± 0.96 μmol L–1 in AdW (Table 2). In June and August, nutrient concentrations were more strongly related to freshening when rivers and glaciers were a source of dissolved (Figure 5A) and particulate (Figure 5D) nitrogen (N) to Isfjorden. The partitioning of the N pool between particulate and dissolved phases also varied along the freshwater-marine gradient. In June and August, partN made up 60 ± 23 and 53 ± 28% of the total N pool in river samples and 30 ± 8 and 28 ± 14% in fjord SW and 23 ± 12 and 21 ± 12% of the total N pool in AdW respectively.
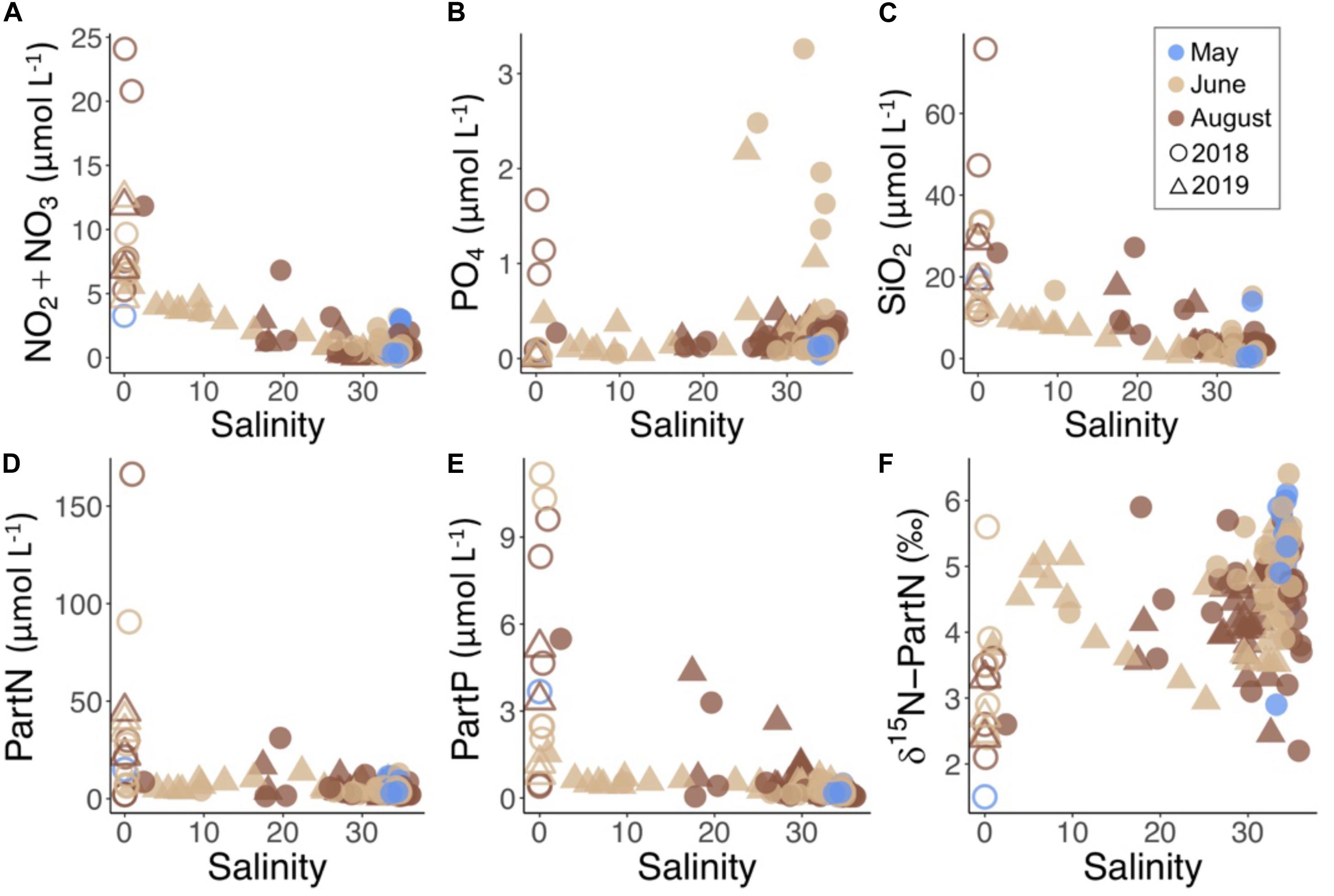
Figure 5. Concentrations of dissolved nutrients (A) NO2 + NO3, (B) PO4, (C) SiO2 and particulate nutrients (D) particulate nitrogen (PartN), (E) particulate phosphorus (PartP), and (F) δ15N-PartN vs. salinity for each water sample by year (symbol) and month (color). River samples are further distinguished by open symbols.
Rivers were also a source of phosphorus (P) in August (Figure 5B and Table 2). Mean concentrations of PO4 were 0.56 ± 0.67 μmol L–1 in river water samples, 0.22 ± 0.09 in fjord SW and 0.26 ± 0.07 μmol L–1 in AdW. Rivers had high concentrations of partP in both June and August, which were exponentially higher than concentrations in Fjord SW (Figure 5E). Similar to N, P concentrations were higher in the particulate fraction in rivers, but then partitioned more toward the dissolved phase across the salinity gradient. In June and August, partP made up 97 ± 3 and 70 ± 26% of total P in river samples and 38 ± 20 and 42 ± 23% in fjord SW and 19 ± 10 and 19 ± 11% of total P in AdW respectively.
Seasonal changes in DOM properties overwhelmed spatial differences within the fjord. In May, steep spectral slopes (S275–295) and high SR (Figure 6) indicated marine-derived, low molecular weight mDOM in the fjord. In both June and August, DOM properties in fjord waters were consistent between river and glacier-influenced parts of the fjord where low S275–295 values indicated the dominance of terrestrially derived OM (Figure 6). However, despite terrestrial OM dominating in both freshwater-influenced months, there was a distinct difference between tDOM in June and August, largely driven by differing concentrations of aCDOM(375) and slope ratio (SR). The higher levels of aCDOM(375) and S350–400 in June indicated that terrestrial cDOM dominated the DOM pool at all fjord stations. High concentrations of DOC in several high salinity samples in the outer fjord in June (Figure 4A) were accompanied by low values of δ13C (Figure 4D), high S350–400 (Figure 6D), and low SR (Figure 6E) values similar to river samples (Figure 6F). Meanwhile, in August, DOM properties reflected a terrestrial (low S275–295), aromatic (high SUVA254) source of DOM, which was of low molecular weight (high SR; Figure 6E) across all fjord stations.
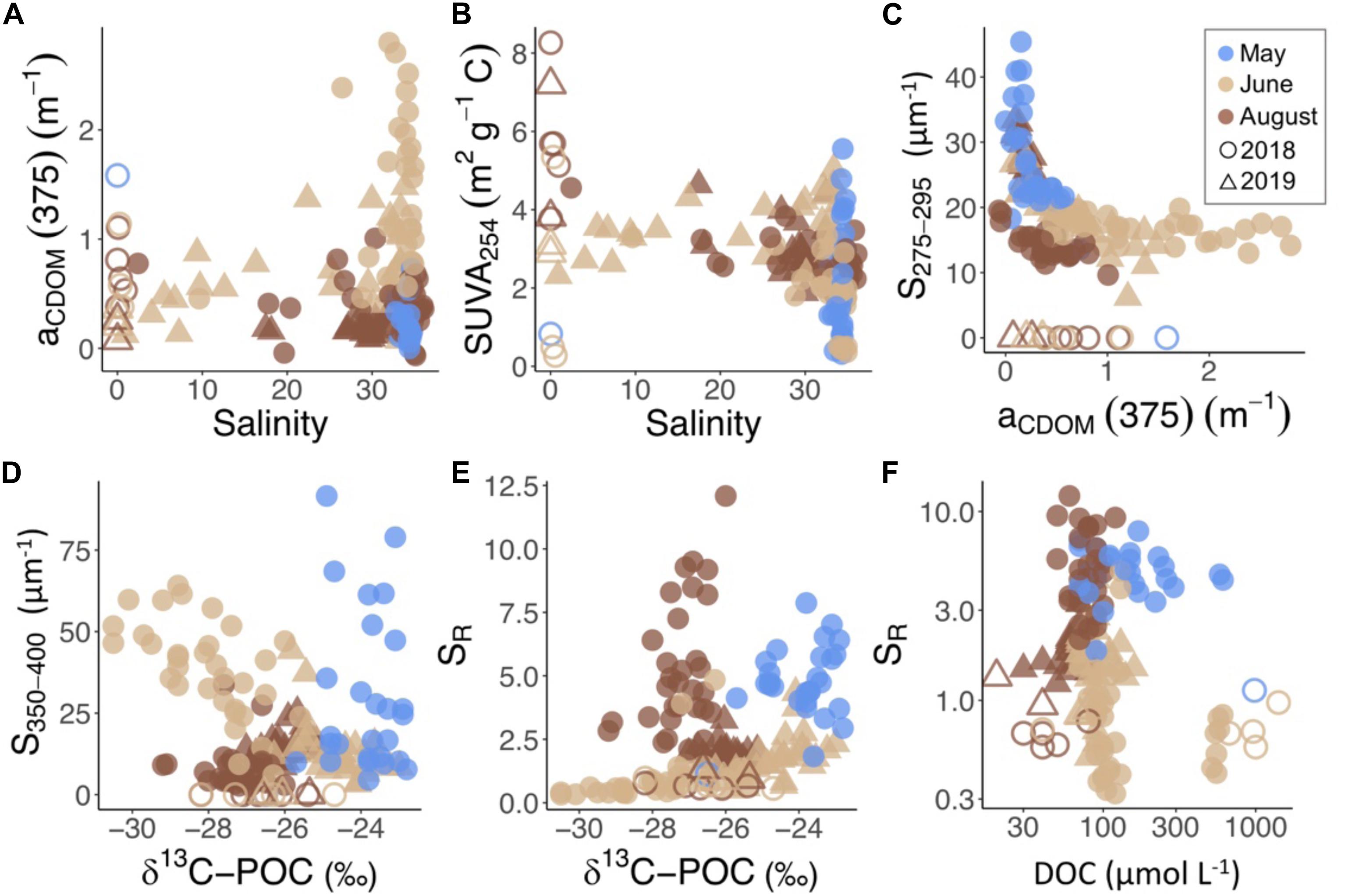
Figure 6. cDOM absorption characteristics including (A) aCDOM(375) and (B) SUVA254 vs. salinity, (C) S275–295 vs. aCDOM(375) and (D) S350–400 and (E) slope ratio (SR) vs. δ13C-POC and (F) SR vs. DOC for all water samples by year (symbol) and month (color). River samples are further distinguished by open symbols.
Results of redundancy analysis illustrated the importance of salinity, turbidity, sampling month and temperature in explaining variation in water chemistry parameters and sampling month, temperature and salinity for explaining variation in OM source and quality in both sampling years (Figure 7). Of the constraining variables, salinity and turbidity explained 31% of the total variation in the water chemistry parameters while sampling month explained the greatest amount of variation in the OM dataset (19% of the total variation; Figure 7).
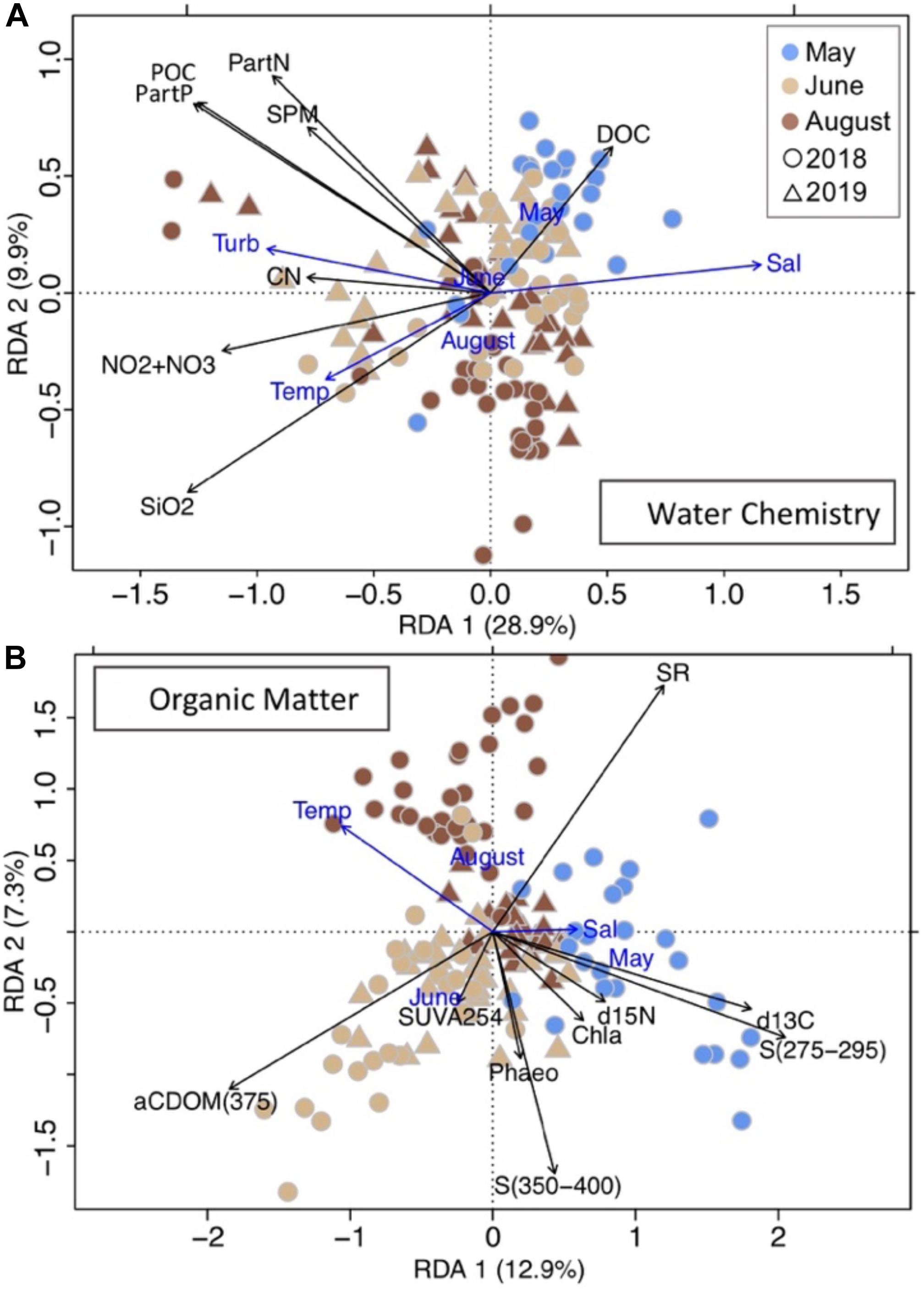
Figure 7. Redundancy analyses (RDA) of (A) water chemistry parameters constrained by salinity, turbidity, temperature and sampling month (in blue), and (B) organic matter properties constrained by salinity, temperature and sampling month (in blue). Response variables unrelated to main axes (p > 0.05) are not shown.
We observed seasonal changes in organic matter properties and water column structure from May to August along the terrestrial to marine gradient (Figure 8). Changes in water column structure can be attributed to two main drivers: freshwater discharge from land and the advection of Atlantic and Arctic water masses from the shelf into the fjord (Figure 2). In Isfjorden, the main source of freshwater is from melting marine-terminating glaciers, and river runoff sustained by land-terminating glacial meltwater and snow melt (Nilsen et al., 2008). Meanwhile, TAW and AW, largely driven by local wind conditions, enter the fjord in the deep and subsurface waters from the shelf. These two endmembers (terrestrial inputs and marine advected water) as well as local autochthonous production, represent the main sources of OM and inorganic nutrients to Isfjorden. The terrestrial endmember, represented here by river samples, shifted seasonally, with high DOC concentrations in June and high dissolved nutrient (NO2 + NO3 and PO4) concentrations measured in August. Thus, from the spring phytoplankton bloom in May to spring freshet in June and late-season melt in August, we observed strong seasonal changes in nutrients and OM properties in the fjord, with potential implications for coastal biogeochemistry and carbon pathways. The fate of these terrestrial carbon and nutrients in the marine system is likely linked to the physical effects of freshwater, including light attenuation and stratification, as well as the bioavailability of the delivered terrestrial material to marine biological communities.
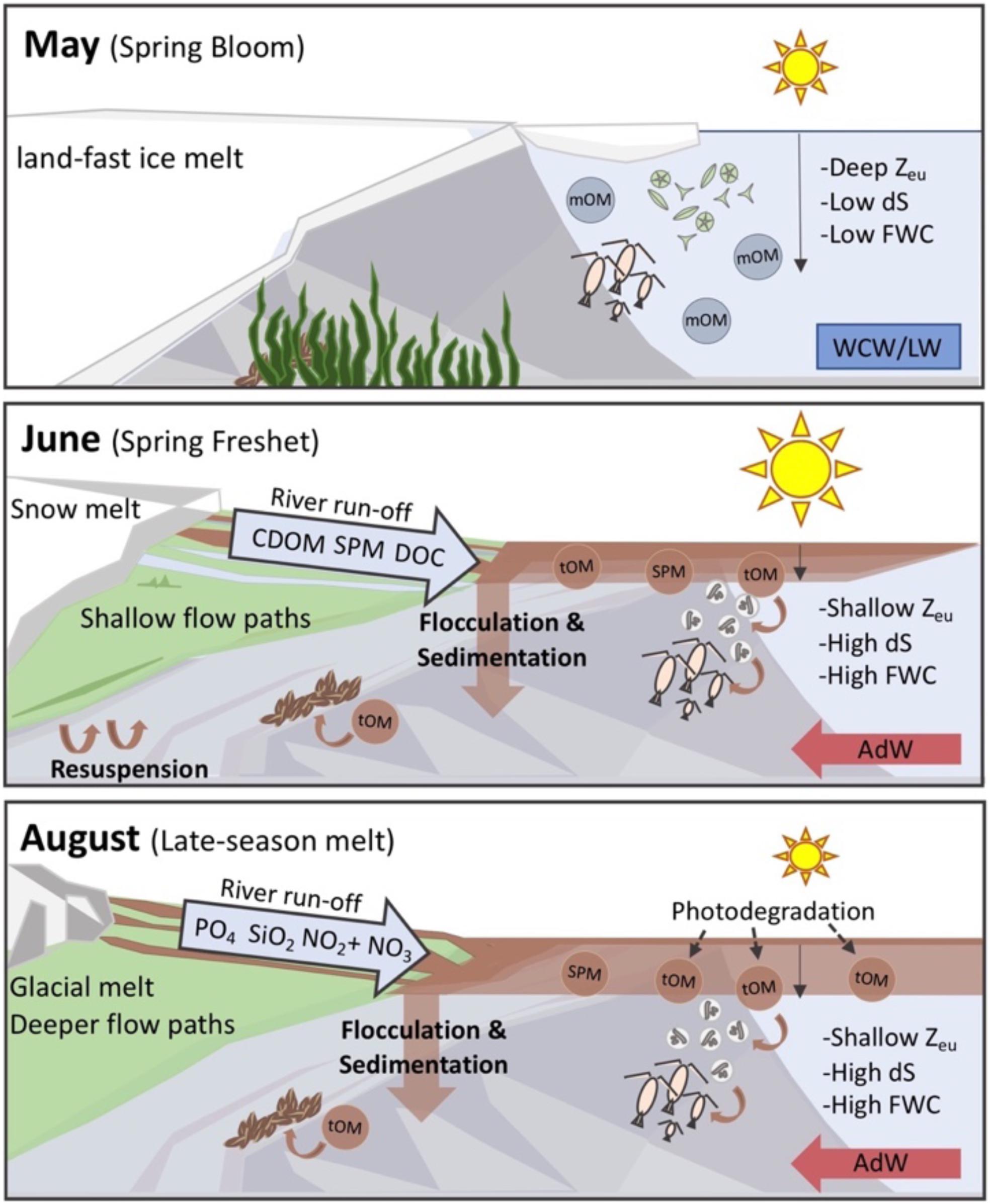
Figure 8. A conceptual diagram summarizing main findings and future perspectives. In May, ice still covered the inner fjord arms of Isfjorden and marine OM (mOM) was present throughout the water column following the spring phytoplankton bloom, when there was a deep euphotic zone (Zeu). The spring freshet in June was a source of terrestrial DOC and SPM to Isfjorden, and while some of these materials were removed from the water column through flocculation and sedimentation, terrestrial OM (tOM) was observed throughout the highly stratified (dS) and turbid fjord surface waters. In August, glacier-fed rivers with deeper flowpaths were a source of nutrients including nitrogen and phosphorus to Isfjorden. Surface waters also had increased fresh water content (FWC) and degraded OM dominated throughout the fjord in August.
Seasonal variation in river water chemistry from May through August reflects changing flow paths in the catchments. River samples collected during the spring freshet in May/June had concentrations of DOC similar to values observed during spring freshet for permafrost dominated catchments in the Siberian and North American Arctic (Holmes et al., 2011; Amon et al., 2012), and much higher than observations from glacier-dominated catchments elsewhere on Svalbard (Zhu et al., 2016) and in Greenland (Paulsen et al., 2017). In fact, concentrations in Sassenelva (a river draining a permafrost-rich valley; Figure 1) in June reached 1400 μmol L–1 while samples from Gipsdalselva and Ebbaelva (both heavily glaciated catchments) were as high as 670 and 1000 μmol L–1, respectively. Adventelva was the only river with low concentrations of DOC in June (40 μmol L–1), but this river was flowing already in May, with a DOC concentration of 980 μmol L–1 at that time (Table 2), confirming that the melt progression occurred earlier in Adventdalen. These high concentrations of riverine DOC draining into Isfjorden in June are consistent with other studies in the Arctic that show that approximately half of Arctic river DOC flux occurs during snow melt (Finlay et al., 2006) and high flow events (Rember and Trefry, 2004; Raymond et al., 2007; Raymond and Saiers, 2010; Coch et al., 2018) when surficial and shallow flow paths (Barnes et al., 2018) and high catchment connectivity (Johnston et al., 2019) help to flush modern, plant-derived OM (Feng et al., 2013) into aquatic systems. Permafrost also plays an important role in mobilization and transport of DOC from C-rich surface soils during snowmelt by sustaining near surface water tables and inhibiting deep percolation (Carey, 2003). Moreover, high discharge periods lead to reduced residence time in the catchment, reducing the potential for processing of DOC during transport from the catchment to coastal areas (Koch et al., 2013; Raymond et al., 2016). Thus, the high concentrations of DOC and increased cDOM observed throughout fjord surface waters in June is likely a result of increased transport of terrestrial OM during the spring freshet.
On Svalbard, late-season run-off is driven by glacial melt (Nowak and Hodson, 2015), which was characterized by much lower concentrations of DOC, but higher concentrations of N and P. Decreases in DOC post-freshet has also been found for the Yukon river (Striegl et al., 2005) and Siberian rivers (Neff et al., 2006) as flow paths deepen. Depending on the geology of the catchment, deeper flow paths can potentially drain nutrient-rich mineral soils, transporting N and P to aquatic systems (Barnes et al., 2018). Alternatively, microbial processes, including nitrification, on catchment glaciers have also been linked to N and P -rich meltwater (Hodson et al., 2004; Telling et al., 2011; Wadham et al., 2016). It is estimated that approximately half of glacially exported N is sourced from microbial activity within glacial sediments at the surface and bed of the ice, doubling N fluxes in runoff (Wadham et al., 2016). However, both glacial and soil-derived nutrients may also be heavily sediment bound (P; Hodson et al., 2004), or retained in the catchment through further microbial processing or uptake by terrestrial vegetation (N; Nowak and Hodson, 2015). Even so, concentrations of NO2 + NO3 in our river samples (sampled close to the river outlet) reached 24 μmol L–1 in August, with estuary surface waters still high at 11.8 μmol L–1. Concentrations of PO4 were also high, reaching 1.7 μmol L–1 in river samples in August. These concentrations are higher than concentrations measured from AW advected from the shelf (maximum of 2 μmol L–1 for NO2 + NO3 and 0.4 μmol L–1 for PO4 in this study, but other observations from Svalbard show 6–11 μmol L–1 for N and 0.8 μmol L–1 for P (Chierici et al., 2019; Halbach et al., 2019). While SiO2 has been associated with glacial meltwater from contact with silica-rich bedrock in Isfjorden (Fransson et al., 2015), Kongsfjorden (Halbach et al., 2019) and Greenland fjords (Meire et al., 2016; Kanna et al., 2018; Hendry et al., 2019), N and P have been linked primarily to advected deep water. In contrast to glacial meltwaters in Kongsfjorden (Halbach et al., 2019) and Greenland (Paulsen et al., 2017), the rivers sampled in our study had comparably high concentrations of N and P in addition to SiO2. While these increased solute concentrations were observed during a relatively low discharge period, the extensive freshwater presence in the fjord in late summer and associated physical effects on the water column could enhance their importance for biological processes.
The physical effects of freshwater and suspended sediments associated with glacial and riverine inputs have implications for the fate of terrestrial OM in the marine system. When river inputs meet the coast, the slowing of the current can cause large particles, including sediment-associated particulate nutrients, to settle out of the water column. In addition, increased salinity causes flocculation and sedimentation of finer particles and dissolved components (Sholkovitz, 1976). These processes are reflected in the exponential decrease in SPM, carbon and nutrients from rivers to estuary stations observed in this study. In the Adventelva estuary, this has been known to lead to the rapid removal of 25% of the suspended sediments from surface waters to the benthos, where hyperpycnal flows transport sediment along the bottom (Zajączkowski, 2008). Despite these losses, concentrations of nutrients in terrestrially influenced surface waters were higher than in subsurface fjord waters, which suggests that these nutrients could support excess coastal production.
Freshwater runoff to surface waters combined with warm, saline water masses transported from the shelf in the deeper waters resulted in seasonally increasing stratification throughout Isfjorden in 2018 and 2019 (Figure 5). As noted in previous studies, strong stratification weakens vertical mixing of the water column and in extreme cases can prevent bottom water renewal (Boone et al., 2017; Torsvik et al., 2019), which can lead to nutrient limitation, especially when nutrients from advected deep waters are important (Bergeron and Tremblay, 2014; Coupel et al., 2015; Yun et al., 2016; Holding et al., 2019). However, in this study, surface waters were influenced by nutrient-rich terrestrial runoff, so the stratification could be an effective physical barrier keeping these nutrients suspended in the euphotic zone, and thus available for primary production. While the fresh surface layer was very thin (and very fresh) in June, mixing of this layer with deeper water can occur through tidal or wind action (Cottier et al., 2010). In August, the fresh surface layer had mixed with the upper water column, resulting in a higher FWC. The deeper mixed layer in August is likely important for the biological utilization of the associated terrestrial nutrients delivered during this period.
High concentrations of SPM are not unusual for coastal waters influenced by runoff from heavily glaciated catchments, where these particles rapidly attenuate light needed for photosynthesis (Murray et al., 2015; Pavlov et al., 2019). In this study, the shallowest mean euphotic depth was observed at estuary stations, where the rapid attenuation of light (max Kd PAR in the top 1 m was 5.40 m–1) resulted in euphotic depths of just over 5 m in June, and 1.55 m in August. Meanwhile, the finer particles, which can remain suspended and, in some cases, can be transported several kilometers from the meltwater plumes (Cowan and Powell, 1991; Meslard et al., 2018), are likely responsible for the far-reaching effects on light attenuation, which reached the fjord transect stations in June. At outer fjord stations, the lowest mean Kd(PAR) was 0.27–0.38 m–1 in August, which is comparable to Kd(PAR) values previously reported in surface waters of WSC in autumn (Pavlov et al., 2015). These corresponded to mean Zeu exceeding 25–30 m. Thus, in August, increased FWC but reduced turbidity may allow for increased photodegradation of terrestrial OM in surface waters. Thus, the fate of transported terrestrial OM is closely tied to the physical effects of terrestrial runoff. Terrestrial carbon and nutrients can be exported to the sediments when reaching the marine system, or transported further out into the fjord where they are largely confined to the mixed layer due to stratification and could potentially be photodegraded or utilized for primary production where turbidity is low enough that sufficient light is available.
The fate of terrestrial OM in the coastal system is also linked to its nutritional value and bioavailability for microbial communities. The seasonality in OM composition observed in this study is linked to the progression from a spring phytoplankton bloom (before spring freshet) to impacts of terrestrial inputs, the geochemical nature of which shifted from freshet to late summer. These seasonal changes, in both the rivers and the fjord, had strong effects on the quality and quantity of DOM throughout the entire fjord and provide insights into the potential for processing of terrestrial carbon in the water column.
In May, the quantity and quality of OM is related to the spring phytoplankton bloom. Monthly chlorophyll a concentrations measured in outer Adventfjorden in 2018 confirm that the spring bloom occurred in early May, roughly a week before the sampling for this study was carried out (Nyeggen, 2019). While the spring bloom was over in the nearshore stations (low concentrations of N and Chl a), the outer fjord stations were characterized by high abundances of Phaeocystis (pers. com; Dąbrowska, 2020). High δ13C values indicate that POC was dominated by marine phytoplankton, and DOM properties (Table 1) also reflect a predominantly marine source of OM. The high S275–295 and SR (Helms et al., 2008) indicate that this freshly produced marine mDOM is of low molecular weight, and is presumably quite bioavailable to bacterial communities. This is in line with a recent study in Isfjorden which highlighted the importance of marine OM, and ice algae for bacterial production following the spring phytoplankton bloom (Holding et al., 2017).
Meanwhile, in June, terrestrially derived OM dominated surface waters throughout the fjord. High DOC concentrations in the rivers in June were found alongside increased aCDOM(375) at all fjord stations, even for more saline samples collected from the outer fjord. Results of stable isotope analysis confirm that the POM at the highly turbid estuary and glacier stations was dominated by terrestrial particles. In fact, terrestrially derived POC was present even in the outer fjord. Surprisingly, while δ13C-POC values from river samples ranged from −24 to −28 ‰, estuary and outer fjord stations had values as low as −30.5 ‰ in June, 2018. Studies in Kongsfjorden have also reported similarly low δ13C values for POC (Kędra et al., 2012; Calleja et al., 2017; Jain et al., 2019), which no clear explanation. We suggest that the low δ13C-POC values here could represent the finer organic fraction of terrestrial POC transported farther from glacial fronts and river outlets, or the transport of material from diffuse runoff, coastal erosion and sediment resuspension in the nearshore (Zajączkowski, 2008). While the outer fjord stations were further from the glacier fronts and river outlets, they were still in close proximity to shore (Figure 1). The low δ15N values of PartN also imply a terrestrial source of POM (Figure 5F). DOM absorption properties for these outer fjord samples with low δ13C-POC further support a terrestrial origin for OM at these sites. As also observed in Kongsfjorden (Calleja et al., 2017), low δ13C values were found alongside steep spectral slopes at the longer wavelengths (S350–400) and low SR (Figure 6), both indications of high molecular weight terrestrial material (Weishaar et al., 2003; Helms et al., 2008). On the other hand, Jain et al. (2019), suggest that low δ13C values can also be observed for marine POC on Svalbard, where increased lipids (which are depleted in 13C) due to the presence of cryophytes in the water column, lead to lower δ13C values in the POM. While cryptophytes and other lipid-rich plankton were present in the water column in June, 2018 (pers. com. Dąbrowska, 2020), no relationship was found between lipid content of POM and δ13C values for POC in our dataset (M. McGovern, unpublished data).
In August, river runoff is driven by glacial meltwater, which was characterized by low DOC concentrations, similar to concentrations found in Bayelva in Kongsfjorden (Zhu et al., 2016). DOM absorption characteristics of these samples reflected a terrestrial yet highly aromatic (high SUVA254) source of DOM (Weishaar et al., 2003). While a high proportion of ancient, glacial OM can be quite labile (Hood et al., 2009) and thus an important resource for microbial processing as glaciers recede, our study indicates that for Isfjorden, terrestrial OM mobilized during freshet (high concentrations of presumably modern, plant-derived DOM from surficial flowpaths), may be more important when considering coastal processes. In fjord surface waters, DOM absorption characteristics in August indicate that while the DOM was terrestrial (and humic), it was also of low molecular weight. Low SR values in August are consistent with previously observed changes in DOM properties associated with photochemical or microbial processing in the marine environment (Moran et al., 2000; Granskog et al., 2012; Asmala et al., 2018). In fact, the decrease in SUVA254 from river to fjord in August, and in S350–400 from June to August in surface waters indicate that photochemical degradation of terrestrial OM, presumably from freshet, could be largely responsible for the observed changes in SR from June to August (Hansen et al., 2016). This photochemical alteration of DOM from larger molecules to smaller labile photoproducts impacts the potential cycling of DOM (Hansen et al., 2016) in Isfjorden, as it could lead to the removal of DOM by volatilization or microbial utilization (Wetzel et al., 1995; Moran and Zepp, 1997). This is in line with the rapid photodegradation of freshet OM to a more bioavailable form readily remineralized by microbial communities in the Mackenzie delta (Gareis and Lesack, 2018) and Kolyma river basin (Mann et al., 2012), and thus may represent an important pathway driving remineralization of terrestrial OM delivered to Isfjorden during freshet.
With air temperatures projected to increase upwards of 10°C by 2100, Svalbard, which is covered by more than 53% glaciers (Nuth et al., 2013), is facing rapid changes (Adakudlu et al., 2019), and the effects are already evident. Pronounced glacier mass loss, changes in precipitation patterns, permafrost warming, and subsequent increases in freshwater runoff have been documented in the last decades (Adakudlu et al., 2019; Błaszczyk et al., 2019; van Pelt et al., 2019) and are expected to continue during this century. The results of this study highlight the spatial and seasonal variability in riverine runoff as a source of OM and inorganic nutrients to Isfjorden, and suggest that in Svalbard, terrestrial DOC inputs could be systematically underestimated due to lack of field sampling during freshet, or following increasingly frequent intense rainfall events (Adakudlu et al., 2019). Since these high DOC concentrations in the river samples are likely due to the flushing of vegetative layer with snow melt, this young terrigenous carbon is presumably semi-bioavailable to fjord microbial communities (Raymond et al., 2007). Moreover, expected increases in vegetative biomass (Myneni et al., 1997; Ju and Masek, 2016) will likely enhance DOC export during periods of high discharge while further permafrost degradation will likely lead to increased POC (Guo and Macdonald, 2006) and nutrient export later in the summer. Higher sediment loads in rivers across the Arctic, including in Adventelva tributaries (Bogen and Bønsnes, 2003) are also expected due to increased erosion with amplified discharge (Syvitski, 2002). Thus, expected future changes in Arctic catchments paired with increased runoff will likely lead to enhanced land-ocean connectivity and increased transport of carbon, nutrients and SPM to coastal areas (Figure 8).
In Svalbard fjords, changes in the timing and geochemical nature of freshwater inputs are occurring alongside increases in Atlantic water advection (Spielhagen et al., 2011), and the disappearance of sea-ice (Muckenhuber et al., 2016) and associated ice algae. Thus, increased freshwater inputs are likely to both limit marine production as the turbid melt season may eventually overlap with the spring phytoplankton bloom, while also providing a potential terrestrial carbon subsidy to marine food-webs. With greener catchments and reductions in sea-ice, terrestrial carbon could become increasingly important for coastal zooplankton and benthos, especially in heavily impacted parts of the fjord where increased light attenuation could limit phytoplankton and macroalgal growth. However, more detailed characterization of the terrestrial DOM pool both seasonally and also between glacial and riverine/permafrost sources is required to better predict the fate of terrestrial material in the marine system. If bioavailable, terrestrial OM can provide heterotrophic bacteria with substrate that allows them to out-compete phytoplankton for nutrients (Sipler et al., 2017), driving shifts in lower food-web structure (Joli et al., 2018; Kellogg et al., 2019) and autotrophic: heterotrophic balance (Wikner and Andersson, 2012). Thus, increasingly persistent and turbid freshwater plumes could lead to changes in basal production, food-web structure and carbon balance in Isfjorden and other Arctic areas facing enhanced land-ocean connectivity (Figure 8).
While it’s evident that terrestrial inputs have profound physical, chemical and biological implications for the fjord, these freshwater plumes are highly variable in space and time. The spatial extent of freshwater plumes is driven by freshwater discharge, the Coriolis effect, tides, ice cover, and the wind direction and strength (Granskog et al., 2005; Forwick et al., 2010). Observed changes in extent and duration of sea ice in Isfjorden (Muckenhuber et al., 2016), and the expected future reduction in sea ice, will also affect the spatial extent of freshwater plumes (Granskog et al., 2005), especially in spring in combination with earlier snow melt (Adakudlu et al., 2019). However, considering the strong explanatory power of turbidity when constraining physicochemical parameters for both sampling years, our results indicate that the use of ocean color data from satellite or airborne platforms has great potential for assessing and quantifying the spatial extent and associated impacts of terrestrial inputs on coastal surface waters. However, the importance of seasonality for constraining OM quality and quantity also emphasizes the need for high temporal resolution data to capture seasonal changes as well as dynamic local events in the catchments and water column.
Seasonality in the magnitude and geochemistry of terrestrial inputs drive strong gradients in light availability, nutrient concentrations, and DOM properties in Isfjorden (Figure 8). Large differences between glacial rivers and marine surface water concentrations indicate that flocculation and sedimentation is an efficient removal pathway for particulate and dissolved carbon and nutrients associated with riverine and glacial SPM. Despite high removal at the land-ocean interface, terrestrial OM was observed throughout Isfjorden’s surface waters in June and August. The physical effects of freshwater on the water column, including retention of terrestrial carbon and nutrients within the euphotic zone due to stratification, may indicate that riverine OM and inorganic nutrients are particularly biologically relevant in coastal systems where vertical mixing is limited during the most productive season. Seasonal shifts in optical properties of DOM further suggest that the photodegradation of terrestrial OM delivered during the spring freshet could lead to increased bioavailability for microbial communities. Climate-change driven increases in freshwater discharge can be expected to lead to increased suspended sediment loads, and the mobilization and transport of terrestrial carbon and nutrients from thawing and greening watersheds, with important implications for future Arctic coastal ecosystems.
The datasets generated for this study are available on request to the corresponding author.
AEP and MM developed the study design. MM, JS, AEP, and AD carried out fieldwork in 2018 and 2019. AKP performed calculations of Kd, FWC, and dS. MM analyzed the data, made the figures, and wrote the manuscript with contributions from all co-authors. All authors contributed to the article and approved the submitted version.
This research was supported by the Norwegian Research Council (TerrACE; project number: 268458), the Fram Center Flagship “Fjord and Coast” grant (FreshFate; project number 132019), and the Svalbard Science Forum’s Arctic Field Grant (RIS number: 10914).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank Espen Lund and Emelie Skogsberg for running the cDOM samples and the NIVA lab (especially Anne Luise Ribeiro and Tina Brytensen) for water chemistry analyses. We would like to thank the students and fellow scientists who helped us with the fieldwork including Uta Brandt, Nathalie Carrasco, Ulrike Dietrich, Cathrine Gundersen, Sverre Johansen, Hannah Miller, Sarah Nelson, Emelie Skogsberg, Liv Sletten, Tobias Vonnahme, and Emilie Hernes Vereide. We also thank Laura de Steur for a discussion about freshwater content calculations. Additional gratitude goes to UNIS logistics and the crew of the RV Helmer Hansen and Clione for their help during the field campaigns.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2020.542563/full#supplementary-material
Adakudlu, M., Andresen, J., Bakke, J., Beldring, S., Benestad, R., Bilt, W., et al. (2019). Climate in Svalbard 2100 – A Knowledge Base for Climate Adaptation. Norway: Norwegian Environmental Agency.
Amon, R., Rinehart, A., Duan, S., Louchouarn, P., Prokushkin, A., Guggenberger, G., et al. (2012). Dissolved organic matter sources in large arctic rivers. Geochim. Cosmochim. Acta 94, 217–237. doi: 10.1016/j.gca.2012.07.015
Asmala, E., Haraguchi, L., Markager, S., Massicotte, P., Riemann, B., Staehr, P. A., et al. (2018). Eutrophication leads to accumulation of recalcitrant autochthonous organic matter in coastal environment. Glob. Biogeochem. Cycles 32, 1673–1687. doi: 10.1029/2017GB005848
Barnes, R. T., Butman, D. E., Wilson, H. F., and Raymond, P. A. (2018). Riverine export of aged carbon driven by flow path depth and residence time. Environ. Sci. Technol. 52, 1028–1035. doi: 10.1021/acs.est.7b04717
Bergeron, M., and Tremblay, J.-E. (2014). Shifts in biological productivity inferred from nutrient drawdown in the southern Beaufort Sea (2003–2011) and the northern Baffin Bay (1997–2011), Canadian Arctic. Geophys. Res. Lett. 41, 3979–3987. doi: 10.1002/2014GL059649
Bianchi, T. S., Arndt, S., Austin, W. E. N., Benn, D. I., Bertrand, S., Cui, X., et al. (2020). Fjords as aquatic critical zones (ACZs). Earth Sci. Rev.ews 203:103145. doi: 10.1016/j.earscirev.2020.103145
Bianchi, T. S., Cui, X., Blair, N. E., Burdige, D. J., Eglinton, T. I., and Galy, V. (2018). Centers of organic carbon burial and oxidation at the land-ocean interface. Organ. Geochem. 115, 138–155. doi: 10.1016/j.orggeochem.2017.09.008
Biskaborn, B. K., Smith, S. L., Noetzli, J., Matthes, H., Vieira, G., Streletskiy, D. A., et al. (2019). Permafrost is warming at a global scale. Nat. Commun. 10:264. doi: 10.1038/s41467-018-08240-4
Blanchet, F. G., Legendre, P., and Borcard, D. (2008). Forward selection of explanatory variables. Ecology 89, 2623–2632. doi: 10.1890/07-0986.1
Błaszczyk, M., Ignatiuk, D., Uszczyk, A., Cielecka-Nowak, K., Grabiec, M., Jania, J. A., et al. (2019). Freshwater input to the Arctic fjord Hornsund (Svalbard). Polar Res. 38:3506. doi: 10.33265/polar.v38.3506
Bogen, J., and Bønsnes, T. E. (2003). Erosion and sediment transport in high arctic rivers, Svalbard. Polar Res. 22, 175–189. doi: 10.1111/j.1751-8369.2003.tb00106.x
Boone, W., Rysgaard, S., Kirillov, S., Dmitrenko, I., Bendtsen, J., Mortensen, J., et al. (2017). Circulation and fjord-shelf exchange during the ice-covered period in young sound-Tyrolerfjord, Northeast Greenland (74° N). Estuar. Coast. Shelf Sci. 194, 205–216. doi: 10.1016/j.ecss.2017.06.021
Calleja, M. L., Kerhervé, P., Bourgeois, S., Kędra, M., Leynaert, A., Devred, E., et al. (2017). Effects of increase glacier discharge on phytoplankton bloom dynamics and pelagic geochemistry in a high Arctic fjord. Prog. Oceanogr. 159, 195–210. doi: 10.1016/j.pocean.2017.07.005
Carey, S. K. (2003). Dissolved organic carbon fluxes in a discontinuous permafrost subarctic alpine catchment. Permafrost Periglac. Process. 14, 161–171. doi: 10.1002/ppp.444
Chierici, M., Vernet, M., Fransson, A., and Børsheim, K. Y. (2019). Net community production and carbon exchange from winter to summer in the Atlantic water inflow to the Arctic Ocean. Front. Mar. Sci. 6:528. doi: 10.3389/fmars.2019.00528
Christiansen, H. H., French, H. M., and Humlum, O. (2005). Permafrost in the Gruve-7 mine, Adventdalen, Svalbard. Norwegian J. Geogr. 59, 109–115. doi: 10.1080/00291950510020592
Coch, C., Lamoureux, S. F., Knoblauch, C., Eischeid, I., Fritz, M., Obu, J., et al. (2018). Summer rainfall dissolved organic carbon, solute, and sediment fluxes in a small Arctic coastal catchment on Herschel Island (Yukon Territory, Canada). Arctic Sci. 4, 750–780. doi: 10.1139/as-2018-0010
Cottier, F. R., Nilsen, F., Skogseth, R., Tverberg, V., Skarðhamar, J., and Svendsen, H. (2010). Arctic fjords: a review of the oceanographic environment and dominant physical processes. Geol. Soc. Lond. Spec. Publ. 344, 35–50. doi: 10.1144/SP344.4
Coupel, P., Ruiz-Pino, D., Sicre, M. A., Chen, J. F., Lee, S. H., Schiffrine, N., et al. (2015). The impact of freshening on phytoplankton production in the Pacific Arctic Ocean. Prog. Oceanogr. 131, 113–125. doi: 10.1016/j.pocean.2014.12.003
Cowan, E. A., and Powell, R. D. (1991). “Ice-proximal sediment accumulation rates in a temperate glacial fjord, southeastern Alaska,” in Glacial Marine Sedimentation: Paleoclimatic Significance, eds J. B. Anderson and G. M. Ashley (Boulder, CO: Geological Society America), 61–73. doi: 10.1130/spe261-p61
Dąbrowska, A. M. (2020). Institute of Oceanology, Polish Academy of Sciences Sopot, Poland. Spring: Personal communication.
Feng, X., Vonk, J. E., Van Dongen, B. E., Gustafsson, Ö, Semiletov, I. P., Dudarev, O. V., et al. (2013). Differential mobilization of terrestrial carbon pools in Eurasian Arctic river basins. Proc. Natl. Acad. Sci. U.S.A. 110, 14168–14173. doi: 10.1073/pnas.1307031110
Finlay, J., Neff, J., Zimov, S., Davydova, A., and Davydov, S. (2006). Snowmelt dominance of dissolved organic carbon in high-latitude watersheds: implications for characterization and flux of river DOC. Geophys. Res. Lett. 33:L10401. doi: 10.1029/2006GL025754
Forwick, M., Vorren, T. O., Hald, M., Korsun, S., Roh, Y., Vogt, C., et al. (2010). Spatial and temporal influence of glaciers and rivers on the sedimentary environment in Sassenfjorden and Tempelfjorden, Spitsbergen. Geol. Soc. Lond. Spec. Publ. 344, 163–193. doi: 10.1144/sp344.13
Fransson, A., Chierici, M., Nomura, D., Granskog, M. A., Kristiansen, S., Martma, O., et al. (2015). Effect of glacial drainage water on the CO2 system and ocean acidification state in an Arctic tidewater-glacier fjord during two contrasting years. J. Geophys. Res. Ocean. 120, 2413–2429. doi: 10.1002/2014JC010320
Fraser, N. J., Skogseth, R., Nilsen, F., and Inall, M. E. (2018). Circulation and exchange in abroad Arctic fjord using glider-based observations. Polar Res. 37:1485417. doi: 10.1080/17518369.2018.1485417
Gareis, J. A., and Lesack, L. F. (2018). Photodegraded dissolved organic matter from peak freshet river discharge as a substrate for bacterial production in a lake-rich great Arctic delta. Arctic Sci. 4, 557–583. doi: 10.1139/as-2017-0055
Granskog, M. A., Ehn, J., and Niemelä, M. (2005). Characteristics and potential impacts of under-ice river plumes in the seasonally ice-covered Bothnian Bay (Baltic Sea). J. Mar. Syst. 53, 187–196. doi: 10.1016/j.jmarsys.2004.06.005
Granskog, M. A., Stedmon, C. A., Dodd, P. A., Amon, R. M., Pavlov, A. K., de Steur, L., et al. (2012). Characteristics of colored dissolved organic matter (CDOM) in the Arctic outflow in the Fram Strait: assessing the changes and fate of terrigenous CDOM in the Arctic Ocean. J. Geophys. Res. Oceans 117:C12021. doi: 10.1029/2012JC008075
Grosse, G., Goetz, S., McGuire, A. D., Romanovsky, V. E., and Schuur, E. A. (2016). Changing permafrost in a warming world and feedbacks to the Earth system. Environ. Res. Lett. 11:040201. doi: 10.1088/1748-9326/11/4/040201
Guerrero, J. L. (2019). Norwegian Institute for Water Research. Oslo: Personal communication related to Isfjorden catchments.
Guo, L., and Macdonald, R. W. (2006). Source and transport of terrigenous organic matter in the upper Yukon River: evidence from isotope (d13C,D14C, and d15N) composition of dissolved, colloidal, and particulate phases. Glob. Biogeochem. Cycles 20:GB2011. doi: 10.1029/2005GB002593
Halbach, L., Assmy, P., Vihtakari, M., Hop, H., Duarte, P., Wold, A., et al. (2019). Tidewater glaciers and bedrock characteristics control the phytoplankton growth environment in an Arctic fjord. Front. Mar. Sci. 6:254. doi: 10.3389/fmars.2019.00254
Hansen, A. M., Kraus, T. E. C., Pellerin, B. A., Fleck, J. A., Downing, B. D., and Bergamaschi, B. A. (2016). Optical properties of dissolved organic matter (DOM): effects of biological and photolytic degradation. Limnol. Oceanogr. 61, 1015–1032 doi: 10.1002/lno.10270
Harris, C. N., McTigue, N. D., McClelland, J. W., and Dunton, K. H. (2018). Do high Arctic coastal food webs rely on a terrestrial carbon subsidy? Food Webs 15, 2352–2496.
Helms, J. R., Stubbins, A., Ritchie, J. D., Minor, E. C., Kieber, D. J., and Mopper, K. (2008). Absorption spectral slopes and slope ratios as indicators of molecular weight, source, and photobleaching of chromophoric dissolved organic matter. Limnol. Oceanogr. 53, 955–969. doi: 10.4319/lo.2008.53.3.0955
Hendry, K. R., Huvenne, V. A. I., Robinson, L. F., Annett, A., Badger, M., Jacobel, A. W., et al. (2019). The biogeochemical impact of glacial meltwater from Southwest Greenland. Prog. Oceanogr. 176:102126. doi: 10.1016/j.pocean.2019.102126
Hodson, A., Mumford, P., and Lister, D. (2004). Suspended sediment and phosphorus in proglacial rivers: bioavailability and potential impacts upon the P status of ice-marginal receiving waters. Hydrol. Process. 18, 2409–2422. doi: 10.1002/hyp.1471
Hodson, A., Nowak, A., and Christiansen, H. (2016). Glacial and periglacial floodplain sediments regulate hydrologic transfer of reactive iron to a high arctic fjord. Hydrol. Process. 30, 1219–1229. doi: 10.1002/hyp.10701
Holding, J. M., Duarte, C. M., Delgado-Huertas, A., Soetaert, K., Vonk, J. E., Agustí, S., et al. (2017). Autochthonous and allochthonous contributions of organic carbon to microbial food webs in Svalbard fjords. Limnol. Oceanography 62, 1307–1323. doi: 10.1002/lno.10526
Holding, J. M., Markager, S., Juul-Pedersen, T., Paulsen, M. L., Moller, E. F., Meire, L., et al. (2019). Seasonal and spatial patterns of primary production in a high-latitude fjord affected by Greenland Ice Sheet run-off. Biogeosciences 16, 3777–3792. doi: 10.5194/bg-16-3777-2019
Holinde, L., and Zielinski, O. (2016). Bio-optical characterization and light availability parameterization in Uummannaq Fjord and Vaigat-Disko Bay (West Greenland). Ocean Sci. 12, 117–128. doi: 10.5194/os-12-117-2016
Holmes, R. M., McClelland, J. W., Peterson, B. J., Tank, S. E., Bulygina, E., Eglinton, T. I., et al. (2011). Seasonal and annual fluxes of nutrients and organic matter from large rivers to the Arctic Ocean and surrounding seas. Estuar. Coasts 35, 369–382. doi: 10.1007/s12237-011-9386-6
Holmes, R. M., McClelland, J. W., Raymond, P. A., Frazer, B. B., Peterson, B. J., and Stieglitz, M. (2008). Lability of DOC transported by Alaskan rivers to the Arctic Ocean. Geophys. Res. Lett. 35:L03402. doi: 10.1029/2007GL032837
Hood, E., Fellman, J., Spencer, R. G. M., Hernes, P. J., Edwards, R., D’Amore, D., et al. (2009). Glaciers as a source of ancient and labile organic matter to the marine environment. Nature 462, 1044–1047. doi: 10.1038/nature08580
Hu, C., Muller-Karger, F. E., and Zepp, R. G. (2002). Absorbance, absorption coefficient, and apparent quantum yield: a comment on common ambiguity in the use of these optical concepts. Limnol. Oceanogr. 47, 1261–1267. doi: 10.4319/lo.2002.47.4.1261
IPCC (2014). “Climate change 2014: synthesis report,” in Proceedings of the Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change[Core Writing Team], eds R. K. Pachauri and L. A. Meyer (Geneva: IPCC).
Jain, A., Krishnan, K. P., Singh, A., Thomas, F. A., Begum, N., Tiwari, M., et al. (2019). Biochemical composition of particles shape particle-attached bacterial community structure in a high Arctic fjord. Ecol. Indic. 102, 581–592. doi: 10.1016/j.ecolind.2019.03.015
Johnston, S. E., Bogard, M. J., Rogers, J. A., Butman, A., Striegl, R. G., and Dornblaser et al. (2019). Constraining dissolved organic matter sources and temporal variability in a model sub-Arctic lake. Biogeochemistry 146, 271–292. doi: 10.1007/s10533-019-00619-9
Joli, N., Gosselin, M., Ardyna, M., Babin, M., Onda, D. F., Tremblay, J. E., et al. (2018). Need for focus on microbial species following ice melt and changing freshwater regimes in a Janus Arctic Gateway. Sci. Rep. 8:9405. doi: 10.1038/s41598-018-27705-6
Ju, J., and Masek, J. G. (2016). The vegetation greenness trend in Canada and US Alaska from 1984–2012 Landsat data. Remote Sens. Environ. 176, 1–16. doi: 10.1016/j.rse.2016.01.001
Kanna, N., Sugiyama, S., Ohashi, Y., Sakakibara, D., Fukamachi, Y., and Nomura, D. (2018). Upwelling of macronutrients and dissolved inorganic carbon by a subglacial freshwater driven plume in Bowdoin Fjord, northwestern Greenland. J. Geophys. Res. Biogeosci. 123, 1666–1682. doi: 10.1029/2017jg004248
Kaste, Ø, Skarbøvik, E., Greipsland, I., Gundersen, C. B., Austnes, K., Skancke, L. B., et al. (2018). The Norwegian River Monitoring Programme–Water Quality Status and Trends 2017. NIVA-Repport M-no 1168. Norway: Miljødirektoratet.
Kędra, M., Kuliński, K., Walkusz, W., and Legeżyńska, J. (2012). The shallow benthic food web structure in the high Arctic does not follow seasonal changes in the surrounding environment. Estuar. Coast. Shelf Sci. 114, 183–191. doi: 10.1016/J.ECSS.2012.08.015
Kellogg, C. T. E., McClelland, J. W., Dunton, K. H., and Crump, B. C. (2019). Strong seasonality in arctic estuarine microbial food webs. Front. Microbiol. 10:2628. doi: 10.3389/fmicb.2019.02628
Kirk, J. T. O. (2010). Light and Photosynthesis in Aquatic Ecosystems. Cambridge: Cambridge University Press.
Koch, J. C., Runkel, R. L., Striegl, R., and McKnight, D. M. (2013). Hydrologic controls on the transport and cycling of carbon and nitrogen in a boreal catchment underlain by continuous permafrost. J. Geophys. Res. Biogeo. 118, 698–712. doi: 10.1002/jgrg.20058
Koziol, K. A., Moggridge, H. L., Cook, J. M., and Hodson, A. J. (2019). Organic carbon fluxes of a glacier surface: a case study of Foxfonna, a small Arctic glacier. Earth Surf. Process. Landf. 44, 405–416. doi: 10.1002/esp.4501
Luckmann, A., Benn, D. I., Cottier, F., Bevan, S., Nilsen, F., and Inall, M. (2015). Calving rates at tidewater glaciers vary strongly with oceantemperature. Nat. Comm. 6:8566. doi: 10.1038/ncomms9566
Mann, P. J., Davydova, A., Zimov, N., Spencer, R. G. M., Davydov, S., Bulygina, E., et al. (2012). Controls on the composition and lability of dissolved organic matter in Siberia’s Kolyma River basin. J. Geophys. Res. 117:G01028. doi: 10.1029/2011jg001798
McClelland, J. W., Déry, S. J., Peterson, B. J., Holmes, R. M., and Wood, E. F. (2006). A pan-arctic evaluation of changes in river discharge during the latter half of the 20th century. Geophys. Res. Lett. 33:L06715. doi: 10.1029/2006gl025753
Meire, L., Mortensen, J., Rysgaard, S., Bendtsen, J., Boone, W., Meire, P., et al. (2016). Spring bloom dynamics in a subarctic fjord influenced by tidewater outlet glaciers (Godthåbsfjord, SW Greenland). J. Geophys. Res. Biogeosci. 121, 1581–1592. doi: 10.1002/2015JG003240
Meslard, F., Bourrin, F., Many, G., and Kerhervé, P. (2018). Suspended particle dynamics and fluxes in an Arctic fjord (Kongsfjorden, Svalbard). Estuar. Coast. Shelf Sci. 204, 212–224. doi: 10.1016/j.ecss.2018.02.020
Moran, M. A., Sheldon, W. M. Jr., and Zepp, R. G. (2000). Carbon loss and optical property changes during long-term photochemical and biological degradation of estuarine dissolved organic matter. Limnol. Oceanogr. 45, 1254–1264. doi: 10.4319/lo.2000.45.6.1254
Moran, M. A., and Zepp, R. G. (1997). Role of photoreactions in the formation of biologically labile compounds from dissolved organic matter. Limnol. Oceanogr. 42, 1307–1316. doi: 10.4319/lo.1997.42.6.1307
Muckenhuber, S., Nilsen, F., Korosov, A., and Sandven, S. (2016). Sea ice cover in Isfjorden and Hornsund, Svalbard (2000–2014) from remote sensing data. Cryosphere 10, 149–158. doi: 10.5194/tc-10-149-2016
Murray, C., Markager, S., Stedmon, C. A., Juul-Pedersen, T., Sejr, M. K., and Bruhn, A. (2015). The influence of glacial melt water on bio-optical properties in two contrasting Greenlandic fjords. Estuar. Coast. Shelf Sci. 163, 72–83. doi: 10.1016/j.ecss.2015.05.041
Myneni, R. B., Keeling, C. D., Tucker, C. J., Asrar, G., and Nemani, R. R. (1997). Increased plant growth in the northern high latitudes from 1981-1991). Nature 386, 698–701. doi: 10.1038/386698a0
Neff, J. C., Finaly, J. C., Zimov, S. A., Davydov, S. P., Carrasco, J. J., Schuur, E. A. G., et al. (2006). Seasonal changes in the age and structure of dissolved organic carbon in Siberian rivers and streams. Geophy. Res. Lett. 33:L10401. doi: 10.1029/2006GL028222
Nilsen, F., Cottier, F., Skogseth, R., and Mattsson, S. (2008). Fjord-shelf exchanges controlled by ice and brine production: the interannual variation of Atlantic Water in Isfjorden, Svalbard. Contin. Shelf Res. 28, 1838–1853. doi: 10.1016/j.csr.2008.04.015
Nilsen, F., Skogseth, R., Vaardal-Lunde, J., and Inall, M. (2016). A simple shelf circulation model: intrusion of Atlantic Water on the West Spitsbergen Shelf. J. Phys. Oceanogr. 46, 1209–1230. doi: 10.1175/jpo-d-15-0058.1
Nowak, A., and Hodson, A. (2015). On the biogeochemical response of a glacierized High Arctic watershed to climate change: revealing patterns, processes and heterogeneity among micro-catchments. Hydrol. Process. 29, 1588–1603. doi: 10.1002/hyp.10263
Nuth, C., Kohler, J., König, M., von Deschwanden, A., Hagen, J. O., and Kääb, A. (2013). Decadal changes from a multi-temporal glacier inventory of Svalbard. Cryosphere 7, 1603–1621. doi: 10.5194/tc-7-1603-2013
Nyeggen, M. U. (2019). Seasonal Zooplankton Dynamics in Svalbard Coastal Waters: The Shifting Dominance of Mero- and Holoplankton and Timing of Reproduction in Three Species of Copepoda. Master thesis. Bergen: University of Bergen.
Oksanen, J., Blanchet, F. G., Friendly, M., Kindt, R., Legendre, P., McGlinn, D., et al. (2018). vegan: Community Ecology Package. R Package Version 2.5-2. 2018.
Parmentier, F. J. W., Christensen, T. R., Rysgaard, S., Bendtsen, J., Glud, R. N., Else, B., et al. (2017). A synthesis of the arctic terrestrial and marine carbon cycles under pressure from a dwindling cryosphere. Ambio 46, S53–S69. doi: 10.1007/s13280-016-0872-8
Parsons, T. R. (2013). A Manual of Chemical & Biological Methods for Seawater Analysis. Kent: Elsevier.
Parsons, T. R., Webb, D. G., Rokeby, B. E., Lawrence, M., Hopkey, G., and Chiperzak, D. (1989). Autotrophic and heterotrophic production in the Mackenzie River/Beaufort Sea estuary. Polar Biol. 9, 261–266. doi: 10.1007/bf00263774
Paulsen, M. L., Nielsen, S. E. B., Müller, O., Møller, E. F., Stedmon, C. A., Juul-Pedersen, T., et al. (2017). Carbon bioavailability in a high arctic fjord influenced by glacial meltwater, NE Greenland. Front. Mar. Sci. 4:176. doi: 10.3389/fmars.2017.00176
Pavlov, A. K., Granskog, M. A., Stedmon, C. A., Ivanov, B. V., Hudson, S. R., and Falk-Petersen, S. (2015). Contrasting optical properties of surface waters across the Fram Strait and its potential biological implications. J. Mar. Syst. 143, 62–72. doi: 10.1016/j.jmarsys.2014.11.001
Pavlov, A. K., Leu, E., Hanelt, D., Bartsch, I., Karsten, U., Hudson, S. R., et al. (2019). “The underwater light climate in Kongsfjorden and its ecological implications,” in The Ecosystem of Kongsfjorden, Svalbard, eds H. Hop and C. Wiencke (Cham: Springer International Publishing), 137–170. doi: 10.1007/978-3-319-46425-1_5
Peterson, B. J., and Fry, B. (1987). Stable isotopes in ecosystem studies. Annu. Rev. Ecol. Syst. 18, 293–320. doi: 10.1146/annurev.es.18.110187.001453
Peterson, B. J., Holmes, R. M., McClelland, J. W., Vörösmarty, C. J., Lammers, R. B., Shiklomanov, A. I., et al. (2002). Increasing river discharge to the Arctic Ocean. Science 298, 2171–2173. doi: 10.1126/science.1077445
Proshutinsky, A., Krishfield, R., Timmermans, M.-L., Toole, J., Carmack, E., McLaughlin, F., et al. (2009). Beaufort gyre freshwater reservoir: state and variability from observations. J. Geophys. Res. Oceans 114:C00A10. doi: 10.1029/2008jc005104
R Core Team (2017). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Raymond, P., Saiers, J., and Sobczak, W. (2016). Hydrological and biogeochemical controls on watershed dissolved organic matter transport: pulse-shunt concept. Ecology 97, 5–16. doi: 10.1890/14-1684.1
Raymond, P. A., McClelland, J. W., Holmes, R. M., Zhulidov, A. V., Mull, K., Peterson, B. J., et al. (2007). Flux and age of dissolved organic carbon exported to the Arctic Ocean: a carbon isotopic study of the five largest arctic rivers. Glob. Biogeochem. Cycles 21:GB401. doi: 10.1029/2007gb002934
Raymond, P. A., and Saiers, J. E. (2010). Event controlled DOC export from forested watersheds. Biogeochemistry 100, 197–209. doi: 10.1007/s10533-010-9416-7
Rember, R. D., and Trefry, J. H. (2004). Increased concentrations of dissolved trace metals and organic carbon during snowmelt in rivers of the Alaskan Arctic. Geochim. Cosmochim. Acta 68, 477–489. doi: 10.1016/s0016-7037(03)00458-7
Sholkovitz, E. R. (1976). Flocculation of dissolved organic and inorganic matter during the mixing of river water and seawater. Geochim. Cosmochim. Acta 40, 831–845. doi: 10.1016/0016-7037(76)90035-1
Sipler, R. E., Kellogg, C. T. E., Connelly, T. L., Roberts, Q. N., Yager, P. L., and Bronk, D. A. (2017). Microbial community response to terrestrially derived dissolved organic matter in the coastal Arctic. Front. Microbiol. 8:1018. doi: 10.3389/fmicb.2017.01018
Smith, R. W., Bianchi, T. S., Allison, M., Savage, C., and Galy, V. (2015). High rates of organic carbon burial in fjord sediments globally. Nat. Geosci. 8, 450–U446. doi: 10.1038/ngeo2421
Spencer, R. G. M., Aiken, G. R., Wickland, K. P., Striegl, R. G., and Hernes, P. J. (2008). Seasonal and spatial variability in dissolved organic matter quantity and composition from the Yukon River basin, Alaska. Glob. Biogeochem. Cycles 22:GB4002. doi: 10.1029/2008gb003231
Spielhagen, R. F., Werner, K., Sørensen, S. A., Zamelczyk, K., Kandiano, E., Budeus, G., et al. (2011). Enhanced modern heat transfer to the arctic by warm atlantic water. Science 331, 450–453. doi: 10.1126/science.1197397
Stedmon, C., and Markager, S. (2001). The optics of chromophoric dissolved organic matter (CDOM) in the Greenland Sea: an algorithm for differentiation between marine and terrestrially derived organic matter. Limnol. Oceanogr. 46, 2087–2093. doi: 10.4319/lo.2001.46.8.2087
Striegl, R. G., Aiken, G. R., Dornblaser, M. M., Raymond, P. A., and Wickland, K. P. (2005). A decrease in discharge-normalized DOC export by the Yukon River during summer through autumn. Geophys. Res. Lett. 32:L21413. doi: 10.1029/2005GL024413
Syvitski, J. P. M. (2002). Sediment discharge variability in Arctic rivers: implications for a warmer future. Polar Res. 21:323. doi: 10.3402/polar.v21i2.6494
Tarnocai, C., Canadell, J. G., Schuur, E. A. G., Kuhry, P., Mazhitova, G., and Zimov, S. (2009). Soil organic carbon pools in the northern circumpolar permafrost region. Glob. Biogeochem. Cycles 23:GB2023. doi: 10.1029/2008gb003327
Telling, J., Anesio, A. M., Tranter, M., Irvine-Fynn, T., Hodson, A., Butler, C., et al. (2011). Nitrogen fixation on Arctic glaciers, Svalbard. J. Geophys. Res. Biogeosci. 116:G03039. doi: 10.1029/2010jg001632
Torsvik, T., Albretsen, J., Sundfjord, A., Kohler, J., Sandvik, A. D., Skarðhamar, J., et al. (2019). Impact of tidewater glacier retreat on the fjord system: modeling present and future circulation in Kongsfjorden, Svalbard. Estuar. Coast. Shelf Sci. 220, 152–165. doi: 10.1016/j.ecss.2019.02.005
van Pelt, W., Pohjola, V. A., Pettersson, R., Marchenko, S., Kohler, J., Luks, B., et al. (2019). A long-term dataset of climatic mass balance, snow conditions, and runoff in Svalbard (1957-2018). Cryosphere 13, 2259–2280. doi: 10.5194/tc-13-2259-2019
van Pelt, W. J. J., Pohjola, V. A., and Reijmer, C. H. (2016). The changing impact of snow conditions and refreezing on the mass balance of an idealized Svalbard Glacier. Front. Earth Sci. 4:102. doi: 10.3389/feart.2016.00102
Vihtakari, M. (2019). PlotSvalbard: PlotSvalbard – Plot Research Data From Svalbard on Maps. Rpackage version 0.8.5.
Wadham, J. L., Hawkings, J., Telling, J., Chandler, D., Alcock, J., O’Donnell, E., et al. (2016). Sources, cycling and export of nitrogen on the Greenland Ice Sheet. Biogeosciences 13, 6339–6352. doi: 10.5194/bg-13-6339-2016
Weishaar, J. L., Aiken, G. R., Bergamaschi, B. A., Fram, M. S., Fujii, R., and Mopper, K. (2003). Evaluation of specific ultraviolet absorbance as an indicator of the chemical composition and reactivity of dissolved organic carbon. Environ. Sci. Technol. 37, 4702–4708. doi: 10.1021/es030360x
Wetzel, R. G., Hatcher, P. G., and Bianchi, T. S. (1995). Natural photolysis by ultraviolet irradiance of recalcitrant dissolved organic matter to simple substrates for rapid bacterial metabolism. Limnol. Oceanogr. 40, 1369–1380. doi: 10.4319/lo.1995.40.8.1369
Wikner, J., and Andersson, A. (2012). Increased freshwater discharge shifts the trophic balance in the coastal zone of the northern Baltic Sea. Glob. Chang. Biol. 18, 2509–2519. doi: 10.1111/j.1365-2486.2012.02718.x
Yun, M. S., Whitledge, T. E., Stockwell, D., Son, S. H., Lee, J. H., Park, J. W., et al. (2016). Primary production in the Chukchi Sea with potential effects of freshwater content. Biogeosciences 13, 737–749. doi: 10.5194/bg-13-737-2016
Zajączkowski, M. (2008). Sediment supply and fluxes in glacial and outwash fjords, Kongsfjorden and Adventfjorden, Svalbard. Polish Polar Res. 29, 59–72.
Keywords: climate change, coastal biogeochemistry, dissolved organic matter, freshwater inputs, glacier runoff, light climate, permafrost, land-ocean interactions
Citation: McGovern M, Pavlov AK, Deininger A, Granskog MA, Leu E, Søreide JE and Poste AE (2020) Terrestrial Inputs Drive Seasonality in Organic Matter and Nutrient Biogeochemistry in a High Arctic Fjord System (Isfjorden, Svalbard). Front. Mar. Sci. 7:542563. doi: 10.3389/fmars.2020.542563
Received: 13 March 2020; Accepted: 17 August 2020;
Published: 08 September 2020.
Edited by:
Susana Agusti, King Abdullah University of Science and Technology, Saudi ArabiaReviewed by:
Maria Lund Paulsen, Aarhus University, DenmarkCopyright © 2020 McGovern, Pavlov, Deininger, Granskog, Leu, Søreide and Poste. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Maeve McGovern, bWFldmUubWNnb3Zlcm5Abml2YS5ubw==; Amanda E. Poste, YW1hbmRhLnBvc3RlQG5pdmEubm8=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.