 E. Mabragaña
E. Mabragaña M. González-Castro
M. González-Castro V. Gabbanelli
V. Gabbanelli D. M. Vazquez
D. M. Vazquez J. M. Díaz de Astarloa
J. M. Díaz de Astarloa- Laboratorio de Biotaxonomía Morfológica y Molecular de Peces, Instituto de Investigaciones Marinas y Costeras (IIMyC), CONICET, Universidad Nacional de Mar del Plata, Mar del Plata, Argentina
Skeletal structures, especially the claspers, neurocrania, and scapulocoracoids have been widely used to characterize and describe new species of elasmobranchs. Intra and interspecific variation of scapulocoracoids in 3 species of Psammobatis (n = 94) from the Southwest Atlantic Ocean were analyzed to assess its utility as a diagnostic character in skates. Moreover, based on scapulocoracoids and body morphometric analyses, the validity of Psammobatis parvacauda McEachran, 1983 was evaluated. A remarkable sexual dimorphism in scapulocoracoids was observed in all species in accordance with original descriptions, which was also evident in the principal component (PCA) and Discriminant Analyses. The number and arrangement of post-dorsal and post-ventral fenestra were highly variable at intra-specific level, in contrast with original descriptions. Particularly, one of the morphotypes observed in females of P. normani was identical to the one reported in P. parvacauda. Results showed intraspecific variation in this structure, as was previously observed in Sympterygia species. Therefore, this structure should not be used as a diagnostic character in skates, at least for these genera. Regarding body morphometry the PCA showed that P. parvacauda grouped with P. normani females and the cross-validated analysis showed that P. parvacauda was classified in the group of P. normani females. P. parvacauda is only known from its original description (one female) and a likely second specimen, but our analyses revealed a lack of diagnostic characteristics. Based on observations and measurements of the holotype of P. parvacauda, and comparisons with congenerics, P. normani is regarded a senior synonym of P. parvacauda.
Introduction
Skates (Chondrichthyes, Rajiformes), constitute a monophyletic and cosmopolitan group of cartilaginous fishes, with representatives in all oceans. The Order includes approximately 290 species and is composed by two highly diverse families (Rajidae and Arhynchobatidae) and two smaller families (Anacanthobatidae and Gurgesiellidae) (Last et al., 2016a, b; Séret et al., 2016; Weigmann et al., 2016). The endoskeleton of skates -like all chondrichthyan fishes- is comprised predominantly of a hyaline-like cartilage that persists throughout the entire life and is not replaced by bone (Compagno, 1999; Seidel et al., 2017). During ontogeny the matrix undergoes a distinctive calcification, developing an outer calcified rind of hydroxyapatite or tesserae, located between the cartilaginous core and the outer, perichondrium (Seidel et al., 2017).
Skeletal structures, especially the claspers, neurocrania and scapulocoracoids have been widely used not only to characterize and describe new species of skates (McEachran and Compagno, 1982; Stehmann and Seret, 1983; McEachran and Last, 1994; de Carvalho et al., 2005; Last and McEachran, 2006; Last and Gledhill, 2007; Jeong and Nakabo, 2009), but also for conducting comprehensive systematic studies (Stehmann, 1970; Hulley, 1972; McEachran and Miyake, 1990; McEachran and Dunn, 1998). The scapulocoracoid or pectoral girdle is located just posterior to the branchial arches and is attached to the vertebral column dorsally (Compagno, 1999). The scapulocoracoid consists basically of a ventral and transverse coracoid bar, a dorsolateral scapular process on each side and, in some species an articulated suprascapula above the scapular process. The lateral face of the scapulocoracoid has an articular surface for the pectoral basal cartilages and foramina for blood vessels and nerves (Compagno, 1999). Fenestrae on the lateral face of the scapulocoracoid of primitive batoids include anterodorsal and anteroventral fenestrae between the pro and mesocondyles, and postdorsal and posventral fenestrae between the meso and metacondyles (Compagno, 1999).
McEachran (1982, 1983) conducted a comprehensive review of skate species from the Sympterygia and Psammobatis genera, and described their skeletal structures including scapulocoracoids. He found a strong sexual dimorphism in this part of the skeleton in Psammobatis species, and defined the overall morphological pattern of this structure for each species, endorsing it as a diagnostic character. Recent studies have called into question the usefulness of scapulocoracoids as a diagnostic character at the specific level. Jurado et al. (2017), reported a high variability in this structure at the intraspecific level in Sympterygia species, especially in the number of postdorsal and postventral fenestrae. On this basis, it would be relevant to investigate potential morphological variability in scapulocoracoids within Psammobatis, a closely related genus (McEachran and Dunn, 1998), to determine whether a similarly non-conservative pattern is observed.
The genus Psammobatis (Rajiformes, Arhynchobatidae) is endemic of South America and is comprised of eight small to medium sized skates. Thorough taxonomic revision of the genus was conducted by McEachran (1983) and completed by de Carvalho and Figueiredo (1994). The genus currently includes two amphioceanic species, P. rudis Gunther, 1870 and P. normani McEachran, 1983, both occurring in the Southwest Atlantic Ocean (SWA) and the South-east Pacific Ocean (SEP), one species exclusively found in the SEP, P. scobina (Philippi, 1857), and 5 species exclusively distributed in the SWA, P. bergi Marini, 1932, P. rutrum Jordan, 1890, P. extenta (Garman, 1913), P. lentiginosa McEachran, 1983, and Psammobatis parvacauda McEachran, 1983. The latter was described based on a single female collected around Malvinas/Falkland Islands, whose external morphological characteristics and skeletal structures, particularly the scapulocoracoid, were unique (McEachran, 1983). After the revision of McEachran (1983), several studies were conducted on almost all Psammobatis species including reproductive biology (Braccini and Chiaramonte, 2002b; Mabragaña and Cousseau, 2004; San Martín et al., 2005, Mabragaña, 2007; Perier et al., 2011; Mabragaña et al., 2012; Martins and Oddone, 2017), feeding ecology (Braccini and Perez, 2005; Mabragaña, 2007; Mabragaña and Giberto, 2007; San Martín et al., 2007; Barbini and Lucifora, 2012), morphology (Braccini and Chiaramonte, 2002a; Mabragaña, 2007), egg cases (Concha et al., 2009; Mabragaña et al., 2011; Vazquez et al., 2016), and their parasites community (Irigoitia et al., 2019). Strikingly, no further studies were conducted in P. parvacauda. In fact, no new records of this species were published since McEachran’s (1983) original description.
The objectives of this study are (1) to analyze the morphological variability of scapulocoracoids in three Psammobatis species from the SWA, (2) to assess the usefulness of scapulocoracoid as a diagnostic character in this genus, and (3) in the light of objectives 1 and 2, to assess the validity of P. parvacauda McEachran, 1983 based on morphological and skeletal analyses.
Materials and Methods
Sample Collection
Skeletal material was obtained from specimens of P. rudis (males n = 13, females n = 18) and P. normani (males n = 18, females n = 19), collected by trawl fishing during research cruises conducted by the National Institute for Fisheries Research and Development (INIDEP) in southern Patagonian waters (49°28′S, 67°03′W to 52°00′S, 53°49′W, and from 84 to 182 m) in April 2000, and from specimens of P. lentiginosa (males n = 13, females n = 13) collected off Buenos Aires Province (36°23′S, 63°59′W to 39°05′S, 58°04′W, from 66 to 99 m depth) in October 2006.
Total length (TL) and disc width (DW) in millimeters, sex and maturity (following Mabragaña and Cousseau, 2004) were recorded for each specimen in the laboratory. Scapular girdles were dissected and kept frozen for subsequent study. In order to reveal scapulocoracoid structure (shape, number, and fenestrae pattern), dissected material was submerged in water at 90–95°C for easy flesh removal. Once cartilaginous structures were cleaned, they were stored in ethanol 70%.
Scapulocoracoid Morphometric Analysis
Morphometric (linear morphometric measurements; LMM) were taken following McEachran and Compagno (1979). Morphometric variables included greatest length and height, anterior and posterior length to mesocondyle (pre- and postmesocondyle, respectively), and height of rear corner. These variables were taken on the left scapulocoracoid side. Given that in P. lentiginosa only 10 scapulocoracoids could be measured, the total of observations was 78. A digital caliper with a 0.01 mm error was used for taking each measurement. Digital photographs were taken from the lateral face of scapulocoracoids for illustrative purposes.
Statistical and mathematical procedures for the LMM analysis followed González-Castro et al. (2012, 2016). The morphometric characters were organized by sexes and species. A normalization technique to scale the data that exhibit allometric growth was employed following Lleonart et al. (2000). Scapulacoracoid length (ScL) was used as the independent variable. ScL0 represents a references value of ScL (23 mm) to which all scapulocoracoids were either reduced or amplified (Lombarte and Lleonart, 1993). After transformation, a principal component analysis (PCA) was performed using MULTIVARIADO®software (Salomón et al., 2004). Finally, principal component scores (PCs) were submitted to cross-validated discriminant analysis (DA) using SPSS®vers. 13.0 (Nie et al., 2004), in order to build a predictive model of group membership based on the observed characteristics of each case. This procedure generates a set of discriminant functions based on linear combinations of the predictor variables that provide the best discrimination between groups.
Scapulocoracoid Morphological Analysis
Morphological variables were taken following McEachran and Compagno (1979). The variables included number of postdorsal and postventral fenestrae, and were taken in both left and right sides of scapulocoradoids. Therefore the total of observations arise to 188. Morphological variation was quantitatively and qualitatively evaluated by comparing number, shape and arrangement of posterior fenestrae, both dorsal and ventral. The number of fenestrae was analyzed through a Mann–Whitney test. Statistical analyses were performed using Statistica 7.0 (Stat. Soft. Inc). For illustrative purposes, digital photographs were taken from the lateral face of scapulocoracoids.
Morphological and Morphometric Analyses of P. parvacauda
To assess the validity of P. parvacauda, the holotype was examined. The specimen is stored at Zoologisches Museum Hamburg [ZMH 25234 (ex ISH 1671-1966 ISH Aussenstelle Ichthyologie des Instituts für Seefischerei)] Germany. Morphological features as well as body morphometric LMM of the specimen were taken.
For comparison purposes, 12 morphometric variables LMM were selected and measured on dorsal and ventral sides of specimens of P. normani (n = 81), P. rudis (n = 34) and P. lentiginosa (n = 35) following Last et al. (2008), and compared to those of P. parvacauda. The variables were: DW, disc length, snout length, preoral length, prenasal length, orbit diameter, interorbital distance, mouth width, distance between first gill openings, distance between fifth openings, distance snout to cloaca, and distance cloaca to caudal tip. As in scapulocoracoid analysis, a normalization technique to scale the data that exhibit allometric growth was employed following Lleonart et al. (2000). In this case, DW was used as the independent variable and DW0 represents a reference value of DW (230 mm) to which all specimens were either reduced or amplified. Even though TL was also measured in all individuals, this variable was removed from the analysis because of the damage tail of the holotype of P. parvacauda.
Results and Discussion
Scapulocoracoid Morphometric Analysis
A sexual dimorphism has been observed in the three Psammobatis species analyzed, according to the original descriptions (McEachran, 1983), being the scapulocoracoids of females more rectangular than those of males, which were more triangular shaped. The PCA of the correlation matrix of LMM of scapulocoracoid, generated by the normalization procedure, produced 2 eigenvalues of >1 (data not shown) and 8 PCs. Correlations between variables and components >0.59 were considered significant (data not shown). The PCA based on LMM allowed a clear differentiation along the PC1 of the females from the males of the three species analyzed (Figure 1). Males showed higher loadings for pre-mesocondyle, greatest height, height of rear corner, and pre-dorsal fenestrae height, whereas higher loadings of variables, post-mesocondyle, and post-dorsal fenestrae length characterized females of the three species analyzed (data not shown). Moreover, the characterization of the species by sexes was only partial, because some overlap between them was detected (Figure 1). The DA on the variations in the 78 individuals of Psammobatis classified by sexes and species were explained by 5 canonical discrimination functions, of which the first 2 explained 93.4% (63.3 and 30.1%, respectively) of the total variance in the data, (Wilks’ lambda = 0.076, p < 0.000). The DA correctly classified 74.4% of the original grouped cases, whereas the cross-validated analysis correctly classified only the 59.4% of the fishes according to their LMM of scapulocoracoid. Moreover, the cross-validated analysis showed that misclassifications varied from 5.3 to 33.3%, depending on the group analyzed (Table 1A). Again, the sexual dimorphism was evidenced as a variable equal, or even stronger than species influence in the group discrimination.
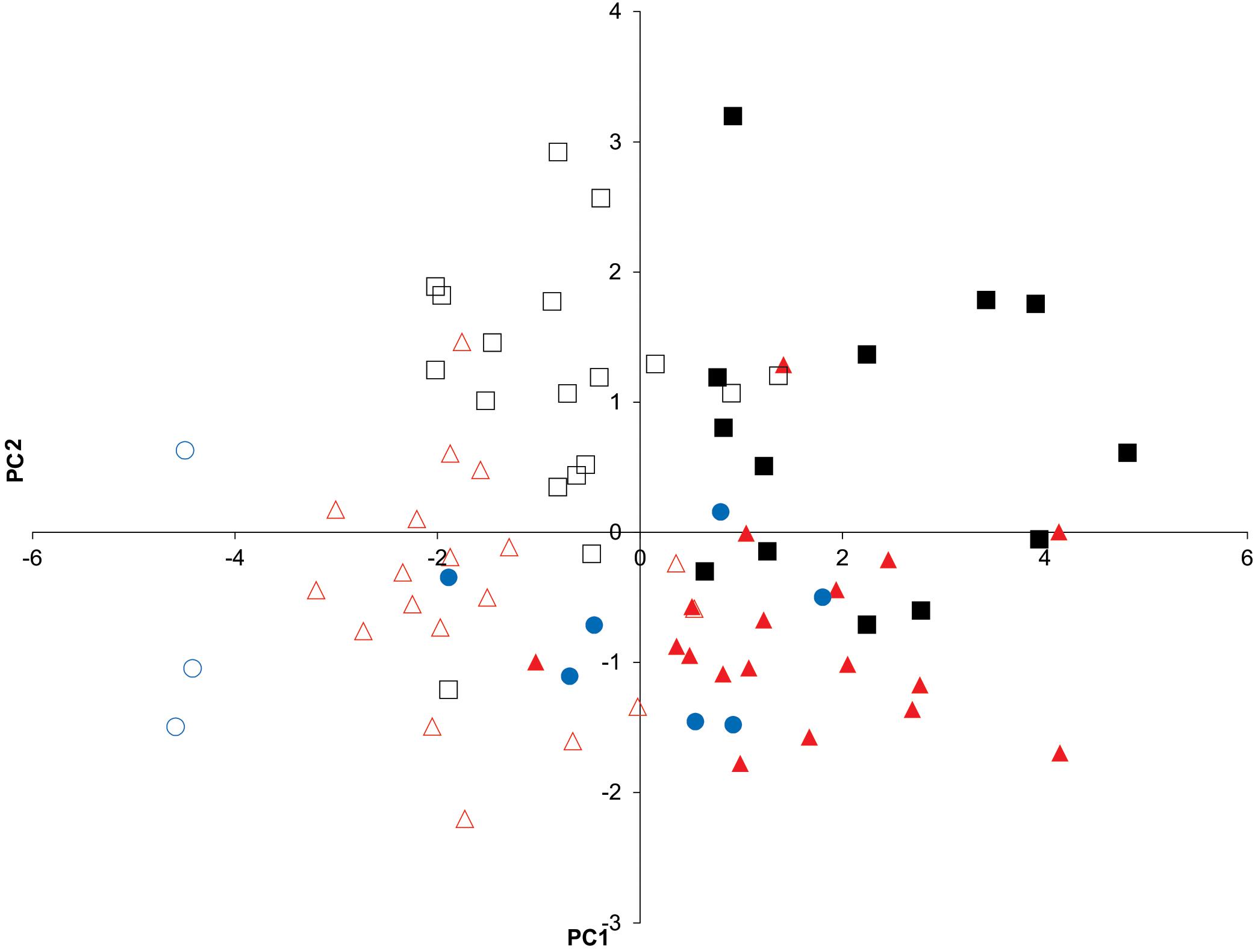
Figure 1. Principal component (PC) analysis (PC1 vs. PC2) based on linear morphometric measurements of scapulocoracoids. The 1st 2 PCs explained 71.3% of the variance in the data. Psammobatis rudis males (black squares); P. rudis females (white squares); P. normani males (red triangles); P. normani females (white triangles); P. lentiginosa males (black circles); and P. lentiginosa females (white circles).
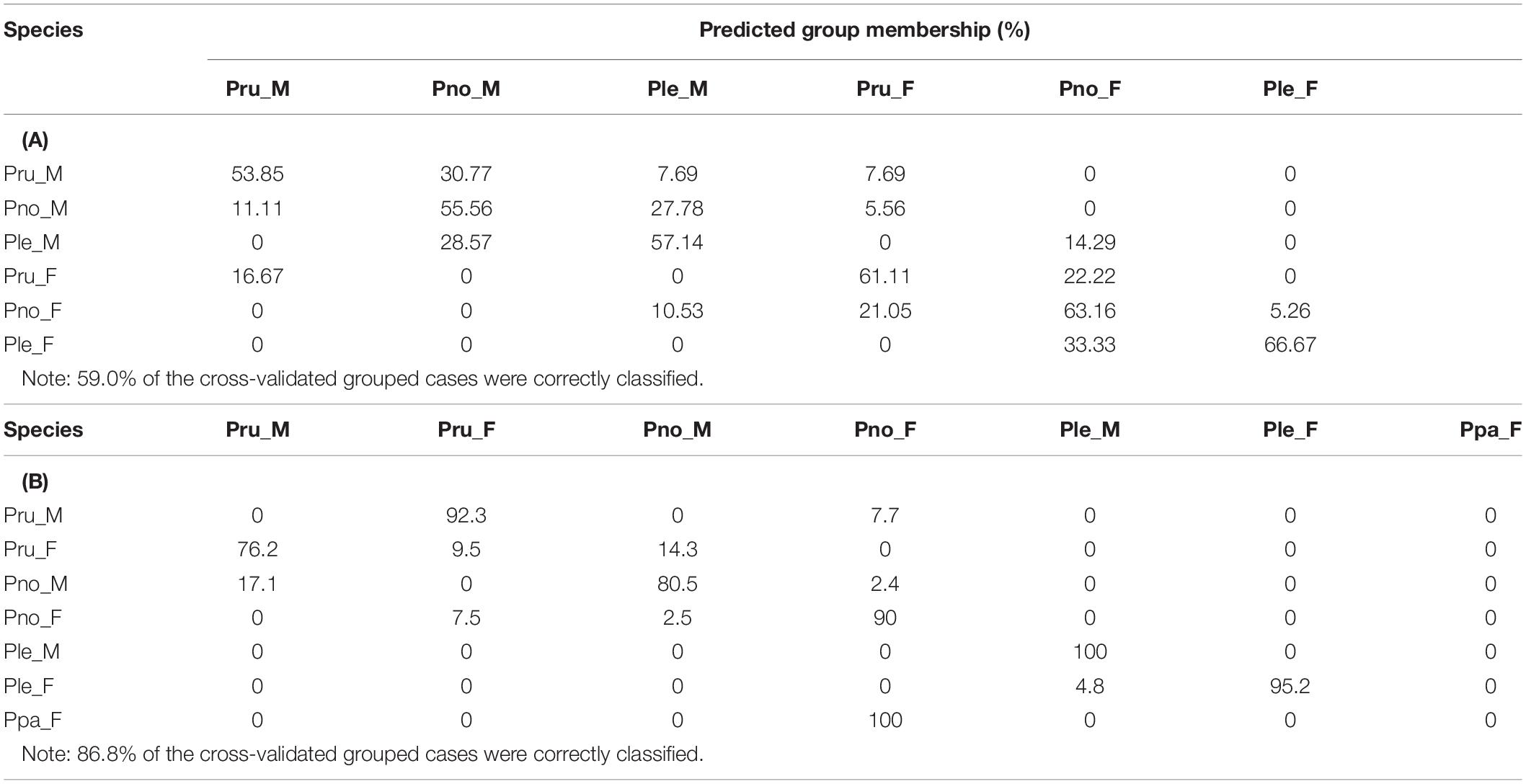
Table 1. Percent values of the cross-validated discriminant analysis, based on the principal component (PC) scores of: (A) Scapulocoracoid linear morphometric measurements, (B) Body linear morphometric. Group codes are: Pru = Psammobatis rudis, Pno = P. normani, Ple = P. lentiginosa, Ppa = P. parvacauda, M = males, F = females.
Scapulocoracoids Morphological Analysis
The number of pre-dorsal and pre-ventral fenestra observed were identical to those described by McEachran (1983). However, a high variability in the number and arrangement of post-dorsal and post-ventral fenestra (pdf and pvf, respectively) was observed, in contrast with original descriptions. This variation was also shown when comparing left and right sides. Both, in P. rudis and P. normani, about 16% of specimens showed differences in the number of postdorsal or postventral fenestrae on each side, and 31% in P. lentiginosa.
Psammobatis rudis showed no differences between males and females in number of pdf and pvf. On the contrary, P. normani showed sexual differences in number of left pvf (p = 0.013). On the other hand, P. lentiginosa showed differences between males and females in both, number of pdf (p = 0.015 left side, p = 0.006 right side), and pvf (p = 0.003 left side, p < 0.001).
Overall, no differences in the number of pdf between males of the three species were observed. Similarly, no differences in the number of pvf between males of P. rudis and P. normani were observed. On the contrary, females of P. rudis presented more pvf than those of P. normani (p = 0.015 left side, p = 0.035 right side). On the other hand, males of P. lentiginosa presented more pvf than those of P. rudis (p < 0.001 left side, p = 0.013 right side). Similarly, females of P. lentiginosa had more pdf (p = 0.004 left side, p < 0.001 right side) and pvf (p < 0.001 left side, p = 0.013 right side) than those of P. rudis. Finally, males of P. lentiginosa had more pvf (p = 0.001 left side, p = 0.007 right side) than those of P. normani, and females of P. lentiginosa presented more pdf (p = 0.003 left side), and pvf (p < 0.001 left side, p < 0.001 right side) than those of P. normani.
Regarding arrangement of fenestrae, in males of P. rudis, three different morphotypes were observed. Twenty-three percent of the samples were identical to those reported by McEachran (1983), (i.e., with one pdf and two pvf), 73% of the samples possessed 1 pdf and 1 pvf, whereas 4% had 1 pdf and 3 pvf (Figure 2A). In females of P. rudis two morphotypes were observed, 67% were coincident with original description, (i.e., with one pdf and one pvf), and 33% presented 2 pvf (Figure 2A).
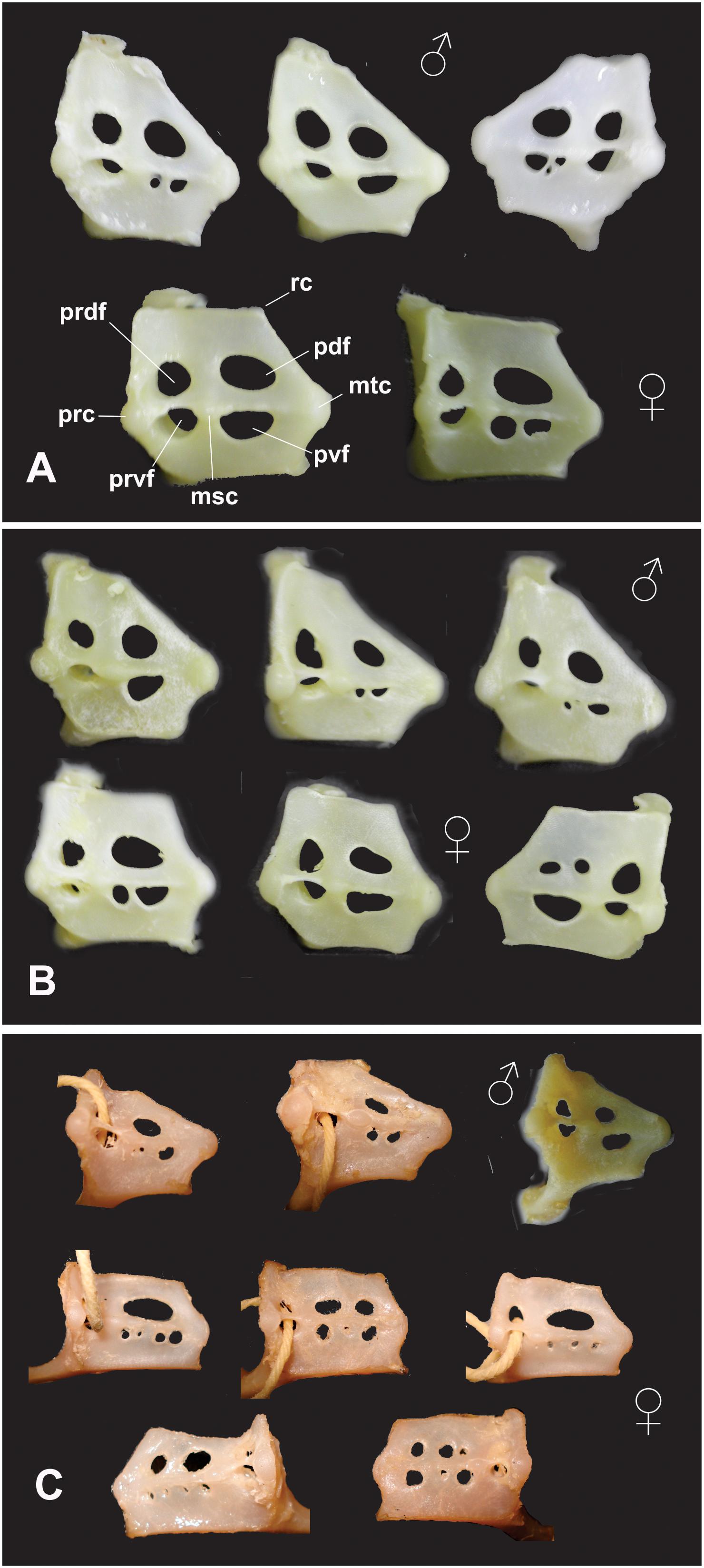
Figure 2. Scapulocoracoid morphotypes observed in 3 species of Psammobatis: (A) P. rudis (above males, below females); (B) P. normani (above males, below females); and (C) P. lentiginosa (above males, below females. Msc = mesocondyle, mtc = metacondyle, pdf = postdorsal foramina or fenestra, prc = procondyle, prdf = predorsal fenestra, prvf = preventral fenestra, pvf = postventral foramina or fenestra, and rc = rear corner.
In males of P. normani, three morphotypes were observed, 72% of the samples were identical to those described by McEachran (1983), having one pdf and one pvf, but 22% possessed 2 pvf and the remaining 6% possessed 3 pvf (two bigger and between them a third little foramina; Figure 2B). In females of P. normani three morphotypes were also observed. 71% of the samples were identical to those described by McEachran (1983), 21% possessed 1 pvf, and 8% (3 specimens) possessed two pdf and one pvf (Figure 2B).
In males of P. lentiginosa three morphotypes were observed, 27% of the specimens are coincident with original descriptions made by McEachran (1983), with one pdf and three pvf, whereas 58% possessed 2 pvf, and 15% had only one pvf (Figure 2C). Finally, in females of P. lentiginosa at least 5 fenestrae arrangement patterns were observed, and only 20% of the samples were coincident with original descriptions, with one large pdf and 4 pvf. The remaining morphotypes were distributed as follows: 38% has one large pdf and 3 pvf, 27% had 2 pdf and 3 pvf, 12% possessed 2 pdf and 4 pvf, and 4% possessed 3 pdf and 3 pvf (Figure 2C).
The aforementioned results showed a high variability in fenestrae arrangement at intraspecific level at least within the genus Psammobatis, similar to what have been observed in Sympterygia spp. (Jurado et al., 2017). Both results call into question the conservative character of this structure and also indicate the lack of taxonomic value, at least in this skates’ genera. In this respect, one of the morphotypes observed in females of P. normani (with two pdf and one pvf, Figure 2B) was identical to the one reported in P. parvacauda by McEachran (1983) who stated “scapulocoracoids of P. parvacauda are derived in possessing: two postdorsal foramina and an expanded postventral fenestra.”
Morphological and Morphometric Analysis of Psammobatis spp.
The morphometric analysis of the holotype of P. parvacauda and specimens of P. normani, P. rudis, and P. lentiginosa (present study) yielded the following results: the PCA of the correlation matrix of LMM of body, generated by the normalization procedure, produced 3 eigenvalues of >1 (data not shown) and 11 PCs. Correlations between variables and components of >0.59 were considered significant (data not shown). As in the scapulocoracoid morphometric analysis, differences between sexes but also species were recorded. Notably, the PCA showed that P. parvacauda (a female) grouped with P. normani females in the PC1-PC2 plot, denoting higher loadings of distance between first gill openings and distance between fifth openings, variables. The characterization of the species by sexes was only partial, because some overlap between them was detected (Figure 3). The DA on the variations in the 151 individuals of Psammobatis classified by sexes and species were explained by 6 canonical discrimination functions, of which the first two explained 84.4% (58.6 and 25.8%, respectively) of the total variance in the data, (Wilks’ lambda = 0.007, p < 0.000). The DA correctly classified 94.7% of the original grouped cases, whereas the cross-validated analysis correctly classified 86.8% of the fishes according to their LMM of body. Moreover, the cross-validated analysis showed that the specimen of P. parvacauda was classified by cross validation analysis in the group of P. normani females (Table 1B).
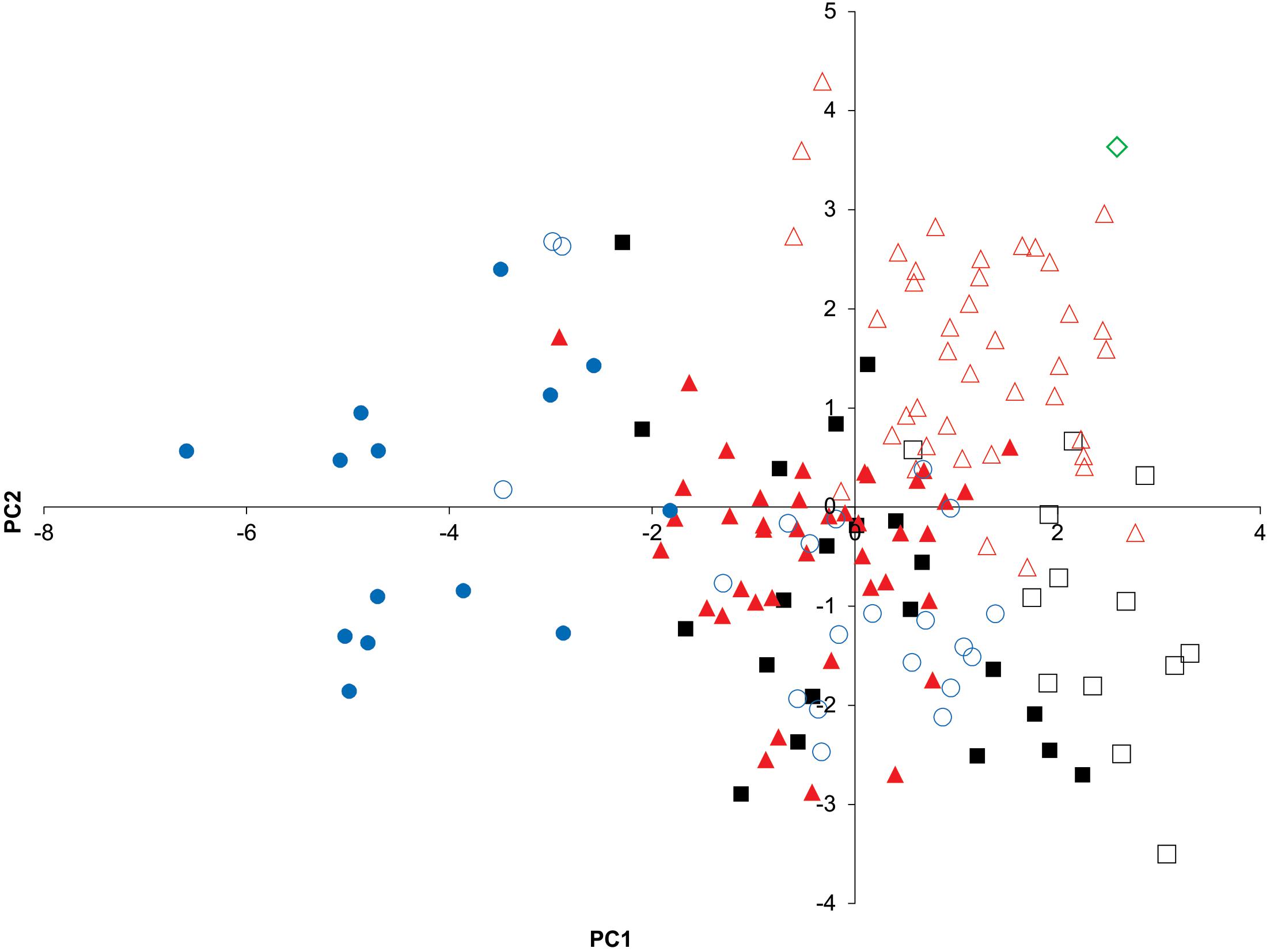
Figure 3. Principal component (PC) analysis (PC1 vs. PC2) based on linear morphometric measurements of body. The 1st 2 PCs explained 55.6% of the variance in the data. Psammobatis rudis males (black squares); P. rudis females (white squares); P. normani males (red triangles); P. normani females (white triangles); P. lentiginosa males (blue circles); and P. lentiginosa females (white circles). Holotype of P. parvacauda (white diamond).
Is Psammobatis parvacauda a Valid Species?
Psammobatis parvacauda was described based on a single female collected around Malvinas/Falkland Islands (McEachran, 1983). According to the author, “P. parvacauda most closely resembles P. rudis and P. scobina, but can be distinguished from the latter by morphometrics, in addition to skeletal structures. In P. parvacauda the preorbital length is less than the preoral length (preorbital snout length is equal to or greater than preoral snout length in P. rudis), the tail length is less than the distance from tip of the snout to the center of the cloaca (the tail length is greater than the distance from the tip of the snout to the center of the cloaca in P. rudis and P. normani, and in all other known species of Psammobatis).” McEachran (1983) also reported 37 tooth rows in the upper jaw and described its spinulation pattern: two suprascapular and two scapular thorns, not forming distinct triangular patch; tail with three irregular rows of thorns on dorsal and dorsolateral surfaces. Finally, coloration was described as dorsal surface tan and scattered with small, faint, white blotches. However, none of these features are diagnostic for P. parvacauda, indeed, the only character used in the key of species provided by McEachran (1983, p. 50) was the length of the tail.
Regarding skeletal structures, as no males are known from this species, the claspers features and their components are obviously unknown. The neurocranium and the scapulocoracoid are the only skeletal structure available for this species. As noticed by McEachran (1983, p. 42) neurocrania of P. parvacauda and P. normani are similar, but, the scapulocoracoid of P. parvacauda is distinct from the other Psammobatis species in possessing two pdf and an expanded pvf (McEachran, 1983). However, as previously demonstrated, the scapulocoracoid in Psammobatis species is highly variable not only among species but also within species. Moreover, as previously indicated the scapulocoracoid pattern of P. parvacauda was observed in three specimens of P. normani (present study). Interestingly, the spinulation pattern and morphometrics of these specimens were in accordance with original descriptions and the remaining P. normani analyzed in the present study. Particularly, they did not have a short tail (length from tip of snout to center of cloaca was similar to length from center of cloaca to tail tip), and eye diameter was not large (4.4% of TL). Therefore, there is no skeletal structures with exclusive characteristic of P. parvacauda.
With respect to P. parvacauda, Weigmann (2016) stated “Possibly an aberrant specimen of another Psammobatis species as the postdorsal tail section of the holotype is apparently incomplete and, despite precise registrations of catches in this area, only one further specimen has been found (Stehmann, 2014, pers. comm.) Nevertheless, the apparently undamaged tail of the second specimen and the very large eyes of the species indicate that the species indeed might be valid despite its close similarity to P. rudis and P. scobina (Weigmann, unpubl. data).” In the present work, the holotype was examined by one of the co-authors and in fact, the tail section is incomplete (the caudal fin base measured 1.8 mm). Indeed, McEachran (1983) mentioned that epicordal lobe of caudal fin was barely developed in P. parvacauda. There is no reliable record of the second specimen reported, because it was not published. Therefore, no information regarding this specimen is available (photos, measurements, meristic, and genetics), only a personal communication by Stehmann to Weigmann is offered. Weigmann highlighted “the very large eyes of the species.” However, McEachran (1983) did not regard this feature as a diagnostic characteristic for the species. He only reported its value as 5.6% of TL (TL 347 mm, OD 19.4 mm).
In addition, Last et al. (2016b) pointed out that “P. parvacauda is very poorly known and its distinction from some sandskates has been questioned. However, based on its relatively large eyes (5.6–6.7% of TL), compared to other members of the genus, we provisionally consider it to be valid.” The authors also indicated that the species is distributed in northeast Malvinas/Falkland Islands (type locality) and in northern Patagonian waters (130 m). However, no record of the latter specimen has been published. On the other hand, the features observed in the “new” sample are not totally in accordance with the holotype. Indeed, Last et al. (2016b) mentioned “Tail rather slender and short, tapering strongly, its length ∼0.9–1 times precloacal length.” Therefore, in the “new specimen” tail length is equal to precloacal length and no shorter as was diagnosed in the first description made by McEachran (1983). Regarding the eye diameter, the values reported by McEachran (1983) for the holotype was 5.6% of TL, slightly higher than those reported for the holotype of P. normani (5.4% TL; McEachran, 1983, table 3, p. 53), whereas for P. rudis and P. scobina the author provided only the mean values and those were lower than 4.5%. From 152 specimens of P. normani analyzed, Mabragaña (2007) showed that this characteristic could reach 5.6% of TL, a value observed in the holotype of P. parvacauda. Therefore, there would be no diagnostic characters for the species.
It is suspicious that after almost 40 years from its original description, no novel specimens from this species were reported so far, and a likely second specimen quoted by Last et al. (2016b) and Weigmann (2016) would be another report of the species, even more if we consider that species from this genus are conspicuous and abundant in the Patagonian shelf (Cousseau et al., 2007). Furthermore, Mabragaña (2007) in spite of revising more than 600 specimens of its congeners P. rudis and P. normani along their entire distribution in the SWA, only found males with clasper morphology typical of P. rudis or P. normani.
The morphological features of the holotype of P. parvacauda, specially the spinulation pattern of tail with 3 irregular rows of tail thorns and no dermal denticles or minute spines on dorsolateral aspects of tail, resemble that of P. normani more than P. rudis (McEachran, 1983; Mabragaña, 2007). In the same way, the number of tooth rows on upper jaw (37) is within the range of both P. rudis (31–37, mean = 34, and n = 34) and P. normani (33–44, mean = 39, and n = 46; Mabragaña, 2007). Orbital nuchal and scapular thorns are also large and visible as in P. normani, and similarly the dorsal surface of disk is not covered with coarse dermal denticles as it happens in P. rudis (McEachran, 1983; Mabragaña, 2007). Also, as was already demonstrated, the specimen of P. parvacauda was statistically classified (cross-validated analysis of DA) in the P. normani female group. The external similarities between P. parvacauda and P. normani, as well as the presence of that particular fenestrae arrangement in the scapulocoracoid of some P. normani similar to that reported for P. parvacauda, the lack of a particular diagnostic character in the holotype, and the absence of a reported male for this species, strongly suggest that P. parvacauda is a junior synonym of P. normani. As Weigmann (2016) stated, this specimen surely constitutes “an aberrant female of other Psammobatis species.” Both, P. rudis and P. normani are simpatrically distributed in the SWA, and although P. rudis is more abundant in the southern south-west Atlantic Ocean, P. normani inhabits also in Patagonian waters (Mabragaña and Cousseau, 2004; Mabragaña and Giberto, 2007).
In conclusion our data strongly indicate that P. parvacauda and P. normani correspond to the same species. Given that both species were described in the same moment, but considering that description of P parvacauda is possible based in an aberrant specimen, P. normani McEachran should be the senior synonym.
Data Availability Statement
The raw data supporting the conclusions of this article are available on Figshare at doi: 10.6084/m9.figshare.12142302.v1.
Ethics Statement
Ethical review and approval was not required for the animal study because It was based on specimen collected with bottom trawls, in research cruises conducted more than 10 years ago, and specimens were landed already dead.
Author Contributions
EM conceived the study. EM, VG, DV, and JD process the sample material. MG-C performed the statistical analyses. EM and MG-C interpreted the results and wrote the first draft of the manuscript. All authors wrote and revised the final draft of the manuscript.
Funding
The work was partially supported by Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET, PIP 0339), MINCYT (PICT-2018-0790), and Facultad de Ciencias Exactas y Naturales, Universidad Nacional de Mar del Plata (EXA767/16 and EXA 867/18).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank the skippers, crews and scientists of the R.V. O. Balda and R.V. E. L. Holmberg from the Instituto Nacional de Investigación y Desarrollo Pesquero for their assistance in the collection of samples. We also thank Patrice Pruvost and Romain Causse from Muséum national d’Histoire naturelle (Paris) for their effort and assistance in the acquisition of the holotype of P. parvacauda.
References
Barbini, S. A., and Lucifora, L. O. (2012). Ontogenetic diet shifts and food partitioning between two small sympatric skates (Chondrichthyes, Rajidae) in the Southwestern Atlantic. Mar. Freshw. Res. 63, 905–913.
Braccini, J. M., and Chiaramonte, G. E. (2002a). Reproductive biology of Psammobatis extenta. J. Fish. Biol. 61, 272–288.
Braccini, J. M., and Chiaramonte, G. E. (2002b). Intraspecific variation in the external morphology of the sand skate. J. Fish. Biol. 61, 959–972.
Braccini, J. M., and Perez, J. E. (2005). Feeding habits of the sandskate Psammobatis extenta (Garman, 1913): sources of variation in dietary composition. Mar. Freshw. Res. 56, 395–403.
Compagno, L. J. V. (1999). “Endoskeleton,” in Sharks, Skates and Rays. The Biology of Elasmobranch Fishes, Chap. 3, ed. W. C. Hamlett, (Maryland: Johns Hopkins Press), 69–92.
Concha, F., Hernández, S., and Oddone, M. C. (2009). Egg capsules of the raspthorn sandskate, Psammobatis scobina (Philippi, 1857)(Rajiformes, Rajidae). Rev. Biol. Mar. Oceanogr. 44, 253–256.
Cousseau, M. B., Figueroa, D. E., Díaz de Astarloa, J. M., Mabragaña, E., and Lucifora, L. O. (2007). Rayas, Chuchos y Otros Batoideos del Atlántico Sudoccidental. Buenos Aires: INIDEP.
de Carvalho, M. R., and Figueiredo, J. L. (1994). Psammobatis extenta (Garman, 1913): a senior synonym of Psammobatis glansdissimilis McEachran, 1983(Chondrichthyes, Rajidae). Copeia 1994, 1029–1033.
de Carvalho, M. R., Gomes, U. L., and Gadig, O. B. F. (2005). Description of a new species of skate of the genus Malacoraja Stehmann, 1970: the first species from the southwestern Atlantic Ocean, with notes on generic monophyly and composition (Chondrichthyes: Rajidae). Neotrop. Ichthyol. 3, 239–258. doi: 10.1590/S1679-62252005000200002
González-Castro, M., Ibáñez, A. L., Heras, S., Roldán, M. I., and Cousseau, M. B. (2012). Assesment of lineal versus landmarks-based morphometry for discriminating species of Mugilidae (Actinopterygii). Zool. Stud. 51, 1515–1528.
González-Castro, M., Rosso, J. J., Mabragaña, E., and Díaz de Astarloa, J. M. (2016). Surfing among species, populations and morphotypes: inferring boundaries between two species of new world silversides (Atherinopsidae). CR Biol. 339, 10–23. doi: 10.1016/j.crvi.2015.11.004
Hulley, P. A. (1972). The origin, interrelationships and distribution of southern African Rajidae (Chondrichthyes, Batoidei). Ann. S. Afr. Mus. 60, 1–103.
Irigoitia, M. M., Braicovich, P. E., Rossin, M. A., Canel, D., Levy, E., Farber, M. D., et al. (2019). Diversity of Empruthotrema Johnston and Tiegs, 1992 parasitizing batoids (Chondrichthyes: Rajiformes and Myliobatiformes) from the Southwest Atlantic Ocean, with description of three new species. Parasitol. Res. 118, 3113–3127. doi: 10.1007/s00436-019-06456-x
Jeong, C. H., and Nakabo, T. (2009). Hongeo, a new skate genus (Chondrichthyes: Rajidae), with redescription of the type species. Ichthyol. Res. 56, 140–155. doi: 10.1007/s10228-008-0083-9
Jurado, C. D., Mabragaña, E., and Díaz de Astarloa, J. M. D. (2017). Morphological variation in a conservative structure: the scapulocoracoids in Sympterygia acuta Garman, 1837 and Sympterygia bonapartii Müller & Henle, 1841 (Chondrichthyes: Rajidae). Zootaxa 4318, 157–166.
Last, P. R., and Gledhill, D. C. (2007). The Maugean Skate, Zearaja maugeana sp. nov. (Rajiformes: Rajidae) –A micro-endemic, gondwanan relict from Tasmanian estuaries. Zootaxa 1494, 45–65.
Last, P. R., and McEachran, J. D. (2006). Notoraja hirticauda, a new species of skate (Chondrichthyes: Rajoidei) from the south-eastern Indian Ocean. Mem. Mus. Vic. 63, 65–75. doi: 10.24199/j.mmv.2006.63.9
Last, P. R., Stehmann, M. F. W., Séret, B., and Weigmann, S. (2016a). “Skates, family rajidae,” in Rays of the World, eds P. R. Last, W. T. White, M. R. Carvalho, B. de Séret, M. F. W. Stehmann, and G. J. P. Naylor, (Melbourne: CSIRO Publishing), 204–363.
Last, P. R., Stehmann, M. F. W., Séret, B., and Weigmann, S. (2016b). “Softnose skates, family arhynchobatidae,” in Rays of the World, eds P. R. Last, W. T. White, M. R. Carvalho, B. de Séret, M. F. W. Stehmann, and G. J. P. Naylor, (Melbourne: CSIRO Publishing), 364–472.
Last, P. R., White, W. T., Pogonoski, J. J., and Gledhill, D. C. (2008). “New Australian skates (Batoidea: Rajoidei) – background and methodology,” in Description of New Australian Skates (Batoidea: Rajoidei), eds P. R. Last, W. T. White, J. J. Pogonoski, and D. C. Gledhill (Canberra: CSIRO Marine and Atmospheric Research), 1–8.
Lleonart, J., Salat, G., and Torres, J. (2000). Removing allometrics effects of body size in morphological analysis. J. Theor. Biol. 205, 85–93. doi: 10.1006/jtbi.2000.2043
Lombarte, A., and Lleonart, J. (1993). Otolith size changes related with body growth, habitat depth and temperature. Environ. Biol. Fish. 37, 297–306.
Mabragaña, E. (2007). Las Rayas del Género Psammobatis de la Plataforma Argentina: Biología y Ecología. Ph.D. Thesis, Universidad Nacional de Mar del Plata, Mar del Plata.
Mabragaña, E., and Cousseau, M. B. (2004). Reproductive biology of two sympatric skates in the south-west Atlantic: Psammobatis rudis and Psammobatis normani. J. Fish. Biol. 65, 559–573.
Mabragaña, E., Figueroa, D. E., Scenna, L. B., Díaz de Astarloa, J. M., Colonello, J. H., and Delpiani, G. (2011). Chondrichthyan egg cases from the south-west Atlantic Ocean. J. Fish. Biol. 79, 1261–1290. doi: 10.1111/j.1095-8649.2011.03111.x
Mabragaña, E., and Giberto, D. A. (2007). Feeding ecology and abundance of two sympatric skates, the shortfin sand skate Psammobatis normani McEachran, and the smallthorn sand skate P. rudis Günther (Chondrichthyes, Rajidae), in the southwest Atlantic. ICES J. Mar. Sci. 64, 1017–1027.
Mabragaña, E., Ibáñez, P., and Cousseau, M. B. (2012). Reproductive biology and abundance of the freckled sandskate Psammobatis lentiginosa McEachran, 1983 in the southwest Atlantic. Mar. Biol. Res. 8, 292–299.
Martins, M. F., and Oddone, M. C. (2017). Reproductive biology of Psammobatis rutrum (Chondrichthyes: Arhynchobatidae) in south Brazil, south-west Atlantic. J. Fish. Biol. 91, 443–459. doi: 10.1111/jfb.13350
McEachran, J. D. (1982). Revision of the South American skate genus Sympterygia (Elasmobranchii, Rajiformes). Copeia 4, 867–890. doi: 10.2307/1444098
McEachran, J. D. (1983). Results of the research cruises of FRV “Walter Herwig” to South America LXI. Revision of the South American skate genus Psammobatis Günther, 1873 (Elasmobranchii, Rajiformes, Rajidae). Arch. Fischereiwiss 34, 23–80.
McEachran, J. D., and Compagno, L. J. V. (1979). A further description of Gurgesiella furvesens with comments on the interrelationships of Gurgesiellidae and Pseudorajidae (Pisces, Rajoidei). Bull. Mar. Sci. 29, 530–553.
McEachran, J. D., and Compagno, L. J. V. (1982). Interrelationships of and within Breviraja based on anatomical structures (Pisces: Rajoidei). Biol. Mar. Sci. 32, 399–425.
McEachran, J. D., and Dunn, K. A. (1998). Phylogenetic analysis of skates, a morphologically conservative clade of elasmobranchs (Chondrichthyes: Rajidae). Copeia 1998, 271–290. doi: 10.2307/1447424
McEachran, J. D., and Last, P. R. (1994). New species of Skate, Notoraja ochroderma, from off Queensland, Australia, with comments on the taxonomic limits of Notoraja (Chondrichthyes, Rajoidei). Copeia 1994, 413–421. doi: 10.2307/1446989
McEachran, J. D., and Miyake, T. (1990). “Phylogenetic interrelationships of skates: a working hypothesis (Condrichthyes: Rajoidei),” in Elasmobranchs as Living Resources: Advances in the Biology, Ecology, Systematics and the Status of Fisheries, eds H. L. Pratt, S. H. Gruber, and T. Taniuchi, (Silver Spring, MA: NOAA), 285–304.
Perier, R., Estalles, M., Coller, M., and Di Giacomo, E. E. (2011). Reproductive biology of the endemic skate Psammobatis lentiginosa in the San Matías Gulf (south-western Atlantic). J. Mar. Biol. Assoc. U.K. 91, 1165–1173.
Salomón, R., Melo, S., and Winzer, N. (2004). MULTIVARIADO Software. Bahía Blanca: Departamento de Matemática, Univ. Nacional del Sur.
San Martín, M. J., Braccini, J. M., Tamini, L. L., Chiaramonte, G. E., and Perez, J. E. (2007). Temporal and sexual effects in the feeding ecology of the marbled sand skate Psammobatis bergi Marini, 1932. Mar. Biol. 151, 505–513.
San Martín, M. J., Perez, J. E., and Chiaramonte, G. E. (2005). Reproductive biology of the south west Atlantic marbled sand skate Psammobatis bergi Marini, 1932 (Elasmobranchii, Rajidae). J. Appl. Ichthyol. 21, 504–510.
Seidel, R., Blumer, M., Pechriggl, E. J., Lyons, K., Hall, B. K., Fratzl, P., et al. (2017). Calcified cartilage or bone? Collagens in the tessellated endoskeletons of cartilaginous fish (sharks and rays). J. Struct. Biol. 200, 54–71. doi: 10.1016/j.jsb.2017.09.005
Séret, B., Last, P. R., Weigmann, S., and Stehmann, M. F. W. (2016). “Legskates, family Anacanthobatidae,” in Rays of the World, eds P. R. Last, W. T. White, M. R. Carvalho, B. de Séret, M. F. W. Stehmann, and G. J. P. Naylor, (Melbourne: CSIRO Publishing), 494–508.
Stehmann, M. (1970). Vergleichend morphologische und anatomische Untersuchungen zur Neuordnung der Systematik der nordostatlantischen Rajidae (Chondrichthyes, Batoidei). Arch. Fischereiwiss 21, 73–164.
Stehmann, M., and Seret, B. (1983). A new species of deep-water skate, Breviraja africana sp. n. (Pisces, Batoidea, Wajidae), from the Eastern Central Atlantic slope, and remarks on the taxonomic status of Breviraja Bigelow & Schroeder, 1948. B Mus. Natl. Hist. Nat. 5, 903–925.
Vazquez, D. M., Mabragaña, E., Gabbanelli, V., and Díaz de Astarloa, J. M. (2016). Exploring nursery sites for oviparous chondrichthyans in the Southwest Atlantic (36° S–41° S). Mar. Biol. Res. 12, 715–725.
Weigmann, S. (2016). Annotated checklist of the living sharks, batoids and chimaeras (Chondrichthyes) of the world, with a focus on biogeographical diversity. J. Fish. Biol. 88, 837–1037. doi: 10.1111/jfb.13234
Keywords: Rajiformes, Psammobatis, scapulocoracoids, intraspecific polymorphism, P. parvacauda, synonymization
Citation: Mabragaña E, González-Castro M, Gabbanelli V, Vazquez DM and Díaz de Astarloa JM (2020) Polymorphism in Conservative Structures? The Scapulocoracoids in Skates Genus Psammobatis (Chondrichthyes, Arhynchobatidae) and the Validity of P. parvacauda. Front. Mar. Sci. 7:291. doi: 10.3389/fmars.2020.00291
Received: 13 December 2019; Accepted: 14 April 2020;
Published: 20 May 2020.
Edited by:
Alejandra Vanina Volpedo, University of Buenos Aires, ArgentinaReviewed by:
William Driggers, National Marine Fisheries Service (NOAA), United StatesPaul Brickle, South Atlantic Environmental Research Institute, Falkland Islands
Copyright © 2020 Mabragaña, González-Castro, Gabbanelli, Vazquez and Díaz de Astarloa. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: E. Mabragaña, ZW1hYnJhZ2FAbWRwLmVkdS5hcg==