 Isis Rojas1*
Isis Rojas1* Mariana M. M. de Mello2
Mariana M. M. de Mello2 Fábio S. Zanuzzo1†
Fábio S. Zanuzzo1† Rebeccah M. Sandrelli1Ellen de Fátima C. Peroni1†
Rebeccah M. Sandrelli1Ellen de Fátima C. Peroni1† Jennifer R. Hall3
Jennifer R. Hall3 Matthew L. Rise1Elisabeth C. Urbinati2
Matthew L. Rise1Elisabeth C. Urbinati2 Anthony K. Gamperl1*
Anthony K. Gamperl1*- 1Department of Ocean Sciences, Memorial University of Newfoundland, St. John’s, NL, Canada
- 2Aquaculture Center of Universidade Estadual Paulista (UNESP), São Paulo State University, Jaboticabal, São Paulo, Brazil
- 3Aquatic Research Cluster, Core Research Equipment & Instrument Training Network (CREAIT) Network, Ocean Sciences Centre, Memorial University of Newfoundland, St. John’s, NL, Canada
Chronic hypoxia events are a common occurrence in Atlantic salmon (Salmo salar) sea-cages, especially during the summer, and their frequency and severity are predicted to increase with climate change. Although hypoxia is considered a very important fish health and welfare issue by the aquaculture industry, few studies have investigated the impact of chronic hypoxia on the fish immune system and its response to pathogen exposure. We exposed post-smolt Atlantic salmon to hypoxia (40% air sat.) for 6 weeks. Thereafter, we sampled fish prior to (i.e., at Time 0, to assess constitutive immune function), and after they were intraperitoneally injected with PBS (phosphate buffered saline) or formalin-killed Aeromonas salmonicida. We measured several innate immune parameters including: hematological immune responses [respiratory burst (RB), hemolytic activity of alternate complement system and plasma lysozyme concentration], and the relative percentage of circulating blood cells (erythrocytes/immature erythrocytes, monocytes, and granulocytes and lymphocytes) at Time 0 and at 24 hours post-injection (hpi); and the transcript expression levels of 8 anti-bacterial biomarkers in the head kidney [interleukin-1 beta (il1b), interleukin-8a (il8a), cyclooxygenase-2 (cox2), toll-like receptor 5, secreted (strl5), CC chemokine-like 19b (ccl19b), serum amyloid A5 (saa5), hepcidin anti-microbial peptide a (hampa) and cathelicidin anti-microbial peptide b (campb)] at Time 0 and at 6 and 24 hpi. In addition, we measured serum immunoglobulin (IgM) levels at Time 0 and at 8 weeks post-injection (4 weeks after a ‘boost’ injection). Fish exposed to chronic hypoxia had greater numbers of monocytes, which was consistent with the increase in RB, plasma lysozyme concentration and upregulated head kidney anti-bacterial gene expression (i.e., campb, ccl19b, hampa, il8a, stlr5). In contrast, chronic hypoxia: reduced RB and leukocyte numbers at 24 hpi in Asal compared to PBS-injected fish, and the transcript levels of campb, il1b, saa5, il8a and stlr5 at 6- and/or 24- hpi; but had no effect on constitutive or post-stimulation serum IgM titers. Overall, our results indicate that chronic hypoxia has differential effects on salmon constitutive innate immune function vs. following antigen exposure, and thus, it is still unclear how chronic hypoxia will impact the capacity of fish to defend against pathogens.
1 Introduction
Atlantic salmon (Salmo salar) are farmed at coastal sea-cages sites, where they are vulnerable to changes in environmental conditions (1–3). For example, decreases in water oxygen levels (hypoxia) are a critical stressor (1, 4), and oxygen saturation levels of ≤ 60-70% are not that atypical in the summer (1, 5–7). Additionally, average sea surface temperatures have been increasing over the past several decades (8). This will also reduce water oxygen levels, and some models predict a further increase of 1.5-2°C in ocean temperatures as well as an increase in the incidence and severity of hypoxia (8, 9). Therefore, it is very likely that farmed Atlantic salmon will face lower levels of oxygen for longer periods if climate change continues unabated.
Hypoxia has been reported to induce a stress response that may modulate and/or compromise the innate immune system. Several studies have investigated the impacts of hypoxia on the fish immune system (10–16). However, most of these authors studied the effects of acute (short-term) or intermittent hypoxia (1, 16–26). Although, acute and intermittent hypoxia may also occur at salmon farms, only a few studies have examined the effects of prolonged exposure to low oxygen levels on fish immune function (12–15, 27–31). For instance, Martinez et al. (15) exposed Coho salmon (Oncorhynchus kisutch) to 100%, 60%, 50%, 35% and 25% air saturation (air sat.) for 28 days and reported increased toll-like receptor and cytokine transcript levels in fish held at oxygen levels below 60% air sat. In contrast, Leeuwis et al. (29) acclimated sablefish (Anoplopoma fimbria) for 16 weeks to normoxia (100% air sat.) and hypoxia (40% air sat.) at 10°C, and injected the fish with formalin-killed Aeromonas salmonicida (Asal) bacterin during the sixth and the tenth weeks. At the end of the experiment, whereas hypoxia had no/limited effects on the innate immune system, this condition prevented the increase in immunoglobulin (IgM) in those fish exposed to bacterin, suggesting that the adaptive immunity of the sablefish was compromised at low oxygen concentrations. Finally, Kvamme et al. (14) acclimated Atlantic salmon to hypoxia (52% air sat.) for 58 days and injected them with poly (I:C) or a Vibrio water-based vaccine, and reported that chronic hypoxia increased the stress response and reduced or delayed the expression of important immune-related transcripts in the head kidney. However, these results are difficult to interpret (i.e., what was the magnitude of immune suppression?) as the control group was exposed to water with an air sat. of only 74 ± 3.6%, and thus, these fish would already have been considered hypoxic (13).
Understanding how prolonged severe hypoxia affects the fish’s immune system, and the underlying mechanisms, is critical to predicting the impact of climate change on fish immune function and disease resistance, and may assist the aquaculture industry in developing strategies to mitigate production losses. Therefore, in this study, we exposed Atlantic salmon to 40% and 100% air sat. for 6 weeks before the injection of formalin-killed Asal or phosphate buffered saline (PBS), and measured a number of innate immune parameters prior to the injections (Time 0) and at 6 and 24 hours post-injection. These included: the respiratory burst (RB) of leukocytes, the hemolytic activity of the alternative pathway of the complement system, lysozyme concentration, the number of several types of leukocytes, and the transcript expression levels of 8 key immune-related genes in the head kidney. In addition, we measured serum IgM titers both pre-injection (at Time 0) and 8 weeks after PBS and Asal injection.
2 Materials and methods
The experimental procedures in this study were approved by the Institutional Animal Care Committee of Memorial University of Newfoundland (MUN; Protocol #17-94-KG) and performed in accordance with the guidelines of the Canadian Council on Animal Care (32).
2.1 Animals and experimental design
Atlantic salmon were obtained from Cape D’or Sustainable Seafood (Nova Scotia, Canada) and initially held at 10°C in seawater at the Dr. Joe Brown Aquatic Research Building (JBARB; Ocean Sciences Centre, MUN) in a 6,000 L circular tank for 1 month. During this period, the fish were fed with a commercial salmonid diet (Optiline MicroBalance, Skretting, St. Andrews, NB, Canada) at a ration of 1.5% biomass daily (body weight·day−1), and photoperiod was 12 h light:12 h dark. After acclimation, 140 fish were netted, anesthetized in seawater containing tricaine methanesulfonate (TMS; AquaLife TMS, 0.1 g·L−1; Syndel Laboratories Ltd, Nanaimo, BC, Canada), weighed, and had a PIT (Passive Integrated Transponder) tag implanted in their peritoneal cavity for identification. After tagging, the fish (117.4 ± 28.1 g) were randomly distributed in two 250 L tanks at the Cold-Ocean and Deep-Sea Research Facility (CDRF; Ocean Sciences Centre, MUN) to recover for 3 weeks before the experiment started. Thereafter, a WitroxCTRL Oxygen System (Loligo® Systems, Viborg, Denmark) was used to monitor and reduce dissolved oxygen (DO) levels in one tank (hypoxia treatment) by 5% every two days until they reached 40% air saturation, this the lowest level of hypoxia that the salmon eat enough to maintain their body weight (33). After 6 weeks at these oxygen levels, 8 fish from both the normoxic (100% air sat.) and hypoxic tanks were sampled (initial sampling, Time 0) for blood and head kidney, and the remaining fish were anaesthetized (AquaLife TMS, 0.1 g·L−1) and intraperitoneally injected with 1 μL·g-1 of formalin-killed Asal or phosphate buffered saline (PBS) (29), and returned to their tanks. Seven (see Results) to eight fish per treatment were sampled at 6 and 24 hours post-injection (hpi; for both the PBS and Asal groups) for measures of innate immunity. Finally, four weeks after the initial injection, the remaining fish were again anaesthetized (AquaLife TMS, 0.1 g·L−1) and given a booster (1 μL·g-1 intraperitoneal injection of Asal or PBS). These fish were sampled for blood 4 weeks later [i.e. 14 weeks after the hypoxic fish reached 40% air sat. so that IgM titers could be measured (see below)]. During the experimental period, the water temperature was 10.3 ± 0.4°C, photoperiod was 12 h light:12 h dark and fish were fed at 1% body mass day−1. For a schematic diagram of the experimental protocol see Figure 1.
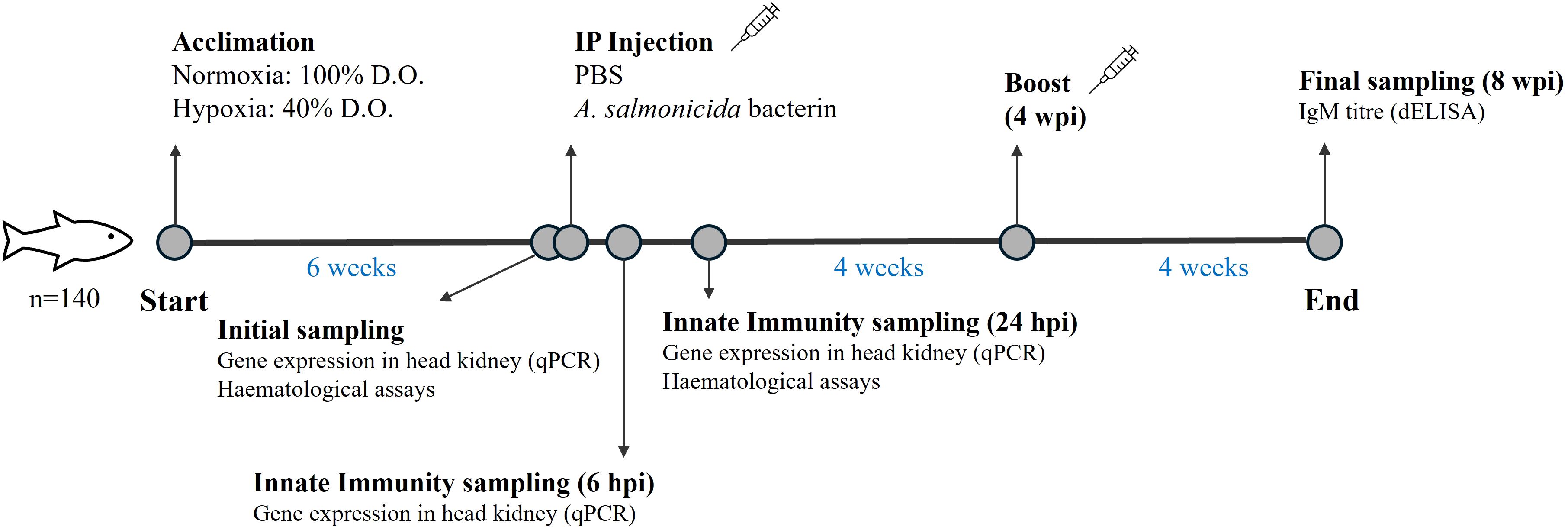
Figure 1. Schematic diagram showing the design of the experiment.
2.2 Bacteria preparation and injection
The Asal strain used in this study was isolated by Vasquez et al. (34). The bacteria were cultured in Trypticase Soy Broth (TSB) media (Difco, Franklin Lakes, NJ), and the obtained bacterial cell suspension was washed three times with PBS (pH 7.2, Gibco) by centrifugation at 4200×g for 10 min at 4°C. The Asal cells were then inactivated with 6% formaldehyde for 3 days at room temperature with gentle agitation. Formalin was removed by centrifugation at 4200×g for 10 min at 4°C and the cells were re-suspended in PBS. The bacterin cell concentrate was quantified using a Bacteria Counting Kit (Invitrogen, Thermo Fisher Scientific®, Mississauga, ON, Canada) for flow cytometry based on the manufacturers’ instructions. The bacterin was stored at 4°C at a concentration of 3.5 × 1010 CFU mL-1 until use, and then diluted in PBS to 108 CFU mL−1 prior to injection. This is the same procedure used in (29), and thus allows us to directly compare the immune responses of the Atlantic salmon and sablefish to this bacterin formulation.
2.3 Sampling
At each sampling, fish were quickly euthanized in seawater containing TMS (Aqualife TMS, 0.4 g·L−1), measured and weighed. The blood was collected by puncture of the caudal vessels using a heparinized (1,000 IU·mL-1) and a non-heparinized (with 21-G needle) syringe, and the head kidney was removed using RNase AWAY®- (Cat. No. 83931, Sigma-Aldrich®, Oakville, ON, Canada) cleaned surgical instruments. The blood collected with the non-heparinized syringe was maintained in a refrigerator overnight and centrifuged the next morning for 10 min at 10,000×g to obtain serum. The serum was stored in 1.5 mL RNase-free tubes at -80°C prior to the determination of IgM concentration. An aliquot of blood collected with a heparinized syringe at 24 hpi was immediately used in the leukocyte respiratory burst (RB) activity assay, and another aliquot was used for blood cell counting using flow cytometry. The remaining blood was centrifuged for 1 min at 10,000×g to separate the plasma. The plasma was then distributed into different 1.5 mL RNase-free tubes, immediately frozen in liquid nitrogen, and stored at -80°C prior to the determination of lysozyme concentration and the hemolytic activity of the alternative pathway of the complement system. At 6 and 24 hpi, a portion of the head kidney was sampled and placed into 1.5 mL RNase-free tubes, snap-frozen in liquid nitrogen, and stored at -80°C until preparation of RNA for immune-related transcript expression analyses.
2.4 Innate immune indicators
Innate humoral immune parameters were measured on initial (Time 0) samples and those collected 24 hpi in PBS- and formalin-killed Asal-injected fish to evaluate the effect of chronic hypoxia, and PBS vs. Asal injection, on innate immune indicators.
2.4.1 Respiratory activity of leukocytes
The production of reactive oxygen species by salmon blood leukocytes was measured using chemiluminescence following the methods described by Marnila et al. (35) and Nikoskelainen (36). Briefly, whole blood (2 µL·mL-1) was added to a mixture of 1 mM luminol (5-amino-2,3-dihydro-1,4-phthalazinedione, Cat. No. 820071, Sigma-Aldrich®) in 0.2 M sodium borate and 1X PBS. Then, 225 µL of this solution was added to triplicate wells of a 96-well white cliniplate (Cat. No. 28298-610, VWR International®, Mississauga, ON, Canada) followed by 25 µL of 20 mg·mL-1 zymosan from Saccharomyces cerevisiae (Cat. No. Z4250, Sigma-Aldrich®). Finally, chemiluminescence was measured over 60 min using a microplate reader (SpectraMax M5; Molecular Devices, Sunnydale, CA, USA) at room temperature. From this, a curve of luminescence counts per second (LCS) was obtained, and peak values were taken to represent the respiratory burst of each sample.
2.4.2 Hemolytic activity of the alternative pathway of the complement system
This parameter was determined according to Zanuzzo et al. (37). A serum sample was diluted (1:8) in TEA-EGTA-Mg2+ buffer (triethanolamine ethylene glycol tetraacetic acid; 8 mM, with 2 mM of Mg2+ and 0.1% gelatin, pH 7.4), and used to resuspend rabbit erythrocytes (RaRBC) in TEA-EGTA/Mg2+/gelatin 0.1%. A kinetic assay was then used to determine the time (in seconds) necessary for the serum sample to lyse 50% of the RaRBC suspension by measuring the absorbance of the sample at 700 nm every 20 sec for 20 min.
2.4.3 Lysozyme concentration
Plasma lysozyme concentration was determined according to Zanuzzo et al. (37) and Demers et al. (38) based on lysis of a Micrococcus lysodeikticus suspension (Cat. No. M3770, Sigma®). The samples were incubated for 30 min at 40°C to inactivate complement proteins, and then added to the M. lysodeikticus suspension. The reduction in absorbance of this solution was measured at 450 nm for 10 min using a plate reader.
2.5 Blood cell counts
The counting of blood cells was performed on Time 0 and 24 hpi PBS- and formalin-killed Asal-injected fish, following the protocol described in Inoue et al. (39). A stock solution of 3,3-dihexyloxacarbocyanine (DiOC6, Cat. No. D273, ThermoFisher Scientific®) was diluted using 1:10 Hanks balanced salt solution (HBSS) and added to a whole blood aliquot diluted in deionized water (1:19). The solution was filtered through a 100 μM mesh, centrifuged for 5 min at 500×g, and the obtained pellet was resuspended in Fluorescence-activated cell sorting (FACs) buffer. The samples were read using a BD FACSCalibur® Flow Cytometer (Becton Dickinson, San Jose, CA, USA) using BD FACSDiva software, which identifies each blood cell population by its location in a Forward scatter (FSC) vs. Side scatter (SSC) plot (see Figure 3E).
2.6 Immune-related gene expression analyses
To further evaluate the effects of chronic hypoxia and bacterial antigen administration on fish immunity, we selected 8 genes (cathelicidin anti-microbial peptide b – campb, CC chemokine-like 19b – ccl19b, cyclooxygenase-2 – cox2, hepcidin anti-microbial peptide a – hampa, interleukin-1 beta – il1b, interleukin-8a – il8a, serum amyloid A5 – saa5 and toll-like receptor 5, secreted – strl5) based on their functional activity, and these include immune receptors, cytokines, antimicrobial effectors and inflammatory mediators (40–46). Transcript expression levels of these genes in head kidney samples from the normoxia- and hypoxia-acclimated fish pre-injection, and at 6 and 24 hpi with either PBS or formalin-killed Asal, were assessed using real-time quantitative polymerase chain reaction (qPCR) analyses. All of the primer pairs utilized in the qPCR analyses conducted herein have been previously reported and subjected to quality assurance testing (see Table 1 for detailed primer information, including references).

Table 1. qPCR primer sequences used to examine the effects of chronic hypoxia on the immune response of Atlantic salmon.
Head kidney samples (approximately 100 mg of tissue) were homogenized in 400 µL of TRIzol Reagent (Cat. No. 15596-018, Invitrogen/ThermoFisher Scientific®) using a motorized Kontes RNase-Free Pellet Pestle Grinder (Kimble Chase, Vineland, NJ, USA). An additional 400 µL of TRIzol Reagent (Invitrogen/ThermoFisher Scientific®) were then added, mixed by pipetting, and the homogenates were frozen in dry ice and stored at -80°C. Frozen homogenates were further processed by slowly thawing them on ice, and then passing them through a QIAshredder (QIAGEN, Mississauga, ON, Canada) spin column following the manufacturer’s instructions. Two-hundred µL of TRIzol (Invitrogen/ThermoFisher Scientific®) were then added to each sample to make a total homogenate volume of approximately 1 mL, and the TRIzol total RNA extractions were then completed following the manufacturer’s instructions. Total RNA samples (45 µg) were treated with 6.8 Kunitz units of DNaseI (RNase-Free DNase Set, Cat. No. 79254, QIAGEN) with the manufacturer’s buffer (1X final concentration) at room temperature for 10 min to degrade any residual genomic DNA. DNase-treated RNA samples were column-purified using the RNeasy Mini Kit (Cat. No. 74106, QIAGEN) following the manufacturer’s instructions. RNA integrity was verified by 1% agarose gel electrophoresis, and RNA purity was assessed using A260/280 and A260/230 ratios determined using a NanoDrop™ UV-Vis spectrophotometer (Thermo Scientific) for both the pre-cleaned and the column-purified RNA samples. Column-purified RNA samples had A260/280 ratios between 2.0 and 2.3 and A260/230 ratios between 1.9 and 2.3.
First-strand cDNA templates for qPCR were synthesized in 20 μL reactions from 1 µg of DNaseI-treated, column-purified, total RNA using random primers (250 ng; Cat. No. 48190-011, Invitrogen/Thermo Fisher Scientific®), dNTPs (0.5 mM final concentration; Cat. No. 10297-018, Invitrogen/Thermo Fisher Scientific®) and M-MLV reverse transcriptase (200 U; Cat. No. 28025-013, Invitrogen/Thermo Fisher Scientific®) with the manufacturer’s first strand buffer (1X final concentration) and DTT (10 mM final concentration) at 37°C for 50 min.
All PCR amplifications were performed in 13 µL reactions using 1X Power SYBR Green PCR Master Mix (Cat. No. A25776, Applied Biosystems/ThermoFisher Scientific®, Waltham, MA, USA), 50 nM of both the forward and reverse primers, and cDNA representing 4 ng of input total RNA. Amplifications were performed using the ViiA 7 Real Time PCR system (384-well format) (Applied Biosystems/Thermo Fisher Scientific®). The real-time analysis program consisted of 1 cycle of 50°C for 2 min, 1 cycle of 95°C for 10 min, and 40 cycles of 95°C for 15 sec and 60°C for 1 min, with fluorescence detection at the end of each 60°C step, and was followed by dissociation curve analysis.
Expression levels of the transcripts of interest (TOIs) were normalized to transcript levels of two endogenous controls. These endogenous controls were selected from six candidate normalizers [60S ribosomal protein L32 (rpl32), β-actin (actb), elongation factor 1-alpha 1 (ef1a1), elongation factor 1-alpha 2 (ef1a2), eukaryotic translation initiation factor 3 subunit D (eif3d) and polyadenylate-binding protein cytoplasmic 1 (pabpc1)]. Briefly, the fluorescence threshold cycle (CT) values of 30 samples (i.e. 3 samples from each of the 10 groups) were measured (in triplicate) for each of these transcripts, and then analyzed using geNorm (51). Based on this analysis, rpl32 (geNorm M = 0.129) and pabpc1 (geNorm M = 0.131) were selected as the two endogenous controls.
The qPCR analyses of expression levels of the 8 TOIs in the 78 head kidney samples were then performed using the ViiA 7 Real Time PCR system (384-well format) (Applied Biosystems/ThermoFisher Scientific®). On each plate, for every sample, the TOIs and endogenous controls were tested in triplicate, and a plate linker sample (i.e., a sample that was run on all plates in each study) and a no-template control were included. The relative quantity (RQ) of each transcript was determined using the ViiA 7 Software Relative Quantification Study Application (Version 1.2.3) (Applied Biosystems/ThermoFisher Scientific®), with normalization to both rpl32 and pabpc1 transcript levels, and with amplification efficiencies incorporated. For each TOI, the sample with the lowest normalized expression (mRNA) level was set as the calibrator sample (i.e., assigned an RQ value = 1.0).
2.7 Adaptative immunity
2.7.1 IgM concentration
Salmon IgM purification was performed according to Vasquez et al. (34) with minor modifications. IgM was purified from pooled serum (~100 mL) collected from ~2 kg salmon, using an immobilized mannan binding protein (MBP) column kit (Cat. No. 44897, Pierce™, Thermo Scientific, USA) according to the manufacturer’s instructions. The integrity and purity of IgM in the elution fractions were evaluated by 10% SDS-PAGE (52) against a marker (PageRuler Plus Prestained Protein Ladder, Thermo Scientific) followed by Coomassie staining, while the concentration was estimated with DirectUV (absorbance at 280 nm) using a Genova-Nano spectrophotometer (Jenway, UK) and a bicinchoninic acid (BCA) assay (BCA protein assay kit, Pierce™, Thermo Scientific) using a plate reader (SpectraMax M5). High quality IgM fractions were pooled and dialyzed twice against 20 mM Tris buffer (pH 8.0) at 4°C with gentle stirring using a dialysis cassette or dialysis bag (3-20 kDa molecular weight cut-off, Thermo Scientific). Subsequently, the purified salmon IgM was lyophilized (EdwarDS-Super Modulyo, Boc Ltd, UK), re-suspended in 20 mM Tris buffer (pH 8.0), and re-evaluated for integrity and concentration using 10% SDS-PAGE and DirectUV. Finally, salmon IgM was stored at -20°C in aliquots to protect against repeated freeze-thaw cycles.
The production of chicken anti-salmon-IgM IgY was done commercially at Somru BioScience Inc. (Charlottetown, PEI, Canada), and involved affinity purification of the antibodies from chicken eggs using the antigen, followed by biotinylation of the antibodies.
We measured total IgM titers using dELISA according to Vasquez et al. (34) with some modifications. Serum samples were initially diluted 100,000x in carbonate-bicarbonate coating buffer (15 mM Na2CO3; 35 mM NaHCO3; pH 9.8) and heat-treated at 45°C for 30 min to inactivate complement activity. Then, 100 µL of each sample was pipetted into triplicate wells of high protein-binding polystyrene 96-well plates (Nunc MaxiSorp™, Thermo Fisher Scientific). For each plate, a pool of samples was diluted a further 30x in carbonate-bicarbonate coating buffer, and then added into duplicate wells (final dilution of 3,000,000x). The optical density (OD) of the pool was used to account for the non-specific background signal, and this value was deduced from each sample’s OD. Each plate included a standard curve with purified salmon IgM diluted in coating buffer to 3, 1.5, 0.75, 0.375, 0.1875, 0.0937 and 0.04687 µg·mL-1, and a negative control with only coating buffer, in duplicate wells. The plates were incubated at 4°C overnight and washed 4 times with 200 μL of 0.1% v/v Tween 20 in 1x PBS (PBS-T 0.1%) using a microplate washer (BioTek™ 50TS, Thermo Fisher Scientific) on the next day. Subsequently, the plates were blocked by adding 150 µL of ChonBlock™ (Cat. No. 9068, Chondrex Inc. WA, USA) in each well for 1 h at 37°C, followed by washing according to the procedure described previously. Then, plates were incubated with 100 μL of chicken anti-salmon-IgM IgY per well (diluted 1,000x in PBS-T 0.05%) for 1 h at 37°C. After washing, 100 μL of streptavidin-horseradish peroxidase (Cat. No. 405210, BioLegend, CA, USA; diluted 500X in PBS-T 0.05%) was added to each well, and the plates were incubated for 1 h at 37°C. Following washing, the plates were incubated with 100 μL of TMB solution (Kementec Solutions Inc. 4850, NH, USA) per well for 30 min at RT in the dark. The color/reaction was stopped by adding 100 μL of 0.3 M H2SO4. The bottom of the plates was wiped, and the plate read at 450 nm (SpectraMax M5; Molecular Devices, CA, USA). Based on the OD of the standards, serum IgM concentration was calculated using a 4-parameter sigmoidal regression using SoftMax Pro software.
2.8 Statistical analyses
All the data are reported as means ± 1 standard error of the mean (n=7 - 8), and P < 0.05 was used as the level of statistical significance. The data were analyzed using RStudio v. 2024.09.0 + 375 and R CRAN v. 4.4.1 (R Core Team, 2024, https://www.r-project.org/). All graphs were made using GraphPad Prism© v. 10.0.0 for Windows (MA, USA). The interquartile range method was applied to identify outliers. Based on this analysis, 3 samples were removed: one fish in the hypoxia Asal 24 hpi group; one fish in the normoxia Asal 6 hpi group; and one fish in the normoxia Asal 24 hpi group. The data were inverse or log2 transformed if necessary to meet the ANOVA normality (Shapiro-Wilk test) and homoscedasticity (Fligner test) assumptions. If not, Kruskal-Wallis tests were performed, and Dunn’s post-hoc tests [package ‘dunn.test’ v. 1.3.6 (53)] were used to identify differences between groups.
We performed two different analyses of variance (ANOVAs) as follows: a 2 (hypoxia and normoxia) x 2 (PBS and Asal) factorial test to examine the innate immune responses, the cellular blood count data and serum IgM levels; and a 2 (hypoxia and normoxia) x 2 (PBS and Asal) x 2 (6 and 24 hpi) factorial test to analyze gene expression. Both of these analyses were followed by Tukey’s [package ‘agricolae’ v. 1.3-7 (54)] post-hoc tests to enable pairwise comparisons. To compare the basal (constitutive) gene expression between treatments, a t-test was used. Dunnett’s test [package ‘FSA’ v. 0. 95 (55)] was performed to examine if values were different from basal values within each oxygen condition (treatment).
3 Results
In this study, we evaluated the effects of 6+ weeks of severe hypoxia (40% air sat.) on the constitutive (basal) and antigen-stimulated innate and acquired immune function of Atlantic salmon. However, this level of hypoxia had a number of other effects on the salmon. Salmon maintained under hypoxia were less active, spent time at the water’s surface, had an increased gill ventilation rate, and grew much slower than fish held in normoxic water (de Mello et al., unpubl). Two fish from the hypoxia treatment died after injection with formalin-killed A. salmonicida, however, these fish were without any external signs of disease/ill health.
3.1 Innate immune response
Constitutive (basal) leukocyte respiratory burst activity (RB) and lysozyme concentration were significantly greater (by 52 and 78%, respectively) in hypoxia-exposed vs. normoxic fish (Figure 2; Supplementary Table S1). At 24 hpi with PBS or attenuated Asal there was an approximately 2.5-fold increase in RB in normoxia-acclimated fish (Figure 2A). However, there were no significant differences between the initial sampling and both PBS and Asal-injected values for fish held at 40% air saturation. This difference in responsiveness resulted in RB in normoxia-acclimated fish being nearly significantly (P = 0.057) or significantly higher at 24 hpi in normoxia- vs. hypoxia-acclimated fish following PBS and Asal injection, respectively. A similar pattern was observed for lysozyme concentration (Figure 2C), where Asal injection into normoxia-acclimated, but not hypoxia-acclimated, fish resulted in an increase in this parameter as compared to initial values. However, in this case, the differences in PBS and Asal injected fish did not reach significance. No significant differences in initial or post-injection hemolytic activity of the complement system were found (Figure 2B).
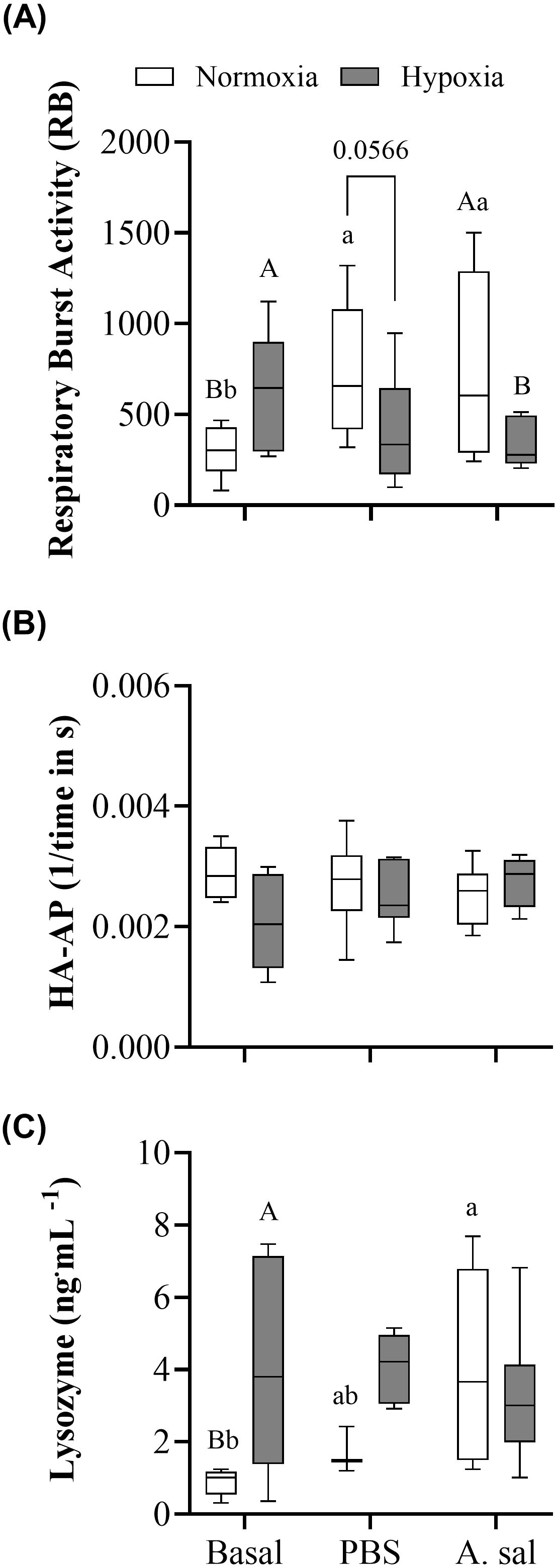
Figure 2. Blood respiratory burst activity (RB) of leukocytes (A), hemolytic activity of the alternative pathway of the complement system (HAAP); (B) and plasma lysozyme concentration (C) in Atlantic salmon after 6 weeks of normoxia or hypoxia (40% air saturation), and 24 hours after these groups were injected with PBS or formalin-killed A. salmonicida. Different capital letters indicate a significant (P < 0.05) difference between treatments at the same sampling time. Different lowercase letters indicate differences between sampling times within the same treatment. Symbols were omitted when there was no statistically significant difference. Values are presented as means ± 1 standard error (n=7-8).
3.2 Blood cells counts
The percentage of circulating monocytes and granulocytes decreased significantly after the injection of PBS or Asal in hypoxia-acclimated, but not normoxia-acclimated, fish compared to initial values (Figure 3C; Supplementary Table S1). This resulted in this parameter being lower (by 0.1 to 0.15%) in hypoxia- vs. normoxia-acclimated fish in the PBS-injected group at 24 hpi. The percentage of lymphocytes was not different pre-injection. The percentage of these cells fell in both groups following PBS and Asal injection. However, the decrease was greater in hypoxia-acclimated fish, and this resulted in lower values (by approx. 1%) in both injection groups at 24 hpi.

Figure 3. Percentage of erythrocytes (A), immature erythrocytes (B), monocytes and granulocytes (C) and lymphocytes (D) in Atlantic salmon after 6 weeks of hypoxia (40% air saturation) or normoxia, and 24 hours after these groups were injected with PBS or formalin-killed A. salmonicida. Different capital letters indicate a difference (P < 0.05) between treatments at the same sampling time. Different lowercase letters indicate differences between sampling times within the same treatment. Symbols were omitted when there was no statistically significant difference. Values are presented as means ± 1 standard error (n=7-8). Image (E) shows the different populations of cells obtained through flow cytometry in samples from normoxia- vs. hypoxia-acclimated fish.
Hypoxia-acclimation had no effect on the initial percentage of erythrocytes, however, this parameter was slightly higher in hypoxia-acclimated fish independent of whether they were injected with PBS or Asal (Figure 3A; Supplementary Table S1). Circulating immature erythrocytes increased in hypoxia-acclimated fish at 24 hpi when given either injection. However, these values were not significantly different from those measured in normoxia-acclimated fish at this time point (Figure 3B; Supplementary Table S1).
3.3 Gene expression
Transcript expression levels of campb, ccl19b, hampa, il8a and stlr5 were significantly higher (by 1.6-, 1.6-, 1.5-, 2.5- and 7-fold, respectively) at the initial sampling in the hypoxia- compared to normoxia-acclimated fish (Figures 4A, B, D, F, H, respectively; Supplementary Table S2). However, no significant differences were observed in cox2, il1b or saa5 expression levels (Figures 4C, E, G, respectively; Supplementary Table S2).
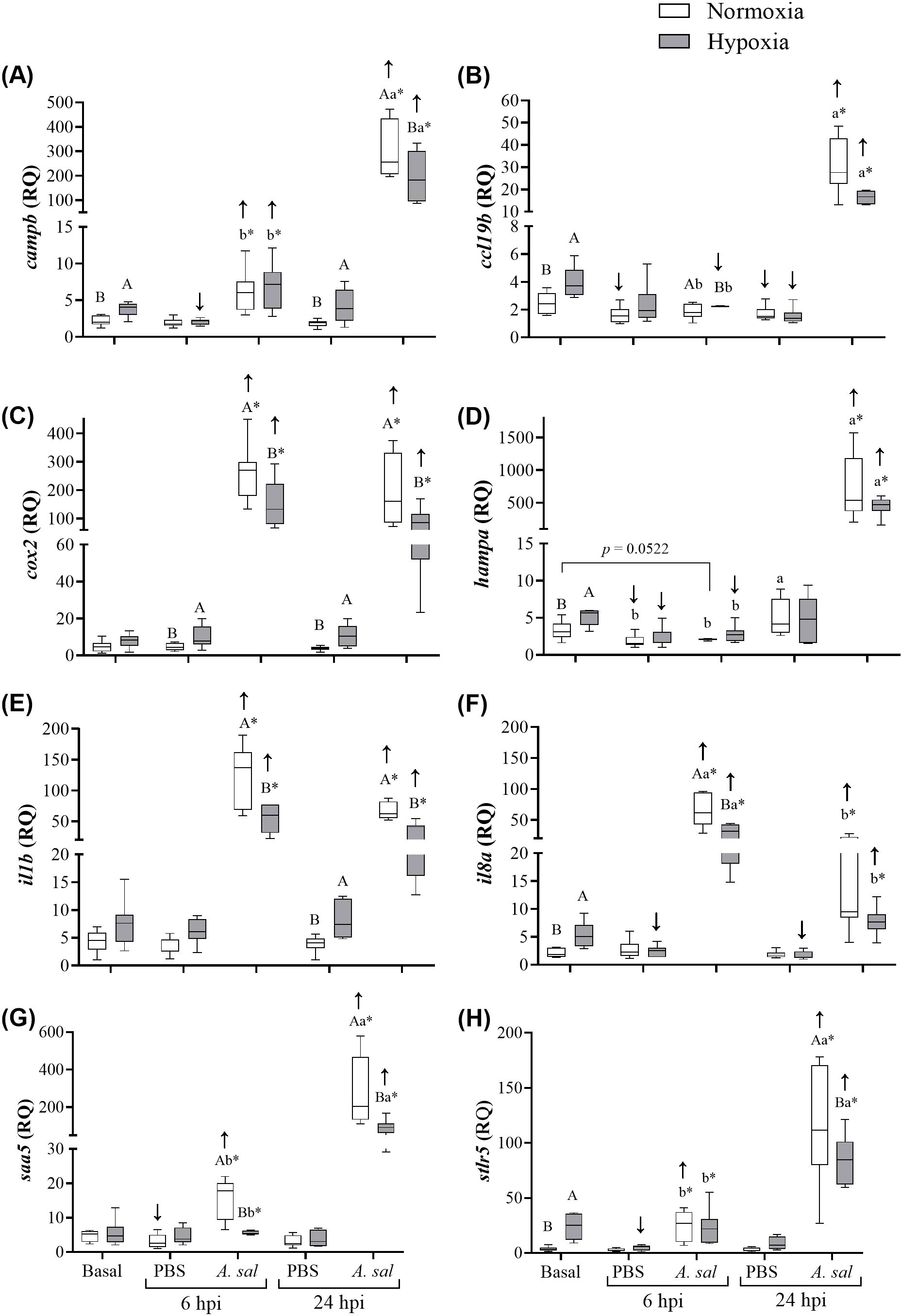
Figure 4. Transcript (mRNA) expression of 8 antibacterial biomarker genes in Atlantic salmon head kidney after 6 weeks of hypoxia (40% air saturation) or normoxia, and in these fish 6 and 24 hours post-injection (hpi) with PBS or formalin-killed A. salmonicida. (A) campb, (B), ccl19b, (C) cox2, (D) hampa, (E) il1b, (F) il8a, (G) saa5, (H) stlr5. Different capital letters indicate significant (P < 0.05) differences between treatments at the same sampling time. Different lowercase letters indicate differences between sampling times (6 and 24 hpi) within the same treatment. Asterisks (*) indicate a difference between injection groups (PBS and A. salmonicida). Arrows indicate difference between initial sampling and each injection group at each sampling time. Symbols were omitted when there was no statistical difference. Values are presented as means ± 1 standard error (n=7-8).
In hypoxia-acclimated fish injected with PBS at 6 hpi, levels of campb, hampa, il8a and stlr5 were significantly lower (by 45, 53, 54 and 81%, respectively) compared to basal expression levels. In contrast, in the normoxia-acclimated fish injected with PBS, levels of ccl19b, hampa and saa5 were significantly lower (by 45, 42 and 17%, respectively) compared to basal expression levels. With the exception of ccl19b and il8a for the hypoxic fish, these values were not significantly lower at 24 hpi, and several genes (i.e., campb, cox2, il1b) were again more highly expressed in hypoxia- than normoxia-acclimated fish.
Injection of Asal into normoxia-acclimated Atlantic salmon substantially increased the expression of 6 of the 8 genes at 6 hpi (ccl19b and hampa were the exception), and all of the genes at 24 hpi. These changes ranged from 2.7- to 53-fold. A similar pattern of gene expression was observed in hypoxia-acclimated fish at the two post-injection sampling points. However, in hypoxia- compared to normoxia-acclimated fish, the magnitude of the increase in transcript expression (by 1.8- to 7.1-fold) was significantly lower at both sampling time points for cox2, il1b and saa5, and at 6 hpi or 24 hpi for il8a and campb/stlr, respectively, and a similar (but not statistically-significant) decrease was evident for ccl19b and hampa (53 and 62%, respectively) at 24 hpi.
3.4 Adaptive immune response
Despite serum IgM levels being 39% higher in the normoxia-acclimated fish after the PBS injection compared to the initial sampling, and 33% higher than in the hypoxia-acclimated fish following PBS injection (P < 0.10), no significant differences were found in constitutive IgM levels between the normoxia- and hypoxia-acclimated groups, or for either injection group as compared to pre-injection (initial) values (Figure 5; Supplementary Table S1).
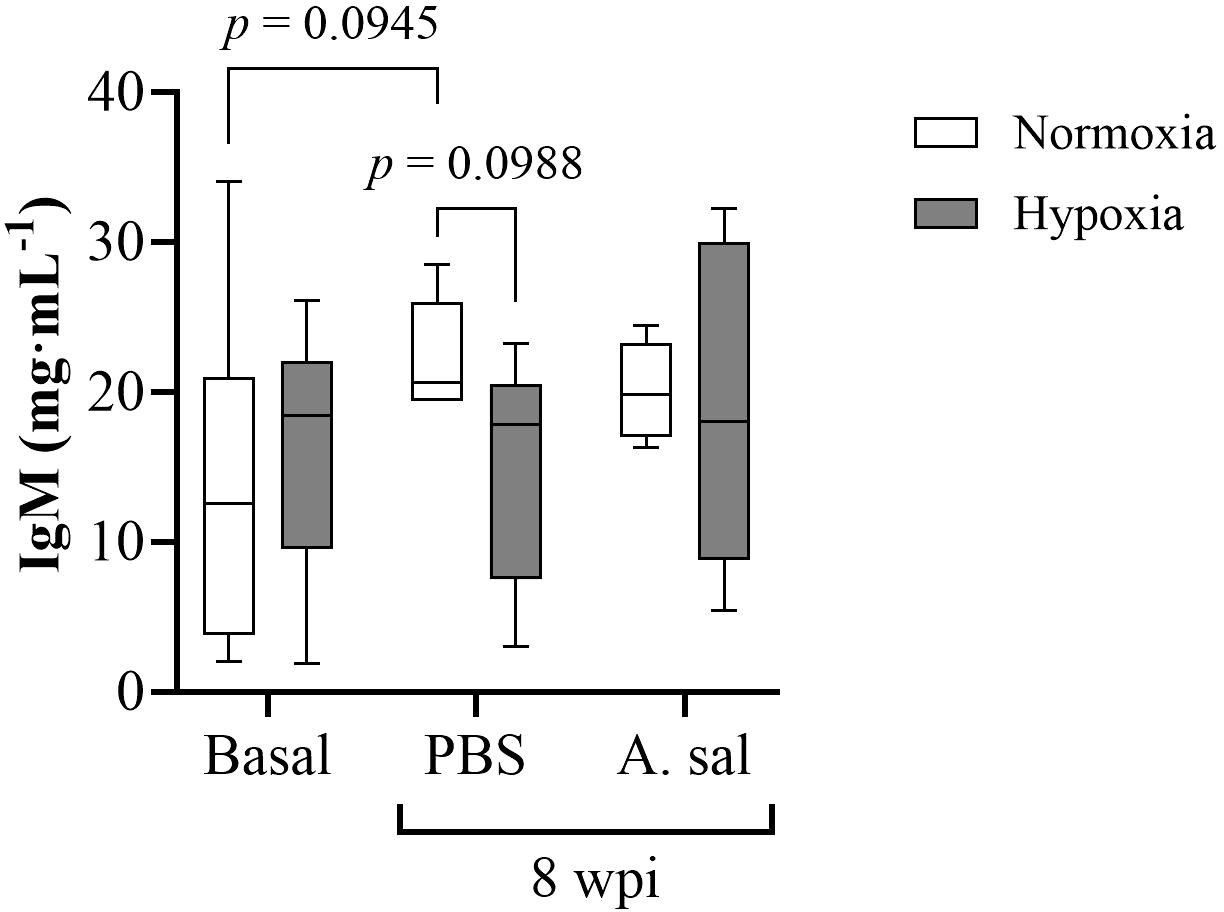
Figure 5. Serum IgM concentration in Atlantic salmon after 6 weeks of hypoxia (40% air saturation) or normoxia, and 8 weeks postinjection (wpi) with PBS or formalin-killed A. salmonicida. Values are presented as means ± 1 standard error (n=7-8).
4 Discussion
4.1 Effect of chronic hypoxia on basal immune function
4.1.1 Humoral immunity and blood cell populations
The innate, or non-specific, immune system provides the first line of defense against pathogens given its rapid response to non-specific antigens (56, 57). In this study, we measured three humoral parameters related to innate immunity: (a) RB activity, (b) hemolytic activity of the alternate complement system, and (c) plasma lysozyme concentration (Figure 2). Salmon held at 40% air sat. for six weeks had increased RB activity compared to fish maintained under normoxia (100% air sat.) by more than 50%. This finding is in contrast to data on Atlantic salmon exposed to intermittent hypoxia (3.5 times day-1 with a mean duration of 69 min) which showed a decrease in head kidney leukocyte RB (10), and on sablefish [Anoplopoma fimbria (29)] held at the same oxygen level as in this study where no effect of hypoxia was observed. However, this response is consistent with what Zanuzzo et al. (58) reported for Atlantic salmon chronically exposed to 60-70% air sat at 20°C. Collectively, these data suggest that hypoxia has different effects on this parameter depending on species and whether the period of oxygen deprivation is short-term (acute) or prolonged. Lysozyme concentration can be used as an indicator of the innate immune response (56, 59), since it is rapidly induced in fish exposed to stressors and has potent bacteriolytic activity (60). There is limited data on the effects of hypoxia on plasma lysozyme activity, although Ni et al. (61) did observe significant increases in total lysozyme protein levels after 6 h of acute hypoxia (5 mg L-1 and 3 mg L-1) in amur sturgeon (Acipenser schrenckii). In our study the increase in RB and lysozyme activity may have been primarily related to the increase in monocyte and granulocyte numbers as these cells produce both reactive oxygen species and lysozyme (62, 63). What resulted in the proliferation of these cells is not known. However, basal cortisol levels were much lower in this population of salmon when held under severe hypoxia vs. normoxia [i.e. ~ 8 vs. 16 ng mL-1; de Mello et al. (unpubl)], and this finding is consistent with Kvamme et al. (14) who also reported lower cortisol levels in hypoxia- vs. normoxia-acclimated Atlantic salmon when oxygen deprivation was prolonged. However, a difference in cortisol levels in normoxic vs. hypoxic sablefish was not observed in our previous study (29), and this finding might explain the disparity in results with regards to alternative complement activity and lysozyme concentration between the two species. Elevated cortisol levels have been reported to result in decreased lymphocyte counts and disease resistance in fishes (64, 65).
In contrast to the results for RB and lysozyme activity, we did not detect any difference in the activity of the alternate complement system (Figure 2B), which plays a crucial role in the host defense and is composed of a wide range of plasma proteins that lyse pathogens (66). However, this was not unexpected. Previous studies have reported that hypoxia does not change the activity of the alternate complement system. For example, both Magnoni et al. (67) and Leeuwis et al. (29) failed to detect any changes in alternative complement activity when rainbow trout (Oncorhynchus mykiss) and sablefish, respectively, were held in hypoxic conditions for prolonged periods. This may be because alternative complement is a basal/constitutive component of the defense against pathogens (i.e., is always active) (68). So much so that complement proteins are transferred from parents to progeny (eggs) (69).
Erythrocytes (RBC) are the predominant cells in circulation, and an increase in erythrocyte numbers is a typical response to the lack of oxygen as this will/may help to acquire more oxygen at the gills and optimize oxygen transport to the tissues (70). This was not observed in our fish, and suggests that at the temperature that the fish were held (10°C) blood oxygen transport capacity was sufficient to meet the Atlantic salmon’s basal requirements.
4.1.2 Immune-related gene expression
To examine the effects of chronic hypoxia on the Atlantic salmon’s immune system at the molecular level, we measured the expression of eight immune-related genes in the head kidney, including campb, ccl19b, cox2, hampa, il1b, il8a, saa5 and strl5. Overall, our results suggest that salmon held under conditions of chronic severe hypoxia have an enhanced constitutive (basal) innate immune capacity, as shown by the higher expression of biomarkers related to bacterial recognition (stlr5), the inflammatory process (il8a), antibacterial activities (campb, hampa) and lymphocyte recruitment/proliferation (ccl19b) (Figure 4). TLRs have traditionally been associated with pathogen recognition (71), however, these molecules can also recognize hypoxia-related molecules such as heat shock proteins (HSPs) and hypoxia inducible factors (HIFs). This has been demonstrated in human endothelial cells and murine macrophages (72, 73), and not surprisingly, prolonged hypoxia resulted in a significant upregulation of this recognition factor in catla (Catla catla) (74). This observation is also in accordance with our results, as basal (constitutive) stlr5 expression was ~7-fold higher in hypoxia-acclimated salmon (Figure 4H), and liver hsp47 (also known as serpinh1), but not hif1ac (hypoxia inducible factor 1 alpha c), transcript expression levels were significantly higher in salmon chronically exposed to 40% air saturation (de Mello et al., unpubl.). However, it is contrary to the data for hypoxic vs. normoxic sablefish, and thus, a response to lower levels of cortisol cannot be excluded for stlr5 expression, or the transcript expression of any of the other genes that we report to be upregulated by chronic exposure to 40% hypoxia (see below). It has been proposed that TLR expression is also linked to the concomitant upregulation of some interleukins in channel catfish (Ictalurus punctatus) exposed to hypoxic conditions (75). Interleukins are known to act as pro-inflammatory cytokines that mediate immune responses (76). Although we did not see any changes in basal il1b expression, the basal expression of il8a was significantly upregulated (~2.7-fold change; Figure 4F) in the hypoxia-acclimated fish. Interleukin-8 is a chemokine that specifically attracts neutrophils and basophils (both granulocytes), and lymphocytes, to sites of infection (77, 78). Therefore, not surprisingly, the higher proportion of leukocytes in the hypoxia-acclimated fish is in accordance with an up-regulation of constitutive levels of il8a. In contrast, however, neither il8a expression nor leukocyte counts were different in sablefish acclimated to hypoxia (29), and again this likely reflects the much greater hypoxia tolerance, and the differential effects of hypoxia on the stress and physiology, of these fishes [(29, 79); de Mello et al., unpubl.].
The gene ccl19b is a proinflammatory chemokine that plays a central role in homeostasis and in the immune response (80), and is one of the most evolutionary conserved members of the chemokine family (81). Hypoxia-acclimated Atlantic salmon showed an upregulation of this gene under basal conditions (by ~1.6-fold; Figure 4B). Importantly, this chemokine is expressed and regulated by leukocytes [reviewed by (82)], and thus, its upregulation was not surprising given the higher leukocyte counts in the hypoxia-acclimated fish. In fishes, ccl19 paralogues have been traditionally linked to the immune response to pathogens (81, 83), while other CC chemokines-like molecules have been related to physiological changes, including those associated with hypoxia (82). Interestingly, ccl19 is negatively related to angiogenesis, a common mechanism of adaptation to low oxygen conditions (83, 84), but regulatory mechanisms need to be further investigated. Another mechanism of adaptation to hypoxia includes alterations in glycolysis [reviewed by (80)]. Despite this, we did not measure metabolic parameters in the present study, and it would be relevant to investigate possible interactions between adaptive mechanisms to low oxygen conditions in the Atlantic salmon and the trade-off with immune status.
Cathelicidin (CAMP) and hepcidin (HAMP) are cationic anti-microbial peptides (AMPs) which have been traditionally associated with the direct lysis of pathogens (85, 86), but also with immunomodulatory functions and inflammation (87–89). We report an upregulation of both campb and hampa (by 1.6- and 1.5-fold, respectively) basal expression in hypoxia-acclimated salmon (Figures 4A, D). HAMP activity is regulated by iron storage status and systemic inflammation, and is a major inducer of hepcidin expression (90). In humans, hypoxic conditions suppress HAMP activity, and hypoxia promotes erythropoiesis to enhance oxygen transport by inducing the production of erythroferrone, a suppressor of HAMP (91, 92). However, we observed the opposite. Hypoxia-acclimated salmon upregulated the expression of hampa, and furthermore, the proportion of immature erythrocytes did not differ between salmon under basal conditions. This suggests that either (1): these fish adapted to hypoxic conditions by lowering their metabolism and not by increasing oxygen uptake capacity; and/or (2) they are in a state of chronic inflammation, as showed by the increased leukocyte numbers and inflammatory biomarkers. On the other hand, CAMP can stimulate angiogenesis to increase oxygen delivery to tissues, and inhibit the inflammatory response by protecting the endoplasmic reticulum from stress (93–96). It is expressed in leukocytes (97), and thus, the increase in its expression is not surprising given the hypoxia-induced increase in these cells.
4.1.3 Serum IgM levels
It is known that IgM levels depend on a number of factors including species, age, weight, gender, season, and infection and vaccination status [reviewed by (98)]. Previous studies have shown that IgM titers in Atlantic salmon are generally low as compared to other species like rainbow trout (9 mg mL-1), halibut (Hippoglossus hippoglossus, 4 mg mL-1) and Atlantic cod (Gadus morhua, 11.5 mg mL-1) (99, 100). However, we report serum IgM levels of ~13 to 16 mg mL-1 in hypoxia and normoxia, respectively; these values more than 10 times higher than reported by Magnadottir et al. (101) for 2-5 kg Atlantic salmon reared in land based tanks at 2-8°C (~1 mg mL-1). We cannot explain this discrepancy at this time. However, the important finding here is that prolonged hypoxia did not affect serum concentrations of IgM in our Atlantic salmon. Immunoglobulins enhance antigen-specific opsonization and phagocytosis, and are synthesized by lymphocytes (102), and thus, this finding is in agreement with the lack of a change in lymphocyte counts in this study and in pikeperch [Sander luciopera (30)] and sablefish (29) when exposed to hypoxia.
4.2 Hypoxia-related impacts on antigen-stimulated immune function
4.2.1 Humoral immunity and blood cell populations
While pre-injection numbers (the percentage) of mature or immature RBCs did not differ between acclimation conditions in this study, the percentage of the former was higher in hypoxia-acclimated fish following the injection of both PBS and Asal, and an increase in immature erythrocytes following injection was only seen in this group (Figures 3A, B). These data suggest that chronic exposure to 40% air saturated water resulted in greater storage of these cells in the spleen, and/or a greater release of these cells. In support of this hypothesis, it has been reported for many fish species that stress causes splenic contractions and the release of RBCs into the circulation to deal with increased oxygen demands [e.g., in rainbow trout (O. mykiss) (103), tambaqui (Colossoma macropomum) (104) and amur sturgeon (A. schrenckii) (61)]. However, it is also possible that this increase in post-injection mature and immature RBC’s in the hypoxia-acclimated fish simply reflected the larger decrease in post-injection leukocyte numbers in this group (Figures 3C, D; see below). In contrast to the results of this study, there was no effect of this environmental challenge on sablefish post-injection (stress) blood erythrocyte numbers/percentages. The sablefish is an extremely hypoxia- tolerant fish (79), and it is possible that such increases in blood oxygen capacity are not needed in this species until hypoxia becomes more severe.
After both PBS and Asal injection, RB and lysozyme activity increased in fish held under normoxia, while no increase was observed in fish acclimated to hypoxia (Figures 2A, C). This suggests that the latter group was unable to increase these parameters in the blood after an acute stimulus (stressor). Again, changes in RB, lysozyme activity and the number/percentage of circulating leukocytes (especially granulocytes and monocytes) were similar in magnitude (Figures 3C, D), and thus, we hypothesize that these differences between the two groups were due to a reduction in leukocyte production/activation in fish kept under hypoxia, and not to the migration of white blood cells to the inflammation site. In this case, however (as opposed to under basal conditions), the injection of PBS and Asal did not result in a difference in plasma cortisol levels in the two groups (de Mello et al., unpubl.), and this strongly suggests that the reduction in leukocyte numbers, RB and lysozyme activity in hypoxia-acclimated fish was not stress-related, but directly linked to the lower oxygen levels to which these fish were exposed. Our results agree with data on Nile tilapia (Oreochromis niloticus) held at low (0.1-1.5 mg L-1), medium (3.0-3.5 mg L-1) and high (6.5-7.0 mg L-1) dissolved oxygen (DO) levels for 12 weeks, where fish injected with Aeromonas hydrophila had a reduced lysozyme concentration at the lower DO levels (17). On the contrary, Leeuwis et al. (29) reported no differences in lysozyme activity after sablefish exposed for 16 weeks to hypoxic conditions were injected Asal. Overall, their results suggest that chronic hypoxia may have little effect on the innate immune system in teleost species that are tolerant of low water oxygen levels.
Regardless of the injection (i.e., PBS or Asal), the post-injection activity of the alternative complement pathway was not different between the two groups (Figure 2B). Again, this finding contrasts with the results for sablefish, where the activity of this pathway decreased in hypoxia-acclimated sablefish injected with Asal (29).
4.2.2 Immune-related gene expression
Phosphate buffered saline injection resulted in relatively minor increases in transcript expression at 6 or 24 hpi, however, campb, cox2 and il1b expression were significantly higher at 24 hpi in hypoxia- as compared to normoxia-acclimated fish (by 2.1- to 2.8-fold, Figure 4). In contrast, Asal injection resulted in very large increases in transcript expression, and this is consistent with other studies that have used formalin-killed formulations of this pathogen (29, 105) or other bacterial antigens (58) to stimulate immune-related gene expression or other physiological responses in Atlantic salmon. Importantly, however, the expression of 6 of the 8 genes was significantly lower in the hypoxia-acclimated fish at 6 and/or 24 hpi as compared to normoxia-acclimated fish, and the two where differences were not significant (ccl19b and hampa) showed a similar reduction in expression after Asal injection at 24 hpi (Figure 4). These results clearly show that antigen-stimulated gene expression is suppressed in hypoxia-acclimated salmon, and suggest that these fish would be more susceptible to bacterial infections during prolonged periods of low water oxygen levels. However, the reason(s) for this reduced antigen-stimulated gene expression is/are not known. Post-injection cortisol levels were only marginally (and not significantly) elevated in these Atlantic salmon, and no difference in post-injection cortisol levels was found between hypoxia- and normoxia-acclimated fish 6 or 24 hpi with PBS or Asal (de Mello et al., unpubl.). Further, although some authors suggest that mounting an immune response to pathogens/antigens is an energy-intensive endeavor [e.g., see (106)], and thus that redirection of energy to maintain homeostasis in the face of a stressor may reduce that available to deal with a pathogen (107), Zanuzzo et al. (105) did not detect any difference in the rainbow trout’s oxygen consumption when injected with Asal vs. PBS and this agrees with recent work on the coral reef damselfish Pomacentrus amboinensis (108). Overall, these data suggest that any metabolic expenditures associated with the innate immune response are minor, and not likely to impact the immune response at varied water oxygen levels. Despite this, we show that ccl19b expression was reduced by approximately ~50% in fish held at 40% air sat., and this suggests a reduced inflammatory response and is consistent with the lower number/percentage of leukocytes.
In a previous study, expression of six of the eight genes (il1b, campb, il8a, hampa, cox2 and stlr5) we studied were measured in Atlantic salmon kept at 20°C under normoxic and hypoxic (60-70% air sat.) conditions and then vaccinated against bacterial pathogens (58). While the addition of moderate hypoxia did not affect peak post-vaccination transcript expression in these fish, the data suggested that hypoxia increased the duration of elevated immune-relevant transcript expression when fish were exposed to high temperatures. This finding is in contrast to our study and that of Kvamme et al. (14) who showed that long-term hypoxia reduced or delayed the expression of these genes in the salmon head kidney when these fish were injected with poly (I:C) or a water-based V. anguillarum vaccine at 10°C. Collectively, these studies suggest that hypoxia’s effect on innate immune gene expression is temperature dependent. However, studies directly investigating this question do not support this hypothesis. For example, Wang et al. (23) investigated the effect of 14 days of intermittent hypoxia at two temperatures on the immune response of Nile tilapia (O. niloticus) vaccinated with Streptococcus agalactiae, and showed that il-1β, tnf-α and ifn-γ were strongly down-regulated in hypoxic fish independent of temperature. As with other data we report in this paper, the decrease in antigen-stimulated gene expression in hypoxia-acclimated salmon does not agree with our previous study on the sablefish at similar DO levels and temperature. In the sablefish, for the vast majority of genes studies (i.e., 15 out of 16), hypoxia had no effect or increased Asal-stimulated gene expression, with tlr5b the only gene showing a decrease in transcript expression.
4.2.3 Serum IgM
Contrary to the results for innate gene expression, we did not see a significant increase in IgM serum titers, or a difference in IgM titers in hypoxia- vs. normoxia-acclimated salmon, following the two injections of Asal (initial and boost; Figure 5). The reason(s) for the lack of IgM increase following Asal injection is/are not known as this bacteria’s antigens clearly stimulated the innate immune system. However, it may be due to the post-injection sampling point chosen. Using a similar Asal preparation, Leeuwis et al. (29) reported an increase in serum IgM titers in normoxic-acclimated sablefish at 10 weeks post-injection (wpi), but not at six wpi. Thus, it is possible that we would have been able to detect differences in serum IgM levels both post-injection and following hypoxic acclimation [as in (29)] if the fish had been sampled a few weeks later at 10°C. However, it may also be that the adaptive immune system of hypoxia-tolerant and intolerant fish species responds differently to prolonged hypoxia. For example, the tilapia, like the sablefish, is a very hypoxia-tolerant fish species, and Gallage et al. (109) showed that serum antibody titer was significantly lower in vaccinated Nile tilapia kept at 55% air saturation as compared to under normoxic conditions.
4.2.4 Conclusions and perspectives
In this experiment we showed that: 1) chronic severe hypoxia improves basal/constitutive humoral immunity and immune-related gene expression in the head kidney; and 2) that while this condition limits the capacity of the innate immune system to respond to bacterial antigens, it did not appear to affect the adaptive immune system as measured by serum IgM titers. These data add considerably to our understanding of how the immune system of Atlantic salmon, and potentially other salmonids, responds to prolonged periods of severe oxygen limitation. However, it is unclear how these changes would influence the ability of these fish to defend against live pathogens. This is because: of the varied responses indicated above; there are very few studies that have examined the effects of chronic hypoxia on disease resistance in this taxon [but see (13)]; to our knowledge no other studies have examined how both the innate and acquired immune system respond to pathogens when challenged at low oxygen levels; and chronic hypoxia may cause an imbalance in the microbiome of the fish, which could lead to an increase in the abundance of opportunistic pathogens (110). Further, based on the few studies that have been conducted [e.g., this study (29, 109)], it appears that the immune system of hypoxia-tolerant fishes (e.g., sablefish, tilapia, carp) may respond differently to pathogens encountered when oxygen levels are low. Clearly, this is an area that demands further study, as are: the mechanisms by which hypoxia depresses the fish’s immune system; and the interactive effects of temperature and hypoxia on fish immune function. Post-injection cortisol levels were not different in this population of Atlantic salmon (de Mello et al., unpubl.), and there is little evidence to support the belief that the immune system requires large metabolic expenses and must be curtailed when oxygen availability is limited. Thus, it is still unclear how hypoxia mediates its effects on the piscine immune system. Further, environmental effects on fish immune function were recently reviewed by Franke et al. (111), and the limited data do not provide many valuable insights into how the fish immune system (and potentially survival) will be impacted by simultaneous exposure to these two climate change-related environmental challenges. Such studies should be a priority as average ocean temperatures are rising (8), heat waves are becoming more frequent and severe (112, 113), and some believe that the greatest challenge to fishes in the current era of climate change is not high temperatures, but hypoxia (114).
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
Ethics statement
The animal study was approved by Memorial University Institutional Animal Care Committee. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
IR: Writing – original draft, Formal analysis. MM: Data curation, Formal analysis, Methodology, Writing – original draft. FZ: Conceptualization, Investigation, Methodology, Writing – original draft. RS: Methodology, Project administration, Writing – review & editing. EC: Writing – review & editing. JH: Formal analysis, Writing – review & editing. MR: Conceptualization, Writing – review & editing. EU: Conceptualization, Writing – review & editing. AG: Conceptualization, Funding acquisition, Methodology, Project administration, Resources, Supervision, Writing – original draft.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This study was conducted as part of the project ‘Mitigating the Impact of Climate-Related Challenges on Salmon Aquaculture (MICCSA)’ and was funded by the Atlantic Canada Opportunities Agency (781–9658–205222), Innovate NL (5404–1209–104), Innovative PEI and The Huntsman Marine Science Centre.
Acknowledgments
The authors thank the staff of the Dr. Joe Brown Aquatic Research Building (JBARB, Memorial University, Canada) for assistance with fish husbandry, and Ms. Tasha Harrold for managing various aspects of the MICCSA research program.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2025.1545754/full#supplementary-material
References
1. Johansen JL, Herbert NA, Steffensen JF. The behavioural and physiological response of Atlantic cod Gadus morhua L. @ to short-term acute hypoxia. J Fish Biol. (2006) 68:1918–24. doi: 10.1111/j.1095-8649.2006.01080.x
2. Solstorm D, Oldham T, Solstorm F, Klebert P, Stien LH, Vagseth T, et al. Dissolved oxygen variability in a commercial sea-cage exposes farmed Atlantic salmon to growth limiting conditions. Aquaculture. (2018) 486:122–9. doi: 10.1016/j.aquaculture.2017.12.008
3. Oppedal F, Dempster T, Stien LH. Environmental drivers of Atlantic salmon behaviour in sea-cages: A review. Aquaculture. (2011) 311:1–18. doi: 10.1016/j.aquaculture.2010.11.020
4. Dempster T, Wright D, Oppedal F. Identifying the nature, extent and duration of critical production periods for Atlantic salmon in Macquarie Harbour, Tasmania, during summer (2016). Available online at: www.frdc.com.au.
5. Burt K, Hamoutene D, Perez-Casanova J, Kurt Gamperl A, Volkoff H. The effect of intermittent hypoxia on growth, appetite and some aspects of the immune response of Atlantic salmon (Salmo salar). Aquac Res. (2013) 45:124–37. doi: 10.1111/j.1365-2109.2012.03211.x
6. Hvas M, Folkedal O, Oppedal F. Fish welfare in offshore salmon aquaculture. Rev Aquac. (2021) 13:836–52. doi: 10.1111/raq.12501
7. Johansson D, Juell J-E, Oppedal F, Stiansen J-E, Ruohonen K. The influence of the pycnocline and cage resistance on current flow, oxygen flux and swimming behaviour of Atlantic salmon (Salmo salar L.) in production cages. Aquaculture. (2007) 265:271–87. doi: 10.1016/j.aquaculture.2006.12.047
8. IPCC, Pörtner HO, Roberts DC, Poloczanska ES, Mintenbeck K, Tignor M, Alegría A, et al. Climate change 2022: impacts, adaptation and vulnerability working group II contribution to the sixth assessment report of the intergovernmental panel on climate change. Portner H, Roberts D, Tignor M, Poloczanska E, Mintenbeck K, Alegria A, Craig M, Langsdorf S, Löschke S, Möller V, et al, editors. Cambridge, UK and New York, NY, USA: Cambridge University Press (2022). 3056 p. doi: 10.1017/9781009325844.Front
9. Breitburg D, Levin LA, Oschlies A, Grégoire M, Chavez FP, Conley DJ, et al. Declining oxygen in the global ocean and coastal waters. Sci (1979). (2018) 359. doi: 10.1126/science.aam7240
10. Burt K, Hamoutene D, Mabrouk G, Lang C, Puestow T, Drover D, et al. Environmental conditions and occurrence of hypoxia within production cages of Atlantic salmon on the south coast of Newfoundland. Aquac Res. (2012) 43:607–20. doi: 10.1111/j.1365-2109.2011.02867.x
11. Evans JJ, Shoemaker CA, Klesius PH. Effects of sublethal dissolved oxygen stress on blood glucose and susceptibility to Streptococcus agalactiae in Nile Tilapia Oreochromis niloticus. J Aquat Anim Health. (2003) 15:202–8. doi: 10.1577/H03-024
12. Gallage S, Katagiri T, Endo M, Futami K, Endo M, Maita M. Influence of moderate hypoxia on vaccine efficacy against Vibrio Anguillarum in Oreochromis niloticus (Nile tilapia). Fish Shellfish Immunol. (2016) 51:271–81. doi: 10.1016/j.fsi.2016.02.024
13. Krasnov A, Burgerhout E, Johnsen H, Tveiten H, Bakke AF, Lund H, et al. Development of Atlantic salmon (Salmo salar L.) under hypoxic conditions induced sustained changes in expression of immune genes and reduced resistance to Moritella viscosa. Front Ecol Evol. (2021) 9:722218. doi: 10.3389/fevo.2021.722218
14. Kvamme BO, Gadan K, Finne-Fridell F, Niklasson L, Sundh H, Sundell K, et al. Modulation of innate immune responses in Atlantic salmon by chronic hypoxia-induced stress. Fish Shellfish Immunol. (2013) 34:55–65. doi: 10.1016/j.fsi.2012.10.006
15. Martinez D, De Lazaro O, Cortes P, Oyarzun-Salazar R, Paschke K, Vargas-Chacoff L. Hypoxia modulates the transcriptional immunological response in Oncorhynchus kisutch. Fish Shellfish Immunol. (2020) 106:1042–51. doi: 10.1016/j.fsi.2020.09.025
16. Oldham T, Dempster T, Crosbie P, Adams M, Nowak B. Cyclic hypoxia exposure accelerates the progression of amoebic gill disease. Pathogens. (2020) 9:597. doi: 10.3390/pathogens9080597
17. Abdel-Tawwab M, Hagras AE, Elbaghdady HAM, Monier MN. Effects of dissolved oxygen and fish size on Nile tilapia, Oreochromis niloticus (L.): growth performance, whole-body composition, and innate immunity. Aquacult Int. (2015) 23:1261–74. doi: 10.1007/s10499-015-9882-y
18. Dagoudo M, Qiang J, Bao J-W, Tao Y-F, Zhu H-J, Tumukunde EM, et al. Effects of acute hypoxia stress on hemato-biochemical parameters, oxidative resistance ability, and immune responses of hybrid yellow catfish (Pelteobagrus fulvidraco × P. vachelli) juveniles. Aquacult Int. (2021) 29:2181–96. doi: 10.1007/s10499-021-00742-1
19. Polymeropoulos ET, Plouffe D, LeBlanc S, Currie S, Elliott NG, Frappell PB. Growth hormone transgenesis and polyploidy increase metabolic rate, alter the cardiorespiratory response and influence HSP expression to acute hypoxia in Atlantic salmon (Salmo salar) yolk-sac alevins. J Exp Biol. (2014) 217:2268–76. doi: 10.1242/jeb.098913
20. Qi D, Xia M, Chao Y, Zhao Y, Wu R. Identification, molecular evolution of toll-like receptors in a Tibetan schizothoracine fish (Gymnocypris eckloni) and their expression profiles in response to acute hypoxia. Fish Shellfish Immunol. (2017) 68:102–13. doi: 10.1016/j.fsi.2017.07.014
21. Soldatov AA, Golovina IV, Kolesnikova EE, Sysoeva IV, Sysoev AA, Kukhareva TA, et al. Activity of energy metabolism enzymes and ATP content in the brain and gills of the black sea scorpionfish Scorpaena porcus under short-term hypoxia. J Evol Biochem Physiol. (2020) 56:224–34. doi: 10.1134/S0022093020030059
22. Varghese T, Mishal P, Gupta G, Kumar M, Pal AK, Dasgupta S. Temporal changes in behavioural responses and serum metabolites of Cirrhinus mrigala exposed to acute hypoxia. J Environ Biol. (2019) 40:641–7. doi: 10.22438/jeb/40/4/MRN-1004
23. Wang J, Lu D-Q, Jiang B, Luo H-L, Lu G-L, Li A-X. The effect of intermittent hypoxia under different temperature on the immunomodulation in Streptococcus agalactiae vaccinated Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. (2018) 79:181–92. doi: 10.1016/j.fsi.2018.04.040
24. Wang M, Wu F, Xie S, Zhang L. Acute hypoxia and reoxygenation: Effect on oxidative stress and hypoxia signal transduction in the juvenile yellow catfish (Pelteobagrus fulvidraco). Aquaculture. (2021) 531:735903. doi: 10.1016/j.aquaculture.2020.735903
25. Wood AT, Taylor RS, Quezada-Rodriguez PR, Wynne JW. Hydrogen peroxide treatment of Atlantic salmon temporarily decreases oxygen consumption but has negligible effects on hypoxia tolerance and aerobic performance. Aquaculture. (2021) 540:736676. doi: 10.1016/j.aquaculture.2021.736676
26. Xia JH, Li HL, Li BJ, Gu XH, Lin HR. Acute hypoxia stress induced abundant differential expression genes and alternative splicing events in heart of tilapia. Gene. (2018) 639:52–61. doi: 10.1016/j.gene.2017.10.002
27. Beemelmanns A, Zanuzzo FS, Sandrelli RM, Rise ML, Gamperl AK. The Atlantic salmon’s stress- and immune-related transcriptional responses to moderate hypoxia, an incremental temperature increase, and these challenges combined. G3 Genes|Genomes|Genetics. (2021) 11. doi: 10.1093/g3journal/jkab102
28. Beemelmanns A, Zanuzzo FS, Xue X, Sandrelli RM, Rise ML, Gamperl AK. The transcriptomic responses of Atlantic salmon (Salmo salar) to high temperature stress alone, and in combination with moderate hypoxia. BMC Genomics. (2021) 22:261. doi: 10.1186/s12864-021-07464-x
29. Leeuwis RHJ, Hall JR, Zanuzzo FS, Smith N, Clow KA, Kumar S, et al. Climate change can impair bacterial pathogen defenses in sablefish via hypoxia-mediated effects on adaptive immunity. Dev Comp Immunol. (2024) 156:105161. doi: 10.1016/j.dci.2024.105161
30. Schafer N, Matoušek J, Rebl A, Stejskal V, Brunner RM, Goldammer T, et al. Effects of chronic hypoxia on the immune status of pikeperch (Sander lucioperca Linnaeus, 1758). Biol (Basel). (2021) 10:649. doi: 10.3390/biology10070649
31. Vega B, Toro-Araneda T, Alvarado JF, Cárcamo CB, Guzmán F, Acosta F, et al. Effects of hypoxia on the antibacterial activity of epidermal mucus from Chilean meagre (Cilus gilberti). Animals. (2024) 14. doi: 10.3390/ani14132014
32. Canadian Council of Animal Care. Guidelines on the care and use of fish in research, teaching and testing. Canadian Council of Animal Care, Ed. Ottawa, ON, Canada: Canadian Council of Animal Care (2005). Available at: https://ccac.ca/Documents/Standards/Guidelines/Fish.pdf.
33. Carnevale C, Roberts JC, Syme DA, Gamperl AK. Hypoxic acclimation negatively impacts the contractility of steelhead trout (Oncorhynchus mykiss) spongy myocardium. Am J Physiol Regul Integr Comp Physiol. (2020) 318:214–26. doi: 10.1152/ajpregu.00107.2019
34. Vasquez I, Cao T, Hossain A, Valderrama K, Gnanagobal H, Dang M, et al. Aeromonas salmonicida infection kinetics and protective immune response to vaccination in sablefish (Anoplopoma fimbria). Fish Shellfish Immunol. (2020) 104:557–66. doi: 10.1016/j.fsi.2020.06.005
35. Marnila P, Tiiska A, Lagerspetz K, Lilius E-M. Phagocyte activity in the frog Rana temporaria: whole blood chemiluminescence method and the effects of temperature and thermal acclimation. Comp Biochem Physiol A Physiol. (1995) 111:609–14. doi: 10.1016/0300-9629(95)00054-B
36. Nikoskelainen S. Effect of environmental temperature on rainbow trout (Oncorhynchus mykiss) innate immunity. Dev Comp Immunol. (2004) 28:581–92. doi: 10.1016/j.dci.2003.10.003
37. Zanuzzo FS, Sabioni RE, Montoya LNF, Favero G, Urbinati EC. Aloe vera enhances the innate immune response of pacu (Piaractus mesopotamicus) after transport stress and combined heat killed Aeromonas hydrophila infection. Fish Shellfish Immunol. (2017) 65:198–205. doi: 10.1016/j.fsi.2017.04.013
38. Demers NE, Bayne CJ. The immediate effects of stress on hormones and plasma lysozyme in rainbow trout. Dev Comp Immunol. (1997) 21:363–73. doi: 10.1016/S0145-305X(97)00009-8
39. Inoue T, Moritomo T, Tamura Y, Mamiya S, Fujino H, Nakanishi T. A new method for fish leucocyte counting and partial differentiation by flow cytometry. Fish Shellfish Immunol. (2002) 13:379–90. doi: 10.1006/fsim.2002.0413
40. Caballero-Solares A, Hall JR, Xue X, Eslamloo K, Taylor RG, Parrish CC, et al. The dietary replacement of marine ingredients by terrestrial animal and plant alternatives modulates the antiviral immune response of Atlantic salmon (Salmo salar). Fish Shellfish Immunol. (2017) 64:24–38. doi: 10.1016/j.fsi.2017.02.040
41. Caballero-Solares A, Umasuthan N, Xue X, Katan T, Kumar S, Westcott JD, et al. Interacting Effects of Sea Louse (Lepeophtheirus salmonis) Infection and Formalin-Killed Aeromonas salmonicida on Atlantic Salmon Skin Transcriptome. Front Immunol. (2022) 13:804987. doi: 10.3389/fimmu.2022.804987
42. Eslamloo K, Kumar S, Caballero-Solares A, Gnanagobal H, Santander J, Rise ML. Profiling the transcriptome response of Atlantic salmon head kidney to formalin-killed Renibacterium salmoninarum. Fish Shellfish Immunol. (2020) 98:937–49. doi: 10.1016/j.fsi.2019.11.057
43. Eslamloo K, Kumar S, Xue X, Parrish KS, Purcell SL, Fast MD, et al. Global gene expression responses of Atlantic salmon skin to Moritella viscosa. Sci Rep. (2022) 12:4622. doi: 10.1038/s41598-022-08341-7
44. Soto-Dávila M, Valderrama K, Inkpen SM, Hall JR, Rise ML, Santander J. Effects of vitamin D2 (ergocalciferol) and D3 (cholecalciferol) on Atlantic salmon (Salmo salar) primary macrophage immune response to Aeromonas salmonicida subsp. salmonicida infection. Front Immunol. (2020) 10:3011. doi: 10.3389/fimmu.2019.03011
45. Smith NC, Christian SL, Taylor RG, Santander J, Rise ML. Immune modulatory properties of 6-gingerol and resveratrol in Atlantic salmon macrophages. Mol Immunol. (2018) 95:10–9. doi: 10.1016/j.molimm.2018.01.004
46. Zanuzzo FS, Sandrelli RM, Peroni E de FC, Hall JR, Rise ML, Gamperl AK. Atlantic Salmon (Salmo salar) bacterial and viral innate immune responses are not impaired by florfenicol or tetracycline administration. Fish Shellfish Immunol. (2022) 123:298–313. doi: 10.1016/j.fsi.2022.02.034
47. Xue X, Hixson SM, Hori TS, Booman M, Parrish CC, Anderson DM, et al. Atlantic salmon (Salmo salar) liver transcriptome response to diets containing Camelina sativa products. Comp Biochem Physiol D Genomics Proteomics. (2015) 14:1–15. doi: 10.1016/j.cbd.2015.01.005
48. Caballero-Solares A, Xue X, Parrish CC, Foroutani MB, Taylor RG, Rise ML. Changes in the liver transcriptome of farmed Atlantic salmon (Salmo salar) fed experimental diets based on terrestrial alternatives to fish meal and fish oil. BMC Genomics. (2018) 19:796. doi: 10.1186/s12864-018-5188-6
49. Katan T, Caballero-Solares A, Taylor RG, Rise ML, Parrish CC. Effect of plant-based diets with varying ratios of ω6 to ω3 fatty acids on growth performance, tissue composition, fatty acid biosynthesis and lipid-related gene expression in Atlantic salmon (Salmo salar). Comp Biochem Physiol Part D Genomics Proteomics. (2019) 30:290–304. doi: 10.1016/j.cbd.2019.03.004
50. Xu Q, Feng CY, Hori TS, Plouffe DA, Buchanan JT, Rise ML. Family-specific differences in growth rate and hepatic gene expression in juvenile triploid growth hormone (GH) transgenic Atlantic salmon (Salmo salar). Comp Biochem Physiol D Genomics Proteomics. (2013) 8:317–33. doi: 10.1016/j.cbd.2013.09.002
51. Vandesompele J, De Preter K, Pattyn F, Poppe B, Van Roy N, De Paepe A, et al. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. (2002) 3:research0034.1. doi: 10.1186/gb-2002-3-7-research0034
52. Sambrook J, Russell W. Molecular cloning: a laboratory manual. 3rd ed. NY: Cold Spring Harbor Press (2001).
53. Dinno A. Dunn’s test of multiple comparisons using rank sums v. 1.3.6 (2024). Available online at: https://cran.r-project.org/web/packages/dunn.test/dunn.test.pdf (Accessed Oct 30, 2024).
54. De Mendiburu F. R package “agricolae”: Statistical Procedures for Agricultural Research (2023). Available online at: https://cran.r-project.org/web/packages/agricolae/agricolae.pdf (Accessed Oct 30, 2024).
55. Ogle DH, Doll J, Wheeler A, Dinno A. “FSA” v. 0.9.5 (2023). Available online at: https://cran.r-project.org/web/packages/FSA/FSA.pdf (Accessed Oct 30, 2024).
56. Tort L. Stress and immune modulation in fish. Dev Comp Immunol. (2011) 35:1366–75. doi: 10.1016/j.dci.2011.07.002
57. Uribe C, Folch H, Enriquez R, Moran G. Innate and adaptive immunity in teleost fish: a review. Vet Med. (2011) 56:486–503. doi: 10.17221/3294-VETMED
58. Zanuzzo FS, Beemelmanns A, Hall JR, Rise ML, Gamperl AK. The innate immune response of Atlantic salmon (Salmo salar) is not negatively affected by high temperature and moderate hypoxia. Front Immunol. (2020) 11:1009. doi: 10.3389/fimmu.2020.01009
59. Rojas I, Cárcamo CB, Defranchi Y, Jeno K, Rengel J, Araya M, et al. A diet rich in HUFAs enhances the energetic and immune response capacities of larvae of the scallop Argopecten purpuratus. Animals. (2023) 13. doi: 10.3390/ani13081416
60. Mu Y, Li W, Wu B, Chen J, Chen X. Transcriptome analysis reveals new insights into immune response to hypoxia challenge of large yellow croaker (Larimichthys crocea). Fish Shellfish Immunol. (2020) 98:738–47. doi: 10.1016/j.fsi.2019.11.021
61. Ni M, Wen H, Li J, Chi M, Bu Y, Ren Y, et al. The physiological performance and immune responses of juvenile Amur sturgeon (Acipenser schrenckii) to stocking density and hypoxia stress. Fish Shellfish Immunol. (2014) 36:325–35. doi: 10.1016/j.fsi.2013.12.002
62. Biller JD, Takahashi LS. Oxidative stress and fish immune system: Phagocytosis and leukocyte respiratory burst activity. Acad Bras Cienc. (2018) 90:3403–14. doi: 10.1590/0001-3765201820170730
63. Zhao Y, Jiang X, Kong X, Di G, Nie G, Li X. Effects of hypoxia on lysozyme activity and antioxidant defences in the kidney and spleen of Carassius auratus. Aquac Res. (2017) 48:223–35. doi: 10.1111/are.12876
64. Maule AG, Schreck CB. Changes in numbers of leukocytes in immune organs of juvenile Coho salmon after acute stress or cortisol treatment. J Aquat Anim Health. (1990) 2:298–304. doi: 10.1577/1548-8667(1990)002<0298:CINOLI>2.3.CO;2
65. Pickering AD, Pottinger TG. Cortisol can increase the susceptibility of brown trout, Salmo trutta L., to disease without reducing the white blood cell count. J Fish Biol. (1985) 27:611–9. doi: 10.1111/j.1095-8649.1985.tb03206.x
66. Nayak S, Portugal I, Zilberg D. Analyzing complement activity in the serum and body homogenates of different fish species, using rabbit and sheep red blood cells. Vet Immunol Immunopathol. (2018) 199:39–42. doi: 10.1016/j.vetimm.2018.03.008
67. Magnoni LJ, Novais SC, Eding E, Leguen I, Lemos MFL, Ozório ROA, et al. Acute stress and an electrolyte- imbalanced diet, but not chronic hypoxia, increase oxidative stress and hamper innate immune status in a rainbow trout (Oncorhynchus mykiss) isogenic line. Front Physiol. (2019) 10:453. doi: 10.3389/fphys.2019.00453
68. Merle NS, Church SE, Fremeaux-Bacchi V, Roumenina LT. Complement system part I - molecular mechanisms of activation and regulation. Front Immunol. (2015) 6:262. doi: 10.3389/fimmu.2015.00262
69. Wang Z, Zhang S, Tong Z, Li L, Wang G. Maternal transfer and protective role of the alternative complement components in zebrafish Danio rerio. PloS One. (2009) 4. doi: 10.1371/journal.pone.0004498
70. Wu Z, You F, Wen A, Ma D, Zhang P. Physiological and morphological effects of severe hypoxia, hypoxia and hyperoxia in juvenile turbot (Scophthalmus maximus L.). Aquac Res. (2016) 47:219–27. doi: 10.1111/are.12483
71. González R, Muñoz K, Brokordt K, Schmitt P. Scallop immunology. In: Reference Module in Life Sciences. Amsterdan, Netherlands: Elsevier (2019). doi: 10.1016/b978-0-12-809633-8.20896-0
72. Kuhlicke J, Frick JS, Morote-Garcia JC, Rosenberger P, Eltzschig HK. Hypoxia inducible factor (HIF)-1 coordinates induction of toll-like receptors TLR2 and TLR6 during hypoxia. PloS One. (2007) 2. doi: 10.1371/journal.pone.0001364
73. Yu L, Wang L, Chen S. Endogenous toll-like receptor ligands and their biological significance. J Cell Mol Med. (2010) 14:2592–603. doi: 10.1111/j.1582-4934.2010.01127.x
74. Basu M, Paichha M, Lenka SS, Chakrabarty R, Samanta M. Hypoxic stress: impact on the modulation of TLR2, TLR4, NOD1 and NOD2 receptor and their down-stream signalling genes expression in catla (Catla catla). Mol Biol Rep. (2016) 43:1–9. doi: 10.1007/s11033-015-3932-4
75. Xiao K, Wang X, Wang M, Guo H-X, Liu W-B, Jiang G-Z. Metabolism, antioxidant and immunity in acute and chronic hypoxic stress and the improving effect of vitamin C in the channel catfish (Ictalurus punctatus). Fish Physiol Biochem. (2024) 50:183–96. doi: 10.1007/s10695-023-01205-5
76. Laing KJ, Zou JJ, Wang T, Bols N, Hirono I, Aoki T, et al. Identification and analysis of an interleukin 8-like molecule in rainbow trout Oncorhynchus mykiss. Dev Comp Immunol. (2002) 26:433–44. doi: 10.1016/S0145-305X(01)00092-1
77. Haugland GT, Rønneseth A, Gundersen L, Lunde HS, Nordland K, Wergeland HI. Neutrophils in Atlantic salmon (Salmo salar L.) are MHC class II+ and secret IL-12p40 upon bacterial exposure. Aquac Fish. (2024) 9:144–53. doi: 10.1016/j.aaf.2022.07.002
78. Mukaida N, Harada A, Matsushima K. Interleukin-8 (IL-8) and monocyte chemotactic and activating factor (MCAF/MCP-1), chemokines essentially involved in inflammatory and immune reactions. Cytokine Growth Factor Rev. (1998) 9:9–23. doi: 10.1016/S1359-6101(97)00022-1
79. Leeuwis RHJ, Nash GW, Sandrelli RM, Zanuzzo FS, Gamperl AK. The environmental tolerances and metabolic physiology of sablefish (Anoplopoma fimbria). Comp Biochem Physiol A Mol Integr Physiol. (2019) 231:140–8. doi: 10.1016/j.cbpa.2019.02.004
80. Korbecki J, Kojder K, Barczak K, Simińska D, Gutowska I, Chlubek D, et al. Hypoxia alters the expression of CC chemokines and CC chemokine receptors in a tumor–a literature review. Int J Mol Sci. (2020) 21:1–32. doi: 10.3390/ijms21165647
81. Fu Q, Yang Y, Li C, Zeng Q, Zhou T, Li N, et al. The chemokinome superfamily: II. The 64 CC chemokines in channel catfish and their involvement in disease and hypoxia responses. Dev Comp Immunol. (2017) 73:97–108. doi: 10.1016/j.dci.2017.03.012
82. Bosco MC, Puppo M, Santangelo C, Anfosso L, Pfeffer U, Fardin P, et al. Hypoxia modifies the transcriptome of primary human monocytes: modulation of novel immune-related genes and identification of CC-chemokine ligand 20 as a new hypoxia-inducible gene. J Immunol. (2006) 177:1941–55. doi: 10.4049/jimmunol.177.3.1941
83. Qiao D, Zhao Y, Pei C, Zhao X, Jiang X, Zhu L, et al. Two Cc CCL19b’s orchestrate an antibacterial immune response in Yellow River carp (Cyprinus carpio haematopterus). Fish Shellfish Immunol. (2023) 140. doi: 10.1016/j.fsi.2023.108987
84. Xu Z, Zhu C, Chen C, Zong Y, Feng H, Liu D, et al. CCL19 suppresses angiogenesis through promoting miR-206 and inhibiting Met/ERK/Elk-1/HIF-1α/VEGF-A pathway in colorectal cancer. Cell Death Dis. (2018) 9. doi: 10.1038/s41419-018-1010-2
85. Bridle A, Nosworthy E, Polinski M, Nowak B. Evidence of an antimicrobial-immunomodulatory role of Atlantic salmon cathelicidins during infection with Yersinia ruckeri. PloS One. (2011) 6. doi: 10.1371/journal.pone.0023417
86. Solstad T, Larsen AN, Seppola M, Jørgensen T. Identification, cloning and expression analysis of a hepcidin cDNA of the Atlantic cod (Gadus morhua L.). Fish Shellfish Immunol. (2008) 25:298–310. doi: 10.1016/j.fsi.2008.05.013
87. Álvarez CA, Santana PA, Salinas-Parra N, Beltrán D, Guzmán F, Vega B, et al. Immune modulation ability of Hepcidin from teleost fish. Animals. (2022) 12. doi: 10.3390/ani12121586
88. Chang CI, Zhang YA, Zou J, Nie P, Secombes CJ. Two cathelicidin genes are present in both rainbow trout (Oncorhynchus mykiss) and Atlantic salmon (Salmo salar). Antimicrob Agents Chemother. (2006) 50:185–95. doi: 10.1128/AAC.50.1.185-195.2006
89. Seppola M, Johnsen H, Mennen S, Myrnes B, Tveiten H. Maternal transfer and transcriptional onset of immune genes during ontogenesis in Atlantic cod. Dev Comp Immunol. (2009) 33:1205–11. doi: 10.1016/j.dci.2009.06.013
90. Liu Q, Davidoff O, Niss K, Haase VH. Hypoxia-inducible factor regulates hepcidin via erythropoietin-induced erythropoiesis. J Clin Invest. (2012) 122:4635–44. doi: 10.1172/JCI63924
91. Kiers D, Van Eijk LT, van der Hoeven JG, Swinkels DW, Pickkers P, Kox M. Hypoxia attenuates inflammation-induced hepcidin synthesis during experimental human endotoxemia. Haematologica. (2019) 104:e230–2. doi: 10.3324/haematol.2018.202796
92. Lee FS. At the crossroads of oxygen and iron sensing: hepcidin control of HIF-2α. J Clin Invest. (2019) 129:72–74. doi: 10.1172/JCI125509
93. Bei Y, Pan LL, Zhou Q, Zhao C, Xie Y, Wu C, et al. Cathelicidin-related antimicrobial peptide protects against myocardial ischemia/reperfusion injury. BMC Med. (2019) 17. doi: 10.1186/s12916-019-1268-y
94. Li CH, Lu XJ, Li MY, Chen J. Cathelicidin modulates the function of monocytes/macrophages via the P2X7 receptor in a teleost, Plecoglossus altivelis. Fish Shellfish Immunol. (2015) 47:878–85. doi: 10.1016/j.fsi.2015.10.031
95. Liu J, Xue W, Xiang H, Zheng J, Zhao Y, Jiao L, et al. Cathelicidin PR-39 peptide inhibits hypoxia/reperfusion-induced kidney cell apoptosis by suppression of the endoplasmic reticulum-stress pathway. Acta Biochim Biophys Sin (Shanghai). (2016) 48:714–22. doi: 10.1093/abbs/gmw061
96. Wu J, Parungo C, Wu G, Kang PM, Laham RJ, Sellke FW, et al. PR39 inhibits apoptosis in hypoxic endothelial cells: role of inhibitor apoptosis protein-2. Circulation. (2004) 109:1660–7. doi: 10.1161/01.CIR.0000124067.35915.E0
97. Santana PA, Guzmán F, Forero JC, Luna OF, Mercado L. Hepcidin, Cathelicidin-1 and IL-8 as immunological markers of responsiveness in early developmental stages of rainbow trout. Dev Comp Immunol. (2016) 62:48–57. doi: 10.1016/j.dci.2016.04.014
98. Hordvik I. Immunoglobulin isotypes in Atlantic salmon, Salmo salar. Biomolecules. (2015) 5:166–77. doi: 10.3390/biom5010166
99. Magnadóttir B. Comparison of immunoglobulin (IgM) from four fish species. Ice Agr Sci. (1998) 12:47–59.
100. Sanchez C, Babin M, Tomillo J, Ubeira FM, Dominguez J. Quantification of low levels of rainbow trout immunoglobulin by enzyme immunoassay using two monoclonal antibodies. Vet Immunol Immunopathol. (1993) 36:65–74. doi: 10.1016/0165-2427(93)90006-p
101. Magnadottir B, Gudmundsdottir BK. A comparison of total and specific immunoglobulin levels in healthy Atlantic salmon (Salmo salar L.) and in salmon naturally infected with Aeromonas salmonicida subsp. achromogenes. Vet Immunol Immunopathol. (1992) 32:179–89. doi: 10.1016/0165-2427(92)90078-5
102. Parra D, Takizawa F, Sunyer JO. Evolution of B cell immunity. Annu Rev Anim Biosci. (2013) 1:65–97. doi: 10.1146/annurev-animal-031412-103651
103. Tetens V, Lykkeboe G. Acute exposure of rainbow trout to mild and deep hypoxia: O2 affinity and O2 capacitance of arterial blood. Respir Physiol. (1985) 61:221–35. doi: 10.1016/0034-5687(85)90128-8
104. Affonso EG, Polez VLP, Corrêa CF, Mazon AF, Araújo MRR, Moraes G, et al. Blood parameters and metabolites in the teleost fish Colossoma macropomum exposed to sulfide or hypoxia. Comp Biochem Physiol C Toxicol Pharmacol. (2002) 133:375–82. doi: 10.1016/S1532-0456(02)00127-8
105. Zanuzzo FS, Urbinati EC, Nash GW, Gamperl AK. Steelhead trout Oncorhynchus mykiss metabolic rate is affected by dietary Aloe vera inclusion but not by mounting an immune response against formalin-killed Aeromonas salmonicida. J Fish Biol. (2015) 87:43–53. doi: 10.1111/jfb.12690
106. Bennoit NR, Craig PM. Increased metabolic rate associated with immune stimulation of heat-killed Vibrio Anguillarum at different temperatures in zebrafish (Danio rerio). Comp Biochem Physiol B Biochem Mol Biol. (2020) 250. doi: 10.1016/j.cbpb.2020.110489
107. Douxfils J, Fierro-Castro C, Mandiki SNM, Emile W, Tort L, Kestemont P. Dietary β-glucans differentially modulate immune and stress-related gene expression in lymphoid organs from healthy and Aeromonas hydrophila-infected rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. (2017) 63:285–96. doi: 10.1016/j.fsi.2017.02.027
108. Levet M, Roche DG, Killen SS, Colosio S, Bshary R, Miest JJ, et al. Energetic costs of mounting an immune response in a coral reef damselfish (Pomacentrus amboinensis). Can J Zool. (2024) 102:380–92. doi: 10.1139/cjz-2023-0033
109. Gallage S, Katagiri T, Endo M, Maita M. Comprehensive evaluation of immunomodulation by moderate hypoxia in S. agalactiae vaccinated Nile tilapia. Fish Shellfish Immunol. (2017) 66:445–54. doi: 10.1016/j.fsi.2017.05.041
110. Xu S, Wang Y, Gao C, Babu VS, Li J, Liu Q, et al. Effects of dissolved oxygen on intestinal bacterial community and immunity of Atlantic salmon Salmo salar. J Oceanol Limnol. (2023) 41:364–75. doi: 10.1007/s00343-021-1336-y
111. Franke A, Beemelmanns A, Miest JJ. Are fish immunocompetent enough to face climate change? Biol Lett. (2024) 20. doi: 10.1098/rsbl.2023.0346
112. Oliver ECJ, Benthuysen JA, Darmaraki S, Donat MG, Hobday AJ, Holbrook NJ, et al. Marine heatwaves. Ann Rev Mar Sci. (2021) 13:313–42. doi: 10.1146/annurev-marine-032720-095144
113. Frölicher TL, Fischer EM, Gruber N. Marine heatwaves under global warming. Nature. (2018) 560:360–4. doi: 10.1038/s41586-018-0383-9
Keywords: hypoxia, immunity, climate change, aquaculture, Aeromonas salmonicida
Citation: Rojas I, de Mello MMM, Zanuzzo FS, Sandrelli RM, Peroni EdFC, Hall JR, Rise ML, Urbinati EC and Gamperl AK (2025) Chronic hypoxia has differential effects on constitutive and antigen-stimulated immune function in Atlantic salmon (Salmo salar). Front. Immunol. 16:1545754. doi: 10.3389/fimmu.2025.1545754
Received: 15 December 2024; Accepted: 03 February 2025;
Published: 19 February 2025.
Edited by:
Jiong Chen, Ningbo University, ChinaReviewed by:
Yinnan Mu, Fujian Agriculture and Forestry University, ChinaTor Gjøen, University of Oslo, Norway
Copyright © 2025 Rojas, de Mello, Zanuzzo, Sandrelli, Peroni, Hall, Rise, Urbinati and Gamperl. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Isis Rojas, aXJvamFzQG11bi5jYQ==; Anthony K. Gamperl, a2dhbXBlcmxAbXVuLmNh
†Present addresses: Fábio S. Zanuzzo, Onda, Victoria, PE, Canada
Ellen de Fátima C. Peroni, Onda, Victoria, PE, Canada