
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
HYPOTHESIS AND THEORY article
Front. Mar. Sci. , 10 January 2017
Sec. Global Change and the Future Ocean
Volume 3 - 2016 | https://doi.org/10.3389/fmars.2016.00290
 Manuel Barange1*†
Manuel Barange1*† Momme Butenschön1
Momme Butenschön1 Andrew Yool2
Andrew Yool2 Nicola Beaumont1
Nicola Beaumont1 Jose A. Fernandes1
Jose A. Fernandes1 Adrian P. Martin2
Adrian P. Martin2 J. Icarus Allen3
J. Icarus Allen3To predict the impacts of climate change it is essential to understand how anthropogenic change alters the balance between atmosphere, ocean, and terrestrial reservoirs of carbon. It has been estimated that natural atmospheric concentrations of CO2 are almost 200 ppm lower than they would be without the transport of organic material produced in the surface ocean to depth, an ecosystem service driven by mechanisms collectively referred to as the biological carbon pump. Here we quantify potential reductions in carbon sequestration fluxes in the North Atlantic Ocean through the biological carbon pump over the twenty-first century, using two independent biogeochemical models, driven by low and high IPCC AR5 carbon emission scenarios. The carbon flux at 1000 m (the depth at which it is assumed that carbon is sequestered) in the North Atlantic was estimated to decline between 27 and 43% by the end of the century, depending on the biogeochemical model and the emission scenario considered. In monetary terms, the value of this loss in carbon sequestration service in the North Atlantic was estimated to range between US$170–US$3000 billion in abatement (mitigation) costs and US$23–US$401 billion in social (adaptation) costs, over the twenty-first century. Our results challenge the frequent assumption that coastal habitats store more significant amounts of carbon and are under greater threat. We highlight the largely unrecognized economic importance of the natural, blue carbon sequestration service provided by the open ocean, which is predicted to undergo significant anthropogenic-driven change.
Anthropogenic emissions of CO2, particularly through fossil-fuel combustion and land-use changes, have driven increases in its atmospheric concentration from pre-industrial conditions (Hartmann et al., 2013). To date, ~25% of anthropogenic CO2 emissions have entered the ocean (Khatiwala et al., 2013), driven primarily by physicochemical processes, contributing to the reservoir of dissolved inorganic carbon (DIC). A significant but less well-understood component of the ocean's natural carbon cycle is that driven by marine biota. Through a suite of biogeochemical processes collectively known as the biological carbon pump (BCP; Broecker, 1982; Volk and Hoffert, 1985), a fraction of the organic carbon produced in sunlit surface waters is transported to the deep ocean via gravitational sinking and vertical migration. The strength of this pump depends on both the amount of primary production and how this is attenuated down the water column by respiration. The depth and location of this respiration controls how quickly the released DIC comes back into contact with the atmosphere, where it can potentially outgas as CO2 (Robinson et al., 2014). The net effect of the biological pump is to elevate the ocean's reservoir of DIC above that expected from CO2 solubility alone. About 1% of primary production is thought to sink into the deep ocean interior or into seafloor sediments (Sanders et al., 2014), where it may be sequestered for hundreds to thousands of years.
As a component of the Earth's contemporary carbon cycle, carbon sequestration driven by the biological pump plays a role in producing the Earth's current, equable climate by contributing to the regulation of the balance of Carbon between the ocean and atmosphere. As such, the biological pump provides an ecosystem service, whose importance may be curtailed by anthropogenic climate change. Recent IPCC reports indicate that climate change will lead to a more stable density stratification in the ocean and a general slowing down of large-scale mixing and circulation, with both a reduced ocean uptake of anthropogenic carbon and a reduced supply of nutrients to ultimately drive the BCP (Sarmiento et al., 1998; Bopp et al., 2001; Heinze et al., 2015). The service provided by the BCP has an economic significance that is often poorly represented in the development of policy and management narratives (Luisetti et al., 2013). Monetary and non-monetary valuation can facilitate this narrative by allowing a comparison of costs with alternative ecosystem service provision and across future socio-economic scenarios, or with mitigation and adaptation interventions. There are currently very few studies which value offshore ecosystem services, and little effort has been devoted to understanding the economic cost of such a potential reduction in carbon sequestration service provision (e.g., Lutz and Martin, 2014; Rogers et al., 2014).
Here we computed changes in carbon export beyond 500 and 1000 m deep in the North Atlantic Ocean, as a function of low and high IPCC AR5 Representative Concentration Pathways (RCP 2.6 and 8.5; Rogelj et al., 2012), and accumulated these over the twenty-first century. We use the magnitude of the flux associated with the biological pump at 1000 m depth as a proxy for the ecosystem service provided by marine life in storing carbon in the North Atlantic Ocean, defining organic carbon as sequestered when it cannot return to the atmosphere for >100 years (e.g., within the span of our model projection; but see Robinson et al., 2014) or when it reaches depths >1000 m (IPCC, WGI, 2007; chapter 5; Lampitt et al., 2008; Guidi et al., 2015). The differences at depth from present-day fluxes are translated into economic values, using the social (adaptation) costs of carbon (Tol, 2005) and an abatement (mitigation) cost approach (DECC, 2011), to estimate the economic cost of changes in the North Atlantic ocean BCP.
We use two different state of the art biogeochemical ecosystem models of the North Atlantic, MEDUSA (Yool et al., 2013b) and ERSEM (Butenschön et al., 2016), simulated under identical climate forcing and with common geographical coverage (but with differential spatial resolution), to estimate the functioning of the BCP. We deliberately use two different models to provide a measure of uncertainty in our results. Both models have an appropriate description of the carbon cycle, i.e., primary and secondary production, respiration, the carbonate system, sedimentation, decomposition of organic matter, and air-sea fluxes of CO2, but with different representation of the microbial food-web, reflecting the lack of robust mechanistic understanding of underlying processes. Projections from both models were normalized by the present “baseline period” (1990–2009), to ensure we captured the strong interannual variability observed in North Atlantic carbon fluxes (Gruber et al., 2002).
MEDUSA (Model of Ecosystem Dynamics, nutrient Utilization, Sequestration and Acidification, Yool et al., 2013b) is an intermediate complexity plankton ecosystem model designed to have sufficient complexity to study key feedbacks between anthropogenically-driven changes and oceanic biogeochemistry. MEDUSA resolves a size-structured ecosystem of small (nanophytoplankton and microzooplankton) and large (microphytoplankton and mesozooplankton) components that explicitly includes the biogeochemical cycles of nitrogen, silicon, and iron nutrients as well as the cycles of carbon, alkalinity, and oxygen. Export production in MEDUSA-2 comprises explicit, slow-sinking particles with temperature-dependent remineralization, and implicit, fast-sinking particles that use biogenic silicon, and a parameterization of the calcification rain ratio (Ridgwell et al., 2007) to ballast sinking organic material (Armstrong et al., 2002). The entire World Ocean is simulated and the model output post-processed to the North Atlantic cut-out shown here. The model was initialized using standard biogeochemical climatological fields (Garcia et al., 2010; Key et al., 2014) and simulated under surface atmospheric forcing derived from HadGEM2-ES simulations performed for CMIP5. A single medium resolution (1°) NEMO-MEDUSA simulation ran for the “historical period” from 1860 to 2005, and was then split into two separate simulations running under IPCC RCPs 2.6 and 8.5 from 2006 to 2099 (Yool et al., 2013a). MEDUSA's export of organic material to the ocean interior is vulnerable both to stratification-mediated climate change and to calcification-mediated ocean acidification (Yool et al., 2013a).
The ERSEM (European Regional Seas Ecosystem Model, Butenschön et al., 2016) model provides a comparatively detailed description. It uses three phytoplankton size classes complemented by diatoms, three size classes of zooplankton, a class of heterotrophic bacteria, three size classes of particulate organic matter and three classes of dissolved matter with distinct degradation time scales. The model resolves the dynamical cycles of the major constituents, carbon, nitrogen, phosphorus, silicate, and iron, independently, and includes a sub-model for the carbonate system providing the fields of alkalinity. The sinking of organic material occurs in relation to the size of the organic material with correspondingly distinct remineralization time scales and the same parameterization of the rain ratio as used in MEDUSA. The NEMO-ERSEM projection is performed on a dedicated North Atlantic domain at a spatial resolution of 0.25°, extending from 20°S to 70–80°N allowing for some spatial buffer toward the model boundaries with respect to the core of the domain used for the analysis of this work in order to discard any artificial boundary effects. The simulation has used the same initial and atmospheric boundary conditions as the NEMO-MEDUSA simulation.
Both models were initialized from observationally derived climatological fields of inorganic nutrients and DIC taken from the World Ocean Atlas and GLODAP Carbon Synthesis Project. However, they differed in the path leading to the modeled present day conditions: the MEDUSA simulation was initialized in 1860 and run continuously up to the present day, while ERSEM was initialized in 1958 in hindcast mode. The 1970 conditions from the hindcast simulation where then used to initialize the climate model forced projection in 1970.
In order to identify any spurious model drift control simulations for both MEDUSA and ERSEM models were performed exposing the model system to external forcing that is not affected by climate change. In the case of ERSEM this was achieved by performing a simulation of the model under the atmospheric conditions of the decade 1970–1979 as used in the projection, but periodically repeated from 1980 to the end of the simulation period (2099) with the partial pressure of CO2 kept at a constant level corresponding to the initial value of the projection (1971). For MEDUSA a similar experiment was conducted, which due to the reduced resource requirements of this system was possible to extend to a longer period. In this case the control run was started as the projection in 1860, but exposed to the periodically repeated atmospheric conditions of 1860–1889 and the atmospheric pCO2 kept constant at the starting value of 1860 throughout the simulation. Results (see Supplementary Material) show that for both simulations the trend in vertical flux is clearly dominated by the climate change signal and not by model drift. Compared to the average present day fluxes of the control simulations the estimated drift is below 10% for both models, while relative to the change signal of the projection simulation it amounts to 10% for the MEDUSA simulation, while for ERSEM it is around 20% due to the smaller change signal in the projection of this model. Thus, for both model systems the projected changes can be attributed to ecosystem process changes under the climate scenarios.
To reflect the variability in valuation methodology two carbon prices are applied to carbon export fluxes below 1000 m. Firstly, a social cost of carbon (SCC, Tol, 2005) of $50 per tC (2005 prices), uprated each year by 2% to reflect the incremental damage of each unit of carbon as temperature rises (Stern, 2006; Defra, 2007). While the price could potentially be uprated by a higher percentage under the RCP8.5 scenario, this is not fully supported by current literature. Thus, the established rate of 2% is applied to both RCP scenarios. SCC is a shadow price of carbon emissions and corresponds to the net present value of the cumulative, worldwide impact of one additional t of C emitted to the atmosphere today over its residence time in the atmosphere (van den Bergh and Botzen, 2015). Secondly, a non-traded mid marginal abatement cost (MAC) of £55 tCO2 (2014 prices) is applied (DECC, 2011), converted to US$/tC and also increasing over time at the same rate. The MAC method is a mitigation cost, assuming measures and related costs required to meet a specific emission reduction target, which are forecasted into the future. There is significant literature on the most appropriate discount rate to use (HM Treasury, 2011; van den Bergh and Botzen, 2015), as this rate is essential to determine what climate policy to pursue. In this analysis we used the current UK recommended social discount rate of 3.5% diminishing over time (Defra, 2007), consistent with recent valuation exercises (Luisetti et al., 2013).
Trends in North Atlantic carbon fluxes over the twenty-first century, normalized to 1990 estimates, indicate a continuous decline in export at 500 m and 1000 m depth. At 500 m deep the decline is consistent until ~2040, at which point the decline is larger for emissions scenario RCP8.5 and larger for MEDUSA than ERSEM (Figure 1). At 1000 m deep the differences between models and emissions are also consistent, but the decline is particularly evident in MEDUSA RCP8.5 (Figure 1). As discussed later, the differences between MEDUSA and ERSEM emerge from the absence of detailed mechanistic understanding of processes in the twilight zone (Sanders et al., 2014).
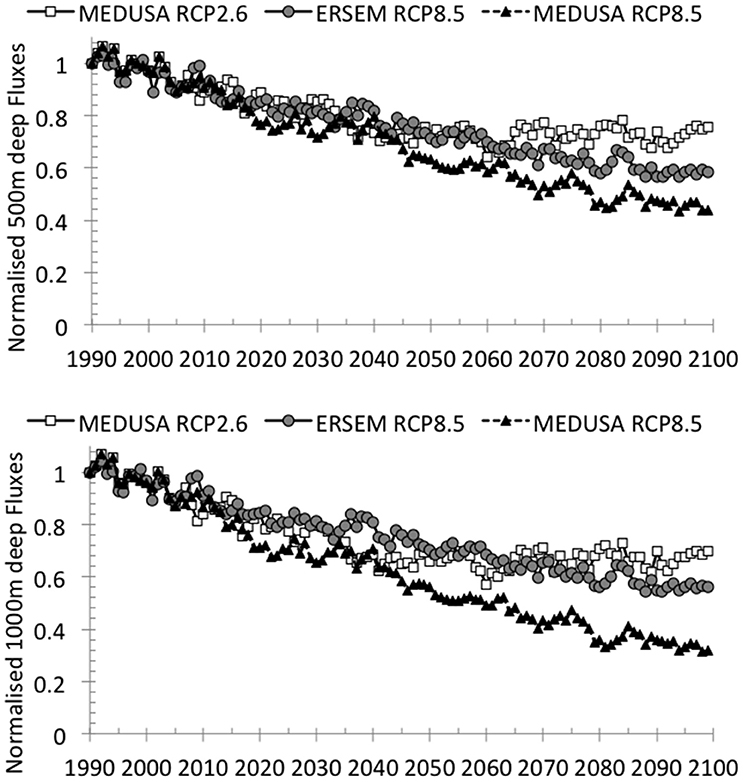
Figure 1. Five hundred meter deep (top) and 1000 m (bottom) annual C fluxes (PgC year−1), normalized according to 1990 estimates, by biogeochemical model and IPCC RCP emissions scenario.
Over the baseline period (1990–2009), the North Atlantic operated as a significant sink of organic carbon to the deep ocean (Figures 2A,B), but the declines in carbon export by the end of the century (Figure 1) are not geographically consistent. By the decade 2090–2099 carbon sequestration under both greenhouse gas emission scenarios is predicted to see a reduction compared to the period 1990–2009 (Figures 2C–F). This reduction is particularly evident at 500 m deep between 45 and 65°N, and off the North African coast (Figures 2C–E). Export fluxes at 1000 m depth are significantly lower and geographically more homogeneous, partially due to the exponential nature of POC decay, but with similar reductions in export to the ocean interior in the central North Atlantic by the end of the twenty-first century (Figures 2D–F).
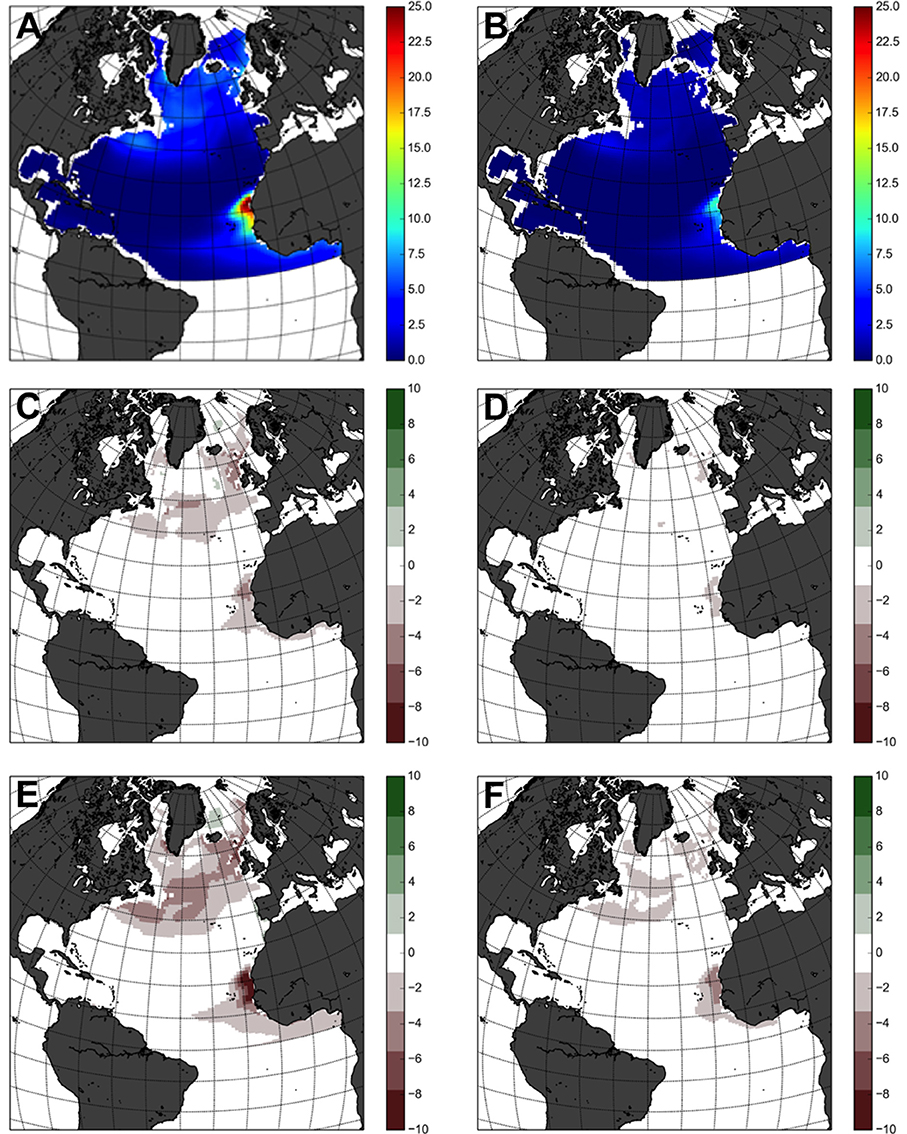
Figure 2. Maps of Carbon fluxes in the North Atlantic based on MEDUSA (1° × 1°). Present fluxes (1990–2009, in g.m−2.year−1) (A) 500 m (B) 1000 m. Changes in 2090–2099 according to the RCP 2.6 emission scenario, referenced to the present (C) 500 m (D) 1000 m. Changes in 2090–2099 according to the RCP 8.5 emission scenario, referenced to the present (E) 500 m (F) 1000 m. Positive values indicate C uptake to the ocean.
To estimate the economic value of the reduction in carbon sequestration we first computed baseline (1990–2009) carbon export fluxes in the North Atlantic at 500 m and 1000 m deep (Table 1). These range between 0.08 Pg C.year−1 for MEDUSA and 0.56 Pg C.year−1 for ERSEM at 500 m deep, and between 0.02 Pg C.year−1 for MEDUSA and 0.45 Pg C. year1 for ERSEM at 1000 m deep (Table 1). MEDUSA export fluxes drop ~70% between both depths while ERSEM fluxes drop by only 20% (see below). We compared the cumulative (2010–2099) difference in carbon export between model outputs and a projection of the fluxes for the baseline period until the end of the century. The cumulative reduction in carbon export by the biological pump in the North Atlantic between 2010 and 2100 ranges, at 500 m, between 1.5 PgC (MEDUSA RCP2.6) and 12.7 PgC (ERSEM RCP8.5). At 1000 m these reductions range between 0.6 Pg C (MEDUSA RCP2.6) to 10.8 Pg C (ERSEM RCP8.5, Table 1). These losses, while significantly different in absolute values, correspond approximately to a 25% reduction in carbon exports over the rest of the twenty-first century compared to present fluxes projected over the same period.
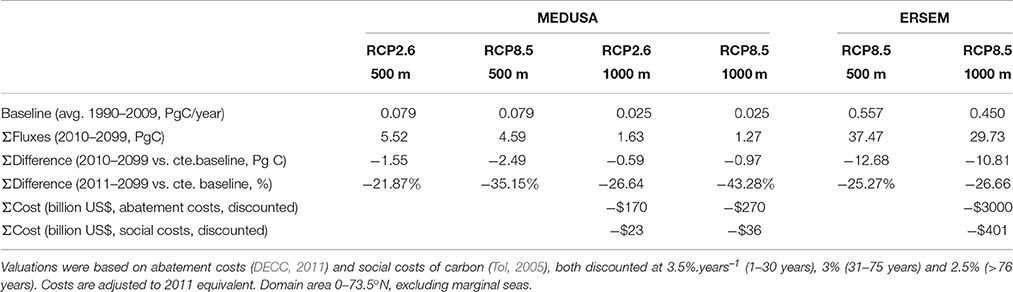
Table 1. Economic value of the reduction in the ecosystem service of carbon sequestration over the period 2011–2099, compared to the baseline fluxes in the period 1990–2009, according to RCP emission scenarios and biogeochemical model.
Monetary valuation of changes in carbon fluxes enables a transparent debate to compare costs and benefits to other ecosystem services, and discuss various intervention or management options. Carbon uptake and sequestration are important components in the regulation of climate, and these benefits can be valued using carbon price as a proxy.
We translated the differences in export fluxes at 1000 m in our scenarios into economic values, using the social (adaptation) and abatement (mitigation) costs approaches. Organic carbon sinking below this level is unlikely to return to the surface and enter in contact with the atmosphere within the span of our model projection. Discounted social costs, for the period 2010–2099 amount to US$23-US$401 billion, the highest figure corresponding to emission scenario RCP8.5 under ERSEM. Discounted abatement costs, also discounted, bring figures between US$170 billion (MEDUSA RCP2.6) and US$3000 billion (ERSEM RCP8.5). This indicates the higher costs of mitigation (abatement) compared to the adaptation costs of resultant negative impacts on society and the economy (social costs) (Table 1).
In this paper we estimated the potential reduction in the biological carbon pump in the North Atlantic over the twenty-first century, using two well-recognized biochemical modeling frameworks for two extreme-end greenhouse gas emission scenarios, and calculate the impacts of these reductions on the BCP carbon sequestration service of the North Atlantic Ocean, using two alternative cost models. This ecological-economic assessment highlights the significant contribution of marine biota to blue carbon sequestration, computes the cost of the loss of ecosystem service provision, and provides a level of uncertainty estimation.
Our results show expected declines in the North Atlantic carbon budgets over the twenty-first century of ~25% from baseline figures projected over the same period. This is qualitatively consistent with other model projections which expect that climate change will reduce oceanic carbon uptake in most oceanic regions, with the North Atlantic predicted to show a larger reduction in ocean carbon sink in response to climate change compared to other ocean basins (Roy et al., 2011).
We have focused exclusively on costing the loss of the ecosystem service provided by the North Atlantic Ocean in terms of storing carbon over the full twenty-first century; carbon that cannot return to the atmosphere for >100 years (e.g., within the span of our model projection) or when it reaches depths >1000 m. This is a critical assumption that would not be valid in all regions (e.g., Robinson et al., 2014), but has been used before to describe carbon sequestration thresholds in the North Atlantic (Lampitt et al., 2008). Guidi et al. (2015) suggest a density surface of 1027.6 (or a more stringent 2000 m) as a more accurate estimate of the carbon sequestration depth. With the exception of the northern margin of our domain, this surface is deeper than seasonal mixing and shallower than our selected horizon. Thus, we would argue that our choice of sequestration depth generally represents a conservative horizon. Furthermore, although it is possible that ocean ventilation may actually decline into the future, the fraction of the carbon flux to >1000 m that may return to surface waters can be assumed to be similar between the present-day and the future. Changes in the long-term storage pool of carbon will be reflected in the air-sea exchange budget (Kwon et al., 2009).
Our results indicate significant differences in the projection trends between MEDUSA and ERSEM runs, particularly toward the end of the century (Figure 1). Our models estimate a baseline export at 1000 m of 0.450 PgC.year−1 in ERSEM and 0.025 PgC.year−1 in MEDUSA (Table 1), thus significantly different. Model discrepancies could be caused by the different mineralization rate in the mesopelagic domain or by different surface production and export. Significant model differences in fluxes between 500 m and 1000 m were also observed. For example, MEDUSA export fluxes drop ~70% between both depths while ERSEM fluxes drop by only 20%. Sedimentation rates based on North Pacific traps suggest a drop of ~45%, thus intermediate to both models (Martin et al., 1987).
To better understand the origin of the discrepancies we first compared the modeled fluxes at 1000 m depth for the period 2000–2009 and compared them with observations from sediment traps from Honjo et al. (2008), using only measurements that covered a full seasonal cycle, and projected to the 1000 m depth assuming a Martin curve with an exponent of −0.86 (Martin et al., 1987, Figure 3). Averaging Honjo data for the North Atlantic model domain, suggest an export flux of 0.12 Gt C.year−1, closer to the averaged MEDUSA export values for the whole North Atlantic domain, but still five times larger than the model run. Calculations also based on observational data from Guidi et al. (2015) give an export estimate at 1000 m depth of 0.33 Pg C.year−1 globally, without taking into account lateral fluxes from land sources (Regnier et al., 2013). Downscaling this estimate to the surface area of the North Atlantic, assuming constant average between ocean basins, would result in North Atlantic fluxes of 0.046 Pg C.year−1, twice the MEDUSA estimates but 10 times smaller than ERSEM.
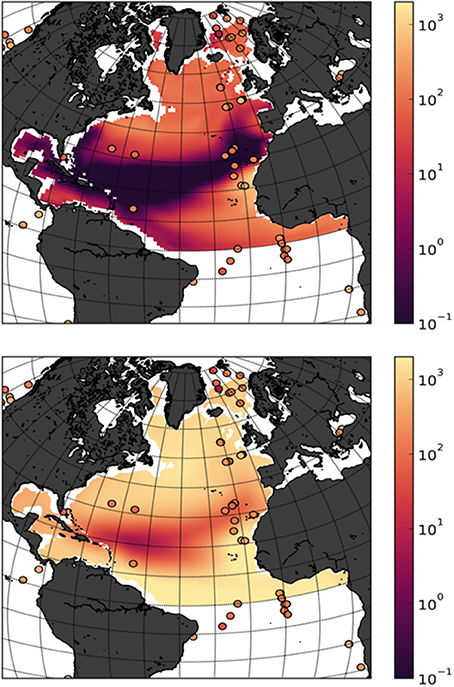
Figure 3. Downward flux estimates of particulate organic carbon (log10 mmol m−2 year−1) at 1000 m. Mean field from the outputs of MEDUSA (top panel) and ERSEM (bottom panel) for the period (2000–2009). Overlaying circles are observations from sediment data from Honjo et al. (2008). Only points where measurements covered a full seasonal cycle were considered. Values were projected to the 1000 m depth assuming a Martin curve with an exponent of −0.86 (Martin et al., 1987).
At this point it must be noted that MEDUSA and ERSEM parameterize the exponential decay of sinking organic carbon and the vertical patterns of remineralization differently. In MEDUSA the remineralization is represented by a parametric description of slow and fast sinking detritus, while in ERSEM it is directly mediated by the scavenging of zooplankton and uptake and release of the functional type for bacteria. Yool et al. (2013a) report that the parametric description in MEDUSA shows an average b of 1.342 in the 1990 s, and 1.524 in the 2090 s, indicating that the sinking flux of organic material is attenuated more rapidly with depth than observed profiles would suggest, an effect that is even increased when progressing into the future. The ERSEM simulation on the contrary show significantly lower values of only 0.3 on average, with simulations showing an increase in relative attenuation in the future (0.33 for the 2090 s). The original assigned value of b is 0.858, although from a relatively limited dataset Buesseler et al. (2007) report values of 1.33 and 0.51 for North Pacific sites, while global synthesis reports a broad geographical range from 0.24 to 1.18 (Henson et al., 2012). This shows that ERSEM and MEDUSA are essentially at both extremes of the plausible remineralization range. Differences between both models therefore can most likely be attributed to the increased metabolic activity of the remineralizing bacteria, which are explicitly included in the ERSEM case, while for MEDUSA the temperature dependence is directly imposed on the remineralization rate.
We also explored whether differences in NPP and export from the photic zone may be responsible for the model differences. Figure 4 shows the estimated C fluxes at 100 m from both models, compared to estimates using the algorithm of Henson et al. (2011), which uses satellite-based Primary Production, the Vertical Generalized Production Model (VGPM, Behrenfeld and Falkowski, 1997), and Advanced Very High Resolution Radiometer (AVHRR) Sea Surface Temperature, for the period 2000–2009. The integrated fluxes at 100 m were 0.46 Pg/y for the satellite-based estimate, compared to 0.87 Pg/y for ERSEM and 0.65 Pg/y for MEDUSA. Thus, while model outputs appear to be higher than the satellite-based Henson dataset, both ERSEM and MEDUSA provide reasonably comparable results.
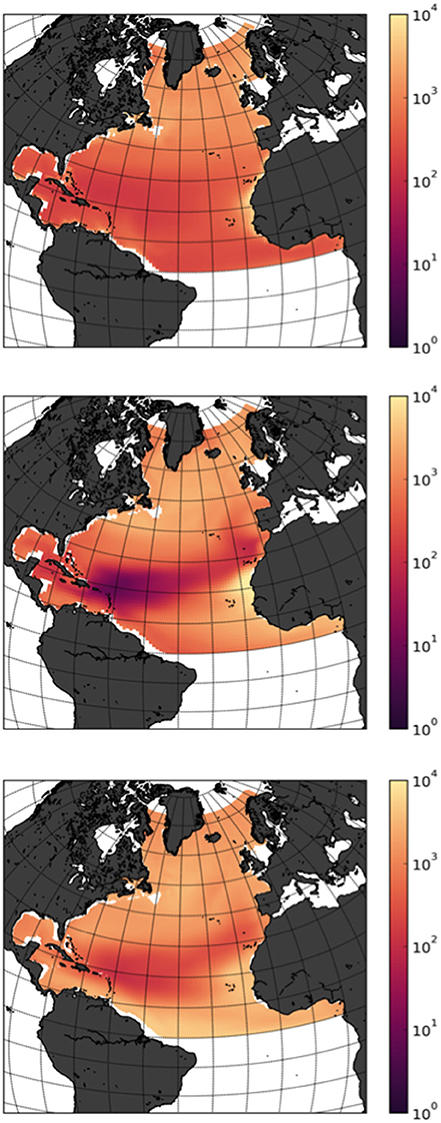
Figure 4. Downward flux estimates of particulate organic carbon (log10 mmol m−2 year−1) at 100 m. Estimated fluxes using the Henson et al. (2011) algorithm, which uses Primary Production derived from satellite data and the Vertical Generalized Production Model (VGPM, Behrenfeld and Falkowski, 1997), and Advanced Very High Resolution Radiometer (AVHRR) Sea Surface Temperature (top panel), and outputs from the MEDUSA (middle panel) and ERSEM (bottom panel) models for the period (2000–2009).
It is therefore evident that model differences emerge from a fundamentally different description of the remineralization process used in the two models. MEDUSA employs an implicit remineralization form, whereby POM is directly remineralized to bioavailable nutrients according to a prescribed rate (Yool et al., 2011). In contrast ERSEM uses a fully explicit microbial model whereby bacteria are explicitly modeled and remineralize POM and DOM (Butenschön et al., 2016). It has been shown that phytoplankton community and food-web type play a central role in determining the carbon fluxes in the marine ecosystem and how carbon export is attenuated by the system (Legendre and Rassoulzadegan, 1995; Daniels et al., 2006). Consequently, substantial difference in the variability of the export is to be expected when the modeled food webs differ significantly. In MEDUSA the correlation between NPP and export is rather straightforward and almost linear, driven by particle dynamics. This is not the case for ERSEM, where export production can reach much higher values for relatively low NPP (see Supplementary Material). The link between NPP and export flux at depth has been debated since both fluxes have been measured or estimated at sea (Antia et al., 2001; Henson et al., 2012).
The large difference in sequestration fluxes must have some impact on ocean tracer fields. However, we have not explored how the models differ in the time evolution of key fields like DIC, DIN, and oxygen export, because given the limited observations available it would be impossible to deduce which export field is more credible. Nevertheless, this is worth exploring in future model comparisons.
Finally, it must be acknowledged that the extent to which the export of organic material from the surface ocean results in oceanic carbon storage is not simply controlled by the magnitude of the export flux or the depth of remineralization, but on the extent to which nutrients in the deep ocean are associated with carbon, and the extent to which this carbon is able to escape from the deep ocean (Gnanadesikan and Marinov, 2008). Further work is clearly needed before more conclusive evidence can obtained.
Finally, the percentage reductions in carbon fluxes at depth over the twenty-first century by ERSEM and MEDUSA (21–43%, Table 1) are larger but consistent with Jones et al. (2014), who estimated that the flux of particulate carbon to the seafloor would reduce by 15% in the North Atlantic by the end of the twenty-first century.
We computed economic valuations using the social (adaptation) and abatement (mitigation) costs approaches, resulting in social (adaptation) costs for the period 2010–2099 of US$23-US$401 billion, and abatement (mitigation) costs, also discounted, bring figures between US$170- US$3000 billion. For comparison, the CO2 emissions resulting from conversion and degradation of vegetated coastal ecosystems equates to $US 6–42 billion annually (US$534-US$3738 billion over the same time period as in our simulation, Pendleton et al., 2012). The value of carbon sequestration over the Mediterranean basin attributable to the biological pump has been recently estimated to be €281 million per year, equivalent to US$29 billion over the same period as this study (Melaku Canu et al., 2014). There are no other comparable values for offshore habitats as the majority of current blue carbon research is focused on coastal environments (Luisetti et al., 2013; Thomas, 2014; Schaafsma and Turner, 2015).
The role of the ocean biota in carbon storage is one of the services that is more difficult to assess in monetary terms, both in terms of incomplete understanding and because it is subject to significant uncertainties not only in the underpinning ecosystem modeling but also in carbon price estimates and discounting rates (HM Treasury, 2011; van den Bergh and Botzen, 2015). Furthermore, the concept of “permanent carbon removal” can be questioned at scales larger than the model runs. Notwithstanding these uncertainties, our results suggest that the ecosystem service loss provided by the North Atlantic biological pump can reach hundreds of billion US$ in mitigation costs, and thousands of billion US$ in adaptation costs, over the twenty-first century, depending on the economic valuation methodology used.
Interest in ecosystem service quantification has led to numerous ecological assessments of ecosystem services, mostly to assist in marine spatial planning (e.g., Börger et al., 2014), or to contribute to national assessments in support of Biodiversity policies. Quantifying ecosystem services is useful for demonstrating the links between humans and the environment, allowing services to be recognized, as well as impacts and trade-offs between management options. In the marine environment, the majority of valuation studies have been applied to coastal and near-shore ecosystems (e.g., Hynes et al., 2013; Beaumont et al., 2014). Here we highlight the potential economic, environmental, and social cost of damaging the ecosystem services provided by the open ocean through anthropogenic climate change, as well as some of the uncertainty associated with projecting the value of ecosystem services into the future.
MBa designed the study, MBu, AY, AM, and JA run, analyzed and/or interpreted the biochemistry models, NB led, and JF and MBa implemented and analyzed the carbon valuation algorithms. All authors contributed to the text.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
This is a contribution to the European Commission 7th Framework programme Integrated Project EURO-BASIN, European Basin-Scale Analysis, Synthesis and INtegration (FP7/2007–2013, ENV.2010.2.2.1–1; grant agreement number 264933, www.euro-basin.eu), and the UK Earth System Modelling Project (UKESM, grant number NE/N018036/1). We would like to that Dr. Stephanie Henson (NOC) for providing estimates of C fluxes at 100 m deep from Henson et al. (2011), using satellite-based Primary Production (Figure 4).
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmars.2016.00290/full#supplementary-material
Antia, A. N., Koeve, W., Fischer, G., Blanz, T., Schulz-Bull, D., Scholten, J., et al. (2001). Basin-wide particulate carbon flux in the Atlantic Ocean: Regional export patterns and potential for atmospheric CO2 sequestration. Global Biogeochem. Cycles 15, 845–862. doi: 10.1029/2000GB001376
Armstrong, R. A., Lee, C., Hedges, J. I., Honjo, S., and Wakeham, S. G. (2002). A new, mechanistic model for organic carbon fluxes in the ocean: based on the quantitative association of POC with ballast minerals. Deep Sea Res. 49, 219–236. doi: 10.1016/S0967-0645(01)00101-1
Beaumont, N. J., Jones, L., Garbutt, A., Hansom, J. D., and Toberman, M. (2014). The value of carbon sequestration and storage in coastal habitats. Est. Coast. Shelf. Sci. 137, 32–40. doi: 10.1016/j.ecss.2013.11.022
Behrenfeld, M. J., and Falkowski, P. G. (1997). Photosynthetic rates derived from satellite-based chlorophyll concentration. Limnol. Oceanogr. 42, 1–20.
Börger, T., Beaumont, N. J., Pendleton, L., Boyle, K. J., Cooper, P., Fletcher, S., et al. (2014). Incorporating ecosystem services in marine planning: the role of valuation. Mar. Policy 46, 161–170. doi: 10.1016/j.marpol.2014.01.019
Bopp, L., Monfray, P., Aumont, O., Dufresne, J.-L., Le Treut, H., Madec, G., et al. (2001). Potential impact of climate change on marine export production. Global Biogeochem. Cycles 15, 81–100. doi: 10.1029/1999GB001256
Broecker, W. S. (1982). Glacial to interglacial changes in ocean chemistry. Prog. Oceanogr. 11, 151–197. doi: 10.1016/0079-6611(82)90007-6
Buesseler, K. O., Lamborg, C. H., Boyd, P. W., Lam, P. J., Trull, T. W., Bidigare, R. R., et al. (2007). Revisiting carbon flux through the ocean's twilight zone. Science 316, 567–570. doi: 10.1126/science.1137959
Butenschön, M., Clark, J., Aldridge, J. N., Allen, J. I., Artioli, Y., Blackford, J., et al. (2016). ERSEM 15.06: a generic model for marine biogeochemistry and the ecosystem dynamics of the lower trophic levels. Geosci. Model Dev. Discuss. 9, 1293–1339. doi: 10.5194/gmd-9-1293-2016
Daniels, R. M., Richardson, T. L., and Ducklow, H. W. (2006). Food web structure and biogeochemical processes during oceanic phytoplankton blooms: an inverse model analysis. Deep Sea Res. Part II 53, 532–554. doi: 10.1016/j.dsr2.2006.01.016
DECC (2011). A Brief Guide to the Carbon Valuation Methodology for UK Policy Appraisal. London: Department of Energy and Climate Change. URN 11D/877.
Defra (2007). The Social Cost of Carbon and the Shadow Price of Carbon: What They Are, and How to Use Them in Economic Appraisal in the UK. London: Defra Economics group.
Garcia, H. E., Locarnini, R. A., Boyer, T. P., Antonov, J. I., Zweng, M. M., Baranova, O. K., et al. (2010). “Volume 4: Nutrients (phosphate, nitrate, silicate),” in World Ocean Atlas 2009, ed S. Levitus (Washington, DC: NOAA Atlas NESDIS 71, US Government Printing Office), 398.
Gnanadesikan, A., and Marinov, I. (2008). Export is not enough: nutrient cycling and carbon sequestration. Mar. Ecol. Prog. Ser. 364, 289–294. doi: 10.3354/meps07550
Gruber, N., Keeling, C. D., and Bates, N. R. (2002). Interannual variability in the North Atlantic Ocean carbon sink. Science 298, 2374–2378. doi: 10.1126/science.1077077
Guidi, L., Legendre, L., Reygondeau, G., Uitz, J., Stemmann, L., and Henson, S. A. (2015). A new look at ocean carbon remineralization for estimating deepwater sequestration. Global Biogeochem. Cycles 29, 1044–1059. doi: 10.1002/2014GB005063
Hartmann, D. L., Klein Tank, A. M. G., Rusticucci, M., Alexander, L. V., Brönnimann, S., Charabi, Y., et al. (2013). “Observations: Atmosphere and Surface,” in Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change, eds T. F. Stocker, D. Qin, G.-K. Plattner, M. Tignor, S. K. Allen, J. Boschung, A. Nauels, Y. Xia, V. Bex, and P. M. Midgley (Cambridge; New York, NY: Cambridge University Press), 159–252.
Heinze, C. S., Meyer, N., Goris, L., Anderson, R., Steinfeldt, N., Bakker, D. E. C., et al. (2015). The ocean carbon sink – impacts, vulnerabilities and challenges. Earth Syst. Dyn. 6, 327–358. doi: 10.5194/esd-6-327-2015
Henson, S. A., Sanders, R., and Madsen, E. (2012). Global patterns in efficiency of particulate carbon export and transfer to the deep ocean. Glob. Biogeochem. Cycles 26, GB1018. doi: 10.1029/2011GB004099
Henson, S. A., Sanders, R., Madsen, E., Morris, P. J., Le Moigne, F., and Quartly, G. D. (2011). A reduced estimate of the strength of the ocean's biological carbon pump. Geophys. Res. Lett. 38:L04606. doi: 10.1029/2011GL046735
HM Treasury (2011). The Green Book. Appraisal and Evaluation in Central Government. London: UK Government.
Honjo, S., Manganini, S. J., Krishfield, R. A., and Francois, R. (2008). Particulate organic carbon fluxes to the ocean interior and factors controlling the biological pump: a synthesis of global sediment trap programs since 1983. Prog. Oceanogr. 76, 217–285. doi: 10.1016/j.pocean.2007.11.003
Hynes, S., Tinch, D., and Hanley, N. (2013). Valuing improvements to coastal waters using choice experiments: an application to revisions of the EU bathing waters directive. Mar. Policy 40, 137–144. doi: 10.1016/j.marpol.2012.12.035
Intergovernmental Panel on Climate Change (IPCC), Working Group I (WGI). (2007). “Observations: Ocean climate change and sea level”, in Climate Change 2007: The Physical Science Basis, eds S. Solomon, D. Qin, M. Manning, Z. Chen, M. Marquis, K. B. Averyt, M. Tignor and H. L. Miller (Cambridge: Cambridge University Press), 385–432.
Jones, D. O. B., Yool, A., Wei, C.-L., Henson, S. A., Ruhl, H. A., Watson, R. A., et al. (2014). Global reductions in seafloor biomass in response to climate change. Global Change Biol. 20, 1861–1872. doi: 10.1111/gcb.12480
Key, R. M., Kozyr, A., Sabine, C. L., Lee, K., Wanninkhof, R., Bullister, J. L., et al. (2014). A global ocean carbon climatology: results from Global Data Analysis Project (GLODAP). Global Biogeochem. Cycles 18, GB4031. doi: 10.1029/2004GB002247
Khatiwala, S., Tanhua, T., Mikaloff Fletcher, S., Gerber, M., Doney, S. C., Graven, H. D., et al. (2013). Global ocean storage of anthropogenic carbon. Biogeosciences 10, 2169–2191. doi: 10.5194/bg-10-2169-2013
Kwon, E. Y., Primeau, F., and Sarmiento, J. L. (2009). The impact of remineralization depth on the air-sea carbon balance. Nat. Geosci. 2, 630–635. doi: 10.1038/ngeo612
Lampitt, R. H., Achterberg, E. P., Anderson, T. R., Hughes, J. A., Iglesias-Rodriguez, M. D., Kelly-Gerreyn, B. A., et al. (2008). Ocean fertilization: a potential means of geoengineering? Philos. Trans. R. Soc. A 366, 3919–3945. doi: 10.1098/rsta.2008.0139
Legendre, L., and Rassoulzadegan, F. (1995). Plankton and nutrient dynamics in marine waters. Ophelia 41, 153–172. doi: 10.1080/00785236.1995.10422042
Luisetti, T., Jackson, E. L., and Turner, R. K. (2013). Valuing the European 'coastal blue carbon' storage benefit. Mar. Poll. Bull. 71, 101–106. doi: 10.1016/j.marpolbul.2013.03.029
Lutz, S. J., and Martin, A. H. (2014). Fish Carbon: Exploring Marine Vertebrate Carbon Services. Arendal: GRID-Arendal.
Martin, J. H., Knauer, G. A., Karl, D. M., and Broenkow, W. W. (1987). VERTEX: carbon cycling in the northeast Pacific. Deep Sea Res. Part A. 34, 267–285.
Melaku Canu, D., Ghermandi, A., Nunes, P. A. L. D., Lazzari, P., Cossarini, G., and Solidoro, C. (2014). Estimating the value of carbon sequestration ecosystem services in the Mediterranean Sea: an ecological economics approach. Global Environ. Change 32, 87–95. doi: 10.1016/j.gloenvcha.2015.02.008
Pendleton, L., Donato, D. C., Murray, B. C., Crooks, S., Jenkins, W. A., Sifleet, S., et al. (2012). Estimating global “Blue carbon” emissions from conversion and degradation of vegetated coastal ecosystems. PLoS ONE 7:e43542. doi: 10.1371/journal.pone.0043542
Regnier, P., Friedlingstein, P., Ciais, P., Mackenzie, F. T., Gruber, N., Janssens, I. A., et al. (2013). Anthropogenic perturbation of the carbon fluxes from land to ocean. Nat. Geosci. 6, 597–607. doi: 10.1038/ngeo1830
Ridgwell, A., Zondervan, I., Hargreaves, J. C., Bijma, J., and Lenton, T. M. (2007). Assessing the potential long-term increase of oceanic fossil fuel CO2 uptake due to CO2-calcification feedback. Biogeosciences 4, 481–492. doi: 10.5194/bg-4-481-2007
Robinson, J., Popova, E. E., Yool, A., Srokosz, M., Lampitt, R. S., and Blundell, J. R. (2014). How deep is deep enough? Ocean iron fertilization and carbon sequestration in the Southern Ocean. Geophys. Res. Lett. 41, 2489–2495. doi: 10.1002/2013GL058799
Rogelj, J., Meinshausen, M., and Knutti, R. (2012). Global warming under old and new scenarios using IPCC climate sensitivity range estimates. Nat. Clim. Change 2, 248–253. doi: 10.1038/nclimate1385
Rogers, A. D., Sumaila, U. R., Hussain, S. S., and Baulcomb, C. (2014). The High Seas and Us: Understanding the Value of High-Seas Ecosystems. Oxford: Global Ocean Commission.
Roy, T., Bopp, L., Gehlen, M., Schneider, B., Cadule, P., Frölicher, T. L., et al. (2011). Regional impacts of climate change and atmospheric CO2 on future ocean carbon uptake: a multi-model linear feedback analysis. J. Clim. 24, 2300–2318. doi: 10.1175/2010JCLI3787.1
Sanders, R., Henson, S. A., Koski, M., De La Rocha, C. L., Painter, S. C., Poulton, A. J., et al. (2014). The biological carbon pump in the North Atlantic. Prog. Oceanogr. 129, 200–218. doi: 10.1016/j.pocean.2014.05.005
Sarmiento, J. L., Hughes, T. M. C., Stouffer, R. J., and Manabe, S. (1998). Simulated response of the ocean carbon cycle to anthropogenic warming. Nature 393, 245–249. doi: 10.1038/30455
Schaafsma, M., and Turner, R. K. (2015). “Valuation of coastal and marine ecosystem services: a literature review,” in Coastal Zones Ecosystem Services: From Science to Values to Decision Making, eds R. K. Turner and M. Schaafsma (Springer), 103–126.
Stern, N. (2006). The stern review on the economic effects of climate change. Popul. Dev. Rev. 32, 793–798. doi: 10.1111/j.1728-4457.2006.00153.x
Thomas, S. (2014). Blue Carbon: Knowledge Gaps, critical issues and novel approaches. Ecol. Econ. 107, 22–38. doi: 10.1016/j.ecolecon.2014.07.028
Tol, R. S. J. (2005). The marginal damage costs of carbon dioxide emissions: an assessment of the uncertainties. Energy Policy 33, 2064–2074. doi: 10.1016/j.enpol.2004.04.002
van den Bergh, J. C. J. M., and Botzen, W. J. W. (2015). Monetary valuation of the social cost of CO2 emissions: a critical survey. Ecol. Econ. 114, 33–46. doi: 10.1016/j.ecolecon.2015.03.015
Volk, T., and Hoffert, M. I. (1985). “Ocean carbon pumps: analysis of relative strengths and efficiencies in ocean-driven atmospheric CO2 changes,” in The Carbon Cycle and Atmospheric CO2: Natural Variations Archean to Present, eds E. T. Sundquist and W. S. Broecker (Washington, DC: American Geophysical Union), 99–110.
Yool, A., Popova, E. E., and Anderson, T. R. (2011). MEDUSA: a new intermediate complexity plankton ecosystem model for the global domain. Geosci. Model Dev. 3, 381–341. doi: 10.5194/gmd-4-381-2011
Yool, A., Popova, E. E., and Anderson, T. R. (2013b). MEDUSA-2.0: an intermediate complexity biogeochemical model of the marine carbon cycle for climate change and ocean acidification studies. Geosci. Model Dev. 6, 1767–1811. doi: 10.5194/gmd-6-1767-2013
Keywords: climate change, ocean biochemistry, North Atlantic, carbon sequestration, economic valuation
Citation: Barange M, Butenschön M, Yool A, Beaumont N, Fernandes JA, Martin AP and Allen JI (2017) The Cost of Reducing the North Atlantic Ocean Biological Carbon Pump. Front. Mar. Sci. 3:290. doi: 10.3389/fmars.2016.00290
Received: 03 November 2015; Accepted: 23 December 2016;
Published: 10 January 2017.
Edited by:
Robinson W. (Wally) Fulweiler, Boston University, USAReviewed by:
Donata Melaku Canu, OGS Istituto Nazionale di Oceanografia e di Geofisica Sperimentale, ItalyCopyright © 2017 Barange, Butenschön, Yool, Beaumont, Fernandes, Martin and Allen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Manuel Barange, bS5iYXJhbmdlQHBtbC5hYy51aw==
†Present Address: Manuel Barange, Food and Agriculture Organization of the United Nations, Rome, Italy
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.