 Juliana López-Angarita1,2*
Juliana López-Angarita1,2* Alexander Tilley2,3
Alexander Tilley2,3 Juan M. Díaz4,5
Juan M. Díaz4,5 Julie P. Hawkins1E. Fernando Cagua6
Julie P. Hawkins1E. Fernando Cagua6 Callum M. Roberts1
Callum M. Roberts1- 1Environment Department, University of York, York, United Kingdom
- 2Fundación Talking Oceans, Bogotá, Colombia
- 3WorldFish, Dili, Timor-Leste
- 4Fundación Marviva, Bogotá, Colombia
- 5Department of Geography, Universidad Nacional de Colombia, Bogotá, Colombia
- 6Centre for Integrative Ecology, School of Biological Sciences, University of Canterbury, Christchurch, New Zealand
The Pacific coast of Colombia has some of the most extensive mangrove forests in South America. As an isolated region and one of the country's poorest, coastal communities rely on fishing as a main source of animal protein and income. In an attempt to reverse declining trends of fisheries resources, in 2008, an Exclusive Zone of Artisanal Fishing closed to industrial fishing, was established by stakeholders in the Northern Chocó region. Here we present a case study to investigate the effects of this area-based management on fisheries productivity and catch composition. Fishery landings data from 2010 to 2013 are compared to those of a neighbouring region with no fisheries management. Catch per unit effort, mean weight landed, and number of landed individuals were calculated for mangrove and non-mangrove associated species by boat type and fishing gear. A set of mixed effects models were used to unpack the effects of multiple factors and their interactions on response variables. Results show that across fishing gears and time, mean catch per unit effort increased by 50% in the Exclusive Zone of Artisanal Fishing within 3 years. Fisheries here focused on offshore resources with 61% more fishing trips associated with motorised boats than in the unmanaged region, where fishing was predominantly in mangroves and close to the coast. This suggests that fisheries management may have played a role in reducing pressure on mangrove resources. However, area-based management may have also driven the displacement of fishing effort by excluding industrial trawlers, which then concentrated their activity in neighbouring areas.
Introduction
Fisheries are important to the national economies of many developing countries, through contributions to food security and supply, employment, livelihoods, and poverty alleviation (Béné, 2009; Finegold, 2009; Mills et al., 2011). However, as fisheries are declining worldwide all these are threatened by that loss (Pauly and Zeller, 2015). Diverse strategies have been proposed to reverse fisheries decline, and include the creation of protected areas, regulation of fishing activity (e.g., total allowable catch, gear restrictions, temporal restrictions), and protection of essential fish habitat (Gell and Roberts, 2003). In the tropics, mangrove habitat offers critical refuge and food to associated fish and invertebrate species (Nagelkerken et al., 2001; Aburto-Oropeza et al., 2008) before they migrate elsewhere (Mumby et al., 2004). Consequently, mangrove protection may serve to safeguard or restore fisheries productivity (Aburto-Oropeza et al., 2008; Carrasquilla-Henao and Juanes, 2016). Mangroves in Colombia host a great diversity of important fishery species such as cockles, prawns, shrimp, crabs and fish such as catfish, snook, snapper, and tarpon (Villalba, 2005). Mangroves are abundant on the Pacific coastline around its extensive sheltered estuaries, bays, and rivers (Prahl et al., 1990). By contrast, on the Atlantic coast, wave climate is often too energetic for mangroves (Álvarez-León and Polanía, 1996). On the Pacific coast, mangroves occur in a broad band in the south, becoming patchy in the north where there are fewer big rivers (Prahl, 1989).
The Colombian Pacific fisheries are highly productive, sustaining 80% of Colombia's total fish catch (Díaz et al., 2011), however, since 1990s catch levels have been steadily decreasing (Lindop et al., 2015). Evidence suggests that many fisheries have exceeded their maximum sustainable yield and that resources are overexploited (Díaz et al., 2011). This was highlighted by the overexploitation of the white shrimp (Litopenaeus occidentalis) and titi shrimp (Xiphopenaeus riveti) fisheries in the Pacific since 1990s (Rueda et al., 2001). In response, industrial fishing fleets were drastically reduced by the diminishing resources (Baos Estupiñán and Zapata, 2011). This situation led to further encroachment from industrial fishers into coastal waters to boost their catches (García, 2010) which in turn fuelled self-mobilised, artisanal fishers groups of the Northern Chocó region to create the Exclusive Zone of Artisanal Fishing (ZEPA) in 2008 (Vieira et al., 2016). This area excluded industrial fishing within 4.6 km of the coast, and incorporated management measures such as the prohibition or gillnets and beach seines to allow the recovery of overfished populations (Vieira et al., 2016).
In this study, we investigate how area-based fisheries management has affected catch composition by comparing landings data from the ZEPA, with a comparable unmanaged area. Additionally, given the need for more information on the contribution of mangroves to subsistence fisheries (Saenger and Funge-Smith, 2013), we examine the importance of mangrove-associated species in the small-scale fisheries of Northern Chocó between 2010 and 2013.
Methods
Study Site
This study took place in the Northern Chocó region of Colombia's Pacific coast (Figure 1), a global biodiversity hotspot with high levels of endemism in flora and fauna (Myers et al., 2000; Díaz and Gast, 2009). The area has some of the world's highest rainfall with figures of 5,000 to 7,700 mm per year, and an average relative humidity of ~89% (Díaz and Galeano, 2016). Normally a rainy season occurs between May and November with dryer weather from December to April, but this varies according to the El Niño–Southern Oscillation (Wang and Fiedler, 2006). The coastline is characterised by rocky hills and precipices and receives input from many rivers. Large estuaries occur in Juradó, Cupicá, Tribugá and Nuquí, where the main mangrove forests occur, and these cover approximately 4,978 ha. The population of ~19,000, are predominantly Afro-descendants and indigenous people from the “Emberá” culture (Camacho et al., 2000) inhabiting small villages in the rainforest. The Colombian Pacific has remained isolated from the economic growth in the rest of the country due to weak presence of government institutions and a lack of infrastructure development, and hence, accessibility (Camacho et al., 2000; Díaz and Galeano, 2016). More than half the population lives in poverty (Camacho et al., 2000; Blanco et al., 2011), with economic activities in these isolated communities restricted to small-scale agriculture and fishing (Blanco et al., 2011). Fishing is mainly artisanal, with commercial activities very limited due to high transport costs for national distribution and export (Díaz and Galeano, 2016).
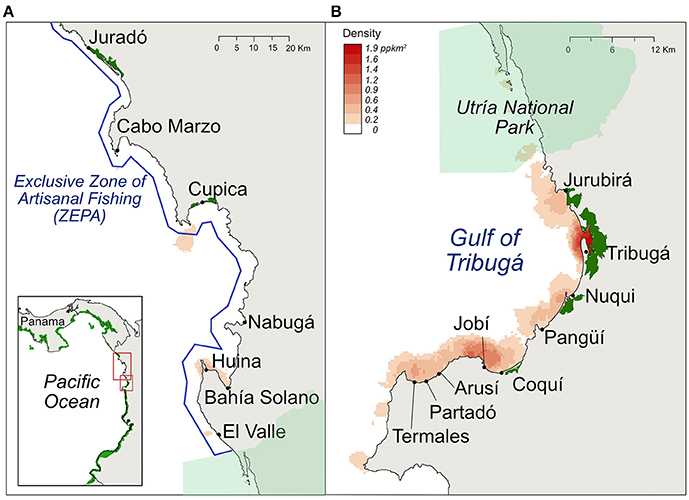
Figure 1. (A) Study site location in the Northern Chocó region on the Pacific coast of Colombia (red square, inset). Solid black circles represent artisanal fisheries landing sites included in this study. Blue line represents the Exclusive Zone of Artisanal Fishing (ZEPA). Mangroves are shown in green. Red shading corresponds to density estimation of fishing grounds showing the distribution of fishing sites per km2 in (A) the ZEPA and (B) the Gulf of Tribugá. Dark red areas represent high fishing ground density and light red represents low density.
The region has two distinct fishery management zones: the Gulf of Tribugá (hereafter referred to as Tribugá) where there are no fishing regulations in place; and the Exclusive Zone of Artisanal Fishing (ZEPA) (García, 2010), where fisheries management exists. In Tribugá there is one protected area called Utría National Park, which was declared in 1986 (García, 2010; Figure 1). However, since this Park does not have distinct regulations to restrict fishing activities, we treated it as part as of Tribugá. In the ZEPA, fishing regulations prohibit the use of gillnets and only allow hand lines and long lines (Vieira et al., 2016). Gear and area restrictions are actively enforced by local fishing cooperatives and fishers in the ZEPA, but some level of non-compliance remains evidenced by gillnet landings reported from the ZEPA.
Data Sampling
To compare the fisheries of the ZEPA and Tribugá, we examined data collected between March 2010 and September 2013 from 16 coastal communities in the two areas by the Marviva Foundation (Marviva). This organization trained local people to collect fishery information using surveys designed for a national information system of landings (Neira et al., 2016). Data recorders were community members familiar with fishing practices such as ex-fishermen, fisher's wives, and fisher family members. They received training by Marviva in both areas with support of marine research institutions in Colombia and government fishing authorities, and their performance was evaluated once a month by a Marviva officer to ensure standardised sampling procedures. Sites where the information was collected were chosen within all landing areas such that all fishing gears used would be considered. Given that most communities are very small (i.e., <400 people per community) and isolated, we do not consider that the distance of landing site from market had a significant effect on the type or quality of fish landed. Sampling frequency varied according to budget availability and local conditions. In general, catch and effort data were recorded 3–4 times per week. Sampling was uniform throughout the year apart from where local religious holidays or restrictive inclement weather created gaps. The information we examined from the Marviva dataset was: catch weight and number, species landed, type of boat used, number of fishers, trip length, fishing gear, and fishing location. Landed weight was recorded using the same model of hook scales in all landing sites. Fisheries independent trips were undertaken by Marviva data recorders to map the location of named fishing grounds (i.e., sites where people regularly fish) recording a single central GPS point for different localities.
Characterizing Artisanal Fisheries of Northern Chocó
Landings data for the ZEPA and Tribugá were analysed to characterise fisheries according to gear type, species composition, and fishing location. Temporal change in landings over the study period was then calculated for these fisheries. Geographic locations of fishing grounds were mapped in ArcGIS 10.3.1 as point data, and interpolated to create a spatial density map of the fishing grounds in each area.
Relative importance of species and gear types were calculated using total weight landed. Catch per unit effort (CPUE) was used as a measure of relative fish abundance across time, and to compare gear types and management zones. CPUE by gear type for the ZEPA and Tribuga were compared using a Wilcoxon test, and a Tukey-Kramer test was used for post-hoc comparisons. Catch was summed in kilograms and fishing effort was calculated as fisher days whereby the number of days fishing was multiplied by the number of fishers on board the vessel.
To understand spatial patterns in the fisheries we calculated the distance from each fishing ground to the coast using ArcGIS 10.3.1. To do this we overlaid the map of points of fishing grounds over the map of the coastline of Northern Chocó, and used the spatial analyst extension in ArcGIS to calculate the most direct distance from each fishing ground to the closest point on the coastline. Mean distances from fishing grounds to the coast were compared between the ZEPA and Tribugá using a Kruskal-Wallis test. Proximity of fishing grounds to the coast was also calculated per boat type, with mean and standard error values estimated taking into account the boat ID because values taken from the same boat at different times are not independent. Boat type was classified as motorised or non-motorised with motorised boats further classified according to engine sizes of: <16, 16–40, and >40 HP. To determine the relative importance of each boat type for the fisheries, we calculated the proportion of each boat type in the total number of trips recorded for each area. Temporal trajectories of fisheries from the ZEPA and Tribugá were estimated by comparing annual CPUE (kg/fisher/day) of finfish species by summing weight landed per month and dividing it for the monthly effort values (kg/fisher/day) for each of the years. To determine if CPUE changes over time were significant, years were compared using a Wilcoxon test, and a Tukey-Kramer test was used for post-hoc comparisons. This was performed for the ZEPA and Tribugá independently, taking into account boat ID. All variables were tested for normality. Statistical analyses were performed in JMP ver. 13.
Exploring Management Effects on Mangrove Dependent Species
To analyse the proportion of mangrove dependent species in landings we defined mangrove dependence following the criteria of Blaber et al. (1989) as: “Species for which estuaries or similar habitats are the principal environment for at least one part of their life cycle.” Information was obtained from Fishbase (www.fishbase.org), The IUCN Red List of Threatened Species (www.iucnredlist.org), and Shorefishes of the Tropical Eastern Pacific (http://biogeodb.stri.si.edu/sftep/en/pages). Given there is incomplete knowledge of the life cycle of many eastern Pacific species we attributed mangrove dependence according to recognised information for similar species (Supplementary Table 1).
Temporal trajectories of relative abundance of mangrove dependent and non-mangrove dependent fish species were compared between Tribugá and the ZEPA using the proxy measure of CPUE. CPUE was calculated by summing the weight of landings per month sampled and dividing it by the monthly effort (fisher·days) for each of the years. Boat type was interpreted as proxy variable for access to financial capital, as more wealthy fishers or owners will have access to powered boats in comparison with less wealthy fishers who will use man-powered boats. Boat type was grouped into non-motorised and motorised. Distance from fishing grounds to the coast was not included in this analysis given its auto-correlation with boat type.
To explore the role of management interventions and mangrove dependence in the fisheries of Northern Chocó, we analysed the response variables of CPUE, catch weight, and number of individuals landed using linear mixed effects models. We first determined the distribution that best described each response variable. CPUE was tested using a normal distribution with a logarithmic link, a gamma and a lognormal distribution, whereas catch weight and number of individuals were tested with poisson, quasipoisson, and lognormal distributions. In all models, the lognormal distribution fitted the response variables best and met the model assumptions adequately. We included as fixed effects: management zone (ZEPA/Tribugá), fishing gears, species mangrove dependence (dependent/independent), and boat type (motorised/non-motorised). The interactions between the management area and all other explanatory variables were also included. Additional variables from the dataset were incorporated in the model as random effects to account for other potential sources of variability in the response variables. These included the potential correlation of observations taken from the same boat/fishers, the year, data recorder ID, fishing grounds and species name. The latter three were included to take into account recorder's bias, spatial variability of different fishing locations, and the species level variation in catchability. To account for differences in catch weight, CPUE and number of landed individuals between taxa (teleosts, mollusca, and crustacea), species were nested within each taxa. Unidentified species were not added as a category in the mixed effects model because they could not be classified as mangrove dependent and non-dependent. However, they were included in all other analyses where mangrove and non-mangrove species were not being compared.
We then selected the best set of models using an information theoretical approach with two stages. Firstly, we evaluated the relevance of the random effects, and then after fixing the random structure, we evaluated the relevance of the fixed effects. In both stages, we compared the Akaike information criterion (AIC) of each competing model. The lower the AIC, the higher the likelihood that the candidate model is the optimal model. We interpreted models with a ΔAIC (the difference between the model's AIC and the minimum AIC among all models) smaller than two, as there is considerably less support for models with larger ΔAIC (Burnham and Anderson, 2002). In the first stage, models were fitted using restricted maximum likelihood using the R package lme4 (Bates et al., 2015). In the second stage, models were fitted using log-likelihood. In both stages, all possible term combinations were generated. With the exception of the CPUE model set, the second best model had large ΔAICs and hence a multi-model inference approach was not necessary.
An important consideration is that since landings were not monitored before the implementation of regulations in the ZEPA, it is not entirely possible to disentangle the relationships between the presence of management and other factors that may contribute to the response variables. An ideal experimental approach would have been to apply a complete before-after-control-impact approach, but in our case, this was restricted by the lack of such data in the study region.
Results
Fisheries of Northern Chocó
Fisheries landings data were recorded in seven fishing communities in the ZEPA and nine in Tribugá. 270 fishing grounds were used in total with 179 located in Tribugá, and 91 in the ZEPA. These were more densely concentrated in Tribugá than in the ZEPA, where the distribution was predominantly uniform (Figure 1). The maximum distance fishers ventured from the coast was 13.8 km. Data were recorded for 6,054 fishing trips within the ZEPA and 30,394 within Tribugá. In the ZEPA, mean monthly fishing effort for 3 years of sampling was 557 fisher-days, vs. 1,832 for Tribugá across 4 years of sampling. The mean annual fishing effort for the ZEPA was 6,128 fisher-days compared to 21,071 for Tribugá. In the last population census in 2005, 8,475 people lived in the ZEPA and 7,089 Tribugá (Supplementary Table 2). Experts familiar with the region consider these numbers still reflect the current demography (C. Vieira, pers. comm., Marviva).
In Tribugá, 116 species were identified in landings, with 80 species from the ZEPA. This combined value increased to a total of 284 when including species with unknown scientific nomenclature. Fifty-five percent of species were classified as mangrove dependent. In the ZEPA, when total weight landed by species was calculated for the entire sampling period, Thunnus albacares was most the most important species with Sphyraena ensis for Tribugá (Figure 2). By this metric, other species that were important to both areas were: Brotula clarkae, Caranx caninus, Caranx caballus, Seriola rivoliana, Lutjanus guttatus, Lutjanus peru, and Scomberomorus sierra (Figure 2).
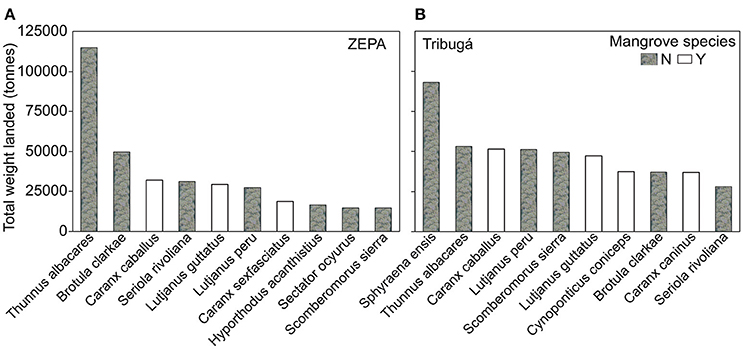
Figure 2. Top 10 species by weight landed (tones) during all the sampling period between 2010 and 2013 in the (A) Exclusive Zone of Artisanal Fishing (ZEPA) and (B) the Gulf of Tribugá. White bars represent mangrove dependent species and mottled grey bars represent species not dependent on mangroves.
For both the ZEPA and Tribugá, the main gear types by mean annual weight of landings were: hand lines, followed by long lines, gillnets, spear guns, manual collection for molluscs and beach seine (Table 1). Landings associated with cast net were only present in Tribugá. However, the relative importance of gear types changes when it is calculated using catch per unit effort, with beach seine showing the highest yields in both regions, followed by long line (Table 1). Annual mean CPUE was significantly different by gear type for Tribugá (Wilcoxon, χ2 = 922.7, DF = 6, P = < 0.001) and the ZEPA (Wilcoxon, χ2 = 122.8, DF = 5, P = < 0.001) (Supplementary Table 3). The most frequently used gear type was hand line, used on 71% of trips in Tribugá and 69% in the ZEPA. In Tribugá, gill nets were the second most important gear type, used on 18% of trips, followed by long line used on 9%. In the ZEPA, long lines were the second most important gear, used on 17% of trips, followed by gillnets which were used on 13%.
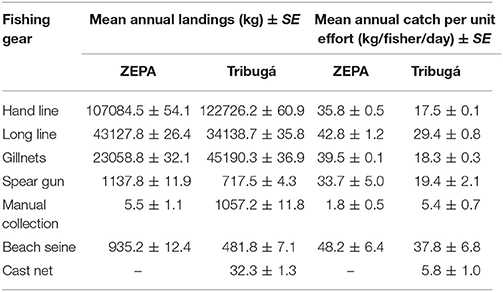
Table 1. Fishing gears used in Northern Chocó region between 2010 and 2013, showing mean weight ± standard error of annual catch and annual mean catch per unit effort.
In Tribugá, most fishing trips (75%) were conducted with non-motorised vessels, whereas in the ZEPA motorised boats accounted for 86% of boat trips of which 88% were 16 HP or less (Table 2). Mean distance from the coast to fishing grounds was significantly different for the ZEPA and Tribugá (Kruskal-Wallis, Z = 13.5, P < 0.001), where average figures were 2.28 km ± 0.07 within the ZEPA and 1.25 km ± 0.03 in Tribugá (Figure 3A), with a maximum distance of 13.8 and 7.9 km, respectively. In Tribugá, 32 fishing grounds were located inside mangroves, with none for the ZEPA. Figure 3B shows that mean distance travelled increased with engine size.
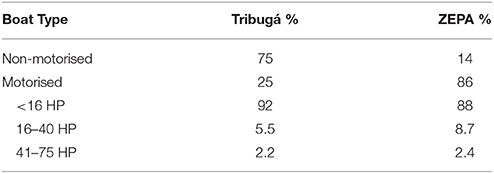
Table 2. Proportion of fishing trips associated with different types of boats in the fisheries of the Exclusive Zone of Artisanal Fishing (ZEPA) and the Gulf of Tribugá between 2010 and 2013.
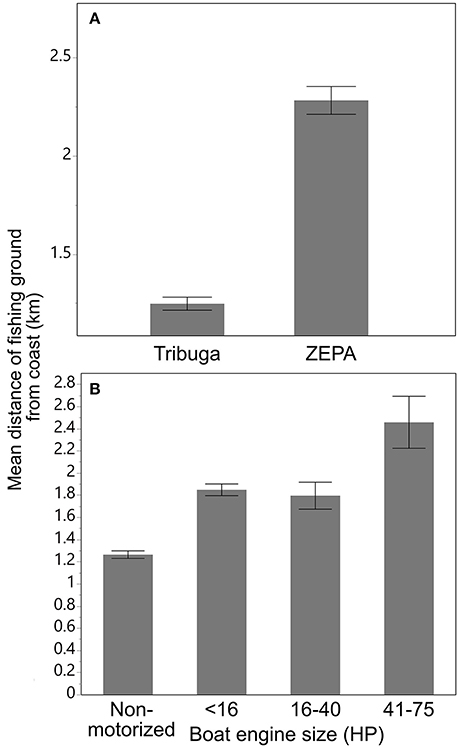
Figure 3. (A) Mean distance from fishing grounds to the coast for the Exclusive Zone of Artisanal Fishing (ZEPA) and the Gulf of Tribugá, and (B) proximity of fishing grounds to the coast in relation to boat engine size in Northern Chocó region between 2010 and 2013. Error bars represent the SE of the mean.
Mean annual CPUE (kg/fisher/day) indicate that the ZEPA and Tribugá have diverged over time, with CPUE increasing 50% in 3 consecutive years in the ZEPA after management was introduced in 2008, compared with a stable trend following an initial decline across the 4 years of sampling in Tribugá (Figure 4). Mean monthly CPUE was significantly different between years sampled in ZEPA (Wilcoxon, χ2 = 19.33, DF = 2, P = < 0.001), and similarly for Tribugá (Wilcoxon, χ2 = 22.93, DF = 3, P = < 0.001) with 2010 showing the largest difference.
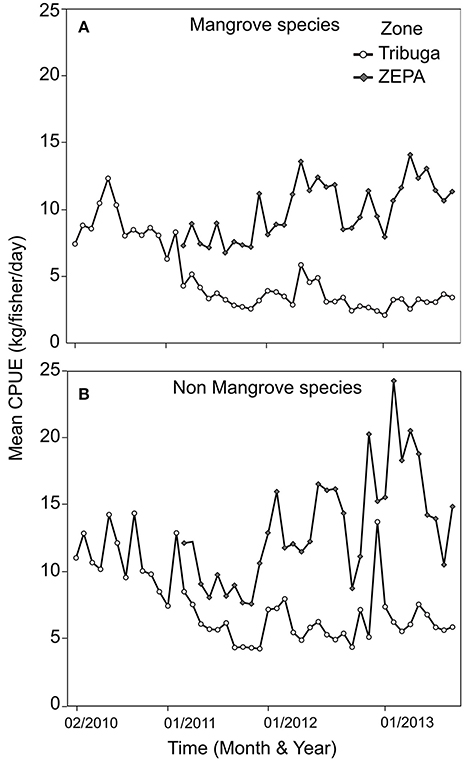
Figure 4. Temporal changes in mean Catch per Unit Effort (CPUE) for (A) mangrove and (B) non-mangrove associated species, showing separate trend lines for the Exclusive Zone of Artisanal Fishing (ZEPA) (grey diamonds) and the Gulf of Tribugá (white circles) between 2010 and 2013.
Exploring Management Effects on Mangrove Dependent Species
In the linear mixed effects models, for all three response variables (CPUE, number of individuals and weight landed) the random structure that best supported the data was the one that accounted for all proposed random effects (Supplementary Table 4) as it had the lowest AIC value. Model selection tables are included as Supplementary Tables 4, 5, and the final model summary table in Supplementary Table 6. The most parsimonious fixed effects models, as determined by AIC, also contained all main effects (fishing zone, gear, mangrove dependency, and boat type), and the interactions between management zone and the other independent variables (Table 3, Supplementary Table 5). Nevertheless, for CPUE the second-best model, as determined by the AIC, does not include the interaction between management zone and mangrove dependence (Supplementary Table 5), which might suggest a relatively weak explanatory power of that interaction.
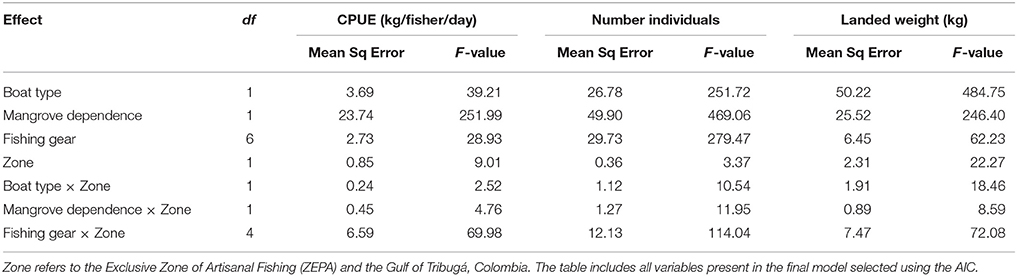
Table 3. Final mixed effects models for the response variables catch per unit effort (CPUE), number of individuals landed (N), and weight of individuals landed (kg).
In the model for CPUE, mangrove dependence and the interaction between management zone and fishing gear had the strongest effect explaining the variation in the data (Table 3). In the case of number of individuals, mangrove dependence had the strongest overall effect but all variables contributed similarly to explaining the variation, in particular we observed important differences to the use of different fishing gear across both management zones (Table 3). Finally, in the model constructed for weight landed, boat type and the interaction between management zone and fishing gear were the variables that explained most of the variation with comparable contribution of mangrove dependence (Table 3). Models of CPUE, number of individuals, and weight landed had a conditional R2 value of 0.56, 0.85, and 0.59, and a marginal R2 value of 0.11, 0.07, and 0.18, respectively.
CPUE and landed weight of non-mangrove species showed higher values than mangrove species (Figure 4). This was corroborated by the predictions of the mixed effect model (Figure 5A, Supplementary Table 7). However, the model also highlighted important differences in fishing methods, with manual collection and seine net showing the highest values for all variables. Predicted mean landings weight and CPUE were consistently higher for the ZEPA than Tribugá in all cases (Figures 5A,E). Motorised boats exhibited higher mean landing weight and CPUE than non-motorised crafts across both management zones (Figures 5B,F). Mean number of individuals landed followed similar trends, with higher number of individuals landed in the ZEPA and by motorised boats. However, an exception was found in non-motorised boats of Tribugá landing a higher number of individuals than the ZEPA (Figures 5D). This can be attributed to the effect of the large numbers of mangrove species captured by seine net and manual collection (Figure 5C). Ark clam (Anadara tuberculosa) was the most abundant species landed by non-motorised boats with 39% of the total, followed by spotted rose snapper (Lutjanus guttatus) with 9.7%, green jack (Caranx caballus) with 9.1%, and flathead grey mullet (Mugil cephalus) with 8.6%.
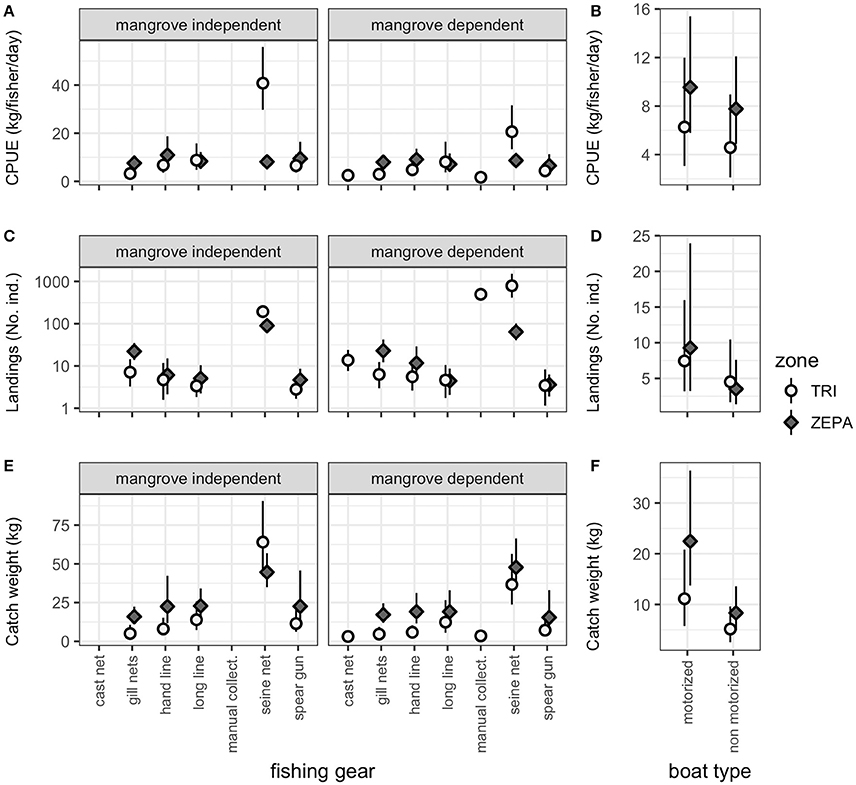
Figure 5. Predicted mean values from linear mixed effects models across management zone, fishing method, and species mangrove dependence for (A) CPUE (kg/fisher/day), (C) number of individuals landed, and (E) catch weight landed (kg). Predicted mean values are also presented across management zone and boat type for (B) CPUE (kg/fisher/day), (D) number of individuals landed, and (F) catch weight landed (kg). Data shows trends for fisheries landings collected between 2010 and 2013 from the Exclusive Zone of Artisanal Fishing (ZEPA) (grey diamonds) and the Gulf of Tribugá (white circles), Colombia. Number of individuals landed is given on a log scale. Vertical lines represent confidence intervals and are asymmetrical because data was modelled using log-normal distributions back transformed to a linear scale for representation. Thus, confidence intervals are symmetric in the logarithmic space but appear asymmetric in a linear scale.
Discussion
Small-scale fisheries in Northern Chocó target a diverse array of inshore and offshore species using hand lines as the main gear. The ZEPA, where area-based management was established in 2008, showed significantly higher fisheries productivity than Tribugá, with CPUE increasing by 50% within the 3 years of sampling, despite evidence of gillnets still in use. Use of motorised boats was more prevalent in the ZEPA and were used to fish further offshore, contrasting with Tribugá, where non-motorised boats fishing closer to the shore were predominant. Non-mangrove species dominated landings by weight in the ZEPA and had higher CPUE than mangrove species. However, mangrove species were particularly important by number of landed individuals for paddle boats in Tribugá. Hence, it appears that in the ZEPA, management has driven fishing effort away from coastal habitats, and into pelagic zones, thereby increasing productivity and reducing pressure on mangrove resources. Results from the mixed effects model suggested that area-based management in the ZEPA is related to the increase in CPUE. However, the effects of displacement of the industrial shrimp trawling fleet to Tribugá, cannot be ignored as a potential factor in driving a reduced CPUE of the artisanal fleet there.
Temporal trajectories indicate that CPUE increased in the ZEPA through to the end of the sampling period, whereas the opposite was true from the first year of sampling in Tribugá, where there was no management (Figure 4). This may suggest that the establishment of the ZEPA in 2008 has led to improvements in fish stocks following past overfishing through (i) exclusion of industrial fishing vessels and (ii) prohibition of gillnets. A similar recovery pattern was observed in Chile, where fisheries management was implemented in the sea snail (Concholepas concholepas) fishery following its collapse between 1989 and 1992 (San Martin et al., 2010). Ten years after management was established, sea snail abundance and mean sizes of individual organisms increased, stabilizing catches and leading to increased public and private benefits (González et al., 2006; San Martin et al., 2010). In contrast, Tribugá fishers claimed that the establishment of the ZEPA caused the displacement of the industrial fleet, after witnessing an increase in shrimp trawling boats in their waters (Díaz and Caro, 2016). Fishing displacement is a known social consequence of the implementation of protected areas (Sen, 2010; Cinner et al., 2014; Bennett et al., 2015; Chollett et al., 2015), when their designation excludes a group of fishers from access to their former fishing grounds (Charles, 2009). Therefore, the greater pressure (fishing effort) displaced to the area left open to fishing can generate resource depletion, habitat degradation and/or socioeconomic consequences because of lower fishery profits (Hiddink et al., 2006; Gimpel et al., 2013). Hiddink et al. (2006) investigated the effects of area closures on benthic communities in the North Sea, and found that through displacement of effort some closures had negative effects on the overall biomass, production, and species richness. This displacement of the industrial fleet's fishing activity may have caused the decline of CPUE of artisanal fisheries in Tribugá.
Former access to the ZEPA by shrimp trawlers likely resulted in significant habitat degradation and bycatch, as seen elsewhere (Freese et al., 1999; Hutchings and Reynolds, 2004; Myers and Worm, 2005; Hinz et al., 2009). Conflicts between industrial and artisanal fisheries in the same geographical space are a common issue in small-scale fisheries because of competition for resources (Bennett et al., 2001; Salas et al., 2007), and along with the use of gillnets, is one of the top problems faced by fishers in Colombia (Saavedra-Díaz et al., 2015, 2016). Declining trends of CPUE seen in the whitemouth croaker fishery in Uruguay were attributed to industrial fisheries activities overlapping spatially with small-scale fisheries (Horta and Defeo, 2012). Given the indication that shrimp trawling may have intensified in Tribugá following exclusion from the ZEPA (Díaz and Caro, 2016), research is needed to establish how this displaced fishing effort contributed to the decline in fish stocks in Tribugá. In the ZEPA, the formalisation of co-management structures and the associated reduction in fishing pressure from industrial boats, may have enhanced success of gear-based management measures (Díaz and Caro, 2016).
When fisheries move from open access conditions to organized entities and ultimately territorial use rights are applied, associated economic revenues have been recorded up to five or 10 times higher than those obtained in open access resource use (Wilen et al., 2012). In Japan and Chile for example, a significant amount of the rent generated in the fisheries comes from market value improvements and better marketing opportunities via organization and cooperation (Cancino et al., 2007). Harvesting decisions in Chile's sea snail fishery moved away from unstructured deals with buyers to predictable sales, based not only on product quantity, but quality and market price (Cancino et al., 2007). In Colombia, a commercial connection between restaurants in Bogotá and fisheries cooperatives was established soon after the ZEPA's formation, and has developed over recent years (Sáenz Pacheco, 2014) with the support of many non-governmental organizations and institutions (Cobos et al., 2016). Restaurants buy “responsibly-caught fish” directly from cooperatives in the ZEPA at a much more profitable price for fishers (Sáenz Pacheco, 2014). This shorter market chain creates an incentive to diversify the fishery, targeting higher value species, with larger, more powerful boats (Cobos et al., 2016). In Northern Chocó, offshore sites harbour large pelagic species such as tunas, where productivity peaks in certain seasons according to specific fisheries (e.g., sardines) (Pereira Velásquez, 1993; Zapata et al., 2007). According to the mixed effects model, the high mean weight landed of non-mangrove dependent species seen in the ZEPA is likely to be related to the dominance of motorised boats in this region (Table 2, Figure 5) as they operate further from the coast than non-motorised boats (Figure 3B). Tuna dominated landings by weight in the ZEPA, representing more than 50% of the total weight landed during the study period (Figure 2). Given that mangrove dependence was an important factor explaining CPUE in the mixed model, and targeting valuable pelagic fish is incentivized in the ZEPA, the increase in CPUE may represent the combined effects of shifting target species and management.
Tribugá's fishers have voiced their concerns over the decline of their catches and the increased sightings of trawlers. Following conflict with industrial fishers, stakeholders in the Gulf of Tribugá established the “Regional District of Integrated Management Golfo de Tribugá-Cabo Corrientes” in 2014 (Díaz and Caro, 2016), introducing a spatial and temporal control over the activity of trawlers. The exclusion of the industrial shrimp trawler fleet from Tribugá may bring about a stock recovery, but the implications of a ban on gillnets for fisher communities with limited livelihood options and adaptive capacity are unknown, and likely to be variable (López Angarita et al., 2014; Tilley and López-Angarita, 2016). Furthermore, the ZEPA is also the result of financial investment to push fishing effort offshore and the development of fair trade style market chains between cooperatives and restaurants. Further studies in Northern Chocó should focus on comparing the effects of both management approaches (i.e. complete exclusion of trawlers vs. regulation of their activity) in the productivity of small-scale fisheries.
For the Gulf of California, Aburto-Oropeza et al. (2008), found that 32% of commercial fishery species were mangrove dependent, while this figure was 80% for commercial and recreational species captured in a study in Florida by Hamilton and Snedaker (1984). In this study around half of species landed were classified as mangrove dependent. Landings of non-motorised boats in Tribugá were dominated by mangrove dependent species in terms of number of individuals across most gear types, though their contribution by weight was low compared to non-mangrove species (Figure 5E). The limited range of these boats means they operate near the coast and inside mangroves in Tribugá, so landed individuals of mangrove dependent species are likely to be juveniles, considering the nursery function of mangroves (Nagelkerken et al., 2002; Mumby et al., 2004; Saenger and Funge-Smith, 2013). Given the declining trends of fish abundance in Tribugá, it may be beneficial to reduce fishing pressure around mangroves to avoid growth overfishing. Since non-motorised boats represent 75% of the fleet in Tribugá, nearshore and mangrove fish populations are clearly an important resource, so fostering their recovery is necessary. If boat type is considered a proxy for financial capacity the consequences of fishing displacement may have disproportionally large negative social and economic effects on the livelihoods of already vulnerable local communities, who rely heavily on coastal resources for food security.
The higher catch rates demonstrated for ZEPA may also be due to an underlying geographical bias, as the Gulf of Tribugá has more estuaries compared to the ZEPA (Prahl et al., 1990). Despite the fishing location/ground being included as random factor in the mixed effects model, more studies are needed to determine how the zones differ in terms of fisheries productivity. Unraveling the effects of management from other factors, such as sampling effort and environmental variability is challenging given the lack of replication in this case study, where only two regions have been compared and no data prior to management exists. The large difference in sampling effort between the regions may also affect comparisons, with more trips sampled in Tribugá. The small population size and higher fishing effort of Tribugá communities compared to the ZEPA, may suggest limited economic opportunities outside of fishing. Greater research on the socio-economic drivers of fishing pressure would provide valuable insight into management alongside these data. Regardless, the dataset used is unique in terms of length of sampling and detail of collected information for this region. Furthermore, in the new management area established in Tribugá, these data and analyses will prove invaluable as a baseline against which to track management performance.
Ecological, economic, and social trade-offs commonly emerge among multiple ecosystem goods and services as a result of fisheries management (Voss et al., 2014). In area based fisheries management, the challenge is conciliating potential conflicting interests related to resource use, conservation, and in the case of the ZEPA, their distributional equity. For Tribugá a local community initiative and the support of government and private organizations, have allowed the protection of artisanal fishers' livelihoods in the face of the increased activity by industrial fishers, but in other regions where fishing displacement occurs, this might not be the case. In the process of management planning, it is necessary to contemplate the spatial and temporal redistribution of effort and use this information to predict potential impacts on livelihoods and natural resources (Hiddink et al., 2006). This planning, should prioritize small-scale fisheries over industrial fisheries given that the artisanal sector not only generates less impact with lower catches and higher selectivity, but also provides social benefits to a wider sector of the community through job creation and contributes more to local economies than the industrial sector (Belhabib et al., 2017).
Leadership, social cohesion and co-management have been shown to improve well-being of small-scale fishing communities (Cinner et al., 2012) and promote successful fisheries (Gutiérrez et al., 2011). In the isolated fishing communities from Northern Chocó, where government resources are limited, the ZEPA has empowered local fishers through a system of fishing rights, management responsibilities, and rewards. Regardless of if the ZEPA is successful in terms of enhancing fisheries productivity, its importance lies in facilitating the organization and cooperation between local community members and leaders. In the ZEPA, the management system may have reduced fishing pressure on coastal stocks, such as mangrove fisheries, by incentivising fishers to target offshore resources. As such, diverting fishing effort to higher productivity species offshore, a management system can effectively enhance mangrove fisheries protection for poor coastal fishers who rely on them for subsistence livelihoods. Finally, further socio-economic research needs to accompany fishery assessments to understand the effects of management on food security and poverty alleviation, which are areas of particular concern for the region studied.
Author Contributions
JL-A and CR designed the study. JL-A, AT, and EC analysed data. JD was responsible for designing and implementing the program of participatory monitoring in fishing communities. JL-A, AT, JH, JD, EC, and CR wrote sections of the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This research was funded by fellowships awarded to JL-A by the Colombian Department of Science and Technology—COLCIENCIAS (Crédito—Beca Francisco José de Caldas), the Schlumberger Foundation Faculty for the Future Programme, the World Wildlife Fund's Russell E. Train Education for Nature fellowship, and the Save our Seas Foundation (SOSF2013APP2N). Thanks to C. Viera and G. Gonzalez from Marviva Foundation for their collaboration. Special thanks to communities in Chocó for their willingness to participate and contribute to this work. Finally, we gratefully acknowledge valuable comments from Lewis Le Vay and Robert Marchant on this manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2018.00023/full#supplementary-material
References
Aburto-Oropeza, O., Ezcurra, E., Danemann, G., Valdez, V., Murray, J., and Sala, E. (2008). Mangroves in the Gulf of California increase fishery yields. Proc. Natl. Acad. Sci. U.S.A. 105, 10456–10459. doi: 10.1073/pnas.0804601105
Álvarez-León, R., and Polanía, J. (1996). Los Manglares del Caribe Colombiano: síntesis de su Conocimiento. Rev. Acad. Colomb. Cienc. 20, 447–464.
Baos Estupiñán, R. A., and Zapata, L. A. (2011). “Análisis de la flota pesquera industrial del Pacífico colombiano radicada en el puerto de Buenaventura durante los a-os 2006 a 2009,” in Diagnóstico de las Principales Pesquerías del Pacífico Colombiano, eds J. M. Díaz, C. A. Vieira and G. J. Melo (Bogotá: Fundación Marviva), 31–48.
Bates, D., Mächler, M., Bolker, B., and Walker, S. (2015). Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67, 1–48. doi: 10.18637/jss.v067.i01
Belhabib, D., Greer, K., and Pauly, D. (2017). Trends in industrial and artisanal catch per effort in West African fisheries. Conserv. Lett. 107, 131–110. doi: 10.1111/conl.12360
Béné, C. (2009). Are fishers poor or vulnerable? Assessing economic vulnerability in small-scale fishing communities. J. Dev. Stud. 45, 911–933. doi: 10.1080/00220380902807395
Bennett, E., Neiland, A., Anang, E., Bannerman, P., Atiq Rahman, A., Huq, S., et al. (2001). Towards a better understanding of conflict management in tropical fisheries: evidence from Ghana, Bangladesh and the Caribbean. Mar. Policy 25, 365–376. doi: 10.1016/S0308-597X(01)00022-7
Bennett, N. J., Govan, H., and Satterfield, T. (2015). Ocean grabbing. Mar. Policy 57, 61–68. doi: 10.1016/j.marpol.2015.03.026
Blaber, S. J. M., Brewer, D. T., and Salini, J. P. (1989). Species composition and biomasses of fishes in different habitats of a tropical Northern Australian estuary: their occurrence in the adjoining sea and estuarine dependence. Estuar. Coast. Shelf Sci. 29, 509–531. doi: 10.1016/0272-7714(89)90008-5
Blanco, J. F., Quiceno, P. A., Jiménez, L. F., and Turbay, S. (2011). Estrategias de Manejo Sostenibles de la Actividad Pesquera en el PNN Utría. Unidad Administrativa Especial del Sistema de Parques Nacionales Naturales (UAESPENN), Fondo de Patrimonio Natural y Corporación Académica Ambiental de la Universidad de Antioquia.
Burnham, K. P., and Anderson, D. R. (2002). Information and Likelihood Theory: A Basis for Model Selection and Inference. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd Edn. New York, NY: Springer.
Camacho, J., Gómez, C. E., Campos, C., and Vieira, C. A. (2000). Proyecto “Comunidades Indígenas y Negras: Desarrollo Regional para la Conservación de los Recursos Naturales en el Chocó Biogeográfico - Colombia.” Fundación Natura, Santa Fe de Bogotá.
Cancino, J. P., Uchida, H., and Wilen, J. E. (2007). TURFs and ITQs: collective vs. individual decision making. Mar. Res. Econ. 22, 391–406. doi: 10.1086/mre.22.4.42629569
Carrasquilla-Henao, M., and Juanes, F. (2016). Mangroves enhance local fisheries catches: a global meta-analysis. Fish Fish. 18, 79–93. doi: 10.1111/faf.12168
Charles, A. (2009). Human dimensions of marine protected areas. ICES J. Mar. Sci. 66, 6–15. doi: 10.1093/icesjms/fsn182
Chollett, I., Box, S. J., and Mumby, P. J. (2015). Quantifying the squeezing or stretching of fisheries as they adapt to displacement by marine reserves. Conserv. Biol. 36, 166–175. doi: 10.1111/cobi.12573
Cinner, J. E., Daw, T., Huchery, C., Thoya, P., Wamukota, A., Cedras, M., et al. (2014). Winners and losers in marine conservation: fishers' displacement and livelihood benefits from marine reserves. Soc. Nat. Resour. 27, 994–1005. doi: 10.1080/08941920.2014.918229
Cinner, J. E., McClanahan, T. R., Graham, N. A., Daw, T. M., Mukminin, A., Feary, D. A., et al. (2012). Comanagement of coral reef social-ecological systems. Proc. Natl. Acad. Sci. U.S.A. 109, 5219–5222. doi: 10.1073/pnas.1121215109
Cobos, A. J., Due-as, J. D., and Velandia, M. C. (2016). “Aspectos económicos de la pesca artesanal,” in La pesca Artesanal en el Norte del Pacífico Colombiano: Un Horizonte Ambivalente, eds J. M. Díaz, L. Guillot and C. M. Velandia (Bogotá: Fundación MarViva), 91–103.
Díaz, J. M., and Caro, N. (2016). “El mar como territorio y la pesca como actividad tradicional en el territorio Chocoano,” in La Pesca Artesanal en el Norte del Pacífico Colombiano: Un Horizonte Ambivalente, eds J. M. Díaz, L. Guillot and C. M. Velandia (Bogotá: Fundación MarViva), 29–43.
Díaz, J. M., and Galeano, J. (2016). “El entorno biogeofísico,” in La Pesca Artesanal en el Norte del Pacífico Colombiano: Un Horizonte Ambivalente, eds J. M. Díaz, L. Guillot and C. M. Velandia (Bogota: Fundación MarViva), 15–27.
Díaz, J. M., Vieira, C., and Melo, G. J. (2011). Diagnóstico de las Principales Pesquerías del Pacífico Colombiano. Bogotá: Fundación MarViva.
Finegold, C. (2009). The Importance of Fisheries and Aquaculture to Development. Stockholm: The Royal Swedish Academy of Agriculture and Forestry.
Freese, L., Auster, P. J., Heifetz, J., and Wing, B. L. (1999). Effects of trawling on seafloor habitat and associated invertebrate taxa in the Gulf of Alaska. Mar. Ecol. Prog. Ser. 182, 119–126. doi: 10.3354/meps182119
García, C. (2010). Diagnóstico de las Áreas Marinas y Costeras Protegidas, y de las Áreas de Manejo en el Pacífico Colombiano. Colombia: Fundación MarViva.
Gell, F. R., and Roberts, C. M. (2003). Benefits beyond boundaries: the fishery effects of marine reserves. Trends Ecol. Evol. 18, 448–455. doi: 10.1016/S0169-5347(03)00189-7
Gimpel, A., Stelzenmüller, V., Cormier, R., Floeter, J., and Temming, A. (2013). A spatially explicit risk approach to support marine spatial planning in the German, EEZ. Mar. Environ. Res. 86, 56–69. doi: 10.1016/j.marenvres.2013.02.013
González, J., Stotz, W., Garrido, J., Orensanz, J. M., Parma, A. M., Tapia, C., et al. (2006). The Chilean TURF system: how is it performing in the case of the loco fishery? Bull. Mar. Sci. 78, 499–527. Available online at: http://www.ingentaconnect.com/contentone/umrsmas/bullmar/2006/00000078/00000003/art00007
Gutiérrez, N. L., Hilborn, R., and Defeo, O. (2011). Leadership, social capital and incentives promote successful fisheries. Nature 470, 386–389. doi: 10.1038/nature09689
Hamilton, S. L., and Snedaker, S. C. (1984). Handbook for Mangrove Area Management. UNEP and East West Center. Honolulu: Environment and Policy Institute.
Hiddink, J., Hutton, T., Jennings, S., and Kaiser, M. (2006). Predicting the effects of area closures and fishing effort restrictions on the production, biomass, and species richness of benthic invertebrate communities. ICES J. Mar. Sci. 63, 822–830. doi: 10.1016/j.icesjms.2006.02.006
Hinz, H., Prieto, V., and Kaiser, M. J. (2009). Trawl disturbance on benthic communities: chronic effects and experimental predictions. Ecol. Appl. 19, 761–773. doi: 10.1890/08-0351.1
Horta, S., and Defeo, O. (2012). The spatial dynamics of the whitemouth croaker artisanal fishery in Uruguay and interdependencies with the industrial fleet. Fish. Res. 125–126, 121–128. doi: 10.1016/j.fishres.2012.02.007
Hutchings, J. A., and Reynolds, J. D. (2004). Marine fish population collapses: consequences for recovery and extinction risk. Bioscience 54, 297–309. doi: 10.1641/0006-3568(2004)054[0297:MFPCCF]2.0.CO2
Lindop, A. M. M., Chen, T., Zylich, K., and Zeller, D. (2015). A Reconstruction of Colombia's Marine Fisheries Catches. Working Paper Series. Fisheries Centre. Vancouver: University of British Columbia, 1–16.
López Angarita, J., Moreno-Sánchez, R., Maldonado, J. H., and Sánchez, J. A. (2014). Evaluating linked social-ecological systems in marine protected areas. Conserv. Lett. 7, 241–252. doi: 10.1111/conl.12063
Mills, D. J., Westlund, L., Graaf, G., de Kura, Y., Willman, R., and Kelleher, K. (2011). “Under-reported and undervalued: small-scale fisheries in the developing world,” in Small-scale Fisheries Management: Frameworks and Approaches for the Developing World, eds R. S. Pomeroy and N. L. Andrew (Wallingford: CABI), 1–15
Mumby, P. J., Edwards, A. J., Arias-González, J. E., Lindeman, K. C., Blackwell, P. G., Gall, A., et al. (2004). Mangroves enhance the biomass of coral reef fish communities in the Caribbean. Nature 427, 533–536. doi: 10.1038/nature02286
Myers, N., Mittermeier, R. A., Mittermeier, C. G., da Fonseca, G. A. B., and Kent, J. (2000). Biodiversity hotspots for conservation priorities. Nature 403, 853–858. doi: 10.1038/35002501
Myers, R. A., and Worm, B. (2005). Extinction, survival or recovery of large predatory fishes. Philos. Trans. R. Soc. Lond. B Biol. Sci. 360, 13–20. doi: 10.1098/rstb.2004.1573
Nagelkerken, I., Kleijnen, S., Klop, T., Van Den Brand, R., La de Moriniere, E. C., and Van Der Velde, G. (2001). Dependence of Caribbean reef fishes on mangroves and seagrass beds as nursery habitats: a comparison of fish faunas between bays with and without mangroves/seagrass beds. Mar. Ecol. Prog. Ser. 214, 225–235. doi: 10.3354/meps214225
Nagelkerken, I., Roberts, C. M., Van Der Velde, G., Dorenbosch, M., Van Riel, M. C., Cocheret de la Morinière, E., et al. (2002). How important are mangroves and seagrass beds for coral-reef fish? The nursery hypothesis tested on an island scale. Mar. Ecol. Progr. Series 244, 299–305. doi: 10.3354/meps244299
Neira, A., Díaz, J. M., Gonzalez, G., Velandia, C. M., and Melo, G. J. (2016). “Diagnóstico de las pesquerías artesanales en el norte del Pacífico chocoano,” in La pesca Artesanal en el Norte del Pacífico Colombiano: Un Horizonte Ambivalente, eds J. M. Díaz, L. Guillot, and C. M. Velandia (Bogotá: Fundación MarViva), 69–89.
Pauly, D., and Zeller, D. (2015). Catch reconstructions reveal that global marine fisheries catches are higher than reported and declining. Nat. Commun. 7, 1–9. doi: 10.1038/ncomms10244
Pereira Velásquez, F. (1993). “La pesca en el Pacífico,” in Colombia Pacífico, ed P. Leyva (Bogota: Fondo para la Protección del Medio Ambiente José Celestino Mutis).
Prahl von, H., Cantera, J. R., and Contreras, R. (1990). Manglares y Hombres del Pacífico Colombiano. 1st Edn. Colombia: Editorial Presencia.
Rueda, M., Rico-Mejía, F., and Angulo, W. (2001). “Evaluación y manejo de la pesquería industrial de camarón de aguas someras en el Pacífico colombiano,” in Diagnóstico de las Principales Pesquerías del Pacífico Colombiano, eds J. M. Díaz, C. A. Vieira, and G. J. Melo (Bogotá: Fundación MarViva), 91–126.
Saavedra-Díaz, L. M., Pomeroy, R., and Rosenberg, A. A. (2016). Managing small-scale fisheries in Colombia. Marit. Stud. 15:621. doi: 10.1186/s40152-016-0047-z
Saavedra-Díaz, L. M., Rosenberg, A. A., and Martín-López, B. (2015). Social perceptions of Colombian small-scale marine fisheries conflicts: insights for management. Mar. Policy 56, 61–70. doi: 10.1016/j.marpol.2014.11.026
Saenger, P., and Funge-Smith (2013). A Review of Mangrove and Seagrass Ecosystems and Their Linkage to Fisheries and Fisheries Management. Bangkok: Food and Agriculture Organization of the United Nations regional office for Asia and the Pacific.
Sáenz Pacheco, G. (2014). Marviva, Pesca Responsable y Comunicación Para la Gobernanza en Bahía Solano y Nuquí (Pacifico Norte Colombiano). Barranquilla: Editorial Universidad del Norte.
Salas, S., Chuenpagdee, R., Seijo, J. C., and Charles, A. (2007). Challenges in the assessment and management of small-scale fisheries in Latin America and the Caribbean. Fish. Res. 87, 5–16. doi: 10.1016/j.fishres.2007.06.015
San Martin, G., Parma, A. M., and Orensanz, J. M. (2010). “The Chilean experience with territorial use rights in fisheries,” in Handbook of Marine Fisheries Conservation and Management, eds R. Q. Grafton, R. Hilborn, D. Squires, M. Tait, and M. Williams (New York, NY: Oxford University Press), 324–337.
Sen, S. (2010). Developing a framework for displaced fishing effort programs in marine protected areas. Mar. Policy 34, 1171–1177. doi: 10.1016/j.marpol.2010.03.017
Tilley, A., and López-Angarita, J. (2016). Predicting vulnerability to management changes in data-limited, small-scale fisheries. Mar. Policy 72, 211–218. doi: 10.1016/j.marpol.2016.05.034
Vieira, C. A., Diaz Granados, M. A., and Díaz, J. M. (2016). “Ordenamiento y manejo pesquero en la costa norte del Pacifico Chocoano,” in La Pesca Artesanal en el Norte del Pacífico colombiano: Un horizonte ambivalente, eds J. M. Díaz, L. Guillot, and C. M. Velandia (Bogotá: Fundación MarViva), 45–57.
Villalba, J. C. (2005). Los Manglares en el Mundo y en Colombia. Bogotá: Sociedad Geográfica de Colombia. Academia de Ciencias Geográficas.
Voss, R., Quaas, M. F., Schmidt, J. O., Tahvonen, O., Lindegren, M., et al. (2014). Assessing social – ecological trade-offs to advance ecosystem-based fisheries management. PLOS ONE 9:e107811. doi: 10.1371/journal.pone.0107811
Wang, C., and Fiedler, P. C. (2006). ENSO variability and the eastern tropical Pacific: a review. Prog. Oceanogr. 69, 239–266. doi: 10.1016/j.pocean.2006.03.004
Keywords: artisanal fisheries, mangroves, Tropical Eastern Pacific, fishing displacement, fisheries management, catch per unit effort
Citation: López-Angarita J, Tilley A, Díaz JM, Hawkins JP, Cagua EF and Roberts CM (2018) Winners and Losers in Area-Based Management of a Small-Scale Fishery in the Colombian Pacific. Front. Mar. Sci. 5:23. doi: 10.3389/fmars.2018.00023
Received: 02 August 2017; Accepted: 17 January 2018;
Published: 06 February 2018.
Edited by:
Steven W. Purcell, Southern Cross University, AustraliaReviewed by:
Louise Teh, University of British Columbia, CanadaOmar Defeo, Universidad de la República de Uruguay, Uruguay
Copyright © 2018 López-Angarita, Tilley, Díaz, Hawkins, Cagua and Roberts. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Juliana López-Angarita, anVsaWFuYWxvcDE0QGdtYWlsLmNvbQ==