 Yokomi N. Lozano-Sardaneta1
Yokomi N. Lozano-Sardaneta1 Nadia Joselyne Soto-Olguín1
Nadia Joselyne Soto-Olguín1 Jorge J. Rodríguez-Rojas2
Jorge J. Rodríguez-Rojas2 Sokani Sánchez-Montes1,3
Sokani Sánchez-Montes1,3 Eduardo A. Rebollar-Téllez4*
Eduardo A. Rebollar-Téllez4* Ingeborg Becker1*
Ingeborg Becker1*- 1Centro de Medicina Tropical, Unidad de Medicina Experimental, Facultad de Medicina, Universidad Nacional Autónoma de México, Ciudad de México, Mexico
- 2Universidad Autónoma de Nuevo León, Centro de Investigación y Desarrollo en Ciencias de la Salud, Unidad de Patógenos y Vectores, Monterrey, Mexico
- 3Facultad de Ciencias Biológicas y Agropecuarias, Región Tuxpan, Universidad Veracruzana, Tuxpan de Rodríguez Cano, Mexico
- 4Universidad Autónoma de Nuevo León, Facultad de Ciencias Biológicas, Departamento de Zoología de Invertebrados, Laboratorio de Entomología Médica, San Nicolás de los Garza, Mexico
Phlebotomine sand flies are vectors of Leishmania spp., Bartonella bacilliformis, and several arboviruses worldwide. In Mexico, the presence of Bartonella species is associated sporadically with arthropods and little is known on the diversity of insects that could be incriminated with its transmission. The aim of this study was to perform a molecular detection of Bartonella DNA in sand fly species collected in northeastern Mexico. Sand flies were collected at the states of Nuevo Leon and Tamaulipas from June to August 2010, using 16 light traps per night. Sand fly species were morphologically identified, and for Bartonella detection, we amplified ~378 bp of the citrate synthase gene (gltA). DNA sequences were compared in a phylogenetic reconstruction based on maximum likelihood. A total of 532 specimens from seven sand fly species were morphologically identified, where 11 specimens from Tamaulipas tested positive for the presence of a new lineage of Bartonella sp. associated with Psathyromyia shannoni and Lutzomyia cruciata. This work represents the second record of Bartonella-associated with sand flies outside of the endemic area of Carrion’s disease. More studies are necessary to understand their life cycle, transmission dynamics, and their relationship with sand fly species.
Introduction
The phlebotomine sand flies (Diptera: Psychodidae: Phlebotominae) are insects of great relevance due to their role as vectors of several emerging and reemerging diseases caused by pathogens such as Leishmania, Bartonella bacilliformis, and some arboviruses worldwide (1).
The bacterium Bartonella bacilliformis is an endemic species of the Andean valleys of Peru, Colombia, and Ecuador and considered the causal agent of the Carrion’s disease or human bartonellosis, which is transmitted by several sand fly species (2). The genus Bartonella encompasses approximately 33 species of Gram-negative, facultative intracellular, and hemotropic bacteria, which can cause chronic intraerythrocytic infections in their hosts. These bacteria are transmitted by several taxa of hematophagous arthropods (fleas, sucking lice, and sand flies) worldwide (3–5). These pathogenic species are highly prevalent and can cause severe infection patterns, which could be related to the increasing number of emerging and reemerging diseases. Thus, it is important to elucidate the possible transmission routes to combat transmission and infections (3, 5).
In Mexico, Bartonella species have been reported in 10 states, and the record of new genetic lineages associated with louse, fleas, and rodents has increased over the last years (6, 7). Yet, studies of this bacterium associated with arthropods have been conducted sporadically and little is known on the insects involved in its transmission (8–10). Recently, our group reported the first study of Bartonella sp. in sand flies from the state of Veracruz, which is outside of the known endemic zone (11). Due to the relevance of the finding, more studies were warranted to detect this bacterium in sand flies and to elucidate whether it could be of relevance in the transmission of these pathogens in the country.
Until now, 54 sand fly species have been reported in Mexico, with Chiapas, Quintana Roo, Campeche, Yucatan, and Veracruz being the states with the highest number of species. Yet, the analysis for Bartonella detection was generally not performed (11, 12). The aim of this study was to perform a molecular detection and to determine the prevalence of Bartonella species in phlebotomine sand flies collected in the states of Nuevo Leon and Tamaulipas, Mexico.
Material and Methods
Study Area and Sampling Collection
The study sites were located in Santiago and Cadereyta Jiménez of the state of Nuevo Leon and in Gómez Farías of the state of Tamaulipas (Figure 1). In the municipality of Santiago, the mean annual temperature is 20.9°C, with a mean annual precipitation of 1,015 mm. Its vegetation is characterized by pine and oak trees (e.g., Quercus rysophylla, Quercus laeta, Pinus teocote, and Pinus pseudostrobus). In contrast, in the municipality of Cadereyta Jiménez, the climate is a dry, warm steppe with an average annual temperature of 23°C and an average annual rainfall of 601–800 mm. The sampling place consisted of areas of abandoned crops, secondary vegetation (herbaceous and grasses), and primary vegetation composed of species of Prosopis glandulosa, Acacia farnesiana, Ehretia anacua, and Pithecellobium ebano. The municipality of Gómez Farías is located in a transition zone in the state of Tamaulipas, where notable neotropical and Nearctic biotic elements overlap. It has a warm sub-humid climate and an average annual temperature of 25°C, with precipitation ranging from 1,558 to 1,778 mm. It is characterized by a tropical deciduous and sub-deciduous forest, with the predominance of Mangifera indica, Ceiba pentandra, Brosimum alicastrum, and Mirandaceltis monoica (13).
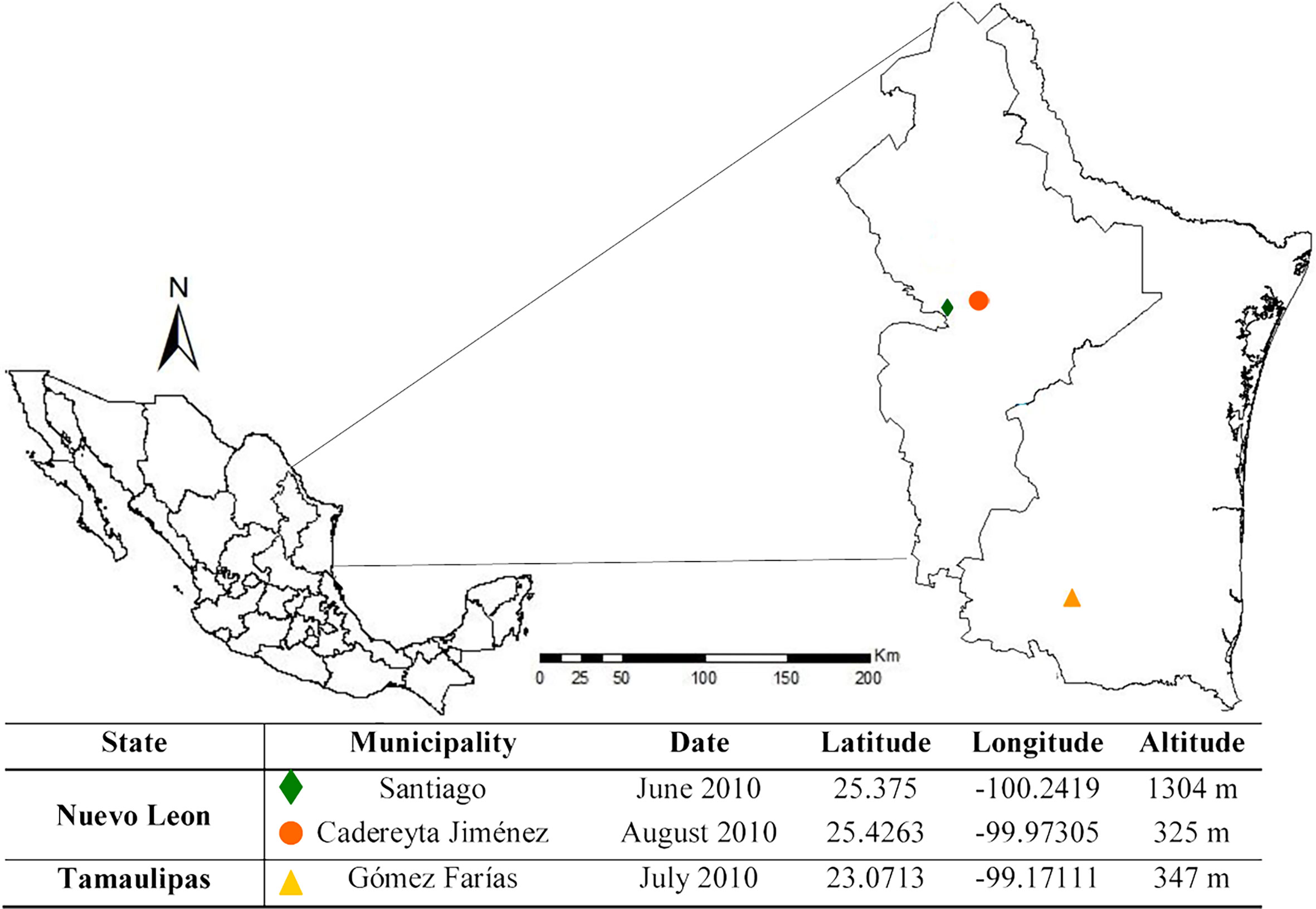
Figure 1 Map showing the study areas of Nuevo Leon and Tamaulipas. The diamond, circle, and triangle depict the municipalities.
Sand flies were collected during three consecutive nights from June to August 2010. Sampling was carried out from 18:00 to 07:00 h, using 16 incandescent light traps (model 512; John W. Hock Co., Gainesville, FL, USA) per night. A transect of 300 m, with at least a 25-m distance between each trap, was used. Traps were hung from trees or branches, approximately 1.5 m above the ground. The specimens were collected daily and were killed using ether vapors. The collected sand fly specimens were sorted from other insects, preserved in vials containing 70% ethanol, and kept at 4°C until their molecular analysis.
Identification of Sand Fly Species
The sand flies were mounted on microscope glass slides using Euparal® medium (BioQuip Products, Rancho Dominguez, CA, USA). The collected female sand flies were dissected, to separate the head from the rest of the body, while the last segments of the abdomen were also severed and preserved for later species identification. Male sand flies were slide-mounted laterally, the head was placed on the side, and the species were identified using morphological characters. Phlebotomine sand fly species were classified according to the classification proposal by Galati (14) and the abbreviation system proposed by Marcondes (15). All the sand fly specimens were deposited in the collection of the Laboratorio de Entomología Médica of the Universidad Autónoma de Nuevo León (UANL) (codes: RECL-NL-20-06-2010; RECL-NL-28-07-2010; RECL-NL-27-08-2010).
DNA Extraction and Polymerase Chain Reaction
Genomic DNA was extracted from the thorax and abdomen of each specimen analyzed, using the phenol chloroform isoamyl alcohol technique (16). For detection of Bartonella species, we amplified a segment of ~378 bp of the citrate synthase (gltA) gene, using the primers BhCS871.p and BhCS1137.n (17). The polymerase chain reactions (PCRs) were performed under the following conditions: initial denaturation at 95°C for 3 min, followed by 35 cycles at 95°C for 30 s, 51°C for 30 s, 72°C for 30 s, and a final extension at 72°C for 7 min (18). The reaction mixture was prepared in a final volume of 25 µl containing 12.5 µl GoTaq® Green Master Mix 2X Promega Corporation (Madison, WI, USA), 1 µl of each primer (100 ng each), 10 µl DNA template (~50 ng), and 0.5 µl nuclease-free water. For the positive control, we used DNA of Bartonella vinsonni obtained from Hoplopleura hirsuta (GenBank code KP9972050.1). The negative control consisted of ultrapure water instead of DNA. The amplified products were analyzed by electrophoresis in 1.5% agarose gels stained with SYTO™ (Invitrogen, CA, USA). Positive PCR products were sequenced at the Laboratorio Nacional de Biodiversidad (Laboratorio de Secuenciación Genómica de la Biodiversidad y de la Salud, Instituto de Biología, UNAM).
Molecular Analysis
The electropherograms were visualized and edited in the Chromas software. Each sequence was compared with available sequences in the GenBank database using the BLASTn tool as a preliminary confirmation of Bartonella amplification. The DNA sequences were aligned with other reference sequences of Bartonella species deposited in GenBank using MEGA version 6.0. We made a phylogenetic reconstruction based on the maximum likelihood (ML) inference method in MEGA 6.0 with 1,000 non-parametric bootstraps, using the Tamura three-parameter model (T92) + gamma distribution and a BIC score of 3295.044. All alignment sites with less than 95% site coverage were eliminated from the analysis. We used sequences of Rickettsia rickettsii (KJ588069.1) as outgroup. The genetic distances were calculated in MEGA 6.0. The obtained sequences were deposited in GenBank under the following accession numbers MZ832303–MZ832304.
Results
Specimens Collected
A total of 532 specimens from seven sand fly species were morphologically identified (Table 1). Psathyromyia shannoni and Lutzomyia cruciata were the most abundant species in the municipality of Gómez Farías in the state of Tamaulipas, Mexico, whereas for the state of Nuevo Leon, Psathyromyia texana was the most abundant species in the municipality Cadereyta Jiménez, and Lutzomyia diabolica in the municipally of Santiago. Only a total of 81 sand fly specimens were available for individual analysis by PCR (13 from Nuevo Leon and 68 from Tamaulipas) (Table 1).
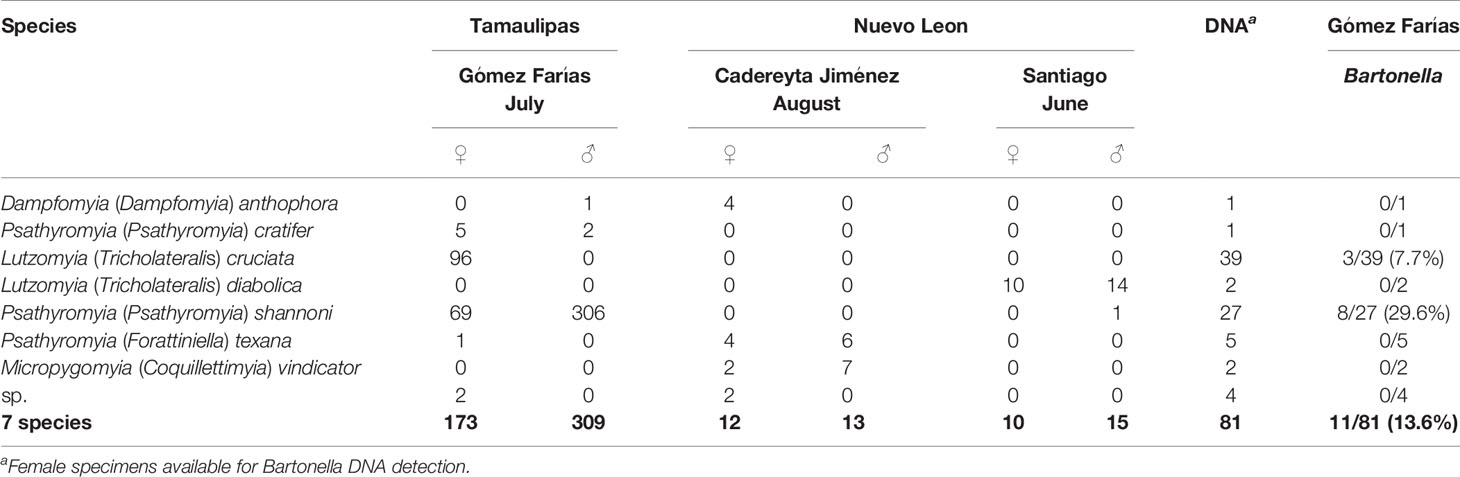
Table 1 Sand fly species collected in two states located at north of Mexico and Bartonella DNA detection.
Bartonella DNA Detection
A total of 11 specimens from Gómez Farías, Tamaulipas, Mexico, collected in July 2010, tested positive for Bartonella DNA using the gltA gene (Table 1). The positive specimens were identified as Pa. shannoni (8/27 prevalence 29.62%) and Lu. cruciata (3/39 prevalence 7.7%). Although the 11 specimens were positive for Bartonella DNA, we only obtained three complete sequences to carry out the phylogenetic analysis (Figure 2).
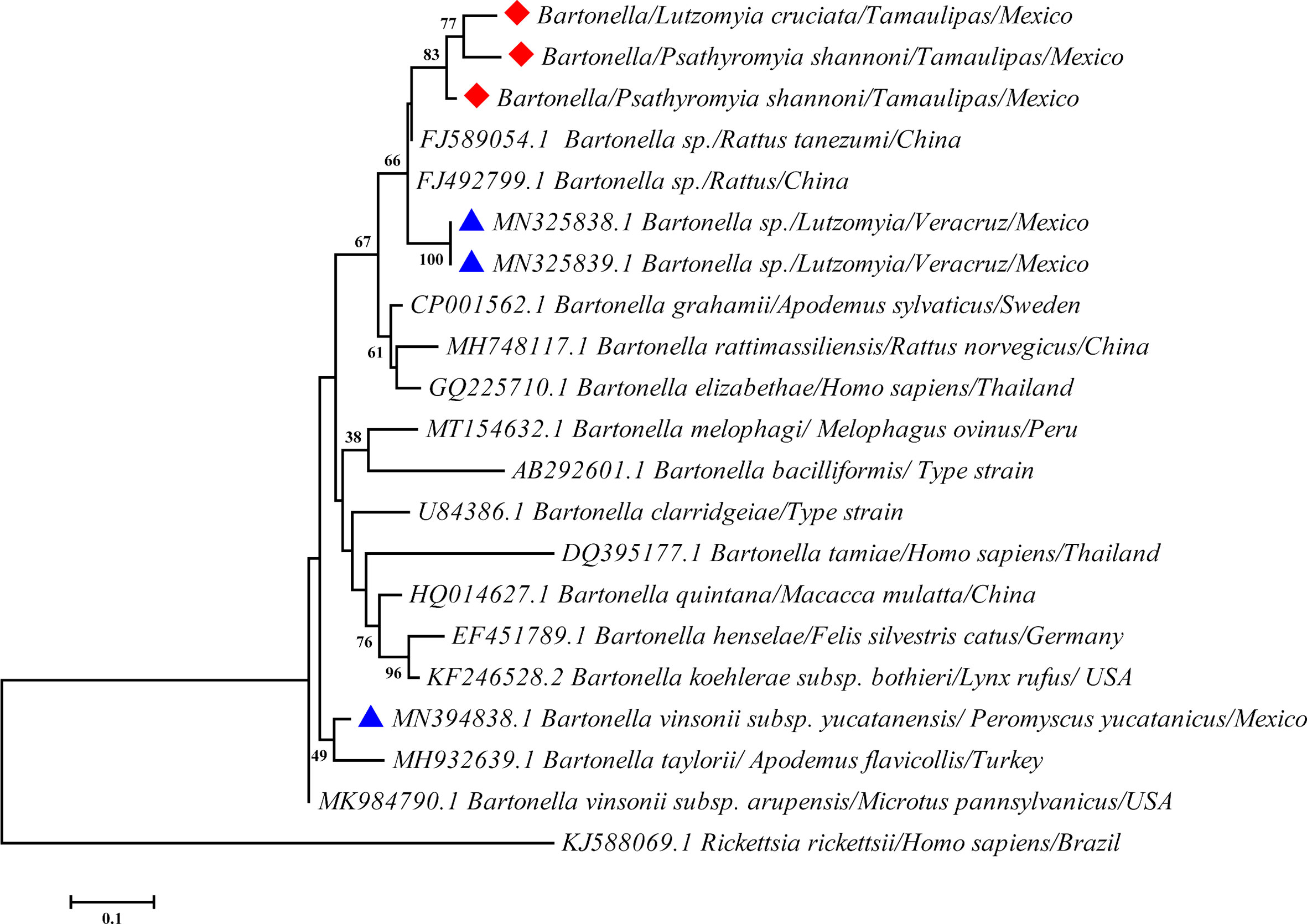
Figure 2 Maximum likelihood analysis for the gltA gene amplified from Bartonella sp. The diamond highlights the sequences generated in this study, and the triangle highlights other Bartonella species present in Mexico.
The gltA sequences obtained in this study were 94% similar between each other and exhibited a 95% similarity with sequences of Bartonella sp. of Rattus tanezumi flavipectus from China (FJ589054.1 and FJ492799.1) and 93% of similarity with Bartonella sp. of Lutzomyia sp. from Veracruz, Mexico (MN325839.1 and MN325838.1). The ML analysis revealed the presence of a putative new lineage of a Bartonella sp., associated with Pa. shannoni and Lu. cruciata, with a bootstrap support value of 83%. According to the genetic distances, the nucleotides showed 89% similarities with the sequences of uncultivated Bartonella from Mexico and 92% with the sequences from China. Based on the proposal that Bartonella species can be classified as new, if they exhibit less than ≥96.0% nucleotide similarity using the gltA gene with other validated species (19), the Bartonella detected in the current study represents a new lineage, although complementary studies are necessary.
This lineage is related to other Bartonella species associated with rodents and humans such as B. grahamii, B. elizabethae, and B. rattimassiliensis (Figure 2). According to the analysis, there are probably three haplotypes (one associated with Lu. cruciata and two with Pa. shannoni) of this new lineage of Bartonella sp.
Discussion
Apart from the Andean valleys, the detection of Bartonella species in sand flies from other parts of the American Continent is not recorded frequently. It could be possible that climate changes may extend the distribution area of sand fly species and thereby also increase the spread of Bartonella (5). A recent study conducted in the state of Veracruz, Mexico, showed that this bacterium was related with sand fly species outside of the known endemic areas (11). Our current work now adds further evidence of this association and also shows the first record of a new strain of Bartonella sp. in the sand fly species Lu. cruciata and Pa. shannoni from Tamaulipas, Mexico.
These sand fly species have anthropophilic habits that are relevant due to their role as vectors of Leishmania spp., and their high abundance and dominance throughout the Gulf and Caribbean of Mexico (12, 13). In this study, the dominance of Pa. shannoni and Lu. cruciata over the other sand fly species was possibly related with the climatic conditions and the type of vegetation of Gómez Farías, Tamaulipas, due to its being a transition zone of the Neotropical and Nearctic biotic provinces. The field work was carried out during the summer, with previous light rains, reporting an average temperature of 25°C and 88% average relative humidity. This characteristic agrees with previous records from Chiapas and Quintana Roo, which showed a positive correlation with the abundance of some sand fly species and these ranges of temperature (<25°C) and humidity (>88%) (20, 21). However, it is advisable to carry out a systematic study in an annual cycle to prove the population dynamic patterns of the phlebotomine sand fly assemblage and thus correlate with the type of vegetation and climatic variables, as well as identify their blood meal preferences to elucidate possible routes of transmission of Bartonella sp. in Gómez Farías, Tamaulipas. Moreover, it is noteworthy that the same sand fly specimens analyzed in the current study had been previously examined for Leishmania DNA showing negative results (13). Due to the possibility of coinfection between both parasites (Bartonella and Leishmania), future studies should take into account the interactions between sand fly species, their pathogens, their hosts, and the environment.
According to sequences obtained, the new strains of an uncultivated Bartonella sp. that we now report are similar to the sequences of Bartonella species detected in sand flies from Veracruz, Mexico (MN325839.1) and rats from China (FJ589054.1) (Figure 2). The identity of the obtained sequences and the phylogenetic analysis suggested that it could be a Bartonella species related to rodents or humans, such as B. grahamii and B. elizabethae. The prevalence of Bartonella sp. in this study was 13.58% (11/81), which could be considered high when compared with other areas (11, 22, 23). Nonetheless, more studies on this bacterium in arthropods and vertebrates are necessary, in order to understand their life cycle and their relationship with sand fly species.
With this report, the number of known sand fly species harboring Bartonella increases to five: Pintomyia verrucarum, Lutzomyia noguchii, Lu. peruensis, Lu. Cruciata, and Pa. shannoni (23). However, at present we do not have sufficient evidence to confirm whether Lu. cruciata or Pa. shannoni are competent vectors for the transmission of this pathogen, since the detection of Bartonella spp. in arthropods using PCR does not represent a confirmation of their vector competence (5). Since Bartonella spp. parasitizes the blood stream or erythrocytes of the infected hosts, their dissemination is facilitated through the blood meal ingestion by several hematophagous arthropods (5, 24, 25), as occurs when sand flies feed on infected blood from a vertebrate host (24).
The record of new sand flies species related to Bartonella sp. is not unexpected, since other studies have suggested that more sand fly species could be potential vectors of this bacterium in endemic and non-endemic areas (5, 22, 24). It is noteworthy that the 16 Bartonella species that have been confirmed as pathogenic were found to be ubiquitously distributed, whereas species adapted to a single host usually lack pathogenicity. It is therefore is likely that the Bartonella species associated with sand flies are not necessarily pathogenic to their host (25).
We now showed that a higher diversity of Bartonella lineages (pathogenic or non-pathogenic) is to be expected in Mexico, than previously reported (4), of which some could be related only with sand fly species. This information is very important, since still missing confirm their possible role as vectors of this bacterium in Mexico, to thereby avoid possible chronic or asymptomatic infections caused by this pathogen (5). To enable future classifications of new lineages of Bartonella species associated with sand flies, the parameters proposed by LaScola et al. (19) are recommended, as our study showed.
Data Availability Statement
The original contributions presented in the study are publicly available. These data can be found here: MZ832303 and MZ832304.
Author Contributions
All authors contributed to the study conception and design. Material preparation, data collection, and analysis were performed by YL-S, NS-O, JR-R, and SS. The first draft of the manuscript was written by YL-S, and all authors commented on previous versions of the manuscript. All authors contributed to the article and approved the submitted version.
Funding
Field work was conducted from grant PAICYT, UANL SA341-10. The sponsorship for this work was supported by grants CONACyT: 6682 and PAPIIT: AG201221.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank Dr. Miriam Berzunza-Cruz for her valuable support in the molecular processing of the samples.
References
1. Akhoundi M, Kuhls K, Cannet A, Votýpka J, Marty P, Delaunay P, et al. Historical Overview of the Classification, Evolution, and Dispersion of Leishmania Parasites and Sandflies. PloS Negl Trop Dis (2016) 10:e0004349. doi: 10.1371/journal.pntd.0004349
2. Serrano C, García E, León V, Rosiles M. Enfermedad De Carrión. Rev Latinoam Patol Clin Med Lab (2014) 61:246–52.
3. Chomel BB, Boulouis H-J, Breitschwerdt EB, Kasten RW, Vayssier-Taussat M, Birtles RJ, et al. Ecological Fitness and Strategies of Adaptation of Bartonella Species to Their Hosts and Vectors. Vet Res (2009) 40:1–22. doi: 10.1051/vetres/2009011
4. Kosoy M, Hayman DTS, Chan KS. Bartonella Bacteria in Nature: Where Does Population Variability End and a Species Start? Infect Genet Evol (2012) 12:894–904. doi: 10.1016/j.meegid.2012.03.005
5. Regier Y, O’Rourke F, Volkhard AJK. Bartonella Spp.- a Chance to Establish One Health Concepts in Veterinary and Human Medicine. Parasit Vectors (2016) 9:1–12. doi: 10.1186/s13071-016-1546-x
6. Sánchez-Montes S, Cabrera-Garrido MY, Ríos-Muñoz CA, Lira-Olguin AZ, Acosta-Gutiérrez R, Mata-Galindo M, et al. Detection of Bartonella and Rickettsia in Small Mammals and Their Ectoparasites in México. THERYA (2019) 10:69–79. doi: 10.12933/therya-19-722
7. Zapata-Valdés C, Avila-Flores R, Gage K, Holmes J, Montenierri J, Kosoy M, et al. Mammalian Hosts, Vectors, and Bacterial Pathogens in Communities With and Without Black-Tailed Prairie Dogs (Cynomys Ludovicianus) in Northwestern Mexico. J Wildl Dis (2018) 54:26–33. doi: 10.7589/2016-09-214
8. Sánchez-Montes S, Guzmán-Cornejo C, Herrera-Montalvo LG, Richman AD, Flores-Martínez JJ, García-Ruiz GF, et al. First Record of Bartonella Vinsonii in the Sucking Louse Hoplopleura Hirsuta Collected From Hispid Cotton Rats, Sigmodon Hispidus, in Mexico. Southwest Entomol (2016) 41:1031–6. doi: 10.3958/059.041.0403
9. López-Pérez AM, Osikowicz L, Bai Y, Montenieri J, Rubio A, Moreno K, et al. Prevalence and Phylogenetic Analysis of Bartonella Species of Wild Carnivores and Their Fleas in Northwestern Mexico. Ecohealth (2017) 14:1–14. doi: 10.1007/s10393-017-1216-2
10. Moskaluk AE, Stuckey MJ, Jaffe DA, Kasten RW, Aguilar-Setién A, Olave-Leyva JI, et al. Molecular Detection of Bartonella Species in Blood-Feeding Bat Flies From Mexico. Vector Borne Zoonotic (2018) 18:258–65. doi: 10.1089/vbz.2017.2213
11. Lozano-Sardaneta YN, Colunga-Salas P, Sánchez-Montes S, Cáceres A, Becker I. First Report of Bartonella Sp. In Sand Flies (Diptera: Psychodidae: Phlebotominae) From Southern Mexico. J Am Mosq Control Assoc (2019) 35:224–7. doi: 10.2987/19-6854.1
12. Lozano-Sardaneta YN, Sánchez-Montes S, Sánchez-Cordero V, Becker I, Paternina L. Molecular Detection of Leishmania Infantum in Sand Flies (Diptera: Psychodidae: Phlebotominae) From Veracruz, Mexico. Acta Trop (2020) 207:1–7. doi: 10.1016/j.actatropica.2020.105492
13. Rodríguez-Rojas J, Rodríguez-Moreno Á, Berzunza-Cruz M, Gutiérrez-Granados G, Becker-Fauser I, Sánchez-Cordero V, et al. Ecology of Phlebotomine Sandflies and Putative Reservoir Hosts of Leishmaniasis in a Border Area in Northeastern Mexico: Implications for the Risk of Transmission of Leishmania Mexicana in Mexico and the USA. Parasite (2017) 24:1–17. doi: 10.1051/parasite/2017034
14. Galati E. Morfologia E Terminologia De Phlebotominae (Diptera: Psychodidae). In: Classificação E Identificação De Táxons Das Américas. Vol I Apostila Da Disciplina Bioecologia E Identificação De Phlebotominae do Programa De Pós-Graduação Em Saúde Pública. São Paulo: Faculdade de Saúde Pública da Universidade de São Paulo (2019). p. 133p.
15. Marcondes B. A Proposal of Generic and Subgeneric Abbreviations for Phlebotomine Sandflies (Diptera: Psychodidae: Phlebotominae) of the World. Entomol News (2007) 118:351–6. doi: 10.3157/0013-872X(2007)118[351:APOGAS]2.0.CO;2
16. Kato H, Uezato H, Katakura K, Calvopiña M, Marco JD, Barroso PA, et al. Detection and Identification of Leishmania Species Within Naturally Infected Sand Flies in the Andean Areas of Ecuador by a Polymerase Chain Reaction. Am J Trop Med Hyg (2005) 72:87–93. doi: 10.4269/ajtmh.2005.72.87
17. Norman F, Regnery R, Jameson P, Greene C, Krause DC. Differentiation of Bartonella-Like Isolates at the Species Level by PCR-Restriction Fragment Length Polymorphism in the Citrate Synthase Gene. J Clin Microbiol (1995) 33:1797–803. doi: 10.1128/jcm.33.7.1797-1803.1995
18. Rubio AV, Ávila-Flores R, Osikowicz LM, Bai Y, Suzán G, Kosoy MY. Prevalence and Genetic Diversity of Bartonella Strains in Rodents From Northwestern Mexico. Vector-Borne Zoonotic Dis (2014) 14:838–45. doi: 10.1089/vbz.2014.1673
19. La Scola B, Zeaiter Z, Khamis A, Raoult D. Gene-Sequence-Based Criteria for Species Definition in Bacteriology: The Bartonella Paradigm. TRENS Microbiol (2003) 11:318–21. doi: 10.1016/S0966-842X(03)00143-4
20. Pérez J, Virgen A, Rojas J, Rebollar-Téllez E, Castillo A, Infante F, et al. Species Composition and Seasonal Abundance of Sandflies (Diptera: Psychodidae: Phlebotominae) in Coffee Agroecosystems. Mem Inst Oswaldo Cruz (2014) 109:80–6. doi: 10.1590/0074-0276130224
21. Rebollar-Téllez E, Reyes-Villanueva F, Fernandez-Salas I, Andrade-Narvaez F. Population Dynamics and Biting Rhythm of the Anthropophilic Sandfly Lutzomyia Cruciata (Diptera: Psychodidae) in Southeast, Mexico. Rev Inst Med Trop Sao Paulo (1996) 38:29–33. doi: 10.1590/S0036-46651996000100006
22. Ellis BA, Rotz LD, Leake JAD, Samalvides F, Bernable J, Ventura G, et al. An Outbreak of Acute Bartonellosis (Oroya Fever) in the Urubamba Region of Peru, 1998. Am J Trop Med Hyg (1999) 61:344–9. doi: 10.4269/ajtmh.1999.61.344
23. Villaseca P, Padilla C, Ventura G, Samalvides F, Yañez H, Chevarría L, et al. Importancia De La Lutzomyia Peruensis En La Transmisión De La Enfermedad De Carrión En El Valle Sagrado De Los Incas. Urubamba-Cusco, Perú. Rev Peru Med Exp Salud Publica (1999) 15:28–30.
24. Billeter SA, Levy MG, Chomel BB, Breitschwerd TE. Vector Transmission of Bartonella Species With Emphasis on the Potential for Tick Transmission. Med Vet Entomol (2008) 22:1–15. doi: 10.1111/j.1365-2915.2008.00713.x
Keywords: Bartonella sp., sand flies, Psathyromyia shannoni, Lutzomyia cruciata, Mexico
Citation: Lozano-Sardaneta YN, Soto-Olguín NJ, Rodríguez-Rojas JJ, Sánchez-Montes S, Rebollar-Téllez EA and Becker I (2021) Molecular Detection of Bartonella sp. in Psathyromyia shannoni and Lutzomyia cruciata From Northeastern Mexico. Front. Trop. Dis 2:780808. doi: 10.3389/fitd.2021.780808
Received: 21 September 2021; Accepted: 25 October 2021;
Published: 18 November 2021.
Edited by:
Yara M. Traub-Csekö, Oswaldo Cruz Foundation (Fiocruz), BrazilReviewed by:
Shih Keng Loong, University of Malaya, MalaysiaZubaidah Ya’cob, University of Malaya, Malaysia
Copyright © 2021 Lozano-Sardaneta, Soto-Olguín, Rodríguez-Rojas, Sánchez-Montes, Rebollar-Téllez and Becker. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ingeborg Becker, YmVja2VyQHVuYW0ubXg=; Eduardo A. Rebollar-Téllez, ZWR1YXJkby5yZWJvbGxhcnRsQHVhbmwuZWR1Lm14