 Giuseppe G. F. Leite1*
Giuseppe G. F. Leite1* Mônica Bragança Sousa1
Mônica Bragança Sousa1 Larissa de Oliveira C. P. Rodrigues1
Larissa de Oliveira C. P. Rodrigues1 Milena Karina Colo Brunialti1
Milena Karina Colo Brunialti1 José Medina-Pestana2,3
José Medina-Pestana2,3 Joe M. Butler4
Joe M. Butler4 Hessel Peters-Sengers4,5
Hessel Peters-Sengers4,5 Lúcio Requião-Moura2,3
Lúcio Requião-Moura2,3 Reinaldo Salomão1*
Reinaldo Salomão1*- 1Division of Infectious Diseases, Escola Paulista de Medicina, Universidade Federal de São Paulo, São Paulo, Brazil
- 2Division of Nephrology, Escola Paulista de Medicina, Universidade Federal de São Paulo, São Paulo, Brazil
- 3Hospital do Rim, Fundação Oswaldo Ramos, São Paulo, Brazil
- 4Center for Experimental and Molecular Medicine (CEMM), Amsterdam UMC, location University of Amsterdam, Amsterdam, Netherlands
- 5Department of Epidemiology and Data Science, Amsterdam UMC, location Vrije Universiteit, Amsterdam, Netherlands
The COVID-19 pandemic has significantly impacted global health, especially in vulnerable populations like kidney transplant recipients (KTRs). Recently, mass spectrometry-based proteomics has emerged as a powerful tool to shed light on a broad spectrum of dysregulated biological processes in KTRs with COVID-19. In this study, we prospectively collected blood samples from 17 COVID-19-positive KTRs and 10 non-infected KTRs between May and September 2020. Using tandem mass tag-based quantitative proteomics, we analyzed peripheral blood mononuclear cells (PBMCs), plasma protein biomarkers, and lymphocyte counts, followed by bioinformatics analysis. Our results revealed significant proteomic alterations in COVID-19-infected KTRs, particularly in pathways related to glycolysis, glucose metabolism, and neutrophil degranulation. Additionally, we observed an altered immune response characterized by elevated cytokines and decreased lymphocyte counts. Notably, KTRs with AKI exhibited worse clinical outcomes, including higher rates of ICU admission and mechanical ventilation. Comparative analysis of PBMC proteomic profiles between AKI and non-AKI patients identified distinct immune-related pathways, with AKI patients showing marked changes in innate immune responses, particularly neutrophil degranulation. Furthermore, we observed a negative correlation between T cell counts and neutrophil degranulation, suggesting a role for immune dysregulation in COVID-19. Our findings provide critical insights into the immune and metabolic responses in COVID-19-infected KTRs, especially those with AKI, highlighting the need for focused research and therapeutic strategies targeting immune dysregulation in this high-risk population.
Introduction
The Coronavirus disease 2019 (COVID-19) pandemic, which emerged in 2020, rapidly became a global health crisis, profoundly affecting healthcare systems, economies, and overall quality of life (1, 2). Prior to the achievement of widespread vaccination coverage, COVID-19 presented with a wide range of clinical manifestations, from asymptomatic or presymptomatic states to varying degrees of severity, including mild, moderate, severe, and, in some cases, life-threatening complications (3).
Individuals with compromised immune systems, especially those undergoing immunosuppressive therapy, were particularly vulnerable. Among these high-risk groups, kidney transplant recipients (KTRs) demonstrated notably higher case-fatality rates, even after vaccination (4, 5), and exhibited lower seroconversion rates following vaccination (6). This elevated risk underscores the compounded vulnerabilities associated with immunosuppressive treatments and the presence of multiple comorbidities in these patients (7–9).
The cumulative number of comorbidities associated with an immune system impairment has led to diverse responses to COVID-19 among KTRs compared with non-transplanted patients (7–9). Investigating a panel of circulating cytokines and vascular mediators in sequential samples, we previously demonstrated that both non-KTRs and KTRs infected with COVID-19 exhibited a similar transition from admission to convalescent samples. However, the key mediators differentiating patients from their respective controls (i.e., healthy volunteers and non-infected KTRs) were distinct. For non-KTRs, the top differentiators were inflammatory cytokines, whereas for KTRs, endothelial response patterns and levels of Neutrophil Gelatinase-Associated Lipocalin (NGAL) (10), an early marker of acute kidney injury (AKI), were the most significant. AKI, commonly observed in numerous systemic conditions (11), has been linked to poor outcomes, a trend that persisted in COVID-19 patients. A multicenter study involving 1,680 KTRs with COVID-19 revealed that the mortality rate was 36.0% in those with AKI, compared to 19.1% in patients maintaining normal renal function, highlighting renal function as a key predictor of survival in this population (12).
Recently, mass spectrometry (MS)-based proteomics, primarily focused on plasma, serum, and urine, has emerged as a powerful tool to shed light on a broad spectrum of dysregulated biological processes in KTRs with COVID-19 (13, 14). However, the influence of SARS-CoV-2 infection on peripheral blood mononuclear cells (PBMCs) in KTRs remains a subject of ongoing investigation.
In this study, we employed a tandem mass tag (TMT)-based quantitative proteomic approach, coupled with bioinformatics analyses, to investigate altered proteins associated with dysregulated pathways and biological processes in kidney transplant recipients (KTRs) with COVID-19. We focused on patients with moderate to severe illness admitted to hospital wards who presented with or without AKI, as well as patients discharged from the hospital. Our aim was to provide insights into proteomic changes during the early stages of the disease and their association with AKI and post-clinical recovery.
Materials and methods
Study design and population
This prospective cohort study was conducted at Hospital Sao Paulo, the University Hospital of the Federal University of Sao Paulo, located in Sao Paulo, Brazil (15). The study was submitted to and approved by the National Research Ethics Committee (Comitê Nacional de Ética em Pesquisa – CONEP), Process number 3.978.709. All volunteers provided written informed consent prior to enrolment. The study included adult patients (≥18 years old) diagnosed with COVID-19 based on positive results from quantitative reverse transcription polymerase chain reaction (RT-qPCR) testing of nasopharyngeal swabs. These patients were admitted to the hospital wards between May 10 and September 26, 2020. This period coincided with the first wave of the COVID-19 pandemic in Brazil (February 25, 2020, to November 5, 2020) (16), when the predominant variants observed were B.1.1.28 (20–30%) and B.1.1.33 (10–35%) (16). Patients included in the cohort had moderate to severe illness, as defined by guidelines from the National Institutes of Health and the World Health Organization (17, 18). Exclusion criteria encompassed patients referred to outpatient clinics or admitted directly to the ICU. Among the initial 68 COVID-19 patients in the cohort, 17 KTRs were selected for this study [for a comprehensive overview of the entire cohort, please refer to Peçanha-Pietrobom et al. (15)]. Additionally, a control group comprised 10 KTR volunteers without COVID-19 symptoms (KTR controls), matched to the patient group for time since transplantation, immunosuppressive therapy, sex, and age. None of the participants in this study were vaccinated due to the unavailability of vaccines at the time of enrolment.
AKI was prospectively defined according to the Kidney Disease Improving Global Outcomes (KDIGO) criteria (19, 20). Baseline graft function was assessed using creatinine levels recorded three months prior to COVID-19 diagnosis, and AKI was identified based on creatinine levels at hospital admission.
Sample collection
Blood samples were collected from both patients and KTR controls into ethylenediaminetetraacetic acid (EDTA)-treated tubes (BD Biosciences, San Diego, CA, USA). Plasma and peripheral blood mononuclear cells (PBMCs) were separated using a Ficoll gradient method (Ficoll-Paque PLUS, GE Healthcare Biosciences, Uppsala, Sweden). Plasma samples were stored at –80°C, and PBMCs were preserved in liquid nitrogen for subsequent use.
Measurements
For detailed information on the proteomics experiments, data processing protocols, plasma biomarker assays, and the corresponding analysis methods, please refer to the supplementary methods in Supplementary Material 1.
Statistical analysis
Statistical analyses were performed using R (version 4.3.0). Normality of data was assessed using the Shapiro–Wilk test and Quantile–Quantile (Q-Q) plots. Non-normally distributed data were analyzed using the Mann–Whitney U test or the Kruskal–Wallis test, while normally distributed data were analyzed using Welch’s t-test or analysis of variance (ANOVA). For post hoc testing of non-normally distributed data, Dunn’s test of multiple comparisons using rank sums was employed; for normally distributed continuous data, Tukey’s post hoc test was conducted. Categorical variables were compared using the chi-square test or Fisher’s exact test, as appropriate. Biomarker data were log-transformed to normalize their distribution. To visualize the overall differences among plasma biomarkers, principal component analysis (PCA) was conducted following previously established method (21, 22). Correlation analyses were performed using Spearman’s rank correlation coefficient (Rho). Differences between groups were quantified using standardized mean differences (SMD) (23).
Bioinformatics
Differential protein abundance analysis was performed using the R/Bioconductor package limma. The model was adjusted for additional covariates: age, sex, time after transplantation (months), maintenance immunosuppressive therapy (class), and donor status (deceased or living). Differences in PBMC proteome profiling between the two groups were detected using the empirical Bayes moderated t-statistics, and Benjamini-Hochberg corrections were applied to all p-values to calculate the false discovery rates (FDR). Protein set enrichment analysis (PSEA) was performed using the fgsea package, with annotation information from the Reactome pathway database and hallmark gene sets from MSigDB. Additionally, single-sample PSEA (ssPSEA) was performed using the hacksig package and the Reactome pathway database. We focused on Reactome-defined immune-related pathways implicated in the host response to infection: innate immune system, adaptive immune system, cytokine signaling in the immune system, hemostasis, programmed cell death, and metabolism.
Results
Clinical and epidemiological features of the study participants
Seventeen kidney transplant recipients (KTRs) with COVID-19, a subset of a hospital-admitted cohort with diverse clinical outcomes (15) were analyzed in this study (Table 1). According to the KDIGO criteria for staging AKI, we observed that six patients (35%) did not present with AKI (stage 0), while five (29%) were classified as stage 1, three (17%) as stage 2, and three (17%) as stage 3. The mean age was 54.0 years, and most were male (n = 10, 59%). The time between COVID-19 symptom onset and hospital admission was 7.5 ± 4.6 days. Primary symptoms included dyspnea (n = 12, 71%), cough (n = 12, 71%), fever (n = 7, 41%), diarrhea (n = 7, 41%), and nausea/vomiting (n = 7, 41%). Arterial hypertension was the most prevalent comorbidity (n = 13, 76%). Additionally, 53% of patients required ICU admission, 41% underwent mechanical ventilation, 29% required renal replacement therapy, and 41% died. Among the survivors, six participated in a clinical follow-up at an average of 29.3 ± 5.0 days after hospital discharge (designated as Convalescent Sample 30 [CS30]). No significant differences were observed in demographic data, maintenance immunosuppressive therapy class, or donor status when comparing KTRs with COVID-19 and KTR controls (Table 1).
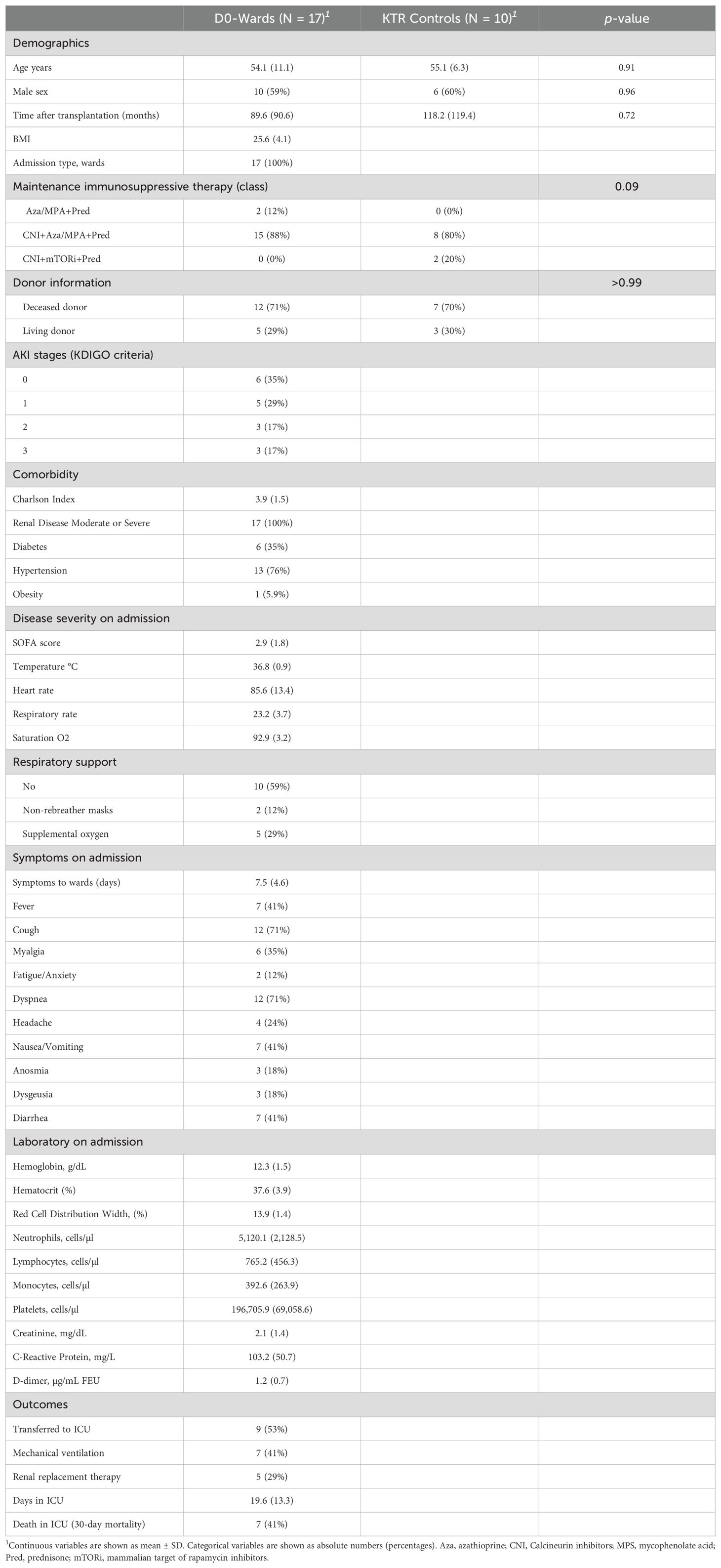
Table 1. Baseline characteristics and clinical outcomes of KTR COVID-19 patients at ward admission and KTR Controls.
Global COVID-19 perturbation of the PBMC proteome, plasma proteins, and lymphocyte counts in kidney transplant recipients
A TMT-based quantitative proteomic approach combined with liquid chromatography-tandem mass spectrometry (LC-MS/MS) was employed to quantify differentially abundant proteins (DAPs) among KTRs. The cohort was categorized based on the timing of blood sampling: patients sampled during their hospital stay (D0-wards) and those sampled approximately 30 days after discharge (CS30). These groups were compared with KTRs non-COVID-19 (KTR controls). Across seven TMT batches, a total of 2,330 proteins were identified. Focusing on proteins quantified in at least 50% of samples reduced this number to 1,403 proteins, with an average of 1,242 proteins per sample (Supplementary Material 1: Supplementary Figure S1). Among these 1,403 proteins, 363 were differentially abundant between D0-wards and KTR controls (Supplementary Material 1: Supplementary Figure S2A and Supplementary Material 2), while 111 showed differential abundance between CS30 and KTR controls (Supplementary Material 1: Supplementary Figure S2B and Supplementary Material 3).
Reactome pathway enrichment analysis revealed that D0-wards patients exhibited alterations in 15 pathways compared with KTR controls, characterized by positive normalized enrichment scores (NES) in pathways associated with interferon response, innate immune system, and neutrophil degranulation. Conversely, pathways related to glycolysis and glucose metabolism exhibited negative NES. Similar NES patterns were observed in hallmark gene set enrichment analysis, with positive NES in pathways related to interferon response and inflammatory processes, and negative NES in metabolism-related pathways (Figure 1A). Notably, only metabolism-related pathways remained altered in CS30 patients (Figure 1A).
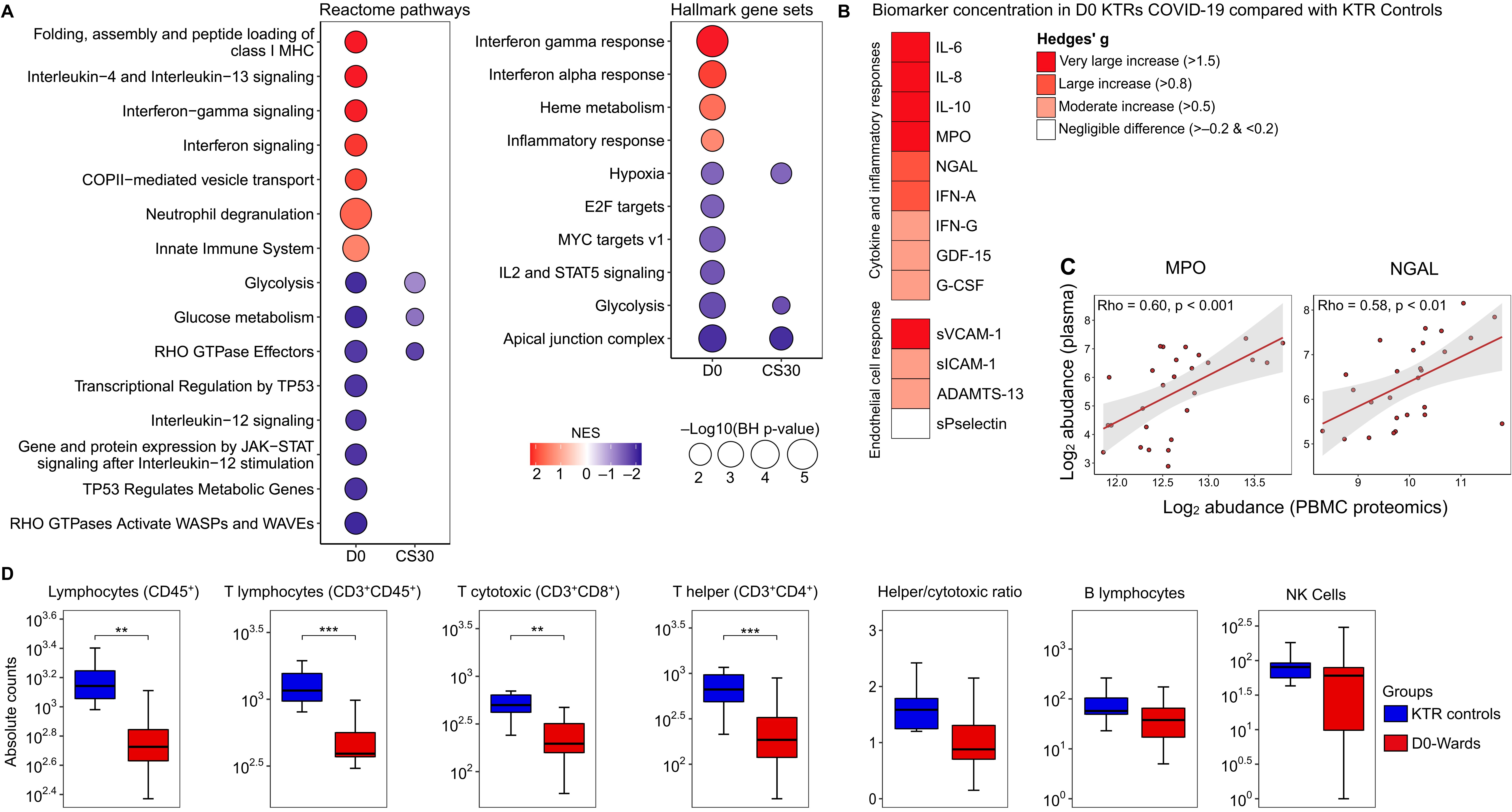
Figure 1. Comprehensive assessment of COVID-19 impact on PBMC proteome, plasma proteins, and lymphocyte counts. (A) Protein set enrichment analysis (PSEA) demonstrating protein perturbations in D0-wards patients (N = 17) versus KTR controls (N = 10), and CS30 patients (N = 6) versus KTR controls. Circle size represents the -log10(BH-adjusted p-value), while color denotes the normalized enrichment score (NES). Red indicates increased NES, and blue indicates decreased NES. (B) Heatmap showing the magnitude of biomarker differences between D0-wards patients and KTR controls, quantified using Hedges’ (g) The red gradient highlights elevated biomarker levels in D0-wards patients. (For pairwise comparisons, see Supplementary Figure S3). (C) Scatter plot illustrating Spearman’s rank correlation coefficients (Rho) between myeloperoxidase (MPO) and neutrophil gelatinase-associated lipocalin (NGAL) abundances in plasma and PBMCs (proteomic data). P-values are BH-adjusted, and (D) Box plot comparing absolute lymphocyte counts between D0-wards COVID-19 patients and KTR controls. Statistical analyses were performed using Welch’s t-test, with significance levels indicated by **p ≤ 0.01 and ***p ≤ 0.001.
Plasma biomarker levels indicative of cytokine/inflammation and endothelial cell activation were significantly elevated in D0-wards patients (Figure 1B, Supplementary Material 1: Supplementary Figure S3). Additionally, two proteins, NGAL and myeloperoxidase (MPO), were identified in both plasma and PBMCs (proteomics), demonstrating a positive moderate correlation (Figure 1C). Furthermore, the absolute number of lymphocytes, specifically T lymphocytes (CD3+CD45+), T cytotoxic (CD3+CD8+), and T helper (CD3+CD4+) were lower in D0-wards compared to KTR controls (Figure 1D). In contrast to admission samples, all plasma biomarkers (Supplementary Material 1: Supplementary Table S1) and absolute lymphocyte counts were comparable between CS30 and KTR controls (Supplementary Material 1: Supplementary Table S2), suggesting a potential return to a homeostatic state approximately 30 days after hospital discharge.
Host response associated with acute kidney injury in KTR COVID-19 patients
We compared KTR COVID-19 patients who presented with AKI (n = 6) to those who did not (n = 11) during ward admission (Table 2). Patients with AKI presented with higher Sequential Organ Failure Assessment (SOFA) scores (SMD: 0.90), increased C-Reactive Protein (CRP) levels (SMD: 0.60), and elevated creatinine levels (SMD: 1.40) upon ward admission. The AKI group experienced worse outcomes, including a higher rate of ICU transfers (SMD: 0.64), increased need for mechanical ventilation (SMD: 0.86), and greater hospital mortality (SMD: 0.86). Interestingly, creatinine showed a strong positive correlation with CRP (Rho = 0.50, p-values < 0.05) and SOFA respiratory subscores (Rho = 0.68, p-values = 0.002) (Supplementary Material 1: Supplementary Figure S4).
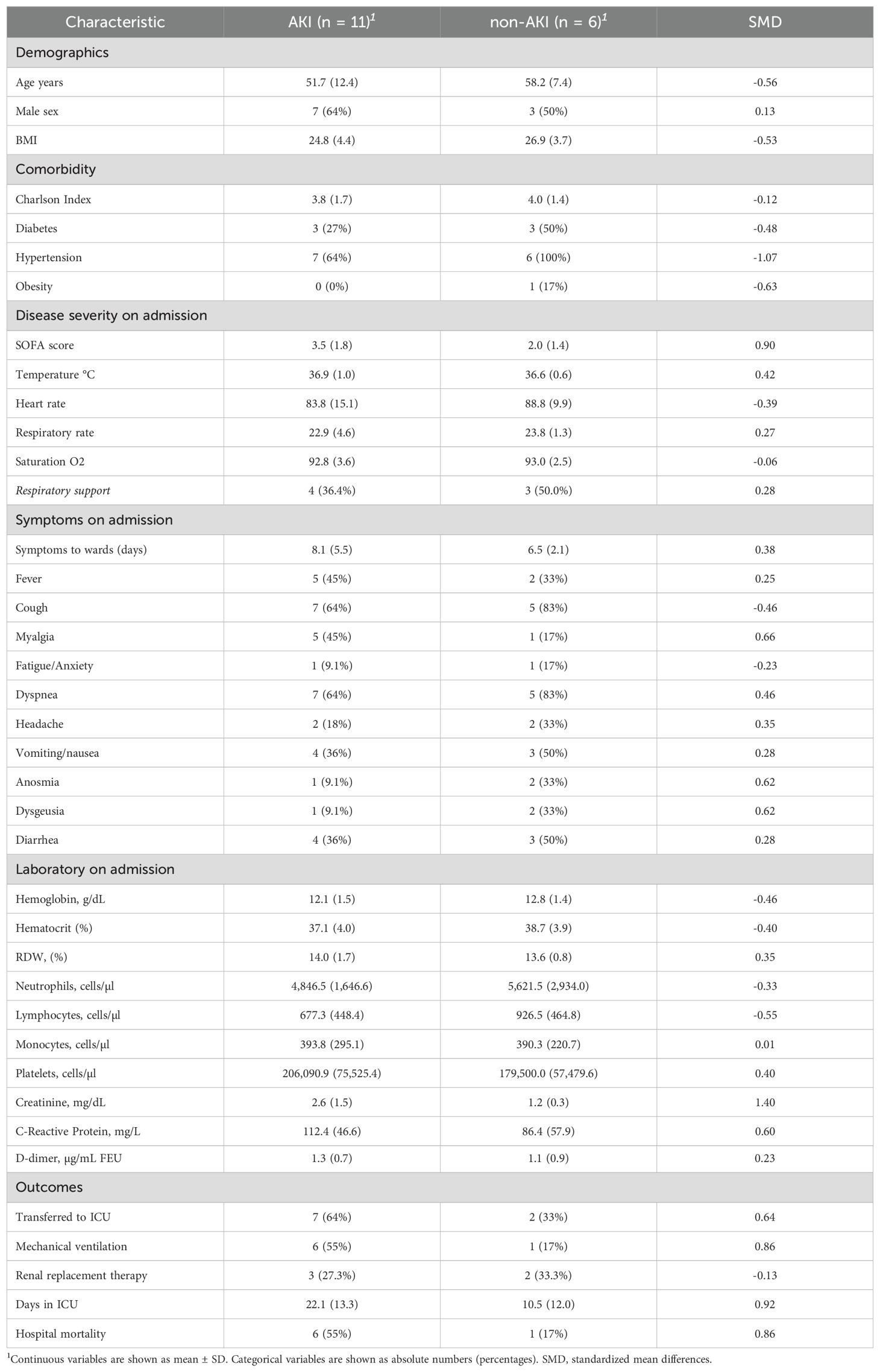
Table 2. Demography, comorbidities, clinical and laboratory admission data, and outcomes stratified according to the presence of acute kidney injury.
We compared the PBMC proteomic alterations in AKI and non-AKI COVID-19 patients using ssPSEA for Reactome-defined immune-related pathways. Our analysis revealed alterations in seven innate immune-related pathways, three adaptive immune system pathways, and five cytokine signaling pathways in the immune system. Additionally, no alterations were observed in hemostasis, while four metabolic pathways and one programmed cell death pathway showed alterations (Figure 2A). Indeed, the innate immune-related pathways exhibited the most differences when comparing patients with AKI and non-AKI; these pathways also demonstrated a pattern of positive correlation with creatinine, SOFA, and CRP levels (Figure 2B).
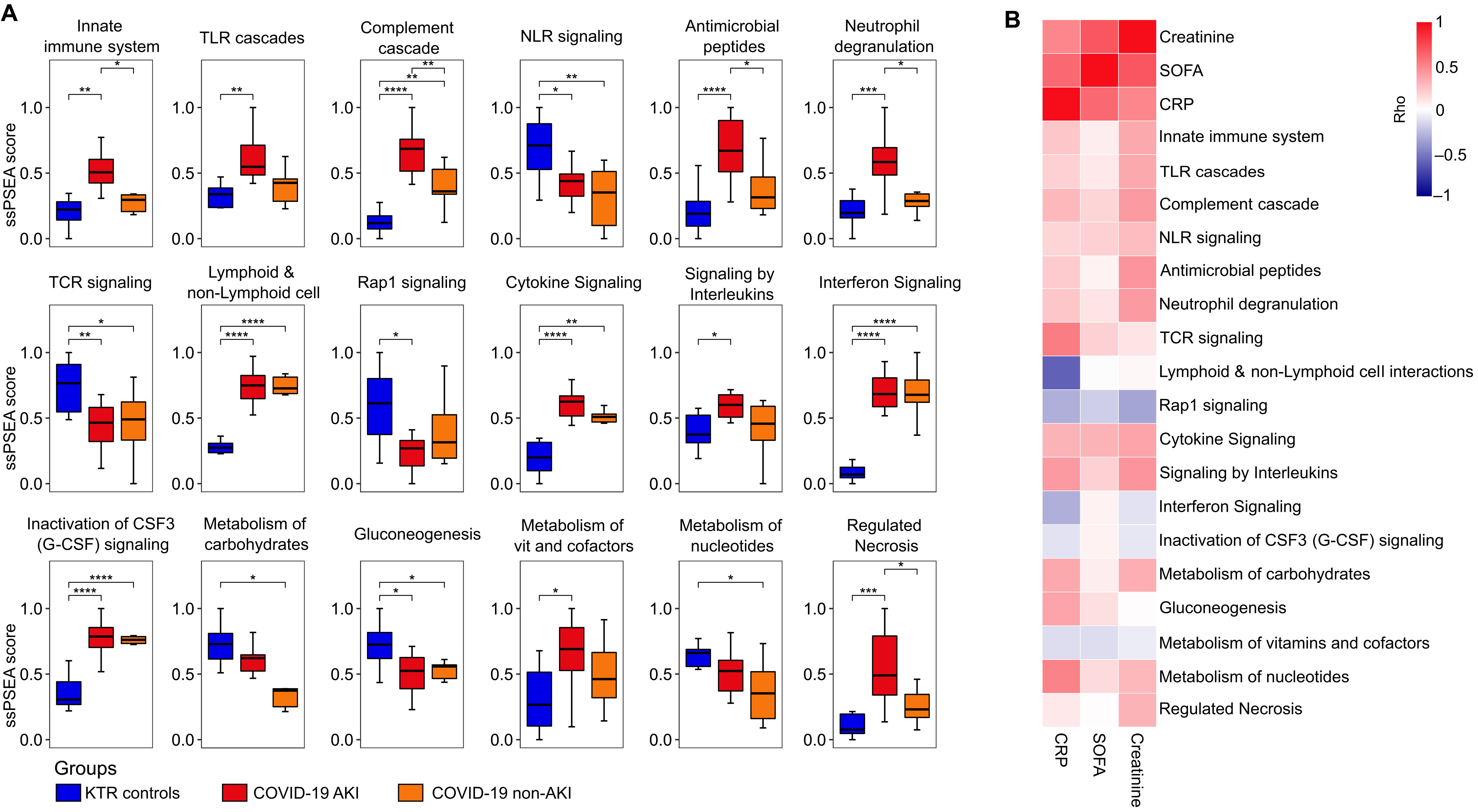
Figure 2. Alterations in immune-related pathways in COVID-19 patients stratified according to the presence of acute kidney injury. (A) Single-sample protein set enrichment analysis (ssPSEA) was performed on D0-wards COVID-19 patients stratified by the presence of acute kidney injury (AKI, n = 11) and non-AKI (n = 6), as well as KTR controls (N = 10). The figure presents only immune-related pathways that showed significant differences between the groups. Statistical analyses were conducted using ANOVA with Tukey’s post-hoc test, corrected by the Benjamini-Hochberg method. Significance is indicated as follows: *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001, and ****p ≤ 0.0001, and (B) Heatmap showing Spearman’s rank correlation coefficients (Rho) between the sequential organ failure assessment (SOFA) score, C-reactive protein (CRP) levels, creatinine levels, and ssPSEA pathway scores in D0-wards COVID-19 patients (N = 17). TLR cascades, Toll-like receptor cascades; NLR signaling, Nucleotide-binding domain, leucine rich repeat containing receptor signaling pathway; TCR signaling, T cell receptor signaling; Immunoregulatory between Lymphoid & non-Lymphoid cell, Immunoregulatory interactions between a Lymphoid and a non-Lymphoid cell; Rap1 signaling, Ras-proximate-1 signaling and Metabolism of vit and cofactors; Metabolism of vitamins and cofactors.
Next, we explored the differences between KTR COVID-19 patients with AKI and non-AKI based on 15 host response biomarkers reflective of two key pathophysiological domains (cytokine/inflammatory response and endothelial cell activation/procoagulant responses) in plasma obtained on admission to the wards. First, we generated domain-specific PCA plots to compare the groups (Figure 3). There were significant differences in plasma biomarkers grouped in the cytokine/inflammatory response domain between patients with AKI and non-AKI. We also identified a substantial overlap in the endothelial cell activation/procoagulant responses between the AKI and non-AKI groups (Figure 3A). The complete contribution of each biomarker to a principal component (PC) score is depicted in Supplementary Material 1: Supplementary Table S3.
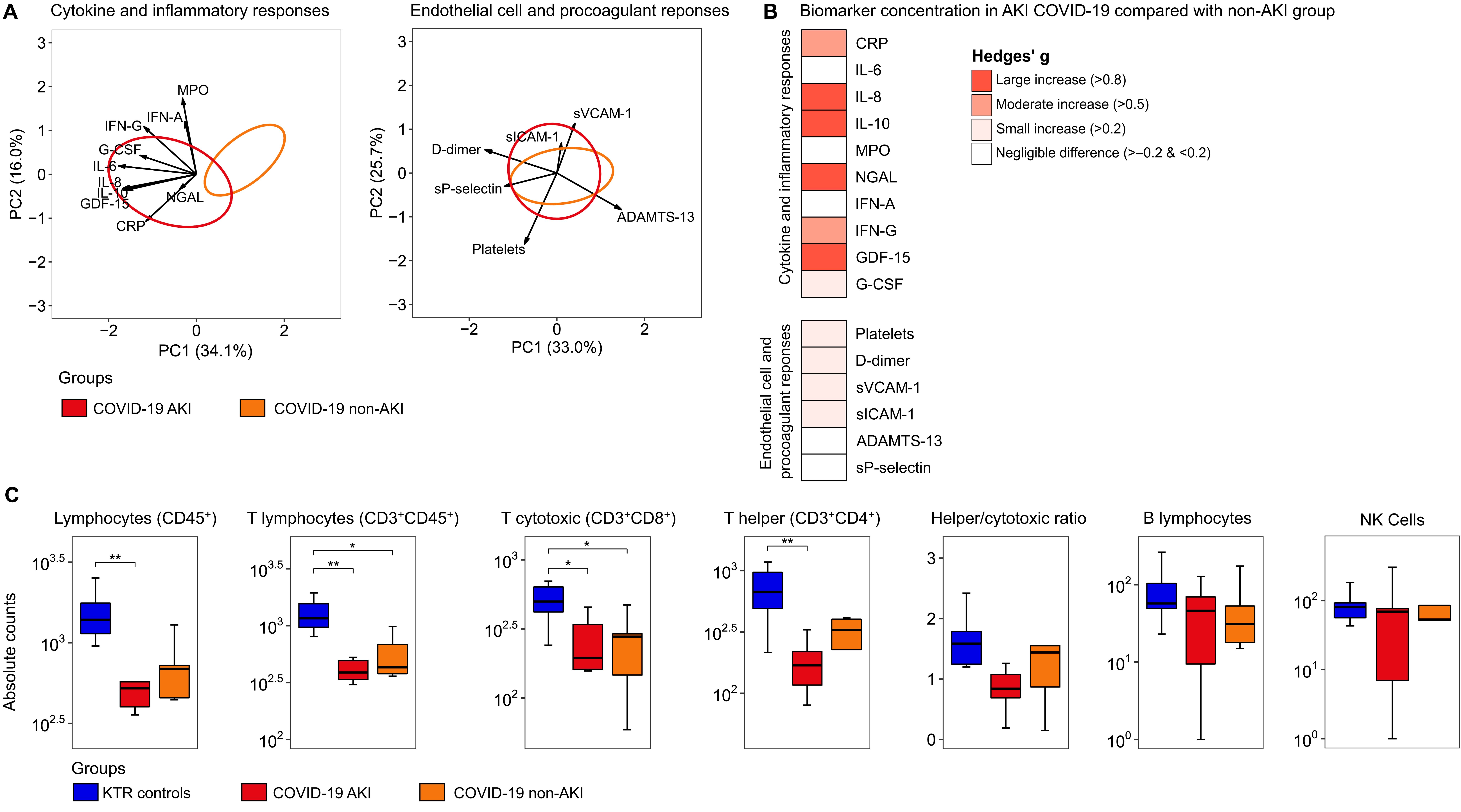
Figure 3. Host response biomarkers and lymphocyte count alterations in COVID-19 patients stratified according to the presence of acute kidney injury. (A) Principal component analysis of cytokine/inflammatory responses and endothelial cell activation/procoagulant responses. The X-axis label shows the percentage of explained variance on principal component 1, while the Y-axis label shows the percentage of explained variance on principal component 2. The ellipse indicates the central 25% of the groups. The arrows indicate the direction (arrow orientation) and strength (arrow length) of the correlation between each biomarker and the PCs. The complete contribution of each biomarker to a PC score is depicted in Supplementary Table S3. (B) Heatmap representing the Hedges’ g effect size between the AKI COVID-19 group compared with the non-AKI group (for pairwise comparison see Supplementary Table S4), and (C) Box plot illustrating differences in lymphocyte absolute counts between D0-wards COVID-19 patients stratified according to the presence of acute kidney injury (AKI, n = 11, and non-AKI, n = 6) and KTR controls (N = 10). Statistical analyses were performed using the ANOVA test with Tukey post-hoc test corrected by the Benjamini-Hochberg method. * Tukey post-hoc test p ≤ 0.05 and ** p ≤ 0.01. IL, interleukin; GDF, growth/differentiation factor; G-CSF, granulocyte-colony stimulating factor; IFN, interferon; CRP, C-Reactive Protein; NGAL, neutrophil gelatinase-associated lipocalin; MPO, Myeloperoxidase; ADAMTS-13, ADAM metallopeptidase with thrombospondin type 1 motif 13; sP-selectin, Soluble platelet selectin; sVCAM-1, Serum vascular cell adhesion molecule-1 and sICAM-1, Soluble Intercellular Adhesion Molecule-1.
When comparing the magnitude of individual biomarker differences expressed as Hedges’ g, we observed moderate to large increases in cytokine/inflammatory markers among AKI patients, while endothelial cell activation/procoagulant markers showed small or negligible differences (Figure 3B). Subsequently, we evaluated the absolute number of total lymphocytes, which showed statistical differences when comparing the KTR control and COVID-19 AKI groups (Figure 3C). We also observed differences between the control and COVID-19 AKI groups for T helper cells. For T lymphocytes and T cytotoxic cells, differences were found in both comparisons between the KTR control vs. COVID-19 AKI groups, as well as between the KTR control vs. non-AKI groups. No differences were found in the comparison between the groups for the helper/cytotoxic ratio, B lymphocytes, and NK cells. Interestingly, for total lymphocytes, T lymphocytes, T cytotoxic cells, and T helper cells, the AKI group consistently showed lower absolute values than the non-AKI group, but with no statistical difference in the pairwise comparison.
Previous studies suggest an increase in immature cells, such as low-density neutrophils (LDNs) or granulocytic/polymorphonuclear myeloid-derived suppressor cells (PMN-MDSCs), in COVID-19 patients, which correlate with T cell suppression (24, 25). LDNs commonly co-purify with PBMCs in various infections, enriching pathways like “neutrophil degranulation” in omics studies (25–30). Therefore, we tested the correlation between the absolute values of T lymphocytes and the “neutrophil degranulation” ssPSEA scores, which showed a moderate negative correlation between T lymphocytes (Rho = –0.43, p-values = 0.02), T helper cells (Rho = –0.45, p = 0.03), and the “neutrophil degranulation” score (Supplementary Material 1: Supplementary Figure S5). A weak negative and not statistically significant correlation was found between T cytotoxic cells and the “neutrophil degranulation” score (Rho = –0.35, p-values = 0.08).
Discussion
In this study, we investigated KTRs infected with SARS-CoV-2 during the initial wave of the COVID-19 pandemic in São Paulo, Brazil. Based on the reported increased severity of COVID-19 in KTRs (4, 5, 7–9), we aimed to investigate the changes in proteomics within PBMCs in this specific population. Additionally, we explored the reported immune dysregulation and endothelial cell activation/procoagulant alterations in COVID-19 (10, 31, 32), particularly emphasizing the early stages of hospitalization while patients are in the ward. Furthermore, we investigated the reported reductions in absolute counts of lymphocytes, especially T lymphocytes and their subsets, during the acute phase of COVID-19 (33, 34), with a specific focus on KTRs.
The proteomic response during the acute phase of COVID-19 has been widely explored (28–30, 35), while the proteomic alterations within PBMCs of KTRs were negligible. Our study revealed proteomic alterations linked to inflammatory/immune response pathways, indicated by positive NES. Interestingly, the majority of these pathways reverted to a status similar to that of non-COVID-19 KTRs after 30 days. This pattern aligns with findings from our previous cohort of non-KTR COVID-19 patients (28) and is consistent with other omics studies (25, 29, 30), particularly the immune-related pathways such as neutrophil degranulation, a finding that has been well documented in studies of PBMCs during infection (36).
Notably, the glycolysis and glucose metabolism pathways exhibited a negative NES and remained altered after 30 days. This metabolic alteration may affect immune cell function in pathogen recognition, as glycolysis plays a pivotal role in this process (37). This dysfunctional glycolysis profile shares similarities with findings from proteomic studies of PBMCs from critically ill COVID-19 patients (28), transcriptomic/metabolic profiling of CD14+ monocytes from patients with moderate COVID-19 (38), and single-cell RNA sequencing in monocytes and macrophages from severe patients with COVID-19-associated pulmonary aspergillosis (39). The impaired glycolysis observed in this context may serve as an indicator of the dysfunctional host response, potentially contributing to heightened morbidity and mortality rates among KTRs.
Next, we investigated the proteomic alterations based on the presence of AKI, a prevalent condition in systemic illnesses affecting a significant portion of hospitalized COVID-19 patients (40). Analysis of immune-related pathways revealed increased NES in the innate immune system, complement cascade, regulated necrosis, antimicrobial peptides, and neutrophil degranulation in COVID-19 AKI patients. These results are in line with studies discussing the pathophysiology of COVID-19–associated AKI, which is believed to be multifaceted (40), involving factors such as local and systemic inflammatory/immune response, and activation of coagulation cascades (41, 42). This is further evidenced when examining host response biomarkers, where the PCA corresponding to this cytokine/inflammatory response domain clearly separates patients with AKI from those without, a distinction not observed in the endothelial domain. Plasma biomarker levels, such as NGAL, were significantly elevated in AKI patients. The identification of this protein in both plasma and PBMCs, with a moderate positive correlation, underscores its central role in immune activation and systemic inflammation during COVID-19. Plasma proteomic profiling has further demonstrated that AKI in COVID-19 involves tubular injury, hemodynamic perturbation, neutrophil activation, and inflammatory responses, as evidenced by elevated NGAL and other markers (43).
The observed inflammatory responses were accompanied by immune cell dysregulation, particularly affecting lymphocyte populations. Although lymphocyte counts did not show statistically significant differences between the AKI and non-AKI groups, it is likely that these disturbances were more severe in the AKI group, as illustrated by the finding that absolute counts of total lymphocytes and T helper cells were lower in the AKI group but not in the non-Aki group compared to the controls. T lymphocytes, which play a critical role in viral clearance (44, 45), are known to decrease during acute COVID-19 (33, 34, 46). Interestingly, the increase in LDNs/PMN-MDSCs may directly correlate with the decrease in T cell numbers (24, 25).
Despite their immunosuppressive properties, LDNs/PMN-MDSCs have also been associated with a pro-inflammatory phenotype, characterized by elevated levels of cytokine secretion and formation of neutrophil extracellular traps (27, 47, 48). The presence of LDNs has been observed in previous studies (25, 26, 28), and may be linked to the severity of infectious diseases (25, 29, 36). Overall, our results are consistent with transcriptomic evidence of enhanced neutrophil activation and cytokine signaling, alongside downregulated adaptive and T cell responses in KTRs with increasing COVID-19 severity (49).
In fact, the AKI group is more severe than the non-AKI, showing higher SOFA scores, CRP levels, and worse outcomes. This may be associated with high scores for LDNs, as well as inflammatory/immune responses. Our findings suggest a relationship between T-cell suppression and elevated levels of LDN-related proteins in PBMCs during COVID-19, particularly in patients with AKI. This association may carry important therapeutic implications for managing immune dysregulation in KTRs, highlighting the need for targeted interventions aimed at mitigating these immune alterations.
This study has some limitations. First, the sample size was relatively small, and all patients were enrolled at a single center. Additionally, the absence of a validation cohort from another hospital in a different geographical region limits the generalizability of our findings. Moreover, we did not investigate the influence of viral load or the roles of virus variants on the results obtained. Our study was conducted in 2020, so the relevance to current variants is less clear, since SARS-CoV2 variants exhibit increasing fitness, and declining pathogenicity and induced-inflammatory response (50). Furthermore, functional assays to validate glycolysis metabolism changes and/or the potential contribution of LDNs in AKI were not conducted, which could have provided further insight into our results. Despite these limitations, our study also has notable strengths. We investigated PBMCs, a cell type that is not often explored in COVID-19 proteomics research. Our study of KTRs infected with SARS-CoV-2, along with a well-established cohort of controls, provides valuable insights and is relatively unexplored. Additionally, all individuals were unvaccinated, allowing for a clearer assessment of proteomic alterations in the absence of vaccination. Furthermore, the samples obtained in the wards and after discharge provided a comprehensive understanding of cellular proteomic changes throughout the initial symptoms of the disease and convalescence.
Future studies with larger, more diverse cohorts are needed to validate our findings and evaluate their long-term clinical implications. As a next step, we aim to explore patients with community-acquired infections, including both KTRs and non KTRs, to investigate broader mechanisms of immune dysregulation. These insights lay the groundwork for the development of targeted therapeutic interventions to address immune dysregulation in vulnerable populations, particularly kidney transplant recipients.
In summary (Figure 4), our study offers preliminary insights into the proteomic alterations within PBMCs of KTRs infected with SARS-CoV-2 during the early phase of the COVID-19 pandemic. We identified significant dysregulation in key biological pathways, particularly those involved in glycolysis, glucose metabolism, and neutrophil degranulation, which may reflect the heightened inflammatory state associated with COVID-19 in this vulnerable population. While our findings suggest an altered immune response—characterized by elevated cytokines, inflammatory mediators, and decreased lymphocyte counts—these results must be interpreted cautiously due to the small sample size. The observed association between T-cell suppression and increased levels of LDN-related proteins in AKI patients warrants further investigation, as it may hold therapeutic implications for mitigating immune dysregulation in COVID-19-infected KTRs.
Figure 4. Summary of proteomic and immune profiling in kidney transplant recipients (KTRs) with COVID-19, highlighting immune dysregulation, metabolic alterations, and the impact of acute kidney injury (AKI).
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: PXD051702 (ProteomeXchange).
Ethics statement
The study was submitted to and approved by the National Research Ethics Committee (Comitê Nacional de Ética em Pesquisa – CONEP), Process number 3.978.709. The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
Author contributions
GL: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Software, Visualization, Writing – original draft, Writing – review & editing. MS: Methodology, Writing – review & editing, Investigation. LR: Investigation, Methodology, Writing – review & editing. MB: Investigation, Methodology, Writing – review & editing. JM-P: Investigation, Methodology, Writing – review & editing. JB: Investigation, Methodology, Writing – review & editing. HP-S: Investigation, Methodology, Writing – review & editing. LR-M: Conceptualization, Investigation, Methodology, Project administration, Supervision, Writing – original draft, Writing – review & editing. RS: Conceptualization, Funding acquisition, Investigation, Methodology, Project administration, Resources, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was funded by FAPESP (Grants 2017/21052-0 and 2020/05110-2) to RS. GL has a scholarship from FAPESP (2019/20532-3).
Acknowledgments
The authors thank Nancy Bellei, Universidade Federal de São Paulo, for the SARS-CoV-2 PCR test; Paulo R. Abrão Ferreira, Jaquelina Sonoe Ota-Arakaki and Paula M. Peçanha-Pietrobom for the support with the selection and enrollment of patients. We thank Paolo Di Mascio and Graziella E. Ronsein for the mass spectrometry analyses performed at the Redox Proteomics Core of the Mass Spectrometry Resource at Chemistry Institute, University of São Paulo (FAPESP grant numbers 2012/12663-1 and 2016/00696-3, CEPID Redoxoma 2013/07937-8). The Figure 4 was created using NIAID NIH BIOART Source (https://bioart.niaid.nih.gov/).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2024.1508110/full#supplementary-material
Supplementary Material 1 | Methods, Tables, and Figures.
Supplementary Material 2 | List of differentially abundant proteins in between D0-wards and KTR controls.
Supplementary Material 3 | List of differentially abundant proteins in CS30 and KTR controls.
Abbreviations
CS30, Convalescent Sample 30 (30 days post-discharge); LDNs, Low-Density Neutrophils; MDSCs, Myeloid-Derived Suppressor Cells; NES, Normalized Enrichment Score; PMN-MDSCs, Polymorphonuclear Myeloid-Derived Suppressor Cells; SMD, Standardized Mean Difference; ssPSEA, Single-Sample Protein Set Enrichment Analysis; TMT, Tandem Mass Tag.
References
1. Naseer S, Khalid S, Parveen S, Abbass K, Song H, Achim MV. COVID-19 outbreak: Impact on global economy. Front Public Health. (2022) 10:1009393. doi: 10.3389/fpubh.2022.1009393
2. Squires N, Garfield R, Mohamed-Ahmed O, Iversen BG, Tegnell A, Fehr A, et al. Essential public health functions: the key to resilient health systems. BMJ Glob Health. (2023) 8(7). doi: 10.1136/bmjgh-2023-013136
3. Cascella M, Rajnik M, Aleem A, Dulebohn SC, Di Napoli R. Features, evaluation, and Treatment of Coronavirus (COVID-19). (2022). Available at: https://www.ncbi.nlm.nih.gov/books/NBK554776 (Accessed March 20, 2024).
4. Weiss A, Hendrickx R, Stensgaard E, Jellingsø M, Sommer MOA. Kidney transplant and dialysis patients remain at increased risk for succumbing to COVID-19. Transplantation. (2023) 107:1136–8. doi: 10.1097/tp.0000000000004462
5. Demir E, Ucar ZA, Dheir H, Danis R, Yelken B, Uyar M, et al. COVID-19 in kidney transplant recipients: A multicenter experience from the first two waves of pandemic. BMC Nephrol. (2022) 23:183. doi: 10.1186/s12882-022-02784-w
6. Sanders JF, Messchendorp AL, de Vries RD, Baan CC, van Baarle D, van Binnendijk R, et al. Antibody and T-cell responses 6 months after coronavirus disease 2019 messenger RNA-1273 vaccination in patients with chronic kidney disease, on dialysis, or living with a kidney transplant. Clin Infect Dis. (2023) 76:e188–e99. doi: 10.1093/cid/ciac557
7. Medina-Pestana J, Cristelli MP, Foresto RD, Tedesco-Silva H, Requião-Moura LR. The higher COVID-19 fatality rate among kidney transplant recipients calls for further action. Transplantation. (2022) 106:908–10. doi: 10.1097/tp.0000000000004086
8. Callaghan CJ, Mumford L, Curtis RMK, Williams SV, Whitaker H, Andrews N, et al. Real-world effectiveness of the pfizer-bioNTech BNT162b2 and oxford-astraZeneca chAdOx1-S vaccines against SARS-coV-2 in solid organ and islet transplant recipients. Transplantation. (2022) 106:436–46. doi: 10.1097/tp.0000000000004059
9. Choudhary D, Kenwar D, Sharma A, Bhalla A, Singh S, Singh MP, et al. Risk factors for mortality in kidney transplant recipients with COVID-19: a single centre experience and case–control study. BMC Nephrol. (2022) 23:241. doi: 10.1186/s12882-022-02821-8
10. Brunialti MKC, Leite GGF, Eburneo GS, de Araujo OR, Peçanha-Pietrobom PM, Ferreira PRA, et al. Patterns of circulating cytokines and vascular markers’ Response in the presence of COVID-19 in kidney transplant recipients compared with non-transplanted patients. Viruses. (2023) 15:2166. doi: 10.3390/v15112166
11. Devarajan P. Neutrophil gelatinase-associated lipocalin: a promising biomarker for human acute kidney injury. biomark Med. (2010) 4:265–80. doi: 10.2217/bmm.10.12
12. Requião-Moura LR, Sandes-Freitas TV, Viana LA, Cristelli MP, Andrade LGM, Garcia VD, et al. High mortality among kidney transplant recipients diagnosed with coronavirus disease 2019: Results from the Brazilian multicenter cohort study. PloS One. (2021) 16:e0254822. doi: 10.1371/journal.pone.0254822
13. Gisby J, Clarke CL, Medjeral-Thomas N, Malik TH, Papadaki A, Mortimer PM, et al. Longitudinal proteomic profiling of dialysis patients with COVID-19 reveals markers of severity and predictors of death. Elife. (2021) 10. doi: 10.7554/eLife.64827
14. Lin H, Ma X, Xiao F, Su H, Shi Y, Liu Y, et al. Identification of a special cell type as a determinant of the kidney tropism of SARS-CoV-2. FEBS J. (2021) 288:5163–78. doi: 10.1111/febs.16114
15. Peçanha-Pietrobom PM, Leite GGF, Hunter J, Ferreira PRA, Burattini MN, Bellei N, et al. The clinical course of hospitalized moderately ill COVID-19 patients is mirrored by routine hematologic tests and influenced by renal transplantation. PloS One. (2021) 16:e0258987. doi: 10.1371/journal.pone.0258987
16. Zeiser FA, Donida B, da Costa CA, Ramos G, Scherer JN, Barcellos NT, et al. First and second COVID-19 waves in Brazil: A cross-sectional study of patients&x2019; characteristics related to hospitalization and in-hospital mortality. Lancet Regional Health – Americas. (2022) 6. doi: 10.1016/j.lana.2021.100107
17. National Institutes of Health. COVID-19 Treatment Guidelines Panel. Coronavirus Disease 2019 (COVID-19) Treatment Guidelines. Available at: https://www.covid19treatmentguidelines.nih.gov/ (Accessed November 15, 2023).
18. World Health Organization. Clinical management of COVID-19: living guideline, 15 September 2022 (2022). Available at: https://apps.who.int/iris/handle/10665/362783 (Accessed November 15, 2023).
19. KDIGO AKI Work Group. KDIGO clinical practice guideline for acute kidney injury. Kidney Int Suppl. (2012) 2:1–141. doi: 10.1159/000339789
20. Bellomo R, Ronco C, Kellum JA, Mehta RL, Palevsky P. Acute Dialysis Quality Initiative w. Acute renal failure - definition, outcome measures, animal models, fluid therapy and information technology needs: the Second International Consensus Conference of the Acute Dialysis Quality Initiative (ADQI) Group. Crit Care. (2004) 8:R204–12. doi: 10.1186/cc2872
21. Michels EHA, Appelman B, Brabander Jd, Amstel R, Chouchane O, Linge C, et al. Age-related changes in plasma biomarkers and their association with mortality in COVID-19. Eur Respir J. (2023), 2300011. doi: 10.1183/13993003.00011-2023
22. Peters-Sengers H, Butler JM, Uhel F, Schultz MJ, Bonten MJ, Cremer OL, et al. Source-specific host response and outcomes in critically ill patients with sepsis: a prospective cohort study. Intensive Care Med. (2022) 48:92–102. doi: 10.1007/s00134-021-06574-0
23. Lin L, Aloe AM. Evaluation of various estimators for standardized mean difference in meta-analysis. Stat Med. (2021) 40:403–26. doi: 10.1002/sim.8781
24. Morrissey SM, Geller AE, Hu X, Tieri D, Ding C, Klaes CK, et al. A specific low-density neutrophil population correlates with hypercoagulation and disease severity in hospitalized COVID-19 patients. JCI Insight. (2021) 6(9). doi: 10.1172/jci.insight.148435
25. Carapito R, Li R, Helms J, Carapito C, Gujja S, Rolli V, et al. Identification of driver genes for critical forms of COVID-19 in a deeply phenotyped young patient cohort. Sci Transl Med. (2022) 14:eabj7521. doi: 10.1126/scitranslmed.abj7521
26. Leite GGF, Ferreira BL, Tashima AK, Nishiduka ES, Cunha-Neto E, Brunialti MKC, et al. Combined transcriptome and proteome leukocyte’s profiling reveals up-regulated module of genes/proteins related to low density neutrophils and impaired transcription and translation processes in clinical sepsis. Front Immunol. (2021) 12:744799. doi: 10.3389/fimmu.2021.744799
27. Eisfeld AJ, Halfmann PJ, Wendler JP, Kyle JE, Burnum-Johnson KE, Peralta Z, et al. Multi-platform ‘Omics analysis of human ebola virus disease pathogenesis. Cell Host Microbe. (2017) 22:817–29.e8. doi: 10.1016/j.chom.2017.10.011
28. Leite GGF, Brunialti MKC, Peçanha-Pietrobom PM, Ferreira PRA, Ota-Arakaki JS, Cunha-Neto E, et al. Understanding COVID-19 progression with longitudinal peripheral blood mononuclear cell proteomics: Changes in the cellular proteome over time. iScience. (2023) 26:107824. doi: 10.1016/j.isci.2023.107824
29. Potts M, Fletcher-Etherington A, Nightingale K, Mescia F, Bergamaschi L, Calero-Nieto FJ, et al. Proteomic analysis of circulating immune cells identifies cellular phenotypes associated with COVID-19 severity. Cell Rep. (2023) 42:112613. doi: 10.1016/j.celrep.2023.112613
30. Li J, Guo M, Tian X, Wang X, Yang X, Wu P, et al. Virus-host interactome and proteomic survey reveal potential virulence factors influencing SARS-coV-2 pathogenesis. Med (N Y). (2021) 2:99–112.e7. doi: 10.1016/j.medj.2020.07.002
31. Cron RQ, Caricchio R, Chatham WW. Calming the cytokine storm in COVID-19. Nat Med. (2021) 27:1674–5. doi: 10.1038/s41591-021-01500-9
32. Sims JT, Krishnan V, Chang CY, Engle SM, Casalini G, Rodgers GH, et al. Characterization of the cytokine storm reflects hyperinflammatory endothelial dysfunction in COVID-19. J Allergy Clin Immunol. (2021) 147:107–11. doi: 10.1016/j.jaci.2020.08.031
33. Cantenys-Molina S, Fernández-Cruz E, Francos P, Lopez Bernaldo de Quirós JC, Muñoz P, Gil-Herrera J. Lymphocyte subsets early predict mortality in a large series of hospitalized COVID-19 patients in Spain. Clin Exp Immunol. (2020) 203:424–32. doi: 10.1111/cei.13547
34. Huang M, Wang Y, Ye J, Da H, Fang S, Chen L. Dynamic changes of T-lymphocyte subsets and the correlations with 89 patients with coronavirus disease 2019 (COVID-19). Ann Transl Med. (2020) 8:1145. doi: 10.21037/atm-20-5479
35. Gu X, Wang S, Zhang W, Li C, Guo L, Wang Z, et al. Probing long COVID through a proteomic lens: a comprehensive two-year longitudinal cohort study of hospitalised survivors. eBioMedicine. (2023) 98. doi: 10.1016/j.ebiom.2023.104851
36. de Toledo MA, de Lima JVS, Salomão R, Leite GGF. Characterizing a Low-Density Neutrophil gene signature in acute and chronic infections and its impact on disease severity. bioRxiv. (2024). doi: 10.1101/2024.11.01.621545. 2024.11.01.621545.
37. Kelly B, O’Neill LA. Metabolic reprogramming in macrophages and dendritic cells in innate immunity. Cell Res. (2015) 25:771–84. doi: 10.1038/cr.2015.68
38. Maher AK, Burnham KL, Jones EM, Tan MMH, Saputil RC, Baillon L, et al. Transcriptional reprogramming from innate immune functions to a pro-thrombotic signature by monocytes in COVID-19. Nat Commun. (2022) 13:7947. doi: 10.1038/s41467-022-35638-y
39. Feys S, Vanmassenhove S, Kraisin S, Yu K, Jacobs C, Boeckx B, et al. Lower respiratory tract single-cell RNA sequencing and neutrophil extracellular trap profiling of COVID-19-associated pulmonary aspergillosis: a single centre, retrospective, observational study. Lancet Microbe. (2024) 5:e247–e60. doi: 10.1016/s2666-5247(23)00368-3
40. Nugent J, Aklilu A, Yamamoto Y, Simonov M, Li F, Biswas A, et al. Assessment of acute kidney injury and longitudinal kidney function after hospital discharge among patients with and without COVID-19. JAMA Netw Open. (2021) 4:e211095. doi: 10.1001/jamanetworkopen.2021.1095
41. Legrand M, Bell S, Forni L, Joannidis M, Koyner JL, Liu K, et al. Pathophysiology of COVID-19-associated acute kidney injury. Nat Rev Nephrol. (2021) 17:751–64. doi: 10.1038/s41581-021-00452-0
42. Wu H, Larsen CP, Hernandez-Arroyo CF, Mohamed MMB, Caza T, Sharshir M, et al. AKI and collapsing glomerulopathy associated with COVID-19 and APOL 1 high-risk genotype. J Am Soc Nephrol. (2020) 31:1688–95. doi: 10.1681/asn.2020050558
43. Paranjpe I, Jayaraman P, Su C-Y, Zhou S, Chen S, Thompson R, et al. Proteomic characterization of acute kidney injury in patients hospitalized with SARS-CoV2 infection. Commun Med. (2023) 3:81. doi: 10.1038/s43856-023-00307-8
44. Zhao J, Zhao J, Van Rooijen N, Perlman S. Evasion by stealth: inefficient immune activation underlies poor T cell response and severe disease in SARS-CoV-infected mice. PloS Pathog. (2009) 5:e1000636. doi: 10.1371/journal.ppat.1000636
45. Qin C, Zhou L, Hu Z, Zhang S, Yang S, Tao Y, et al. Dysregulation of immune response in patients with coronavirus 2019 (COVID-19) in wuhan, China. Clin Infect Dis. (2020) 71:762–8. doi: 10.1093/cid/ciaa248
46. Iannetta M, Buccisano F, Fraboni D, Malagnino V, Campogiani L, Teti E, et al. Baseline T-lymphocyte subset absolute counts can predict both outcome and severity in SARS-CoV-2 infected patients: a single center study. Sci Rep. (2021) 11:12762. doi: 10.1038/s41598-021-90983-0
47. Silvestre-Roig C, Fridlender ZG, Glogauer M, Scapini P. Neutrophil diversity in health and disease. Trends Immunol. (2019) 40:565–83. doi: 10.1016/j.it.2019.04.012
48. Carmona-Rivera C, Kaplan MJ. Low-density granulocytes: a distinct class of neutrophils in systemic autoimmunity. Semin Immunopathol. (2013) 35:455–63. doi: 10.1007/s00281-013-0375-7
49. Sun Z, Zhang Z, Banu K, Azzi YA, Reghuvaran A, Fredericks S, et al. Blood transcriptomes of SARS-coV-2-infected kidney transplant recipients associated with immune insufficiency proportionate to severity. J Am Soc Nephrol. (2022) 33:2108–22. doi: 10.1681/asn.2022010125
50. Barh D, Tiwari S, Rodrigues Gomes LG, Ramalho Pinto CH, Andrade BS, Ahmad S, et al. SARS-coV-2 variants show a gradual declining pathogenicity and pro-inflammatory cytokine stimulation, an increasing antigenic and anti-inflammatory cytokine induction, and rising structural protein instability: A minimal number genome-based approach. Inflammation. (2023) 46:297–312. doi: 10.1007/s10753-022-01734-w
Keywords: acute kidney injury, KTRs, mass spectrometry, PBMC, SARS-CoV-2, systems biology
Citation: Leite GGF, Sousa MB, Rodrigues LdOCP, Brunialti MKC, Medina-Pestana J, Butler JM, Peters-Sengers H, Requião-Moura L and Salomão R (2024) Proteomic profiling of peripheral blood mononuclear cells reveals immune dysregulation and metabolic alterations in kidney transplant recipients with COVID-19. Front. Immunol. 15:1508110. doi: 10.3389/fimmu.2024.1508110
Received: 08 October 2024; Accepted: 02 December 2024;
Published: 16 December 2024.
Edited by:
Pei-Hui Wang, Shandong University, ChinaReviewed by:
Georgia Damoraki, National and Kapodistrian University of Athens, GreecePrasanti Kotagiri, Monash University, Australia
Copyright © 2024 Leite, Sousa, Rodrigues, Brunialti, Medina-Pestana, Butler, Peters-Sengers, Requião-Moura and Salomão. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Giuseppe G. F. Leite, Z2l1c2VwcGUuZ2lhbmluaUB1bmlmZXNwLmJy; Reinaldo Salomão, cnNhbG9tYW9AdW5pZmVzcC5icg==