 Zdravko Petanjek
Zdravko Petanjek Ivan Banovac
Ivan Banovac Dora Sedmak
Dora Sedmak Matija Vid Prkačin
Matija Vid Prkačin Ana Hladnik
Ana Hladnik- 1Department of Anatomy and Clinical Anatomy, School of Medicine, University of Zagreb, Zagreb, Croatia
- 2Department of Neuroscience, Croatian Institute for Brain Research, School of Medicine, University of Zagreb, Zagreb, Croatia
- 3Center of Excellence for Basic, Clinical and Translational Neuroscience, School of Medicine, University of Zagreb, Zagreb, Croatia
By studying human cortical cytoarchitecture, von Economo noticed large spindle-shaped-neurons within layer Vb in the anterior-cingulate and fronto-insular cortex. Those neurons had such extremely elongated stick-like or corkscrew-like soma shape that appeared to him as a pathological alteration. Eventually, he realized that this was a specialized-type of neuron which he described as distinct from the main cortical cell populations, including the commonly found spindle cells. Data from recent studies suggest that specialized-stick-corkscrew-neurons may have first developed in the fronto-insular cortex before the division of hominids and Old World monkeys, and that they have become abundant in the anterior-cingulate cortex only in the hominid line. Golgi analysis found that they have distinctive somato-dendritic morphology with a characteristic very distal position of their axon origin. Many additional studies claimed to find cells similar to the specialized cells described by von Economo in other non-primate species, even in functionally unrelated cortical regions and layers. However, these studies did not provide sufficient evidence that the cells they described are indeed distinct from common spindle-shaped-neurons, and that they truly correspond to the specialized-stick-corkscrew-cells described by von Economo. We believe that present evidence primarily supports the presence of specialized-stick-corkscrew-neurons in hominids, with a seeming increase in their number in humans compared to other primates. The functional significance of such neuronal specialization within specific areas of the human cerebral cortex remains to be elucidated.
1 Introduction
By studying the cytoarchitecture of the human cerebral cortex, von Economo (1918) noticed and described large elongated neurons in the anterior cingulate cortex (ACC) and in the fronto-insular cortex (FI). The systematical analysis by von Economo and Koskinas (von Economo and Koskinas, 1925) of all cortical regions using Nissl staining, which visualizes the cell body and most proximal parts of the dendrites, revealed that these cells have an extremely elongated corkscrew-like or stick-like soma and are located within layer Vb of the ACC and FI grouped into cell clusters (usually 3–5 neurons). Since von Economo first found these cells in patients with encephalitis lethargica and thought of them as a pathological alteration due to their peculiar morphology in 1918, it wasn’t until later that they were recognized as a unique neuron subtype found only in distinct cortical areas and referred to as “corkscrew cells” (von Economo, 1926; Seeley et al., 2012).
After these initial descriptions, specialized stick-corkscrew neurons came back into in the focus of research after it was shown that patients with Alzheimer’s disease might have a decreased number of this neurons in ACC (Nimchinsky et al., 1995), even though this finding was not confirmed in later studies (Seeley et al., 2006; Kim et al., 2012; Gami-Patel et al., 2019; Gami-Patel et al., 2022; Singleton et al., 2022). A following comparative study found such specialized cells in the ACC of hominids (Nimchinsky et al., 1999). Analysis of Nissl staining of the ACC indicated that the specialized stick-corkscrew neurons were found primarily in hominids but not in other primates (Nimchinsky et al., 1999). Furthermore, these specialized cells seemed to be especially abundant in bonobos and common chimpanzees, less numerous, but still frequently observed in gorillas and relatively sparse in orangutans (Nimchinsky et al., 1999). Only in bonobos they formed clusters similar to those found in the human ACC, while in other apes they were found as individual cells.
In the study by Nimchinsky et al. (1999) specialized stick-corkscrew neurons were not found in the ACC of any of the other 23 analyzed primate species, including the gibbon, several New World (Ceboidea) and Old World (Cercopithecidae) monkey species and prosimians. These findings were largely supported by a later study by Allman et al. (2010). This suggested that specialized stick-corkscrew neurons appeared in the ACC during hominid (Hominidae, great apes) evolution, and that they may have become more numerous and possibly more complex with an increase in encephalization level (Nimchinsky et al., 1999). Somewhat in contrast to these findings, a more recent study found such specialized cells in the FI of the rhesus and cynomolgus monkeys using both Nissl and Golgi staining (Evrard et al., 2012).
We would like to emphasize that in the manuscript by Nimchinsky et al. (1999), specialized stick-corkscrew neurons were referred to as “spindle cells”, whereas in the manuscript by Evrard et al. (2012) they were referred to as “von Economo neurons”. The term “von Economo neurons” was first used by Watson et al. (2006) describing the Golgi morphology of spindle-shaped cells within the human ACC that they considered to be the specialized cells described by von Economo. Using the terms spindle cells and von Economo neurons interchangeably led to subsequent researchers describing the elongated specialized cells of the ACC and FI as “von Economo neurons”.
After introducing the term “von Economo neurons” (VENs), many studies claimed to have found cells similar to the specialized cells of von Economo in other non-primate species, even in regions outside the FI and ACC, and not only in layer V, but also in layers II and III. Most importantly, von Economo explicitly stated that these special cells with a stick- or corkscrew-shaped cell body were clearly distinguishable from other spindle-shaped cells found throughout the cerebral cortex. Therefore, most of the recent studies (see Banovac et al., 2021) have not dedicated sufficient attention to describe and notice a clear distinction between the specialized von Economo neurons and the spindle cells commonly found in the deep layers of the cerebral cortex. This has caused significant confusion in research on fusiform cells and spindle transformation. Furthermore, in this paper we intend to discuss if the present evidence supports the notion that cells observed in other species, areas and layers, truly correspond to the specialized stick-corkscrew neurons described by von Economo (von Economo, 1918).
2 Early descriptions of specialized stick-corkscrew neurons in the human anterior cingulate and fronto-insular cortex
Von Economo and Koskinas (1925) described the three main groups of cortical cells: pyramidal, granule, and spindle (fusiform) cells. Spindle cells were recognized as a major cell population even before von Economo and Koskinas (see Seeley et al., 2012; Banovac et al., 2021), and many studies even specifically emphasized spindle-shaped cells in the ACC, but none of these studies recognized the cells found in the ACC as a separate neuron class. Thus, until von Economo’s work, the elongated cells of the ACC were not recognized as special cells, as already established giant pyramidal neurons in the primary motor (Betz cells) and the Meynert cells in the visual cortex. To distinguish the specialized cells in the ACC and FI from common fusiform (spindle) cells, von Economo (von Economo and Koskinas, 1925; von Economo, 1926; von Economo, 1927) marked them as “corkscrew cells” and “stick cells”.
The first comprehensive modern cyto- and pygmento-architectonic study that aimed to provide a detailed classification of human cortical neurons was performed by Braak (1980). Braak briefly mentioned extremely elongated spindle-shaped cells of layer V of the ACC and referred to them as corkscrew or stick cells of von Economo and Koskinas (1925). Importantly, Braak gave a detailed description of common bipolar fusiform neurons – a subclass of modified pyramidal neurons. These neurons have vertically oriented dendrites with a spindle-shaped soma and are the dominant neuron population of layer VI within most human cortical regions. Note that common bipolar fusiform neurons of layer VI are characterized by a clear demarcation between the soma and the dominant dendrites, and that the axon mainly arises from the soma (von Economo and Koskinas, 1925; Braak, 1980; Petanjek and Kostović, 1994), which is not the case for the specialized stick-corkscrew neurons of the ACC and FI.
The aforementioned data showed that both classical and more recent anatomical studies recognized that vertically oriented spindle-shaped (fusiform) neurons are a common neuron type found throughout the human cerebral cortex, particularly in layer VI. Such cells were referred to as “spindle cells”. Studies published between von Economo’s and Braak’s work (Sanides, 1962; Stephan, 1964; Stephan, 1975) emphasized that the neurons in ACC and FI have a characteristic large and highly elongated cell body located within layer V. Therefore, using the term “spindle cell” to describe the specialized neurons of the ACC and FI might cause misinterpretation because it is conflating commonly found fusiform cells with specialized cells that have distinct morphology and regional/laminar distribution.
Von Economo and Koskinas in their comprehensive overview (von Economo and Koskinas, 1925; von Economo, 1927; von Economo and Triarhou, 2009) on the morphology of different spindle-shaped cells found in the cerebral cortex, have also introduced the term “spindle transformation”. The term refers to morphological changes of pyramidal cells of layer V, and less frequently pyramidal cells of layer III, to establish a spindle-like form. It is important to note that von Economo did not describe layer III and V elongated cells as spindle cells, but as spindle-transformed pyramidal cells. This is in line with Braak (1980) description of numerous spindle-shaped cell body layer III modified pyramidal neurons of the human cortex.
3 Somato-dendritic features and axon origin marks specialized stick-corkscrew neurons
Before von Economo’s description of specialized stick-corkscrew neurons, using Golgi staining, Cajal (1899); Cajal (1995) described the morphology of such cells present in the FI of a 1-month-old infant (Figure 1A). He noticed their distinctive somato-dendritic morphology. This observation was almost fully neglected, even though Cajal’s figure shows that large spindle-shaped layer V neurons have the body shape identical to that of the specialized stick-corkscrew neurons described by von Economo (Figure 1B). Cajal’s drawing also clearly showed the most distinct feature of the large layer V spindle cells in the FI – the very distant origin site of the axon, which arose from the ending of the basal extension. We have to mention that using the Bielchowsky silver staining method, von Economo concluded that VENs had an axon arising near the middle of the soma, directed laterally and possibly branching in the same cortical layer (von Economo, 1926). This is not in line with data from any other studies, and therefore we found that the process observed by von Economo actually represented a thin side dendrite (for a more detailed elaboration see fig. 8 and corresponding text in Banovac et al., 2021).

Figure 1 Comparison of specialized stick-corkscrew neurons of von Economo (VEN) and common spindle-shaped modified pyramidal neurons (MPN) in the fronto-insular (FI), anterior cingulate cortex (ACC), and dorsolateral prefrontal cortex (PFC) on Golgi and Nissl staining. Axons on Golgi staining are marked by “a”. The magnification for all microphotographs is indicated by the 50 μm scale bar in the lower right corner. (A) Drawing of a VEN in the FI of a 1-month-old human, Golgi staining. Note the distant axon origin and the brush-like terminal branching of the prominent basal dendrite. Image modified from Cajal (1899). (B) Microphotograph showing a cluster of VENs in the FI of an adult human, Nissl staining. Image modified from von Economo and Koskinas (1925). (C) Microphotograph of a VEN in the ACC of an adult human, Golgi staining. Note the distant axon origin and the brush-like terminal branching of the prominent basal dendrite. Image modified from Banovac et al. (2019). (D) Microphotograph showing a cluster of VENs in the ACC of an adult human, Nissl staining. Image modified from von Economo and Koskinas (1925). (E) Microphotograph of a common MPN with a spindle-shaped cell body found throughout the PFC, Golgi staining. Note the axon origin close to the cell body and the lack of the brush-like terminal branching typical for VENs. Image modified from Banovac et al. (2019). (F) Microphotograph showing several common MPNs with a spindle-shaped cell body found throughout the PFC, Nissl staining. Image taken from Banovac et al. (2021), licensed under CC BY 4.0.
After Cajal, the Golgi study by Watson et al. (2006) described spindle-shaped cells in the ACC and FI of a 23-year-old human male, without observing the position of axon origin (nor indicating it on the figures). Similarly, the axon origin was neglected in the Golgi study by Correa-Júnior (Correa-Júnior et al., 2020) where the ACC of 4 adult human specimens was analyzed as well as in the study by Fuentealba-Villarroel et al. (2022) describing spindle-shaped cells in the human precuneus.
In addition, the dendritic morphology of most neurons shown in these papers resembles common spindle-shaped, bipolar fusiform neurons (Braak, 1980; Petanjek and Kostović, 1994) and lacks the distinct dendritic morphology characteristic for the special cells shown by Cajal in layer V of the FI. We intended to clarify this issue by performing a Golgi study (Banovac et al., 2019), where the neuron morphology of modified pyramidal neurons in the ACC of 5 adult human specimens was analyzed. We found that specialized stick-corkscrew neurons have a distinct somato-dendritic morphology (Figures 1C, D) that clearly separates them from other modified pyramidal neurons, including common spindle-shaped, bipolar fusiform neurons (Figures 1E, F). Specialized stick-corkscrew neurons were perpendicularly oriented. Their stick-shaped core part consisting of the cell body that extended in both directions without a sharp decrease in thickness. We defined these extensions as a basal and an apical stem. The length of the core part (soma and extensions) was between 150 and 250 μm. The thickness varied from 10 to 21 μm with numerous horizontally oriented, thin and spiny dendrites. The basal extension commonly ended by branching into several smaller dendrites, branching in a brush-like pattern. The topology of apical extension was similar to those characterizing pyramidal neurons apical dendrite.
Therefore, the unique somato-dendritic morphology of specialized stick-corkscrew neurons of the FI and ACC allows for their reliable identification based on soma shape and dendritic topology. In addition, a very distal axon origin appears to a unique feature of the stick and corkscrews cells of layer Vb of the ACC and FI, that is in human very rarely found in cells of other cortical regions (Braak, 1980). These unique features make it easy to delineate specialized stick-corkscrew neurons from common spindle-shaped bipolar neurons (Figure 2). Since these criteria were not taken into account in the manuscript by Watson (Watson et al., 2006) which introduced the term von Economo neurons (VENs), it is questionable how this term was actually interpreted in later studies and whether authors truly described the same type of special cell that was first described by Cajal and von Economo.
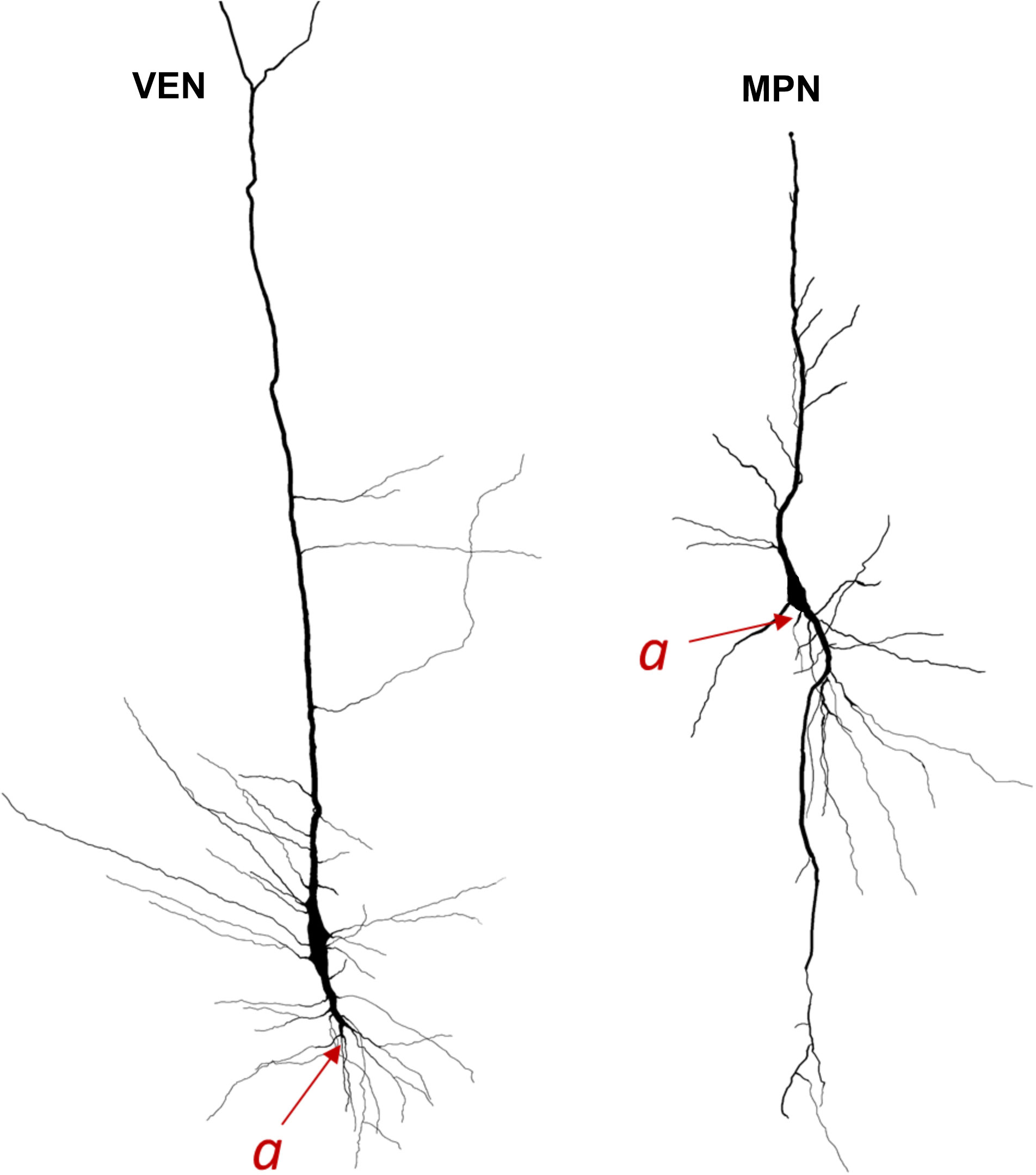
Figure 2 Neurolucida 3D reconstructions of specialized stick-corkscrew neurons of von Economo (VEN) and common spindle-shaped modified pyramidal neurons (MPN) found in the human cerebral cortex. The axon origin is marked by “a” and the axon is traced in red. Note the differences in axon origin between VENs and common MPNs as well as the brush-like terminal branching of the basal dendrite in VENs that is not present in common MPNs. Image modified from Banovac et al. (2021).
Only two Golgi studies intended to identify von Economo neurons were performed in non-human species (Evrard et al., 2012; Butti et al., 2014). In the FI of the adult macaque monkey, Evrard et al. (Evrard et al., 2012) demonstrated the distal axon origin, which was in line with Cajal’s descriptions. However, the authors haven’t noted this morphological feature. The cell body size of the described neurons was smaller and the dendritic morphology seemed less complex when compared to specialized cells of human ACC and FI, but had very similar dendritic topology (Cajal, 1899; von Economo, 1918; von Economo, 1926; Evrard, 2018; Banovac et al., 2019).
4 Discussion
After Nimchinsky’s work, research on spindle-shaped cells in the cerebral cortex greatly expanded (for review see Banovac et al., 2021) with studies in neuropathology or neurodivergent states (e.g. Lin et al., 2019; Nana et al., 2019), comparative and molecular studies (including immunohistochemical characterization and transcriptomics). Many of these studies claimed that von Economo neurons could be found in species other than monkeys, apes and humans, as well as in regions and layers out of FI and ACC layer Vb. The most comprehensive study was done by Raghanti et al. (2015) where Nissl-stained sections were taken from the occipital and frontal pole as well as FI and ACC of the sheep, bowhead whale, cow, pig, deer, rock hyrax, horse, and human. This study found that spindle cells resembling von Economo neurons were present in all analyzed areas and species, not only in layer V, but also in upper cortical layers II and III. Before this study, it was proposed that von Economo neurons might have separately evolved in phylogenetically distant species that have brain size over 300 g, and a sophisticated social behavior (Nimchinsky et al., 1999; Hof and van der Gucht, 2007; Butti et al., 2009; Hakeem et al., 2009; Allman et al., 2011). The study by Raghanti et al. (2015) concluded that von Economo neurons were not restricted to socially complex and highly encephalized species. They concluded that von Economo neurons are present in distantly related species as a result of convergent evolution, possibly representing a typical morphological response to common functional requirements.
However, most of the presented microphotographs and drawings in these manuscripts do not depict the peculiar morphology presented by von Economo in the human brain. Moreover, the described features mainly correspond to common spindle-shaped bipolar neurons. The authors even clearly stated that they defined von Economo neurons based only on the presence of a spindle-shaped soma larger than the surrounding pyramidal cells. In addition, no demonstration of their dendritic morphology and position of axon origin was given. This shows that using a severely watered-down definition of von Economo neurons doesn’t benefit the further discourse on these cells, and particularly not if every large spindle-shaped cell becomes classified in the same category as the specialized stick-corkscrew neurons described by von Economo. This leads to inconsistency in defining these neurons and to contradictory results. For instance, by using anti-NeuN staining the studies, Fajardo et al. (2008) and González-Acosta et al. (2018) claimed to have found von Economo neurons in human Brodmann area 9. Interestingly, in Brodmann area 10, von Economo neurons weren’t identified in the eight subjects analyzed by Fajardo et al. (2008). In contrast, González-Acosta et al. (2018) claimed to have found von Economo neurons in all five analyzed human subjects in Brodmann area 10. Both studies claimed that von Economo neurons appeared to be far less abundant in these regions than in the ACC and FI (Fajardo et al., 2008; González-Acosta et al., 2018). Although, none of these studies provided clear evidence about the abundant presence of cells with similar features as von Economo’s specialized stick-corkscrew neurons, they provide valuable data about the proportion of spindle-shaped neurons as well as “spindle transformation” within various species and layers. A combination of morphological and molecular characteristics (see Dijkstra et al., 2018) might help distinguishing between common spindle-shaped neurons and specialized types of neurons as those described by von Economo.
The functional implications of the “spindle transformation” of principal neurons are still unclear and research on the density and distribution of spindle-shaped neurons in different species, cortical regions and layers is necessary for understanding the importance of this process. There was a recent study (Hodge et al., 2020) performing single nucleus RNA-sequencing of FI layer 5 identified a transcriptomically-defined large spindle cell cluster. By performing patch clamp recordings, it was shown that these neurons have distinctive intrinsic membrane properties relative to neighboring pyramidal neurons. It might be expected, that due to their very special morphology (Banovac et al., 2019), and their unique molecular features (Dijkstra et al., 2018), von Economo neurons might have unique functional properties.
A limiting factor in most studies identifying specialized von Economo neurons is a lack of standardized classification. Hence, some authors even claim that spindle-shaped von Economo-like neurons are present in rat layers III and V, and that these cells express nNOS and NADPH (Pellicer et al., 2022). Based on the morphology and molecular characteristic of these cells, they more likely correspond to the large GABAergic fusiform somatostatin interneurons, which co-express nNOS (Banovac et al., 2022). Such neurons seem to have little in common with von Economo’s specialized cells, which are even larger, have no demarcation between the soma and dendrites and express glutamatergic and projection neuron markers. Therefore, it is clear that there is a need to define objective classification criteria to make the distinction between common spindle bipolar cells and specialized stick-corkscrew neurons easier and more consistent. Thus, finding the right balance between straying too far from the original descriptions and still allowing for appropriate level of variety in non-human species is an imperative for any future studies.
In summary, our point of view is that the present evidence suggests that specialized stick-corkscrew neurons may have evolved before the division of hominids from Old World monkeys and that they appear to be far more abundant in the hominid line. The functional significance of such neuronal specialization and a high number of specialized neurons present only in two areas of the human cerebral cortex remains to be elucidated (Watson and Allman, 2007; Butti et al., 2013; Bruton, 2021).
Data availability statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Author contributions
Conceptualization, study design, and writing the manuscript: ZP. Drafting the manuscript, figure preparation and reference sampling: IB. Drafting the manuscript: DS, AH, MP. All authors contributed to the article and approved the submitted version.
Funding
The research was co-financed by CSF-project “Brain extracellular matrix in development and in perinatal hypoxia” (acronym – BrainECM, IP-2019-04-3182); the Scientific Centre of Excellence for Basic, Clinical and Translational Neuroscience, project “Experimental and clinical research of hypoxic-ischemic damage in perinatal and adult brain”; GA KK01.1.1.01.0007 funded by the European Union through the European Regional Development Fund; Bilateral France-Croatia project COGITO – Program Hubert Curein “Functional and molecular characteristics of the calretinin neuron in primates”.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author ZP declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Allman J. M., Tetreault N. A., Hakeem A. Y., Manaye K. F., Semendeferi K., Erwin J. M., et al. (2010). The von Economo neurons in frontoinsular and anterior cingulate cortex in great apes and humans. Brain Struct. Funct. 214, 495–517. doi: 10.1007/s00429-010-0254-0
Allman J. M., Tetreault N. A., Hakeem A. Y., Park S. (2011). The von Economo neurons in apes and humans. Am. J. Hum. Biol. 23, 5–21. doi: 10.1002/ajhb.21136
Banovac I., Sedmak D., Džaja D., Jalšovec D., Jovanov Milošević N., Rašin M. R., et al. (2019). Somato-dendritic morphology and axon origin site specify von Economo neurons as a subclass of modified pyramidal neurons in the human anterior cingulate cortex. J. Anat. 235, 651–669. doi: 10.1111/joa.13068
Banovac I., Sedmak D., Esclapez M., Petanjek Z. (2022). The distinct characteristics of somatostatin neurons in the human brain. Mol. Neurobiol. 59 (8), 4953–4965. doi: 10.1007/s12035-022-02892-6
Banovac I., Sedmak D., Judaš M., Petanjek Z. (2021). Von economo neurons - primate-specific or commonplace in the mammalian brain? Front. Neural Circuits 15. doi: 10.3389/fncir.2021.714611
Bruton O. J. (2021). Is there a “g-neuron”? Establishing a systematic link between general intelligence (g) and the von Economo neuron. Intelligence 86, 101540. doi: 10.1016/j.intell.2021.101540
Butti C., Ewan Fordyce R., Ann Raghanti M., Gu X., Bonar C. J., Wicinski B. A., et al. (2014). The cerebral cortex of the pygmy hippopotamus, Hexaprotodon liberiensis (Cetartiodactyla, Hippopotamidae): MRI, cytoarchitecture, and neuronal morphology. Anat. Rec. (Hoboken) 297, 670–700. doi: 10.1002/ar.22875
Butti C., Santos M., Uppal N., Hof P. R. (2013). Von Economo neurons: clinical and evolutionary perspectives. Cortex 49, 312–326. doi: 10.1016/j.cortex.2011.10.004
Butti C., Sherwood C. C., Hakeem A. Y., Allman J. M., Hof P. R. (2009). Total number and volume of Von Economo neurons in the cerebral cortex of cetaceans. J. Comp. Neurol. 515, 243–259. doi: 10.1002/cne.22055
Cajal S. R. (1899). Textura del Sistema Nervioso del Hombre y de los Vertebrados, Tomo II (Madrid: Nicolas Moya).
Cajal S. R. (1995). Histology of the Nervous System of Man and Vertebrates (New York, NY: Oxford University Press).
Correa-Júnior N. D., Renner J., Fuentealba-Villarroel F., Hilbig A., Rasia- Filho A. A. (2020). Dendritic and spine heterogeneity of von Economo neurons in the human cingulate cortex. Front. Synaptic Neurosci. 12. doi: 10.3389/fnsyn.2020.00025
Dijkstra A. A., Lin L. C., Nana A. L., Gaus S. E., Seeley W. W. (2018). Von economo neurons and fork cells: A neurochemical signature linked to monoaminergic function. Cereb. Cortex 28, 131–144. doi: 10.1093/cercor/bhw358
Evrard H. C. (2018). Von Economo and fork neurons in the monkey insula, implications for evolution of cognition. Curr. Opin. Behav. Sci. 21, 182–190. doi: 10.1016/j.cobeha.2018.05.006
Evrard H. C., Forro T., Logothetis N. K. (2012). Von Economo neurons in the anterior insula of the macaque monkey. Neuron 74, 482–489. doi: 10.1016/j.neuron.2012.03.003
Fajardo C., Escobar M. I., Buriticá E., Arteaga G., Umbarila J., Casanova M. F., et al. (2008). Von Economo neurons are present in the dorsolateral (dysgranular) prefrontal cortex of humans. Neurosci. Lett. 435, 215–218. doi: 10.1016/j.neulet.2008.02.048
Fuentealba-Villarroel F. J., Renner J., Hilbig A., Bruton O. J., Rasia-Filho A. A. (2022). Dendritic and spine heterogeneity of von economo neurons in the human cingulate cortex. Front. Synaptic Neurosci. 12. doi: 10.3389/fnsyn.2020.00025
Gami-Patel P., Scarioni M., Bouwman F. H., Boon B. D. C., van Swieten J. C., Brain Bank N., et al. (2022). The severity of behavioural symptoms in FTD is linked to the loss of GABRQ-expressing VENs and pyramidal neurons. Neuropathol. Appl. Neurobiol. 48, e12798. doi: 10.1111/nan.12798
Gami-Patel P., van Dijken I., van Swieten J. C., Pijnenburg Y. A. L., Netherlands Brain Bank, Rozemuller A. J. M., et al. (2019). Von Economo neurons are part of a larger neuronal population that are selectively vulnerable in C9orf72 frontotemporal dementia. Neuropathol. Appl. Neurobiol. 45, 671–680. doi: 10.1111/nan.12558
González-Acosta C. A., Escobar M. I., Casanova M. F., Pimienta H. J., Buriticá E. (2018). Von economo neurons in the human medial frontopolar cortex. Front. Neuroanat. 12. doi: 10.3389/fnana.2018.00064
Hakeem A. Y., Sherwood C. C., Bonar C. J., Butti C., Hof P. R., Allman J. M. (2009). Von Economo neurons in the elephant brain. Anat. Rec. (Hoboken) 292, 242–248. doi: 10.1002/ar.20829
Hodge R. D., Miller J. A., Novotny M., Kalmbach B. E., Ting J. T., Bakken T. E., et al. (2020). Transcriptomic evidence that von Economo neurons are regionally specialized extratelencephalic-projecting excitatory neurons. Nat. Commun. 11, 1172. doi: 10.1038/s41467-020-14952-3
Hof P. R., van der Gucht E. (2007). Structure of the cerebral cortex of the humpback whale, Megaptera novaeangliae (Cetacea, Mysticeti, Balaenopteridae). Anat. Rec. (Hoboken) 290, 1–31. doi: 10.1002/ar.20407
Kim E. J., Sidhu M., Gaus S. E., Huang E. J., Hof P. R., Miller B. L., et al. (20122012). Selective frontoinsular von Economo neuron and fork cell loss in early behavioral variant frontotemporal dementia. Cereb. Cortex 22, 251–259. doi: 10.1093/cercor/bhr004
Lin L. C., Nana A. L., Hepker M., Hwang J. L., Gaus S. E., Spina S., et al. (2019). Preferential tau aggregation in von Economo neurons and fork cells in frontotemporal lobar degeneration with specific MAPT variants. Acta Neuropathol. Commun. 7, 159. doi: 10.1186/s40478-019-0809-0
Nana A. L., Sidhu M., Gaus S. E., Hwang J. L., Li L., Park Y., et al. (2019). Neurons selectively targeted in frontotemporal dementia reveal early stage TDP-43 pathobiology. Acta Neuropathol. 137, 27–46. doi: 10.1007/s00401-018-1942-8
Nimchinsky E. A., Gilissen E., Allman J. M., Perl D. P., Erwin J. M., Hof P. R. (1999). A neuronal morphologic type unique to humans and great apes. Proc. Natl. Acad. Sci. U.S.A. 96, 5268–5273. doi: 10.1073/pnas.96.9
Nimchinsky E. A., Vogt B. A., Morrison J. H., Hof P. R. (1995). Spindle neurons of the human anterior cingulate cortex. J. Comp. Neurol. 355, 27–37. doi: 10.1002/cne.903550106
Pellicer F., Singh S., Sanchez-Islas E., Leon-Olea M. (2022). Identification of spindle-shaped von Economo-like neurons in the rodent Rattus norvegicus. Res. Square. doi: 10.21203/rs.3.rs-1623986/v1
Petanjek Z., Kostović I. (1994). Golgi morphology of neurons in the adult human prefrontal cortex: II. modified pyramidal neurons. Period. Biol. 96, 93–94.
Raghanti M. A., Spurlock L. B., Treichler F. R., Weigel S. E., Stimmelmayr R., Butti C., et al. (2015). An analysis of von Economo neurons in the cerebral cortex of cetaceans, artiodactyls, and perissodactyls. Brain Struct. Funct. 220, 2303–2314. doi: 10.1007/s00429-014-0792-y
Sanides F. (1962). Die Architektonik des Menschlichen Stirnhirns. Vol. 98 of Monographien aus dem Gesamtgebiete der Neurologie und Psychiatrie (Berlin: Springer).
Seeley W. W., Carlin D. A., Allman J. M., Macedo M. N., Bush C., Miller B. L., et al. (2006). Early frontotemporal dementia targets neurons unique to apes and humans. Ann. Neurol. 60, 660–667. doi: 10.1002/ana.21055
Seeley W. W., Merkle F. T., Gaus S. E., Craig A. D. B., Allman J. M., Hof P. R., et al. (2012). Distinctive neurons of the anterior cingulate and frontoinsular cortex: a historical perspective. Cereb. Cortex 22, 245–250. doi: 10.1093/cercor/bhr005
Singleton E. H., Pijnenburg Y. A. L., Gami-Patel P., Boon B. D. C., Bouwman F., Papma J. M., et al. (2022). The behavioral variant of Alzheimer's disease does not show a selective loss of Von Economo and phylogenetically related neurons in the anterior cingulate cortex. Alzheimers Res. Ther. 14, 11. doi: 10.1186/s13195-021-00947-9
von Economo C. (1926). Eine neue Art Spezialzellen des Lobus cinguli und Lobus insulae. Z. Ges Neurol. Psychiatr. 100, 706–712. doi: 10.1007/bf02970950
von Economo C. F., Koskinas G. N. (1925). Die Cytoarchitektonik der Hirnrinde des Erwachsenen Menschen. (Wien: J. Springer).
von Economo C., Triarhou L. C. (2009). Cellular Structure of the Human Cerebral Cortex. (Basel: Karger).
Watson K. K., Allman J. M. (2007). Role of Spindle Cells in the Social Cognition of Apes and Humans. (Amsterdam: Elsevier Inc).
Keywords: higher cognitive functions, cortical circuitry, human-specific neurons, modified pyramidal neurons, schizophrenia, autism
Citation: Petanjek Z, Banovac I, Sedmak D, Prkačin MV and Hladnik A (2023) Von Economo neurons as a specialized neuron class of the human cerebral cortex. Front. Mamm. Sci. 2:1242289. doi: 10.3389/fmamm.2023.1242289
Received: 19 June 2023; Accepted: 15 September 2023;
Published: 04 October 2023.
Edited by:
Ruth Benavides-Piccione, Spanish National Research Council (CSIC), SpainReviewed by:
William Seeley, University of California, San Francisco, United StatesCopyright © 2023 Petanjek, Banovac, Sedmak, Prkačin and Hladnik. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Zdravko Petanjek, emRyYXZrby5wZXRhbmpla0BtZWYuaHI=