
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
BRIEF RESEARCH REPORT article
Front. Malar., 14 January 2025
Sec. Antimalarial Drug Resistance
Volume 2 - 2024 | https://doi.org/10.3389/fmala.2024.1516733
 Alan Oyarce1*†
Alan Oyarce1*† Lizbeth Huaccha1†
Lizbeth Huaccha1† Esteban M. Cordero1
Esteban M. Cordero1 Bárbara Parra1
Bárbara Parra1 Jorge Fernández1
Jorge Fernández1 Mario E. Soto-Marchant2
Mario E. Soto-Marchant2 Natalia T. Santis-Alay2
Natalia T. Santis-Alay2 María Isabel Jercic1
María Isabel Jercic1A key challenge in the global effort to combat malaria is the emergence of drug resistance. Malaria-free countries must also address issues such as limited access to antimalarial drugs and treatment failures caused by drug resistance. Chile, as a malaria-free country, faces the risk of malaria being reintroduced due to the presence of the malaria vector in its continental territory. This study aims to analyze the genetic profile associated with antimalarial drug resistance in the pfcytb, pfdhfr, and pvmrp1 genes. A total of ninety blood samples from 55 individuals who had been diagnosed with malaria in Chile between 2019 and 2021 were subjected to mutational analysis. The parasites target genes were amplified by polymerase chain reaction (PCR) out of total DNA extracted from patient blood samples and the amplicons submitted to DNA sequencing. All the genes analyzed had at least one mutation. In the pfdhfr gene, three mutations were observed (S108N/N51I/C59R). In the pfcytb gene, the Y268C mutation, found in post-treatment samples, was associated with treatment failure. In the pvmrp1 gene, five distinct mutations were identified. Of these, the Y1393D (100%) and V1478I (95.2%) were the most common. Our findings indicate that both P. falciparum and P. vivax samples from travelers and migrants in Chile carry mutations in genes linked to malaria resistance. The circulation of parasites with potential drug resistance in non-endemic countries further complicates the challenge of ensuring adequate treatment. It is crucial to continue genetic surveillance and expand the search for new resistance markers for Plasmodium species.
Malaria continues to pose a significant challenge to global public health. It is estimated that there were 249 million malaria cases in 2022, resulting in 600,000 deaths in 85 countries (World Health Organization, 2023a). Increased international migration has led to the spread of cases to malaria-free countries, complicating the situation in areas where Anopheles vectors are still present (Tatem et al., 2017).
Of the five main Plasmodium species affecting humans, Plasmodium falciparum and Plasmodium vivax exhibit the highest rates of infection and morbidity. P. falciparum is the predominant species in sub-Saharan Africa and is responsible for the majority of malaria-related deaths. In contrast, P. vivax is the predominant species in Asia-Pacific and the Americas, and its impact on public health is increasingly recognized (Price et al., 2009; Phyo et al., 2022).
The World Health Organization (WHO) has certified Chile as malaria-free country since 1968, within the category of “Countries where malaria never existed or disappeared without specific measures” (World Health Organization, 2024). However, there are imported cases of malaria - caused by different species of Plasmodium - each year among travelers and migrants from different endemic countries around the world, with P. vivax and P. falciparum as predominant species (Escobar et al., 2020). Timely and appropriate diagnosis and treatment of cases are essential to prevent disease transmission from being reestablished due to the presence of the Anopheles pseudopunctipennis vector in northern Chile (Escobar et al., 2020; Valderrama et al., 2021).
Nowadays, a key challenge in controlling and eliminating malaria is the emergence and spread of drug-resistant strains (World Health Organization, 2021). P. falciparum has been observed to have developed resistance to most of the available antimalarial drugs, including quinoline drugs, antifolates, and even artemisinin derivatives (Balikagala et al., 2021; Ippolito et al., 2021). P. vivax has also shown increased tolerance to drugs, such as chloroquine and primaquine (Vinetz, 2006; Krudsood et al., 2008). Research on resistance in P. vivax has evolved gradually, and molecular markers have mainly been sought in homologous genes previously described in P. falciparum (Buyon et al., 2021). In the meantime, resistance to several antimalarial drugs has led to changes in the WHO’s treatment guidelines and recommendations (World Health Organization, 2000; World Health Organization, 2023b).
Access to antimalarial drugs is limited in Chile and the acquisition is conducted at the central level within the country. Local technical guidelines recommend atovaquone/proguanil (AP) as the first-line treatment for uncomplicated P. falciparum cases and chloroquine/primaquine (CP) is recommended for P. vivax infections to prevent relapses due to dormant stages (hypnozoites) in the liver (MINSAL, 2015). Gaining insight into the epidemiology of malaria in Chile and the patterns of different genetic markers of resistance to antimalarials would enable more efficient management of drug selection and distribution in the country. Furthermore, it would enable the evaluation and updating of current treatment regimens with the aim of ensuring timely and appropriate treatment for all cases.
Atovaquone inhibits the cytochrome bc1 complex (cytbc1), an integral membrane protein that catalyzes transmembrane electron transfer (ET) pathway and maintains mitochondrial membrane potential (Korsinczky et al., 2000). Point mutations in the mitochondrial cytochrome b gene of P. falciparum (pfcytb) have been identified as being associated with resistance to this drug (Sutherland et al., 2008). In addition, cycloguanil, the active metabolite of proguanil, is a pfdhfr inhibitor (Fidock et al., 1998), altering pyrimidine biosynthesis, thereby affecting nucleic acid replication in the parasite. The accumulation of mutations in the pfdhfr gene has been associated with resistance to drugs such as proguanil and pyrimethamine (Peterson et al., 1988, 1990). On the other hand, P. vivax confirmed molecular markers for resistance have yet to be identified due to factors such as the difficulty in distinguishing between recrudescence and relapse, or the difficulty in confirming resistance phenotypes ex vivo because in vitro culture of P. vivax is challenging (Buyon et al., 2021). Studying the P. vivax multidrug resistance protein 1 (pvmrp1) gene has drawn interest because it encodes a transmembrane protein member of the ATP-binding cassette (ABC) transporters. Its ortholog in P. falciparum is associated with resistance to multiple drugs such as mefloquine, chloroquine, quinine, artemisinin, piperaquine, and primaquine (Raj et al., 2009; Buyon et al., 2021; Yin et al., 2022). Genetic studies on pvmrp1 demonstrate that it displays high haplotype diversity, a high proportion of non-synonymous SNPs, low nucleotide diversity, and is under purifying selection, making it a putative marker of drug resistance in P. vivax (Dharia et al., 2010; Buyon et al., 2021; Yin et al., 2022).
The National Parasitology Reference Laboratory of the Instituto de Salud Pública (ISP) has been conducting molecular analyses on P. falciparum to identify drug resistance markers in imported malaria cases. Mutant genotypes have been identified in the P. falciparum chloroquine resistance transporter (pfcrt) gene and P. falciparum multidrug resistance associated protein 1 (pfmdr1) gene (Escobar et al., 2020). Molecular markers associated with drug resistance need to be monitored as they provide early warning of resistance emergence; it is therefore necessary to extend the analysis to new markers associated with the main antimalarial drugs, targeting the two most commonly diagnosed species in Chile, P. vivax and P. falciparum.
This study proposes analyzing the genetic profile associated with antimalarial drug resistance in the genes pfcytb, pfdhfr and pvmrp1 in samples from malaria-confirmed travelers and migrants in Chile between 2019 and 2022.
The study population comprised all blood samples (N = 90), both pre- and post-treatment, from 55 malaria cases who had been diagnosed at the National Reference Laboratory for Parasitology of the ISP. The samples were submitted according to the mandatory surveillance process outlined in Supreme Decree 7/2019. Malaria status was assessed by the confirmatory protocol, which has the ability to detect the 5 main species of Plasmodium (P. falciparum, P. vivax, P. malariae, P. ovale and P. knowlesi) and comprises the following steps: 1) Microscopic examination (observation of thin and thick smears); 2) rapid diagnostic tests (RDTs) that detect the parasites proteins, and 3) detection of the parasites genetic material by real-time polymerase chain reaction (qPCR) (Rougemont et al., 2004). The cases were confirmed as positive according to the outcome of the ISP algorithm (Supplementary File S1).
Epidemiological data for confirmed cases were collected from the sample submission form filled out by the referring medical institutions. Of the cases analyzed, 82% were male (N = 45) and 18% were female (N = 10). Regarding the Plasmodium species identified, 78% were caused by P. vivax (N = 43), while 22% were caused by P. falciparum (N = 12). No mixed infections were observed. The median age of the cases was 29 years, with the youngest case being two years old and the oldest case being 61 years old. Most cases (78%, N = 43) involved foreign nationals, while only 22% (N = 12) were Chileans who had visited malaria-endemic countries (Supplementary File S2).
Molecular analyses were conducted on total DNA extracted from blood samples using the QIAamp® DNA Mini Kit (Qiagen) according to the manufacturer’s instructions. The purified DNA was either used immediately or stored at -20 °C until needed. The confirmation of Plasmodium infection and species identification were determined by multiplex qPCR as previously described (Rougemont et al., 2004), employing the StepOnePlus™ Real-Time PCR System (Applied Biosystems). The pfcytb, pfdhfr, and pvmrp1 genes were amplified by nested PCR, employing the primers described in Table 1 and conditions detailed in Supplementary File S5. The amplicons were purified from agarose gels using QIAquick Gel Extraction Kit (Qiagen, Cat#28706). Bi-directional Sanger sequencing of amplicons was performed using BigDye terminator v3.1 (Applied Biosystem, Cat# 4337455), and the following cycling profile: 96 °C for 1 minute, 30 cycles at 96 °C for 10 seconds, 52 °C (pfdhfr), 56 °C (pfcytb) or 58 °C (pvmrp1) for 5 seconds and 60 °C for 2 minutes using the primers listed in Table 1. The sequencing fragments were purified by precipitation using 3 M sodium acetate (pH 5.5, Invitrogen, Cat#AM9740) and absolute ethanol (Merck, Cat#1.00983.2500). Purified fragments were resuspended in Hi-Di formamide (Applied Biosystems, Cat#4311320) and submitted to capillary electrophoresis on the ABI 3500 (Applied Biosystems) equipped with 3500 series Data Collection software (Version 3). Sequencing data normalization was carried out using Sequencing Analysis Software (Version 6). Assembled sequences were trimmed and aligned with reference sequences for mutation identification. Sequence alignments were performed using Sequencher software (version 5.4.6) and the reference sequences: AY282930.1 and XM_001351443.1 from P. falciparum; and XM_001612630.1 from P. vivax for detection of mutations in the pfcytb, pfdhfr, and pvmrp1 genes, respectively.
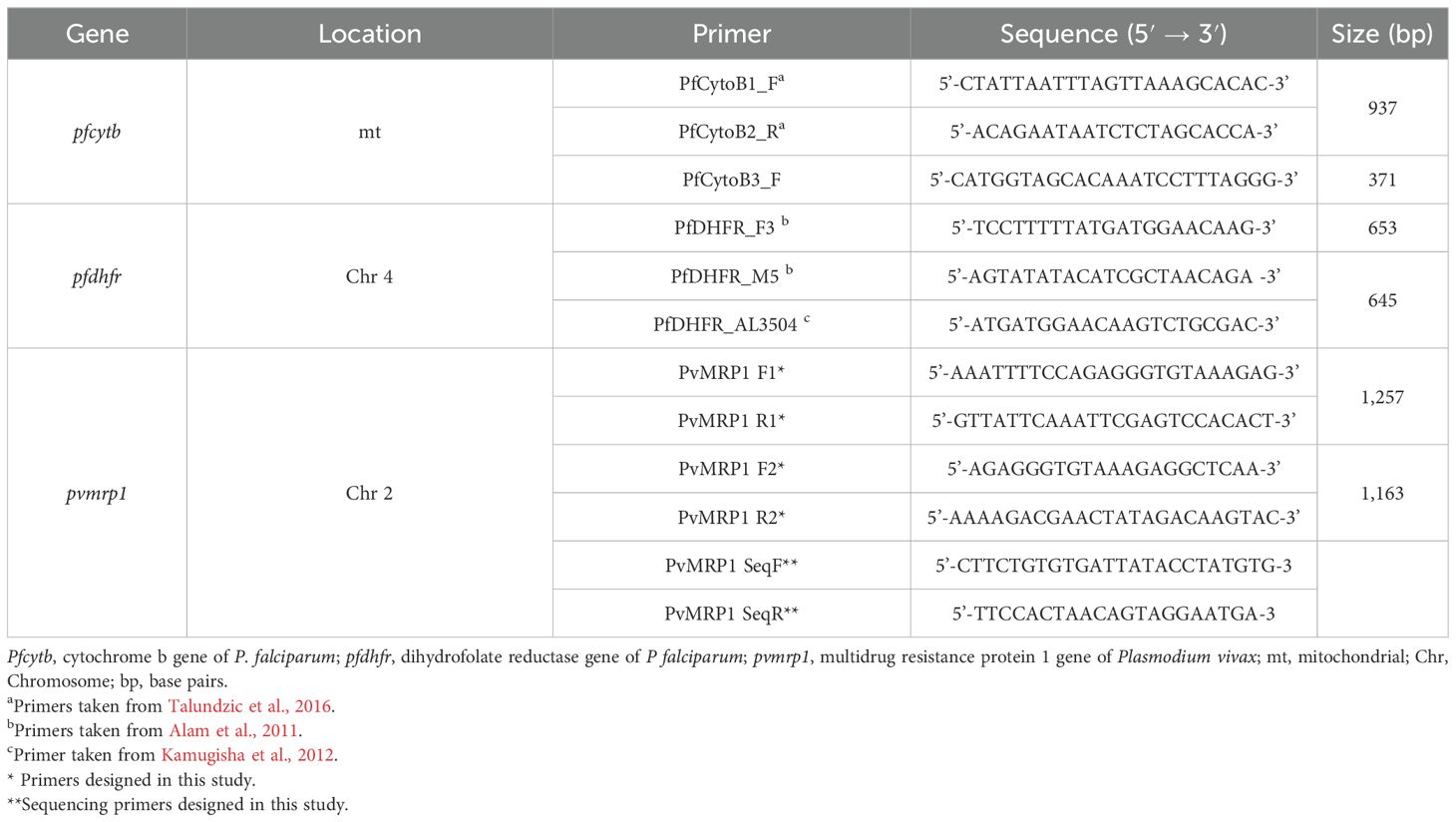
Table 1. Primers used for PCR amplification and DNA sequencing of P. falciparum and P. vivax genes.
Regarding the country of malaria acquisition, a total of 14 countries were identified as epidemiologically linked to the observed cases. Of these, six countries were located in the Americas, which accounted for 82% of cases (N = 45) and 93% (N = 42) of which were attributable to P. vivax. The remaining eight countries were located in Africa, accounting for 18% of cases (N = 10), of which 90% (N = 9) were attributable to P. falciparum. The country with the highest number of cases was Venezuela (N = 28, 50.9%), followed by Colombia (N = 6, 10.9%), Peru (N = 4, 7.3%), Brazil (N = 3, 5.5%), and then Bolivia, Nigeria, and Togo (N = 2, 3.6% each). In addition, Panama, Cameroon, Congo, Ethiopia, Guinea, Ivory Coast, and Uganda, all with one case each (1.8%). Furthermore, there was one case involved travel to both Brazil and Bolivia (1.8%). Of the Latin American countries, Colombia was the only one where travelers contracted P. falciparum (N = 3) infection. Conversely Ethiopia was the only African country where a traveler contracted P. vivax infection (N = 1). Regarding the locations of the cases in Chile, malaria was confirmed in 11 (69%) of the 16 regions that comprise the administrative division. The Metropolitana region accounted for 56% (N = 31) of the cases, followed by Valparaíso with 12.7% (N = 7), Tarapacá with 7.3% (N = 4), and La Araucanía with 5.5% (N = 3). Two cases were reported in each of the Antofagasta, Coquimbo, and Arica y Parinacota regions (3.6%), while one case was reported in each of the Los Lagos, Maule, Ñuble, and O’Higgins regions (1.8%) (Figure 1).

Figure 1. Country of origin/visit and Plasmodium species detected in imported malaria cases in Chile between 2019-2022 by administrative region. The column on the left outlines the Chilean region of malaria onset/diagnosis. The central column indicates the Plasmodium species identified and the country of origin/visit of the cases is listed in column on the right. Numbers indicate the total cases of malaria. Colombia: three cases of P. falciparum and P. vivax malaria each. Metropolitana region: 7 and 24 cases of P. falciparum and P. vivax malaria, respectively. This Sankey diagram was generated using the networkD3 v0.4 package in R v4.1.1 (Allaire et al., 2017).
Among the 90 malaria-confirmed samples received by the ISP between 2019 and 2022, 87 (96.7%), corresponding to 53 patients, met the inclusion criteria and were successfully genotyped for the Plasmodium pfcytb, pfdhfr, and pvmrp1 genes. Two samples were excluded from the genetic study due to the unavailability of backup specimens. Both samples were unique cases of two male travelers; one positive for P. falciparum who visited Ivory Coast and later died, and the other with confirmed P. vivax infection acquired during a trip to Colombia. The third omitted sample was a P. falciparum microscopy-negative post-treatment sample from a male patient that failed to yield amplifiable DNA from the parasite.
Of the 87 sequenced samples, 48 (55.2%) were P. vivax, representing 42 cases, while 39 samples (44.8%) were P. falciparum. One sample positive for P. falciparum failed to yield the expected PCR product for the pfdhfr gene (Supplementary File S3). The total samples sequenced from P. falciparum-positive specimens were 38 (44.2% out of 87) for pfdhfr gene and 39 (44.8% out of 87) for the pfcytb gene, corresponding to 11 cases. All genes sequenced, both in P. falciparum and P. vivax, exhibited evidence of at least one sample with non-synonymous mutation. The pfdhfr gene showed mutations in all 38 samples (11 cases) of P. falciparum. In contrast, only 7.7% (3 out of 39) of the samples exhibited mutations in the pfcytb gene, all of which corresponded to a single patient. Regarding P. vivax, the pfmvrp1 gene was found to be mutated in 100% of the 48 samples (42 cases) (Table 2). Noteworthy, no silent mutations were identified in all the sequences analyzed.

Table 2. Amino acid substitutions in pfdhfr, pfcytb and pvmrp1 genes in samples, cases, country of origin/visit, and destination region.
The mutations in the pfdhfr gene identified in the 11 cases of P. falciparum studied were present in patients who had visited or originated from seven different countries: Colombia (27.2%, N = 3), Nigeria (18.2%, N = 2), Togo (18.2%, N = 2), Congo (9.1%, N = 1), Cameroon (9.1%, N = 1), Uganda (9.1%, N = 1) and Guinea (9.1%, N = 1) (Figure 1. Note the excluded case involved a visit to Ivory Coast).
The imported malaria cases showed three distinct non-synonymous substitutions in the pfdhfr gene i.e., S108N, N51I, and/or C59R. The triple mutation S108N+N51I+C59R were detected in 63.6% (7 out of 11) of the cases, the double mutation S108N+N51I were detected in 27.3% (3 out of 11) of the cases and the unique mutation S108N was identified in 9.1% (1 out of 11) of the cases (Table 2). The S108N substitution was the most common mutation found in all P. falciparum cases (N = 11) and samples (N = 38) tested. This was followed by N51I, which was observed in 90.9% of cases (10 out of 11) and 97.4% of samples (37 out of 38). Finally, the C59R substitution was identified in 66.3% of cases (7 out of 11) and 81.6% of samples (31 out of 38) (Table 2; Supplementary File S3). The mutations were found in both pre- and post-treatment follow-up samples.
Analysis of the pfcytb gene revealed that 90.9% (10 out of 11) of infected cases exhibited the wild-type genotype, with the sole exception of a case involving a patient who had visited Nigeria, which displayed a mutation (Y268C) associated with resistance (Table 2). This de novo mutation was detected during the first post-treatment follow-up. Additionally, this case also showed parasites with the triple mutation N51I+C59R+S108N in the pfdhfr gene.
The mutations in the pvmrp1 gene identified in the 42 cases of P. vivax studied were present in patients who had visited or originated from seven different countries: Venezuela (66.7%, N = 28), Peru (9.5%, N = 4), Brazil (7.1%, N = 3), Colombia (4.8%, N = 2), Bolivia (4,8%, N = 2), Panama (2.4%, N = 1), Ethiopia (2.4%, N = 1), and one case that visited both Brazil and Bolivia (2.4%) (Figure 1. Consider exclusion of one case from Colombia).
A total of five different non-synonymous substitutions (Y1393D/V1478I/G1419A/L1282I/H1586I) were identified in the pvmrp1 gene. These mutations comprise single Y1393D (4.8%, 2 out of 42), double Y1393D+V1478I (52.4%, 22 out of 42), triple Y1393D+V1478I+G1419A (40.5%, 17 out of 42) and quintuple Y1393D+V1478+G1419A+L1282I+H1586I (2.3%, 1 out of 42) (Table 2). The Y1393D mutation was the most common mutation observed in all P. vivax samples tested, followed by V1478I at a frequency of 95.2% (40 out of 42), G1419A at 42.9% (18 out of 42), V1544I at 11.9% (5 out of 42), and finally L1282I and H1586I, both at 2.4% (1 out of 42 each). These mutations were detected in both pre- and post-treatment samples. Refer to Supplementary File S4 for information on the mutations detected in each sample.
The emergence of antimalarial drug resistance is a significant concern for both endemic countries and those that are currently countries certified malaria-free. This study identified mutations in both the pfdhfr and pfcytb genes associated with antimalarial drug resistance in P. falciparum. Additionally, the pvmrp1 gene, which has been proposed as a putative marker of drug resistance in P. vivax, was also found to have mutations. The samples were obtained from travelers and migrants who were confirmed in Chile between 2019 and 2022.
Analysis of the Pfdhfr gene revealed the presence of mutations (N51I+C59R+S108N) associated with different levels of resistance to drugs such as pyrimethamine (Pyr) and cycloguanil (Cyc), an active metabolite of proguanil (Peterson et al., 1990; Sirawaraporn et al., 1997)). The S108N mutation, observed in this study with a high degree of fixation (100%), is associated with low levels of resistance to Pyr and Cyc, and is considered insufficient to result in treatment failure (Rallón et al., 1999). However, it seems necessary for the occurrence of the following mutations (Foote et al., 1990; Peterson et al., 1990; Kublin et al., 1998). The double mutant N51I+S108N, observed in 15.8% of the samples, is associated with moderate levels of resistance of P. falciparum to Pyr and Cyc (Sirawaraporn et al., 1997). Previous reports of this mutant have been documented in parasites from Colombia and Nigeria, which is consistent with the countries of origin/visited by the travelers whose parasites showed these mutations in this study (Zhao et al., 2021; Guerra et al., 2022). The N51I+C59R+S108N genotype, which was the main combination observed (81.6%), has been associated with a high degree of resistance to Pyr and a moderate degree of resistance to Cyc (Sirawaraporn et al., 1997). This triple mutant has been observed mainly in Africa and Asia (Nair et al., 2003; Roper et al., 2003; Sridaran et al., 2010; Yan et al., 2021). No cases of Asian origin were confirmed in Chile during the time period analyzed and, therefore, were not included in this study. The wild-type S108 genotype, which is considered sensitive to both Pyr and Cyc (Foote et al., 1990), was not observed in Pfdhfr, evidencing a high degree of fixation with the S108N mutation. Resistance to Pyr is hypothesized to develop in a stepwise manner, typically commencing with the S108N mutant. Where the number of mutations is greater, the likelihood of treatment failure rises (Bzik et al., 1987; Peterson et al., 1988; Ferlan et al., 2001; Lozovsky et al., 2009). The quadruple mutant N51I+C59R+S108N+I164L, which is associated with high resistance to Pyr and Cyc (Sirawaraporn et al., 1997), was not identified in the samples studied. Furthermore, the S108T mutant was not observed, which, along with A16V, is associated with resistance to Cyc (Foote et al., 1990; Peterson et al., 1990).
In the study of the pfcytb gene, the wild genotype was predominately observed, although the Y268C mutation was also detected, which is associated with resistance to atovaquone (Ato). The mutation was identified in a case of P. falciparum that had been imported from Nigeria and, following treatment with AP, resulted in therapeutic failure. The Y268C mutation was observed exclusively in post-treatment samples (see Supplementary File S3), suggesting that it likely arose de novo during the primary infection due to selective pressure induced by the AP drug used. Previously, cases of Ato resistance associated with the Y268C mutation had been sporadically described in various locations, including in Africa (Musset et al., 2006; Sutherland et al., 2008; Perry et al., 2009; Massamba et al., 2020; Chenet et al., 2021). Moreover, other mutations in codon 268 have been linked to Ato resistance, including Y268N (Fivelman et al., 2002), Y268S (Schwartz et al., 2003), Y268C (Musset et al., 2006) and Y268M (Plucinski et al., 2014). Furthermore, treatment failure with AP has also been observed in the absence of these mutations, indicating that not all mechanisms are fully understood (Wichmann et al., 2004). In Nigeria, the origin of the identified mutants, both Y268N (Fivelman et al., 2002; Happi et al., 2006), Y268S (Plucinski et al., 2014), Y268M (Plucinski et al., 2014) and Y268C (Chenet et al., 2021) have been previously reported. In addition to mutations in the pfcytb gene, the case exhibited the triple mutant N51I, C59R, S108N in the pfdhfr gene, which is associated with resistance to Cyc (Sirawaraporn et al., 1997). A full case report of this AP resistance case has been published (Chenet et al., 2021).
In the analysis of the Pvmrp1 gene of P. vivax, the most frequently observed mutations, Y1393D (100%) and V1478I (95.8%), have been previously reported in Asia, Africa, South America, and Oceania (Benavente et al., 2021; Yin et al., 2022). In areas such as the China-Myanmar border, these mutations also exhibit frequencies approaching fixation (Zeng et al., 2021). The double mutant Y1393D+V1478I, which was the most observed combination (50%), was identified less frequently than in Asia (95.65%) (Zeng et al., 2021). The G1419A mutation, observed in smaller circulation than Y1393D and V1478I, has been reported in South Asia, Africa, and South America (Benavente et al., 2021; Zeng et al., 2021). The triple mutant G1419A+Y1393D+V1478I, detected in 40% of the samples, has been reported with a lower frequency (2.17%) in the Asian population (Zeng et al., 2021). The L1282I and H1586Y mutations have been reported with a low frequency. In the present study, these mutations were observed exclusively in samples from Peru. The H1586Y mutation has been identified in South America, Asia, and Oceania. In contrast, no records of the L1282I mutation have been observed outside the Americas (Benavente et al., 2021; Yin et al., 2022).
Thus far, no mutation in Pvmrp1 has been validated as genetic marker for antimalarial drug resistance. Ex vivo studies of P. vivax isolates have demonstrated a correlation between V1478 and elevated chloroquine (CQ) IC50 values. However, further studies are needed to validate this finding. Conversely, the G1419A and V1478I mutations have been ex vivo linked to reduced susceptibility to artesunate and dihydroartemisinin (Zeng et al., 2021). Additionally, the G1419A mutation has been associated with reduced sensitivity to primaquine, mefloquine, and quinine (Zeng et al., 2021). Further research into resistance genes in primaquine would be beneficial in Pvmrp1 and other putative genes, given the drug’s extensive use in the treatment of P. vivax and the reports of treatment failure (Thomas et al., 2016). Additionally, it would be valuable to assess the patterns that reflect the selective pressure towards drug resistance and the heterogeneity of mutations in different geographic clusters (Ali et al., 2024).
The presence of malaria cases carrying mutations related to resistance in Chile implies a difficulty for ensure the diagnosis and treatment of resistant cases in a country with a low incidence and limited access to antimalarial drugs throughout the territory. Furthermore, the circulation of mutations linked to resistance in areas with the presence of the transmitting vector, such as the regions of Arica y Parinacota and Tarapacá in Chile (Valderrama et al., 2021), would pose significant challenges in managing the risk of malaria reintroduction. This is due to the evidence of species of resistant Plasmodium establishing themselves in new territories (Roper et al., 2004). Moreover, these regions of the country also represent an important area of migratory flow and land connection with the rest of the continent, thereby increasing the risk of autochthonous cases.
The geographical origin of malaria-positive travelers and the low number of malaria cases in Chile, as it is a non-endemic country, may have hindered the observation of other mutations of interest in this study. However, continuous monitoring is essential given Chile’s status as a prominent global tourist destination. It is possible that additional Plasmodium species and novel mutations related with resistance to other drugs could be observed. This work strengthens the strategy for prevention of the reintroduction of malaria in Chile. The strategy encompasses an integrated surveillance system (human, environmental and vector-based), ensuring diagnosis, mandatory reporting, effective treatment and epidemiological investigation of each case. This approach allows for the assessment of drug management in accordance with the local resistance profile and expanding the profile of useful markers for confirming of therapeutic failure due to genetic resistance.
Identifying malaria cases in Chile with the potential to harbor or develop mutations associated with drug resistance should encourage other non-endemic countries to collaborate in global drug-resistance surveillance. This will facilitate the provision of data on the epidemiology of malaria in endemic regions where surveillance systems are weak (Tatem et al., 2017). It is therefore crucial to expand the use and validation of new confirmed antimalarial drug resistance markers. This will facilitate the genetic confirmation of cases of treatment failure and make it possible to monitor the circulation, introduction, and spread of resistance in different territories.
The present study identified mutations in three genes linked/associated to antimalarial resistance i.e., pfdhfr and pfcytb genes in P. falciparum and pvmrp1 gene in P. vivax (putative drug resistance marker). Noteworthy, mutations were present in several samples analyzed ranging from at least one for the pfcytb gene and 100% of the samples for both, pfdhfr and pvmrp1 genes. The emergence of drug resistance represents a threat to the availability of effective treatment options in regions where malaria is not endemic. Furthermore, the vector’s presence in these regions could hinder local efforts to address the emergence of new autochthonous cases. Molecular markers are an invaluable resource for monitoring and confirming resistance to the main antimalarial drugs in use. Further research is needed to expand the investigation of potential resistance markers in additional genes, particularly in P. vivax, the species responsible for the highest number of cases in Chile and a growing public health concern.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
The studies involving humans were approved by Scientific Ethics Committee of the Eastern Metropolitan Health Service of Santiago, Chile (SSMOriente110723). The studies were conducted in accordance with the local legislation and institutional requirements. Written informed consent for participation was not required from the participants or the participants’ legal guardians/next of kin because the samples used come from the national surveillance system and were analyzed anonymously without exposing sensitive data.
AO: Investigation, Project administration, Supervision, Writing – original draft, Writing – review & editing. LH: Formal analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing. EC: Formal analysis, Investigation, Methodology, Writing – review & editing, Conceptualization, Data curation, Writing – original draft. BP: Data curation, Formal analysis, Investigation, Writing – review & editing. JF: Data curation, Formal analysis, Investigation, Writing – review & editing. MS: Project administration, Supervision, Writing – review & editing. NS: Project administration, Supervision, Writing – review & editing. MJ: Funding acquisition, Project administration, Supervision, Writing – original draft, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This research was funded by the Ministry of Science, Technology, Knowledge and Innovation of Chile, within the framework of a transfer agreement to the Institute of Public Health for the purpose of strengthening its technical and research capacities. Providencia N°. 1462. Resolución exenta N°1258, April 2024.
We extend our gratitude to all the people who make up the national epidemiological surveillance network, including health establishments, blood services, health services, Regional Ministerial Secretariats (SEREMI) of Health, the ISP, and the Ministry of Health (MINSAL). Their invaluable contributions were essential to generate the data used in this study.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmala.2024.1516733/full#supplementary-material
Supplementary File S1 | ISP Malaria Laboratory Testing Algorithm.
Supplementary File S2 | Description of confirmed malaria samples and cases in Chile, 2019-2022.
Supplementary File S3 | Mutations identified in codons 51, 59, 108 of pfdhfr and codon 268 of pfcytb gene amplified from 38 and 39 isolates of P. falciparum confirmed malaria cases. Polymorphic amino acids are highlighted in blue for each haplotype. The color pink indicates amino acid residues that are identical to the reference sequences XM_001351443.1 and AY282930.1 for pfdhfr and pfcytb, respectively. The total number of mutations for each sample is indicated in the column on the right. N/A indicates that sample did not amplify by PCR. Samples shaded in the same color are from the same confirmed case.
Supplementary File S4 | Mutations identified in codons 1282, 1393, 1419, 1478, 1525, 1544 and 1586 of the pvmrp1 gene amplified from 48 isolates of confirmed P. vivax malaria. Polymorphic amino acids are highlighted in blue for each haplotype. The color pink indicates amino acid residues that are identical to the reference sequences XM_001612630. The total number of mutations for each sample is indicated in the column on the right. Asterisk indicates co-detection of wild-type copies. Samples shaded in the same color are from the same confirmed case.
Supplementary File S5 | Amplification of Plasmodium falciparum dihydrofolate reductase (dhfr) and cytochrome b (cytb) genes and Plasmodium vivax putative multidrug resistance- associated protein 1 (mrp1) gene by polymerase chain reaction (PCR).
Alam M. T., de Souza D. K., Vinayak S., Griffing S. M., Poe A. C., Duah N. O., et al. (2011). Selective sweeps and genetic lineages of Plasmodium falciparum drug -resistant alleles in Ghana. J. Infect. Dis. 203, 220–227. doi: 10.1093/infdis/jiq038
Ali N. A., Nema S., Schneider K. A., Krishna S., Verma A. K., Das A., et al. (2024). Low prevalence of antimalarial resistance mutations in India during 2014-2015: impact of combining first-line therapy with primaquine. J. Infect. Dis. 229, 1574–1583. doi: 10.1093/infdis/jiad553
Allaire J. J., Ellis P., Gandrud C., Kuo K., Lewis B. W., Owen J., et al. (2017). networkD3: D3 JavaScript network graphs from R. R package version 0.4. Available online at: http://CRAN.R-project.org/package=networkD3 (Accessed December 5, 2024).
Balikagala B., Fukuda N., Ikeda M., Katuro O. T., Tachibana S.-I., Yamauchi M., et al. (2021). Evidence of artemisinin-resistant malaria in Africa. New Engl. J. Med. 385, 1163–1171. doi: 10.1056/NEJMoa2101746
Benavente E. D., Manko E., Phelan J., Campos M., Nolder D., Fernandez D., et al. (2021). Distinctive genetic structure and selection patterns in Plasmodium vivax from South Asia and East Africa. Nat. Commun. 12, 3160. doi: 10.1038/S41467-021-23422-3
Buyon L. E., Elsworth B., Duraisingh M. T. (2021). The molecular basis of antimalarial drug resistance in Plasmodium vivax. Int. J. Parasitol. Drugs Drug Resist. 16, 23–37. doi: 10.1016/J.IJPDDR.2021.04.002
Bzik D. J., Li W. B., Horii T., Inselburg J. (1987). Molecular cloning and sequence analysis of the Plasmodium falciparum dihydrofolate reductase-thymidylate synthase gene. Proc. Natl. Acad. Sci. U.S.A. 84, 8360–8364. doi: 10.1073/PNAS.84.23.8360
Chenet S. M., Oyarce A., Fernandez J., Tapia-Limonchi R., Weitzel T., Tejedo J. R., et al. (2021). Atovaquone/proguanil resistance in an imported malaria case in Chile. Am. J. Trop. Med. Hyg. 104, 1811–1813. doi: 10.4269/ajtmh.20-1095
Dharia N. V., Bright A. T., Westenberger S. J., Barnes S. W., Batalov S., Kuhen K., et al. (2010). Whole-genome sequencing and microarray analysis of ex vivo Plasmodium vivax reveal selective pressure on putative drug resistance genes. Proc. Natl. Acad. Sci. U S A. 107, 20045–20050. doi: 10.1073/PNAS.1003776107
Escobar D. F., Lucchi N. W., Abdallah R., Valenzuela M. T., Udhayakumar V., Jercic M. I., et al. (2020). Molecular and epidemiological characterization of imported malaria cases in Chile. Malar J. 19, 289. doi: 10.1186/S12936-020-03353-Y
Ferlan J. T., Mookherjee S., Okezie I. N., Fulgence L., Sibley C. H. (2001). Mutagenesis of dihydrofolate reductase from Plasmodium falciparum: Analysis in Saccharomyces cerevisiae of triple mutant alleles resistant to pyrimethamine or WR99210. Mol. Biochem. Parasitol. 113, 139–150. doi: 10.1016/S0166-6851(01)00207-9
Fidock D. A., Nomura T., Wellems T. E. (1998). Cycloguanil and its parent compound proguanil demonstrate distinct activities against Plasmodium falciparum malaria parasites transformed with human dihydrofolate reductase. Mol. Pharmacol. 54, 1140–1147. doi: 10.1124/mol.54.6.1140
Fivelman Q. L., Butcher G. A., Adagu I. S., Warhurst D. C., Pasvol G. (2002). Malarone treatment failure and in vitro confirmation of resistance of Plasmodium falciparum isolate from Lagos, Nigeria. Malar J. 1, 1. doi: 10.1186/1475-2875-1-1
Foote S. J., Galatis D., Cowman A. F. (1990). Amino acids in the dihydrofolate reductase-thymidylate synthase gene of Plasmodium falciparum involved in cycloguanil resistance differ from those involved in pyrimethamine resistance. Proc. Natl. Acad. Sci. U S A. 87, 3014–3017. doi: 10.1073/PNAS.87.8.3014
Guerra A. P., Olivera M. J., Cortés L. J., Chenet S. M., Macedo de Oliveira A., Lucchi N. W. (2022). Molecular surveillance for anti-malarial drug resistance and genetic diversity of Plasmodium falciparum after chloroquine and sulfadoxine-pyrimethamine withdrawal in Quibdo, Colombia 2018. Malar J. 21, 306. doi: 10.1186/S12936-022-04328-x
Happi C. T., Gbotosho G. O., Folarin O. A., Milner D., Sarr O., Sowunmi A., et al. (2006). Confirmation of emergence of mutations associated with atovaquone-proguanil resistance in unexposed Plasmodium falciparum isolates from Africa. Malar J. 5, 82. doi: 10.1186/1475-2875-5-82
Ippolito M. M., Moser K. A., Jean-Bertin, Kabuya B., Cunningham C., Juliano J. J. (2021). Antimalarial drug resistance and implications for the WHO global technical strategy. Curr. Epidemiol. Rep. 8, 46–62. doi: 10.1007/S40471-021-00266-5
Kamugisha E., Jing S., Minde M., Kataraihya J., Kongola G., Kironde F., et al. (2012). Efficacy of artemether-lumefantrine in treatment of malaria among under-fives and prevalence of drug resistance markers in Igombe-Mwanza, north-western Tanzania. Malar. J. 11, 58. doi: 10.1186/1475-2875-11-58
Korsinczky M., Chen N., Kotecka B., Saul A., Rieckmann K., Cheng Q. (2000). Mutations in Plasmodium falciparum cytochrome b that are associated with atovaquone resistance are located at a putative drug-binding site. Antimicrob. Agents Chemother. 44, 2100–2108. doi: 10.1128/aac.44.8.2100-2108.2000
Krudsood S., Tangpukdee N., Wilairatana P., Phophak N., Baird J. K., Brittenham G. M., et al. (2008). High-dose Primaquine Regimens against Relapse of Plasmodium vivax Malaria. Am. J. Trop. Med. Hyg. 78, 736–740. doi: 10.4269/ajtmh.2008.78.736
Kublin J. G., Witzig R. S., Shankar A. H., Zurita J. Q., Gilman R. H., Guarda J. A., et al. (1998). Molecular assays for surveillance of antifolate-resistant malaria. Lancet 351, 1629–1630. doi: 10.1016/S0140-6736(98)24022-0
Lozovsky E. R., Chookajorn T., Brown K. M., Imwong M., Shaw P. J., Kamchonwongpaisan S., et al. (2009). Stepwise acquisition of pyrimethamine resistance in the malaria parasite. Proc. Natl. Acad. Sci. U S A. 106, 12025–12030. doi: 10.1073/PNAS.0905922106
Massamba L., Madamet M., Benoit N., Chevalier A., Fonta I., Mondain V., et al. (2020). Late clinical failure associated with cytochrome b codon 268 mutation during treatment of falciparum malaria with atovaquone–proguanil in traveller returning from Congo. Malar J. 19, 37. doi: 10.1186/S12936-020-3126-Y
MINSAL (2015). Orientaciones técnicas para el diagnóstico y tratamiento de la malaria en Chile Santiago (Ministerio de Salud Pública de Chile). Available online at: http://www.repositoriodigital.minsal.cl/handle/2015/833?show=full (Accessed 10 Jan 2019).
Musset L., Bouchaud O., Matheron S., Massias L., Le Bras J. (2006). Clinical atovaquone-proguanil resistance of Plasmodium falciparum associated with cytochrome b codon 268 mutations. Microbes Infect. 8, 2599–2604. doi: 10.1016/J.MICINF.2006.07.011
Nair S., Williams J. T., Brockman A., Paiphun L., Mayxay M., Newton P. N., et al. (2003). A selective sweep driven by pyrimethamine treatment in southeast asian malaria parasites. Mol. Biol. Evol. 20, 1526–1536. doi: 10.1093/molbev/msg162
Perry T. L., Pandey P., Grant J. M., Kain K. C. (2009). Severe atovaquone-resistant Plasmodium falciparum malaria in a Canadian traveller returned from the Indian subcontinent. Open Med. 3, e10–e16.
Peterson D. S., Milhous W. K., Wellems T. E. (1990). Molecular basis of differential resistance to cycloguanil and pyrimethamine in Plasmodium falciparum malaria. Proc. Natl. Acad. Sci. U S A. 87, 3018–3022. doi: 10.1073/PNAS.87.8.3018
Peterson D. S., Walliker D., Wellems T. E. (1988). Evidence that a point mutation in dihydrofolate reductase-thymidylate synthase confers resistance to pyrimethamine in falciparum malaria. Proc. Natl. Acad. Sci. U S A. 85, 9114–9118. doi: 10.1073/PNAS.85.23.9114
Phyo A. P., Dahal P., Mayxay M., Ashley E. A. (2022). Clinical impact of vivax malaria: A collection review. PloS Med. 19, e1003890. doi: 10.1371/journal.pmed.1003890
Plucinski M. M., Huber C. S., Akinyi S., Dalton W., Eschete M., Grady K., et al. (2014). Novel mutation in cytochrome B of plasmodium falciparum in one of two atovaquone-proguanil treatment failures in travelers returning from same site in Nigeria. Open Forum Infect. Dis. 1, ofu059. doi: 10.1093/ofid/ofu059
Price R. N., Douglas N. M., Anstey N. M. (2009). New developments in Plasmodium vivax malaria: severe disease and the rise of chloroquine resistance. Curr. Opin. Infect. Dis. 22, 430–435. doi: 10.1097/QCO.0b013e32832f14c1
Raj D. K., Mu J., Jiang H., Kabat J., Singh S., Sullivan M., et al. (2009). Disruption of a plasmodium falciparum multidrug resistance-associated protein (PfMRP) alters its fitness and transport of antimalarial drugs and glutathione. J. Biol. Chem. 284, 7687–7696. doi: 10.1074/jbc.M806944200
Rallón N. I., Osorio L. E., Giraldo L. E. (1999). Lack of an association between the ASN-108 mutation in the dihydrofolate reductase gene and in vivo resistance to sulfadoxine/pyrimethamine in Plasmodium falciparum. Am. J. Trop. Med. Hyg. 61, 245–248. doi: 10.4269/ajtmh.1999.61.245
Roper C., Pearce R., Bredenkamp B., Gumede J., Drakeley C., Mosha F., et al. (2003). Antifolate antimalarial resistance in southeast Africa: a population-based analysis. Lancet 361, 1174–1181. doi: 10.1016/S0140-6736(03)12951-0
Roper C., Pearce R., Nair S., Sharp B., Nosten F., Anderson T. (2004). Intercontinental spread of pyrimethamine-resistant malaria. Science 305, 1124. doi: 10.1126/science.1098876
Rougemont M., Van Saanen M., Sahli R., Hinrikson H. P., Bille J., Jaton K. (2004). Detection of four Plasmodium species in blood from humans by 18S rRNA gene subunit-based and species-specific real-time PCR assays. J. Clin. Microbiol. 42, 5636–5643. doi: 10.1128/JCM.42.12.5636-5643.2004
Schwartz E., Bujanover S., Kain K. C. (2003). Genetic confirmation of atovaquone-proguanil-resistant Plasmodium falciparum malaria acquired by a nonimmune traveler to East Africa. Clin. Infect. Dis. 37, 450–451. doi: 10.1086/375599
Sirawaraporn W., Sathitkul T., Sirawaraporn R., Yuthavong Y., Santi D. V. (1997). Antifolate-resistant mutants of Plasmodium falciparum dihydrofolate reductase. Proc. Natl. Acad. Sci. U S A. 94, 1124–1129. doi: 10.1073/pnas.94.4.1124
Sridaran S., McClintock S. K., Syphard L. M., Herman K. M., Barnwell J. W., Udhayakumar V. (2010). Anti-folate drug resistance in Africa: meta-analysis of reported dihydrofolate reductase (dhfr) and dihydropteroate synthase (dhps) mutant genotype frequencies in African Plasmodium falciparum parasite populations. Malar J. 9, 247. doi: 10.1186/1475-2875-9-247
Sutherland C. J., Laundy M., Price N., Burke M., Fivelman Q. L., Pasvol G., et al. (2008). Mutations in the Plasmodium falciparum cytochrome b gene are associated with delayed parasite recrudescence in malaria patients treated with atovaquone-proguanil. Malar J. 7, 240. doi: 10.1186/1475-2875-7-240
Talundzic E., Plucinski M. M., Biliya S., Silva-Flannery L. M., Arguin P. M., Halsey E. S., et al. (2016). Advanced molecular detection of malarone resistance. Antimicrob. Agents Chemother. 60, 3821–3823. doi: 10.1128/AAC.00171-16
Tatem A. J., Jia P., Ordanovich D., Falkner M., Huang Z., Howes R., et al. (2017). The geography of imported malaria to non-endemic countries: a meta-analysis of nationally reported statistics. Lancet Infect. Dis. 17, 98–107. doi: 10.1016/S1473-3099(16)30326-7
Thomas D., Tazerouni H., Sundararaj K. G., Cooper J. C. (2016). Therapeutic failure of primaquine and need for new medicines in radical cure of Plasmodium vivax. Acta Trop. 160, 35–38. doi: 10.1016/j.actatropica.2016.04.009
Valderrama L., Ayala S., Reyes C., González C. R. (2021). Modeling the potential distribution of the malaria vector anopheles (Ano.) pseudopunctipennis theobald (Diptera: culicidae) in arid regions of Northern Chile. Front. Public Health 9. doi: 10.3389/fpubh.2021.611152
Vinetz J. M. (2006). Emerging chloroquine-resistant Plasmodium vivax (Benign Tertian) malaria: the need for alternative drug treatment. Clin. Infect. Dis. 42, 1073–1074. doi: 10.1086/501353
Wichmann O., Muehlen M., Gruss H., Mockenhaupt F. P., Suttorp N., Jelinek T. (2004). Malarone treatment failure not associated with previously described mutations in the cytochrome b gene. Malar J. 3, 14. doi: 10.1186/1475-2875-3-14
World Health Organization (2000). Report on antimalarial drug efficacy, resistance and response 10 years of surveillance. Available online at: https://www.who.int/publications/i/item/9789240012813 (Accessed November 19, 2020).
World Health Organization (2021). Global technical strategy for malaria 2016–2030. Available online at: https://apps.who.int/iris/handle/10665/342995 (Accessed July 18, 2023).
World Health Organization (2023a). World malaria report 2023. Available online at: https://www.who.int/publications/i/item/9789240086173 (Accessed December 25, 2023).
World Health Organization (2023b). WHO guidelines for malaria. Available online at: http://apps.who.int/bookorders (Accessed December 26, 2023).
World Health Organization (2024). Countries and territories certified malaria-free by WHO. Available online at: https://www.who.int/teams/global-malaria-programme/elimination/countries-and-territories-certified-malaria-free-by-who (Accessed October 20, 2024).
Yan H., Feng J., Yin J. H., Huang F., Kong X. L., Lin K. M., et al. (2021). High Frequency Mutations in pfdhfr and pfdhps of Plasmodium falciparum in Response to Sulfadoxine-Pyrimethamine: A Cross-Sectional Survey in Returning Chinese Migrants From Africa. Front. Cell Infect. Microbiol. 11. doi: 10.3389/FCIMB.2021.673194
Yin Y., Chen G., Nyunt M. H., Zhang M., Liu Y., Zhu G., et al. (2022). Prevalence of pvmrp1 polymorphisms and its contribution to antimalarial response. Microorganisms 10, 1482. doi: 10.3390/microorganisms10081482/S1
Zeng W., Zhao H., Zhao W., Yang Q., Li X., Li X., et al. (2021). Molecular surveillance and ex vivo drug susceptibilities of plasmodium vivax isolates from the China–Myanmar border. Front. Cell Infect. Microbiol. 11. doi: 10.3389/fcimb.2021.738075/FULL
Keywords: malaria, Plasmodium falciparum, Plasmodium vivax, pfcytb, pfdhfr, pvmrp1, genotyping, drug resistant marker
Citation: Oyarce A, Huaccha L, Cordero EM, Parra B, Fernández J, Soto-Marchant ME, Santis-Alay NT and Jercic MI (2025) Molecular identification of pfcytb, pfdhfr, and pvmrp1 mutations in imported malaria cases in Chile. Front. Malar. 2:1516733. doi: 10.3389/fmala.2024.1516733
Received: 24 October 2024; Accepted: 20 December 2024;
Published: 14 January 2025.
Edited by:
Annette Elizabeth Kaiser, University of Duisburg-Essen, GermanyReviewed by:
Stephen Rogerson, The University of Melbourne, AustraliaCopyright © 2025 Oyarce, Huaccha, Cordero, Parra, Fernández, Soto-Marchant, Santis-Alay and Jercic. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alan Oyarce, YW95YXJjZUBpc3BjaC5jbA==
†These authors share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.