 Maarten P. M. Vanhove1,2*
Maarten P. M. Vanhove1,2* Stephan Koblmüller3
Stephan Koblmüller3 Jorge M. O. Fernandes4
Jorge M. O. Fernandes4 Christoph Hahn3*
Christoph Hahn3* Michelle Plusquin5
Michelle Plusquin5 Nikol Kmentová1,2,6
Nikol Kmentová1,2,6- 1Research Group Zoology: Biodiversity and Toxicology, Centre for Environmental Sciences, Hasselt University, Diepenbeek, Belgium
- 2International Union for Conservation of Nature (IUCN) Species Survival Commission (SSC) Parasite Specialist Group, Diepenbeek, Belgium
- 3Institute of Biology, University of Graz, Graz, Austria
- 4Renewable Marine Resources Department, Institut de Ciències del Mar, Spanish National Research Council, Barcelona, Spain
- 5Research Group Environmental Biology, Center for Environmental Sciences, Hasselt University, Diepenbeek, Belgium
- 6Freshwater Biology, Operational Directorate Natural Environment, Royal Belgian Institute of Natural Sciences, Brussels, Belgium
The “Old Friends Hypothesis” suggests insufficient exposure to symbionts hinders immune development, contributing to increased immune-related diseases in the Global North. The microbiome is often the focus; helminths, potentially also offering health benefits, lack attention. Infection and effect of helminths are influenced and perhaps determined by micro-organisms. Mechanisms behind parasite-microbiome interactions are poorly understood, despite implications on host health. These interactions are typically studied for single helminth species in laboratory animal models, overlooking helminth diversity. Reviewing research on relationships between helminth and microbial diversity yielded 27 publications; most focused on human or other mammalian hosts, relying on natural exposure rather than experimental helminth inoculation. Only about half investigated host health outcomes. Remaining knowledge gaps warrant considering additional candidate model systems. Given the high helminthiasis burden and species diversity of helminths, we propose seeking models in the Global South, where a considerable proportion of research on diversity aspects of helminth-microbiome interactions took place. Low availability of genomic resources for helminths in the Global South, however, necessitates more integrative helminthological research efforts. Given substantial similarities in immune systems, several fishes are models for human health/disease. More effort could be done to establish this for cichlids, whose representatives in the African Great Lakes provide a well-delineated, closed natural system relevant to human health in view of fish-borne zoonoses and other water-borne parasites. A good baseline exists for these cichlids’ genomics, parasitology, and microbiology. We suggest exploring African Great Lake cichlids as model hosts for interactions between microbial diversity, helminth diversity, and host health.
1 Introduction
From birth onwards, humans are colonized by microorganisms including fungi, protozoans and viruses (1). In addition, humans have coexisted with helminths for most of our existence (2). Yet, since the industrial era, countries in the Global North have established a reduction in prevalence of helminths and other parasites (3, 4). Additionally, studies have observed an association between industrial lifestyles and reduced diversity in the gut microbiome (5). Allergic disorders and inflammatory conditions increased throughout the 20th century in the Global North (6, 7). From the late 19th century, hay fever was increasingly reported in wealthy urban populations, while farming communities remained less affected (8). As living conditions have changed, these trends indicate a disturbance in immune regulation, leading to a disbalanced response to harmless allergens or infections.
The “Old Friends Hypothesis” provides an evolutionary view on the rise of immune-related disorders like allergies, asthma, and autoimmune diseases, including a focus on early life (9). It describes that the absence or reduced exposure to specific microorganisms or helminths in early life, which co-evolved with humans as hunter-gatherer omnivores, disrupts normal immune system development. This may lead to a disturbance of immunoregulation due to insufficient exposure to microorganisms or helminths that drive the expansion of components such as regulatory T cells (10). The hypothesis suggests that exposures to specific organisms are critical for properly training the immune system to differentiate between harmful and benign antigens, and preventing excessive immune responses. For example, lipopolysaccharide, a component of the outer membrane of gram-negative bacteria, may protect against allergic responses by inducing the ubiquitin-modifying enzyme A20 in lung epithelial cells (11). Also, helminths seem to fit the narrative of the “Old Friends Hypothesis”. For instance, helminth infections may also confer protective benefits against autoimmune diseases, allergies, and inflammatory disorders by promoting immune tolerance through immune system modulation and microbiome alterations (12–14). In laboratory animal models, helminth infections have been linked to suppression of pro-inflammatory gut bacteria, leading to protection against Crohn’s disease in genetically susceptible mice (15). Similarly, in humans, helminth infections have been shown to increase microbial diversity and alter antibody levels, although the full extent of these effects is still not well understood (16). Infection with pinworms (Enterobius vermicularis) was shown to be associated with a protective effect on the development of clinical malaria in Tanzanian children, while infection with hookworm exacerbates the severity of malaria (17).
Several studies indicate that helminth infections may have a substantial effect and change the diversity and composition of gut microbiota (18–21). These shifts in the microbiome could affect the risk for various diseases, such as asthma, viral infections, and metabolic disorders (22, 23). Furthermore, the microbiome plays a role in the establishment of certain helminth infections and can influence their progression (24). Specific bacterial taxa have been found to affect an individual’s resistance or susceptibility to helminths (25), though the precise mechanisms underlying these helminth-microbiota interactions remain largely unclear. Despite the promising findings that infection with helminth parasites alleviates symptoms of various diseases, our understanding of the mechanisms underlying the interaction between parasites and the microbiome remains limited.
2 The study of microbiome-parasite interactions needs more attention for parasite and host diversity, which relies on natural exposure
Despite their interconnected roles in host health, microbes and parasites have traditionally been studied in isolation, overlooking their collective contribution as part of a “symbiome”. We regard the term “symbiont” (and collectively “symbiome”) as referring to any host-associated organisms (including viruses), which can have negative, neutral, or positive interactions with the host (26).
According to the holobiont concept (27, 28), the symbionts and the host act as a unique biological entity termed “holobiont”, a unit of selection in evolution (29). Consequently, the “hologenome” is the collective of genomes of host and symbiome, with interactions between hologenome and environment determining phenotype and being pivotal to adaptation and evolution (30, 31). Symbiome genomes can change rapidly under environmental stress and therefore buy the host precious time to adapt and survive (32). Hologenomic approaches might be applied as part of the One Health approach for in-depth ecosystem health assessments in real-time (33) and in aquaculture to enhance growth, health and overall production sustainability (34). Unfortunately, in spite of being an integral part of the holobiont, the role of parasites in this intricate evolutionary unit has been largely ignored.
Parasites, including helminths, inhabit diverse niches within their hosts, where they inevitably encounter a plethora of commensal, mutualistic and pathogenic microorganisms. These interactions can significantly influence the host immune response, metabolism, overall health and development, both in animal models and humans (12, 13, 35, 36). Despite recent progress (37–39), our understanding of the molecular and cellular mechanisms governing parasite-microbiome dynamics remains limited. Addressing this gap is crucial, as the interplay between parasites and the microbiome might have far-reaching effects on progression of diseases, effectiveness of therapeutic interventions, and development of novel treatments.
Helminths secrete various immunomodulatory molecules that can impact the host immune system directly or alter the composition and function of the (gut) microbiota (14, 40), potentially creating environments more favorable for the parasite while dampening host immune responses. Conversely, certain microbiota may influence parasite colonization and pathogenicity, implying a bidirectional relationship (38). Exploring these parasite-microbiome interactions is essential not only for advancing our understanding of parasitology but also for unlocking new avenues in microbiome research and therapeutic innovation. These may lead to novel microbiome-targeted strategies for managing parasitic infections and novel interventions/treatments that harness the beneficial effects of helminths. Unfortunately, research on the microbiome associated with helminths or other parasites has not kept track with the general surge in microbiome studies (41).
The importance of microbial community composition and the beneficial role of a diverse microbiome on health outcomes of animals, plants, and ecosystems are well-established (e.g., 42, 43). For instance, they may determine the resilience of a host or ecosystem to environmental perturbations (44, 45). The literature on helminth-microbiota interactions, however, typically focuses on laboratory-kept hosts and single-species infections of helminths (46), overlooking the aspect of parasite diversity at the species and genomic level. To assess the extent to which diversity is taken into account in the study of microbiome-helminth interactions, we performed a search on Web of Science. The search for research on the relationship between host helminth diversity and microbial diversity, resulted in 244 initial hits (Supplementary Table 1) and was filtered to 27 relevant studies based on specific criteria such as focus on empirical work, relevance to parasitic worms, and inclusion of helminth diversity, while categorizing these studies by host taxa, health outcomes, and type of helminth exposure (natural or experimental) (Figure 1). Twenty-seven publications were retained, which indicates that only a minority of studies on microbiome-parasitic worm interactions includes aspects of helminth (species) diversity. Several studies mentioned the presence of multiple species of helminths or other parasite taxa, while investigating the microbial interactions with a single one (47–49). Figure 1 details the state of empirical research about the relationship between the diversity of helminths of a host and microbial diversity. Out of the 27 retained studies, 22 investigated human or other mammal hosts (Figure 1A). Only just over half of the studies verified health outcomes for the host (Figure 1B). Only one publication investigated helminth infections resulting from experimental rather than natural exposure (Figure 1C), and a single study focused on the microbiome of helminths rather than of the host itself (Figure 1D).
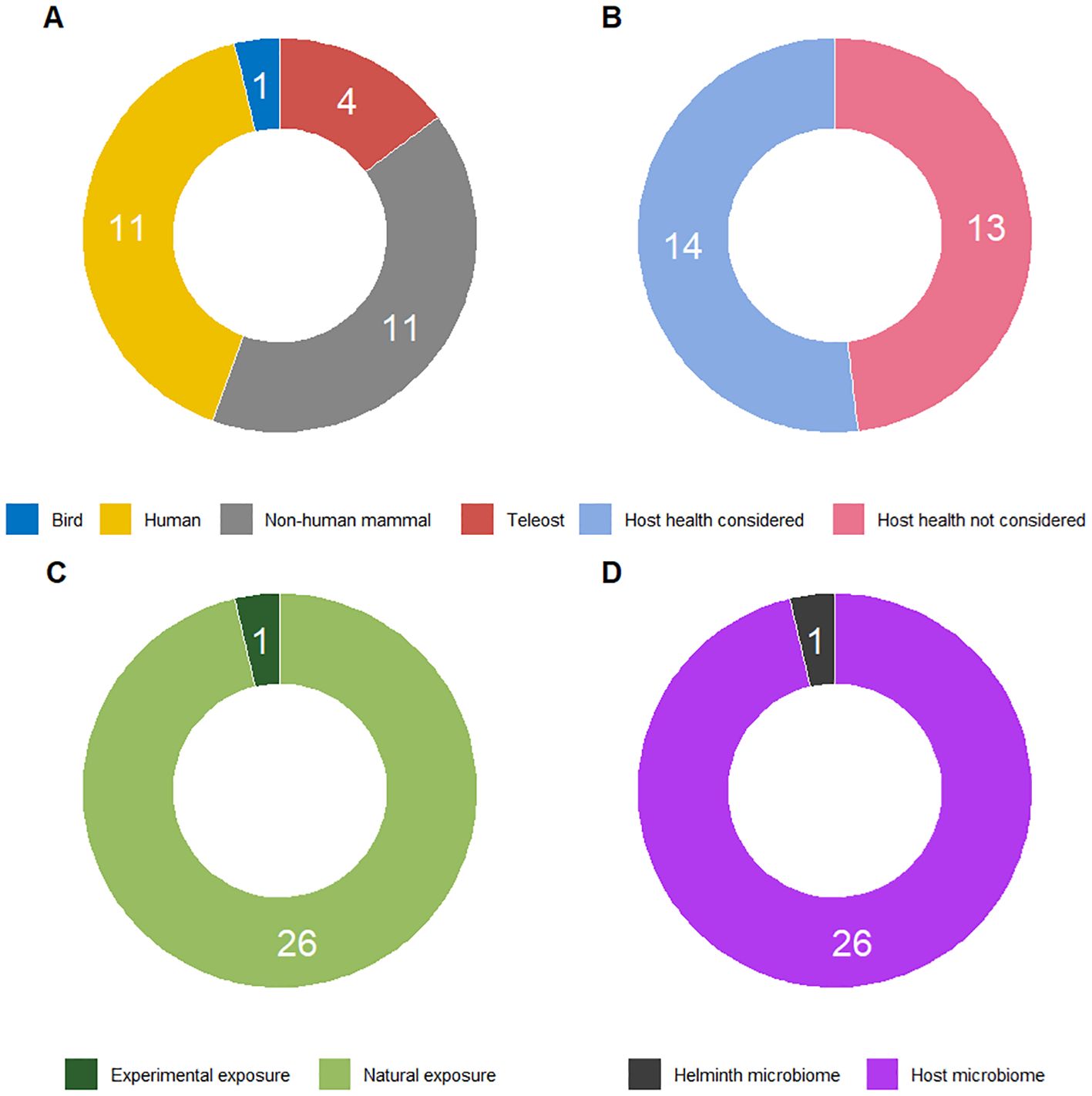
Figure 1. Characterisation of empirical research about the relationship between the diversity of helminths of a host and microbial diversity, based on the following Web of Science search string on 27 August 2024: “diversity AND microb*” (Topic) NOT “Review OR Editorial Material” (Document Type) AND “helminth OR trematod* OR fluke OR cestod* OR tapeworm OR digenean OR monogen* OR nematod* OR acanthocephalan OR worm OR leech OR hirud*” (Topic) AND “health” (Topic). From the resulting 244 hits (Supplementary Table 1), 27 were retained, and 179 were not considered either because they did not contain empirical work (they were e.g. a review paper, perspective paper, proceedings paper, database paper, meta-analysis, or methodological paper), or because they did not deal with parasitic worms at all, or only studied helminths in the context of plant parasites, environmental samples, or waste(water). A further 38 were not retained because helminth diversity was not taken into consideration, i.e. these studies either focused on a single species, considered all helminth infections together, or only lumped several species into a single higher-order taxon. For the 27 studies retained, (A) shows the distribution over focal host taxa; (B) indicates whether studies look at health outcomes for the host, which we considered to be the case when host health was empirically studied, beyond the mere presence or clearance of infection; (C) compares whether studies applied experimental infection or relied on natural exposure to helminths; and (D) illustrates the proportion of studies focusing on the microbiome of helminths rather than on the microbiome of their hosts.
The influence of the host’s environment and behavior on its symbionts makes it hard to mimic a real-life symbiome, or natural exposure to symbionts (50), experimentally. Indeed, the microbiomes of conspecific natural and captive (including laboratory) animal populations are known to differ (e.g., 51, 52), as do the microbial and parasite load between rural and urban populations (53–55). Moreover, in humans and other animals, lifestyle and environment are key factors in determining microbiome composition (56–60). Therefore, approaches going beyond laboratory conditions gained traction in immunology (61) and may be highly relevant to studying the symbiome. For instance, releasing test animals into a semi-natural environment, or oral inoculation, may allow fungal colonization enriching the intestinal microbiota (62). Such approaches are also promising when studying helminth infections and their immune responses: they allow taking into account host and environmental factors that facilitate translation to a natural context. Hence, although experiments are definitely needed to reveal the mechanisms behind helminth-microbiome interactions (63), it remains hard to include the diversity of helminths into experimental approaches. Unsurprisingly, in our literature sample which excluded single-helminth species studies, only a single study applied experimental co-infection: Bonde et al. (64) inoculated pigs with the nematodes Ascaris suum and Oesophagostomum dentatum. This clearly illustrates that investigations of helminth-microbiota interactions rely on natural host populations when the entire helminth community is to be considered. Unfortunately, such studies are scarce (46) and it is timely to propose additional suitable candidate models to study the health and disease outcomes of helminth-microbiome interactions.
3 Candidate models to study helminth-microbiome interactions should be found in the Global South
We mapped the geographical distribution of the helminth-microbiome studies which take into account the diversity of helminths (resulting from the above-mentioned Web of Science search). Figure 2 shows where the empirical research took place, across the regional groupings in the framework of the Sustainable Development Goals1. We compared this with the state of exploration of helminth diversity and the availability of genetic resources (Figure 2A) (65), and with the prevalence of cysticercosis (caused by the tapeworm Taenia solium), as a proxy for the burden of helminthiases (Figure 2B).
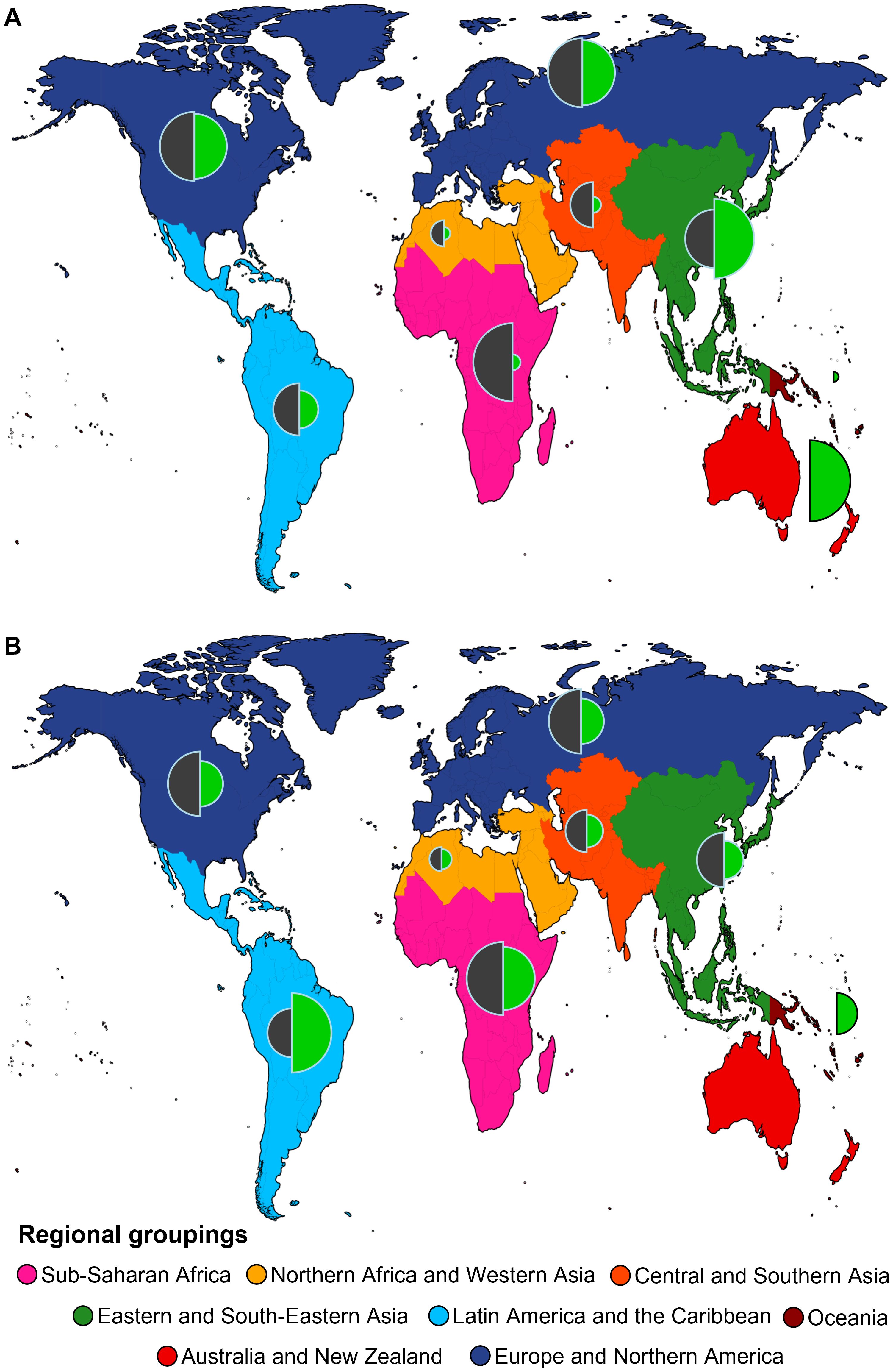
Figure 2. Global distribution of empirical research about the relationship between the diversity of helminths of a host and its microbial diversity (see legend of Figure 1 for search string) across regional groupings used for indicators and reporting regarding the Sustainable Devevelopment Goals, with colours on the map denoting different regional groupings (https://unstats.un.org/sdgs/indicators/regional-groups/). (A) Proportional research effort on helminth and microbial diversity (proportional to the radius of the left half-circles) compared to recent research effort on helminths relative to their known diversity in a given regional grouping (proportional to the radius of the right half-circles). For the latter, we divided the number of available nucleotide sequences on NCBI GenBank per region, by the aggregated total for the region of per-country data of host-helminth records, based on the data and analyses from Poulin et al. (65). This provides an estimate of genetic research effort relative to the regional helminth diversity; in a region where this proportion is low, the helminth diversity has not been adequately studied with recent molecular tools. (B) Proportional research effort on helminth and microbial diversity (proportional to the radius of the left half-circles) compared to aggregated per-country prevalence of cysticercosis (proportional to the radius of the right half-circles) (per 100 000 people, based on data from the Global Burden of Disease study, Institute for Health Metrics and Evaluation, taken from https://ourworldindata.org/grapher/prevalence-cysticercosis). We opted to use data on cysticercosis here, as it is a neglected tropical disease that is also transmitted locally in countries of the Global North, and hence not only a public health challenge in the Global South (66, 67).
While the total number of studies retained in our search is low, they appear quite evenly spread across the globe (Figure 2). It is noteworthy that the Global South has seen a comparable amount of research attention in this field. In this region the known and estimated helminth diversity are highest (Figure 2A), as is the maximal expected proportion of undiscovered helminth species (68). It is also where the highest cysticercosis prevalences are found (Figure 2B). Indeed, although next to cysticercosis there are other helminthiases that occur in the Global North and South (e.g., ascariasis caused by nematode species belonging to Ascaris: 69), the highest toll of helminthiases is typically found in the Global South and Africa in particular (70, 71). Therefore, it is fitting that the latter continent is well-covered in certain areas of helminthology, such as geospatial mapping of human helminthiasis (72).
On the other hand, the relative lack of genetic resources in many parts of the Global South, especially sub-Saharan Africa (Figure 2A) suggests that its considerable helminth diversity has been insufficiently studied, particularly with regard to modern integrative approaches. This complicates helminth microbiome research and its translation into practice, as it limits, for instance, the possibility to diagnose and identify species and strains by molecular means. Globally, metabarcoding initiatives have done little to mitigate this situation, as they contributed rather to our understanding of occurrences of different microbial (e.g., 73) and helminth communities (reviewed in 74) than patterns of their potential reciprocal interactions. Thus, we commend calls for more parasitological research in the Global South (75); needless to say, this includes capacity development and technology transfer (e.g., 76). In particular, the situation in Africa shows the highest contrast between the available helminth genetic resources and helminth disease burden (Figure 2) (see also 77). We believe that work on how helminth diversity patterns influence helminth-microbe interactions therefore also requires more multidisciplinary attention for helminths, including sequencing efforts focusing on host populations under natural exposure.
4 Proposal for a fish host model to study helminth-microbiome interactions in real-life settings
Fish models provide effective platforms for understanding the mechanisms and developing therapeutic agents for various human conditions, for instance cancer, cardiovascular, bone, renal and blood diseases (78), and hypercholesterolemia (79). They are also well-established for infectious diseases (80–82), including those directly influenced by microbial communities (83). A key advantage is that fish gut microbiomes can be manipulated to establish the link between host symbiome diversity, health and disease (84). Alterations of the gut microbiota in fish can be achieved directly by fecal material transplant (85) or through indirect approaches, such as diet and dietary inclusion of prebiotics and probiotics (86, 87) as well as antimicrobials (84). Vargas-Albores et al. (88), Luna et al. (89) and Zhang et al. (90) provided overviews of possibilities and knowledge gaps. For example, Rimoldi et al. (91) showed that diets containing insect meal of Hermetia illucens significantly increased the abundance of beneficial bacteria in the gut and improved intestinal health in gilthead seabream (Sparus aurata) farmed inshore.
Several factors contribute to the establishment of many (tropical) fish species as invaluable laboratory models, namely short generation time (enabling individual monitoring throughout lifespans in experimental settings; in some species even multigenerational studies are possible in the time frame of typical research projects), high fecundity rate and external development, which facilitate genetic and functional studies. Moreover, genome assemblies of many fish species are publicly available2. Overall, the gene complements of fishes and tetrapods, including humans, are sufficiently conserved to allow insights into molecular mechanisms obtained in the former to inform our understanding in the latter, including aspects of the immune system. Remarkably, approximately 82% of the genes that have been linked to human diseases have counterparts (i.e., orthologues) in the zebrafish (Danio rerio) genome (92). Overall, these features facilitate possibilities of precision genome editing to design human disease models. Tessadori et al. (93) introduced patient alleles in their zebrafish orthologues to model specific cardiovascular disorders, enabling the development of novel therapeutic strategies.
The zebrafish is the most widely recognized fish model of human diseases (78) but not the only one. Teleost fishes, the largest and most diverse group of vertebrates, exhibit an extraordinary range of specialised phenotypes that reflect their adaptation to a wide array of ecological niches (94). Due to its unique life cycle, the turquoise killifish (Nothobranchius furzeri), for instance, has emerged as a model for studying ageing and age-related neurological diseases (95). Medaka (Oryzias latipes) serves as a model for neurodegenerative conditions like Parkinson’s disease (96). Recently, unusual fish evolutionary mutant models have gained popularity for phenotypes that mimic maladaptive human diseases but are beneficial for the species’ adaptation to its environment (97). Elephantfishes (mormyrids) and knifefishes (gymnotiforms) have experienced numerous natural mutations in the ion channel gene scn4aa, making them excellent species for modelling channelopathies affecting skeletal muscle (98). Other examples of fish evolutionary mutant models include the hybrid Xiphophorus (X. maculatus x X. helleri) (99), the mummichog (Fundulus heteroclitus) (100) and the Mexican cavefish (Astyanax mexicanus) (101), as models of malignant melanoma, mitochondrial, and diabetes and metabolic diseases, respectively.
We propose that fishes, currently only represented by a single study in our sample (Figure 1A), are an ideal system to observationally and experimentally study helminth-microbiome interactions and their effect on host health. Fishes are the ecologically dominant vertebrates in many aquatic environments (102). Their immune systems, while simpler than those of mammals, share key components with humans (103). This similarity allows for investigating how parasitic infections and microbiome changes impact the immune system, thereby enhancing our general understanding of how such interactions influence health outcomes.
Given the need for natural candidate models and helminthological research in the Global South in general and in Africa in particular (Figure 2), and the above-mentioned potential of fishes, we propose that a well-delineated aquatic system in the tropics, such as the African Great Lakes, could provide an ideal system to investigate helminth-microbiome interactions under natural conditions. Disease burden of tropical water-borne diseases is societally relevant in the region (75) and contact with water as homogenizer exposes the human population to the same naturally occurring parasites as other potential hosts in the ecosystem. The increasing availability of high-quality reference fish genomes facilitates the application of various -omics approaches, especially for cichlid fishes (104–107), the most species-rich fish taxon in these lakes (108). Cichlids, for which also an elaborate experimental toolbox is available (109), have been proposed as models for both non-communicable (110) and infectious diseases, with the African Great Lakes cichlids among the best-studied ones in terms of metazoan parasitological baseline (111). Also, other African cichlid species, especially economically important ones such as Nile tilapia and other tilapias, are well-studied parasitologically (112, 113), though health aspects are rarely considered (114). Elsewhere in the Global South, several Neotropical cichlids are well-studied (109) also in terms of their parasites (115–120). Moreover, also the microbial symbionts of (African Great Lake) cichlids have recently seen a surge in scientific attention (121, 122). In turn, cichlids sourced from such diverse natural study systems, may be well-suited to develop as model organisms for experimentally manipulating both microbiomes and parasitic load under lab conditions, enabling researchers to uncover causal relationships and test potential therapeutic interventions relevant to human health.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
Author contributions
MV: Conceptualization, Data curation, Investigation, Writing – original draft, Writing – review & editing. SK: Conceptualization, Writing – original draft, Writing – review & editing. JF: Conceptualization, Writing – original draft, Writing – review & editing. CH: Conceptualization, Writing – original draft, Writing – review & editing. MP: Conceptualization, Writing – original draft, Writing – review & editing. NK: Conceptualization, Data curation, Investigation, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. The content of this manuscript previously appeared online in a preprint (123). This work was supported by the Special Research Fund of Hasselt University (BOF21INCENT09) and by the AfroWetMaP project of the Belgian Federal Science Policy Office (4255-FED-tWIN-G3 program, Prf-2022-049). This research was funded in part by the Austrian Science Fund (FWF) (grant DOI: 10.55776/P32691). JMOF acknowledges the institutional support of the grant ‘Severo Ochoa Centre of Excellence’ accreditation (CEX2019-000928-S) funded by AEI 10.13039/501100011033 (Spain). For open access purposes, the authors have applied a CC BY public copyright license to any author-accepted manuscript version arising from this submission.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2025.1527184/full#supplementary-material.
Footnotes
References
1. Jackson JA, Friberg IM, Little S, Bradley JE. Review series on helminths, immune modulation and the hygiene hypothesis: immunity against helminths and immunological phenomena in modern human populations: coevolutionary legacies? Immunology. (2009) 126:18–27. doi: 10.1111/j.1365-2567.2008.03010.x
2. Ferreira LF, de Araújo AJ, Confalonieri UE, Nuñez L. The finding of eggs of Diphyllobothrium in human coprolites (4,100-1,950 B.C.) from northern Chile. Mem Inst Oswaldo Cruz. (1984) 79:175–80. doi: 10.1590/s0074-02761984000200004
4. Ryan H, Flammer PG, Nicholson R, Loe L, Reeves B, Allison E, et al. Reconstructing the history of helminth prevalence in the UK. PloS Negl Trop Dis. (2022) 16:e0010312. doi: 10.1371/journal.pntd.0010312
5. De Filippo C, Cavalieri D, Di Paola M, Ramazzotti M, Poullet JB, Massart S, et al. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc Natl Acad Sci USA. (2010) 107:14691–6. doi: 10.1073/pnas.1005963107
6. Eder W, Ege MJ, von Mutius E. The asthma epidemic. N Engl J Med. (2006) 355:2226–35. doi: 10.1056/NEJMra054308
7. Conrad N, Misra S, Verbakel JY, Verbeke G, Molenberghs G, Taylor PN, et al. Incidence, prevalence, and co-occurrence of autoimmune disorders over time and by age, sex, and socioeconomic status: a population-based cohort study of 22 million individuals in the UK. Lancet. (2023) 401:1878–90. doi: 10.1016/S0140-6736(23)00457-9
8. Blackley CH. Experimental researches on the causes and nature of catarrhus aestivus (hay-fever and hay-asthma). London: Baillière, Tindall and Cox (1873).
9. Rook GA. The old friends hypothesis: evolution, immunoregulation and essential microbial inputs. Front Allergy. (2023) 4:1220481. doi: 10.3389/falgy.2023.1220481
10. Rook GA, Adams V, Hunt J, Palmer R, Martinelli R, Brunet LR. Mycobacteria and other environmental organisms as immunomodulators for immunoregulatory disorders. Springer Semin Immunopathol. (2004) 25:237–55. doi: 10.1007/s00281-003-0148-9
11. Schuijs MJ, Willart MA, Vergote K, Gras D, Deswarte K, Ege MJ, et al. Farm dust and endotoxin protect against allergy through A20 induction in lung epithelial cells. Science. (2015) 349:1106–10. doi: 10.1126/science.aac6623
12. Giacomin P, Croese J, Krause L, Loukas A, Cantacessi C. Suppression of inflammation by helminths: a role for the gut microbiota? Phil. Trans R Soc B. (2015) 370:20140296. doi: 10.1098/rstb.2014.0296
13. Versini M, Jeandel P-Y, Bashi T, Bizzaro G, Blank M, Schoenfeld Y. Unraveling the hygiene hypothesis of helminthes and autoimmunity: origins, pathophysiology, and clinical applications. BMC Med. (2015) 13:81. doi: 10.1186/s12916-015-0306-7
14. Maizels RM, Smits HH, McSorley HJ. Modulation of host immunity by helminths: the expanding repertoire of parasite effector molecules. Immunity. (2018) 49:801–18. doi: 10.1016/j.immuni.2018.10.016
15. Ramanan D, Bowcutt R, Lee SC, Tang MS, Kurtz ZD, Ding Y, et al. Helminth infection promotes colonization resistance via type 2 immunity. Science. (2016) 352:608–12. doi: 10.1126/science.aaf3229
16. Yang C-A, Liang C, Lin C-L, Hsiao C-T, Peng C-T, Lin H-C, et al. Impact of Enterobius vermicularis infection and mebendazole treatment on intestinal microbiota and host immune response. PloS Negl Trop Dis. (2017) 11:e0005963. doi: 10.1371/journal.pntd.0005963
17. Salim Masoud N, Knopp S, Lenz N, Lweno O, Abdul Kibondo U, Mohamed A, et al. The impact of soil transmitted helminth on malaria clinical presentation and treatment outcome: a case control study among children in Bagamoyo district, coastal region of Tanzania. PloS Negl Trop Dis. (2024) 18:e0012412. doi: 10.1371/journal.pntd.0012412
18. Rausch S, Midha A, Kuhring M, Affinass N, Radonic A, Kühl AA, et al. Parasitic nematodes exert antimicrobial activity and benefit from microbiota-driven support for host immune regulation. Front Immunol. (2018) 9:2282. doi: 10.3389/fimmu.2018.02282
19. Lee SC, Tang MS, Easton AV, Devlin JC, Chua LL, Cho I, et al. Linking the effects of helminth infection, diet and the gut microbiota with human whole-blood signatures. PloS Pathog. (2019) 15:e1008066. doi: 10.1371/journal.ppat.1008066
20. Colin Y, Molbert N, Berthe T, Agostini S, Alliot F, Decencière B, et al. Dysbiosis of fish gut microbiota is associated with helminths parasitism rather than exposure to PAHs at environmentally relevant concentrations. Sci Rep. (2022) 12:11084. doi: 10.1038/s41598-022-15010-2
21. Rafaluk-Mohr C, Gerth M, Sealey JE, Ekroth AKE, Aboobaker AA, Kloock A, et al. Microbial protection favors parasite tolerance and alters host-parasite coevolutionary dynamics. Curr Biol. (2022) 32:1593–8. doi: 10.1016/j.cub.2022.01.063
22. Sanya RE, Webb EL, Zziwa C, Kizindo R, Sewankambo M, Tumusiime J, et al. The effect of helminth infections and their treatment on metabolic outcomes: results of a cluster-randomized trial. Clin Infect Dis. (2020) 71:601–13. doi: 10.1093/cid/ciz859
23. Zheng D, Liwinski T, Elinav E. Interaction between microbiota and immunity in health and disease. Cell Res. (2020) 30:492–506. doi: 10.1038/s41422-020-0332-7
24. Hayes KS, Bancroft AJ, Goldrick M, Portsmouth C, Roberts IS, Grencis RK. Exploitation of the intestinal microflora by the parasitic nematode Trichuris muris. Science. (2010) 328:1391–4. doi: 10.1126/science.1187703
25. Dea-Ayuela MA, Rama-Iñiguez S, Bolás-Fernandez F. Enhanced susceptibility to Trichuris muris infection of B10Br mice treated with the probiotic Lactobacillus casei. Int Immunopharmacol. (2008) 8:28–35. doi: 10.1016/j.intimp.2007.10.003
26. Bass D, Rueckert S, Stern R, Cleary AC, Taylor JD, Ward GM, et al. Parasites, pathogens, and other symbionts. Trends Parasitol. (2021) 37:875–89. doi: 10.1016/j.pt.2021.05.006
27. Margulis L, Fester R. Symbiosis as a source of evolutionary innovation: speciation and morphogenesis. Cambridge, MA: MIT Press (1991).
28. Baedke J, Fábregas-Tejeda A, Delgado AN. The holobiont concept before Margulis. J Exp Zool B Mol Dev Evol. (2020) 334:149–55. doi: 10.1002/jez.b.22931
29. Zilber-Rosenberg I, Rosenberg E. Role of microorganisms in the evolution of animals and plants: the hologenome theory of evolution. FEMS Microbiol Rev. (2008) 32:723–35. doi: 10.1111/j.1574-6976.2008.00123.x
30. Madhusoodanan J. Do hosts and their microbes evolve as a unit? Proc Natl Acad Sci USA. (2019) 116:14391–4. doi: 10.1073/pnas.1908139116
31. González-Pech RA, Li VY, Garcia V, Boville E, Mammone M, Kitano H, et al. The evolution, assembly, and dynamics of marine holobionts. Ann Rev Mar Sci. (2024) 16:443–66. doi: 10.1146/annurev-marine-022123-104345
32. Rosenberg E, Zilber-Rosenberg I. The hologenome concept of evolution after 10 years. Microbiome. (2018) 6:78. doi: 10.1186/s40168-018-0457-9
33. Urban L, Perlas A, Francino O, Martí-Carreras J, Muga BA, Mwangi JW, et al. Real-time genomics for One Health. Mol Syst Biol. (2023) 19:e11686. doi: 10.15252/msb.202311686
34. Limborg MT, Alberdi A, Kodama M, Roggenbuck M, Kristiansen K, Gilbert MTP. Applied hologenomics: feasibility and potential in aquaculture. Trends Biotechnol. (2018) 36:252–64. doi: 10.1016/j.tibtech.2017.12.006
35. Zaiss MM, Harris NL. Interactions between intestinal microbiome and helminth parasites. Parasite Immunol. (2016) 38:5–11. doi: 10.1111/pim.12274
36. Piazzesi A, Putignani L. Impact of helminth-microbome interaction on childhood health and development - A clinical perspective. Parasite Immunol. (2022) 45:e12949. doi: 10.1111/pim.12949
37. Brosschot TP, Reynolds LA. The impact of a helminth-modified microbiome on host immunity. Mucosal Immunol. (2018) 11:1039–46. doi: 10.1038/s41385-018-0008-5
38. Loke P, Harris NL. Networking between helminths, microbes, and mammals. Cell Host Microbe. (2023) 31:464–71. doi: 10.1016/j.chom.2023.02.008
39. Rooney J, Northcote HM, Williams TL, Cortés A, Cantacessi C, Morphew RM. Parasitic helminths and the host microbiome - a missing ‘extracellular vesicle-sized’ link? Trends Parasitol. (2023) 38:737–47. doi: 10.1016/j.pt.2022.06.003
40. Yeshi K, Ruscher R, Loukas A, Wangchuk P. Immunomodulatory and biological properties of helminth-derived small molecules: potential application in diagnostics and therapeutics. Front Parasitol. (2022) 1:984152. doi: 10.3389/para.2022.984152
41. Salloum PM, Jorge F, Dheilly NM, Poulin R. Eco-evolutionary implications of helminth microbiomes. J Helminthol. (2023) 97:e22. doi: 10.1017/S0022149X23000056
42. Li Y, An J, Dang Z, Lv H, Pan W, Gao Z. Treating wheat seeds with neonicotinoid insecticides does not harm the rhizosphere microbial community. PloS One. (2018) 13:e0205200. doi: 10.1371/journal.pone.0205200
43. Peixoto RS, Harkins DM, Nelson KE. Advances in microbiome research for animal health. Annu Rev Anim Biosci. (2021) 9:289–311. doi: 10.1146/annurev-animal-091020-075907
44. Dunphy CM, Vollmer SV, Gouhier TC. Host–microbial systems as glass cannons: explaining microbiome stability in corals exposed to extrinsic perturbations. J Anim Ecol. (2021) 90:1044–57. doi: 10.1111/1365-2656.13466
45. Potts LD, Douglas A, Perez Calderon LJ, Anderson JA, Witte U, Prosser JI, et al. Chronic environmental perturbation influences microbial community assembly patterns. Environ Sci Technol. (2022) 56:2300–11. doi: 10.1021/acs.est.1c05106
46. Kreisinger J, Bastien G, Hauffe HC, Marchesi J, Perkins SE. Interactions between multiple helminths and the gut microbiota in wild rodents. Philos Trans R Soc Lond B Biol Sci. (2015) 370:20140295. doi: 10.1098/rstb.2014.0295
47. Prommi A, Prombutara P, Watthanakulpanich D, Adisakwattana P, Kusolsuk T, Yoonuan T, et al. Intestinal parasites in rural communities in Nan Province, Thailand: changes in bacterial gut microbiota associated with minute intestinal fluke infection. Parasitology. (2020) 147:972–84. doi: 10.1017/S0031182020000736
48. Martínez-Mota R, Righini N, Mallott EK, Gillespie TR, Amato KR. The relationship between pinworm (Trypanoxyuris) infection and gut bacteria in wild black howler monkeys (Alouatta pigra). Am J Primatol. (2021) 83:e23330. doi: 10.1002/ajp.23330
49. Doolin ML, Weinstein SB, Dearing MD. Pinworms are associated with taxonomic but not functional differences in the gut microbiome of white-throated woodrats (Neotoma albigula). J Parasitol. (2022) 108:408–18. doi: 10.1645/22-11
50. Oyesola OO, Souza COS, Loke PN. The influence of genetic and environmental factors and their interactions on immune response to helminth infections. Front Immunol. (2022) 13:869163. doi: 10.3389/fimmu.2022.869163
51. Rosshart SP, Vassallo BG, Angeletti D, Hutchinson DS, Morgan AP, Takeda K, et al. Wild mouse gut microbiota promotes host fitness and improves disease resistance. Cell. (2017) 171:1015–28. doi: 10.1016/j.cell.2017.09.016
52. Clavere-Graciette AG, McWhirt ME, Hoopes LA, Bassos-Hull K, Wilkinson KA, Stewart FJ, et al. Microbiome differences between wild and aquarium whitespotted eagle rays (Aetobatus narinari). Anim Microbiome. (2022) 4:34. doi: 10.1186/s42523-022-00187-8
53. Rouffaer LO, Strubbe D, Teyssier A, Hudin NS, Van den Abeele A-M, Cox I, et al. Effects of urbanization on host-pathogen interactions, using Yersinia in house sparrows as a model. PloS One. (2017) 12:e0189509. doi: 10.1371/journal.pone.0189509
54. Flandroy L, Poutahidis T, Berg G, Clarke G, Dao M-C, Decaestecker E, et al. The impact of human activities and lifestyles on the interlinked microbiota and health of humans and of ecosystems. Sci Total Environ. (2018) 627:1018–38. doi: 10.1016/j.scitotenv.2018.01.288
55. Vanhove MPM, Thys S, Decaestecker E, Antoine-Moussiaux N, De Man J, Hugé J, et al. Global change increases zoonotic risk, COVID-19 changes risk perceptions: a plea for urban nature connectedness. Cities Health. (2021) 5:S131–9. doi: 10.1080/23748834.2020.1805282
56. Von Hertzen L, Hanski I, Haahtela T. Natural immunity: biodiversity loss and inflammatory diseases are two global megatrends that might be related. EMBO Rep. (2011) 12:1089–93. doi: 10.1038/embor.2011.195
57. Glendinning L, Nausch N, Free A, Taylor DW, Mutapi F. The microbiota and helminths: sharing the same niche in the human host. Parasitology. (2014) 141:1255–71. doi: 10.1017/S0031182014000699
58. Bahrndorff S, Alemu T, Alemneh T, Lund Nielsen J. The microbiome of animals: implications for conservation biology. Int J Genomics. (2016) 2016:5304028. doi: 10.1155/2016/5304028
59. Campbell SC, Wisniewski PJ. Exercise is a novel promoter of intestinal health and microbial diversity. Exerc. Sport Sci Rev. (2017) 45:41–7. doi: 10.1249/JES.0000000000000096
60. Riggs DW, Yeager RA, Bhatnagar A. Defining the human envirome: an omics approach for assessing the environmental risk of cardiovascular disease. Circ Res. (2018) 122:1259–75. doi: 10.1161/CIRCRESAHA.117.311230
61. Graham AL. Naturalizing mouse models for immunology. Nat Immunol. (2021) 22:111–7. doi: 10.1038/s41590-020-00857-2
62. Chen YH, Yeung F, Lacey KA, Zaldana K, Lin JD, Bee GCW, et al. Rewilding of laboratory mice enhances granulopoiesis and immunity through intestinal fungal colonization. Sci Immunol. (2023) 8:eadd6910. doi: 10.1126/sciimmunol.add6910
63. Formenti F, Cortés A, Brindley PJ, Cantacessi C, Rinaldi G. A bug’s life: delving into the challenges of helminth microbiome studies. PloS Negl Trop Dis. (2020) 14:e0008446. doi: 10.1371/journal.pntd.0008446
64. Bonde CS, Mejer H, Myhill LJ, Zhu L, Jensen P, Büdeyri Gökgöz N, et al. Dietary seaweed (Saccharina latissima) supplementation in pigs induces localized immunomodulatory effects and minor gut microbiota changes during intestinal helminth infection. Sci Rep. (2023) 13:21931. doi: 10.1038/s41598-023-49082-5
65. Poulin R, Hay E, Jorge F. Taxonomic and geographic bias in the genetic study of helminth parasites. Int J Parasitol. (2019) 49:429–35. doi: 10.1016/j.ijpara.2018.12.005
66. Butala C, Brook TM, Majekodunmi AO, Welburn SC. Neurocysticercosis: current perspectives on diagnosis and management. Front Vet Sci. (2021) 8:615703. doi: 10.3389/fvets.2021.615703
67. Gonzalez-Alcaide G, Sosa N, Shevy L, Belinchon-Romero I, Ramos-Rincon JM. Global research on cysticercosis and neurocysticercosis: a bibliometric analysis. Front Vet Sci. (2023) 10:1156834. doi: 10.3389/fvets.2023.1156834
68. Carlson CJ, Dallas TA, Alexander LW, Phelan AL, Phillips AJ. What would it take to describe the global diversity of parasites? Proc R Soc B. (2020) 287:20201841. doi: 10.1098/rspb.2020.1841
69. Gazzinelli-Guimarães AC, Gazzinelli-Guimarães P, Weatherhead JE. A historical and systematic overview of Ascaris vaccine development. Parasitology. (2021) 148:1795–805. doi: 10.1017/S0031182021001347
70. Lustigman S, Prichard RK, Gazzinelli A, Grant WN, Boatin BA, McCarthy JS, et al. A research agenda for helminth diseases of humans: the problem of helminthiases. PloS Negl Trop Dis. (2012) 6:e1582. doi: 10.1371/journal.pntd.0001582
71. Torgerson PR, Devleesschauwer B, Praet N, Speybroeck N, Willingham AL, Kasuga F, et al. World Health Organization estimates of the global and regional disease burden of 11 foodborne parasitic diseases 2010: a data synthesis. PloS Med. (2015) 12:e1001920. doi: 10.1371/journal.pmed.1001920
72. Schluth CG, Standley CJ, Bansal S, Carlson CJ. Spatial parasitology and the unmapped human helminthiases. Parasitology. (2023) 150:391–9. doi: 10.1017/S0031182023000045
73. Papaiakovou M, Littlewood DTJ, Doyle SR, Gasser RB, Cantacessi C. Worms and bugs of the gut: the search for diagnostic signatures using barcoding, and metagenomics–metabolomics. Parasite Vector. (2022) 15:118. doi: 10.1186/s13071-022-05225-7
74. Miller ML, Rota C, Welsh A. Transforming gastrointestinal helminth parasite identification in vertebrate hosts with metabarcoding: a systematic review. Parasite Vector. (2024) 17:311. doi: 10.1186/s13071-024-06388-1
75. Robertson LJ, Chitanga S, Mukaratirwa S. Food and waterborne parasites in Africa –threats and opportunities. Food Waterborne Parasitol. (2020) 20:e00093. doi: 10.1016/j.fawpar.2020.e00093
76. Scholz T, Vanhove MPM, Smit N, Jayasundera Z, Gelnar M eds. A guide to the parasites of African freshwater fishes Vol. 18. Brussels: Royal Belgian Institute of Natural Sciences (2018). AbcTaxa.
77. Pullan RL, Smith JL, Jasrasaria R, Brooker SJ. Global numbers of infection and disease burden of soil transmitted helminth infections in 2010. Parasite Vector. (2014) 7:37. doi: 10.1186/1756-3305-7-37
78. Adhish M, Manjubala I. Effectiveness of zebrafish models in understanding human diseases - A review of models. Heliyon. (2023) 9:e14557. doi: 10.1016/j.heliyon.2023.e14557
79. Gora AH, Rehman S, Dias J, Fernandes JMO, Olsvik PO, Sørensen M, et al. Protective mechanisms of a microbial oil against hypercholesterolemia: evidence from a zebrafish model. Front Nutr. (2023) 10:1161119. doi: 10.3389/fnut.2023.1161119
80. Goody MF, Sullivan C, Kim CH. Studying the immune response to human viral infections using zebrafish. Dev Comp Immunol. (2014) 46:84–95. doi: 10.1016/j.dci.2014.03.025
81. Sanders JL, Moulton H, Moulton Z, McLeod R, Dubey JP, Weiss LM, et al. The zebrafish, Danio rerio, as a model for Toxoplasma gondii: an initial description of infection in fish. J Fish Dis. (2015) 38:675–9. doi: 10.1111/jfd.12393
82. Misner M, Zhang M, Sapi E. Establishing a zebrafish model for Borrelia burgdorferi infection using immersion and microinjection methods. Methods Mol Biol. (2024) 2742:131–49. doi: 10.1007/978-1-0716-3561-2_11
83. Flores EM, Nguyen AT, Odem MA, Eisenhoffer GT, Krachler AM. The zebrafish as a model for gastrointestinal tract-microbe interactions. Cell Microbiol. (2019) 22:e13152. doi: 10.1111/cmi.13152
84. Gupta S, Fernandes J, Kiron V. Antibiotic-induced perturbations are manifested in the dominant intestinal bacterial phyla of Atlantic salmon. Microorganisms. (2019) 7:233. doi: 10.3390/microorganisms7080233
85. Ruiz A, Gisbert E, Andree KB. Impact of the diet in the gut microbiota after an inter-species microbial transplantation in fish. Sci Rep. (2024) 14:4007. doi: 10.1038/s41598-024-54519-6
86. Batista S, Medina A, Pires MA, Moriñigo MA, Sansuwan K, Fernandes JMO, et al. Innate immune response, intestinal morphology and microbiota changes in Senegalese sole fed plant protein diets with probiotics or autolyzed yeast. Appl Microbiol Biotechnol. (2016) 100:7223–38. doi: 10.1007/s00253-016-7592-7
87. Dhanasiri AKS, Jaramillo-Torres A, Chikwati EM, Chikwati EM, Forberg T, Krogdahl Å, et al. Effects of dietary supplementation with prebiotics and Pediococcus acidilactici on gut health, transcriptome, microbiota, and metabolome in Atlantic salmon (Salmo salar L.) after seawater transfer. Anim. Microbiome. (2023) 5:10. doi: 10.1186/s42523-023-00228-w
88. Vargas-Albores F, Martínez-Córdova LR, Hernández-Mendoza A, Cicala F, Lago-Lestón A, Martínez-Porchas M. Therapeutic modulation of fish gut microbiota, a feasible strategy for aquaculture? Aquaculture. (2021) 544:737050. doi: 10.1016/j.aquaculture.2021.737050
89. Luna GM, Quero GM, Kokou F, Kormas K. Time to integrate biotechnological approaches into fish gut microbiome research. Curr Opin Biotechnol. (2022) 73:121–7. doi: 10.1016/j.copbio.2021.07.018
90. Zhang Z, Yang Q, Liu H, Jin J, Yang Y, Zhu X, et al. Potential functions of the gut microbiome and modulation strategies for improving aquatic animal growth. Rev Aquac. (2025) 17:e12959. doi: 10.1111/raq.12959
91. Rimoldi S, Di Rosa AR, Oteri M, Chiofalo B, Hasan I, Saroglia M, et al. The impact of diets containing Hermetia illucens meal on the growth, intestinal health, and microbiota of gilthead seabream (Sparus aurata). Fish Physiol Biochem. (2024) 50:1003–24. doi: 10.1007/s10695-024-01314-9
92. Howe K, Clark MD, Torroja CF, Torrance J, Berthelot C, Muffato M, et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature. (2013) 496:498–503. doi: 10.1038/nature12111
93. Tessadori F, Roessler HI, Savelberg SM, Chocron S, Kamel SM, Duran KJ, et al. Effective CRISPR/Cas9-based nucleotide editing in zebrafish to model human genetic cardiovascular disorders. Dis Model Mech. (2018) 11:dmm035469. doi: 10.1242/dmm.035469
94. Parey E, Louis A, Montfort J, Bouchez O, Roques C, Iampietro C, et al. Genome structures resolve the early diversification of teleost fishes. Science. (2023) 379:572–5. doi: 10.1126/science.abq4257
95. Kodera K, Matsui H. Zebrafish, medaka and turquoise killifish for understanding human neurodegenerative/neurodevelopmental disorders. Int J Mol Sci. (2022) 23:1399. doi: 10.3390/ijms23031399
96. Matsui H, Gavinio R, Takahashi R. Medaka fish Parkinson’s disease model. Exp Neurobiol. (2012) 21:94–100. doi: 10.5607/en.2012.21.3.94
97. Beck EA, Healey HM, Small CM, Currey MC, Desvignes T, Cresko WA, et al. Advancing human disease research with fish evolutionary mutant models. Trends Genet. (2022) 38:22–44. doi: 10.1016/j.tig.2021.07.002
98. Saunders AN, Gallant JR. A review of the reproductive biology of mormyroid fishes: an emerging model for biomedical research. J Exp Zool B Mol Dev Evol. (2024) 342:144–63. doi: 10.1002/jez.b.23242
99. Schartl M, Lu Y. Validity of Xiphophorus fish as models for human disease. Dis Model Mech. (2024) 17:dmm050382. doi: 10.1242/dmm.050382
100. Baris TZ, Wagner DN, Dayan DI, Du X, Blier PU, Pichaud N, et al. Evolved genetic and phenotypic differences due to mitochondrial-nuclear interactions. PloS Genet. (2017) 13:e1006517. doi: 10.1371/journal.pgen.1006517
101. McGaugh SE, Kowalko JE, Duboué E, Lewis P, Franz-Odendaal TA, Rohner N, et al. Dark world rises: the emergence of cavefish as a model for the study of evolution, development, behavior, and disease. J Exp Zool B Mol Dev Evol. (2020) 334:397–404. doi: 10.1002/jez.b.22978
102. Helfman GS, Collette BB, Facey DE, Bowen BW. The diversity of fishes: diversity, evolution and ecology. Chichester: Wiley-Blackwell (2009).
103. Rauta PR, Nayak B, Das S. Immune system and immune responses in fish and their role in comparative immunity study: a model for higher organisms. Immunol Lett. (2012) 148:23–33. doi: 10.1016/j.inlet.2012.08.003
104. Brawand D, Wagner CE, Li YI, Malinsky M, Keller I, Fan S, et al. The genomic substrate for adaptive radiation in African cichlid fish. Nature. (2014) 513:375–81. doi: 10.1038/nature13726
105. Irisarri I, Singh P, Koblmüller S, Torres-Dowdall J, Henning F, Franchini P, et al. Phylogenomics uncovers early hybridization and adaptive loci shaping the radiation of Lake Tanganyika cichlid fishes. Nat Commun. (2018) 9:3159. doi: 10.1038/s41467-018-05479-9
106. Matschiner M, Böhne A, Ronco F, Salzburger W. The genomic timeline of cichlid fish diversification across continents. Nat Commun. (2020) 11:5895. doi: 10.1038/s41467-020-17827-9
107. Ronco F, Matschiner M, Böhne A, Boila A, Büscher HH, El Taher A, et al. Drivers and dynamics of a massive adaptive radiation in cichlid fishes. Nature. (2021) 589:76–81. doi: 10.1038/s41586-020-2930-4
108. Salzburger W, Van Bocxlaer B, Cohen AS. Ecology and evolution of the African Great Lakes and their faunas. Annu Rev Ecol Evol Syst. (2014) 45:519–45. doi: 10.1146/annurev-ecolsys-120213-091804
109. Santos ME, Lopes JF, Kratochwil CF. East African cichlid fishes. EvoDevo. (2023) 14:1. doi: 10.1186/s13227-022-00205-5
110. Parsons KJ, McWhinnie K, Armstrong T. An evodevo view of post-genomic African cichlid biology: enhanced models for evolution and biomedicine. In: Abate ME, Noakes DL, editors. The behavior, ecology and evolution of cichlid fishes (Fish & Fisheries series, vol. 40. Springer, Dordrecht (2021). p. 779–802. doi: 10.1007/978-94-024-2080-7_21
111. Vanhove MPM, Kmentová N, Faes C, Fernandes JMO, Hahn C, Hens N, et al. Understanding the influence of host radiation on symbiont speciation through parasites of species flocks. Cold Spring Harb Perspect Biol. (2025) 17:a041450. doi: 10.1101/cshperspect.a041450
112. Cruz-Laufer AJ, Artois T, Pariselle A, Smeets K, Vanhove MPM. The cichlid-Cichlidogyrus network: a blueprint for a model system of parasite evolution. Hydrobiologia. (2021) 848:3847–63. doi: 10.1007/s10750-020-04426-4
113. Shinn AP, Avenant-Oldewage A, Bondad-Reantaso MG, Cruz-Laufer A, García Vásquez A, Hernández-Orts JS, et al. A global review of problematic and pathogenic parasites of farmed tilapia. Rev Aquac. (2023) 15:92–153. doi: 10.1111/raq.12742
114. Shigoley MI, Antoine-Moussiaux N, Jauniaux T, Vanhove MPM. Parasitology of one of the world’s foremost fisheries target species lacks a One Health approach. Hydrobiologia. (2024). doi: 10.1007/s10750-024-05695-z
115. Vidal-Martínez VM, Aguirre-Macedo ML, Scholz T, González-Solís D, Mendoza-Franco EF. Atlas of the helminth parasites of cichlid fish of Mexico. Prague: Academia (2001).
116. Salgado-Maldonado G. Checklist of helminth parasites of freshwater fishes from Mexico. Zootaxa. (2006) 1324:1–357. doi: 10.11646/zootaxa.1324.1.1
117. Salgado-Maldonado G. Helminth parasites of freshwater fish from Central America. Zootaxa. (2008) 1915:29–53. doi: 10.11646/zootaxa.1915.1.2
118. Luque JL, Poulin R. Metazoan parasite species richness in Neotropical fishes: hotspots and the geography of biodiversity. Parasitology. (2007) 134:865–78. doi: 10.1017/S0031182007002272
119. Roche DG, Leung B, Franco EFM, Torchin ME. Higher parasite richness, abundance and impact in native versus introduced cichlid fishes. Int J Parasitol. (2010) 40:1525–30. doi: 10.1016/j.ijpara.2010.05.007
120. Vanhove MPM, Hablützel PI, Pariselle A, Šimková A, Huyse T, Raeymaekers JAM. Cichlids: a host of opportunities for evolutionary parasitology. Trends Parasitol. (2016) 32:820–32. doi: 10.1016/j.pt.2016.07.002
121. Riera JL, Baldo L. Microbial co-occurrence networks of gut microbiota reveal community conservation and diet-associated shifts in cichlid fishes. Anim Microbiome. (2020) 2:36. doi: 10.1186/s42523-020-00054-4
122. Costa VA, Ronco F, Mifsud JC, Harvey E, Salzburger W, Holmes EC. Host adaptive radiation is associated with rapid virus diversification and cross-species transmission in African cichlid fishes. Curr Biol. (2024) 34:1247–57. doi: 10.1016/j.cub.2024.02.008
Keywords: Cichlidae, immunomodulation, Lake Tanganyika, microbial diversity, symbiome, old friends hypothesis, parasite, worm
Citation: Vanhove MPM, Koblmüller S, Fernandes JMO, Hahn C, Plusquin M and Kmentová N (2025) Cichlid fishes are promising underutilized models to investigate helminth-host-microbiome interactions. Front. Immunol. 16:1527184. doi: 10.3389/fimmu.2025.1527184
Received: 13 November 2024; Accepted: 27 January 2025;
Published: 13 February 2025.
Edited by:
Tae Sung Jung, Gyeongsang National University, Republic of KoreaReviewed by:
Priscila Salloum, University of Otago, New ZealandGuillermo Salgado-Maldonado, National Autonomous University of Mexico, Mexico
Copyright © 2025 Vanhove, Koblmüller, Fernandes, Hahn, Plusquin and Kmentová. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Maarten P. M. Vanhove, bWFhcnRlbi52YW5ob3ZlQHVoYXNzZWx0LmJl; Christoph Hahn, Y2hyaXN0b3BoLmhhaG5AdW5pLWdyYXouYXQ=