
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Immunol., 06 March 2025
Sec. Vaccines and Molecular Therapeutics
Volume 16 - 2025 | https://doi.org/10.3389/fimmu.2025.1538453
This article is part of the Research TopicSARS-CoV-2 Vaccines Beyond the Pandemic EraView all 26 articles
 Vitor Gabriel Lopes da Silva1*†
Vitor Gabriel Lopes da Silva1*† Gabriela Justamante Händel Schmitz2†
Gabriela Justamante Händel Schmitz2† Kathleen E. Sullivan3†
Kathleen E. Sullivan3† Júlia Barbate1†Maria Izabel de Haro Azinar1
Júlia Barbate1†Maria Izabel de Haro Azinar1 Carolina Sanchez Aranda1†
Carolina Sanchez Aranda1† Maria Isabel de Moraes-Pinto1*†
Maria Isabel de Moraes-Pinto1*†Objective: Patients with Inborn Errors of Immunity (IEI) are at higher risk of severe SARS-CoV-2 infection. We evaluated humoral and cellular responses to COVID-19 vaccines in Brazilian patients with IEI and healthy controls.
Methods: Fifty-five patients with IEI (13–61 years) and 60 controls (13–71 years) received inactivated SARS-CoV-2 (CoronaVac), non-replicating virus-vectored (ChAdOx1 nCoV-19, AstraZeneca) or monovalent mRNA (Original strain of BNT162b2, Pfizer-BioNTech) and bivalent mRNA (Original/Omicron BA.1, Pfizer-BioNTech) vaccines and were sampled five times. Diagnoses included common variable immunodeficiency (n=25), specific antibody deficiency (n=9), ataxia-telangiectasia (n=5), X-linked agammaglobulinemia (n=4), PIK3CD-related disorders (n=4), hyper-IgM syndrome (n=4), combined immunodeficiency (n=3), and STAT1 gain-of-function (n=1). Humoral immunity was assessed via multiplex microarray for Spike, Nucleocapsid, RBD-Wuhan, RBD-Delta, RBD-BA.1, RBD-BA.2 and RBD-BA.5 neutralizing antibodies. T-cell responses to Spike and Nucleocapsid were assessed using ELISpot.
Results: Patients with IEI exhibited significantly lower levels of Nucleocapsid and RBD-neutralizing antibodies (p < 0.05). Notable differences in RBD-BA.2 (p = 0.008) and IgG-Nucleocapsid (p = 0.010) levels emerged over time. T-cell responses to Spike were stronger in patients with IEI post-booster (405 vs. 149 spot-forming cells/million PBMC; p = 0.002). Both groups showed enhanced Nucleocapsid-specific cellular responses over time (p = 0.017). COVID-19 hospitalization rates among patients with IEI with SARS-CoV-2 diagnosis dropped from 33.3% to zero after the first booster dose.
Conclusions: While humoral responses to SARS-CoV-2 vaccines were weaker in patients with IEI, their cellular immunity was similar to controls. Boosters enhanced both humoral and cellular responses. After completion of the vaccination protocol, none of the patients with IEI were hospitalized with COVID-19. Robust T-cell responses may play a critical role in protecting patients with IEI from severe COVID-19 and mortality.
The COVID-19 pandemic has affected more than 777 million people and killed more than 7.1 million people worldwide as of January 2025. In Brazil alone, there were nearly 38 million confirmed cases and 703,000 deaths in the same period (1). Specific conditions, such as combined immunodeficiencies, immune dysregulation disorders [especially defects in tolerance, such as IPEX (immune dysregulation, polyendocrinopathy, and enteropathy X-linked syndrome), and other “TRegopathies”], and defects in the type I interferon pathway are associated with worse COVID-19 outcomes (2).
Although patients with Inborn Errors of Immunity (IEI) are at increased risk of developing severe COVID-19 (2, 3), they can develop potentially protective immune responses following vaccination, which can be further enhanced by booster doses. The wide range of vaccination response rates may be attributed to different vaccination protocols (4, 5), different underlying conditions and the small sample sizes of published studies (6). Antibody responses alone may not necessarily be correlated with the prevention of COVID-19 hospitalization, as other immunological mediators, such as vaccine-specific T-cell responses, can prevent or reduce the severity of COVID-19 (7–10).
Published studies of responses after two doses of COVID-19 vaccination in patients with IEI indicated that 48.5% to 86.0% of patients developed neutralizing antibodies against SARS-CoV-2 (4, 5, 11–15), whereas 73.1% to 87.0% of patients exhibited T-cell responses (4, 5, 11–13, 15). The COVID-19 vaccines induced substantially lower immune responses in patients with IEI than in healthy controls (4, 11, 12, 16). These differences were especially significant concerning neutralizing antibodies to Omicron variants with relevant specific mutations that induce an immune escape (17).
In Brazil, the vaccination of patients with IEI started in May 2021 after more than 400,000 deaths. Four months later, in September 2021, the administration of the third COVID-19 vaccine dose for immunosuppressed individuals began. The immunization was preferably performed with an original strain of BNT162b2 (Pfizer-BioNTech) or, alternatively, with a viral vector vaccine of Ad26.COV2.S (Janssen) or ChAdOx1 nCoV-19 (AstraZeneca) (18). The administration of bivalent mRNA (Original/Omicron BA.1, Pfizer-BioNTech) vaccine as booster began in March 2023.
The high burden of COVID-19 in Brazil led us to analyze responses in patients with IEI followed at the Immunology Clinic at the Federal University of Sao Paulo. This patient population is vulnerable and the level of protection has not yet been characterized. We concluded that most patients with IEI respond to COVID-19 immunization with a three-dose primary vaccination schedule followed by two booster doses (4th and 5th vaccines doses) although humoral and T cell responses differed.
This study adheres to the principles of the Declaration of Helsinki and was approved by the Brazilian National Research Ethics Committee (number 51535921.2.0000.5505). All participants provided written informed consent before enrollment.
This was a prospective cohort study that took place between October 2021 and November 2023 (Supplementary Figure S1 in the Supplementary Material). Patients with an established IEI diagnosis from a Brazilian reference center, the Immunology Clinic of the Federal University of São Paulo, who received three doses of COVID-19 vaccines, were offered the opportunity to join the study. A control group of healthy individuals of similar sex and age distributions were also invited to participate. One-third of patients with IEI and controls had a basic immunization schedule that consisted of two doses of either CoronaVac, BNT162b2 (Pfizer-BioNTech) or ChAdOx1 nCoV-19 (AstraZeneca), followed by a third dose of BNT162b2 (Pfizer-BioNTech). The fourth and fifth doses varied according to the availability of the different COVID-19 vaccine platforms in both the IEI cohort and the control cohort. Patients with IEI had blood samples collected 1 and 6 months after the fourth COVID-19 vaccine dose (the first booster for the IEI cohort) and 1 month after the fifth COVID-19 vaccine dose (the second booster for the IEI cohort). Blood samples were collected from control individuals 1 and 6 months after the third COVID-19 vaccine dose (the first booster for the control cohort) and 1 month after the fourth COVID-19 vaccine dose (the second booster for the control cohort) (Figure 1).
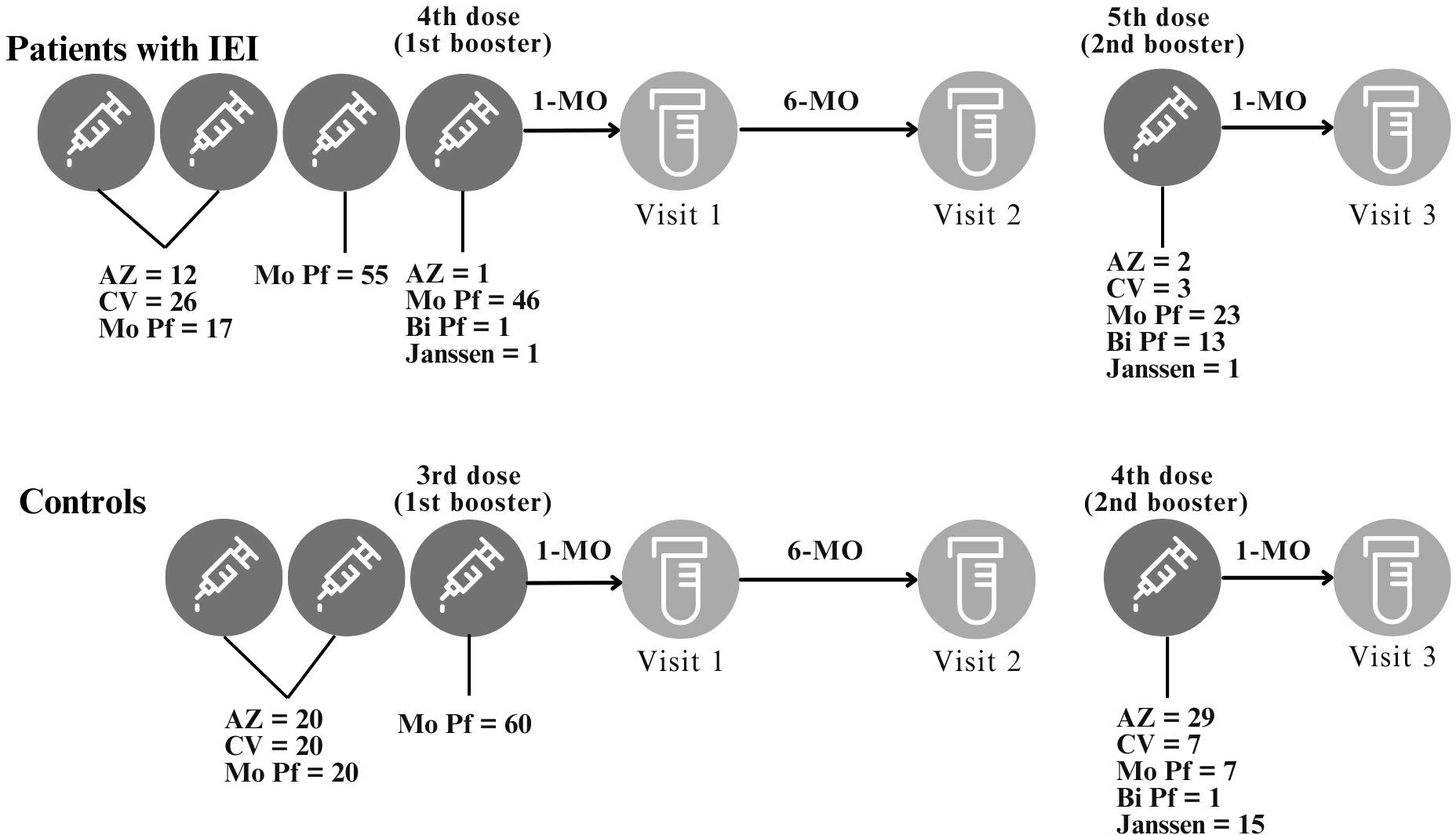
Figure 1. Study design. Study visits and intervals between study visits or between vaccination and study visits are depicted. The BNT162b2 (Mo Pf) vaccine was used as the third vaccine dose. The 4th and 5th vaccine doses were based on local availability. For the control cohort, the 3rd dose is the first booster and the 4th dose is the second booster. For the IEI cohort, the 4th dose is the first booster and the 5th dose is the second booster. This study compared the immune response after the 1st and 2nd booster for both IEI and Control cohorts. AZ = ChAdOx1 nCoV-19, AstraZeneca; CV = CoronaVac; Mo Pf = monovalent mRNA original strain of BNT162b2, Pfizer-BioNTech; Bi Pf = bivalent mRNA Original/Omicron BA.1, Pfizer-BioNTech; Janssen = Ad26.COV2.S.
Lymphocyte subsets (CD3+ T, CD4+ T, CD8+ T, CD19+ and NK cells) were assessed via a single platform (TruCount and Multitest, BD Biosciences, San Jose, CA) on a BD FACSCalibur™ 4-color flow cytometer using CellQuest software (BD Biosciences, San Jose, CA). Data analysis was performed via MultiSET v3.0.2 software (Becton, Dickinson and Company, New Jersey, USA).
Peripheral blood mononuclear cells (PBMCs) were isolated using Ficoll gradient density. Following isolation, the cells were stored in liquid nitrogen for later ELISpot assays to evaluate peptide-induced cytokine production.
Serum samples were separated on the day of blood collection and stored at -80°C for later analysis. The humoral immune response was assessed via the commercial SARS-CoV-2 NTChip® Test (V-NTCGOK) (Viramed Biotech AG, Germany). This is an in vitro qualitative and quantitative multiplex microarray microsystem based on an enzyme immunoassay previously validated against the 50% Plaque Reduction Neutralization Test (PRNT50) to determine the level of neutralizing antibodies against purified specific surface antigens from the RBD portion of the Spike protein, in printed spot triplets in a nitrocellulose membrane, from variants and subvariants of SARS-CoV-2, such as Wuhan – wild type, Delta, Omicron BA.1 – B.1.1.529 and BA.2 and BA.5, which are on each well of a 96-well plate (Supplementary Figure S2).
During the development of the NTChip® (19), in order to validate the PRNT50 – the gold standard to determine the level of neutralizing antibodies for many viral diseases – six groups of sera were evaluated: 1) pre-pandemic (W-O-), 2) previous Omicron variant (W+O-), 3) infected/vaccinated post Omicron (W+O+), 4) naïve Omicron positive (W-O+), 5) Omicron positive 0-2 days and 6) 10 days after infection. Sensitivity and specificity results for RBD-Wuhan, RBD-Omicron BA.1 and Nucleocapsid (N) were: 96.7% and 100.0%; 95.7% and 99.1%, and 95.1% and 94.0%, respectively.
To detect neutralizing antibodies, the assay uses the cellular receptor angiotensin conversion enzyme-2 (ACE-2). Results can be expressed for neutralizing antibodies in percentage of inhibition, which is transformed in IU/mL by the ViraChip® Software after the results are read by the ViraChip® Scanner. The NTChip® test can also assess a previous contact with the virus using Nucleocapsid (N) printed in spots triplets on the nitrocellulose; IgG antibodies from the sera are detected using a conjugated anti-IgG.
The result for captured IgG can be expressed in arbitrary units (AU) and in binding arbitrary units (BAU). For quantitative results, the NTChip® was calibrated against the WHO international standard NIBSC 21/338 (20). The quantitative cutoffs (NIBSC 21/338) for Wuhan, Omicron (BA.1) and Nucleocapsid were: 9.1 IU/mL, 626.5 IU/mL and 16.4 BAU/mL, respectively, which can be translated into the following percentages of inhibition: 9%, 29% and 74 AU, respectively. As only RBD-Wuhan, RBD-BA.1 and Nucleocapsid were validated against PRNT50, there are no cutoffs for RBD-Delta, RBD-BA.2 and RBD-BA.5.
To evaluate the specific T-cell immune response to SARS-CoV-2, the commercial T-SPOT®COVID kit (Oxford Immunotec, Oxford, England) was used. This standardized method detects CD4+ T and CD8+ T cells that secrete interferon-gamma (IFN-γ) in response to stimulation with antigens via two specific peptide pools from SARS-CoV-2, one from the Spike protein and the other from the Nucleocapsid. Each assay was read using the AID EliSpot fluorescence microplate reader (Autoimmun Diagnostika GMBH, Germany) and AID EliSpot 7.0 software. The test result is considered positive if either Spike and/or Nucleocapsid have a count of 8 spots or more per 250,000 PBMC (Supplementary Figure S3). The test result is considered negative if both have a count of 4 spots or less. Results of 5, 6, or 7 spots are considered indeterminate according to the manufacturer. A reactive result indicates the presence of SARS-CoV-2-sensitized effector T cells in the sample. A non-reactive result indicates that SARS-CoV-2-sensitized effector T cells were not detected in the sample.
Associations between two categorical variables were assessed using the Chi-square test or Fisher’s Exact test. Comparisons of means between two groups were performed using Student’s t-test. Normality assumption was checked using Kolmogorov-Smirnov test. In case of violation of this assumption, the nonparametric Mann−Whitney test was used alternatively.
Geometric means (GMT) and 95% confidence intervals (CIs) are presented for antibody assessment. Comparisons of variable levels by group and time were conducted via linear models with random effects (21) on log-transformed variables. This model assessed the effects of three components: time, group, and interaction between group and time. The presence of interaction between group and time indicates that group means evolve differently over time.
For qualitative cellular responses, logistic regression models with random effects were used. The linear model with random effects assumes data normality. However, deviation from normality does not bias estimates (22).
A significance level of 5% was used for all the statistical tests. Analyses were performed using SPSS 20.0 and STATA 17.
All the graphs were generated via GraphPad Prism 10.4.3.
A total of 55 patients with IEI and 60 controls were included in the study. The two groups were similar with respect to sex and age (p >0.05). The median age of patients with IEI was 27.3 years (ranging from 13.3 to 61.9 years) and that of the controls was 25.0 years (13.5 to 71.2 years) (p = 0.991). Patients with IEI patients had a median BMI of 21.91 kg/m² (15.15 to 36.51 kg/m²), which was lower than that of the controls (p = 0.026). The groups were comparable regarding the following comorbidities: systemic arterial hypertension, diabetes mellitus, heart disease, and obesity. Among the patients with IEI, 40% (22/55) had pulmonary diseases (asthma, bronchiectasis, pulmonary fibrosis, and lobectomy) and 10.9% (6/55) had other diseases, such as nephropathy, hepatopathy, neuropathy, and myasthenia gravis. Due to the exclusion criteria for the control group, no individuals in this group had diabetes mellitus, heart disease, pulmonary disease, or other comorbidities (Table 1).
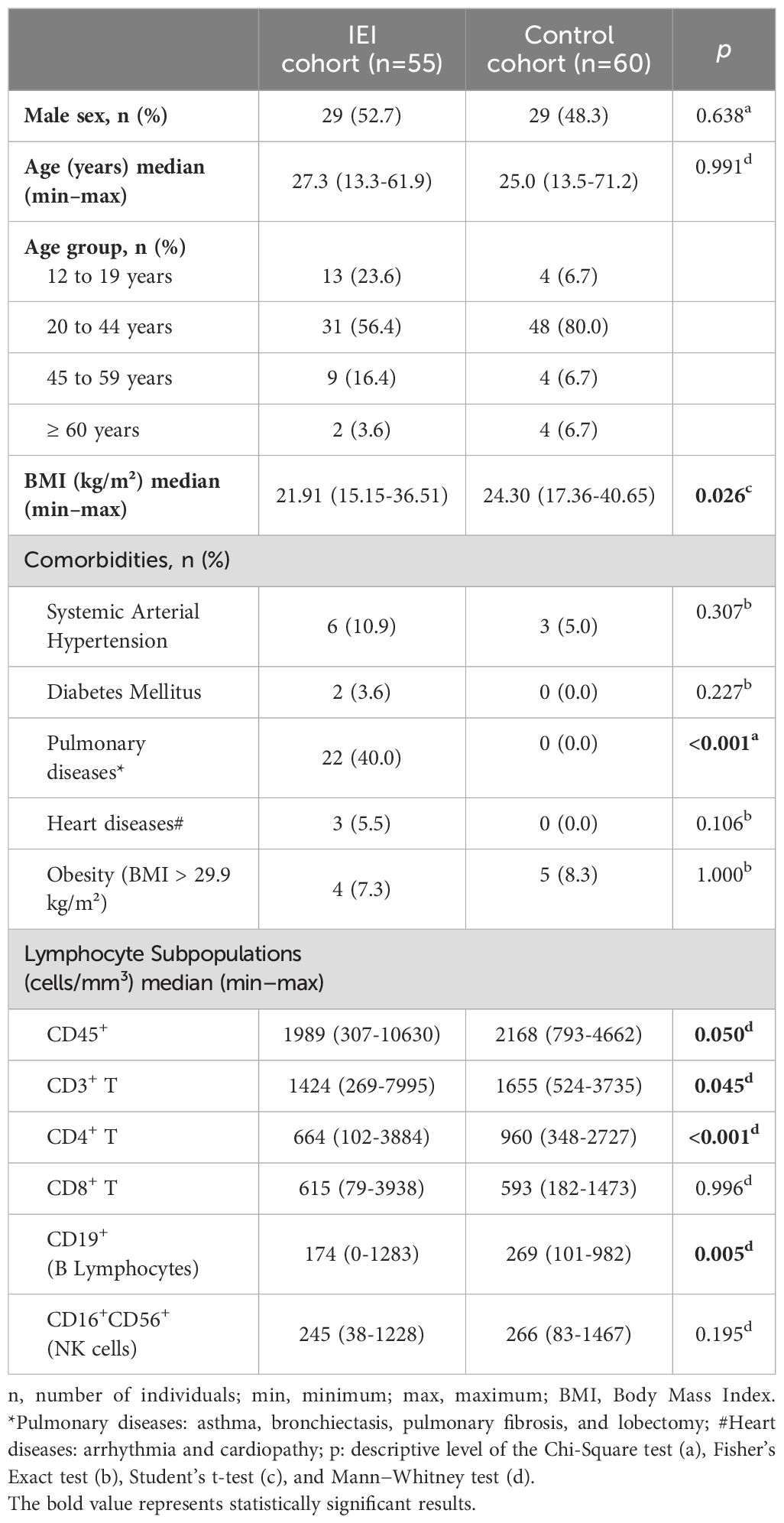
Table 1. Clinical and demographic data of the study participants.
The median numbers of CD3+ T lymphocytes (p = 0.045), CD4+ T lymphocytes (p < 0.001), and B lymphocytes (p = 0.005) were lower in the IEI group than in the control group. No differences were observed for CD8+ T lymphocytes or NK cells (p = 0.996 and p = 0.195, respectively) (Table 1).
All controls (n = 60) and 42 patients with IEI completed all 5 study visits (Figure 1). Among the 13 individuals who did not complete the visits, 6 did not want to receive the 4th vaccine dose, and 6 did not want to receive the 5th vaccine dose. One patient with X-linked Agammaglobulinemia had a history of pyoderma gangrenosum in the left lower limb that worsened during the study period; owing to frequent and long periods of hospitalization, he could not receive the 5th vaccine dose; this event was not considered to be caused by the vaccination.
The distribution of patients with IEI according to the first two COVID-19 vaccine doses administered is depicted in Table 2. Immunoglobulin replacement therapy (IGRT) was used by 48/55 (87.3%) patients with IEI. No differences were noted between the IEI and control groups with respect to the distribution of the types of the first two vaccine doses (p = 0.245). For the first and second doses, 47.3% (26/55) of the patients received CoronaVac, 30.9% (17/55) received BNT162b2 (Pfizer-BioNTech), and 21.8% (12/55) received ChAdOx1 nCoV-19 (AstraZeneca). The controls had an equal distribution of 33.3% for each vaccine (Figure 1).
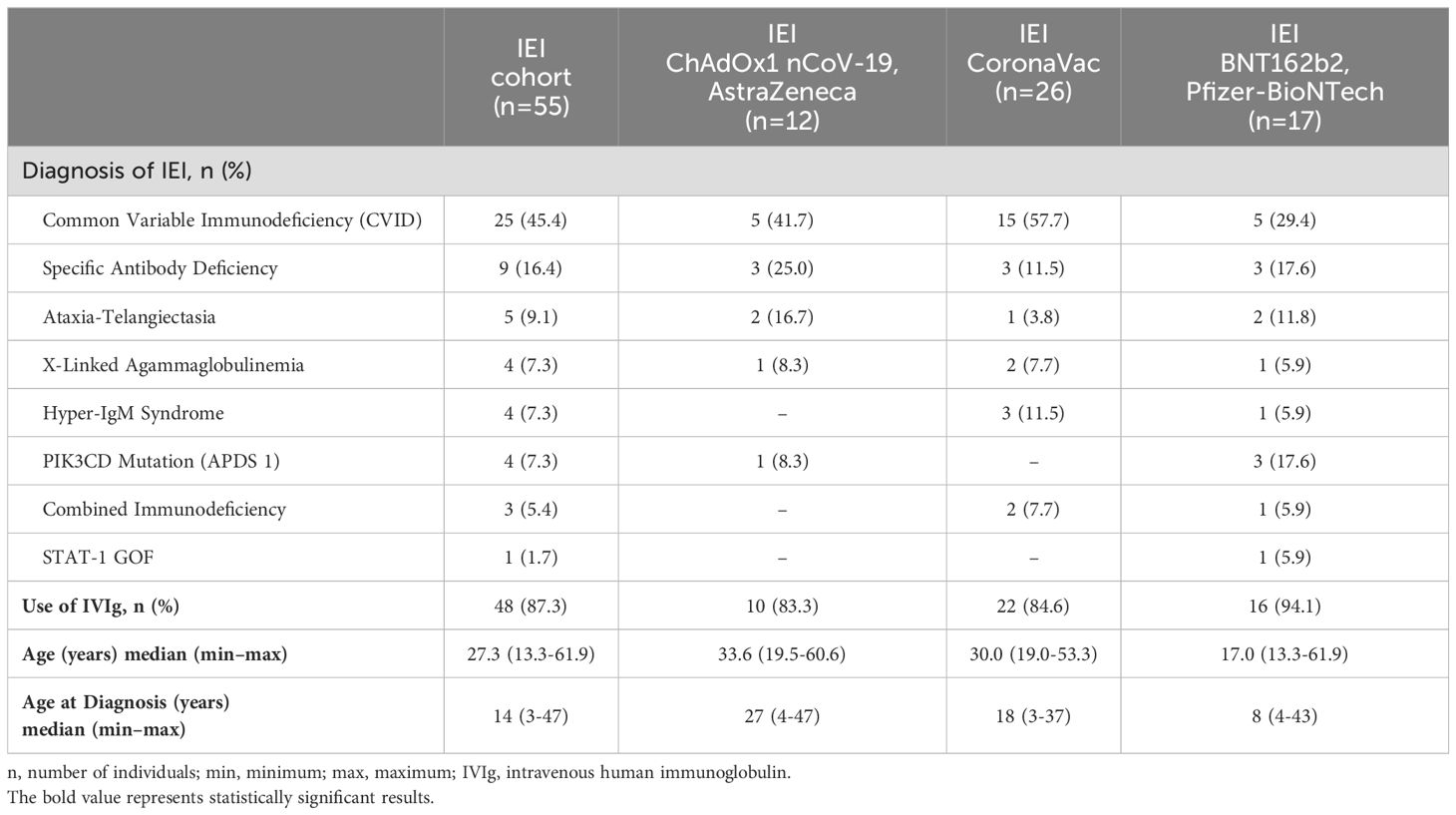
Table 2. Characteristics of the patients with IEI.
In both groups, all participants received original strain of BNT162b2 (Pfizer-BioNTech) as the third vaccine dose, according to the study protocol. For the fourth dose, following vaccination campaign priorities, patients with IEI predominantly received original strain of BNT162b2 (93.9%; 46/49). One patient received ChAdOx1 nCoV-19 (AstraZeneca), one received Ad26.COV2.S (Janssen), and one received bivalent mRNA (Original/Omicron BA.1, Pfizer-BioNTech) vaccine. Among controls, the fourth dose was predominantly with ChAdOx1 nCoV-19 (AstraZeneca) (49.2%; 29/59), followed by Ad26.COV2.S (Janssen) (25.4%; 15/59), CoronaVac, and original strain of BNT162b2 (11.9% each; 7/59), with one person receiving bivalent mRNA (Original/Omicron BA.1, Pfizer-BioNTech) (1.7%) (Figure 1). One control did not receive the fourth dose as he was under 18 years of age, and this vaccine was not yet available for that age group during the study period. Since patients with IEI were prioritized in the vaccination campaign, the mean interval between the third and fourth doses was smaller among patients with IEI compared to controls (150.2 days vs. 234.8 days; p < 0.001, Mann-Whitney test). This also occurred between the second and third doses: 95.5 days in patients with IEI and 174.3 days in controls (p < 0.001, Student’s t test).
Patients with IEI predominantly received the fifth dose (second booster) with original strain of BNT162b2 (54.8%; 23/42), followed by bivalent mRNA (Original/Omicron BA.1, Pfizer-BioNTech) (31%; 13/42). Three patients received CoronaVac (7.1%), two received ChAdOx1 nCoV-19 (AstraZeneca) (4.8%), and one received Ad26.COV2.S (Janssen) (2.4%) (Figure 1). As the bivalent mRNA (Original/Omicron BA.1, Pfizer-BioNTech) vaccine was not yet available at the start of the second booster campaign for patients with IEI, some individuals did not receive it. The mean interval between the fourth and fifth doses was 247 days, ranging from 123 to 514 days.
We assessed neutralizing antibodies to RBD-BA.1, RBD-BA.2, and RBD-BA.5 recognizing that there is some cross-reactivity and some antibodies related to natural infection (23, 24) (Supplementary Figure S1). We noted lower levels of neutralizing antibodies at first assessment for most epitopes tested. Changes in the GMTs of antibodies over time were evaluated via a linear regression model with random effects (Figure 2; see Supplementary Table S1 for seropositivity data and Supplementary Table S2 in the Supplementary Material for detailed data). Compared with controls, lower antibody levels for all variables analyzed were observed in patients with IEI (p < 0.05).

Figure 2. Humoral response. RBD-Wuhan (A), RBD-Delta (B), Nucleocapsid N-specific IgG (C), RBD-BA.1 (D), RBD-BA.2 (E) and RBD-BA.5 (F) antibodies 1 and 6 months after the 1st booster and 1 month after the 2nd booster in IEI and Control. For the control cohort, the 3rd dose is the first booster and the 4th dose is the second booster. For the IEI cohort, the 4th dose is the first booster and the 5th dose is the second booster. GMT values and 95% Confidence Intervals are shown. The dotted horizontal line is the quantitative cut-offs (NIBSC 21/338) for RBD-Wuhan (9.1 IU/mL), RBD-BA.1 (626.5 IU/mL) and Nucleocapsid (16.4 BAU/mL). No cut-offs are available for RBD-Delta, RBD-BA.2 and RBD-BA.5.
The rate of change of antibodies to RBD-Wuhan (p = 0.201) and RBD-Delta (p = 0.343) was similar over time when patients with IEI and controls were compared. Close to significant differences were also observed for antibodies to RBD-BA.1 (p = 0.078) and RBD-BA.5 (p = 0.056) variants. Over time, the two cohorts also showed a significantly distinct response to RBD-BA.2 neutralizing antibodies (p = 0.008). IgG-Nucleocapsid antibodies clearly differed over time with less increase over time apparent in patients with IEI (p = 0.010).
For RBD-BA.2, controls had lower GMT antibodies 6 months after the 1st booster (4266 IU/mL; 95% CI, 3124-5827), but antibodies increased 1 month after the 2nd booster (8402 IU/mL; 95% CI, 6513-10838), as did levels 1 month after the 1st booster (6603 IU/mL; 95% CI, 5159-8451). In the IEI cohort, the GMT was greater only 1 month after the 2nd booster (3002 IU/mL; 95% CI 1690-5329) than it was 1 month after the 1st booster (1975 IU/mL; 95% CI 1217-3203).
For Nucleocapsid, controls had higher GMT antibodies after the 2nd booster (87.8 BAU/mL; 95% CI, 68.1-113.1; p < 0.001) than 1 month and 6 months after the 1st booster, with similar GMTs between these time points [30.4 BAU/mL (95% CI, 23.1-39.9) and 49.5 BAU/mL (95% CI, 35.0-69.8), respectively]. In contrast, patients with IEI had similar GMTs across all evaluated time points [30.7 (95% CI, 22.6-41.7), 32.4 (95% CI, 21.8-48.3) and 42.2 BAU/mL (95% CI, 30.3-58.9)].
The RBD-BA.1, RBD-BA.2, RBD-BA.5 and Nucleocapsid antibodies were significantly increased 1 month after the second booster. For Nucleocapsid, IEI and control cohorts had similar GMTs 1 month after the 1st booster (30.7 vs 30.4 BAU/mL); however, 6 months after the 1st booster and 1 month after the 2nd booster, the IEI cohort had significantly lower GMTs than the controls did (32.4 vs 49.5 BAU/mL and 42.2 vs 87.8 BAU/mL, respectively). No time effect was observed for RBD-Wuhan (p = 0.173) or RBD-Delta antibodies (p = 0.376).
No significant differences were observed in the humoral response between IEIs and controls who received CoronaVac, ChAdOx1 nCoV-19 (AstraZeneca), or BNT162b2 (Pfizer-BioNTech) as their first two doses of COVID-19 vaccines (Supplementary Table S3 in the Supplementary Material).
T cell and humoral responses may be discordant and T cell responses are thought to offer some level of protection in COVID-19. We therefore assessed T cell responses in patients and controls. A similar evolution of cellular response to Nucleocapsid (p = 0.763) and Spike (p = 0.695) was observed over time for IEI and control cohorts (Figure 3, Supplementary Table S4 in the Supplementary Material). The IEI cohort presented a greater T-cell response to Spike in terms of the mean number of spots per million (SFC/106) PBMCs than did the control cohort (p = 0.002), but a similar Nucleocapsid response (p = 0.180).
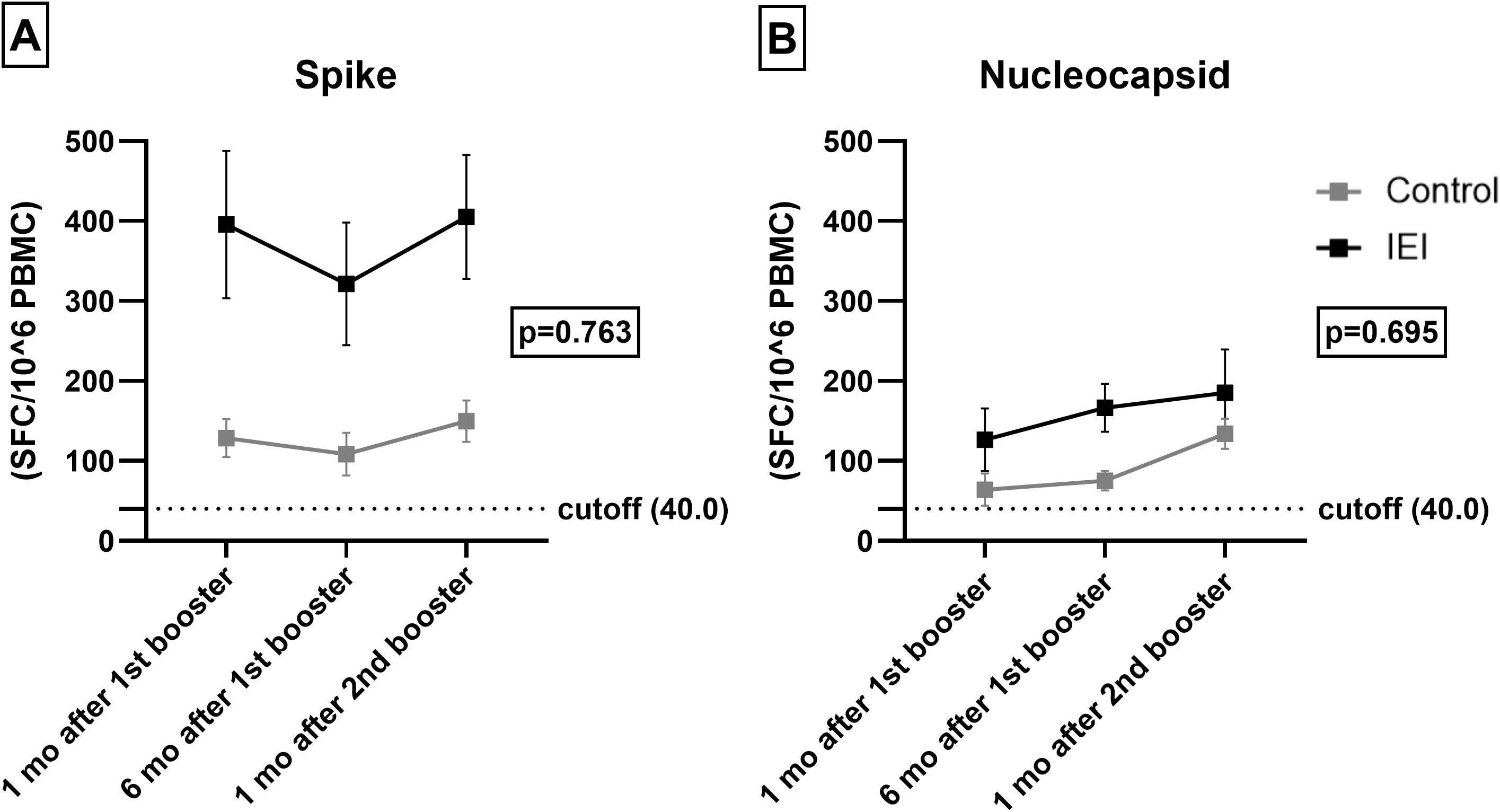
Figure 3. T-cell response. Spike (A) and Nucleocapsid (B) specific T-cell 1 and 6 months after the 1st booster and 1 month after the 2nd booster in IEI and Control. For the control cohort, the 3rd dose is the first booster and the 4th dose is the second booster. For the IEI cohort, the 4th dose is the first booster and the 5th dose is the second booster. Mean values of SFC/106 PBMC with Errors bars are shown. The dotted horizontal line is the responder cut-off of the T-SPOT COVID assay (40 SFC/106 PBMCs).
Within group analysis of change related to time revealed that the 1 and 6 months post-1st booster responses were lower than those at one month post-2nd booster (p = 0.017) for Nucleocapsid. This pattern was not observed for Spike (p = 0.445).
The vaccines received in the first two doses (CoronaVac, ChAdOx1 nCoV-19 or BNT162b2) did not affect the mean number of spots for Nucleocapsid (p = 0.554) and Spike (p = 0.554) (Supplementary Table S5 in the Supplementary Material).
The goal of vaccination is to reduce mortality and hospitalizations. We therefore assessed the severity of infection among vaccinated individuals. Among the 115 participants, 71 were found to have COVID-19 before and during the study, as assessed by SARS-CoV-2 RT−PCR or antigen test detection.
Before immunization, 9/55 (16.3%) of patients with IEI had confirmed COVID-19: 6/55 (10.9%) did not need hospitalization and 3/55 (5.4%) required hospitalization. Between the 1st and 3rd vaccine dose, 3/55 (5.4%) had confirmed COVID-19, of which 2/55 (3.6%) did not need hospitalization and 1/55 (1.8%) had to be hospitalized. Between the 3rd vaccine dose and the 4th dose, during the Omicron circulation, 21/55 (38.2%) developed SARS-CoV-2 infection and only 1/55 (1.8%) was admitted to hospital. After the 4th dose and up to 16 months, 7/49 (14.3%) had confirmed SARS-CoV-2 infection, but none had to be hospitalized (Figure 4).
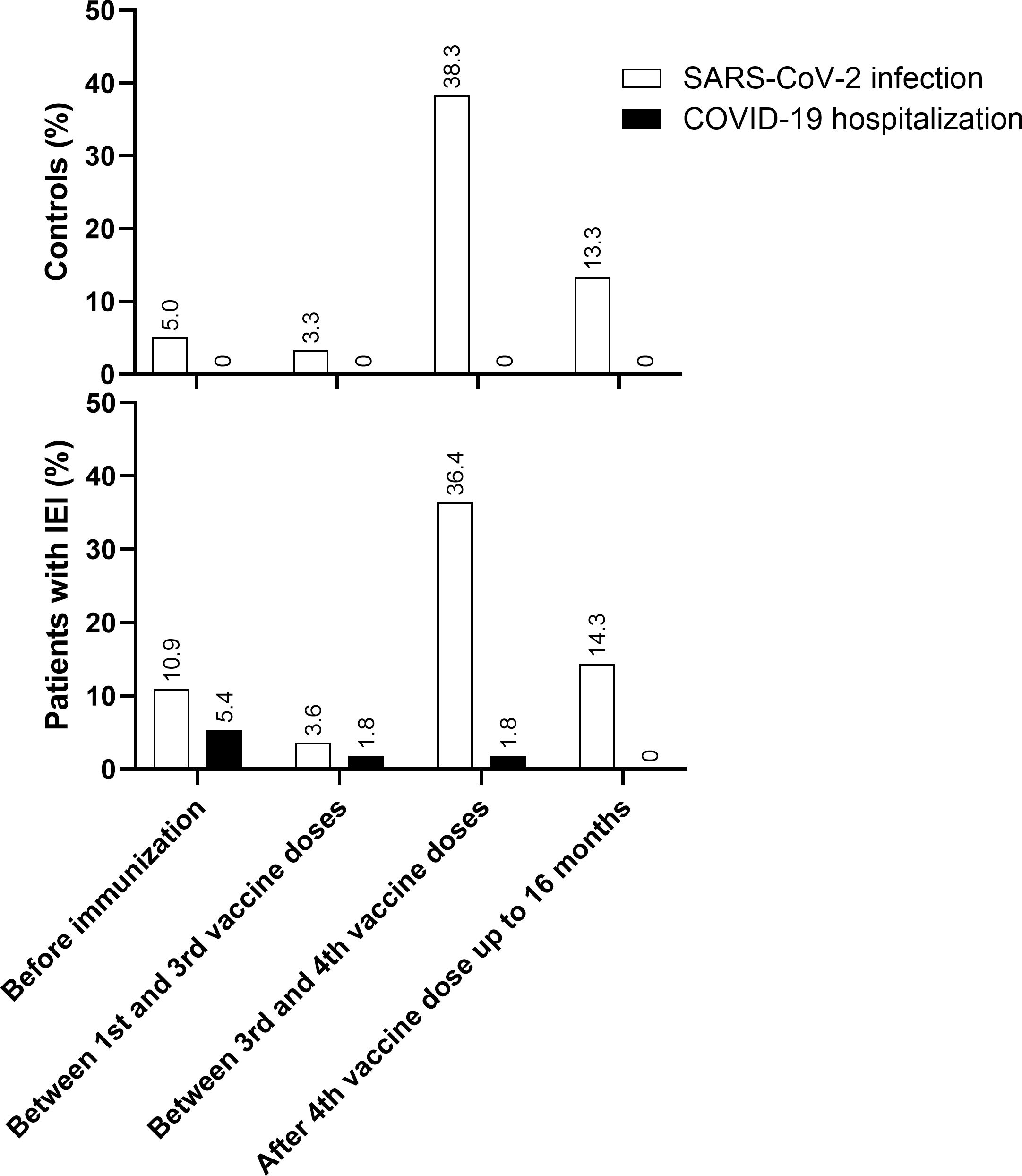
Figure 4. COVID-19 and hospitalizations. SARS-CoV-2 infection and COVID-19 hospitalization in Control and IEI cohorts before immunization, between 1st and 3rd vaccine dose, between 3rd and 4th vaccine dose and after 4th vaccine dose up to 16 months of follow-up. For the control cohort, the 3rd dose is the first booster and the 4th dose is the second booster. For the IEI cohort, the 4th dose is the first booster and the 5th dose is the second booster.
In the control group, 3/60 (5.0%) individuals developed SARS-CoV-2 infection before immunization and 2/60 (3.3%) between the 1st and 3rd vaccine dose; 23/60 (38.3%) were infected between the 3rd dose and the 4th dose, and 8/60 (13.3%) were infected after that up to 16 months of follow-up. None of them needed hospitalization (Figure 4).
The incidence of COVID-19 diagnosis was greater after the 3rd dose, when the Omicron variant was circulating in Brazil (Supplementary Figure S1 in the Supplementary Material). However, although 77.1% (27/35) of the patients with IEI and 86.1% (31/36) of the controls were diagnosed with COVID-19 after the 3rd vaccine dose, all cases were mild, except for one patient with IEI who still required hospitalization but did not need oxygen therapy or ICU admission. Thus, vaccination achieved its overall clinical goal in our IEI cohort.
As there were only 5 patients who were hospitalized in this cohort, with 2 occurring after the start of the study, it is not possible to analyze specific details of the humoral or cellular responses that might have led to a higher probability of hospitalization compared to other patients. However, a qualitative analysis was performed.
After the start of the study, one 38-year-old female patient with CVID, who had a confirmed SARS-CoV-2 infection 12 days after her first dose and very likely without antibodies in her IVIg (June 2021) required oxygen therapy and ICU admission. She had a mild reinfection 8 months later. At study admission (one month after the 3rd vaccine dose, CoronaVac/CoronaVac/BNT162b2 scheme), her immunological status was CD4+ T = 664 cells/mm3, CD8+ T = 849 cells/mm3, CD19+ = 213 cells/mm3, with neutralizing antibodies detected against all SARS-CoV-2 variants (e.g., NAb anti-RBD-BA.5 = 20,925.0 IU/mL) and a robust cellular response (Nucleocapsid T-cell response = 53 SFC/106 PBMC, Spike T-cell response = 367 SFC/106 PBMC).
The other hospitalized patient was a 29-year-old male with AT, who tested positive for SARS-CoV-2 RT-PCR in June 2022, 153 days after receiving his 3rd vaccine dose. Differently from the other patient, he did not require oxygen therapy or ICU admission. At admission to study (one month after the 3rd vaccine dose, ChAdOx1 nCoV-19/ChAdOx1 nCoV-19/BNT162b2 scheme, and 4 months before COVID-19 hospitalization), he had CD4+ T = 202 cells/mm3, CD8+ T = 79 cells/mm3, CD19+ = 6 cells/mm3. He developed a robust cellular response (Spike T-cell response = 163 SFC/106 PBMC), despite lymphopenia and was receiving IVIg, likely containing antibodies (as he had low B cell numbers, albeit lower antibody levels compared to other patients, e.g., NAb anti-RBD-BA.5 = 79.7 IU/mL). He was infected 4 months after vaccination during Omicron circulation (June 2022). Hospitalization was mainly for medical observation, without need for oxygen therapy or ICU admission.
When cellular and humoral immune responses were compared between patients with IEI and controls at three different time points (Figure 5), control individuals displayed a more homogeneous response for both antibodies and the cellular response. In contrast, patients with IEI had a much more variable humoral immune response, and some did not even reach the cutoff level for the antibody response.
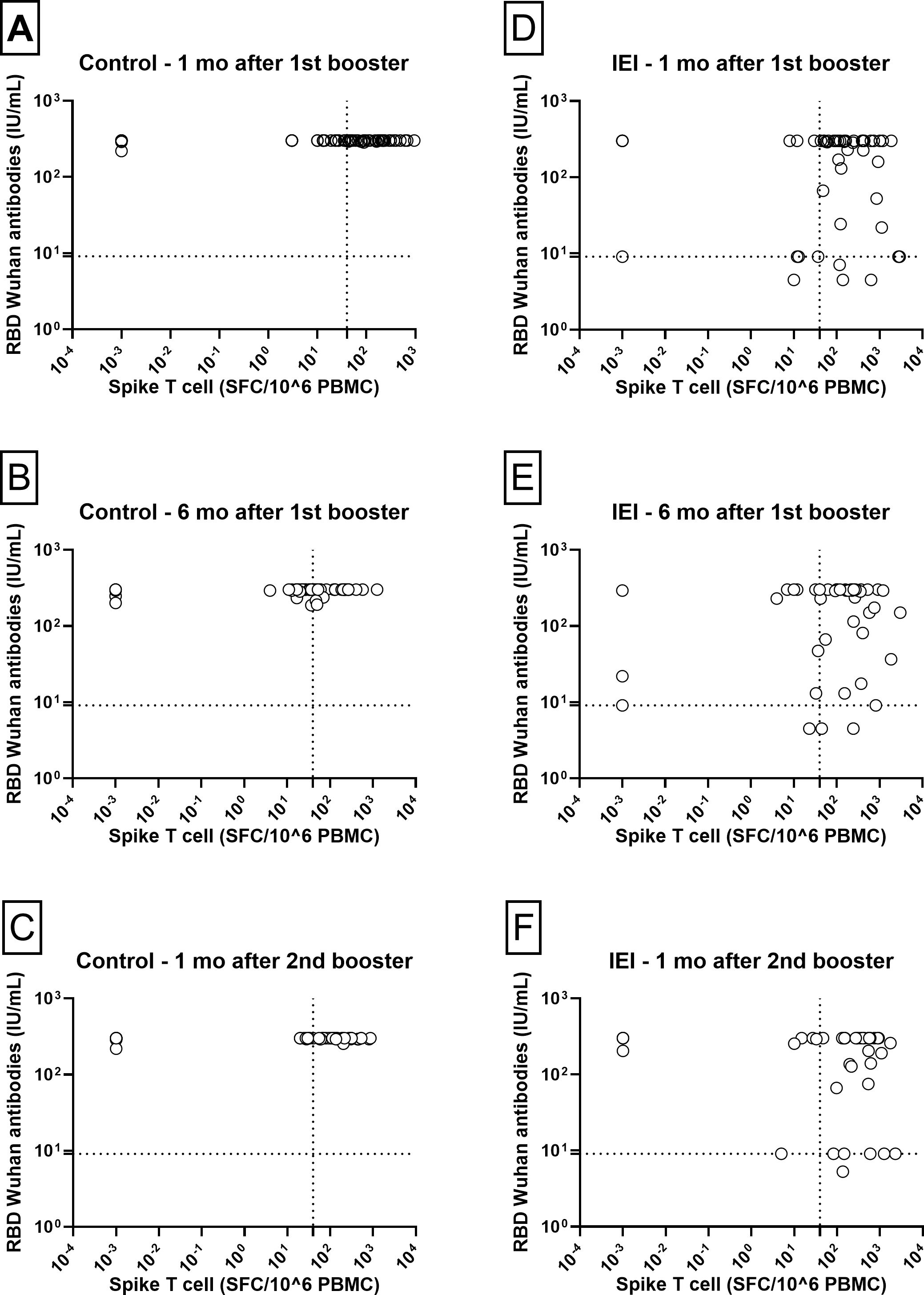
Figure 5. Humoral vs T-cell responses. Correlation between NAb RBD-Wuhan and Spike T-cell responses 1 and 6 months after the 1st booster and 1 month after the 2nd booster in Control [(A–C) respectively] and IEI [(D–F) respectively] cohorts. For the control cohort, the 3rd dose is the first booster and the 4th dose is the second booster. For the IEI cohort, the 4th dose is the first booster and the 5th dose is the second booster. The dotted horizontal line is the responder cut-off of the NTCHIP® assay for RBD-Wuhan (9.0 IU/mL). The dotted vertical line is the responder cut-off of the T-SPOT COVID assay (40 SFC/106 PBMCs).
The results presented in Figure 5 reveal two distinct subgroups of patients with IEI regarding their humoral response at the time points evaluated. At one month post-1st booster, patients with negative antibodies (below the threshold of 9.1 IU/mL) included one case of Hyper-IgM, two of CVID, one of XLA and one of Combined Immunodeficiency. Among those with a borderline response (antibody levels at threshold of 9.1 IU/mL), there were three patients with CVID, one with XLA, one with Hyper-IgM and one with Combined Immunodeficiency.
At one month post-2nd booster, one patient with CVID had negative antibodies (the same who had 24.4 IU/mL after the 1st booster). Among those with a borderline response (antibody levels at threshold of 9.1 IU/mL), there were four patients with CVID and two with XLA. Except for one XLA patient, all the other five patients were previously described at one month post-1st booster.
These patterns suggest heterogeneity in the humoral response, even among patients receiving similar vaccine regimens, with a potential impact of individual characteristics, such as the type and severity of IEI, on the ability to generate neutralizing antibodies and the presence of antibodies in the commercial immunoglobulin preparations administered to these patients.
We compared the humoral and cellular immune responses after SARS-CoV-2 booster vaccination schedules in patients with IEI to those of healthy controls. Patients with IEI responded to a three-dose SARS-CoV-2 immunization with cellular immunity similar to that of controls. However, the IEI had a lower humoral response. Boosters (4th and 5th vaccine doses for IEI group and 3rd and 4th vaccine doses for control group) increased both humoral and cellular immunity. The evaluation of the immune response to SARS-CoV-2 in individuals with IEI is essential to understand vaccine effectiveness and to identify the role of different immune components in controlling the infection.
The interaction between SARS-CoV-2 and the human immune system underscores critical aspects of the host-pathogen interplay, particularly involving innate and adaptive immunity. Type I interferons are essential during the initial stages of infection, serving as a critical line of defense against SARS-CoV-2 (25). Adaptive immunity, orchestrated by T and B cells, plays a pivotal role in modulating disease severity and facilitating viral clearance (26–29). T cells are important for containing the spread of infection, while B cells contribute by producing high-quality antibodies through affinity maturation, somatic mutations, and class switching. These processes enhance the humoral response, culminating in the production of neutralizing antibodies capable of blocking viral entry (30).
The importance of these immune mechanisms is evident in the increased susceptibility of individuals with antibody deficiencies, such as CVID or congenital agammaglobulinemia, who face significantly higher risks of severe COVID-19 and hospitalization compared to the general population (31–37). T-cell responses play a crucial role in protecting patients with IEI from severe COVID-19 and death by compensating for impaired humoral responses and providing broad, cross-reactive immunity (7–10). Although patients with IEI may show diminished responses to immunization, the administration of SARS-CoV-2 vaccines was able to protect Brazilian patients. Recent Brazilian guidelines offer SARS-CoV-2 vaccines with more recent variants for patients with IEI every 6 months (38), similar to the CDC guidelines (39).
Delmonte, Castagnoli, and Notarangelo (2022) (13) suggested that COVID-19 vaccines are effective and safe in patients with IEI, but they underscored the need for additional studies to assess the duration and robustness of immune responses to COVID-19 vaccines in patients with IEI, with larger cohorts and longitudinal assessments for more informed conclusions on vaccine efficacy in this heterogeneous population. Currently, few studies have monitored the long-term immune response to COVID-19 in immunocompromised patients. In this study, we systematically monitored the humoral and cellular immune responses after five COVID-19 vaccinations for up to 22 months.
Several studies have highlighted the role of T-cell responses in mitigating the severity of COVID-19, particularly in individuals with compromised B-cell function. Patients with B-cell deficiencies, such as those receiving anti-CD20 therapy or those with CVID, exhibit elevated T-cell responses to SARS-CoV-2 infection and vaccination. These enhanced T-cell responses, especially within the CD8+ T-cell compartment, are associated with reduced odds of severe COVID-19 (8). The highest risk patients are those with antibodies to type I interferons such as patients with thymoma or autoimmune polyendocrine syndrome type 1 (APS-1). Additional patients with inherited defects in the type I interferon pathway have also been identified, cementing this pathway as critical for the defense against SARS-CoV-2 (40, 41). Moreover, patients with IEIs who received COVID-19 mRNA vaccines demonstrated an increase in the breadth of SARS-CoV-2-specific T-cell clonotypes, even in the absence of seroconversion. These findings suggest that T-cell responses can compensate for the lack of antibody production and provide protection against severe disease (9). Additionally, the presence of cross-reactive T cells from previous exposures to common cold coronaviruses has been associated with protection from symptomatic and fatal SARS-CoV-2 infections. These cross-reactive T cells can recognize conserved epitopes across different coronaviruses, contributing to a robust immune response (10).
Following COVID-19 immunization, both patients with IEI and controls with the first three doses of original strain of BNT162b2 (Pfizer-BioNTech) had samples collected starting in January 2022 during intense circulation of the highly transmissible Omicron variant, which had some immune escape from vaccines then available (42). According to Wang et al. (2023) (42), all current circulating SARS-CoV-2 variants show high antibody evasion levels, suggesting that this is the main reason for the high transmissibility of the virus. Notably, many positive results, both in terms of the cellular response (especially to Nucleocapsid) and in the humoral response in these subgroups, may be due to asymptomatic or untested SARS-CoV-2 infections, since two-thirds of both patients and controls have not received the CoronaVac vaccine, which contains Nucleocapsid in composition.
A slight and steady increase in antibody levels was observed in patients with IEI, possibly indicating an increase related to IGRT, which contrasts with the profile observed in controls. In the control group, antibodies initially declined, followed by a significant increase after booster doses or infections (Figure 2). Thus, immunological protection against severe COVID-19 in patients with IEI with predominantly humoral dysfunction likely depends on vaccine-induced cellular responses (7–10), as well as specific antibodies generated through immunoglobulin replacement therapies (43–46). Vaccination and passive immunity from IGRT may not always be sufficient for protection. Also, new SARS-CoV-2 variants may not be neutralized with the same effectiveness (47) with the XBB1.5 variant, highlighting the importance of booster doses with new vaccines as offered twice a year by the Brazilian Ministry of Health for immunocompromised individuals since May 2024 (38).
Studies with different vaccine platforms have also shown the importance of a third dose in increasing both humoral and cellular immunity in patients with IEI (48–54). There are presently six studies that have evaluated humoral and/or cellular response after the 4th dose: three with 33, 25 and 17 patients with CVID (47, 49, 55) and three with 30, 25 and 19 patients with different IEIs (24, 48, 51). A recent study reinforced our data on the importance of boosters in increasing the percentage of patients with IEI who do not respond to the initial doses of the immunization scheme but do so after the fourth dose (47). The booster effect is of importance in the IEI community. A waning enthusiasm for vaccination and limited ongoing public health efforts directed as COVID-19 requires that clinicians caring for patients with IEI continue to promote boosters.
Before the beginning of SARS-CoV-2 immunization in Brazil, the mortality of patients with IEI was twice that reported in the general population (31). A recent USA study with 823 patients with various IEIs who experienced COVID-19 showed that those who were immunized with at least one vaccine prior to infection had significantly lower rates of hospitalization and intensive care unit admission than nonvaccinated individuals did (56). However, as shown in a Swedish study, after receiving two COVID-19 vaccine doses, patients with IEI are still at greater risk of hospitalization than the general population (57). In the present study, after COVID-19 vaccination with four doses, no patients with IEI who became infected with SARS-CoV-2 developed severe disease. However, at earlier intervals with fewer vaccines administered, there were still hospitalizations in patients with IEI. The rationale of comparing different booster schedules between IEI and Control cohorts as we proposed is the same of observed for hepatitis B vaccine, in which immunocompromised individuals should receive an additional dose to ensure adequate protection against the virus (58), once the vaccination with a 4-dose scheme can be more effective than that 3-dose scheme (59).
This study had some limitations. We did not exclude participants with prior SARS-CoV-2 infection because of the potential impact on sample size. Additionally, it was not possible to evaluate the immune response in participants before the first COVID-19 vaccine dose. Also, we cannot exclude that asymptomatic and/or untested SARS-CoV-2 infections may have occurred. This study included a heterogenous group of patients and represents a real-life cohort with mixed vaccine types administered.
Nevertheless, there are strengths in our study. To our knowledge, this is the only study that has evaluated humoral responses to Spike, Nucleocapsid, Wuhan, Delta and Omicron BA.1, BA.2, and BA.5, in addition to the cellular immune response to Nucleocapsid and Spike, until one month after the 5th COVID-19 vaccine dose (second booster for immunocompromised patients). The high rate of COVID-19 in Brazil allowed for a crucial and robust analysis of COVID-19 hospitalization after vaccination.
In summary, our study demonstrated that patients with IEI exhibit a robust specific T-cell response to SARS-CoV-2 following vaccination, similar to that of healthy controls, but with a significantly lower humoral response. Booster doses (4th and 5th for the IEI group and 3rd and 4th for the control group) enhanced both humoral and cellular immunity. Furthermore, we observed a reduction in hospital admissions due to SARS-CoV-2 infection among vaccinated IEI patients, reinforcing the clinical relevance of our findings. These findings reinforce the importance of booster strategies to improve protection in these patients, who are at higher risk of severe disease and related complications from COVID-19. It is important to acknowledge that the immune response in individuals with IEI may be more heterogeneous and complex than in healthy populations due to the diversity of underlying immune defects.
Future research directions include long-term follow-up of immune responses in patients with IEI and investigation of the role of immunoglobulin replacement therapies in modulating vaccine immune responses. Data collection on responses to new variants of SARS-CoV-2 will also be essential to understand the impact of booster doses, as well as the role of T-cell responses in protection against emerging variants.
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
The studies involving humans were approved by Comissão Nacional de Ética em Pesquisa (CONEP), Brazil (number 51535921.2.0000.5505). The studies were conducted in accordance with the local legislation and institutional requirements. Written informed consent for participation in this study was provided by the participants’ legal guardians/next of kin.
VLGS: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Project administration, Resources, Software, Validation, Visualization, Writing – original draft, Writing – review & editing. GJHS: Data curation, Formal analysis, Investigation, Methodology, Software, Visualization, Writing – review & editing. KS: Formal analysis, Supervision, Validation, Writing – review & editing. JB: Investigation, Writing – review & editing. MIHA: Investigation, Writing – review & editing. CSA: Conceptualization, Funding acquisition, Project administration, Resources, Supervision, Validation, Visualization, Writing – review & editing. MIMP: Conceptualization, Funding acquisition, Data curation, Formal analysis, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This study was financed by the São Paulo Research Foundation (FAPESP), Brasil (Process Number #2021/13419-6 (to CSA)) and by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) – Finance Code 001 (to VGLS). The article publishing charges (APCs) were funded by São Paulo Research Foundation (FAPESP), Brasil (Process Number #2020/08943-5 (to MDM-P)).
We would like to thank our patients and their families for participating in the research study. We are grateful to Prof. Daniela Santoro Rosa, PhD; Juliana Apostólico, PhD; and João Nunes for their help with the ELISpot reads. We also thank Milena Brunialti, PhD; Gabriela Ebúrneo, Janaína Vieira, Nathalia Farias, Melissa Correia and Jaqueline Muniz for their invaluable technical help, Mitti Koyama for performing the statistical analysis, the support of Prof. Ana Marli Sartori, PhD; Dr. Martin Kintrup (Viramed, Germany) and Fábio Morruzzi (NL Diagnóstica, Brazil) and all the people who contributed to this study.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2025.1538453/full#supplementary-material
ACE-2, Angiotensin-Converting Enzyme 2; AU, Arbitrary Units; BAU, Binding Arbitrary Units; BMI, Body Mass Index; CDC, Centers for Disease Control and Prevention; CVID, Common Variable Immunodeficiency; COVID-19, Coronavirus Disease 2019; ELISpot, Enzyme-Linked ImmunoSpot; GMT, Geometric Means; ICU, Intensive Care Unit; IEI, Inborn Errors of Immunity; IGRT, Immunoglobulin Replacement Therapy; IFN-γ, Interferon-Gamma; IPEX, Immune Dysregulation, Polyendocrinopathy, Enteropathy, X-Linked Syndrome; IVIg, Intravenous Human Immunoglobulin; mRNA, Messenger Ribonucleic Acid; Nab, Neutralizing Antibody; NIBSC, National Institute for Biological Standards and Control; N, Nucleocapsid; PBMCs, Peripheral Blood Mononuclear Cells; APDS 1, Activated PI3K-Delta Syndrome 1 (PIK3CD Mutation); PRNT50, Plaque Reduction Neutralization Test (50%); RBD, Receptor-Binding Domain; RT-PCR, Reverse Transcription Polymerase Chain Reaction; SARS-CoV-2, Severe Acute Respiratory Syndrome Coronavirus 2; SFC/106, Spots Forming Cells Per Million; STAT-1 GOF, Gain-of-Function Mutation in STAT-1; WHO, World Health Organization; XLA, X-linked agammaglobulinemia.
1. WHO. COVID-19 dashboard (2024). Available online at: https://data.who.int/dashboards/covid19 (Accessed February 04, 2025).
2. Bucciol G, Tangye SG, Meyts I. Coronavirus disease 2019 in patients with inborn errors of immunity: Lessons learned. Curr Opin Pediatr. (2021) 33:648–56. doi: 10.1097/MOP.0000000000001062
3. Tangye SG, Al-Herz W, Bousfiha A, Cunningham-Rundles C, Franco JL, Holland SM, et al. Human Inborn Errors of Immunity: 2022 update on the classification from the International Union of Immunological Societies Expert Committee. J Clin Immunol. (2022) 42:1473–507. doi: 10.1007/s10875-022-01289-3
4. Göschl L, Mrak D, Grabmeier-Pfistershammer K, Stiasny K, Haslacher H, Schneider L, et al. Reactogenicity and immunogenicity of the second COVID-19 vaccination in patients with inborn errors of immunity or mannan-binding lectin deficiency. Front Immunol. (2022) 13:974987. doi: 10.3389/fimmu.2022.974987
5. Erra L, Uriarte I, Colado A, Paolini MV, Seminario G, Fernández JB, et al. COVID-19 vaccination responses with different vaccine platforms in patients with inborn errors of immunity. J Clin Immunol. (2023) 43:271–85. doi: 10.1007/s10875-022-01382-7
6. Ainsua-Enrich E, Pedreño-Lopez N, Bracke C, Ávila-Nieto C, Rodríguez de la Concepción ML, Pradenas E, et al. Kinetics of immune responses elicited after three mRNA COVID-19 vaccine doses in predominantly antibody-deficient individuals. iScience. (2022) 25(11):105455. doi: 10.1016/j.isci.2022.105455
7. Moss P. The T cell immune response against SARS-CoV-2. Nat Immunol. (2022) 23:186–93. doi: 10.1038/s41590-021-01122-w
8. Zonozi R, Walters LC, Shulkin A, Naranbhai V, Nithagon P, Sauvage G, et al. T cell responses to SARS-CoV-2 infection and vaccination are elevated in B cell deficiency and reduce risk of severe COVID-19. Sci Transl Med. (2023) 15(724):eadh4529. doi: 10.1126/scitranslmed.adh4529
9. Delmonte OM, Oguz C, Dobbs K, Myint-Hpu K, Palterer B, Abers MS, et al. Perturbations of the T-cell receptor repertoire in response to SARS-CoV-2 in immunocompetent and immunocompromised individuals. J Allergy Clin Immunol. (2024) 153:1655–67. doi: 10.1016/j.jaci.2023.12.011
10. Coulon PG, Prakash S, Dhanushkodi NR, Srivastava R, Zayou L, Tifrea DF, et al. High frequencies of alpha common cold coronavirus/SARS-CoV-2 cross-reactive functional CD4+ and CD8+ memory T cells are associated with protection from symptomatic and fatal SARS-CoV-2 infections in unvaccinated COVID-19 patients. Front Immunol. (2024) 15:1343716. doi: 10.3389/fimmu.2024.1343716
11. Amodio D, Ruggiero A, Sgrulletti M, Pighi C, Cotugno N, Medri C, et al. Humoral and cellular response following vaccination with the BNT162b2 mRNA COVID-19 vaccine in patients affected by primary immunodeficiencies. Front Immunol. (2021) 12:727850. doi: 10.3389/fimmu.2021.727850
12. Hagin D, Freund T, Navon M, Halperin T, Adir D, Marom R, et al. Immunogenicity of Pfizer-BioNTech COVID-19 vaccine in patients with inborn errors of immunity. J Allergy Clin Immunol. (2021) 148:739–49. doi: 10.1016/j.jaci.2021.05.029
13. Delmonte OM, Castagnoli R, Notarangelo LD. COVID-19 and inborn errors of immunity. Physiology. (2022) 37(6):0. doi: 10.1152/physiol.00016.2022
14. Durkee-Shock JR, Keller MD. Immunizing the imperfect immune system: Coronavirus disease 2019 vaccination in patients with inborn errors of immunity. Ann Allergy Asthma Immunol. (2022) 129:562–571.e1. doi: 10.1016/j.anai.2022.06.009
15. Pham MN, Murugesan K, Banaei N, Pinsky BA, Tang M, Hoyte E, et al. Immunogenicity and tolerability of COVID-19 messenger RNA vaccines in primary immunodeficiency patients with functional B-cell defects. J Allergy Clin Immunol. (2022) 149:907–911.e3. doi: 10.1016/j.jaci.2021.11.022
16. Ricotta E, Zendt M, Castillo FB, Kelly S, Saturday T, DeGrande M, et al. Immune response to COVID-19 vaccines in people with immunodeficiencies. Res Square. (2023). doi: 10.21203/rs.3.rs-2514984/v1
17. Nadesalingam A, Cantoni D, Aguinam ET, Chan AC, Paloniemi M, Ohlendorf L, et al. Vaccination and protective immunity to SARS-CoV-2 omicron variants in people with immunodeficiencies. Lancet Microbe. (2023) 4(2):24–9. doi: 10.1016/S2666-5247(22)00297-X
18. Brazil Ministry of Health. PLANO NACIONAL DE OPERACIONALIZAÇÃO DA VACINAÇÃO CONTRA A COVID-19 . Available online at: https://www.gov.br/saude/pt-br/assuntos/covid-19/publicacoes-tecnicas/guias-e-planos/plano-nacional-de-operacionalizacao-da-vacinacao-contra-covid-19.pdf (Accessed October 24, 2024).
19. Springer DN, Traugott M, Reuberger E, Kothbauer KB, Borsodi C, Nägeli M, et al. A multivariant surrogate neutralization assay identifies variant-specific neutralizing antibody profiles in primary SARS-coV-2 omicron infection. Diagnostics. (2023) 13:2278. doi: 10.3390/diagnostics13132278
20. WHO international standard NIBSC 21/338 . Available online at: https://nibsc.org/documents/ifu/21-338.pdf (Accessed October 24, 2024).
21. Skrondal A, Rabe-Hesketh S. Generalized Latent Variable Modeling: Multilevel, Longitudinal and Structural Equation Models. Boca Raton, FL: Chapman & Hall/CRC (2004). doi: 10.1201/9780203489437
22. Gelman A, Hill J. Data Analysis Using Regression and Multilevel/Hierarchical Models. Cambridge: Cambridge University Press (2007).
23. Baerends EAM, Reekie J, Andreasen SR, Stærke NB, Raben D, Nielsen H, et al. Omicron variant-specific serological imprinting following BA.1 or BA.4/5 bivalent vaccination and previous SARS-CoV-2 infection: A cohort study. Clin Infect Dis. (2023) 77:1511–20. doi: 10.1093/cid/ciad402
24. Cheng SS, Mok CK, Li JK, Ng SS, Lam BH, Jeevan T, et al. Plaque-neutralizing antibody to BA.2.12.1, BA.4, and BA.5 in individuals with three doses of BioNTech or CoronaVac vaccines, natural infection, and breakthrough infection. J Clin Virol. (2022) 156:105273. doi: 10.1016/j.jcv.2022.105273
25. Sallard E, Lescure FX, Yazdanpanah Y, Mentre F, Peiffer-Smadja N. Type 1 interferons as a potential treatment against COVID-19. Antiviral Res. (2020) 178:104791. doi: 10.1016/j.antiviral.2020.104791
26. Diniz MO, Maini MK, Swadling L. T cell control of SARS-CoV-2: When, which, and where? Semin Immunol. (2023) 70:101828. doi: 10.1016/j.smim.2023.101828
27. Fang M, Sigal LJ. Antibodies and CD8+ T cells are complementary and essential for natural resistance to a highly lethal cytopathic virus. J Immunol. (2005) 175:6829–36. doi: 10.4049/jimmunol.175.10.6829
28. Gros L, Pelegrin M, Michaud HA, Bianco S, Hernandez J, Jacquet C, et al. Endogenous cytotoxic T-cell response contributes to the long-term antiretroviral protection induced by a short period of antibody-based immunotherapy of neonatally infected mice. J Virol. (2008) 82:1339–49. doi: 10.1128/JVI.01970-07
29. Seiler P, Bründler MA, Zimmermann C, Weibel D, Bruns M, Hengartner H, et al. Induction of protective cytotoxic T cell responses in the presence of high titers of virus-neutralizing antibodies: implications for passive and active immunization. J Exp Med. (1998) 187:649–54. doi: 10.1084/jem.187.4.649
30. Portilho AI, Gimenes Lima G, De Gaspari E. Enzyme-linked immunosorbent assay: an adaptable methodology to study SARS-coV-2 humoral and cellular immune responses. J Clin Med. (2022) 11:1503. doi: 10.3390/jcm11061503
31. Goudouris ES, Pinto-Mariz F, Mendonça LO, Aranda CS, Guimarães RR, Kokron C, et al. Outcome of SARS-coV-2 infection in 121 patients with inborn errors of immunity: A cross-sectional study. J Clin Immunol. (2021) 41:1479–89. doi: 10.1007/s10875-021-01066-8
32. Delavari S, Abolhassani H, Abolnezhadian F, Babaha F, Iranparast S, Ahanchian H, et al. Impact of SARS-coV-2 pandemic on patients with primary immunodeficiency. J Clin Immunol. (2021) 41:345–55. doi: 10.1007/s10875-020-00928-x
33. Milito C, Lougaris V, Giardino G, Punziano A, Vultaggio A, Carrabba M, et al. Clinical outcome, incidence, and SARS-CoV-2 infection-fatality rates in Italian patients with inborn errors of immunity. J Allergy Clin Immunology: In Practice. (2021) 9:2904–2906.e2. doi: 10.1016/j.jaip.2021.04.017
34. Pulvirenti F, Salinas AF, Milito C, Terreri S, Mortari EP, Quintarelli C, et al. B cell response induced by SARS-CoV-2 infection is boosted by the BNT162b2 vaccine in primary antibody deficiencies. Cells. (2021) 10:2915. doi: 10.3390/cells10112915
35. Shields AM, Burns SO, Savic S, Richter AG, Anantharachagan A, Arumugakani G, et al. COVID-19 in patients with primary and secondary immunodeficiency: The United Kingdom experience. J Allergy Clin Immunol. (2021) 147:870–875.e1. doi: 10.1016/j.jaci.2020.12.620
36. Ponsford MJ, Shillitoe BMJ, Humphreys IR, Gennery AR, Jolles S. COVID-19 and X-linked agammaglobulinemia (XLA) - Insights from a monogenic antibody deficiency. Curr Opin Allergy Clin Immunol. (2021) 21:525–34. doi: 10.1097/ACI.0000000000000792
37. Nguyen AA, Habiballah SB, LaBere B, Day-Lewis M, Elkins M, Al-Musa A, et al. Rethinking immunological risk: A retrospective cohort study of severe SARS-CoV-2 infections in individuals with congenital immunodeficiencies. J Allergy Clin Immunol Pract. (2023) 11:3391–3399.e3. doi: 10.1016/j.jaip.2023.07.042
38. Brazil Ministry of Health. Informe técnico - vacina COVID-19 XBB. Available online at: https://www.gov.br/saude/pt-br/vacinacao/informes-tecnicos/informe-tecnico-vacina-covid-xbb (Accessed October 24, 2024).
39. CDC. Interim clinical considerations for COVID-19 vaccines in the United States . Available online at: https://www.cdc.gov/vaccines/covid-19/clinical-considerations/interim-considerations-us.html (Accessed October 24, 2024).
40. Bastard P, Rosen LB, Zhang Q, Michailidis E, Hoffmann HH, Zhang Y, et al. Autoantibodies against type I IFNs in patients with life-threatening COVID-19. Science. (2020) 370:eabd4585. doi: 10.1126/science.abd4585
41. Zhang Q, Bastard P, Liu Z, Le Pen J, Moncada-Velez M, Chen J, et al. Inborn errors of type I IFN immunity in patients with life-threatening COVID-19. Science. (2020) 370:eabd4570. doi: 10.1126/science.abd4570
42. Wang Q, Li Z, Guo Y, Mellis IA, Iketani S, Liu M, et al. Evolving antibody evasion and receptor affinity of the Omicron BA. 2.75 sublineage SARS-CoV-2 iScience. (2023) 26:108254. doi: 10.1016/j.isci.2023.108254
43. Karbiener M, Kindle G, Meyts I, Seppänen MRJ, Candotti F, Kamieniak M, et al. Clinical efficacy of SARS-CoV-2 Omicron-neutralizing antibodies in immunoglobulin preparations for the treatment of agammaglobulinemia in patients with primary antibody deficiency. J Med Virol. (2024) 96(6):e29738. doi: 10.1002/jmv.29738
44. Upasani V, Townsend K, Wu MY, Carr EJ, Hobbs A, Dowgier G, et al. Commercial immunoglobulin products contain neutralizing antibodies against severe acute respiratory syndrome coronavirus 2 spike protein. Clin Infect Dis. (2023) 77:950–60. doi: 10.1093/cid/ciad368
45. Zimmerman O, Altman Doss AM, Ying B, Liang CY, Mackin SR, Davis-Adams HG, et al. Immunoglobulin replacement products protect against SARS-CoV-2 infection in vivo despite poor neutralizing activity. JCI Insight. (2024) 9(3):e176359. doi: 10.1172/jci.insight.176359
46. Milota T, Smetanova J, Zelena H, Rataj M, Lastovicka J, Bartunkova J. Content and specificity of the Anti-SARS-CoV-2 antibodies in solutions for immunoglobulin replacement therapy. Int Immunopharmacol. (2023) 125:111159. doi: 10.1016/j.intimp.2023.111159
47. Gutiérrez-Bautista JF, Díaz-Alberola I, Tarriño M, Aguilera M, Cobo F, Reguera JA, et al. Follow-up of immune response in patients with common variable immunodeficiency following SARS-CoV-2 vaccination. Clin Exp Immunol. (2024) 217:253–62. doi: 10.1093/cei/uxae039
48. Hurme A, Jalkanen P, Marttila-Vaara M, Heroum J, Jokinen H, Vara S, et al. T cell immunity following COVID-19 vaccination in adult patients with primary antibody deficiency – a 22-month follow-up. Front Immunol. (2023) 14:1146500. doi: 10.3389/fimmu.2023.1146500
49. Nielsen BU, Drabe CH, Barnkob MB, Johansen IS, Hansen AKK, Nilsson AC, et al. Antibody response following the third and fourth SARS-CoV-2 vaccine dose in individuals with common variable immunodeficiency. Front Immunol. (2022) 13:934476. doi: 10.3389/fimmu.2022.934476
50. Shin JJ, Par-Young J, Unlu S, McNamara A, Park HJ, Shin MS, et al. Defining clinical and immunological predictors of poor immune responses to COVID-19 mRNA vaccines in patients with primary antibody deficiency. J Clin Immunol. (2022) 42:1137–50. doi: 10.1007/s10875-022-01296-4
51. Ibrahim KY, Moreira RM, Santos CFD, Strabelli TMV, Belizário JC, de-Moraes-Pinto MI, et al. Immunogenicity of COVID-19 adsorbed inactivated vaccine (CoronaVac) and additional doses of mRNA BNT162b2 vaccine in immunocompromised adults compared with immunocompetent persons. Rev Inst Med Trop Sao Paulo. (2024) 66:e24. doi: 10.1590/S1678-9946202466024
52. Movahedi M, Movahedi M, Parvaneh N, Abolhassani H, Mahdavi M, Mosavikhorshidi MS, et al. Antibody response before and after the booster dose of inactivated corona vaccine in antibody deficient patients. Iran J Allergy Asthma Immunol. (2024) 23:158–67. doi: 10.18502/ijaai.v23i2.15322
53. van Leeuwen LPM, Grobben M, GeurtsvanKessel CH, Ellerbroek PM, de Bree GJ, Potjewijd J, et al. Immunogenicity of COVID-19 booster vaccination in Patients with IEI and their one year clinical follow-up after start of the COVID-19 vaccination program. Front Immunol. (2024) 15:1390022. doi: 10.3389/fimmu.2024.1390022
54. Rocco JM, Boswell KL, Laidlaw E, Epling B, Anderson M, Serebryannyy L, et al. Immune responses to SARS-CoV-2 mRNA vaccination in people with idiopathic CD4 lymphopenia. J Allergy Clin Immunol. (2024) 153:503–12. doi: 10.1016/j.jaci.2023.10.012
55. Piano Mortari E, Pulvirenti F, Marcellini V, Terreri S, Salinas AF, Ferrari S, et al. Functional CVIDs phenotype clusters identified by the integration of immune parameters after BNT162b2 boosters. Front Immunol. (2023) 14:1194225. doi: 10.3389/fimmu.2023.1194225
56. McDonnell J, Cousins K, Younger MEM, Lane A, Abolhassani H, Abraham RS, et al. COVID-19 vaccination in patients with inborn errors of immunity reduces hospitalization and critical care needs related to COVID-19: a USIDNET report. J Clin Immunol. (2024) 44:86. doi: 10.1007/s10875-023-01613-5
57. Lindahl H, Kahn F, Nilsdotter-Augustinsson Å, Fredrikson M, Hedberg P, Möller IK, et al. Inborn errors of immunity are associated with increased COVID-19-related hospitalization and intensive care compared to the general population. J Allergy Clin Immunol. (2024) 155(2):387–97.e6. doi: 10.1016/j.jaci.2024.10.013
58. Terrault NA, Lok ASF, McMahon BJ, Chang KM, Hwang JP, Jonas MM, et al. Update on prevention, diagnosis, and treatment of chronic hepatitis B: AASLD 2018 hepatitis B guidance. Hepatology. (2018) 67:1560–99. doi: 10.1002/hep.29800
Keywords: COVID-19 vaccines, booster, inborn errors of immunity, primary immunodeficiency disorders, SARS-CoV-2, microarray, immune response, ELISpot enzyme-linked immunospot
Citation: Lopes da Silva VG, Schmitz GJH, Sullivan KE, Barbate J, de Haro Azinar MI, Aranda CS and de Moraes-Pinto MI (2025) Enhanced T-cell immunity and lower humoral responses following 5-dose SARS-CoV-2 vaccination in patients with inborn errors of immunity compared with healthy controls. Front. Immunol. 16:1538453. doi: 10.3389/fimmu.2025.1538453
Received: 02 December 2024; Accepted: 06 February 2025;
Published: 06 March 2025.
Edited by:
Ritthideach Yorsaeng, Chulalongkorn University, ThailandReviewed by:
Emily S. J. Edwards, Monash University, AustraliaCopyright © 2025 Lopes da Silva, Schmitz, Sullivan, Barbate, de Haro Azinar, Aranda and de Moraes-Pinto. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Maria Isabel de Moraes-Pinto, bS5pc2FiZWxtcEBnbWFpbC5jb20=; Vitor Gabriel Lopes da Silva, dml0b3IubG9wZXNAdW5pZmVzcC5icg==
†ORCID: Maria Isabel de Moraes-Pinto, orcid.org/0000-0001-7345-5477
Gabriela Justamante Händel Schmitz, orcid.org/0000-0002-7069-1017
Kathleen E. Sullivan, orcid.org/0000-0002-3577-9021
Júlia Barbate Pintão, orcid.org/0000-0002-6213-2573
Carolina Sanchez Aranda, orcid.org/0000-0002-2187-8192
Maria Isabel de Moraes-Pinto, orcid.org/0000-0001-7345-5477
Vitor Gabriel Lopes da Silva, orcid.org/0000-0002-8802-8117
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.