 Ronghao Zhu
Ronghao Zhu Jing Huang
Jing Huang Fenhong Qian*
Fenhong Qian*
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Immunol. , 26 February 2025
Sec. Cancer Immunity and Immunotherapy
Volume 16 - 2025 | https://doi.org/10.3389/fimmu.2025.1556209
This article is part of the Research Topic Harnessing Molecular Insights for Enhanced Drug Sensitivity and Immunotherapy in Cancer View all 25 articles
Lung cancer remains a leading cause of cancer-related deaths worldwide, necessitating innovative treatments. Tumor-associated macrophages (TAMs) are primary immunosuppressive effectors that foster tumor proliferation, angiogenesis, metastasis, and resistance to therapy. They are broadly categorized into proinflammatory M1 and tumor-promoting M2 phenotypes, with elevated M2 infiltration correlating with poor prognosis. Strategies aimed at inhibiting TAM recruitment, depleting TAMs, or reprogramming M2 to M1 are therefore highly promising. Key signaling pathways, such as CSF-1/CSF-1R, IL-4/IL-13–STAT6, TLRs, and CD47-SIRPα, regulate TAM polarization. Additionally, macrophage-based drug delivery systems permit targeted agent transport to hypoxic regions, enhancing therapy. Preclinical studies combining TAM-targeted therapies with chemotherapy or immune checkpoint inhibitors have yielded improved responses and prolonged survival. Several clinical trials have also reported benefits in previously unresponsive patients. Future work should clarify the roles of macrophage-derived exosomes, cytokines, and additional mediators in shaping the immunosuppressive tumor microenvironment. These insights will inform the design of next-generation drug carriers and optimize combination immunotherapies within precision medicine frameworks. Elucidating TAM phenotypes and their regulatory molecules remains central to developing novel strategies that curb tumor progression and ultimately improve outcomes in lung cancer. Importantly, macrophage-based immunomodulation may offer expanded treatment avenues.
Lung cancer remains the foremost cause of cancer-related mortality globally, responsible for 20.4% of such deaths in the U.S. in 2024, with daily fatalities nearly double those from colorectal cancer, and surpassing combined deaths from breast, prostate, and pancreatic cancers (1–3). The mortality from lung cancer is expected to greatly exceed that of gastric and breast cancers in the coming decades (4, 5). The tumor immune microenvironment, particularly macrophages differentiated into TAMs, critically influences lung cancer progression by enhancing angiogenesis and metastasis (6–11). TAMs correlate strongly with tumor size, differentiation, invasion depth, lymph node metastasis, and TNM stage in lung cancer (12), highlighting their central role in disease dynamics. Besides, in the tumor microenvironment (TME) of cancer, TAMs express characteristic surface markers such as CD68/CD163/CD204/CD206 (13), and their polarization significantly influences tumor progression and therapeutic resistance (14–18). M2-like TAMs, in particular, are associated with a pro-tumoral environment, marked by the secretion of immunosuppressive cytokines like IL-10 and TGF-β, which not only enhance tumor growth but also hinder the immune response (19–21). These macrophages also facilitate PD-L1 expression, diminishing the effectiveness of existing immunotherapies. Strategies targeting TAMs, alongside chemotherapy and PD-1/PD-L1 inhibitors, show promising therapeutic potential in preclinical models (22–24). This review discusses TAM classification, their influence on lung cancer progression, and their therapeutic implications.
TAMs exhibit significant plasticity, adapting to various TME conditions and polarizing into two major phenotypes: the proinflammatory M1 and the tumor-promoting M2 macrophages (25–27). M1 macrophages are typically activated by inflammatory cytokines (e.g., GM-CSF, TNF-α, IFN-γ, LPS) and are characterized by their secretion of proinflammatory cytokines (e.g., IL-1α/β, IL-6, IL-12, IL-23), which enhance cytotoxic immune responses and anti-tumor immunity (28). In contrast, M2 macrophages, which predominantly promote tumor progression, are divided into subtypes based on their cytokine responses and functional roles. These subtypes include M2a, M2b, M2c, and M2d, each associated with different aspects of tumor support. M2a macrophages, driven by IL-4 and IL-13, are primarily involved in tissue repair, promoting fibrosis and immune regulation through the secretion of TGF-β and insulin-like growth factors (29). M2b macrophages, activated by immune complexes and LPS, are characterized by their anti-inflammatory profile, releasing IL-10 and IL-6, which dampen anti-tumor immunity (30). M2c macrophages, induced by IL-10, TGF-β, and glucocorticoids, play a key role in immunosuppression and extracellular matrix remodeling, contributing to the tumor’s immune escape mechanisms (31). M2d macrophages, activated by adenosine and TLRs, secrete IL-10, TGF-β, and VEGF, promoting angiogenesis and immunosuppression, which enhances tumor progression and metastasis (32, 33). The M2a/M2c macrophages facilitate the invasion of lung cancer cells and contribute to tumor progression (34). In A549 cells, the presence of M2c macrophages induced epithelial-mesenchymal transition (EMT), which was characterized by increased expression of vimentin, fibronectin, E-cadherin, NF-kB, and CCL-17 (35). Targeting TAM polarization toward the M1 phenotype, while eliminating the M2a and particularly M2c subtypes, represents a promising approach for effective anti-cancer strategies (36).
The organism utilizes innate and adaptive immune mechanisms to counteract tumor initiation and progression; however, tumor cells can evade these defenses through multiple strategies (37–39). In the TME, TAMs transition from an antitumor to a pro-tumor role, chiefly impeding other immune cells’ immunomodulatory functions. This inhibition occurs via several pathways: (I) M2-type TAMs, influenced by Th2 cells within the pulmonary TME, produce immunosuppressive factors like IL-10 and TGF-β. IL-10 particularly enhances PD-L1 expression in macrophages, diminishing cytotoxic T cell activity and fostering immune tolerance (40); (II) TAMs secrete C-C motif ligand 22 (CCL-22), attracting regulatory T cells (Tregs) to the TME and dampening effector T cell function, thereby cultivating an immunosuppressive environment (41); (III) Hypoxia in the TME upregulates the expression of HIF-1α in macrophages, resulting in decreased CD8+ T-cell activation mediated by macrophages and promoting immune evasion (42–44). While under hypoxic conditions, TAMs produce increased levels of arginase I, which depletes L-arginine in the microenvironment, thereby inhibiting T cells by arresting them in the G0/G1 phase of the cell cycle and preventing their proliferation (45); (IV) CD206 mannose receptors on the surface of TAMs suppress CD45 phosphatase activity, leading to upregulated expression of cytotoxic T-lymphocyte–associated protein 4 (CTLA-4) and ultimately inducing T-cell tolerance (46); (V) IL-4 stimulation activates the PI3Kγ–mTor–S6Ka–C/EBPβ pathway in macrophages and inhibits nuclear factor kappa-B (NF-κB), thereby suppressing immunity and promoting tumor growth (47). In addition, TAMs interact with various immune cells, including T lymphocytes, NK cells, dendritic cells, neutrophils, and MDSCs. Moreover, TAMs suppress the cytotoxic activity of T cells, natural killer T (NKT) cells, and NK cells by expressing ligands for immune checkpoint receptors such as PD-1 and CTLA-4 (48–51).
TAMs secrete proangiogenic factors (e.g., VEGF, CXCL8) into avascular regions, promoting tumor angiogenesis. VEGF strongly stimulates endothelial cell proliferation, neovascularization, and vascular permeability, facilitating tumor cell extravasation (52). TAM-derived IL-8, VEGF, and urokinase-type plasminogen activator (uPA) further contribute to neovascularization (53). Hypoxia in the TME induces HIF-1/2, which upregulate VEGF, platelet-derived growth factor (PDGF), and EGF (54, 55). VEGF also attracts more TAMs, generating a positive feedback loop (56). Additionally, TAMs secrete TGF-β, TNF-α, MMPs, and TIE2, all of which promote intratumoral vascular formation (57–60).
In lung cancer, TAMs foster tumor proliferation, invasion, and metastasis via chemokines and cytokines—TGF-β, IL-10, IL-6, matrix metalloproteinases (MMPs), and VEGF—and may activate cancer stem cells (CSCs) through IL-10 (61). Hypoxia from rapid tumor growth increases M2 macrophages, elevating IL-10, VEGF, and HIF-1α, promoting metastasis and further macrophage infiltration (62). TAMs also facilitate EMT, critical for metastasis (63) by upregulating CRYAB (64) and Ezrin phosphorylation–mediated FUT4/LeY fucosylation (65). TAM-derived IL-6, IL-10, and TGF-β regulate EMT (66) with IL-6 and IL-10 inducing M2 polarization through JAK/STAT3 (67), and TGF-β promoting SOX9 expression via c-jun/SMAD3, enhancing proliferation and invasiveness (68, 69). Furthermore, TAMs secrete MMPs, mainly MMP-9 and MMP-2, to degrade the extracellular matrix and facilitate invasion and metastasis (70). MMP-9 expression is also associated with lymph node metastasis and prognosis (71). Chemokines like CCL18 from TAMs fuel tumor progression (72). In addition, TAMs promote collagen fiber formation, guiding lung cancer cells toward blood vessels and into circulation (73). CSCs promote lung cancer progression as well (61). TAMs interact with CSCs, including TAM recruitment through vascularization, the release of chemokines by TAMs to maintain CSC dormancy, and antigen modification of CSCs to evade immune recognition, which plays a crucial role in tumor progression and metastasis. Besides, CSCs promote TAM polarization from M1 to M2, induce angiogenesis via VEGF, and create supportive niches through tissue repair pathways (74). A potential therapeutic strategy targeting the interactions between TAMs and CSCs could provide an effective approach for lung cancer (75, 76). Exosomes also participate in tumorigenesis and metastasis by mediating material exchange among tumor cells and macrophages (77–79). For instance, exosomes derived from tumors release miR-19b-3p, which promotes the polarization of M2 macrophages, and also secrete LINC00273, facilitating the metastasis of lung adenocarcinoma through the Hippo signaling pathway (80). MiR-501-3p expression was elevated, while WDR82 levels were reduced in lung cancer tissues and cell lines. Additionally, M2-derived exosomes contributed to the further increase in miR-501-3p levels. These exosomes, along with their cargo of miR-501-3p, played a significant role in promoting lung cancer cell proliferation. Exosomal miR-501-3p has been shown to inhibit apoptotic processes, thus further enhancing lung cancer proliferation, invasion, and metastasis (81). Besides, exosomes secreted by M2 TAMs contribute to osimertinib resistance in non-small cell lung cancer by modulating the MSTRG.292666.16-miR-6836-5p-MAPK8IP3 signaling pathway (82).
TAMs facilitate tumor growth, progression, and chemoresistance by supplying cytokines and upregulating anti-apoptotic genes. Treatment of murine models with CTX, PTX, or DOX increases CD206+ TAMs, triggering revascularization and recurrence (83). Chemotherapy-induced IL-34 augments TAM-mediated drug resistance (19). Furthermore, TAM-derived extracellular matrix remodels tumor–macrophage interactions, enhancing chemoradiotherapy resistance (84, 85). M2-polarized TAMs secrete growth factors and suppress cell death signaling, conferring chemoresistance and radioprotection, ultimately leading to poor prognosis (86). Cisplatin-resistant lung cancer cells exhibit enhanced self-renewal and metastatic capacity by secreting macrophage inhibitory factor (MIF), which promotes the M2 polarization of TAMs (87). Near-infrared fluorescence (NIRF) imaging and single-photon emission computed tomography (SPECT) imaging reveal M2 TAM infiltration in recurrent tumors and lymph node metastases, underscoring their role in tumor recurrence and pulmonary spread post-chemotherapy (88). P2X7 activation of STAT6/IRF4 drives M2 polarization, fueling tumor proliferation, angiogenesis, and T-cell suppression. Inhibiting or deleting P2X7 weakens M2 TAMs, restricting lung tumor growth while overcoming immunotherapy and chemotherapy resistance (89). Besides, TAMs activated CSC-related pathways might enhance drug resistance (90).
GM-CSF governs hematopoietic cell generation and differentiation, as well as angiogenesis. CSF-1 binds CSF-1R, triggering the PI3K signaling cascade and further activating protein kinase B (AKT) and mammalian target of rapamycin complex 2 (mTORC2), thereby regulating the M1/M2 polarization axis of macrophages (91). PI3K and AKT overexpression suppresses M1 activation partly by negatively regulating the NF-κB signaling pathway, which otherwise promotes M1 phenotypes (92). CSF-1R also binds IL-34; elevated IL-34 and CSF-1R levels correlate with tumor progression and poorer survival (93). Moreover, CSF-1 can recruit and reprogram TAMs to secrete factors that facilitate tumor growth and metastasis (94).
Inflammatory factors are pivotal in inflammatory diseases progression and significantly influence the efficacy of therapies (95–98). IL-4 and IL-13 participate in Th2-type immune responses (99) and drive TAMs toward an M2 phenotype that promotes aberrant angiogenesis and tumor progression. Both cytokines bind type I or II IL-4 receptors, activating JAKs and subsequently phosphorylating STAT6, which forms dimers, translocates to the nucleus, and upregulates M2-associated genes such as Arg-1, mannose receptor c-type 1 (Mrc-1), Chil3/Ym1, and resistin-like molecule alpha (RELM-α or Fizz-1) (100, 101). STAT6 function is also regulated by additional factors: one study found that TRAF3 enhances STAT6 ubiquitination (K450), thereby boosting transcriptional activity and M2 marker expression (102). In a B16 melanoma model, TRAF3 deficiency in bone marrow suppressed tumor growth and metastasis, underscoring STAT6’s importance in macrophage-mediated immunosuppression.
TLR are crucial in recognizing pathogen-associated molecular patterns (PAMPs) and damage-associated molecular patterns (DAMPs), playing a significant role in activating the immune response (103). In the context of cancer, TLR signaling can influence TAM polarization, promoting tumor progression (104). TLRs, particularly TLR2, TLR4, and TLR7, have been shown to modulate the immune microenvironment by skewing macrophages toward a pro-tumoral M2 phenotype, thus enhancing immune evasion, angiogenesis, and metastasis (105, 106). Another study showed that combining low-toxicity IFN (IFN-γ, IFN-β) with TLR agonists increased M1 induction by up to 100-fold, suggesting novel TLR-targeted strategies for tumor immunotherapy (107). For instance, clinical trials indicated that the intravenous administration of TLR agonists poly(I:C) + R848 reduced the growth of orthotopic lung tumors through demonstrated reprogramming of interstitial macrophages increasing the M1/M2 ratio (104).
CD47 (integrin-associated protein) is a thrombospondin-family receptor that modulates platelet activation, migration, adhesion, and phagocytosis. Overexpressed on tumor cells, CD47 binds SIRPα on macrophages, leading to phosphorylation of immunoreceptor tyrosine-based inhibitory motifs (ITIMs) and inhibiting phagocytosis (108, 109). High CD47 expression is associated with worse prognoses in various solid tumors. In non-small cell lung cancer (NSCLC), approximately two-thirds of tumor samples exhibit CD47 overexpression, correlating with increased SIRPα on TAMs (110). Although CD68+ TAMs typically suggest better outcomes, high SIRPα levels on TAMs reduce FOXP3+ TIL proportions and TIL scores, leading to poor prognosis. In 191 resected NSCLC specimens, CD47 was more frequently overexpressed in females, never-smokers, and lung adenocarcinoma patients, significantly correlating with EGFR mutations but inversely with PD-L1 levels and tumor mutation burden (111). Gefitinib-resistant lung cancer cells upregulate CD47, impairing macrophage phagocytosis. Inhibition of CD47-SIRPα signaling via STAT3 inhibitors enhances TAM phagocytic activity and mitigates EGFR-TKI resistance, offering a novel treatment strategy for patients with acquired EGFR-TKI resistance (112) (Figure 1).
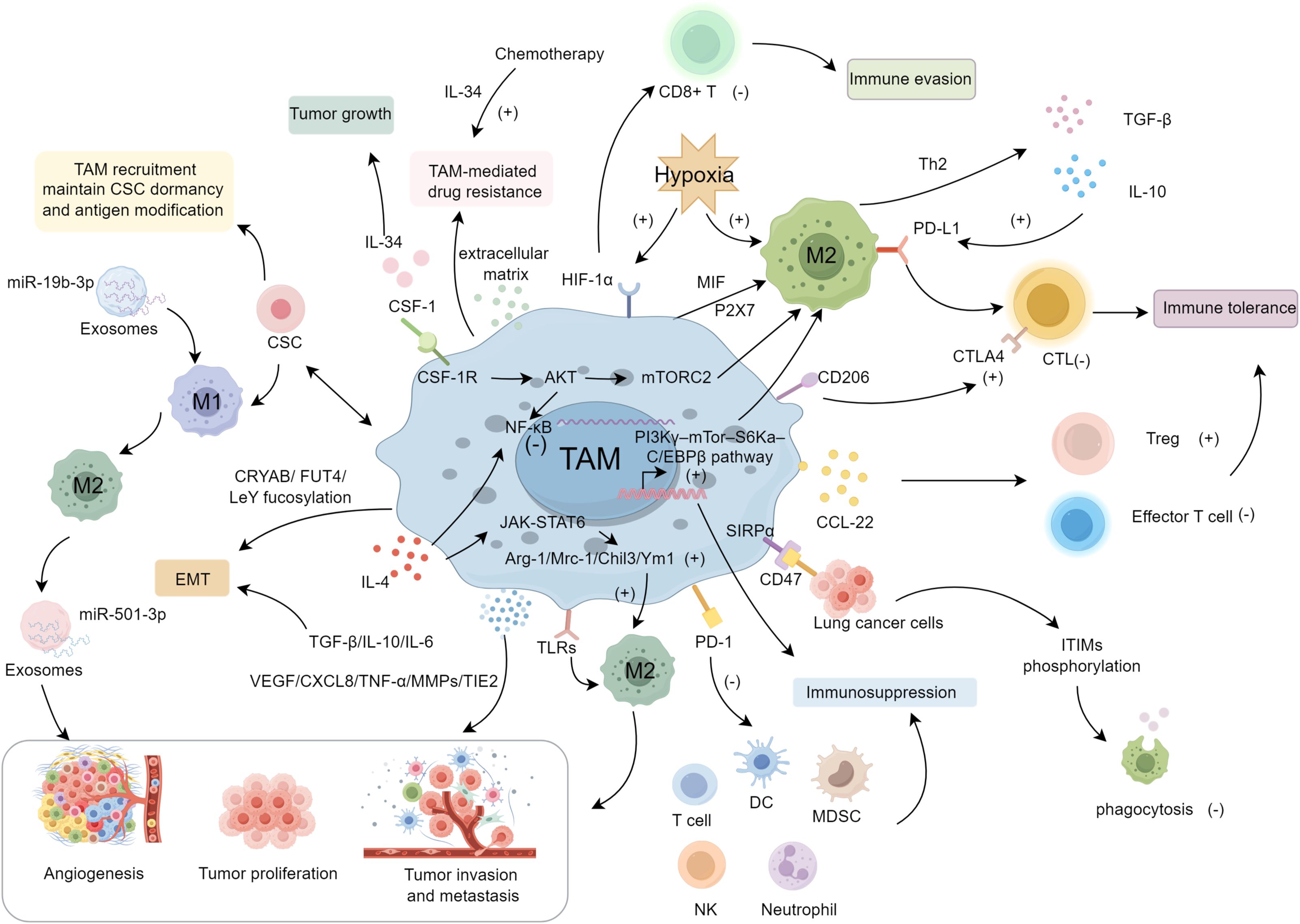
Figure 1. The mechanisms and regulating molecules of TAMs in the progression of lung cancer.
TAMs play a critical role in the formation of the TME, tumor immunity, and responses to immunotherapy, making them important and promising therapeutic targets in lung cancer. Notably, combining therapies targeting TAMs with other immunotherapeutic approaches can yield superior efficacy (113, 114). Below, we introduce four strategies for targeting TAMs.
Limiting TAM infiltration requires restricting the recruitment of circulating monocytes via factors like chemokines (e.g., CCL2), cytokines (e.g., CSF-1), and complement mediators. In particular, the CCL2–CCR2 and CSF-1/CSF-1R axes have garnered significant attention. CCL2–CCR2 signaling recruits TAMs, promoting tumor angiogenesis and growth (115). In a Lewis lung cancer model, CCR2 knockout or the CCR2 inhibitor RS504393 reduced TAM recruitment, shifted TAMs to an M1 phenotype, and suppressed angiogenesis and tumor progression (22). Another approach to prevent TAM infiltration into the tumor microenvironment involves inhibiting the CSF-1–CSF-1R axis, which is vital for TAM differentiation, survival, and recruitment (6). CSF-1R blockade can reduce the infiltration of TAMs mediated by immunosuppressive T cells (116). In animal models, it has been demonstrated that small-molecule CSF-1R inhibitors in combination with immune checkpoint inhibitors are currently in Phase I/II clinical trials for the treatment of advanced breast cancer and other solid tumors (117). This finding provides a novel perspective for targeting and inhibiting TAM recruitment in lung cancer.
High TAM infiltration in lung cancer correlates with poor prognosis, though single-agent depletion strategies have shown limited efficacy (118–120). Specifically targeting M2-like TAMs is preferable, as broad macrophage eradication is suboptimal. For instance, selectively depleting M2 TAMs improved survival in tumor-bearing mice (121). The CSF-1/CSF-1R axis remains the most explored depletion method. Blocking CSF-1R depletes macrophages in both normal and tumor tissues, as seen with BLZ945 and PLX3397 (122). Moreover, combining CSF-1R inhibition with other treatments yields better outcomes, enhancing CD8+ T-cell infiltration and slowing tumor progression, especially when coupled with anti-PD-1 therapy (114, 123). Targeting surface molecules like CD52, scavenger receptor A, folate receptor β, and CD206 can also deplete TAMs (124, 125).
Elevated M2 TAM density correlates with poorer survival, whereas higher M1 density predicts better outcomes (126, 127). Reprogramming M2 TAMs to M1 slows tumor progression (128). TLR7/8 agonist R848, delivered via β-cyclodextrin nanoparticles, can repolarize M2 TAMs, while combining R848 with anti-PD-1 further enhances therapeutic efficacy (129). Besides depleting macrophages, CSF-1/CSF-1R inhibitors can modulate TAM phenotypes (130, 131). Ginsenoside Rh2 also shifts M2 to M1 macrophages, reducing NSCLC metastasis by downregulating VEGF, MMP2, and MMP9 (132). Nanoformulations that load macrophage-targeted agents (e.g., TLR7/8 agonists) can overcome delivery challenges in inaccessible tumors (133, 134). Inhibiting TAM receptors, namely Tyro3, Axl, and MERTK, further counters M2-like polarization (135). Axl and MERTK are overexpressed in lung cancer (136, 137), and MERTK inhibitors (e.g., UNC2025) reduce distant metastases in NSCLC models (138). Additionally, imatinib suppresses M2 polarization by inhibiting STAT6 phosphorylation, preventing metastasis (139, 140). Blocking M2-activating cytokines (IL-13, IL-4, IL-10) also enhances efficacy, including immune checkpoint inhibitors (84), as high IL-10 in TAMs is linked to NSCLC staging (141). Cancer cells release succinate into the extracellular space, promoting macrophage migration and influencing TAM polarization. This suggests that succinate functions as an oncometabolite, potentially serving as a critical target for cancer chemoprevention and therapeutic strategies (142).
Although biocompatible nanomaterials have improved drug delivery, their limited circulation half-life remains a hurdle. Macrophages, with longer half-lives, immune functions, and intrinsic tumor-homing capabilities, offer a promising alternative (143–145). In animal models, doxorubicin-loaded M1 macrophages prolonged survival and inhibited tumor invasion (146). However, directly loading macrophages with anti-cancer drugs may impair their function. To circumvent this, “indirect” loading onto biocompatible nanomaterials allows higher drug capacity while preserving macrophage viability (147, 148). Attaching microparticles to macrophage surfaces, rather than internalization, can further maintain targeting efficiency (149). Photothermal therapy, which uses light to generate cytotoxic heat, is another emerging strategy. Nanomaterials with high photothermal conversion efficiency can be immobilized on macrophage membranes to enhance biocompatibility, immune evasion, and tumor-homing (150). For instance, doxorubicin-loaded graphene oxide on mouse macrophages markedly suppresses tumor growth (151) (Table 1).
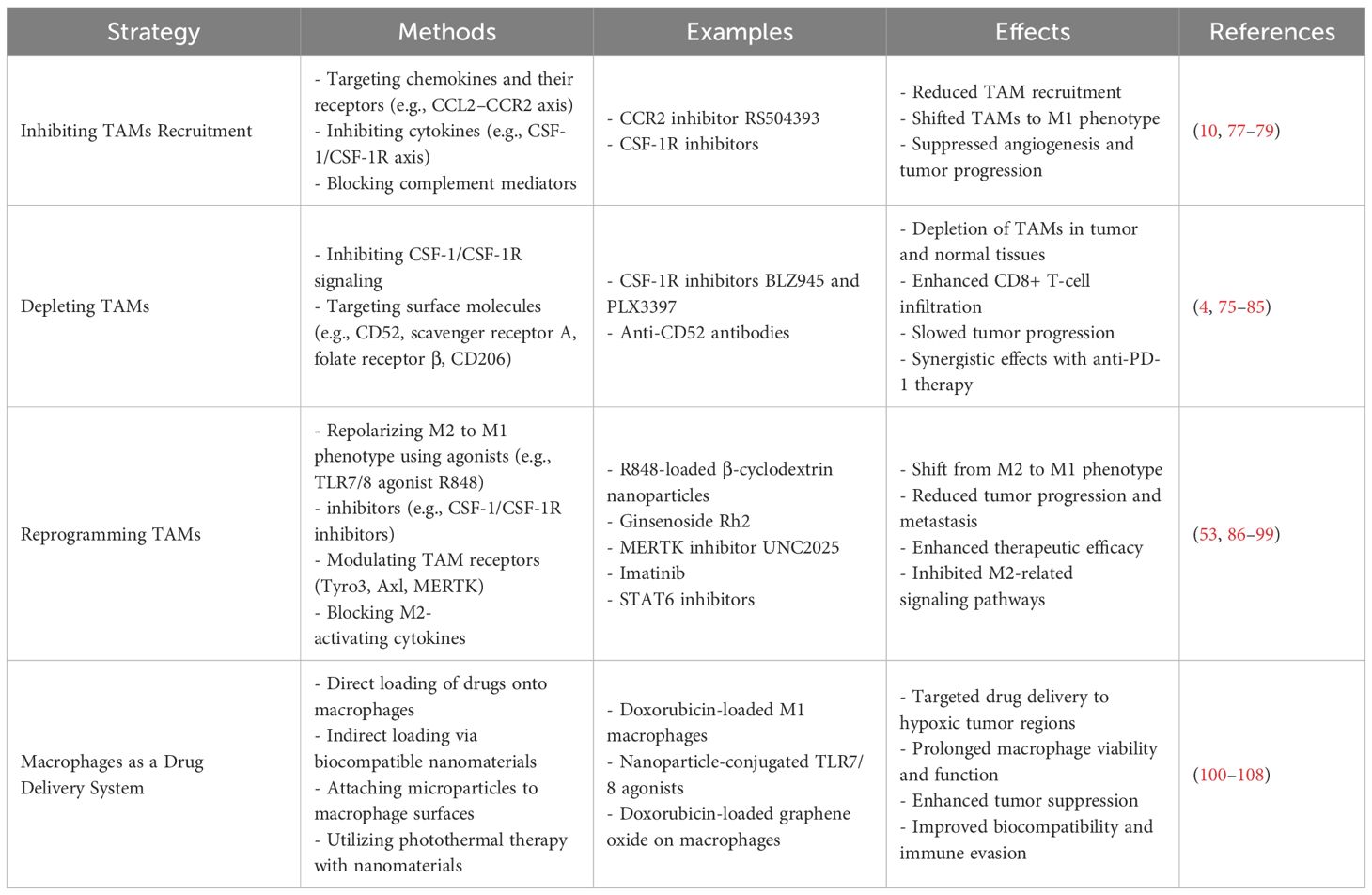
Table 1. Therapeutic strategies targeting TAMs in lung cancer.
Lung cancer remains a leading global malignancy, with persistently low five-year survival despite immunotherapy advances. Increasing evidence identifies TAMs as key immunosuppressive TME mediators driving tumor cell proliferation, angiogenesis, metastasis, immune evasion, and drug resistance. Their polarization state and infiltration levels strongly correlate with prognosis; hence, inhibiting TAM aggregation, depleting TAMs, or reprogramming M2 to M1 is critical for improving outcomes. Clinical trials show that combining TAM-targeting agents with existing therapies enhances efficacy, benefiting previously unresponsive patients. Further research is needed to elucidate macrophage-derived cytokines, exosomes, and other factors within the TME, potentially expanding therapeutic targets and advancing macrophage-based drug delivery. Clarifying TAM phenotypes and their molecular pathways is essential for understanding lung cancer progression, informing novel immunotherapeutic strategies, and providing key theoretical underpinnings for developing next-generation antitumor drugs.
RZ: Writing – original draft. JH: Writing – original draft. FQ: Writing – original draft, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. The present study was supported by a grant from the National Natural Science Foundation of China (No. 81370119). The present study was supported by China International Medical Foundation (Z-2014-08-2209). The present study was supported by Zhenjiang Science and Technology Innovation Fund (SH2023082).
We thank FQ for her guidance and revision of this review. The authors would like to thank all the reviewers who participated in the review.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Siegel RL, Giaquinto AN, Jemal A. Cancer statistics, 2024. CA Cancer J Clin. (2024) 74:12–49. doi: 10.3322/caac.21820
2. Wang Y, Ma L, He J, Gu H, Zhu H. Identification of cancer stem cell-related genes through single cells and machine learning for predicting prostate cancer prognosis and immunotherapy. Front Immunol. (2024) 15:1464698. doi: 10.3389/fimmu.2024.1464698
3. Wang Y, He J, Zhao Q, Bo J, Zhou Y, Sun H, et al. Evaluating the predictive value of angiogenesis-related genes for prognosis and immunotherapy response in prostate adenocarcinoma using machine learning and experimental approaches. Front Immunol. (2024) 15:1416914. doi: 10.3389/fimmu.2024.1416914
4. Li L, Shao M, He X, Ren S, Tian T. Risk of lung cancer due to external environmental factor and epidemiological data analysis. Math Biosci Eng. (2021) 18:6079–94. doi: 10.3934/mbe.2021304
5. Kanwal M, Ding XJ, Cao Y. Familial risk for lung cancer. Oncol Lett. (2017) 13:535–42. doi: 10.3892/ol.2016.5518
6. Mantovani A, Marchesi F, Malesci A, Laghi L, Allavena P. Tumour-associated macrophages as treatment targets in oncology. Nat Rev Clin Oncol. (2017) 14:399–416. doi: 10.1038/nrclinonc.2016.217
7. Li C, Liu T, Liu Y, Zhang J, Zuo D. Prognostic value of tumour microenvironment-related genes by TCGA database in rectal cancer. J Cell Mol Med. (2021) 25:5811–22. doi: 10.1111/jcmm.v25.12
8. Li C, Wirth U, Schardey J, Ehrlich-Treuenstätt VV, Bazhin AV, Werner J, et al. An immune-related gene prognostic index for predicting prognosis in patients with colorectal cancer. Front Immunol. (2023) 14:1156488. doi: 10.3389/fimmu.2023.1156488
9. Zhang C, Sun D, Li C, Liu Y, Zhou Y, Zhang J. Development of cancer-associated fibroblasts subtype and prognostic model in gastric cancer and the landscape of tumor microenvironment. Int J Biochem Cell Biol. (2022) 152:106309. doi: 10.1016/j.biocel.2022.106309
10. Wang Y, Wang J, He J, Ji B, Pang Z, Wang J, et al. Comprehensive analysis of PRPF19 immune infiltrates, DNA methylation, senescence-associated secretory phenotype and ceRNA network in bladder cancer. Front Immunol. (2023) 14:1289198. doi: 10.3389/fimmu.2023.1289198
11. Wang Q, Liu Y, Li Z, Tang Y, Long W, Xin H, et al. Establishment of a novel lysosomal signature for the diagnosis of gastric cancer with in-vitro and in-situ validation. Front Immunol. (2023) 14:1182277. doi: 10.3389/fimmu.2023.1182277
12. Sarode P, Zheng X, Giotopoulou GA, Weigert A, Kuenne C, Gunther S, et al. Reprogramming of tumor-associated macrophages by targeting beta-catenin/FOSL2/ARID5A signaling: A potential treatment of lung cancer. Sci Adv. (2020) 6:eaaz6105. doi: 10.1126/sciadv.aaz6105
13. Larionova I, Tuguzbaeva G, Ponomaryova A, Stakheyeva M, Cherdyntseva N, Pavlov V, et al. Tumor-associated macrophages in human breast, colorectal, lung, ovarian and prostate cancers. Front Oncol. (2020) 10:566511. doi: 10.3389/fonc.2020.566511
14. Li Z, Maeda D, Yoshida M, Umakoshi M, Nanjo H, Shiraishi K, et al. The intratumoral distribution influences the prognostic impact of CD68- and CD204-positive macrophages in non-small cell lung cancer. Lung Cancer. (2018) 123:127–35. doi: 10.1016/j.lungcan.2018.07.015
15. Xiao F, Liu N, Ma X, Qin J, Liu Y, Wang X. M2 macrophages reduce the effect of gefitinib by activating AKT/mTOR in gefitinib-resistant cell lines HCC827/GR. Thorac Cancer. (2020) 11:3289–98. doi: 10.1111/1759-7714.13670
16. Dalton HJ, Pradeep S, McGuire M, Hailemichael Y, Ma S, Lyons Y, et al. Macrophages facilitate resistance to anti-VEGF therapy by altered VEGFR expression. Clin Cancer Res. (2017) 23:7034–46. doi: 10.1158/1078-0432.CCR-17-0647
17. Arlauckas SP, Garris CS, Kohler RH, Kitaoka M, Cuccarese MF, Yang KS, et al. In vivo imaging reveals a tumor-associated macrophage-mediated resistance pathway in anti-PD-1 therapy. Sci Transl Med. (2017) 9:eaal3604. doi: 10.1126/scitranslmed.aal3604
18. Wang Q, Li Z, Zhou S, Li Z, Huang X, He Y, et al. NCAPG2 could be an immunological and prognostic biomarker: From pan-cancer analysis to pancreatic cancer validation. Front Immunol. (2023) 14:1097403. doi: 10.3389/fimmu.2023.1097403
19. Chen Y, Song Y, Du W, Gong L, Chang H, Zou Z. Tumor-associated macrophages: an accomplice in solid tumor progression. J BioMed Sci. (2019) 26:78. doi: 10.1186/s12929-019-0568-z
20. Zhang X, Zhang P, Cong A, Feng Y, Chi H, Xia Z, et al. Unraveling molecular networks in thymic epithelial tumors: deciphering the unique signatures. Front Immunol. (2023) 14:1264325. doi: 10.3389/fimmu.2023.1264325
21. Deng Y, Shi M, Yi L, Naveed Khan M, Xia Z, Li X. Eliminating a barrier: Aiming at VISTA, reversing MDSC-mediated T cell suppression in the tumor microenvironment. Heliyon. (2024) 10:e37060. doi: 10.1016/j.heliyon.2024.e37060
22. Schmall A, Al-Tamari HM, Herold S, Kampschulte M, Weigert A, Wietelmann A, et al. Macrophage and cancer cell cross-talk via CCR2 and CX3CR1 is a fundamental mechanism driving lung cancer. Am J Respir Crit Care Med. (2015) 191:437–47. doi: 10.1164/rccm.201406-1137OC
23. Salvagno C, Ciampricotti M, Tuit S, Hau CS, van Weverwijk A, Coffelt SB, et al. Therapeutic targeting of macrophages enhances chemotherapy efficacy by unleashing type I interferon response. Nat Cell Biol. (2019) 21:511–21. doi: 10.1038/s41556-019-0298-1
24. De Henau O, Rausch M, Winkler D, Campesato LF, Liu C, Cymerman DH, et al. Overcoming resistance to checkpoint blockade therapy by targeting PI3Kgamma in myeloid cells. Nature. (2016) 539:443–7. doi: 10.1038/nature20554
25. Chen D, Zhang X, Li Z, Zhu B. Metabolic regulatory crosstalk between tumor microenvironment and tumor-associated macrophages. Theranostics. (2021) 11:1016–30. doi: 10.7150/thno.51777
26. Zhu C, Sun Z, Wang J, Meng X, Ma Z, Guo R, et al. Exploring oncogenes for renal clear cell carcinoma based on G protein-coupled receptor-associated genes. Discovery Oncol. (2023) 14:182. doi: 10.1007/s12672-023-00795-z
27. Zuo D, Zhang J, Liu T, Li C, Ning G. Claudin-1 is a valuable prognostic biomarker in colorectal cancer: A meta-analysis. Gastroenterol Res Pract. (2020) 2020:4258035. doi: 10.1155/2020/4258035
28. Cassetta L, Pollard JW. Tumor-associated macrophages. Curr Biol. (2020) 30:R246–8. doi: 10.1016/j.cub.2020.01.031
29. Hughes R, Qian BZ, Rowan C, Muthana M, Keklikoglou I, Olson OC, et al. Perivascular M2 macrophages stimulate tumor relapse after chemotherapy. Cancer Res. (2015) 75:3479–91. doi: 10.1158/0008-5472.CAN-14-3587
30. Madsen DH, Jurgensen HJ, Siersbaek MS, Kuczek DE, Grey Cloud L, Liu S, et al. Tumor-associated macrophages derived from circulating inflammatory monocytes degrade collagen through cellular uptake. Cell Rep. (2017) 21:3662–71. doi: 10.1016/j.celrep.2017.12.011
31. Nelson MP, Christmann BS, Dunaway CW, Morris A, Steele C. Experimental Pneumocystis lung infection promotes M2a alveolar macrophage-derived MMP12 production. Am J Physiol Lung Cell Mol Physiol. (2012) 303:L469–475. doi: 10.1152/ajplung.00158.2012
32. Koscso B, Csoka B, Kokai E, Nemeth ZH, Pacher P, Virag L, et al. Adenosine augments IL-10-induced STAT3 signaling in M2c macrophages. J Leukoc Biol. (2013) 94:1309–15. doi: 10.1189/jlb.0113043
33. Zhang H, Liu L, Liu J, Dang P, Hu S, Yuan W, et al. Roles of tumor-associated macrophages in anti-PD-1/PD-L1 immunotherapy for solid cancers. Mol Cancer. (2023) 22:58. doi: 10.1186/s12943-023-01725-x
34. Yuan A, Hsiao YJ, Chen HY, Chen HW, Ho CC, Chen YY, et al. Opposite effects of M1 and M2 macrophage subtypes on lung cancer progression. Sci Rep. (2015) 5:14273. doi: 10.1038/srep14273
35. Pang ZQ, Wang JS, Wang JF, Wang YX, Ji B, Xu YD, et al. JAM3: A prognostic biomarker for bladder cancer via epithelial-mesenchymal transition regulation. Biomol BioMed. (2024) 24:897–911. doi: 10.17305/bb.2024.9979
36. Engur-Ozturk S, Dikmen M. Proteasome inhibitor immunotherapy for the epithelial to mesenchymal transition: assessing the A549 lung cancer cell microenvironment and the role of M1, M2a and M2c ‘hydrocortisone-polarised’ macrophages. Mol Biol Rep. (2022) 49:4777–93. doi: 10.1007/s11033-022-07329-w
37. Wang Y, Wang J, Liu J, Zhu H. Immune-related diagnostic markers for benign prostatic hyperplasia and their potential as drug targets. Front Immunol. (2024) 15:1516362. doi: 10.3389/fimmu.2024.1516362
38. Li Z, Li J, Bai X, Huang X, Wang Q. Tumor microenvironment as a complex milieu driving cancer progression: a mini review. Clin Transl Oncol. (2024). doi: 10.1007/s12094-024-03697-w
39. Wu D, Liu Y, Liu J, Ma L, Tong X. Myeloid cell differentiation-related gene signature for predicting clinical outcome, immune microenvironment, and treatment response in lung adenocarcinoma. Sci Rep. (2024) 14:17460. doi: 10.1038/s41598-024-68111-5
40. Sumitomo R, Hirai T, Fujita M, Murakami H, Otake Y, Huang CL. PD-L1 expression on tumor-infiltrating immune cells is highly associated with M2 TAM and aggressive Malignant potential in patients with resected non-small cell lung cancer. Lung Cancer. (2019) 136:136–44. doi: 10.1016/j.lungcan.2019.08.023
41. Wang D, Yang L, Yue D, Cao L, Li L, Wang D, et al. Macrophage-derived CCL22 promotes an immunosuppressive tumor microenvironment via IL-8 in Malignant pleural effusion. Cancer Lett. (2019) 452:244–53. doi: 10.1016/j.canlet.2019.03.040
42. Doedens AL, Stockmann C, Rubinstein MP, Liao D, Zhang N, DeNardo DG, et al. Macrophage expression of hypoxia-inducible factor-1 alpha suppresses T-cell function and promotes tumor progression. Cancer Res. (2010) 70:7465–75. doi: 10.1158/0008-5472.CAN-10-1439
43. Shen G, Wang Q, Li Z, Xie J, Han X, Wei Z, et al. Bridging chronic inflammation and digestive cancer: the critical role of innate lymphoid cells in tumor microenvironments. Int J Biol Sci. (2024) 20:4799–818. doi: 10.7150/ijbs.96338
44. Yang L, Wang Q, He L, Sun X. The critical role of tumor microbiome in cancer immunotherapy. Cancer Biol Ther. (2024) 25:2301801. doi: 10.1080/15384047.2024.2301801
45. Rodriguez PC, Hernandez CP, Morrow K, Sierra R, Zabaleta J, Wyczechowska DD, et al. L-arginine deprivation regulates cyclin D3 mRNA stability in human T cells by controlling HuR expression. J Immunol. (2010) 185:5198–204. doi: 10.4049/jimmunol.1001224
46. Schuette V, Embgenbroich M, Ulas T, Welz M, Schulte-Schrepping J, Draffehn AM, et al. Mannose receptor induces T-cell tolerance via inhibition of CD45 and up-regulation of CTLA-4. Proc Natl Acad Sci U.S.A. (2016) 113:10649–54. doi: 10.1073/pnas.1605885113
47. Kaneda MM, Messer KS, Ralainirina N, Li H, Leem CJ, Gorjestani S, et al. PI3Kgamma is a molecular switch that controls immune suppression. Nature. (2016) 539:437–42. doi: 10.1038/nature19834
48. Kumar V, Patel S, Tcyganov E, Gabrilovich DI. The nature of myeloid-derived suppressor cells in the tumor microenvironment. Trends Immunol. (2016) 37:208–20. doi: 10.1016/j.it.2016.01.004
49. Noy R, Pollard JW. Tumor-associated macrophages: from mechanisms to therapy. Immunity. (2014) 41:49–61. doi: 10.1016/j.immuni.2014.06.010
50. Xia Z, Chen S, He M, Li B, Deng Y, Yi L, et al. Editorial: Targeting metabolism to activate T cells and enhance the efficacy of checkpoint blockade immunotherapy in solid tumors. Front Immunol. (2023) 14:1247178. doi: 10.3389/fimmu.2023.1247178
51. Xie H, Xi X, Lei T, Liu H, Xia Z. CD8(+) T cell exhaustion in the tumor microenvironment of breast cancer. Front Immunol. (2024) 15:1507283. doi: 10.3389/fimmu.2024.1507283
52. Lewis CE, Harney AS, Pollard JW. The multifaceted role of perivascular macrophages in tumors. Cancer Cell. (2016) 30:18–25. doi: 10.1016/j.ccell.2016.05.017
53. Dallavalasa S, Beeraka NM, Basavaraju CG, Tulimilli SV, Sadhu SP, Rajesh K, et al. The role of tumor associated macrophages (TAMs) in cancer progression, chemoresistance, angiogenesis and metastasis-current status. Curr Med Chem. (2021) 28:8203–36. doi: 10.2174/0929867328666210720143721
54. Magar AG, Morya VK, Kwak MK, Oh JU, Noh KC. A molecular perspective on HIF-1alpha and angiogenic stimulator networks and their role in solid tumors: an update. Int J Mol Sci. (2024) 25:3313. doi: 10.3390/ijms25063313
55. Zheng X, Weigert A, Reu S, Guenther S, Mansouri S, Bassaly B, et al. Spatial density and distribution of tumor-Associated macrophages predict survival in non-Small cell lung carcinoma. Cancer Res. (2020) 80:4414–25. doi: 10.1158/0008-5472.CAN-20-0069
56. Lewis JS, Landers RJ, Underwood JC, Harris AL, Lewis CE. Expression of vascular endothelial growth factor by macrophages is up-regulated in poorly vascularized areas of breast carcinomas. J Pathol. (2000) 192:150–8. doi: 10.1002/1096-9896(2000)9999:9999<::AID-PATH687>3.0.CO;2-G
57. Gao J, Liang Y, Wang L. Shaping polarization of tumor-associated macrophages in cancer immunotherapy. Front Immunol. (2022) 13:888713. doi: 10.3389/fimmu.2022.888713
58. Chen Z, Li H, Li Z, Chen S, Huang X, Zheng Z, et al. SHH/GLI2-TGF-beta1 feedback loop between cancer cells and tumor-associated macrophages maintains epithelial-mesenchymal transition and endoplasmic reticulum homeostasis in cholangiocarcinoma. Pharmacol Res. (2023) 187:106564. doi: 10.1016/j.phrs.2022.106564
59. Moeini P, Niedzwiedzka-Rystwej P. Tumor-associated macrophages: combination of therapies, the approach to improve cancer treatment. Int J Mol Sci. (2021) 22:7239. doi: 10.3390/ijms22137239
60. Zhang M, Liu J, Zhang F, Liang Q, Guo Z. Comprehensive characterization of neddylation related genes in cutaneous melanoma identified a novel prognostic signature for treatment outcomes, immune landscape. Discovery Oncol. (2024) 15:722. doi: 10.1007/s12672-024-01627-4
61. Yang L, Dong Y, Li Y, Wang D, Liu S, Wang D, et al. IL-10 derived from M2 macrophage promotes cancer stemness via JAK1/STAT1/NF-kappaB/Notch1 pathway in non-small cell lung cancer. Int J Cancer. (2019) 145:1099–110. doi: 10.1002/ijc.v145.4
62. Zhang J, Cao J, Ma S, Dong R, Meng W, Ying M, et al. Tumor hypoxia enhances Non-Small Cell Lung Cancer metastasis by selectively promoting macrophage M2 polarization through the activation of ERK signaling. Oncotarget. (2014) 5:9664–77. doi: 10.18632/oncotarget.v5i20
63. Yuan X, Wu H, Han N, Xu H, Chu Q, Yu S, et al. Notch signaling and EMT in non-small cell lung cancer: biological significance and therapeutic application. J Hematol Oncol. (2014) 7:87. doi: 10.1186/s13045-014-0087-z
64. Guo Z, Song J, Hao J, Zhao H, Du X, Li E, et al. M2 macrophages promote NSCLC metastasis by upregulating CRYAB. Cell Death Dis. (2019) 10:377. doi: 10.1038/s41419-019-1618-x
65. Wang A, Lu C, Ning Z, Gao W, Xie Y, Zhang N, et al. Tumor-associated macrophages promote Ezrin phosphorylation-mediated epithelial-mesenchymal transition in lung adenocarcinoma through FUT4/LeY up-regulation. Oncotarget. (2017) 8:28247–59. doi: 10.18632/oncotarget.16001
66. Dehai C, Bo P, Qiang T, Lihua S, Fang L, Shi J, et al. Enhanced invasion of lung adenocarcinoma cells after co-culture with THP-1-derived macrophages via the induction of EMT by IL-6. Immunol Lett. (2014) 160:1–10. doi: 10.1016/j.imlet.2014.03.004
67. Fu XL, Duan W, Su CY, Mao FY, Lv YP, Teng YS, et al. Interleukin 6 induces M2 macrophage differentiation by STAT3 activation that correlates with gastric cancer progression. Cancer Immunol Immunother. (2017) 66:1597–608. doi: 10.1007/s00262-017-2052-5
68. Zhang S, Che D, Yang F, Chi C, Meng H, Shen J, et al. Tumor-associated macrophages promote tumor metastasis via the TGF-beta/SOX9 axis in non-small cell lung cancer. Oncotarget. (2017) 8:99801–15. doi: 10.18632/oncotarget.21068
69. Zuo D, Li C, Liu T, Yue M, Zhang J, Ning G. Construction and validation of a metabolic risk model predicting prognosis of colon cancer. Sci Rep. (2021) 11:6837. doi: 10.1038/s41598-021-86286-z
70. Cardoso AP, Pinto ML, Pinto AT, Pinto MT, Monteiro C, Oliveira MI, et al. Matrix metalloproteases as maestros for the dual role of LPS- and IL-10-stimulated macrophages in cancer cell behaviour. BMC Cancer. (2015) 15:456. doi: 10.1186/s12885-015-1466-8
71. Wang R, Zhang J, Chen S, Lu M, Luo X, Yao S, et al. Tumor-associated macrophages provide a suitable microenvironment for non-small lung cancer invasion and progression. Lung Cancer. (2011) 74:188–96. doi: 10.1016/j.lungcan.2011.04.009
72. Shi L, Zhang B, Sun X, Zhang X, Lv S, Li H, et al. CC chemokine ligand 18(CCL18) promotes migration and invasion of lung cancer cells by binding to Nir1 through Nir1-ELMO1/DOC180 signaling pathway. Mol Carcinog. (2016) 55:2051–62. doi: 10.1002/mc.v55.12
73. Bonnans C, Chou J, Werb Z. Remodelling the extracellular matrix in development and disease. Nat Rev Mol Cell Biol. (2014) 15:786–801. doi: 10.1038/nrm3904
74. Sica A, Schioppa T, Mantovani A, Allavena P. Tumour-associated macrophages are a distinct M2 polarised population promoting tumour progression: potential targets of anti-cancer therapy. Eur J Cancer. (2006) 42:717–27. doi: 10.1016/j.ejca.2006.01.003
75. Feng Q, Fang W, Guo Y, Hu P, Shi J. Nebulized therapy of early orthotopic lung cancer by iron-based nanoparticles: macrophage-regulated ferroptosis of cancer stem cells. J Am Chem Soc. (2023) 145:24153–65. doi: 10.1021/jacs.3c08032
76. Wang J, Zuo Z, Yu Z, Chen Z, Tran LJ, Zhang J, et al. Collaborating single-cell and bulk RNA sequencing for comprehensive characterization of the intratumor heterogeneity and prognostic model development for bladder cancer. Aging (Albany NY). (2023) 15:12104–19. doi: 10.18632/aging.205166
77. Lan J, Sun L, Xu F, Liu L, Hu F, Song D, et al. M2 macrophage-derived exosomes promote cell migration and invasion in colon cancer. Cancer Res. (2019) 79:146–58. doi: 10.1158/0008-5472.CAN-18-0014
78. Kwon Y, Kim M, Kim Y, Jung HS, Jeoung D. Exosomal microRNAs as mediators of cellular interactions between cancer cells and macrophages. Front Immunol. (2020) 11:1167. doi: 10.3389/fimmu.2020.01167
79. Wang Y, Wang J, Liu Y, Wang X, Ren M. Multidimensional pan-cancer analysis of HSPA5 and its validation in the prognostic value of bladder cancer. Heliyon. (2024) 10:e27184. doi: 10.1016/j.heliyon.2024.e27184
80. Chen J, Zhang K, Zhi Y, Wu Y, Chen B, Bai J, et al. Tumor-derived exosomal miR-19b-3p facilitates M2 macrophage polarization and exosomal LINC00273 secretion to promote lung adenocarcinoma metastasis via Hippo pathway. Clin Transl Med. (2021) 11:e478. doi: 10.1002/ctm2.v11.9
81. Lei J, Chen P, Zhang F, Zhang N, Zhu J, Wang X, et al. M2 macrophages-derived exosomal microRNA-501-3p promotes the progression of lung cancer via targeting WD repeat domain 82. Cancer Cell Int. (2021) 21:91. doi: 10.1186/s12935-021-01783-5
82. Huang Q, Wu X, Wang Z, Chen X, Wang L, Lu Y, et al. The primordial differentiation of tumor-specific memory CD8(+) T cells as bona fide responders to PD-1/PD-L1 blockade in draining lymph nodes. Cell. (2022) 185:4049–4066.e4025. doi: 10.1016/j.cell.2022.09.020
83. Larionova I, Cherdyntseva N, Liu T, Patysheva M, Rakina M, Kzhyshkowska J. Interaction of tumor-associated macrophages and cancer chemotherapy. Oncoimmunology. (2019) 8:1596004. doi: 10.1080/2162402X.2019.1596004
84. Ruffell B, Coussens LM. Macrophages and therapeutic resistance in cancer. Cancer Cell. (2015) 27:462–72. doi: 10.1016/j.ccell.2015.02.015
85. Jiang S, Yang X, Lin Y, Liu Y, Tran LJ, Zhang J, et al. Unveiling Anoikis-related genes: A breakthrough in the prognosis of bladder cancer. J Gene Med. (2024) 26:e3651. doi: 10.1002/jgm.v26.1
86. Jayasingam SD, Citartan M, Thang TH, Mat Zin AA, Ang KC, Ch’ng ES. Evaluating the polarization of tumor-associated macrophages into M1 and M2 phenotypes in human cancer tissue: technicalities and challenges in routine clinical practice. Front Oncol. (2019) 9:1512. doi: 10.3389/fonc.2019.01512
87. Huang WC, Kuo KT, Wang CH, Yeh CT, Wang Y. Cisplatin resistant lung cancer cells promoted M2 polarization of tumor-associated macrophages via the Src/CD155/MIF functional pathway. J Exp Clin Cancer Res. (2019) 38:180. doi: 10.1186/s13046-019-1166-3
88. Zhang C, Yu X, Gao L, Zhao Y, Lai J, Lu D, et al. Noninvasive imaging of CD206-positive M2 macrophages as an early biomarker for post-chemotherapy tumor relapse and lymph node metastasis. Theranostics. (2017) 7:4276–88. doi: 10.7150/thno.20999
89. Qin J, Zhang X, Tan B, Zhang S, Yin C, Xue Q, et al. Blocking P2X7-Mediated macrophage polarization overcomes treatment resistance in lung cancer. Cancer Immunol Res. (2020) 8:1426–39. doi: 10.1158/2326-6066.CIR-20-0123
90. Ye J, Wu D, Wu P, Chen Z, Huang J. The cancer stem cell niche: cross talk between cancer stem cells and their microenvironment. Tumour Biol. (2014) 35:3945–51. doi: 10.1007/s13277-013-1561-x
91. Lee AW, States DJ. Colony-stimulating factor-1 requires PI3-kinase-mediated metabolism for proliferation and survival in myeloid cells. Cell Death Differ. (2006) 13:1900–14. doi: 10.1038/sj.cdd.4401884
92. Yadav S, Priya A, Borade DR, Agrawal-Rajput R. Macrophage subsets and their role: co-relation with colony-stimulating factor-1 receptor and clinical relevance. Immunol Res. (2023) 71:130–52. doi: 10.1007/s12026-022-09330-8
93. Baghdadi M, Endo H, Takano A, Ishikawa K, Kameda Y, Wada H, et al. High co-expression of IL-34 and M-CSF correlates with tumor progression and poor survival in lung cancers. Sci Rep. (2018) 8:418. doi: 10.1038/s41598-017-18796-8
94. Maldonado MDM, Schlom J, Hamilton DH. Blockade of tumor-derived colony-stimulating factor 1 (CSF1) promotes an immune-permissive tumor microenvironment. Cancer Immunol Immunother. (2023) 72:3349–62. doi: 10.1007/s00262-023-03496-2
95. Zhai X, Zhang H, Xia Z, Liu M, Du G, Jiang Z, et al. Oxytocin alleviates liver fibrosis via hepatic macrophages. JHEP Rep. (2024) 6:101032. doi: 10.1016/j.jhepr.2024.101032
96. Xiao J, Lin H, Liu B, Xia Z, Zhang J, Jin J. Decreased S1P and SPHK2 are involved in pancreatic acinar cell injury. biomark Med. (2019) 13:627–37. doi: 10.2217/bmm-2018-0404
97. Xiao J, Huang K, Lin H, Xia Z, Zhang J, Li D, et al. Mogroside II(E) inhibits digestive enzymes via suppression of interleukin 9/interleukin 9 receptor signalling in acute pancreatitis. Front Pharmacol. (2020) 11:859. doi: 10.3389/fphar.2020.00859
98. Zhang H, Xia T, Xia Z, Zhou H, Li Z, Wang W, et al. KIF18A inactivates hepatic stellate cells and alleviates liver fibrosis through the TTC3/Akt/mTOR pathway. Cell Mol Life Sci. (2024) 81:96. doi: 10.1007/s00018-024-05114-5
99. Qian BZ, Pollard JW. Macrophage diversity enhances tumor progression and metastasis. Cell. (2010) 141:39–51. doi: 10.1016/j.cell.2010.03.014
100. Jimenez-Garcia L, Herranz S, Luque A, Hortelano S. Critical role of p38 MAPK in IL-4-induced alternative activation of peritoneal macrophages. Eur J Immunol. (2015) 45:273–86. doi: 10.1002/eji.201444806
101. Hu Q, Wu G, Wang R, Ma H, Zhang Z, Xue Q. Cutting edges and therapeutic opportunities on tumor-associated macrophages in lung cancer. Front Immunol. (2022) 13:1007812. doi: 10.3389/fimmu.2022.1007812
102. Shi JH, Liu LN, Song DD, Liu WW, Ling C, Wu FX, et al. TRAF3/STAT6 axis regulates macrophage polarization and tumor progression. Cell Death Differ. (2023) 30:2005–16. doi: 10.1038/s41418-023-01194-1
103. Ayala-Cuellar AP, Cho J, Choi KC. Toll-like receptors: A pathway alluding to cancer control. J Cell Physiol. (2019) 234:21707–15. doi: 10.1002/jcp.28879
104. Anfray C, Varela CF, Ummarino A, Maeda A, Sironi M, Gandoy S, et al. Polymeric nanocapsules loaded with poly(I:C) and resiquimod to reprogram tumor-associated macrophages for the treatment of solid tumors. Front Immunol. (2023) 14:1334800. doi: 10.3389/fimmu.2023.1334800
105. Lee CH, Wu CL, Shiau AL. Toll-like receptor 4 signaling promotes tumor growth. J Immunother. (2010) 33:73–82. doi: 10.1097/CJI.0b013e3181b7a0a4
106. Kim S, Takahashi H, Lin WW, Descargues P, Grivennikov S, Kim Y, et al. Carcinoma-produced factors activate myeloid cells through TLR2 to stimulate metastasis. Nature. (2009) 457:102–6. doi: 10.1038/nature07623
107. Muller E, Speth M, Christopoulos PF, Lunde A, Avdagic A, Oynebraten I, et al. Both type I and type II interferons can activate antitumor M1 macrophages when combined with TLR stimulation. Front Immunol. (2018) 9:2520. doi: 10.3389/fimmu.2018.02520
108. Brown EJ, Frazier WA. Integrin-associated protein (CD47) and its ligands. Trends Cell Biol. (2001) 11:130–5. doi: 10.1016/S0962-8924(00)01906-1
109. Zhang W, Huang Q, Xiao W, Zhao Y, Pi J, Xu H, et al. Advances in anti-tumor treatments targeting the CD47/SIRPalpha axis. Front Immunol. (2020) 11:18. doi: 10.3389/fimmu.2020.00018
110. Giatromanolaki A, Mitrakas A, Anestopoulos I, Kontosis A, Koukourakis IM, Pappa A, et al. Expression of CD47 and SIRPalpha macrophage immune-checkpoint pathway in non-small-cell lung cancer. Cancers (Basel). (2022) 14:1801. doi: 10.3390/cancers14071801
111. Xu Y, Li J, Tong B, Chen M, Liu X, Zhong W, et al. Positive tumour CD47 expression is an independent prognostic factor for recurrence in resected non-small cell lung cancer. ESMO Open. (2020) 5:e000823. doi: 10.2139/ssrn.3539689
112. Lu J, Li J, Lin Z, Li H, Lou L, Ding W, et al. Reprogramming of TAMs via the STAT3/CD47-SIRPalpha axis promotes acquired resistance to EGFR-TKIs in lung cancer. Cancer Lett. (2023) 564:216205. doi: 10.1016/j.canlet.2023.216205
113. Tu MM, Abdel-Hafiz HA, Jones RT, Jean A, Hoff KJ, Duex JE, et al. Inhibition of the CCL2 receptor, CCR2, enhances tumor response to immune checkpoint therapy. Commun Biol. (2020) 3:720. doi: 10.1038/s42003-020-01441-y
114. Peranzoni E, Lemoine J, Vimeux L, Feuillet V, Barrin S, Kantari-Mimoun C, et al. Macrophages impede CD8 T cells from reaching tumor cells and limit the efficacy of anti-PD-1 treatment. Proc Natl Acad Sci U.S.A. (2018) 115:E4041–50. doi: 10.1073/pnas.1720948115
115. Low-Marchelli JM, Ardi VC, Vizcarra EA, van Rooijen N, Quigley JP, Yang J. Twist1 induces CCL2 and recruits macrophages to promote angiogenesis. Cancer Res. (2013) 73:662–71. doi: 10.1158/0008-5472.CAN-12-0653
116. Gomez-Roca CA, Cassier PA, Italiano A, Cannarile M, Ries C, Brillouet A, et al. Phase I study of RG7155, a novel anti-CSF1R antibody, in patients with advanced/metastatic solid tumors. Am Soc Clin Oncol. (2015) 33:15. doi: 10.1200/jco.2015.33.15_suppl.3005
117. Ries CH, Hoves S, Cannarile MA, Ruttinger D. CSF-1/CSF-1R targeting agents in clinical development for cancer therapy. Curr Opin Pharmacol. (2015) 23:45–51. doi: 10.1016/j.coph.2015.05.008
118. O’Brien SA, Orf J, Skrzypczynska KM, Tan H, Kim J, DeVoss J, et al. Activity of tumor-associated macrophage depletion by CSF1R blockade is highly dependent on the tumor model and timing of treatment. Cancer Immunol Immunother. (2021) 70:2401–10. doi: 10.1007/s00262-021-02861-3
119. Fritz JM, Tennis MA, Orlicky DJ, Lin H, Ju C, Redente EF, et al. Depletion of tumor-associated macrophages slows the growth of chemically induced mouse lung adenocarcinomas. Front Immunol. (2014) 5:587. doi: 10.3389/fimmu.2014.00587
120. Sun Z, Wang J, Fan Z, Yang Y, Meng X, Ma Z, et al. Investigating the prognostic role of lncRNAs associated with disulfidptosis-related genes in clear cell renal cell carcinoma. J Gene Med. (2024) 26:e3608. doi: 10.1002/jgm.v26.1
121. Cieslewicz M, Tang J, Yu JL, Cao H, Zavaljevski M, Motoyama K, et al. Targeted delivery of proapoptotic peptides to tumor-associated macrophages improves survival. Proc Natl Acad Sci U.S.A. (2013) 110:15919–24. doi: 10.1073/pnas.1312197110
122. Yan D, Kowal J, Akkari L, Schuhmacher AJ, Huse JT, West BL, et al. Inhibition of colony stimulating factor-1 receptor abrogates microenvironment-mediated therapeutic resistance in gliomas. Oncogene. (2017) 36:6049–58. doi: 10.1038/onc.2017.261
123. Jin W, Yang Q, Chi H, Wei K, Zhang P, Zhao G, et al. Ensemble deep learning enhanced with self-attention for predicting immunotherapeutic responses to cancers. Front Immunol. (2022) 13:1025330. doi: 10.3389/fimmu.2022.1025330
124. Sawa-Wejksza K, Kandefer-Szerszen M. Tumor-associated macrophages as target for antitumor therapy. Arch Immunol Ther Exp (Warsz). (2018) 66:97–111. doi: 10.1007/s00005-017-0480-8
125. Bak SP, Walters JJ, Takeya M, Conejo-Garcia JR, Berwin BL. Scavenger receptor-A-targeted leukocyte depletion inhibits peritoneal ovarian tumor progression. Cancer Res. (2007) 67:4783–9. doi: 10.1158/0008-5472.CAN-06-4410
126. Cao L, Che X, Qiu X, Li Z, Yang B, Wang S, et al. M2 macrophage infiltration into tumor islets leads to poor prognosis in non-small-cell lung cancer. Cancer Manag Res. (2019) 11:6125–38. doi: 10.2147/CMAR.S199832
127. Mei J, Xiao Z, Guo C, Pu Q, Ma L, Liu C, et al. Prognostic impact of tumor-associated macrophage infiltration in non-small cell lung cancer: A systemic review and meta-analysis. Oncotarget. (2016) 7:34217–28. doi: 10.18632/oncotarget.v7i23
128. Zhang L, Zhang K, Zhang J, Zhu J, Xi Q, Wang H, et al. Loss of fragile site-associated tumor suppressor promotes antitumor immunity via macrophage polarization. Nat Commun. (2021) 12:4300. doi: 10.1038/s41467-021-24610-x
129. Rodell CB, Arlauckas SP, Cuccarese MF, Garris CS, Li R, Ahmed MS, et al. TLR7/8-agonist-loaded nanoparticles promote the polarization of tumour-associated macrophages to enhance cancer immunotherapy. Nat BioMed Eng. (2018) 2:578–88. doi: 10.1038/s41551-018-0236-8
130. Cuccarese MF, Dubach JM, Pfirschke C, Engblom C, Garris C, Miller MA, et al. Heterogeneity of macrophage infiltration and therapeutic response in lung carcinoma revealed by 3D organ imaging. Nat Commun. (2017) 8:14293. doi: 10.1038/ncomms14293
131. Liu T, Li C, Zhang J, Hu H, Li C. Unveiling efferocytosis-related signatures through the integration of single-cell analysis and machine learning: a predictive framework for prognosis and immunotherapy response in hepatocellular carcinoma. Front Immunol. (2023) 14:1237350. doi: 10.3389/fimmu.2023.1237350
132. Li H, Huang N, Zhu W, Wu J, Yang X, Teng W, et al. Modulation the crosstalk between tumor-associated macrophages and non-small cell lung cancer to inhibit tumor migration and invasion by ginsenoside Rh2. BMC Cancer. (2018) 18:579. doi: 10.1186/s12885-018-4299-4
133. Weissleder R, Nahrendorf M, Pittet MJ. Imaging macrophages with nanoparticles. Nat Mater. (2014) 13:125–38. doi: 10.1038/nmat3780
134. Bolli E, Scherger M, Arnouk SM, Pombo Antunes AR, Strassburger D, Urschbach M, et al. Targeted repolarization of tumor-associated macrophages via imidazoquinoline-linked nanobodies. Adv Sci (Weinh). (2021) 8:2004574. doi: 10.1002/advs.202004574
135. Myers KV, Amend SR, Pienta KJ. Targeting Tyro3, Axl and MerTK (TAM receptors): implications for macrophages in the tumor microenvironment. Mol Cancer. (2019) 18:94. doi: 10.1186/s12943-019-1022-2
136. Zhang G, Wang M, Zhao H, Cui W. Function of Axl receptor tyrosine kinase in non-small cell lung cancer. Oncol Lett. (2018) 15:2726–34. doi: 10.3892/ol.2017.7694
137. Xie S, Li Y, Li X, Wang L, Yang N, Wang Y, et al. Mer receptor tyrosine kinase is frequently overexpressed in human non-small cell lung cancer, confirming resistance to erlotinib. Oncotarget. (2015) 6:9206–19. doi: 10.18632/oncotarget.v6i11
138. Cummings CT, Zhang W, Davies KD, Kirkpatrick GD, Zhang D, DeRyckere D, et al. Small molecule inhibition of MERTK is efficacious in non-small cell lung cancer models independent of driver oncogene status. Mol Cancer Ther. (2015) 14:2014–22. doi: 10.1158/1535-7163.MCT-15-0116
139. Yao Z, Zhang J, Zhang B, Liang G, Chen X, Yao F, et al. Imatinib prevents lung cancer metastasis by inhibiting M2-like polarization of macrophages. Pharmacol Res. (2018) 133:121–31. doi: 10.1016/j.phrs.2018.05.002
140. Zhou W, Yun Z, Wang T, Li C, Zhang J. BTF3-mediated regulation of BMI1 promotes colorectal cancer through influencing epithelial-mesenchymal transition and stem cell-like traits. Int J Biol Macromol. (2021) 187:800–10. doi: 10.1016/j.ijbiomac.2021.07.106
141. Wang R, Lu M, Zhang J, Chen S, Luo X, Qin Y, et al. Increased IL-10 mRNA expression in tumor-associated macrophage correlated with late stage of lung cancer. J Exp Clin Cancer Res. (2011) 30:62. doi: 10.1186/1756-9966-30-62
142. Wu JY, Huang TW, Hsieh YT, Wang YF, Yen CC, Lee GL, et al. Cancer-Derived succinate promotes macrophage polarization and cancer metastasis via succinate receptor. Mol Cell. (2020) 77:213–227.e215. doi: 10.1016/j.molcel.2019.10.023
143. Pan Y, Yu Y, Wang X, Zhang T. Tumor-associated macrophages in tumor immunity. Front Immunol. (2020) 11:583084. doi: 10.3389/fimmu.2020.583084
144. Zhou J, Kroll AV, Holay M, Fang RH, Zhang L. Biomimetic nanotechnology toward personalized vaccines. Adv Mater. (2020) 32:e1901255. doi: 10.1002/adma.201901255
145. Yu H, Yang Z, Li F, Xu L, Sun Y. Cell-mediated targeting drugs delivery systems. Drug Delivery. (2020) 27:1425–37. doi: 10.1080/10717544.2020.1831103
146. Guo L, Zhang Y, Yang Z, Peng H, Wei R, Wang C, et al. Tunneling nanotubular expressways for ultrafast and accurate M1 macrophage delivery of anticancer drugs to metastatic ovarian carcinoma. ACS Nano. (2019) 13:1078–96. doi: 10.1021/acsnano.8b08872
147. Sakai H, Kokura S, Ishikawa T, Tsuchiya R, Okajima M, Matsuyama T, et al. Effects of anticancer agents on cell viability, proliferative activity and cytokine production of peripheral blood mononuclear cells. J Clin Biochem Nutr. (2013) 52:64–71. doi: 10.3164/jcbn.12-60
148. Li S, Feng S, Ding L, Liu Y, Zhu Q, Qian Z, et al. Nanomedicine engulfed by macrophages for targeted tumor therapy. Int J Nanomedicine. (2016) 11:4107–24. doi: 10.2147/IJN.S110146
149. Doshi N, Swiston AJ, Gilbert JB, Alcaraz ML, Cohen RE, Rubner MF, et al. Cell-based drug delivery devices using phagocytosis-resistant backpacks. Adv Mater. (2011) 23:H105–109. doi: 10.1002/adma.201004074
150. Estelrich J, Busquets MA. Iron oxide nanoparticles in photothermal therapy. Molecules. (2018) 23:1567. doi: 10.3390/molecules23071567
Keywords: tumor-associated macrophages, lung cancer, immunomodulation, immunotherapy, TAMs
Citation: Zhu R, Huang J and Qian F (2025) The role of tumor-associated macrophages in lung cancer. Front. Immunol. 16:1556209. doi: 10.3389/fimmu.2025.1556209
Received: 06 January 2025; Accepted: 10 February 2025;
Published: 26 February 2025.
Edited by:
Minghua Ren, First Affiliated Hospital of Harbin Medical University, ChinaReviewed by:
Jian Liu, Qingpu Branch of Zhongshan Hospital Affiliated to Fudan University, ChinaCopyright © 2025 Zhu, Huang and Qian. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Fenhong Qian, emhhb3FpYW42MDRAMTI2LmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.