
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Mar. Sci. , 06 November 2018
Sec. Marine Ecosystem Ecology
Volume 5 - 2018 | https://doi.org/10.3389/fmars.2018.00411
This article is part of the Research Topic Harmful Algal Blooms (HABs) in Latin America View all 26 articles
 Oscar Amaya1
Oscar Amaya1 Rebeca Quintanilla1*
Rebeca Quintanilla1* Brian A. Stacy2
Brian A. Stacy2 Marie-Yasmine Dechraoui Bottein3
Marie-Yasmine Dechraoui Bottein3 Leanne Flewelling4
Leanne Flewelling4 Robert Hardy4
Robert Hardy4 Celina Dueñas5Gerardo Ruiz1
Celina Dueñas5Gerardo Ruiz1In late October and early November 2013 and 2017, hundreds of sea turtles were found dead along the Pacific coastline of El Salvador. The dead turtles were in good body condition and did not have any injuries or other major anomalies. In order to determine the role of paralytic shellfish toxins (PST) in this mass mortality, tissue samples, including blood, flipper, liver, kidney, stomach and intestinal contents, of dead green turtles (Chelonia mydas) and olive ridley turtles (Lepidochelys olivacea) were analyzed for PST using a radioactive receptor binding assay, enzyme-linked immunosorbent assay, and high performance liquid chromatography. Highest values of PST were detected in enteric contents in the 2013 event (7,304.1 μg STX eq kg−1) and in gastric contents during the 2017 event (16,165.0 μg STX eq kg−1). During these events, remotely-sensed chlorophyll-a and fluorescence line height imagery revealed anomalies suggestive of algal blooms off the coast of El Salvador. In the 2017 event, Pyrodinium bahamense was observed in samples of gastrointestinal contents from affected sea turtles. Seawater from the region where dead sea turtles were found was also analyzed, but saxitoxin-producing species were found in low abundance (5400 cell/L in 2013 and 672 cell/L in 2017), which may reflect limited sampling. Although threshold levels of toxicity in sea turtle species are not well-characterized, our evidence suggests that these large events were the result of PST-producing algal blooms and that these blooms are a major cause of sea turtle mortality in this region.
Harmful algal blooms (HABs) negatively affect not only human health, but also that of marine ecosystems. These effects include hypoxia/anoxia events, decreased water clarity, and altered feeding behavior and toxicosis of marine fauna (Zingone and Enevoldsen, 2000). Toxin-producing algal blooms are recurrent off the coast of El Salvador and are predominately paralytic shellfish poisoning events, which are produced by several paralytic shellfish toxin (PST)-producing dinoflagellates, such as Pyrodinium bahamense var. compressum, Gymnodinium catenatum, and Alexandrium spp. These events have caused human intoxication and death, as well as mortality of marine fauna (Espinoza et al., 2013). Globally, exposure to PST toxins has been associated with mortality of marine mammals (Geraci et al., 1989; Van Dolah et al., 2003; Lefebvre et al., 2016), sea birds (Shumway et al., 2003; Shearn-Bochsler et al., 2014), estuarine turtles (Hattenrath-Lehmann et al., 2017), and sea turtles (Maclean, 1975; Licea et al., 2008).
Four of the seven sea turtle species found in the world nest or forage in El Salvador and its waters. All species are considered vulnerable, endangered, or critically endangered by the International Union for the Conservation of Nature (IUCN, 2018) Red List. In 2005, over 200 sea turtles, including green turtles (Chelonia mydas) and olive ridley turtles (Lepidochelys olivacea), were found dead along the coast of El Salvador. The mortality event was attributed to PST based on concentrations found in brain samples which were as high as of 6278.0 μg STX eq kg−1 (Licea et al., 2008). Additionally, in 2010 two hawksbill turtles (Eretmochelys imbricata) were found dying after a Pyrodinium bahamense bloom. One of the turtles was found with 1212.5 μg STX eq kg−1 in brain tissue, and both individuals had diarrhea and exhibited erratic swimming, disorientation, and reduced activity (Licea et al., 2013). However, the limited toxin analytical data prevented confident attribution of clinical signs to PSP (Licea et al., 2013).
Two additional large sea turtle mortality events occurred in El Salvador waters in recent years. In early October 2013, the Ministry of Environment and Natural Resources (MARN) received a report from fishermen of dozens of dead sea turtles floating far from the coast, drifting northwest. In a subsequent report, 40 dead sea turtles were found on beaches of La Paz and La Libertad during September and October, and an undetermined number on beaches in Usulután. On October 28th, 2017, an estimated 400 dead sea turtles were found floating approximately 12 miles off the coast of Bahía de Jiquilisco. In the following days, 160 turtles were observed around 7 miles off the coast of Puerto Parada, of which 60% were L. olivacea and 40% were C. mydas. Between November 3rd and 6th, another 47 sea turtles were found floating off the coast of Usulután and La Paz and 25 L. olivacea individuals were detected 10 miles off El Cordoncillo in La Paz. No other dead marine organisms were observed during either event. The turtles were in good nutritional condition based on muscle mass and body fat and did not have any apparent injuries or other abnormalities to explain the cause of death.
PST-producing HABs were again suspected and samples of tissues, gastrointestinal contents, and seawater were collected for biotoxin analysis and phytoplankton identification. In addition, data collected by satellite (MODIS-Aqua) were reviewed for evidence of possible algal blooms. Herein, we report the results of these analyses as evidence that both mortality events were caused by PST exposure, demonstrating that PST-producing algal blooms are a substantial cause of sea turtle mortality in the region.
Tissue samples from C. mydas and L. olivacea were collected by the MARN from different sites along the coast during stranding events in 2013 and 2017 (Figure 1) and analyzed for PST at three different laboratories. In 2013, stomach, intestines, liver and kidneys were taken from 13 C. mydas that stranded in El Amatal-Toluca, San Diego, and El Pimental were analyzed by Laboratorio de Toxinas Marinas at the University of El Salvador (LABTOX-UES); 24 samples consisting of brain, enteric content, kidney, liver, and salt gland were analyzed by the International Atomic Energy Agency (IAEA) Environment Laboratories in Monaco; and 7 samples consisting of enteric contents, liver, kidney, and lung from 5 C. mydas that stranded in El Amatal and El Pimental and one L. olivacea that stranded in Barra de Santiago, were analyzed by the Florida Fish and Wildlife Conservation Commission, Fish and Wildlife Research Institute (FWRI). CITES export and import permits were obtained for all samples exported to the U.S.
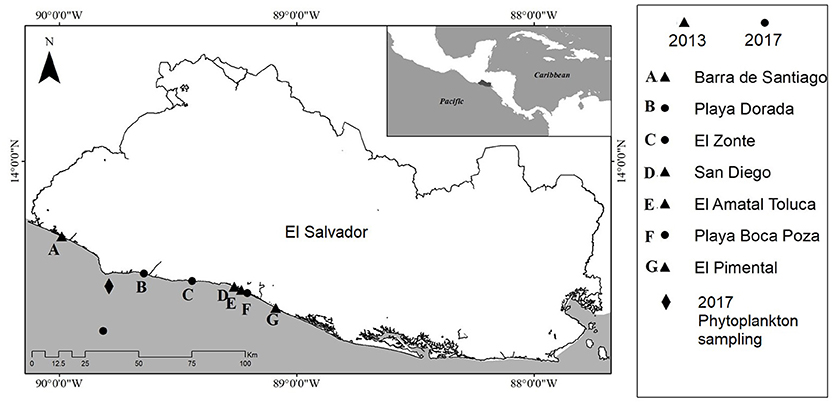
Figure 1. Location of sea turtle strandings in 2013 and 2017 and phytoplankton sampling locations in 2017.
In the 2017 event, 25 samples consisting of whole blood, serum, soft tissue from flippers from 23 C. mydas, and liver and intestinal contents from one L. olivacea were analyzed by LABTOX-UES. Additionally, 28 samples consisting of sea turtle liver, stomach contents, and enteric contents from 14 C. mydas and one L. olivacea were analyzed by FWRI.
Samples were analyzed for PST at LABTOX-UES and IAEA Environment Laboratories using a receptor binding assay (RBA) that is the AOAC official method of analysis (OMA-2011-27) (Amaya et al., 2012; Van Dolah et al., 2012; Dechraoui Bottein and Clausing, 2017). Briefly, the RBA measures competition between a radiolabeled 3H-STX and the PSTs present in samples for binding to voltage gated sodium channels in brain membrane preparations. The quantification is obtained against a standard curve generated using increasing concentrations of unlabeled STX reference material (S. Hall, United States Food and Drug Administration/Center for Food Safety and Applied Nutrition, Washington, DC). Toxin concentration is reported as micrograms of STX equivalents per kilogram of sample (μg STX eq. kg−1). The assay format described in the present study provides quantitative determination of the compound toxicity of PST in turtle extracts using a MicroBeta Trilux 1450 LSC PerkinElmer with 96-well microplate. For analyses conducted by LABTOX-UES, detection limits were 150 μg STX eq. kg−1 in 2013 and 70 μg STX eq. kg−1 in 2017.
FWRI analyzed samples for PST using two methods, enzyme-linked immunosorbent assay (ELISA) and high performance liquid chromatography (HPLC). Some changes in laboratory methodology occurred between 2013 and 2017. Frozen samples were completely thawed and homogenized before sub-sampling. To extract PST, 0.1M HCl was added at a ratio of 1:1 (w:v) in 2013 or 1:10 (w:v) in 2017. The mixture was adjusted to pH 2.5-4, boiled for 5 min in a water bath, and then centrifuged at 3,000 × g for 20 min. The supernatant was retained.
Samples from the 2013 event were analyzed for PST using HPLC with pre-column oxidation and fluorescence detection (Lawrence and Niedzwiadek, 2001) only. The HPLC system consisted of a Shimadzu (Tokyo, Japan) chromatograph equipped with an SCL-10A VP controller, LC-10 AD pump, SIL-10AF autosampler, CTO-10AS VP column oven, and RF-10A XL fluorescence detector. Sample clean-up and PST separations were performed as described in Lawrence and Niedzwiadek (2001). Certified reference standards of STX, NEO, dcSTX, B1, GTX 1/4, GTX 2/3, dcGTX-2/3, and C1/2 (National Research Council, Canada) were used for instrument calibration. Detection limits varied for each congener, ranging between 6 and 18 ng mL−1 extract and 12–36 μg kg−1 tissue. Total HPLC-FL results are expressed as STX equivalents calculated using experimentally derived toxicity factors for each congener (Oshima, 1995).
In 2017, sample extracts were first screened using the Abraxis Saxitoxin (PSP) ELISA. All extracts were diluted with provided sample diluent to 0.01 g tissue equivalent per ml or less prior to loading on the plate. The Abraxis Saxitoxin (PSP) ELISA is a direct competitive ELISA specific to STX, with recognition of other PST to varying degrees. All samples and standards were run in duplicate wells, and the plate was analyzed at a wavelength of 450 nm on a BioTek μQuant™ microplate spectrophotometer. The kit is calibrated using STX, and results are expressed as STX equivalents. The limit of detection for the ELISA as performed was 2 μg STX eq. kg−1. Toxin confirmation and characterization were obtained for a subset of sample extracts using HPLC-FL as described above. Liver samples selected for HPLC confirmation were re-extracted at a ratio of 1:1 (w:v) to yield a more concentrated sample extract.
Sampling campaigns were carried out to monitor toxic microalgae during both mortality events. In 2013, samples were taken in La Libertad and Acajutla in October 15th and 16th. In 2017, samples were taken on November 7th 2017, at Los Cóbanos (Figure 1). This site was selected because the Ministry of the Environment and Natural Resources received reports of sea turtle stranding in the central and west coast of El Salvador, after the dead sea turtles were located floating in Bahía de Jiquilisco. Water samples were collected at five points along a 15 nautical mile transect perpendicular to Los Cóbanos coastline. Samples for phytoplankton quantification were collected using a Niskin bottle and using a phytoplankton net.
Phytoplankton abundance was estimated using an inverted microscope, following the Utermöhl method (Utermöhl, 1958). Additionally, in the 2017 event, intestinal contents of one L. olivacea and one C. mydas were screened for the presence of toxic microalgae using an inverted microscope by LABTOX-UES; and two gastric and two enteric contents were examined by FWRI using light microscopy.
We examined corresponding chlorophyll-a (Chl) and normalized fluorescence line height (NFLH) imagery to visualize the approximate location and spatiotemporal extent of a potential algae bloom. The NFLH imagery provide an alternative source of imagery that is capable of detecting blooms but is less sensitive to interference by non-algal substances common in coastal waters (e.g., dissolved organic matter; Hu et al., 2005).
During both events, PST were detected in most sample types from all sea turtles. Out of the 75 tissue samples analyzed, 63 were positive for PST and 12 were below the detection limits (Figure 2). Those samples that were below detection limits included heart and fat, and well as blood, serum, and flipper soft tissue sampled during 2017.
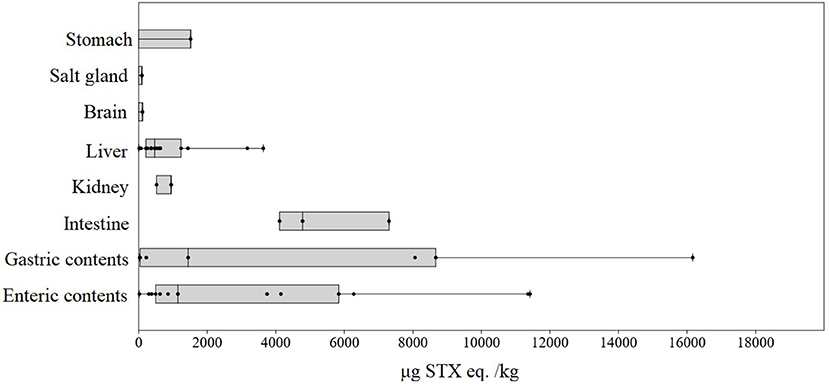
Figure 2. PST concentration (μg STX eq. kg−1) in sea turtles (C. mydas and L. olivacea) per sample type from both 2013 and 2017 mortality events.
Highest PST concentrations were detected in enteric contents in the 2013 event (7304.1 μg STX eq. kg−1 using RBA method) and in gastric contents during the 2017 event (16165.0 μg STX eq. kg−1 using ELISA) (Table 1). Both values were found in C. mydas individuals. In general, PST concentrations found in 2017 were higher than those found in 2013. PST concentrations in tissues from brain, lung and salt glands were generally lower, ranging from 94.1 (RBA) to 136.0 μg (HPLC-FL) STX eq. kg−1 (n = 6) (Figure 1). Particularly in the brain, the concentration was 111.1 μg STX eq. kg−1.
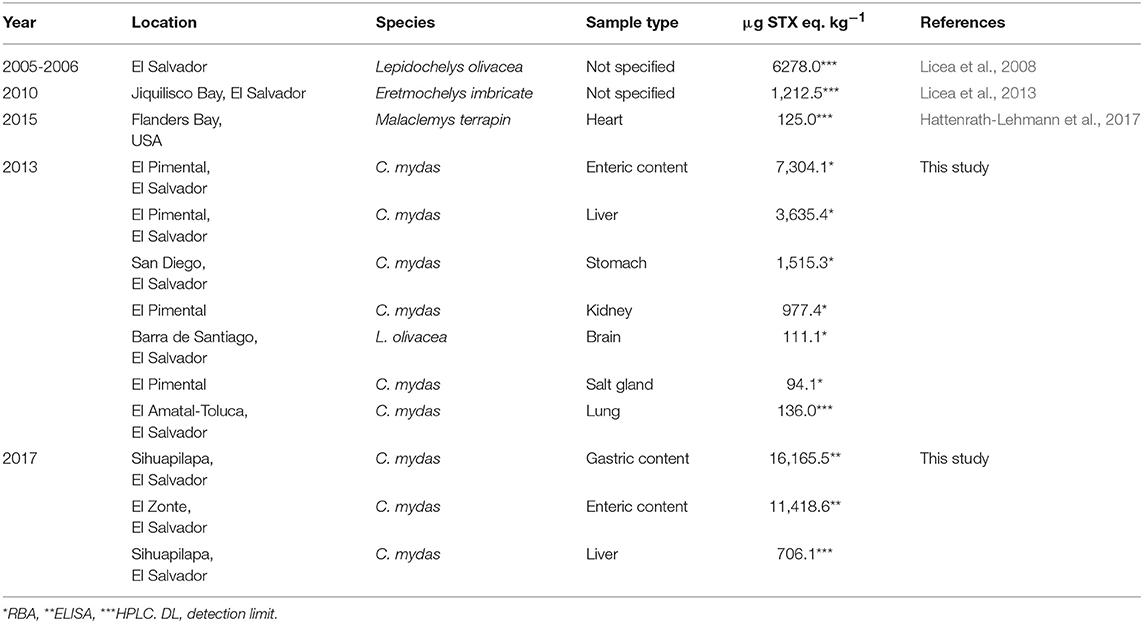
Table 1. Maximum PST concentrations for each sample type using Receptor Binding Assay, HPLC-FL and ELISA in sea turtle tissues from 2013 and 2017 mortality events.
During 2013 event, minimum PST concentrations were found in C. mydas intestinal contents with 18.8 μg STX eq. kg−1; while in 2017, minimum PST concentration were found in C. mydas liver with 4.0 μg STX eq. kg−1.
Ten of the samples analyzed at FWRI in 2017 were selected for HPLC-FL analysis, and the presence of PST was confirmed in all 10 samples. Of the PST monitored for (STX, NEO, dcSTX, B1, GTX 1/4, GTX 2/3, dcGTX-2/3, and C1/2) only STX, B1, and dcSTX were detected (Table 2). The toxin profile was dominated by STX, which represented 63–82% of the toxin present. B1 accounted for 14–33%, and dcSTX was present at low levels (0–7% of the total).
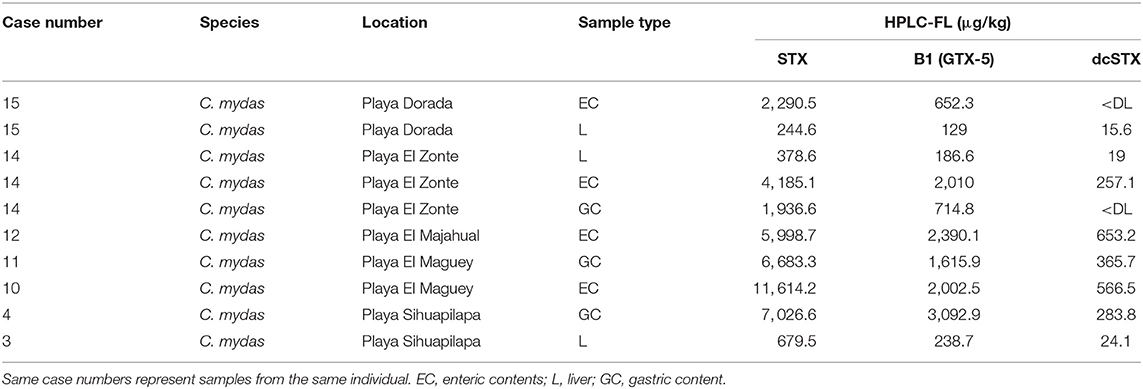
Table 2. Results of analysis for saxitoxin (STX) and gonyautoxins (GTX) using high performance liquid chromatography with fluorescence detection (HPLC-FL) for 2017 mortality event.
The most abundant potential producer of PST found in the phytoplankton samples from 2013 was Gymnodinium catenatum (5,400 cells L−1) in Acajutla. Other PST-producing species were detected in low abundance, such as Pyrodinium bahamense and Alexandrium monilatum (Table 3). During 2017, Alexandrium sp. was the only potential PST-producing microalgae detected in water samples from Los Cóbanos, with a low abundance of 672 cell per liter. The most abundant phytoplankton species found in the area were the diatoms Dactyliosolen fragilissimus and Pseudo-nitzschia spp., with 110,235 cell L−1 and 4,033 cell L−1, respectively (Table 3).
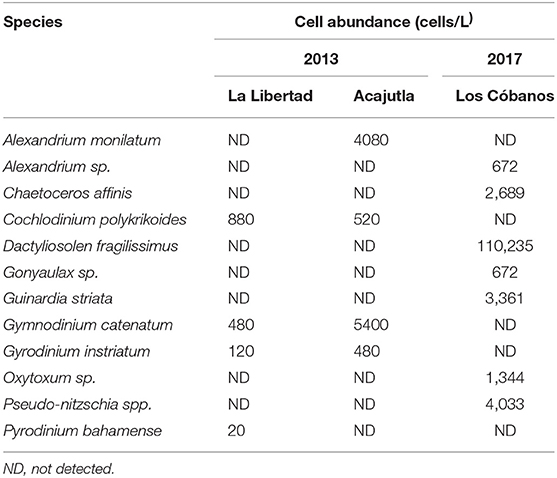
Table 3. Concentration of the most abundant phytoplankton species found in La Libertad and Acajutla during the 2013 event, and in Los Cóbanos during 2017.
In contrast to the water samples, microscopy of gastrointestinal contents of dead turtles from 2017 found Pyrodinium bahamense cells and thecae in gastric and enteric contents from two C. mydas. No PST-producing dinoflagellates were found in the enteric contents of an additional C. mydas and one L. olivacea, but whole cells of Planktoniella sol, Scrippsiella trochoidea, and Prorocentrum compressum were found in low abundance.
During early September 2013, a potential dinoflagellate bloom was present off the western coast of El Salvador (Figures 3A,B). Another suspect bloom appeared during mid-September 2013 along the central coast (Figures 3C,D) and persisted through the end of the month (Figures 3E,F).
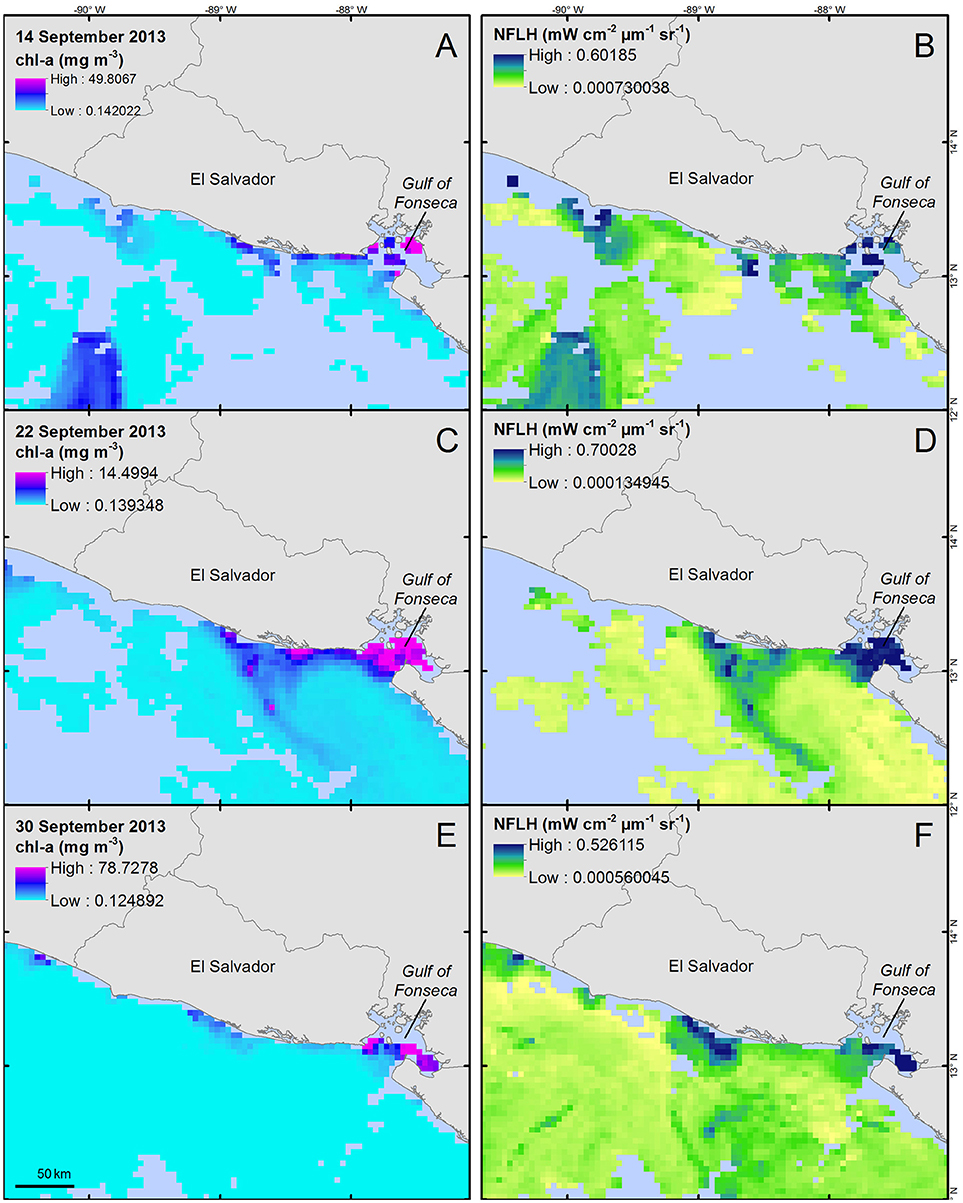
Figure 3. Chlorophyll-a concentration (A,C,E, units = mg m−3) and normalized fluorescence line height (B,D,F, units = mW cm−2 μm−1 sr−1) for three 8-day time periods during September 2013. Data were collected from the MODIS-Aqua sensor and provided by NASA's Giovanni system (Acker and Leptoukh, 2007). Corresponding images represent 8-day means for each parameter ending on the dates displayed in the images; (A,B) represent 6–14 September data, (C,D) represent 14–22 September, and (E,F) represent 22–30 September. All maps are shown at an equal spatial scale.
Throughout October 2017, a potential dinoflagellate bloom was present off El Salvador's central coast (Figure 4). In addition, a potential offshore bloom appeared to the south of the central coast during late October (Figures 4C,D).
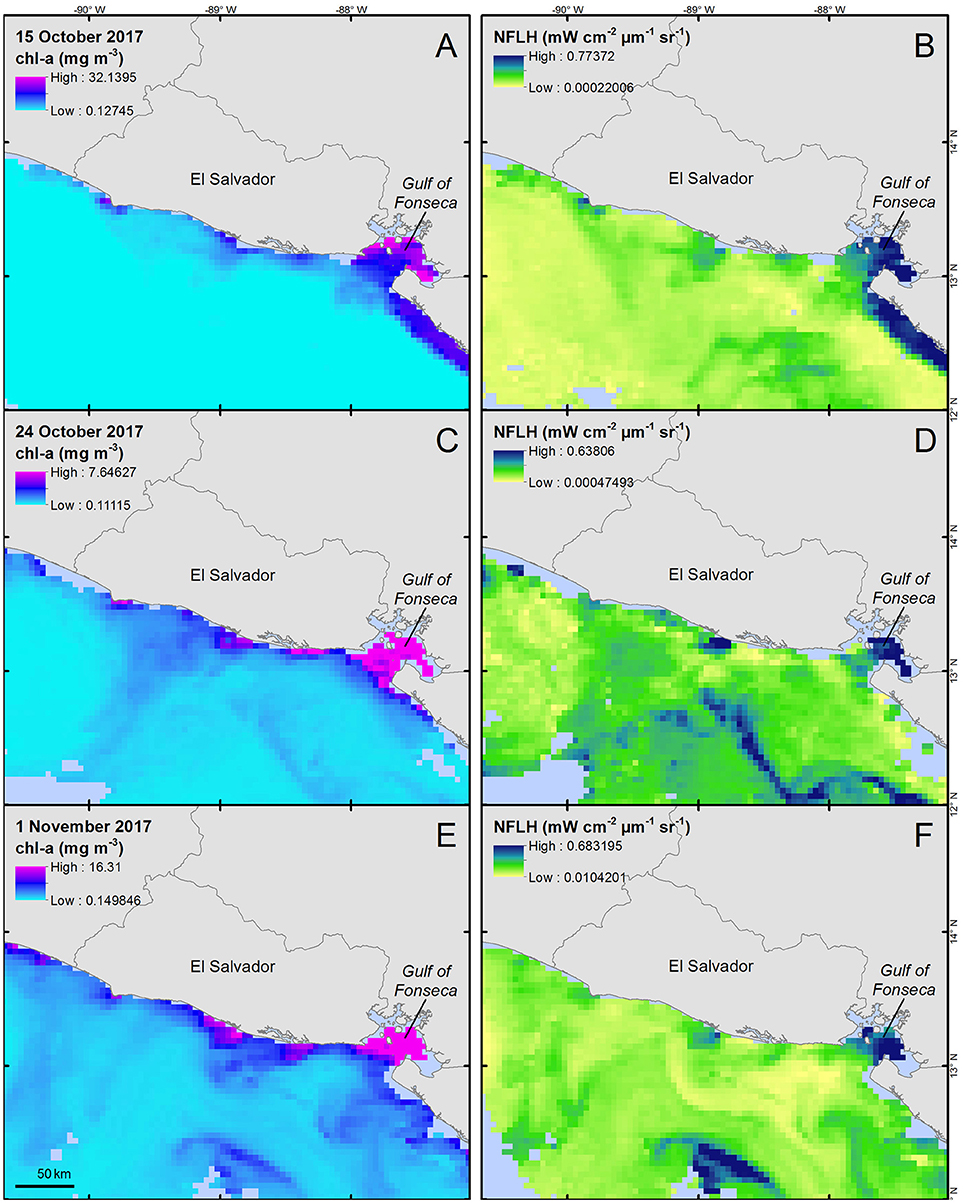
Figure 4. Chlorophyll-a concentration (A,C,E, units = mg m−3) and normalized fluorescence line height (B,D,F, units = mW cm−2 μm−1 sr−1) for three 8-day time periods during October 2017. Data were collected from the MODIS-Aqua sensor and provided by NASA's Giovanni system (Acker and Leptoukh, 2007). Corresponding images represent 8-day means for each parameter ending on the dates displayed in the images; (A,B) represent 8–15 October data, (C,D) represent 16–24 October, and (E,F) represent 24 October−1 November. All maps are shown at an equal spatial scale.
Previous sea turtle mass mortality events suspected to be caused by PST-producing dinoflagellate blooms have been reported on the Pacific coast of Mexico and Central America, and in Papua New Guinea; however, very limited information about these events is available (Maclean, 1975; Licea et al., 2008, 2013; Meave del Castillo et al., 2008).
There are significant challenges associated with investigating sea turtle mortality events in many regions of the world, including the remote nature of many coastal areas, limited logistical resources, and inevitable delays between initial observations and response to reports. Although our sample size and some aspects of our data reflect these challenges, we were able to characterize two relatively large sea turtle mortality events in El Salvador and provide evidence that exposure to PST was the most likely cause. This evidence includes postmortem findings consistent with acute toxicosis, detection of relatively high concentrations of PST in gastrointestinal contents and tissues, observation of PST-producing dinoflagellates within gastrointestinal contents, and remote-sensing data suggestive of dinoflagellate blooms concurrent with sea turtle mortality.
In Chelonia mydas, higher concentrations of STX has been found in the gastro-intestinal tract contents rather than tissues (Capper et al., 2013). Toxicity thresholds have not been defined for PST in chelonians or other reptiles. Given their conservation status, exposure studies in sea turtles are not feasible and laboratory studies using non-imperiled chelonians have not yet been conducted. Attribution of clinical effect and mortality currently relies on exclusion of other possible causes and comparison of toxin exposures associated with mortality during known blooms, as well as exposure in turtles that are clinically stable or those that died from apparently unrelated causes. The dead turtles found during both events described herein were consistent with the general features of sea turtle mortality attributed to brevetoxicosis, another biotoxin that acts on neuronal voltage-gated sodium channels (Fauquier et al., 2013; Walker et al., 2018). These features include strandings of turtles in good nutritional condition without injuries, other abnormalities, or known association with other causes of mortality. Similar to a mortality event involving diamondback terrapins (Malaclemys terrapin) associated with a PST-producing Alexandrium bloom (Hattenrath-Lehmann et al., 2017), all turtles were found dead thus we were unable to ascertain whether affected turtles exhibited signs of neurotoxicosis. However, as previously reviewed, abnormal neurological signs including, erratic swimming, disorientation, and reduced activity were observed in stranded turtles found in 2010, one of which had detectable STX in brain tissue (Licea et al., 2013).
The PST concentrations detected in sea turtles that were found during both events were similar to those reported in 2006 (Licea et al., 2008) and 2010 (Licea et al., 2013), the only other published accounts of sea turtle mortality associated with PSP that includes toxin analyses. Our values were much higher than detected in the aforementioned M. terrapin mortality event (Hattenrath-Lehmann et al., 2017). In the case of PST found in one sample from brain tissue in 2013, the value is lower than found in brain tissue from the 2005 to 2006 mortality event (Licea et al., 2008). Two of the authors (Stacy and Flewelling, unpublished data) have screened sea turtles found stranded in Florida for PST as part of health studies and investigation of mortality events. In those results, concentrations typically are below detectable limits (10 ng/g) or well below those documented in this report, including within areas that experience periodic PST-producing blooms. Thus, although some degree of asymptomatic PST exposure almost certainly occurs in sea turtles, we have not found evidence of widespread background exposure similar to that measured during these events as a complication related to interpretation of analytical results. Nonetheless, broader sampling is needed of sea turtles during mortality events and PST-producing blooms and from unrelated circumstances with which to better understand PST exposure in Central America.
The opportunistic nature of sampling during these events did not allow conclusions to be drawn regarding differences in toxin concentrations among turtles and sample types other than variation is attributable to potential differences in dose, timing of exposure, toxin absorption and metabolism, and analytical methods. There are notable differences in the assays performed, which were determined by logistical considerations during each event. For example, RBA measures overall PST toxicity, whereas HPLC measures concentrations of specific congeners (Costa et al., 2009). ELISA is very sensitive and useful for screening for exposure, and analytical confirmation was achieved for the 2017 animals screened by ELISA. However, we did not have the ability to compare results of the same samples across all three methods. Future efforts should endeavor to standardize both sampling and analytical methods to the degree possible.
We observed anomalies in remotely-sensed imagery suggesting possible dinoflagellate blooms during both events. Unfortunately, available water samples were inadequate to confirm that the anomalies observed in satellite imagery were blooms of PST-producing species; corresponding in situ water samples are essential to confirm the presence of a bloom (Hu et al., 2005). The spatiotemporal extent of water sampling was extremely limited and was not considered representative of the anomalies detected by satellite. Nevertheless, the possibility that the observed anomalies were caused by blooms of Pyrodinium bahamanse and the likelihood that such blooms contributed to these mortality events are strengthened by concurrent detection of PST exposure in turtles, detection of Pyrodinium bahamense in the gastrointestinal contents of some turtles, and the observed toxin profiles. PST toxin profiles reported for P. bahamense vary somewhat from location to location, but generally contain fewer toxins than those described for other PST-producing dinoflagellates (Wiese et al., 2010). Profiles consisting of STX, NEO, dcSTX, GTX5, and GTX6 were reported in P. bahamense clones both from Palau (Harada et al., 1982, 1983) and Malaysia (Usup et al., 1994). Pyrodinium bahamense isolates from the Philippines and the southeastern US contained only STX, GTX5, and dcSTX (Landsberg et al., 2006; Gedaria et al., 2007), similar to the PST profile observed in the turtles analyzed in the present study. Although we were unable to confirm and identify blooms of specific species, the remotely-sensed data provided insight into the possible locations and extent of the blooms that may have resulted in toxin exposure and caused sea turtle mortality. Hopefully, this technology will be used to inform real-time sampling during future events.
Another consideration relevant to collection and interpretation of environmental data is the potential for persistent toxin exposure following dissipation of blooms. In marine animal mortality events attributed to brevetoxicosis, animals are not only exposed to high toxin levels during bloom periods, but also to toxins circulating through food webs for weeks or months after a bloom has dissipated (Landsberg et al., 2009). The same potential may exist for PST.
The number of sea turtles found likely reflects a minority of affected animals. Previous studies of beach strandings have estimated that around 5–20% of sea turtles that die at sea are subsequently found on shore (Hart et al., 2006; Mancini et al., 2011; Koch et al., 2013). Therefore, actual mortality from both events could have numbered in the hundreds or even thousands of sea turtles. Moreover, the 2013 event may have been widespread based on the extent of the potential bloom observed in remotely-sensed imagery and reported concurrent sea turtle strandings in Guatemala (Brittain et al., 2014).
Given the status of sea turtles and efforts that are required to manage significant anthropogenic threats, an understanding of various causes of mortality is important for conservation and management. The number of sea turtle deaths attributed to HABs on the Pacific coast of Central America is notable and warrants further effort to understand the factors that contribute to these events.
OA, RQ, and M-YD contributed conception and design of the study. RQ, LF, and BS organized the database. RQ performed the statistical analysis. OA wrote the first draft of the manuscript. RQ, M-YD, BS, RH, LF, CD, and GR wrote sections of the manuscript. All authors contributed to manuscript revision, read and approved the submitted version.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank the International Agency of Atomic Energy through projects RLA7020 and RL7022, the staff from the Ecosystems and Wildlife Department of the Ministry of the Environment and Natural Resources, the guards of Los Cóbanos Marine Protected Area and Puerto Parada, staff from Governance and Territory Department of the Ministry of the Environment and Natural Resources, the Network of Local Observers of MARN, Civil National Police, NGOs PROCOSTA-ICAPO, FUNZEL and ACOTOMBSAB, and the people who supported in the collection of tissues and stranding information. The International Atomic Energy Agency is grateful to the Government of the Principality of Monaco for the support provided to its Environment Laboratories. We also thank the U.S. Department of State for facilitating diagnostic analyses and April Granholm (FWRI) and Sugandha Shankar (FWRI) for performing toxin analyses. Satellite imagery analyses and visualizations used in this study were produced with the Giovanni online data system, developed and maintained by the NASA GES DISC.
Acker, J. G., and Leptoukh, G. (2007). Online Analysis Enhances Use of NASA Earth Science Data. Eos. Trans. AGU 88, 14–17. doi: 10.1029/2007EO020003
Amaya, O., Ruiz, G., Rivera, W., and Espinoza, J. (2012). Application of a Receptor Binding Assay to the Analyses of PSP Toxins in Four Species of Shellfish in El Salvador. Harmful Algae News 45, 14–15.
Brittain, R., Lucas, S., Handy, S., and Nunny, R. (2014). Sea Turtle Strandings and Mortalities on the Southeast Coast of Guatemala. Marine Turtle Newsl. 143, 10–13.
Capper, A., Flewelling, L. J., and Arthur, K. (2013). Dietary Exposure to Harmful Algal Bloom (HAB) Toxins in the Endangered Manatee (Trichechus Manatus Latirostris) and Green Sea Turtle (Chelonia mydas) in Florida, USA. Harmful Algae 28, 1–9. doi: 10.1016/j.hal.2013.04.009
Costa, P. R., Baugh, K. A., Wright, B., RaLonde, R., Nance, S. L., Tatarenkova, N., et al. (2009). Comparative determination of paralyic shellfish toxins (PSTs) using five different toxin detection methods in shellfish species collected in the Aleutian Islands, Alaska. Toxicon 54, 313–320. doi: 10.1016/j.toxicon.2009.04.023
Dechraoui Bottein, M.-Y., and Clausing, R. J. (2017). “Receptor-Binding Assay for the Analysis of Marine Toxins: Detection and Mode of Action.” in Recent Advances in the Analysis of Marine Toxins, Vol. 78, eds J. Diogene and M. Campas (Amsterdam: Elsevier B. V.), 277–301. doi: 10.1016/bs.coac.2017.08.004
Espinoza, J., Amaya, O., and Quintanilla, R. (2013). Atlas de Fitoplancton Marino. 1st Edn. San Salvador: Universidad de El Salvador.
Fauquier, D. A., Flewelling, L. J., Maucher, J., and Manire, C. A. (2013). Brevetoxin in blood, biological fluids, and tissues of sea turtles naturally exposed to Karenia brevis blooms in central west Florida. J. Zoo Wildl. Med. 44, 364–375. doi: 10.1638/2012-0164R.1
Gedaria, A. I., Luckas, B., Reinhardt, K., and Azanza, R. V. (2007). Growth response and toxin concentration of cultured Pyrodinium bahamense var. compressum to varying salinity and temperature conditions. Toxicon 50, 518–529. doi: 10.1016/j.toxicon.2007.04.021
Geraci, J. R., Anderson, D. M., Timperi, R. J., St. Aubin, D. J., Early, G. A., Prescott, J. H., et al. (1989). Humpback whales (Megaptera novaeangliae) fatally poisoned by dinoflagellate toxin. Can. J. Fish. Aquatic Sci. 46, 1895–1898 doi: 10.1139/f89-238
Harada, T., Oshima, Y., Kamiya, H., and Yasumoto, T. (1982). Confirmation of paralytic shellfish toxins in the dinoflagellate Pyrodinium bahamense var. compressa and bivalves in Palau. Bull. Jpn. Soc. Sci. Fish 48, 821–825. doi: 10.2331/suisan.48.821
Harada, T., Oshima, Y., and Yasumoto, T. (1983). Natural occurrence of decarbamoylsaxitoxin in tropical dinoflagellate and bivalves. Agric. Biol. Chem. 47, 191–193.
Hart, K., Mooreside, P., and Crowder, L. (2006). Interpreting the spatio-temporal patterns of sea turtle strandings: going with the flow. Biol. Conserv. 129, 283–290. doi: 10.1016/j.biocon.2005.10.047
Hattenrath-Lehmann, T. K., Ossiboff, R. J., Burnell, C. A., Rauschenberg, C. D., Hynes, K., Burke, R. L., et al. (2017). The role of a PSP-producing Alexandrium bloom in an unprecedented diamondback terrapin (Malaclemys terrapin) mortality event in Flanders Bay, New York, USA. Toxicon 129, 36–43. doi: 10.1016/j.toxicon.2017.02.006
Hu, C., Muller-Karger, F. E., Taylor, C., Carder, K. L., Kelble, C., Johns, E., et al. (2005). Red tide detection and tracing using MODIS fluorescence data: a regional example in SW Florida coastal waters. Remote Sens. Environ. 97, 311–321. doi: 10.1016/j.rse.2005.05.013
IUCN (2018). The IUCN Red List of Threatened Species. Version 2018-1. Available online at: http://www.iucnredlist.org
Koch, V., Peckham, H., Mancini, A., and Eguchi, T. (2013). Estimating at-sea mortality of marine turtles from stranding frequencies and drifter experiments. PLoS ONE 8:e56776. doi: 10.1371/journal.pone.0056776
Landsberg, J. H., Flewelling, L. J., and Naar, J. (2009). Karenia brevis Red Tides, Brevetoxins in the Food Web, and Impacts on Natural Resources: Decadal Advancements. Harmful Algae 8, 598–607. doi: 10.1016/j.hal.2008.11.010
Landsberg, J. H., Hall, S., Johannessen, J. N., White, K. D., Conrad, S. M., Abbott, J. P. Steidinger, et al. (2006). Saxitoxin puffer fish poisoning in the United States, with the first report of Pyrodinium bahamense as the putative toxin source. Environ. Health Perspect 114, 1502–1507. doi: 10.1289/ehp.8998
Lawrence, J. F., and Niedzwiadek, B. (2001). Quantitative determination of paralytic shellfish poisoning toxins in shellfish by using prechromatographic oxidation and liquid chromatography with fluorescence detection. J. AOAC Inter. 84, 1099–1108.
Lefebvre, K. A., Quakenbush, L., Frame, E., Huntington, K. B., Sheffield, G., Stimmelmayr, R., et al. (2016). Prevalence of algal toxins in Alaskan marine mammals foraging in a changing arctic and subarctic environment. Harmful Algae 55, 13–24. doi: 10.1016/j.hal.2016.01.007
Licea, S., Navarrete, A., Casta-eda, V., and Guzmán, J. B. (2013). “Monitoring Program for Harmful Algal Blooms in Salvadoran Waters: Report of Pyrodinium bahamense from November 2009 to June 2010,” in Proceedings of the 14th International Conference on Harmful Algae, eds K. A. Pagou and G. M. Hallegraeff (International Society for the Study of Harmful Algae and Intergovernmental Oceanographic Commission of UNESCO), 120–22.
Licea, S., Navarrete, A., Rodriguez, R., Bustillos, J., Martínez, B. C., and Ramirez (2008). “Monitoring a Bloom of Pyrodinium Bahamense Var. Compressus Occurring in El Salvador, Guatemala and Mexico (November 2005-March 2006),” in 12th Proceedings of the International Conference on Harmful Algae, eds G. Doucette, H. Enevoldsen, A. Godhe, G. Hallegraeff, B. Luckas, N. Lundholm, J. Lewis, K. Rengefors, K. Sellner, K. Steidinger, P. Tester, and A. Zingone (Copenhague: International Society for the Study of Harmful Algae and Intergovernmental Oceanographic Commission of UNESCO), 219–220.
Mancini, A., Koch, V., Seminoff, J. A., and Madon, B. (2011). Small-scale gill-net fisheries cause massive green turtle Chelonia mydas mortality in Baja California Sur, Mexico. Oryx 46, 69–77. doi: 10.1017/S0030605310001833
Meave del Castillo, E., Rodríguez, R., and Vargas, M. (2008). “Blooms of Pyrodinium bahamense var. compressum along the Pacific Coast of Central America and southern Mexico,” in Proceedings of the 12th International Conference on Harmful Algae, eds G. Doucette, H. Enevoldsen, A. Godhe, G. Hallegraeff, B. Luckas, N. Lundholm, J. Lewis, K. Rengefors, K. Sellner, K. Steidinger, P. Tester, and A. Zingone (Copenhague: International Society for the Study of Harmful Algae and Intergovernmental Oceanographic Commission of UNESCO), 212–215.
Oshima, Y. (1995). Postcolumn derivation liquid chromatographic method for paralytic shellfish toxins. J. AOAC Int. 78, 528–532.
Shearn-Bochsler, V., Lance, E. W., Corcoran, R., Piatt, J., Bodenstein, B., Frame, E., et al. (2014). Fatal paralytic shellfish poisoning in Kittlitz's murrelet (Brachyramphus brevirostris) nestlings, Alaska, USA. J. Wildl. Dis. 50, 933–937. doi: 10.7589/2013-11-296
Shumway, S. E., Allen, S. M., and Dee Boersma, P. (2003). Marine birds and harmful algal blooms: sporadic victims or under-reported events? Harmful Algae 2, 1–17. doi: 10.1016/S1568-9883(03)00002-7
Usup, G., Kulis, D. M., and Anderson, D. M. (1994). Growth and toxin production of the toxic dinoflagellate Pyrodinium bahamense var. compressum in laboratory cultures. Nat. Toxins 2:254–262. doi: 10.1002/nt.2620020503
Utermöhl, H. (1958). Zur Vervollkommnung Der Quantitativen Phytoplankton-Methodik: Mit 1 Tabelle Und 15 Abbildungen Im Text Und Auf 1 Tafel. SIL Commun. 9, 1–38. doi: 10.1080/05384680.1958.11904091
Van Dolah, F. M., Doucette, G. J., Gulland, F. M. D., Rowles, T. L., and Bossart, G. D. (2003). “Impacts of algal toxins on marine mammals,” in Toxicology of Marine Mammals, eds J. G. Vos, G. D. Bossart, M. Fournier, T. J. O'Shea (London: Taylor & Francis), 247–269.
Van Dolah, F. M., Fire, S. E., Leighfield, T. A., Mikulski, C. M., and Doucette, G. J. (2012). Determination of Paralytic Shellfish Toxins in Shellfish by Receptor Binding Assay: Collaborative Study. J. AOAC Int. 95, 795–812. doi: 10.5740/jaoacint.CS2011
Walker, J. S., Shaver, D. J., Stacy, B. A., Flewelling, L. J., Broadwater, M. H., and Wang, Z. (2018). Brevetoxin exposure in sea turtles in south Texas (USA) during Karenia brevis red tide. Dis. Aquat. Organ. 31, 145–150 doi: 10.3354/dao03194
Wiese, M., D'Agostino, P. M., Mihali, T. K., Moffitt, M. C., and Neilan, B. A. (2010). Neurotoxic alkaloids: saxitoxin and its analogs. Mar. Drugs 8, 2185–2211. doi: 10.3390/md8072185
Keywords: sea turtle, paralytic shellfish poisoning, saxitoxin, receptor binding assay, HABs
Citation: Amaya O, Quintanilla R, Stacy BA, Dechraoui Bottein M-Y, Flewelling L, Hardy R, Dueñas C and Ruiz G (2018) Large-Scale Sea Turtle Mortality Events in El Salvador Attributed to Paralytic Shellfish Toxin-Producing Algae Blooms. Front. Mar. Sci. 5:411. doi: 10.3389/fmars.2018.00411
Received: 01 August 2018; Accepted: 16 October 2018;
Published: 06 November 2018.
Edited by:
Juan José Dorantes-Aranda, Institute for Marine and Antarctic Studies (IMAS), AustraliaReviewed by:
Gustaaf Marinus Hallegraeff, University of Tasmania, AustraliaCopyright © 2018 Amaya, Quintanilla, Stacy, Dechraoui Bottein, Flewelling, Hardy, Dueñas and Ruiz. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rebeca Quintanilla, cmViZWtxdWludGFuaWxsYUBnbWFpbC5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.