 Steven Schepanski
Steven Schepanski Gonza B. Ngoumou
Gonza B. Ngoumou Claudia Buss
Claudia Buss Georg Seifert
Georg Seifert
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Immunol. , 29 January 2025
Sec. Immunological Tolerance and Regulation
Volume 16 - 2025 | https://doi.org/10.3389/fimmu.2025.1538920
This article is part of the Research Topic Maternal-fetal-placental Immune Interactions: Implications for Pregnancy Outcomes and Long-term Health View all 16 articles
This review evaluates in-vitro models for studying how maternal influences during pregnancy impact the development of offspring microglia, the immune cells of the central nervous system. The models examined include primary microglia cultures, microglia cell lines, iPSC-derived microglia, PBMC-induced microglia-like cells, 3D brain organoids derived from iPSCs, and Hofbauer cells. Each model is assessed for its ability to replicate the in-vivo environment of the developing brain, with a focus on their strengths, limitations, and practical challenges. Key factors such as scalability, genetic and epigenetic fidelity, and physiological relevance are highlighted. Microglia cell lines are highly scalable but lack genetic and epigenetic fidelity. iPSC-derived microglia provide moderate physiological relevance and patient-specific genetic insights but face operational and epigenetic challenges inherent to reprogramming. 3D brain organoids, derived from iPSCs, offer an advanced platform for studying complex neurodevelopmental processes but require extensive resources and technical expertise. Hofbauer cells, which are fetal macrophages located in the placenta and share a common developmental origin with microglia, are uniquely exposed to prenatal maternal factors and, depending on fetal barrier maturation, exhibit variable epigenetic fidelity. This makes them particularly useful for exploring the impact of maternal influences on fetal programming of microglial development. The review concludes that no single model comprehensively captures all aspects of maternal influences on microglial development, but it offers guidance on selecting the most appropriate model based on specific research objectives and experimental constraints.
The concept of fetal programming refers to the idea that changes within the intrauterine environment can contribute to the offspring’s vulnerability to developing diseases later in life by shaping their responses to future internal and external stimuli (1, 2). Various maternal challenges such as stress (3, 4), infection (5, 6), and exposure to environmental pollutants (7, 8) can alter gestational biology and thereby reshaping the intrauterine environment. These alterations, especially during critical windows of fetal development, have been associated with an increased risk of neurodevelopmental disorders and other long-term health consequences in the offspring (9–11).
Microglia, the resident macrophages of the central nervous system (CNS), have garnered increasing interest as key mediators of neurodevelopmental processes during these sensitive periods in the womb and afterwards. In the developing brain, microglia exhibit a high degree of responsiveness to extracellular stimuli (12, 13), which enables them to shape neural circuits through mechanisms such as synaptic pruning (14), clearing of apoptotic cells (15–17), and supporting neurogenesis (18). Originating from yolk sac progenitor cells, microglial cells colonize the CNS as early as the fourth week of gestation (9, 19, 20) and continue to perform essential functions throughout life. In the mature brain, microglia are maintained through self-renewal over the entire lifespan (21) and are essential for homeostasis, neuroplasticity (22), and responses to pathogens (23). Beyond that, microglia are implicated in cognitive functions (24, 25), underscoring their dual role in supporting healthy brain function and contributing to neuropathology when dysregulated.
Research, primary from animal models, has demonstrated that maternal-derived factors, such as elevated maternal glucocorticoids (26, 27), cytokines (28, 29) or immune cells (30), and exposure to environmental pollutants such as diesel exhaust particles (31, 32), can interfere with fetal microglial development. These maternal factors can affect fetal microglia development either through being directly transferred across the placenta or by triggering placental responses that alter the intrauterine environment. Such environmental changes have been associated with dysregulated microglial reactivity, manifesting as either heightened sensitivity with excessive synaptic pruning or diminished responsiveness resulting in less pruning (31, 33–36). However, recent evidence suggests that microglia may not be universally required for experience-dependent neural circuit maturation (37). Given microglia’s central role in neural circuit formation, disruptions in microglia function due to these prenatal maternal factors have been associated with an increased risk of neurodevelopmental disorders in the offspring.
While findings from animal models have substantially expanded our understanding of how intrauterine conditions shape fetal microglial development, translating these findings to humans remains challenging (38). Differences in the biology of murine and human microglia (39), along with the lack of animal models for certain human CNS disorders, limit the applicability of these insights. Additionally, restricted access to human fetal brain tissue hinders direct investigation at cellular and molecular levels. As a result, in-vitro models have become essential tools for exploring the mechanisms of neurodevelopment.
This review critically evaluates the suitability of various in-vitro models for studying how variation in maternal-derived intrauterine factors affects fetal brain development, assessing their ability to replicate key physiological, genetic, and epigenetic conditions relevant to human development, and discussing their potential for translating findings from animal research into meaningful insights for human studies.
Primary microglia cultures are derived directly from brain tissue, typically sourced from rodent embryos, neonates, or, less commonly, human fetal postmortem tissue (40, 41). The isolation process involves dissecting specific brain regions, dissociating tissues, and purifying microglia using techniques such as density gradient centrifugation, magnetic-activated cell sorting (MACS) (42), fluorescence-activated cell sorting (FACS) (43, 44), or orbital shaking (45). Because these cells are promptly isolated from fresh tissue sources, they retain essential microglial phenotypes and immune functions, allowing for physiological relevance in culture. This preservation of immune characteristics and cellular integrity makes primary microglia valuable for studying baseline microglial responses to various stimuli in a controlled environment.
Despite their physiological relevance, primary microglia cultures face limitations when used to model prenatal in-utero conditions. One major challenge is the impact of postmortem changes in brain cell structure. Studies (46–48) on postmortem brain tissue have shown that cell morphometry can be significantly altered depending on the postmortem interval. Cell death through oncotic necrosis, resulting from adenosine triphosphate depletion, leads to cell swelling, vacuolization, and loss of membrane integrity (46, 49). These changes can impair critical cellular functions, such as metabolic activity or signaling responsiveness (50), reducing the physiological relevance of isolated microglia for primary cultures. Additionally, fluid shifts and vacuolization during the postmortem interval can distort brain tissue structure (46), potentially inducing cellular stress responses that affect microglial behavior in-vitro.
In addition to degradation concerns, variability in prenatal exposures, such as maternal stress or infection history, introduces heterogeneity in microglial immune responsiveness and phenotypic characteristics, complicating reproducibility when facing limited availability of human fetal tissue. This further restricts the ability to obtain appropriately powered sample sizes and limits scalability for high-throughput studies. Moreover, practical and ethical constraints, alongside the labor-intensive nature of microglial isolation and culture, make primary microglia cultures less feasible for large-scale research.
In summary, while primary microglia cultures maintain physiological relevance and can capture certain baseline features of prenatal exposures, challenges related to postmortem cellular degradation, variability in intrauterine exposure, limited scalability, and ethical constraints reduce their suitability for comprehensively modeling mechanisms of fetal brain development.
Microglia cell lines are widely used in neuroimmunological research due to their ease of handling, reproducibility, and scalability. These cell lines are typically derived from primary microglia obtained from brain or spinal cord tissues and are immortalized using viral transduction with oncogenes (51, 52) to ensure continuous proliferation. While this immortalization process allows for large-scale studies and consistent experimental outcomes, it introduces significant limitations that hinder their ability to accurately model the physiological, genetic, and epigenetic conditions necessary for understanding mechanisms of fetal brain development.
From a physiological perspective, microglia cell lines retain some core microglial functions, including adenosine triphosphate responsiveness, expression of macrophage/microglia marker, and basic phagocytic abilities (52). However, critical differences have been reported between primary microglia and cell lines in their responses to inflammatory stimuli (53–55). For example, the HMO6 cell line, developed by transducing embryonic microglia from telencephalon with a v-myc retroviral vector, showed limited ability to secrete a diverse range of inflammatory proteins upon exposure to lipopolysaccharide (LPS) or amyloid-β (52). Specifically, HMO6 cells showed limited secretion of interleukin (IL)-1β, IL-6, and macrophage inflammatory protein-1 α (MIP-α) compared to primary microglia (52). These functional deficits, likely attributable to the immortalization process, limit the utility of microglia cell lines for modeling the dynamic immune responses in the developing brain. Other cell lines, such as HMC3 (51) and C13NJ (56), are derived from the CHME-5 lineage, which has been suggested to originate from non-human sources, raising concerns about their validity for human-centered research (57).
In addition to physiological differences, the immortalization process may fundamentally alter the genetic and epigenetic landscape of microglia cell lines. Studies on cancer cell lines expressing SV40 T-antigen and oncogenic H-RAS demonstrate significant genetic and epigenetic changes over time, including disrupted tumor suppressor pathways, de novo DNA methylation at gene promoters, and transcriptional reprogramming (58). These changes lead to silencing of differentiation-associated genes, activation of cancer-associated signaling pathways, and acquisition of abnormal growth properties. While these findings are specific to cancer cells, the reliance on similar oncogenic transformation processes suggests that microglial cell lines may exhibit comparable alterations, including disrupted chromatin landscapes and gene expression, further reducing their suitability.
Such limitations compromise the utility of microglial cell lines for studying mechanisms of fetal programming, where precise epigenetic regulation is critical for understanding microglial roles in the developing brain. These shortcomings underscore the need for careful interpretation of results and the importance of combining microglia cell lines with other models, such as primary cultures or in-vivo systems, to better capture the complexity of prenatal brain development.
Stem cell technology offers promising opportunities for generating microglial cells in large quantities for in-vitro research. Microglia can be derived from embryonic stem cells (ESCs), harvested from the inner cell mass of a blastocyst, or induced pluripotent stem cells (iPSCs), reprogrammed from somatic cells (59, 60). Differentiating ESCs or iPSCs into microglia is a multi-step process, completed in around 30 days and often using co-culture systems with astrocytes to support microglial maturation (61, 62). However, stem cell-derived microglia often display immature phenotypes, limiting their physiological relevance for modeling prenatal environments. This is of particular importance because it has been previously shown that fetal microglia exposed to maternal inflammation, in a model of maternal immune activation, undergo accelerated maturation (63), a process that in this case stem cell-derived models may not fully replicate.
Further, the reprogramming and differentiation steps alter the cells’ epigenetic landscape (64, 65), which may impact their ability to accurately model in-vivo conditions. This poses a significant challenge for research on fetal programming, where maternal health and environmental exposures are associated with epigenetic modifications that affect long-term brain development. Consequently, stem cell-derived microglia may fail to capture these specific epigenetic changes, thereby limiting their suitability.
Despite these limitations, iPSC-derived microglia offer significant advantages, including patient-specific genetic backgrounds and precise experimental control. These models enable detailed studies of how genetic mutations, such as those associated with schizophrenia, influence microglial behavior, providing insights into neurodevelopmental disorders (66–68). However, operational challenges such as differentiation protocols are labor-intensive, costly, and require specialized conditions, limiting their scalability for high-throughput studies (60).
In summary, while iPSC-derived microglia are valuable tools for investigating molecular pathways and genetic variations, their occasional immature phenotypes, altered epigenetic profiles, and operational challenges limit their use in modeling the complex effects of maternal health and prenatal environmental factors on fetal brain development.
Three-dimensional (3D) brain organoids, developed from iPSCs, represent a major advancement in modeling human brain development and disease (69). These self-organizing structures mimic key features of the human fetal brain, including cytoarchitecture, cellular diversity, and gene expression profiles, resembling the first and second trimesters of development (70–75). Organoids can generate region-specific structures, such as the forebrain, hippocampus, and midbrain, and advances in fused assembloid systems allow researchers to model connectivity between brain regions, an essential feature for studying neurodevelopmental disorders (76–80).
A major strength of 3D brain organoids is their high physiological relevance. Their spatial organization enables the investigation of neurodevelopmental processes, such as cell-cell interactions and responses to environmental influences, in a more lifelike setting. Incorporating iPSC-derived microglia enhances their complexity, allowing for the study of microglial functions, including synaptic pruning and neuroinflammation, within a neural context (81). This makes organoids particularly valuable for exploring how intrauterine environmental factors, like maternal stress, shape brain development.
However, 3D brain organoids face significant limitations. Generating them is technically demanding and resource-intensive, and there is considerable variability in size, cell composition, and maturation between batches, making standardization and reproducibility challenging (72, 82). Additionally, the lack of systemic features like vascularization limits their capacity to sustain long-term growth and fully replicate in-vivo conditions. Additionally, while organoids inherit the genetic background of donor iPSCs, the reprogramming process alters their epigenetic landscape (83, 84). This reduced epigenetic fidelity poses challenges for studying maternal influences that rely on specific epigenetic modifications, such as those induced by maternal stress or inflammation, which are critical to understanding mechanisms of fetal programming.
In summary, 3D brain organoids offer a high degree of physiological relevance and are powerful tools for modeling neurodevelopmental processes. However, their moderate epigenetic fidelity, technical complexity, and inability to replicate systemic features necessitate cautious interpretation of findings. To address these limitations, organoids should be used alongside other in-vitro or in-vivo models for a more comprehensive investigation of how maternal health influences the developing brain of the fetus.
Peripheral blood mononuclear cell (PBMC)-induced microglia-like cells offer an accessible and scalable alternative for studying CNS-resident microglia. Derived from myeloid cells in PBMCs or cord blood mononuclear cells (CBMCs), these cells are differentiated into microglia-like cells within 10–14 days using growth factors such as macrophage colony-stimulating factor (M-CSF), granulocyte-macrophage colony-stimulating factor (GM-CSF), and IL-34 (85–89). This method is advantageous due to the ease of obtaining PBMCs through non-invasive blood draws and the availability of CBMCs collected at birth (90).
While PBMC-induced microglia-like cells express key microglial markers such as IBA1, CX3CR1, PU.1, P2RY12, and TMEM119, their hematopoietic origin fundamentally differs from CNS-resident microglia, which derive from yolk sac progenitors during embryogenesis (68, 90). This difference in lineage results in functional distinctions, as PBMC-derived cells resemble peripheral macrophages more closely than microglia, limiting their ability to model the unique roles of CNS microglia in prenatal brain development (68, 91, 92). Additionally, the epigenetic landscape of PBMC-derived cells is not CNS-specific. While cord blood-derived monocytes may retain some prenatal epigenetic marks, they likely reflect only the influence of maternal factors late in gestation and thus fail to capture the dynamic exposures and modifications occurring over time. This limits their relevance for studying the epigenetic mechanisms underlying fetal programming.
Despite these limitations, PBMC-induced microglia-like cells are highly practical for high-throughput studies due to their rapid and cost-effective generation. Their simple and scalable differentiation protocols make them accessible to labs with limited resources. However, variability in differentiation conditions can impact reproducibility, and the lack of interaction with other brain cell types, such as neurons and astrocytes, reduces their physiological relevance (93). While efforts to address these limitations, such as using simplified co-culture models to study synaptic interactions, have shown promise (68, 94), these cells fall short of comprehensively model neuroimmune interactions in the developing brain.
In summary, PBMC-induced microglia-like cells are valuable for basic microglial studies and large-scale investigations due to their accessibility and rapid generation. However, their distinct developmental origin and limited physiological and epigenetic fidelity restrict their application in studying maternal health impacts and fetal brain development. For a more complete understanding of prenatal programming, these cells should be used in combination with complementary models like iPSC-derived microglia or organoids.
Hofbauer cells (HBCs), fetal macrophages originating from yolk sac progenitors, populate the chorionic villi of the placenta throughout pregnancy and share a developmental origin with CNS-resident microglia (95–97). Both cell types arise from the same progenitor lineage but migrate to distinct tissues, where they perform specialized immune functions. In the placenta, HBCs regulate immune responses, combat infections, and contribute to tissue remodeling, roles analogous to microglia in the developing brain (98, 99). This developmental and functional overlap makes HBCs a potential surrogate for studying mechanisms of fetal programming in humans.
One of the primary strengths of HBCs is their direct and continuous exposure to maternal conditions throughout gestation, allowing them to reflect intrauterine changes from the earliest stages of development. However, maternal factors not crossing placental or fetal barriers may limit HBCs’ fidelity. Previous research (95, 100) using differential gene expression (DEG) and Gene Ontology (GO) enrichment analyses has demonstrated significant alterations in HBC gene expression under maternal inflammatory conditions during pregnancy, affecting pathways related to immune signaling, metabolism, and cellular stress. Canonical pathway analyses have further shown shared responses between HBCs and fetal microglia, such as changes in glycolysis, oxidative stress responses, and inflammatory signaling under maternal obesity (95). These parallels highlight the potential of HBCs as proxies for investigating how maternal influences on fetal development.
Despite shared responses, important differences arise due to the distinct tissue environments in which HBCs and microglia reside. Microglia interact closely with neurons and CNS-specific cell types, contributing to neurodevelopmental processes like synaptic pruning and neural circuit refinement. In contrast, HBCs are influenced by placental functions, such as nutrient exchange, hormone production, and vascular development. For example, GO analyses reveal that mitochondrial metabolism and regulation of body fluid levels are uniquely enriched in HBCs, whereas microglia show enrichment in pathways related to neuron regulation and microtubule polymerization (95). These differences illustrate the tissue-specific adaptations of these macrophages, even under shared maternal health conditions.
HBCs are particularly well-suited for studying the effects of pre-conceptional maternal health, such as obesity, on fetal immune programming, as they are exposed to maternal factors from the earliest stages of development. However, they are less informative for maternal influences arising exclusively during pregnancy, as key developmental events, such as progenitor migration to the brain or placenta and blood-brain-barrier formation may have already occurred. This temporal limitation underscores the differences between HBC and fetal microglial responses to maternal health.
Practical challenges also restrict the broader applicability of HBCs. Their collection requires timely access to placental tissue at delivery, limiting scalability and accessibility. Furthermore, variability among placental samples, driven by maternal health factors such as infection and diet, complicates reproducibility and standardization. This heterogeneity poses challenges for studies requiring uniform cell populations or modeling precise biological processes.
In summary, Hofbauer cells are a valuable model for investigating how maternal health influences fetal brain development, particularly through mechanisms of immune programming shaped by pre-conceptional factors. Their high physiological relevance and potential to reflect early prenatal exposures make them well-suited for long-term developmental studies. However, limitations in scalability, reproducibility, and their ability to capture pregnancy-specific influences highlight the need for complementary models to achieve a comprehensive understanding of prenatal programming and its implications for fetal brain and immune development.
This review critically evaluates the suitability of various in-vitro models for studying the effects of maternal health conditions on fetal brain development, with a focus on their physiological relevance, genetic and epigenetic fidelity, and practical considerations such as scalability, reproducibility, and cost. Each model offers unique advantages and limitations, and none fully replicates the complex interactions between mother and child during pregnancy. Therefore, the choice of model should be guided by the specific research aims, the desired level of biological fidelity, and the available resources.
Table 1 provides a comparative summary of these models, helping researchers select the most appropriate model for their study based on the criteria discussed. This resource highlights each model’s strengths and limitations, facilitating informed decisions in research planning.
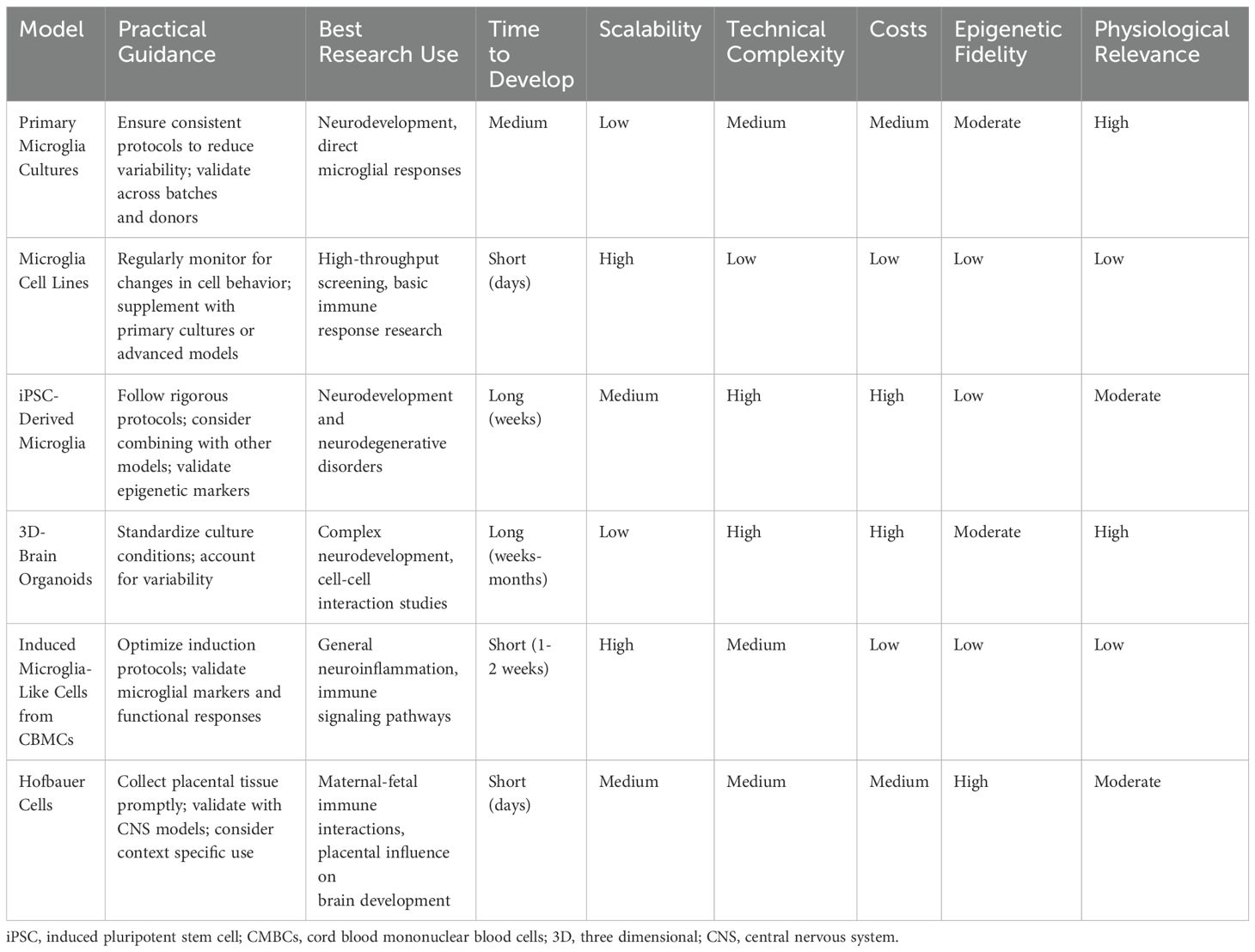
Table 1. Practical considerations for in-vitro microglia models.
SS: Writing – original draft, Writing – review & editing, Conceptualization. GN: Writing – original draft, Writing – review & editing. CB: Writing – original draft, Writing – review & editing. GS: Writing – original draft, Writing – review & editing.
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
We thank Christopher Urbschat for proof reading.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Gluckman PD, Hanson MA. Developmental origins of disease paradigm: a mechanistic and evolutionary perspective. Pediatr Res. (2004) 56:311–7. doi: 10.1203/01.PDR.0000135998.08025.FB
2. Arck PC, Hecher K. Fetomaternal immune cross-talk and its consequences for maternal and offspring’s health. Nat Med. (2013) 19:548–56. doi: 10.1038/nm.3160
3. Hartwig IR, Sly PD, Schmidt LA, van Lieshout RJ, Bienenstock J, Holt PG, et al. Prenatal adverse life events increase the risk for atopic diseases in children, which is enhanced in the absence of a maternal atopic predisposition. J Allergy Clin Immunol. (2014) 134:160–9.e7. doi: 10.1016/j.jaci.2014.01.033
4. Buss C, Davis EP, Shahbaba B, Pruessner JC, Head K, Sandman CA. Maternal cortisol over the course of pregnancy and subsequent child amygdala and hippocampus volumes and affective problems. Proc Natl Acad Sci. (2012) 109:E1312–E9. doi: 10.1073/pnas.1201295109
5. Borren I, Tambs K, Gustavson K, Schjølberg S, Eriksen W, Håberg SE, et al. Early prenatal exposure to pandemic influenza A (H1N1) infection and child psychomotor development at 6months – A population-based cohort study. Early Hum Dev. (2018) 122:1–7. doi: 10.1016/j.earlhumdev.2018.05.005
6. Edlow AG, Castro VM, Shook LL, Kaimal AJ, Perlis RH. Neurodevelopmental outcomes at 1 year in infants of mothers who tested positive for SARS-CoV-2 during pregnancy. JAMA network Open. (2022) 5:e2215787–e. doi: 10.1001/jamanetworkopen.2022.15787
7. Berezovsky E, Kohn E, Britzi M, Efreim S, Berlin M, Oppenheimer S, et al. Possible associations between prenatal exposure to environmental pollutants and neurodevelopmental outcome in children. Reprod Toxicol. (2024) 128:108658. doi: 10.1016/j.reprotox.2024.108658
8. Volk HE, Park B, Hollingue C, Jones KL, Ashwood P, Windham GC, et al. Maternal immune response and air pollution exposure during pregnancy: insights from the Early Markers for Autism (EMA) study. J Neurodev Disord. (2020) 12:42. doi: 10.1186/s11689-020-09343-0
9. Schepanski S, Buss C, Hanganu-Opatz IL, Arck PC. Prenatal immune and endocrine modulators of offspring’s brain development and cognitive functions later in life. Front Immunol. (2018) 9:2186. doi: 10.3389/fimmu.2018.02186
10. Graignic-Philippe R, Dayan J, Chokron S, Jacquet A, Tordjman S. Effects of prenatal stress on fetal and child development: a critical literature review. Neurosci Biobehav Rev. (2014) 43:137–62. doi: 10.1016/j.neubiorev.2014.03.022
11. Suleri A, Rommel A-S, Dmitrichenko O, Muetzel RL, Cecil CA, de Witte L, et al. The association between maternal immune activation and brain structure and function in human offspring: a systematic review. Mol Psychiatry. (2024) 28:1–14. doi: 10.1038/s41380-024-02760-w
12. Paolicelli RC, Ferretti MT. Function and dysfunction of microglia during brain development: consequences for synapses and neural circuits. Front Synaptic Neurosci. (2017) 9:9. doi: 10.3389/fnsyn.2017.00009
13. Dziabis JE, Bilbo SD. Microglia and sensitive periods in brain development. In: Andersen SL. (eds) Sensitive Periods of Brain Development and Preventive Interventions. Current Topics in Behavioral Neurosciences (2021) 53:55–78. (Springer, Cham). doi: 10.1007/7854_2021_242
14. Paolicelli RC, Bolasco G, Pagani F, Maggi L, Scianni M, Panzanelli P, et al. Synaptic pruning by microglia is necessary for normal brain development. Science. (2011) 333:1456–8. doi: 10.1126/science.1202529
15. Chan A, Magnus T, Gold R. Phagocytosis of apoptotic inflammatory cells by microglia and modulation by different cytokines: mechanism for removal of apoptotic cells in the inflamed nervous system. Glia. (2001) 33:87–95. doi: 10.1002/1098-1136(20010101)33:1<87::AID-GLIA1008>3.0.CO;2-S
16. Magnus T, Chan A, Grauer O, Toyka KV, Gold R. Microglial phagocytosis of apoptotic inflammatory T cells leads to down-regulation of microglial immune activation. J Immunol. (2001) 167:5004–10. doi: 10.4049/jimmunol.167.9.5004
17. Magnus T, Chan A, Savill J, Toyka KV, Gold R. Phagocytotic removal of apoptotic, inflammatory lymphocytes in the central nervous system by microglia and its functional implications. J Neuroimmunol. (2002) 130:1–9. doi: 10.1016/S0165-5728(02)00212-6
18. Aarum J, Sandberg K, Haeberlein SL, Persson MA. Migration and differentiation of neural precursor cells can be directed by microglia. Proc Natl Acad Sci U S A. (2003) 100:15983–8. doi: 10.1073/pnas.2237050100
19. Monier A, Adle-Biassette H, Delezoide AL, Evrard P, Gressens P, Verney C. Entry and distribution of microglial cells in human embryonic and fetal cerebral cortex. J Neuropathol Exp Neurol. (2007) 66:372–82. doi: 10.1097/nen.0b013e3180517b46
20. Menassa DA, Gomez-Nicola D. Microglial dynamics during human brain development. Front Immunol. (2018) 9:1014. doi: 10.3389/fimmu.2018.01014
21. Ajami B, Bennett JL, Krieger C, Tetzlaff W, Rossi FM. Local self-renewal can sustain CNS microglia maintenance and function throughout adult life. Nat Neurosci. (2007) 10:1538–43. doi: 10.1038/nn2014
22. Madinier A, Bertrand N, Mossiat C, Prigent-Tessier A, Beley A, Marie C, et al. Microglial involvement in neuroplastic changes following focal brain ischemia in rats. PloS One. (2009) 4:e8101. doi: 10.1371/journal.pone.0008101
23. Cherry JD, Tripodis Y, Alvarez VE, Huber B, Kiernan PT, Daneshvar DH, et al. Microglial neuroinflammation contributes to tau accumulation in chronic traumatic encephalopathy. Acta Neuropathol Commun. (2016) 4:112. doi: 10.1186/s40478-016-0382-8
24. Schalbetter SM, von Arx AS, Cruz-Ochoa N, Dawson K, Ivanov A, Mueller FS, et al. Adolescence is a sensitive period for prefrontal microglia to act on cognitive development. Sci Adv. (2022) 8:eabi6672. doi: 10.1126/sciadv.abi6672
25. Willis EF, MacDonald KPA, Nguyen QH, Garrido AL, Gillespie ER, Harley SBR, et al. Repopulating microglia promote brain repair in an IL-6-dependent manner. Cell. (2020) 180:833–46.e16. doi: 10.1016/j.cell.2020.02.013
26. Rosin JM, Sinha S, Biernaskie J, Kurrasch DM. A subpopulation of embryonic microglia respond to maternal stress and influence nearby neural progenitors. Dev Cell. (2021) 56:1326–45.e6. doi: 10.1016/j.devcel.2021.03.018
27. Gomez-Gonzalez B, Escobar A. Prenatal stress alters microglial development and distribution in postnatal rat brain. Acta Neuropathol. (2010) 119:303–15. doi: 10.1007/s00401-009-0590-4
28. Loayza M, Lin S, Carter K, Ojeda N, Fan LW, Ramarao S, et al. Maternal immune activation alters fetal and neonatal microglia phenotype and disrupts neurogenesis in mice. Pediatr Res. (2023) 93:1216–25. doi: 10.1038/s41390-022-02239-w
29. Gumusoglu SB, Fine RS, Murray SJ, Bittle JL, Stevens HE. The role of IL-6 in neurodevelopment after prenatal stress. Brain Behav Immun. (2017) 65:274–83. doi: 10.1016/j.bbi.2017.05.015
30. Schepanski S, Chini M, Sternemann V, Urbschat C, Thiele K, Sun T, et al. Pregnancy-induced maternal microchimerism shapes neurodevelopment and behavior in mice. Nat Commun. (2022) 13:4571. doi: 10.1038/s41467-022-32230-2
31. Block CL, Eroglu O, Mague SD, Smith CJ, Ceasrine AM, Sriworarat C, et al. Prenatal environmental stressors impair postnatal microglia function and adult behavior in males. Cell Rep. (2022) 40:111161. doi: 10.1016/j.celrep.2022.111161
32. Levesque S, Taetzsch T, Lull ME, Kodavanti U, Stadler K, Wagner A, et al. Diesel exhaust activates and primes microglia: air pollution, neuroinflammation, and regulation of dopaminergic neurotoxicity. Environ Health Perspect. (2011) 119:1149–55. doi: 10.1289/ehp.1002986
33. Hayes LN, An K, Carloni E, Li F, Vincent E, Trippaers C, et al. Prenatal immune stress blunts microglia reactivity, impairing neurocircuitry. Nature. (2022) 610:327–34. doi: 10.1038/s41586-022-05274-z
34. Wu J, Zhang J, Chen X, Wettschurack K, Que Z, Deming BA, et al. Microglial over-pruning of synapses during development in autism-associated SCN2A-deficient mice and human cerebral organoids. Mol Psychiatry. (2024) 29:2424–37. doi: 10.1038/s41380-024-02518-4
35. Chini M, Popplau JA, Lindemann C, Carol-Perdiguer L, Hnida M, Oberlander V, et al. Resolving and rescuing developmental miswiring in a mouse model of cognitive impairment. Neuron. (2020) 105:60–74.e7. doi: 10.1016/j.neuron.2019.09.042
36. Coiro P, Padmashri R, Suresh A, Spartz E, Pendyala G, Chou S, et al. Impaired synaptic development in a maternal immune activation mouse model of neurodevelopmental disorders. Brain Behav Immun. (2015) 50:249–58. doi: 10.1016/j.bbi.2015.07.022
37. Brown TC, Crouse EC, Attaway CA, Oakes DK, Minton SW, Borghuis BG, et al. Microglia are dispensable for experience-dependent refinement of mouse visual circuitry. Nat Neurosci. (2024) 27(8):1462–7. doi: 10.1038/s41593-024-01706-3
38. Smith AM, Dragunow M. The human side of microglia. Trends Neurosci. (2014) 37:125–35. doi: 10.1016/j.tins.2013.12.001
39. Geirsdottir L, David E, Keren-Shaul H, Weiner A, Bohlen SC, Neuber J, et al. Cross-species single-cell analysis reveals divergence of the primate microglia program. Cell. (2019) 179:1609–22.e16. doi: 10.1016/j.cell.2019.11.010
40. Kracht L, Borggrewe M, Eskandar S, Brouwer N, Chuva de Sousa Lopes SM, Laman JD, et al. Human fetal microglia acquire homeostatic immune-sensing properties early in development. Science. (2020) 369:530–7. doi: 10.1126/science.aba5906
41. Menassa DA, Muntslag TAO, Martin-Estebane M, Barry-Carroll L, Chapman MA, Adorjan I, et al. The spatiotemporal dynamics of microglia across the human lifespan. Dev Cell. (2022) 57:2127–39.e6. doi: 10.1016/j.devcel.2022.07.015
42. Bordt EA, Block CL, Petrozziello T, Sadri-Vakili G, Smith CJ, Edlow AG, et al. Isolation of microglia from mouse or human tissue. STAR Protoc. (2020) 1:100035. doi: 10.1016/j.xpro.2020.100035
43. Bennett ML, Bennett FC, Liddelow SA, Ajami B, Zamanian JL, Fernhoff NB, et al. New tools for studying microglia in the mouse and human CNS. Proc Natl Acad Sci U S A. (2016) 113:E1738–46. doi: 10.1073/pnas.1525528113
44. Olah M, Raj D, Brouwer N, De Haas AH, Eggen BJ, Den Dunnen WF, et al. An optimized protocol for the acute isolation of human microglia from autopsy brain samples. Glia. (2012) 60:96–111. doi: 10.1002/glia.21251
45. Tamashiro TT, Dalgard CL, Byrnes KR. Primary microglia isolation from mixed glial cell cultures of neonatal rat brain tissue. J Vis Exp. (2012) 66:e3814. doi: 10.3791/3814-v
46. Krassner MM, Kauffman J, Sowa A, Cialowicz K, Walsh S, Farrell K, et al. Postmortem changes in brain cell structure: a review. Free Neuropathol. (2023) 4:4–10. doi: 10.31219/osf.io/gj29w
47. Kandigian SE, Ethier EC, Kitchen RR, Lam TT, Arnold SE, Carlyle BC. Proteomic characterization of post-mortem human brain tissue following ultracentrifugation-based subcellular fractionation. Brain Commun. (2022) 4:fcac103. doi: 10.1093/braincomms/fcac103
48. Blair JA, Wang C, Hernandez D, Siedlak SL, Rodgers MS, Achar RK, et al. Individual case analysis of postmortem interval time on brain tissue preservation. PloS One. (2016) 11:e0151615. doi: 10.1371/journal.pone.0151615
49. Guo J, Yang WT, Mai FY, Liang JR, Luo J, Zhou MC, et al. Unravelling oncosis: morphological and molecular insights into a unique cell death pathway. Front Immunol. (2024) 15:1450998. doi: 10.3389/fimmu.2024.1450998
50. Donaldson AE, Lamont IL. Metabolomics of post-mortem blood: identifying potential markers of post-mortem interval. Metabolomics. (2015) 11:237–45. doi: 10.1007/s11306-014-0691-5
51. Janabi N, Di Stefano M, Wallon C, Hery C, Chiodi F, Tardieu M. Induction of human immunodeficiency virus type 1 replication in human glial cells after proinflammatory cytokines stimulation: effect of IFNgamma, IL1beta, and TNFalpha on differentiation and chemokine production in glial cells. Glia. (1998) 23:304–15. doi: 10.1002/(SICI)1098-1136(199808)23:4<304::AID-GLIA3>3.0.CO;2-2
52. Nagai A, Nakagawa E, Hatori K, Choi HB, McLarnon JG, Lee MA, et al. Generation and characterization of immortalized human microglial cell lines: expression of cytokines and chemokines. Neurobiol Dis. (2001) 8:1057–68. doi: 10.1006/nbdi.2001.0437
53. Butovsky O, Jedrychowski MP, Moore CS, Cialic R, Lanser AJ, Gabriely G, et al. Identification of a unique TGF-beta-dependent molecular and functional signature in microglia. Nat Neurosci. (2014) 17:131–43. doi: 10.1038/nn.3599
54. Das A, Kim SH, Arifuzzaman S, Yoon T, Chai JC, Lee YS, et al. Transcriptome sequencing reveals that LPS-triggered transcriptional responses in established microglia BV2 cell lines are poorly representative of primary microglia. J Neuroinflammation. (2016) 13:182. doi: 10.1186/s12974-016-0644-1
55. Melief J, Sneeboer MA, Litjens M, Ormel PR, Palmen SJ, Huitinga I, et al. Characterizing primary human microglia: A comparative study with myeloid subsets and culture models. Glia. (2016) 64:1857–68. doi: 10.1002/glia.v64.11
56. Martin S, Vincent JP, Mazella J. Involvement of the neurotensin receptor-3 in the neurotensin-induced migration of human microglia. J Neurosci. (2003) 23:1198–205. doi: 10.1523/JNEUROSCI.23-04-01198.2003
57. Garcia-Mesa Y, Jay TR, Checkley MA, Luttge B, Dobrowolski C, Valadkhan S, et al. Immortalization of primary microglia: a new platform to study HIV regulation in the central nervous system. J Neurovirol. (2017) 23:47–66. doi: 10.1007/s13365-016-0499-3
58. Gordon K, Clouaire T, Bao XX, Kemp SE, Xenophontos M, de Las Heras JI, et al. Immortality, but not oncogenic transformation, of primary human cells leads to epigenetic reprogramming of DNA methylation and gene expression. Nucleic Acids Res. (2014) 42:3529–41. doi: 10.1093/nar/gkt1351
59. Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. (2006) 126:663–76. doi: 10.1016/j.cell.2006.07.024
60. Haenseler W, Sansom SN, Buchrieser J, Newey SE, Moore CS, Nicholls FJ, et al. A highly efficient human pluripotent stem cell microglia model displays a neuronal-co-culture-specific expression profile and inflammatory response. Stem Cell Rep. (2017) 8:1727–42. doi: 10.1016/j.stemcr.2017.05.017
61. Abud EM, Ramirez RN, Martinez ES, Healy LM, Nguyen CHH, Newman SA, et al. iPSC-derived human microglia-like cells to study neurological diseases. Neuron. (2017) 94:278–93.e9. doi: 10.1016/j.neuron.2017.03.042
62. Pandya H, Shen MJ, Ichikawa DM, Sedlock AB, Choi Y, Johnson KR, et al. Differentiation of human and murine induced pluripotent stem cells to microglia-like cells. Nat Neurosci. (2017) 20:753–9. doi: 10.1038/nn.4534
63. Douvaras P, Sun B, Wang M, Kruglikov I, Lallos G, Zimmer M, et al. Directed differentiation of human pluripotent stem cells to microglia. Stem Cell Rep. (2017) 8:1516–24. doi: 10.1016/j.stemcr.2017.04.023
64. Mertens J, Paquola ACM, Ku M, Hatch E, Bohnke L, Ladjevardi S, et al. Directly reprogrammed human neurons retain aging-associated transcriptomic signatures and reveal age-related nucleocytoplasmic defects. Cell Stem Cell. (2015) 17:705–18. doi: 10.1016/j.stem.2015.09.001
65. Prasad A, Manivannan J, Loong DT, Chua SM, Gharibani PM, All AH. A review of induced pluripotent stem cell, direct conversion by trans-differentiation, direct reprogramming and oligodendrocyte differentiation. Regener Med. (2016) 11:181–91. doi: 10.2217/rme.16.5
66. Krencik R, Zhang SC. Directed differentiation of functional astroglial subtypes from human pluripotent stem cells. Nat Protoc. (2011) 6:1710–7. doi: 10.1038/nprot.2011.405
67. Nistor GI, Totoiu MO, Haque N, Carpenter MK, Keirstead HS. Human embryonic stem cells differentiate into oligodendrocytes in high purity and myelinate after spinal cord transplantation. Glia. (2005) 49:385–96. doi: 10.1002/glia.20127
68. Sellgren CM, Gracias J, Watmuff B, Biag JD, Thanos JM, Whittredge PB, et al. Increased synapse elimination by microglia in schizophrenia patient-derived models of synaptic pruning. Nat Neurosci. (2019) 22:374–85. doi: 10.1038/s41593-018-0334-7
69. Kyrousi C, Cappello S. Using brain organoids to study human neurodevelopment, evolution and disease. Wiley Interdiscip Rev Dev Biol. (2020) 9:e347. doi: 10.1002/wdev.347
70. Amiri A, Coppola G, Scuderi S, Wu F, Roychowdhury T, Liu F, et al. Transcriptome and epigenome landscape of human cortical development modeled in organoids. Science. (2018) 362:eaat6720. doi: 10.1126/science.aat6720
71. Camp JG, Badsha F, Florio M, Kanton S, Gerber T, Wilsch-Brauninger M, et al. Human cerebral organoids recapitulate gene expression programs of fetal neocortex development. Proc Natl Acad Sci U S A. (2015) 112:15672–7. doi: 10.1073/pnas.1520760112
72. Kanton S, Boyle MJ, He Z, Santel M, Weigert A, Sanchis-Calleja F, et al. Organoid single-cell genomic atlas uncovers human-specific features of brain development. Nature. (2019) 574:418–22. doi: 10.1038/s41586-019-1654-9
73. Qian X, Jacob F, Song MM, Nguyen HN, Song H, Ming GL. Generation of human brain region-specific organoids using a miniaturized spinning bioreactor. Nat Protoc. (2018) 13:565–80. doi: 10.1038/nprot.2017.152
74. Watanabe M, Buth JE, Vishlaghi N, de la Torre-Ubieta L, Taxidis J, Khakh BS, et al. Self-organized cerebral organoids with human-specific features predict effective drugs to combat zika virus infection. Cell Rep. (2017) 21:517–32. doi: 10.1016/j.celrep.2017.09.047
75. Benito-Kwiecinski S, Lancaster MA. Brain organoids: human neurodevelopment in a dish. Cold Spring Harb Perspect Biol. (2020) 12:a035709. doi: 10.1101/cshperspect.a035709
76. Qian X, Nguyen HN, Song MM, Hadiono C, Ogden SC, Hammack C, et al. Brain-region-specific organoids using mini-bioreactors for modeling ZIKV exposure. Cell. (2016) 165:1238–54. doi: 10.1016/j.cell.2016.04.032
77. Qian X, Song H, Ming GL. Brain organoids: advances, applications and challenges. Development. (2019) 146:dev166074. doi: 10.1242/dev.166074
78. Silva TP, Sousa-Luis R, Fernandes TG, Bekman EP, Rodrigues CAV, Vaz SH, et al. Transcriptome profiling of human pluripotent stem cell-derived cerebellar organoids reveals faster commitment under dynamic conditions. Biotechnol Bioeng. (2021) 118:2781–803. doi: 10.1002/bit.v118.7
79. Velasco S, Kedaigle AJ, Simmons SK, Nash A, Rocha M, Quadrato G, et al. Individual brain organoids reproducibly form cell diversity of the human cerebral cortex. Nature. (2019) 570:523–7. doi: 10.1038/s41586-019-1289-x
80. Lancaster MA, Renner M, Martin CA, Wenzel D, Bicknell LS, Hurles ME, et al. Cerebral organoids model human brain development and microcephaly. Nature. (2013) 501:373–9. doi: 10.1038/nature12517
81. Park DS, Kozaki T, Tiwari SK, Moreira M, Khalilnezhad A, Torta F, et al. iPS-cell-derived microglia promote brain organoid maturation via cholesterol transfer. Nature. (2023) 623:397–405. doi: 10.1038/s41586-023-06713-1
82. Quadrato G, Nguyen T, Macosko EZ, Sherwood JL, Min Yang S, Berger DR, et al. Cell diversity and network dynamics in photosensitive human brain organoids. Nature. (2017) 545:48–53. doi: 10.1038/nature22047
83. Oberlander TF, Weinberg J, Papsdorf M, Grunau R, Misri S, Devlin AM. Prenatal exposure to maternal depression, neonatal methylation of human glucocorticoid receptor gene (NR3C1) and infant cortisol stress responses. Epigenetics. (2008) 3:97–106. doi: 10.4161/epi.3.2.6034
84. Kocher K, Bhattacharya S, Niforatos-Andescavage N, Almalvez M, Henderson D, Vilain E, et al. Genome-wide neonatal epigenetic changes associated with maternal exposure to the COVID-19 pandemic. BMC Med Genomics. (2023) 16:268. doi: 10.1186/s12920-023-01707-4
85. Easley-Neal C, Foreman O, Sharma N, Zarrin AA, Weimer RM. CSF1R ligands IL-34 and CSF1 are differentially required for microglia development and maintenance in white and gray matter brain regions. Front Immunol. (2019) 10:2199. doi: 10.3389/fimmu.2019.02199
86. Wang Y, Szretter KJ, Vermi W, Gilfillan S, Rossini C, Cella M, et al. IL-34 is a tissue-restricted ligand of CSF1R required for the development of Langerhans cells and microglia. Nat Immunol. (2012) 13:753–60. doi: 10.1038/ni.2360
87. Dikmen HO, Hemmerich M, Lewen A, Hollnagel JO, Chausse B, Kann O. GM-CSF induces noninflammatory proliferation of microglia and disturbs electrical neuronal network rhythms in situ. J Neuroinflamm. (2020) 17:235. doi: 10.1186/s12974-020-01903-4
88. Liva SM, Kahn MA, Dopp JM, de Vellis J. Signal transduction pathways induced by GM-CSF in microglia: significance in the control of proliferation. Glia. (1999) 26:344–52. doi: 10.1002/(SICI)1098-1136(199906)26:4<344::AID-GLIA8>3.0.CO;2-L
89. Smith AM, Gibbons HM, Oldfield RL, Bergin PM, Mee EW, Curtis MA, et al. M-CSF increases proliferation and phagocytosis while modulating receptor and transcription factor expression in adult human microglia. J Neuroinflammation. (2013) 10:85. doi: 10.1186/1742-2094-10-85
90. Sheridan SD, Thanos JM, De Guzman RM, McCrea LT, Horng JE, Fu T, et al. Umbilical cord blood-derived microglia-like cells to model COVID-19 exposure. Transl Psychiatry. (2021) 11:179. doi: 10.1038/s41398-021-01287-w
91. Ormel PR, Bottcher C, Gigase FAJ, Missall RD, van Zuiden W, Fernandez Zapata MC, et al. A characterization of the molecular phenotype and inflammatory response of schizophrenia patient-derived microglia-like cells. Brain Behav Immun. (2020) 90:196–207. doi: 10.1016/j.bbi.2020.08.012
92. Ryan KJ, White CC, Patel K, Xu J, Olah M, Replogle JM, et al. A human microglia-like cellular model for assessing the effects of neurodegenerative disease gene variants. Sci Transl Med. (2017) 9:eaai7635. doi: 10.1126/scitranslmed.aai7635
93. Sargeant TJ, Fourrier C. Human monocyte-derived microglia-like cell models: A review of the benefits, limitations and recommendations. Brain Behav Immun. (2023) 107:98–109. doi: 10.1016/j.bbi.2022.09.015
94. Cuní-López C, Stewart R, Oikari LE, Nguyen TH, Roberts TL, Sun Y, et al. Advanced patient-specific microglia cell models for pre-clinical studies in Alzheimer’s disease. J neuroinflammation. (2024) 21:50. doi: 10.1186/s12974-024-03037-3
95. Batorsky R, Ceasrine AM, Shook LL, Kislal S, Bordt EA, Devlin BA, et al. Hofbauer cells and fetal brain microglia share transcriptional profiles and responses to maternal diet-induced obesity. Cell Rep. (2024) 43:114326. doi: 10.1016/j.celrep.2024.114326
96. Chen X, Tang AT, Tober J, Yang J, Leu NA, Sterling S, et al. Mouse placenta fetal macrophages arise from endothelial cells outside the placenta. Dev Cell. (2022) 57:2652–60.e3. doi: 10.1016/j.devcel.2022.11.003
97. Freyer L, Lallemand Y, Dardenne P, Sommer A, Biton A, Gomez Perdiguero E. Erythro-myeloid progenitor origin of Hofbauer cells in the early mouse placenta. Development. (2022) 149:dev200104. doi: 10.1242/dev.200104
98. Reyes L, Golos TG. Hofbauer cells: their role in healthy and complicated pregnancy. Front Immunol. (2018) 9:2628. doi: 10.3389/fimmu.2018.02628
99. Thomas JR, Appios A, Zhao X, Dutkiewicz R, Donde M, Lee CYC, et al. Phenotypic and functional characterization of first-trimester human placental macrophages, Hofbauer cells. J Exp Med. (2021) 218:e20200891. doi: 10.1084/jem.20200891
Keywords: microglia, brain development, in-vitro models, neuroimmunology and microglia, neurodevelopmental disorders, neuropsychiatry, developmental origin of health and diseases, fetal programming
Citation: Schepanski S, Ngoumou GB, Buss C and Seifert G (2025) Assessing in-vitro models for microglial development and fetal programming: a critical review. Front. Immunol. 16:1538920. doi: 10.3389/fimmu.2025.1538920
Received: 03 December 2024; Accepted: 08 January 2025;
Published: 29 January 2025.
Edited by:
Sandra Maria Blois, University Medical Center Hamburg-Eppendorf, GermanyReviewed by:
Alexandre Bonnin, University of Southern California, United StatesCopyright © 2025 Schepanski, Ngoumou, Buss and Seifert. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Steven Schepanski, c3RldmVuLnNjaGVwYW5za2lAY2hhcml0ZS5kZQ==
†These authors share last authorship
‡ORCID: Steven Schepanski, orcid.org/0000-0002-7695-1289
Georg Seifert, orcid.org/0000-0002-7109-9277
Gonza B. Ngoumou, orcid.org/0000-0001-6501-732X
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.