 Christian Zimmermann
Christian Zimmermann Sonja Dähn
Sonja Dähn Anika E. Wagner
Anika E. Wagner
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Immunol. , 13 May 2024
Sec. Comparative Immunology
Volume 15 - 2024 | https://doi.org/10.3389/fimmu.2024.1404086
This article is part of the Research Topic Sulforaphane and Isothiocyanates in Health View all 9 articles
Since infections with antibiotic-resistant bacteria cause increasing problems worldwide, the identification of alternative therapies is of great importance. Plant-derived bioactives, including allyl-isothiocyanate (AITC), have received attention for their antimicrobial properties. The present study therefore investigates the impact of AITC on survival and antimicrobial peptide (AMP) levels in Drosophila melanogaster challenged with the fly pathogenic bacteria Pectobacterium carotovorum subsp. carotovorum and Leuconostoc pseudomesenteroides. AITC, a sulfur-containing compound derived from glucosinolates, exhibits antimicrobial properties and has been suggested to modulate AMP expression. By using D. melanogaster, we demonstrate that AITC treatment resulted in a concentration-dependent decrease of survival rates among female flies, particularly in the presence of the Gram-negative bacterium Pectobacterium carotovorum subsp. carotovorum, whereas AITC did not affect survival in male flies. Despite the ability of isothiocyanates to induce AMP expression in cell culture, we did not detect significant changes in AMP mRNA levels in infected flies exposed to AITC. Our findings suggest sex-specific differences in response to AITC treatment and bacterial infections, underlining the complexity of host–pathogen interactions and potential limitations of AITC as a preventive or therapeutic compound at least in D. melanogaster models of bacterial infections.
The widespread use of antibiotics in livestock farming and medicine causes increasing problems with antibiotic-resistant pathogenic bacteria (1). In 2019, antibiotic-resistant bacteria have been made responsible for 1.27 million deaths worldwide (WHO) (2). The identification and the development of alternative therapies and prevention strategies is, therefore, of high importance for the global fight against severe bacterial infections. In this context, bioactive plant compounds may be a promising alternative in the treatment of diseases caused by antibiotic-resistant bacteria (3). Plant bioactives are compounds produced within the secondary metabolism of plants, which are used either as a defense mechanism against environmental stressors such as UV radiation, predators, and pathogenic microorganisms (4) or as coloring agents and odorants to attract pollinators (5, 6). Various plant bioactives have been shown to exhibit several health-promoting properties including anti-inflammatory, anti-infectious, anticancer, and antibacterial effects (7–10). Especially the sulfur-containing compounds such as isothiocyanates have been suggested to mediate antibacterial properties (8). In the presence of a neutral milieu, isothiocyanates are generated through hydrolysis from glucosinolates (GLS) by the enzymatic action of myrosinase, a thioglucohydrolase. While the GLS can be found in the vacuoles of cruciferous plant cells, myrosinase is generally present in separate myrosin cells and only gets in contact with GLS following the destruction of the plant cell by, e.g., chewing or cutting (11).
Allyl-isothiocyanate (AITC), the compound which is responsible for the pungent taste of, e.g., wasabi, radish, and mustard, is formed during the myrosinase-catalyzed conversion of the GLS sinigrin following cell disruption. In humans, several biological and health-promoting effects of AITC have been postulated, including anti-inflammatory and antioxidant effects as well as anticancer properties and antimicrobial activities (12–14). In particular, AITC has been able to inhibit the growth of different strains of Campylobacter jejuni (8), Escherichia coli, and Listeria monocytogenes in vitro (15). In cell culture studies, it has also been demonstrated that isothiocyanates are able to increase the expression levels of antimicrobial peptides (AMPs). In particular, it has been shown that the isothiocyanate sulforaphane induces the expression of human β-defensin-2 in intestinal epithelial cells (16). AMPs are known to play a critical role in the innate immune response and are produced by a variety of organisms including mammalian species, insects, and plants (17). They are mainly positively charged proteins and can act by membrane- or non-membrane-targeting mechanisms, thereby destroying bacteria, fungi, or viruses. Since recently, AMPs have been suggested as an alternative treatment option in the therapy of bacterial infections (1). Therefore, we suggest that AITC exhibits its antibacterial effect via increasing the expression levels of AMPs. To test this hypothesis, we applied the fruit fly Drosophila melanogaster as a model organism. D. melanogaster is especially known for its use in genetic research but has also been well established as a model to investigate and answer physiological questions (18). Due to its short generation time, the relatively inexpensive and easy maintenance, and the fact that 60% of the genes present have orthologs in mammals, the fruit fly is an ideal model for studying evolutionarily conserved genes and signaling pathways (19, 20). Due to the absence of an adaptive immune response in the fruit fly, processes of the innate immunity can be studied individually (21). In D. melanogaster, AMPs are produced in the fat body following systemic infections but also to a certain extent in the gut epithelia. The formation of AMPs in D. melanogaster is triggered by the activation of the two NFκB pathways, the toll- and the Imd pathway (22). This is highly similar to humans, where the expression of AMPs is also initiated by an activation of toll-like-receptors (23). To activate these pathways in D. melanogaster, we orally applied the two different fly-pathogenic bacterial strains Pectobacterium carotovorum subsp. carotovorum (ECC) and Leuconostoc pseudomesenteroides (LP), which have been shown to induce AMP secretion (24–26). ECC is a Gram-negative bacterium; therefore, it is a potential activator of the Imd pathway. LP is a Gram-positive bacterium, potentially activating the toll pathway (27). To test, whether AITC intervenes in these processes we fed flies AITC-supplemented diets, exposed them to the pathogenic bacteria and checked the flies for health-related parameters including survival and AMP expression.
Female and male w1118 D. melanogaster (Bloomington Drosophila Stock Center, Indiana, USA; #5905) were reared under standard conditions in a climate chamber (Memmert, HPP400, Büchenbach, Germany) at 25°C and 60% humidity with a 12 h/12 h light–dark cycle on Caltech Medium (CT), as described previously (28). For experiments, 3-day-old age-matched flies from synchronized eggs were anesthetized with CO2 or on ice and separated according to their sex. Then, 25 flies were transferred into vials containing either the control medium or bacterial suspension. The control medium comprises 10% sucrose (Carl Roth, Karlsruhe, Germany), 10% inactive yeast (Genesee via Kisker, Steinfurt, Germany), 2% agar (Apex via Kisker), and as preservatives 0.3% propionic acid (Carl Roth) and 1.5% tegosept (Apex via Kisker) (Linford et al., 2013). A 1-M stock solution of AITC (Sigma-Aldrich, Taufkirchen, Germany) was prepared in ethanol (abs.) (Merck, Darmstadt, Germany). To investigate the effect of AITC and/or bacteria on food intake, survival, and gene expression, different concentrations of AITC (0.250 mM, 0.125 mM) were added to the control medium. For control medium, the same amount of ethanol (abs.) was added to the food.
For infection studies, the fly pathogenic bacteria Pectobacterium carotovorum subsp. carotovorum (ECC) (Leibniz Institute DSMZ – German Collection of Microorganisms and Cell Cultures, Braunschweig, Germany) and Leuconostoc pseudomesenteroides (LP) (kind gift from Dr. Kwang-Zin Lee, Fraunhofer Institute for Molecular Biology and Applied Ecology, IME, Giessen, Germany) were grown in LB-Broth (Carl Roth) and MRS-Broth (Carl Roth), respectively. Both bacterial strains used were cultivated in a shaking incubator (B. Braun Biotech International, Melsungen, Germany) under aerobic conditions at 29°C and grown over night. In experiments, both bacterial strains were applied in their stationary growth phase.
For infection experiments, overnight cultures of ECC and LP were adjusted to an OD of 1 with a 100-mM sterile sucrose solution. Subsequently, 1 ml of the bacteria-sucrose suspension or sterile sucrose solution (control solution) was applied to three layers of cellulose paper lining the bottom of the respective vial. Three-day-old age-matched female and male D. melanogaster from synchronized eggs were transferred into these vials (25 flies per vial) and maintained under standard conditions for 18 h (see 2.1).
To check for potential effects of AITC on the feeding behavior of D. melanogaster, the flies’ food intake was determined by applying the gustatory assay according to Deshpande et al. (29). The flies received either a control or an AITC-supplemented diet for 10 days. On day 10, flies were transferred to control medium with and without AITC (0.125 mM and 0.250 mM) supplemented with 0.2% sulforhodamine B sodium salt (Sigma-Aldrich) and kept under standard conditions (see 2.1) for further 8 h. Then, 20 flies per treatment were transferred in 200 µl PBS (pH 7.4) (Thermo Fisher Scientific, Schwerte, Germany) with 1% Triton X-100 (Sigma-Aldrich) and homogenized in a TissueLyser II (Qiagen, Hilden, Germany) at a frequency of 25 s−1 for 6 min. Following, the obtained samples were centrifuged (4,000 g, 5 min) and the fluorescence signal (extinction: 535 nm/emission: 590 nm) of the supernatant was detected in a microplate reader SpectraMax iD3 (Molecular Devices, San Jose, USA). Flies fed unstained food served as controls, and the corresponding values were subtracted from the sample readings. Serial dilutions of sulforhodamine B sodium salt were used to generate a standard curve.
In order to investigate an effect of both, bacterial infection and AITC supplementation, on the survival rate of D. melanogaster, 3-day-old flies from synchronized eggs were separated according to their sex, sorted into different vials, and exposed to the corresponding treatments as depicted in Figure 1. Each experiment was performed two to three times with three vials per sex containing 25 flies each. Every 2–3 days, flies were transferred to new vials with fresh food, whereas dead flies were removed and counted.
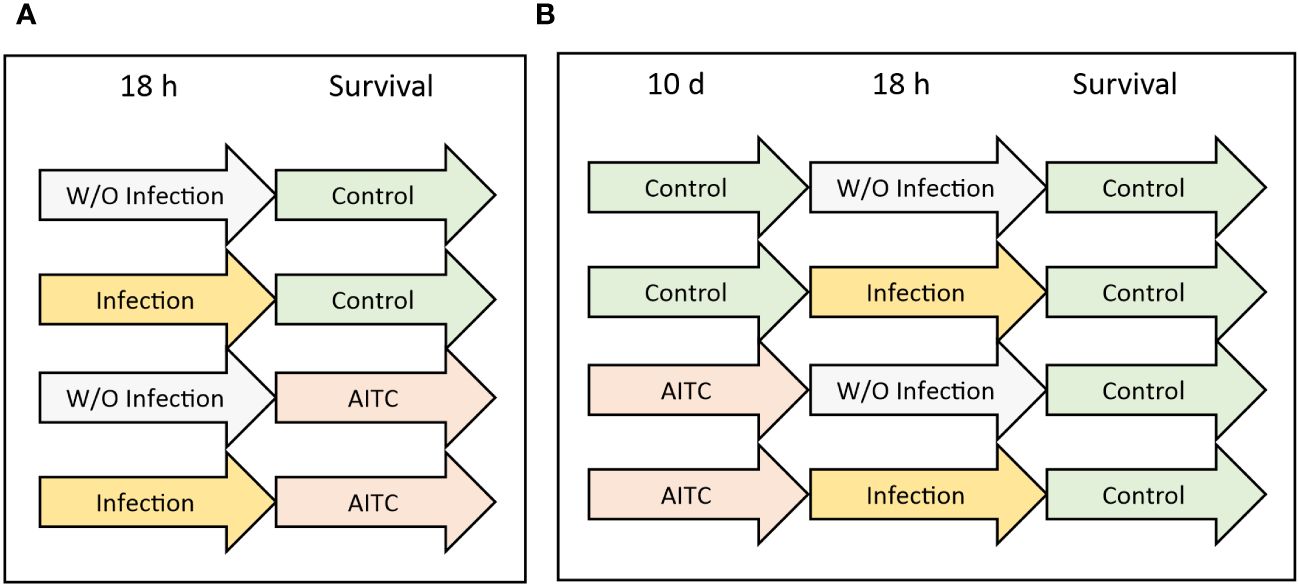
Figure 1 Experimental procedure of survival analysis. (A) D. melanogaster were initially exposed to a 100-mM sucrose solution with or without L. pseudomesenteroides or P. carotovorum subsp. carotovorum for 18 h and were subsequently transferred on either a control or an AITC-supplemented diet (0.125 mM or 0.250 mM) for the rest of their lifetime. (B) D. melanogaster were initially kept on either a control or an AITC-supplemented diet (0.125 mM or 0.250 mM) for 10 days, followed by an exposure to a 100-mM sucrose solution with or without L. pseudomesenteroides or P. carotovorum subsp. carotovorum for 18 h and were subsequently transferred on a control diet for the rest of their lifetime.
In order to investigate an effect of both, bacterial infection and AITC supplementation, on mRNA expression 3-day-old flies were exposed to the corresponding treatments as depicted in Figure 2. RNA was either isolated by applying TRI Reagent (Ambion, Carlsbad, USA) or the Quick-RNA Tissue/Insect kit (Zymo Research, Freiburg, Germany). In case of RNA isolation with TRI Reagent, five flies were put in a microcentrifuge tube containing 1 ml TRI Reagent and homogenized in a TissueLyser II (Qiagen) at a frequency of 25 s−1 for 6 min. After adding 200 µl chloroform, the samples were vortexed and incubated at room temperature for 10 min. Subsequently, the samples were centrifuged at 12,000 g at 4°C for 15 min. Then, the resulting upper, clear phase was transferred into a fresh microcentrifuge tube, mixed with 500 µl ice cold isopropanol, and left on ice for 10 min. Following, the samples were centrifuged at 12,000 g at 4°C for 15 min. The resulting supernatant was discarded, and the remaining pellet was washed twice with 500 µl ice cold 75% ethanol followed by centrifugation at 12,000 g at 4°C for 10 min. After removing the ethanol in the last washing step, the pellets were air-dried at room temperature for 30 min. Following, the dried pellet was resuspended in 50 µl RNase-free-water (Th. Geyer, Renningen, Germany) and dissolved at 55°C for 10 min. Subsequently, RNA samples were subjected to DNase treatment by applying the DNase kit (Sigma-Aldrich) according to the manufacturer’s instructions. In brief, 20 µl of isolated RNA was mixed with 2.5 µl DNase reaction buffer (Sigma-Aldrich) and 2.5 µl DNase I amplification grade (1 U/µl) (Sigma-Aldrich). After 15 min of incubation at room temperature, a 2.5-µl stop solution (Sigma-Aldrich) was added. Then, the samples were heated to 70°C for 10 min. Samples were stored at -80°C until further use.
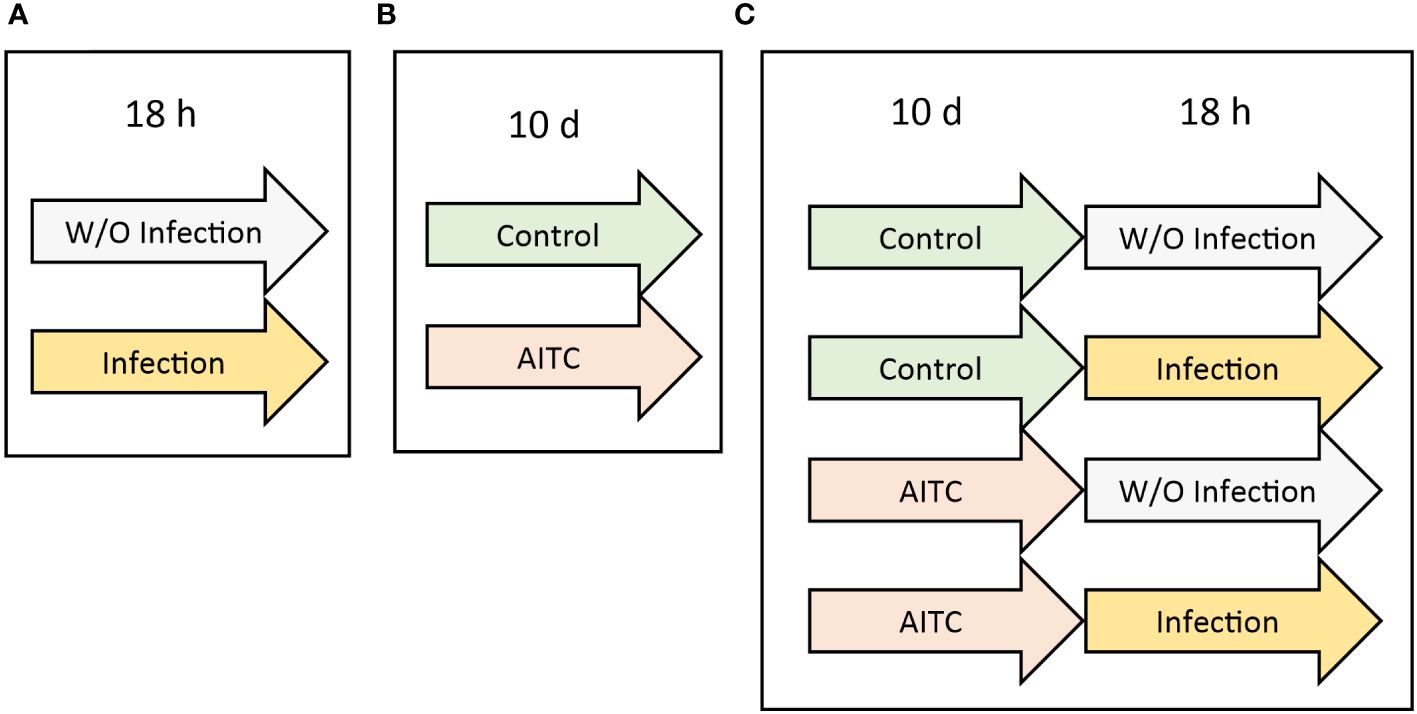
Figure 2 Experimental procedure of generating fly samples for subsequent mRNA analysis. (A) D. melanogaster were initially exposed to a 100-mM sucrose solution with P. carotovorum subsp. carotovorum or L. pseudomesenteroides for 18 h. (B) D. melanogaster were fed either a control diet or an AITC-supplemented diet (0.125 mM or 0.250 mM) for 10 days. (C) D. melanogaster were initially kept on either a control or an AITC-supplemented diet (0.125 mM or 0.250 mM) for 10 days and were subsequently transferred to a 100-mM sucrose solution with or without L. pseudomesenteroides or P. carotovorum subsp. carotovorum for 18 h. .
RNA isolation with the Quick-RNA Tissue/Insect kit (Zymo Research, Freiburg, Germany) was performed according to the manufacturer’s instructions. In brief, 10 flies were homogenized in RNA lysis buffer. After that, the homogenate was transferred into a column tube followed by several washing steps and DNase treatment. Finally, RNA was eluted in DNase/RNase-free water. RNA was stored at −80°C until further use.
The purity of the RNA samples was photometrically detected (260/280 nm) in a UVmini-1240 UV-VIS spectrophotometer (Shimadzu, Duisburg, Germany). Samples with a 260/280-nm ratio lower than 1.7 were excluded from further analysis.
For cDNA synthesis, 1 µg RNA was added to DNase/RNase-free-water, resulting in a total volume of 11.5 µl. Then, 1 µl oligo (dt) primers (20 ng/µl) (Promega, Mannheim, Germany), 4 µl M-MLV RT 5× Buffer (Promega), 0.5 µl RNasin RiboLock (40 U/µl) (Sigma-Aldrich), 2 µl dNTP mix (10 mM) (Promega), and 1 µl M-MLW reverse transcriptase (200 U/µl) (Promega) were added. Afterward, the samples were incubated at 42°C for 60 min and subsequently heated in a thermocycler (T-Gradient ThermoBlock, Biometra, Göttingen, Germany) at 70°C for 10 min. The obtained cDNA was stored at −80°C until further use.
For real-time PCR, cDNA was diluted 1:2 in DNase/RNase-free-water. The master mix contained 10 µl PerfeCTa SYBR Green SuperMix, Low ROX (Quantabio, Beverly, MA, USA), 0.25 µl forward/reverse primer (10 pmol/µl), 7.5 µl DNase/RNase-free-water, and 2 µl cDNA.
Semiquantitative real-time PCR was performed in a 7500 Real-Time PCR system (Applied Biosystems, Heidelberg, Germany). rPl32 (Ribosomal Protein L32), RpS20 (Ribosomal Protein S20), and alphaTub84B (alpha-Tubulin at 84B) were tested as reference genes. alphaTub84B showed treatment-dependent effects, why it was excluded as a reference gene. rPl32 and RpS20 were used as reference genes (see for RpS20 in Supplementary Table S1). Dro (Drosocin) and mtk (Metchnikowin) were measured as target genes (for primer sequences, see Table 1).
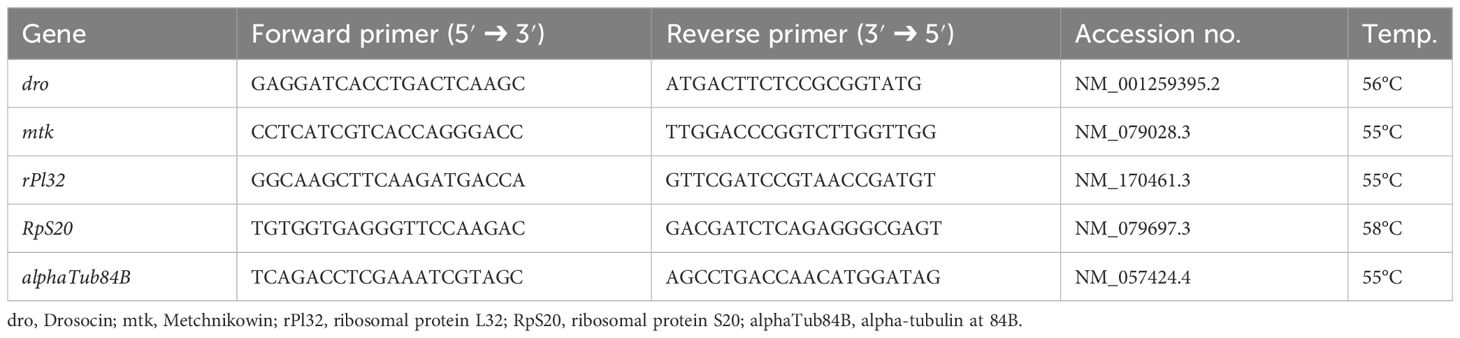
Table 1 Primer sequences for real-time PCR (Drosophila melanogaster).
Statistical analyses were carried out using GraphPad Prism Software (Version 10.1.2, GraphPad Software, LLC, San Diego, CA, USA). Unless otherwise stated, the experiments were performed in triplicates. Results are presented as mean ± standard deviation (SD) unless otherwise stated. Data were tested for normality of distribution (Shapiro–Wilk) and homogeneity of variances (Brown–Forsythe or Bartlett’s). The means of normally distributed data and homogenous variances were compared by one-way ANOVA followed by a post hoc test (Tukey’s) for multiple comparisons. Data with heterogeneous variances were tested with Brown–Forsythe–Welch ANOVA followed by a post hoc test (Dunnet’s T3) for multiple comparisons. Not normally distributed data were tested with a non-parametric test (Kruskal–Wallis). Survival analysis was performed by the Kaplan–Meier approach and a log-rank test to test for significant differences. Median survival times were calculated and tested for significant differences by the Mann–Whitney test. Significance was accepted at p < 0.05.
To test whether AITC enrichment at different concentrations has an effect on food intake in D. melanogaster, a gustatory assay was performed. The application of 0.125 mM AITC for 10 days did not affect the flies’ food intake, either in female or in male D. melanogaster. In contrast, addition of 0.250 mM AITC significantly reduced the food intake in male flies by approximately 7% but not in female flies (Figures 3A, B).
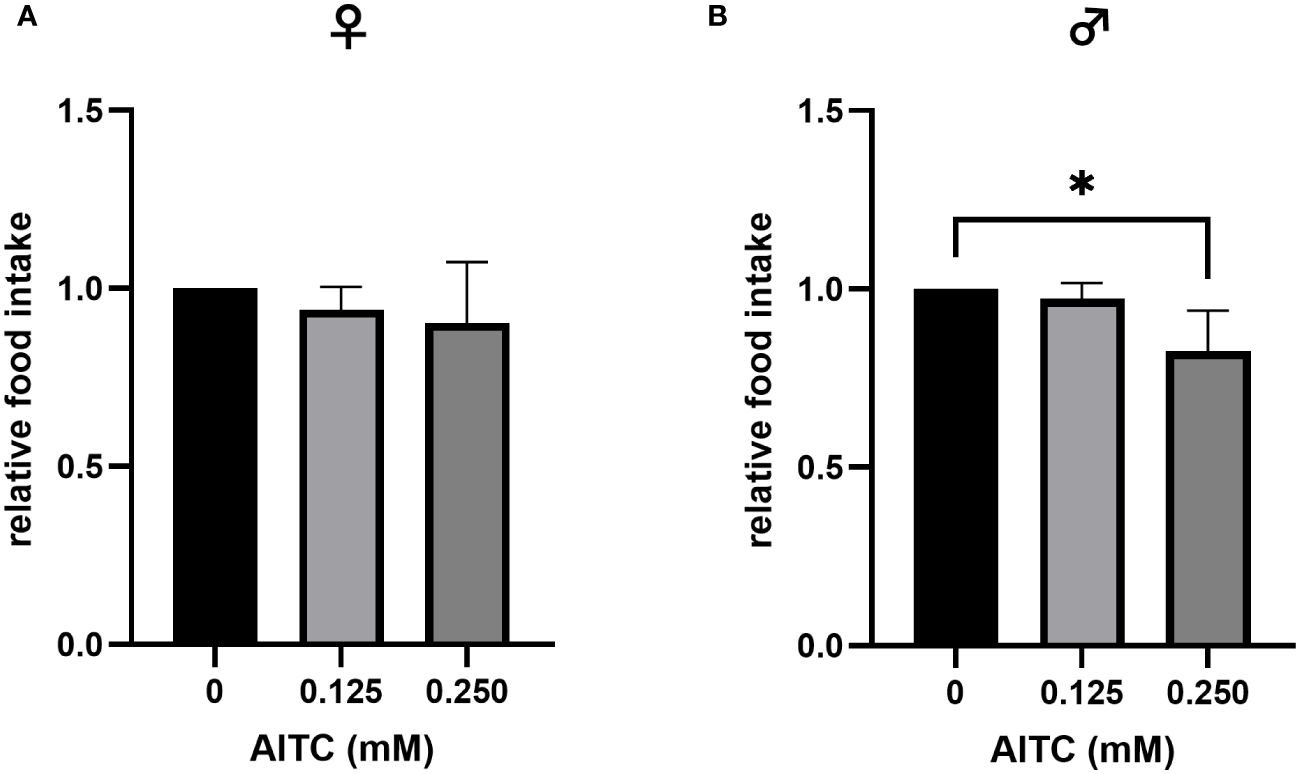
Figure 3 Relative food intake of female (A) and male (B) D. melanogaster fed either a control diet or an AITC-supplemented diet (0.125 mM or 0.250 mM), respectively, for 10 days. Bars show the mean ± SD of three independent experiments. Significant differences between groups were tested by applying one-way ANOVA followed by Dunnett’s multiple-comparison test. Significance was accepted at p < 0.05. *p < 0.05.
A survival assay was performed to investigate potential effects of both bacterial infection and AITC supplementation, on the flies’ survival rates. D. melanogaster initially exposed for 18 h to a 100-mM sucrose solution and subsequently transferred on an AITC-supplemented (Figure 4A) diet led to a significant concentration-dependent decrease of survival time of female flies. In contrast, no effects of AITC on male flies were observed. The medium survival times did not differ between the groups (Figures 4B, C). In female flies, infection with ECC resulted in a decreased survival whereas no significant effect was observed after LP infection. In contrast, LP infection of male flies resulted in an increased survival whereas ECC infection had no effect on the survival of male flies (Figures 4D, E). In D. melanogaster infected with either ECC or LP and subsequent exposure to AITC (Figure 5A), AITC exhibited a significant effect on survival times in ECC-infected female flies only (Figures 5B–E). In this case, the administration of 0.125 mM and 0.250 mM AITC significantly reduced the survival time in a concentration-dependent manner (Figure 5D).
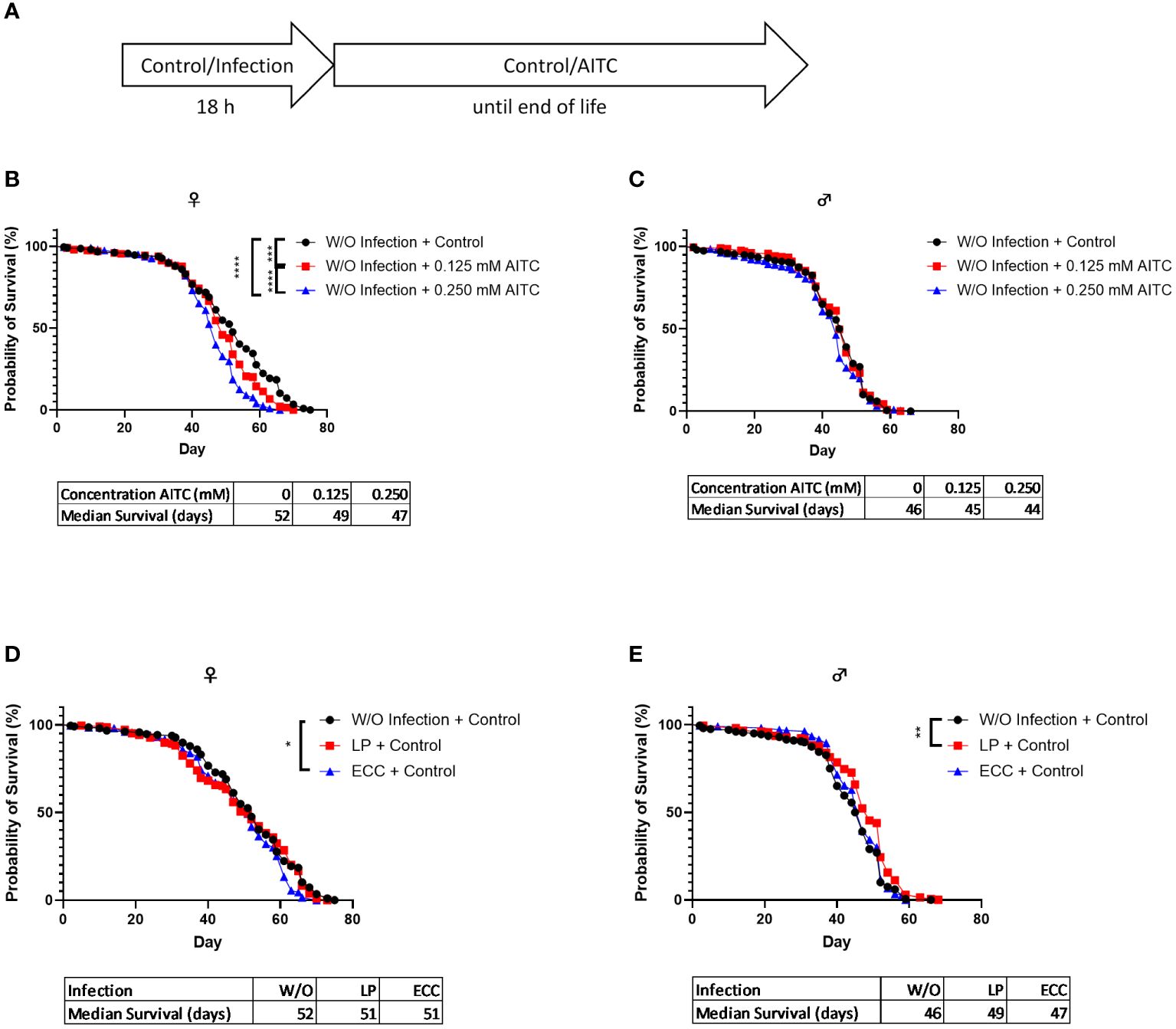
Figure 4 Survival curves and median survival time of female (B, D) and male (C, E) D. melanogaster initially exposed to a control solution (W/O Infection) (B, C) or a solution with L. pseudomesenteroides (LP) or P. carotovorum subsp. carotovorum (ECC) (D, E) for 18 h and subsequently transferred on either a control or an AITC-supplemented diet (0.125 mM or 0.250 mM) for the rest of their lifetime. The experiment procedure is shown in (A). Results show the three independent experiments. Significant differences between treatments were tested by the Kaplan–Meier approach and a log-rank test. Significance was accepted at p < 0.05. **p < 0.01, ***p < 0.001, ****p < 0.0001.
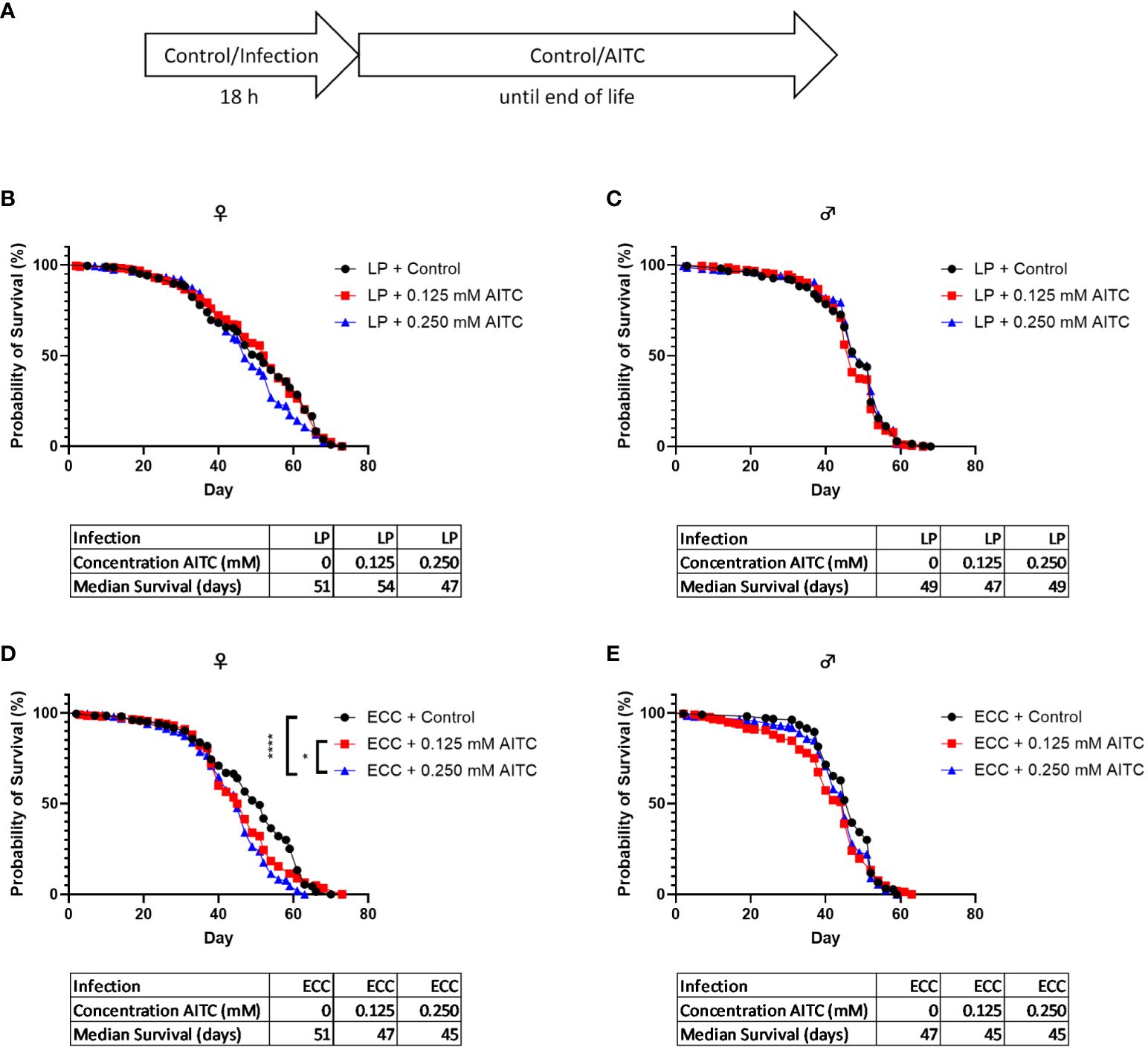
Figure 5 Survival curves and median survival time of female (B, D) and male (C, E) D. melanogaster initially exposed to a solution with L. pseudomesenteroides (LP) (B, C) or P. carotovorum subsp. carotovorum (ECC) (D, E) for 18 h and subsequently transferred on either a control or an AITC-supplemented diet (0.125 mM or 0.250 mM) for the rest of their lifetime. The experiment procedure is shown in (A). Results show three independent experiments. Significant differences between treatments were tested by the Kaplan–Meier approach and a log-rank test. Significance was accepted at p < 0.05. *p < 0.05, ****p < 0.0001.
D. melanogaster initially kept on an AITC-supplemented diet for 10 days (Figure 6A) did not show any difference in survival rates compared with flies reared on a control diet (Figures 6B, C). Both ECC and LP infections of female flies reared on a control diet for 10 days also did not have any effect on the survival rates (Figure 6D). In contrast, infection of male flies with ECC resulted in significantly higher survival rates whereas there was no significant difference in the median survival (Figure 6E). A pre-feeding with AITC for 10 days (Figure 7A) resulted in a concentration-dependent decrease in survival rates of ECC-infected female flies (Figure 7D) whereas in ECC-infected male flies, a reduced survival time was observed following exposure of the higher compared with the lower concentration of AITC (Figure 7E). AITC pre-feeding did not exhibit any effects on flies infected with LP (Figures 7B, C).
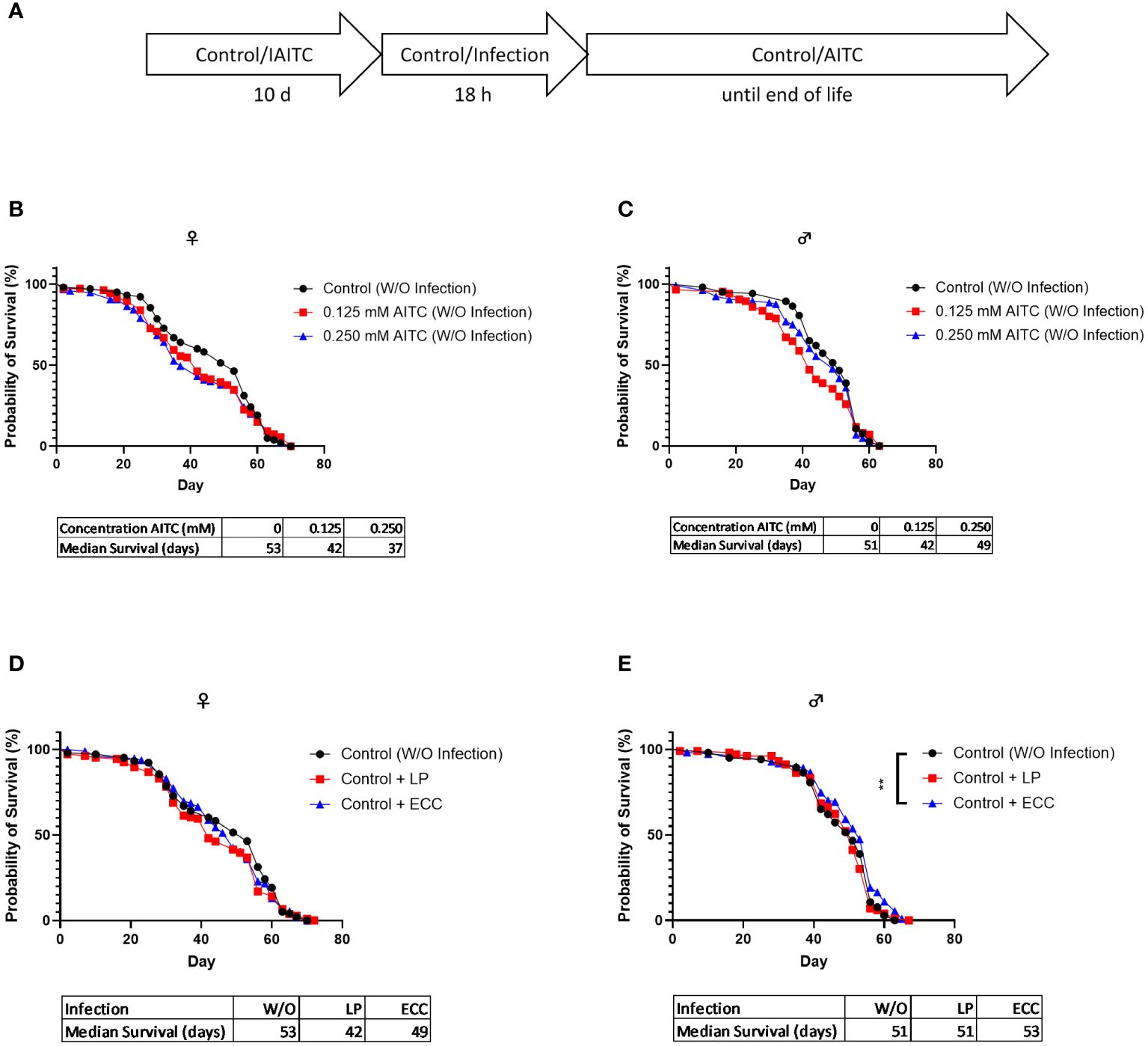
Figure 6 Survival curves and median survival time of female (B, D) and male (C, E) D. melanogaster initially kept on either a control or an AITC-supplemented diet (0.125 mM or 0.250 mM) for 10 days then exposed to a control solution (W/O infection) (B, C) or a solution with L. pseudomesenteroides (LP) or P. carotovorum subsp. carotovorum (ECC) (D, E) for 18 h and subsequently transferred on a control diet for the rest of their lifetime. The experiment procedure is shown in (A). Results show two independent experiments. Significant differences between treatments were tested by the Kaplan–Meier approach and a log-rank test. Significance was accepted at p < 0.05. **p < 0.01.
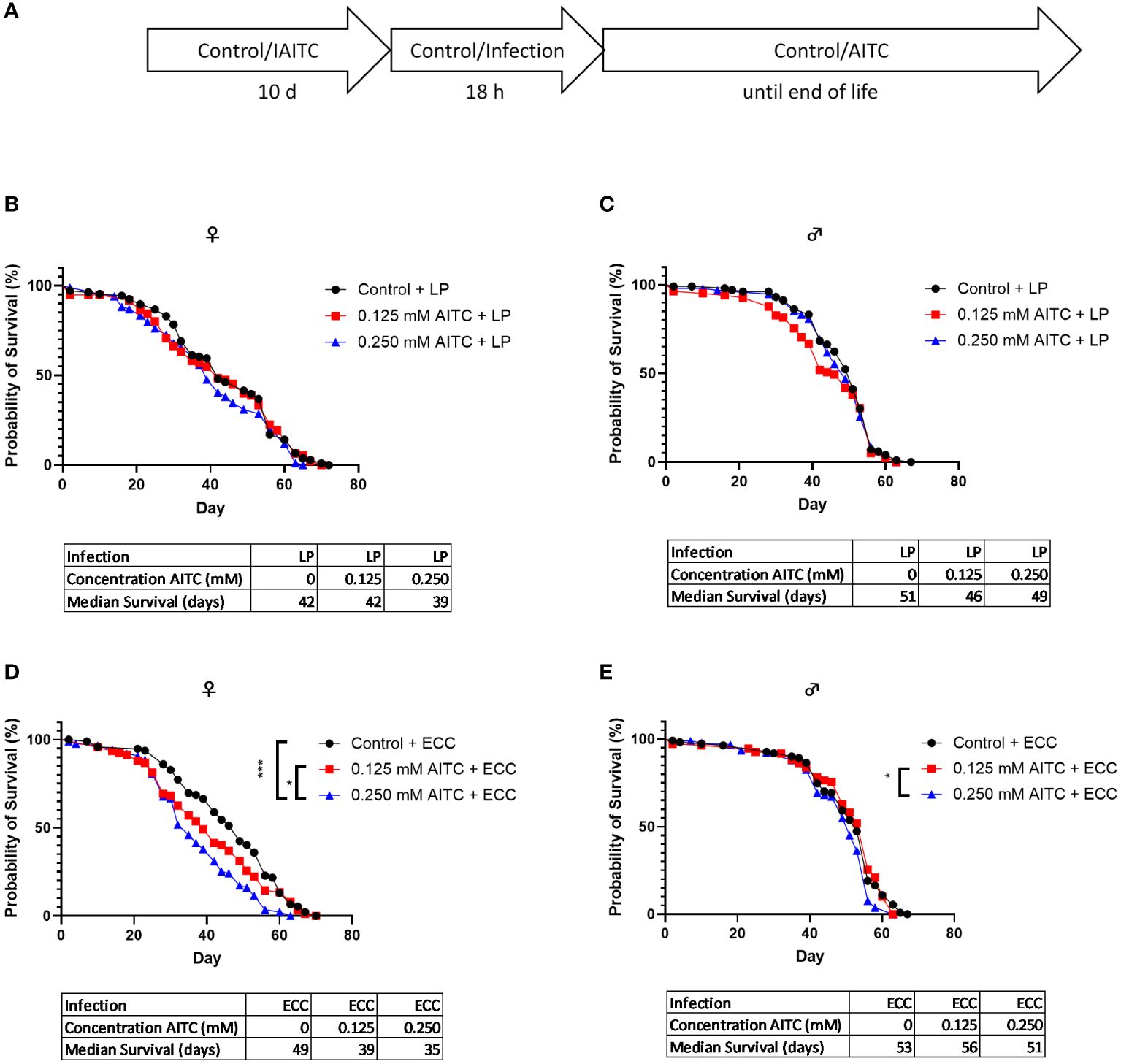
Figure 7 Survival curves and median survival time of female (B, D) and male (C, E) D. melanogaster initially kept either on a control or an AITC-supplemented diet (0.125 mM or 0.250 mM) for 10 days then exposed to a solution with L. pseudomesenteroides (LP) (B, C) or P. carotovorum subsp. carotovorum (ECC) (D, E) for 18 h and subsequently transferred on a control diet for the rest of their lifetime. The experiment procedure is shown in (A). Results show two independent experiments. Significant differences between treatments were tested by the Kaplan–Meier approach and a log-rank test. Significance was accepted at p < 0.05. *p < 0.05, **p < 0.01, ***p < 0.001.
In order to test for potential changes in the production of AMPs, gene expression levels of dro and mtk were examined. Neither in flies infected with ECC or LP for 18 h (Figures 8A, B and 9A, B) nor in flies kept on an AITC-supplemented diet for 10 days (Figures 8C, D and 9C, D) effects on either dro or mtk mRNA expression levels were observed. Flies initially kept on a control diet or an AITC-supplemented diet for 10 days with or without a subsequent bacterial infection did not show any significant changes in dro or mtk mRNA levels. This applied to infections with both bacterial strains and to both sexes (Figures 10A–H and 11B–H). Only ECC-infected female flies, pre-exposed to an AITC-supplemented diet for 10 days, showed a significant 4.5-fold increase of mtk expression levels compared with the uninfected controls (Figure 11A) whereas no significant effect was detected for dro expression levels (Figure 10A).
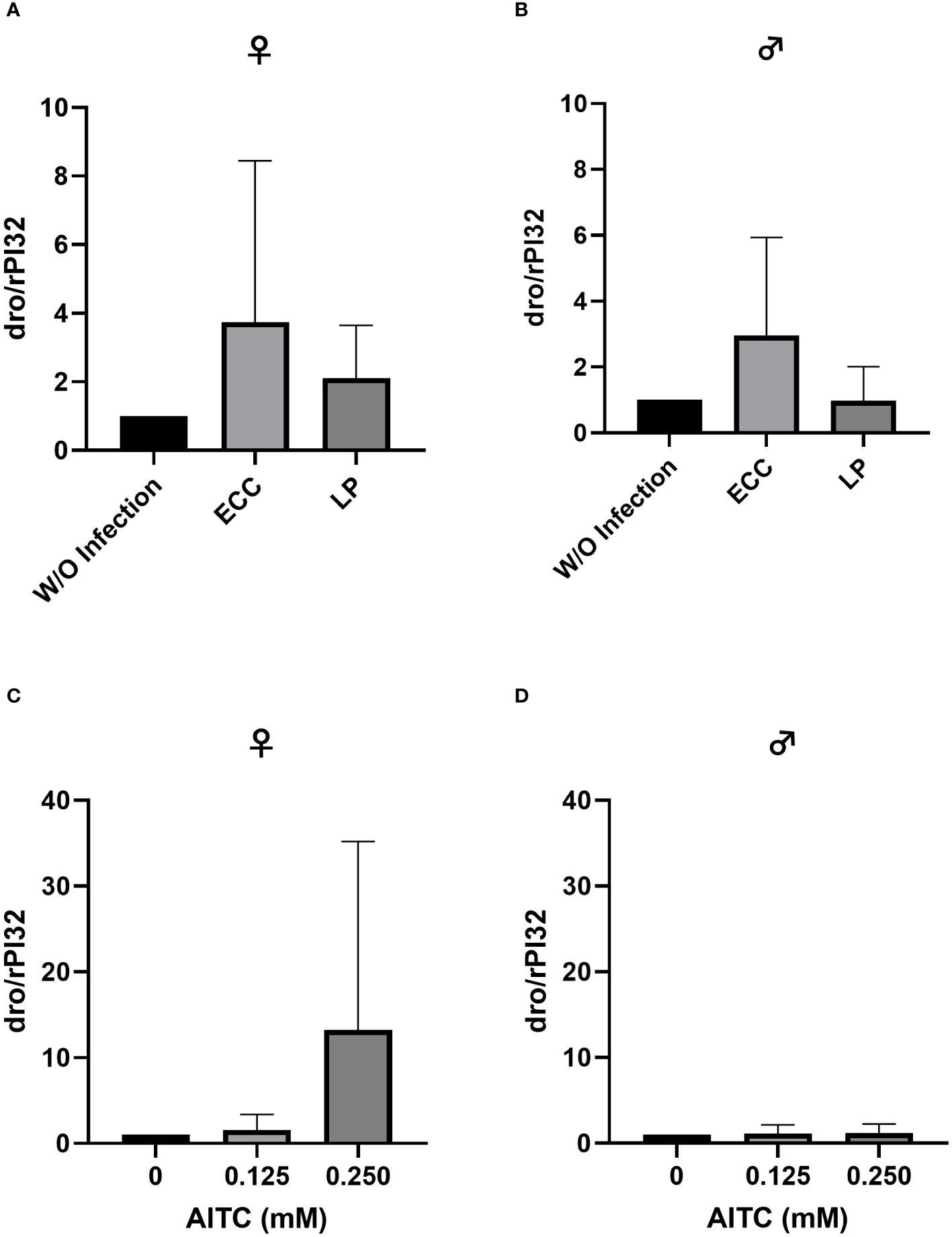
Figure 8 Relative mRNA expression levels of dro in female (A, C) and male (B, D) D. melanogaster exposed to a 100-mM sucrose solution with P. carotovorum subsp. carotovorum (ECC) or L. pseudomesenteroides (LP) for 18 h (A, B) and D. melanogaster fed either a control diet or an AITC-supplemented diet (0.125 mM or 0.250 mM) for 10 days (C, D). mRNA levels were determined in fly samples from three independent experiments with three replicates containing 10 flies each. Bars show the mean ± SD. Significant differences between groups were tested by applying one-way ANOVA followed by Dunnett’s multiple comparison test. Significance was accepted at p < 0.05.
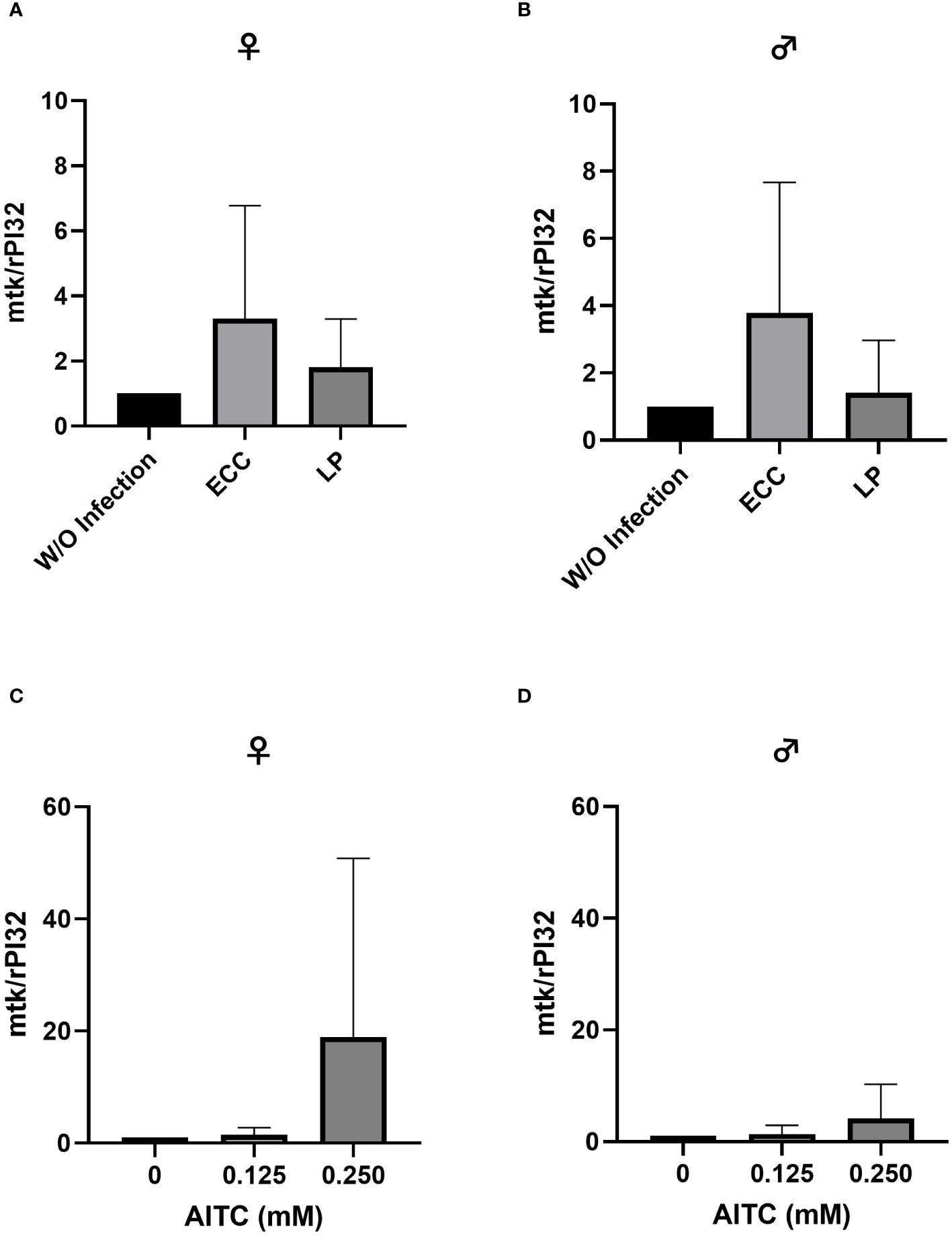
Figure 9 Relative mRNA expression levels of mtk in female (A, C) and male (B, D) D. melanogaster exposed to a 100-mM sucrose solution with P. carotovorum subsp. carotovorum (ECC) or L. pseudomesenteroides (LP) for 18 h (A, B) and D. melanogaster fed either a control diet or an AITC-supplemented diet (0.125 mM or 0.250 mM) for 10 days (C, D). mRNA levels were determined in fly samples from three independent experiments with three replicates containing 10 flies each. Bars show the mean ± SD. Significant differences between groups were tested by applying one-way ANOVA followed by Dunnett’s multiple-comparison test. Significance was accepted at p<0.05.
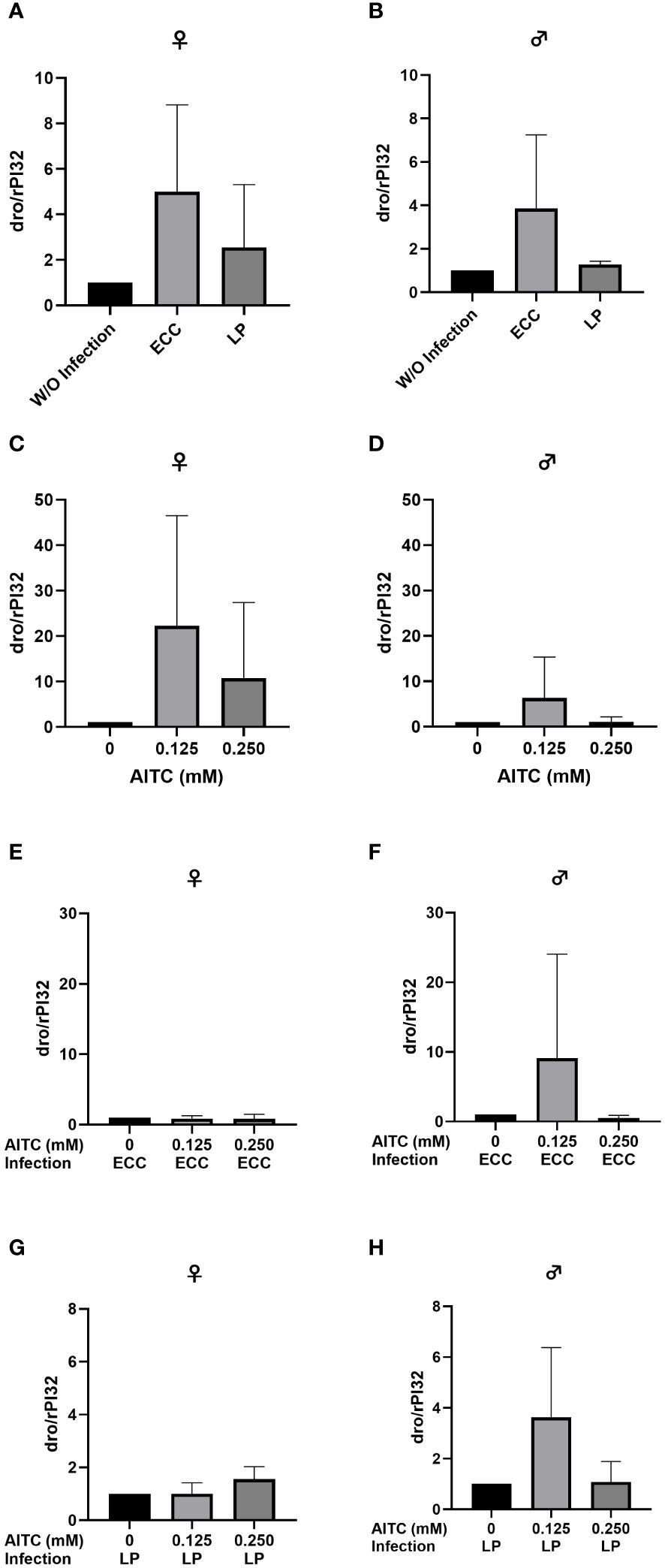
Figure 10 Relative mRNA expression levels of dro in female (A, C, E, G) and male (B, D, F, H) D. melanogaster initially kept on either a control or an AITC-supplemented diet (0.125 mM or 0.250 mM) for 10 days and subsequently exposed to a 100-mM sucrose solution with or without L. pseudomesenteroides (LP) or P. carotovorum subsp. carotovorum (ECC) for 18 h. mRNA levels were determined in fly samples from three independent experiments with three replicates containing 10 flies each. Bars show the mean ± SD. Significant differences between groups were tested by applying one-way ANOVA followed by Dunnett’s multiple-comparison test. Significance was accepted at p < 0.05.
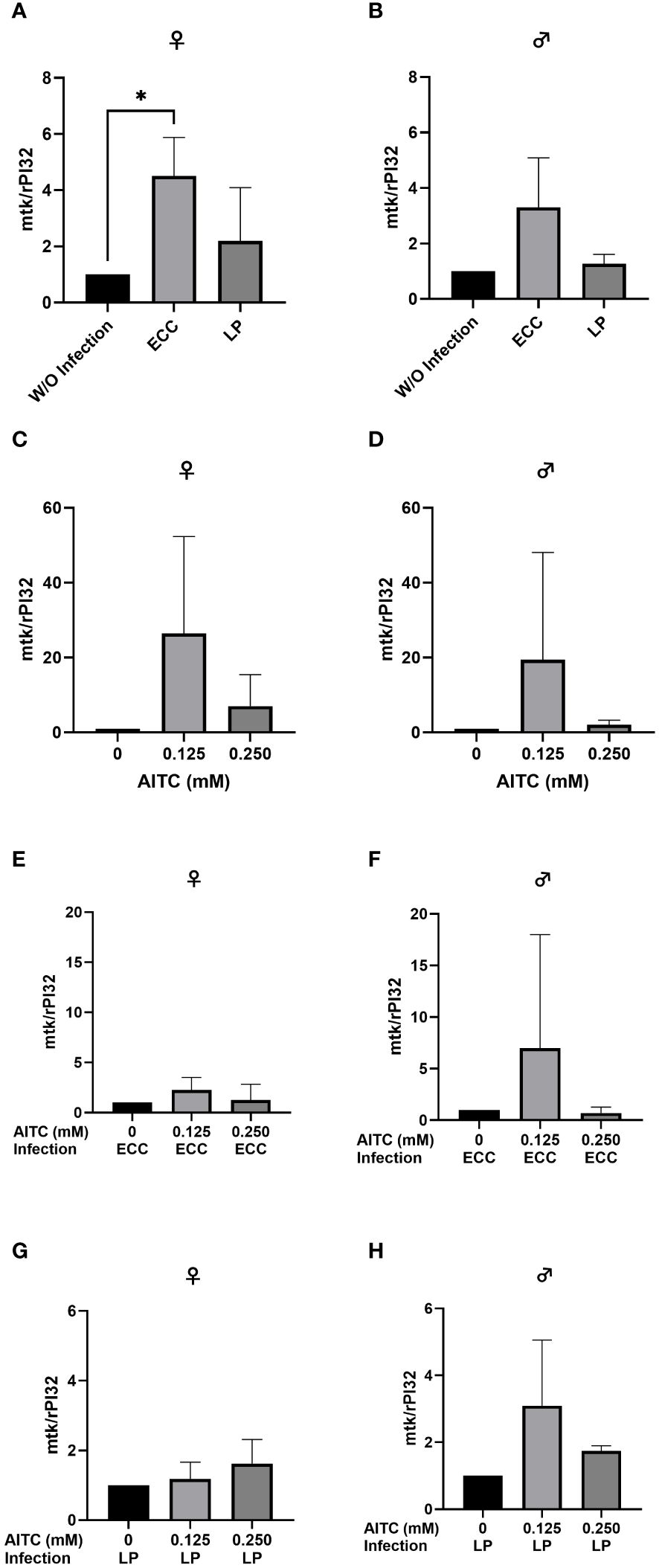
Figure 11 Relative mRNA expression levels of mtk in female (A, C, E, G) and male (B, D, F, H) D. melanogaster initially kept on either a control or an AITC-supplemented diet (0.125 mM or 0.250 mM) for 10 days and subsequently exposed to a 100-mM sucrose solution with or without L. pseudomesenteroides (LP) or P. carotovorum subsp. carotovorum (ECC) for 18 h. mRNA levels were determined in fly samples from three independent experiments with three replicates containing 10 flies each. Bars show the mean ± SD. Significant differences between groups were tested by applying one-way ANOVA followed by Dunnett’s multiple comparison test. Significance was accepted at p < 0.05. *p < 0.05.
The rise of bacterial resistance to antibiotics shows a need to identify alternative therapies. Antimicrobial peptides (AMPs) are emerging as promising candidates due to their potent antimicrobial properties (23, 30). Modulating the production of endogenous AMPs, as being already shown for the isothiocyanate sulforaphane in vitro, may offer a possibility for preventing bacterial infections (16).
We therefore asked whether AITC may be a potential candidate compound in the treatment of bacterial infections and whether it affects the endogenous production of AMPs. To test health-promoting effects, we initially analyzed the survival of fruit flies exposed to AITC and/or pathogenic bacteria. In male flies, we did not detect any effect of both applied AITC concentrations without the presence of pathogenic bacteria on survival time, neither in a short-term nor in a long-term treatment (Figures 4C, 6C). Intriguingly, we did not detect any effect of AITC on the food intake in female flies but in male flies where 0.250 mM AITC in the diet resulted in a significantly lower food intake (Figure 3B). This may explain the unaltered survival curves following AITC exposure in males compared with females where a long-term treatment with AITC resulted in a significant and dose-dependent decrease of the survival rates (Figure 4B). Due to the lower food intake of male flies, they potentially also consume less AITC, which could explain the difference in mortality compared with female flies. In addition, in male flies, AITC may have induced a kind of caloric restriction, which is well documented to increase lifespan in D. melanogaster (31). Interestingly, Merinas-Amo and colleagues (32) tested different cultivars of Brassica rapa with a high difference in their individual GLS and consequently isothiocyanate content on effects on the lifespan of D. melanogaster. The authors observed a significant dose-dependent increase of the flies’ lifespan, which was in contrast to the results of the present study where a significant decrease in the lifespan of female flies (Figure 4B) and no effect on male flies following exposure to AITC (Figure 4C) was detected. These differences may have occurred from the fact that our flies received the isolated isothiocyanate (and not the corresponding precursor sinigrin) via the diet and our results were generated sex-specifically.
One explanation for the detected higher mortality in female flies may have resulted from the applied AITC concentrations that were possibly too high. Insecticidal effects of AITC have been described for other insects but not for D. melanogaster (33–36). However, it should be considered that the increased mortality in most studies was caused by AITC applied as a volatile substance. Numerous positive effects were observed in other organisms after an oral ingestion of AITC or AITC-containing extracts possibly due to the intestinal metabolization of AITC (37–42).
However, Mazari et al., 2014 (43) did not detect any weakness or impaired phenotype in wild-type fruit flies exposed to 1 mM AITC in their diet. However, male flies being transgenic for GSTE7, an enzyme important for detoxifying xenobiotics, had a significantly higher mortality when exposed to phenethyl isothiocyanate for 1 week compared with female GSTE7 transgenic flies (43). This may explain the detected differences in the lifespan between male and female flies according to AITC treatment in the present study. Potentially, our male flies had higher GSTE7 levels compared with female flies resulting in an improved detoxification of AITC, which caused the better survival of male compared with female flies.
Although this does not support a potential toxic effect of AITC in our flies, it may be the reason for unchanged mtk and dro mRNA levels following the exposure to AITC. To examine if AITC exhibited health benefits and affected the mRNA expression of AMPs in pathogen-infected D. melanogaster, we orally applied the fly pathogenic bacteria ECC and LP to both female and male D. melanogaster. In order to study host–pathogen interactions including the production of AMPs, ECC-infected D. melanogaster have already been used (44, 45). Piegholdt et al. (45) showed that ECC induced the mRNA expression levels of the AMPs mtk, dro, attB (attacin B), attC (attacin C), and dpt (diptericin). In addition, ECC exposure reduced the survival of our D. melanogaster under nutrient deficiency (Supplementary Figure S1), as shown for other bacteria (46, 47). For LP-mediated infections, to the best of our knowledge, there is currently only information for bacteriocins produced by LP but no information on a potential effect on AMP expression levels available. However, LP infection also reduced the survival of fruit flies under nutrient deficiency (Supplementary Figure S1), which supports the findings of Hiebert et al. (47). In case of an oral 18-h-infection of 3-day-old female D. melanogaster under sufficient nutrient supply, we observed a significant reduction in survival time following ECC treatment (Figure 4D). In contrast, an infection of 13-day-old female flies had no effect on survival (Figure 6D). In male flies, an infection with LP at the age of 3 days and an infection with ECC at the age of 13 days even resulted in a significant increase in survival time (Figure 4E and Figure 6E). This may be due to a sex-specific response to bacterial infections. It was demonstrated that in general males and females in various species differ in their immunological responses. When challenged with antigens, females generally have a more protective humoral and cell-mediated immune response, whereas for males, more intense inflammatory immune responses were described (48, 49). In D. melanogaster, female flies were described to be more likely than male flies to die from infections with several strains of Beauveria bassiana, which was postulated to be substantiated from sex-specific differences in the toll and the Imd pathway (50). These pathways are activated by bacteria and fungi consequently inducing the production of AMPs in D. melanogaster (27), which only owns an innate immune system (21). The negative effect of AITC on the survival time of female D. melanogaster appeared to reinforce the influence of ECC, as an AITC dose-dependent reduction in survival time was observed in both, in an infection followed by long-term treatment with AITC (Figure 5D) and in a 10-days treatment with AITC followed by a subsequent infection (Figure 7D).
When looking at the expression levels of AMPs, no significant differences were detected for dro and mtk after an 18-h-infection with ECC and LP of 3-day-old female and male flies (Figure 8A and Figure 9A). However, it has to be considered that larger variations between the experimental replicates may have masked a potential increase in AMP expression levels. Since we analyzed the AMP expression in the whole organism, a detection of AMP levels in individual organs might have reduced the observed variations in our data. In contrast, the infection of 13-day-old females with ECC led to a significant higher expression level of mtk (Figure 11A). Rera et al. (51) showed that the expression of AMPs was tightly related to a dysfunction of the intestinal barrier, which increased in the course of life and therefore caused higher AMP levels in older flies (52–54). These adverse effects on the intestinal barrier may be further increased by pathogenic infections. With regard to the AMP expression levels in our flies being exposed to AITC, no effect of both AITC concentrations was observed (Figures 8-11). This was also true for additional bacterial infections, where AITC was applied after oral bacterial infections (Figures 10, 11). Perhaps the larger variations between the experimental replicates, which could be due to differences between the individual flies, masked a potential statistically significant difference between the AMP expression levels.
In female fruit flies, the response to infections also seems to depend on whether they have already mated. If there was only a limited amount of energy available, the fly had to decide where to invest the energy—either in the process of reproduction or in the immune defense. Experiments conducted by Gordon and colleagues (55) support this assumption since infected mated female flies had a lower rate of survival, a higher bacterial load, and lower AMP levels in comparison with unmated females. As mated female flies were used in our experiments this could—at least partly—explain why we did not detect many differences in the expression levels of dro and mtk in the infected flies.
Overall, the results of the present study indicate that AITC treatments caused a significant decrease in survival among females but not males. Furthermore, AITC reduced the survival of females even in the presence of ECC, but not in the presence of LP, whereas no effect of AITC on survival rates were observed in infected male flies. Similarly, AITC did not significantly affect AMP levels in either infected male or infected female flies. The observed differences in survival between female and male flies may depend on sex-specific variations in immune responses as evidenced by the differing outcomes following bacterial infection in males. The high variations in mRNA expression levels of AMPs due to differences between experimental replicates may have obscured significant differences. Therefore, further experiments are needed to investigate the impact of AITC on pathogenic bacterial infections in fruit flies in more depth and with a specific focus on inflammatory signaling pathways.
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements because study included the use of lower invertebrate animals (Drosophila melanogaster).
CZ: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing. SD: Investigation, Methodology, Writing – original draft, Writing – review & editing. AW: Conceptualization, Project administration, Supervision, Writing – original draft, Writing – review & editing.
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
We would like to thank Cordula Becker for excellent technical assistance and Maria Theresia Schminke for experimental support.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2024.1404086/full#supplementary-material
1. Huan Y, Kong Q, Mou H, Yi H. Antimicrobial peptides: Classification, design, application and research progress in multiple fields. Front Microbiol. (2020) 11:582779. doi: 10.3389/fmicb.2020.582779
2. Murray CJL, Ikuta KS, Sharara F, Swetschinski L, Robles Aguilar G, Gray A, et al. Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. Lancet. (2022) 399:629–55. doi: 10.1016/S0140–6736(21)02724–0
3. Álvarez-Martínez FJ, Barrajón-Catalán E, Micol V. Tackling antibiotic resistance with compounds of natural origin: A comprehensive review. Biomedicines. (2020) 8:405. doi: 10.3390/biomedicines8100405
4. Petruk G, Del Giudice R, Rigano MM, Monti DM. Antioxidants from plants protect against skin photoaging. Oxid Med Cell Longev. (2018) 2018:1454936. doi: 10.1155/2018/1454936
5. Davies KM, Jibran R, Zhou Y, Albert NW, Brummell DA, Jordan BR, et al. The evolution of flavonoid biosynthesis: A bryophyte perspective. Front Plant Sci. (2020) 11:7. doi: 10.3389/fpls.2020.00007
6. Zhang P, Zhu H. Anthocyanins in plant food: current status, genetic modification, and future perspectives. Molecules. (2023) 28:866. doi: 10.3390/molecules28020866
7. Zimmermann C, Wagner AE. Impact of food-derived bioactive compounds on intestinal immunity. Biomolecules. (2021) 11:1901. doi: 10.3390/biom11121901
8. Dufour V, Alazzam B, Ermel G, Thepaut M, Rossero A, Tresse O, et al. Antimicrobial activities of isothiocyanates against Campylobacter jejuni isolates. Front Cell Infect Microbiol. (2012) 2:53. doi: 10.3389/fcimb.2012.00053
9. Keyvani-Ghamsari S, Rahimi M, Khorsandi K. An update on the potential mechanism of gallic acid as an antibacterial and anticancer agent. Food Sci Nutr. (2023) 11:5856–72. doi: 10.1002/fsn3.3615
10. Duda-Chodak A, Tarko T, Petka-Poniatowska K. Antimicrobial compounds in food packaging. Int J Mol Sci. (2023) 24:2457. doi: 10.3390/ijms24032457
11. Sturm C, Wagner AE. Brassica-derived plant bioactives as modulators of chemopreventive and inflammatory signaling pathways. Int J Mol Sci. (2017) 18:1890. doi: 10.3390/ijms18091890
12. Rajakumar T, Pugalendhi P. Allyl isothiocyanate regulates oxidative stress, inflammation, cell proliferation, cell cycle arrest, apoptosis, angiogenesis, invasion and metastasis via interaction with multiple cell signaling pathways. Histochem Cell Biol. (2023) 161:211–21. doi: 10.1007/s00418–023-02255–9
13. Tran HT, Stetter R, Herz C, Spöttel J, Krell M, Hanschen FS, et al. Allyl isothiocyanate: A TAS2R38 receptor-dependent immune modulator at the interface between personalized medicine and nutrition. Front Immunol. (2021) 12:669005. doi: 10.3389/fimmu.2021.669005
14. Tarar A, Peng S, Cheema S, Peng C-A. Anticancer activity, mechanism, and delivery of allyl isothiocyanate. Bioengineering (Basel). (2022) 9:470. doi: 10.3390/bioengineering9090470
15. Lin CM, Preston JF, Wei CI. Antibacterial mechanism of allyl isothiocyanate. J Food Prot. (2000) 63:727–34. doi: 10.4315/0362–028x-63.6.727
16. Schwab M, Reynders V, Loitsch S, Steinhilber D, Schröder O, Stein J. The dietary histone deacetylase inhibitor sulforaphane induces human beta-defensin-2 in intestinal epithelial cells. Immunology. (2008) 125:241–51. doi: 10.1111/j.1365-2567.2008.02834.x
17. Mazurkiewicz-Pisarek A, Baran J, Ciach T. Antimicrobial peptides: Challenging journey to the pharmaceutical, biomedical, and cosmeceutical use. Int J Mol Sci. (2023) 24:9031. doi: 10.3390/ijms24109031
18. Droujinine IA, Perrimon N. Interorgan communication pathways in physiology: Focus on drosophila. Annu Rev Genet. (2016) 50:539–70. doi: 10.1146/annurev-genet-121415–122024
19. Baenas N, Wagner AE. Drosophila melanogaster as an alternative model organism in nutrigenomics. Genes Nutr. (2019) 14:14. doi: 10.1186/s12263-019-0641-y
20. Staats S, Lüersen K, Wagner AE, Rimbach G. Drosophila melanogaster as a versatile model organism in food and nutrition research. J Agric Food Chem. (2018) 66:3737–53. doi: 10.1021/acs.jafc.7b05900
21. Yu S, Luo F, Xu Y, Zhang Y, Jin LH. Drosophila innate immunity involves multiple signaling pathways and coordinated communication between different tissues. Front Immunol. (2022) 13:905370. doi: 10.3389/fimmu.2022.905370
22. Hanson MA, Dostálová A, Ceroni C, Poidevin M, Kondo S, Lemaitre B. Synergy and remarkable specificity of antimicrobial peptides in vivo using a systematic knockout approach. Elife. (2019) 8:e44341. doi: 10.7554/eLife.44341
23. Campbell Y, Fantacone ML, Gombart AF. Regulation of antimicrobial peptide gene expression by nutrients and by-products of microbial metabolism. Eur J Nutr. (2012) 51:899–907. doi: 10.1007/s00394–012-0415–4
24. Gendrin M, Welchman DP, Poidevin M, Hervé M, Lemaitre B. Long-range activation of systemic immunity through peptidoglycan diffusion in Drosophila. PloS Pathog. (2009) 5:e1000694. doi: 10.1371/journal.ppat.1000694
25. Sawa N, Okamura K, Zendo T, Himeno K, Nakayama J, Sonomoto K. Identification and characterization of novel multiple bacteriocins produced by Leuconostoc pseudomesenteroides QU 15. J Appl Microbiol. (2010) 109:282–91. doi: 10.1111/j.1365-2672.2009.04653.x
26. Makhloufi KM, Carré-Mlouka A, Peduzzi J, Lombard C, van Reenen CA, Dicks LM, et al. Characterization of leucocin B-KM432Bz from Leuconostoc pseudomesenteroides isolated from boza, and comparison of its efficiency to pediocin PA-1. PloS One. (2013) 8:e70484. doi: 10.1371/journal.pone.0070484
27. Hanson MA, Lemaitre B. New insights on Drosophila antimicrobial peptide function in host defense and beyond. Curr Opin Immunol. (2020) 62:22–30. doi: 10.1016/j.coi.2019.11.008
28. Wagner AE, Piegholdt S, Rabe D, Baenas N, Schloesser A, Eggersdorfer M, et al. Epigallocatechin gallate affects glucose metabolism and increases fitness and lifespan in Drosophila melanogaster. Oncotarget. (2015) 6:30568–78. doi: 10.18632/oncotarget.5215
29. Deshpande SA, Carvalho GB, Amador A, Phillips AM, Hoxha S, Lizotte KJ, et al. Quantifying Drosophila food intake: comparative analysis of current methodology. Nat Methods. (2014) 11:535–40. doi: 10.1038/nmeth.2899
30. Xuan J, Feng W, Wang J, Wang R, Zhang B, Bo L, et al. Antimicrobial peptides for combating drug-resistant bacterial infections. Drug Resist Update. (2023) 68:100954. doi: 10.1016/j.drup.2023.100954
31. Kapahi P, Kaeberlein M, Hansen M. Dietary restriction and lifespan: Lessons from invertebrate models. Ageing Res Rev. (2017) 39:3–14. doi: 10.1016/j.arr.2016.12.005
32. Merinas-Amo T, Lozano-Baena M-D, Obregón-Cano S, Alonso-Moraga Á, de Haro-Bailón A. Role of glucosinolates in the nutraceutical potential of selected cultivars of brassica rapa. Foods. (2021) 10:2720. doi: 10.3390/foods10112720
33. Wu H, Zhang G-A, Zeng S, Lin K. Extraction of allyl isothiocyanate from horseradish (Armoracia rusticana) and its fumigant insecticidal activity on four stored-product pests of paddy. Pest Manag Sci. (2009) 65:1003–8. doi: 10.1002/ps.1786
34. Vilela Ad, Faroni LR, Sousa AH, Pimentel MA, Gomes JL. Toxicological and physiological effects of allyl isothiocyanate upon Callosobruchus maculatus. J Stored Products Res. (2020) 87:101625. doi: 10.1016/j.jspr.2020.101625
35. Shi C-H, Hu J-R, Xie W, Yang Y-T, Wang S-L, Zhang Y-J. Control of bradysia odoriphaga (Diptera: sciaridae) with allyl isothiocyanate under field and greenhouse conditions. J Econ Entomol. (2017) 110:1127–32. doi: 10.1093/jee/tow303
36. Jabeen A, Zaitoon A, Lim L-T, Scott-Dupree C. Toxicity of five plant volatiles to adult and egg stages of drosophila suzukii matsumura (Diptera: Drosophilidae), the spotted-wing drosophila. J Agric Food Chem. (2021) 69:9511–9. doi: 10.1021/acs.jafc.1c01384
37. Hasegawa K, Miwa S, Tsutsumiuchi K, Miwa J. Allyl isothiocyanate that induces GST and UGT expression confers oxidative stress resistance on C. elegans, as demonstrated by nematode biosensor. PloS One. (2010) 5:e9267. doi: 10.1371/journal.pone.0009267
38. Yamasaki M, Ogawa T, Wang L, Katsube T, Yamasaki Y, Sun X, et al. Anti-obesity effects of hot water extract from Wasabi (Wasabia japonica Matsum.) leaves in mice fed high-fat diets. Nutr Res Pract. (2013) 7:267–72. doi: 10.4162/nrp.2013.7.4.267
39. Ahn J, Lee H, Im SW, Jung CH, Ha TY. Allyl isothiocyanate ameliorates insulin resistance through the regulation of mitochondrial function. J Nutr Biochem. (2014) 25:1026–34. doi: 10.1016/j.jnutbio.2014.05.006
40. Thejass P, Kuttan G. Allyl isothiocyanate (AITC) and phenyl isothiocyanate (PITC) inhibit tumour-specific angiogenesis by downregulating nitric oxide (NO) and tumour necrosis factor-alpha (TNF-alpha) production. Nitric Oxide. (2007) 16:247–57. doi: 10.1016/j.niox.2006.09.006
41. Rajakumar T, Pugalendhi P, Thilagavathi S. Protective effect of allyl isothiocyanate on glycoprotein components in 7,12-dimethylbenz(a)anthracene induced mammary carcinoma in rats. Indian J Clin Biochem. (2018) 33:171–7. doi: 10.1007/s12291–017-0663–4
42. Bhattacharya A, Li Y, Wade KL, Paonessa JD, Fahey JW, Zhang Y. Allyl isothiocyanate-rich mustard seed powder inhibits bladder cancer growth and muscle invasion. Carcinogenesis. (2010) 31:2105–10. doi: 10.1093/carcin/bgq202
43. Mazari AM, Dahlberg O, Mannervik B, Mannervik M. Overexpression of glutathione transferase E7 in Drosophila differentially impacts toxicity of organic isothiocyanates in males and females. PloS One. (2014) 9:e110103. doi: 10.1371/journal.pone.0110103
44. Basset A, Khush RS, Braun A, Gardan L, Boccard F, Hoffmann JA, et al. The phytopathogenic bacteria Erwinia carotovora infects Drosophila and activates an immune response. Proc Natl Acad Sci U.S.A. (2000) 97:3376–81. doi: 10.1073/pnas.97.7.3376
45. Piegholdt S, Rimbach G, Wagner AE. Effects of the isoflavone prunetin on gut health and stress response in male Drosophila melanogaster. Redox Biol. (2016) 8:119–26. doi: 10.1016/j.redox.2016.01.001
46. Hiebert N, Carrau T, Bartling M, Vilcinskas A, Lee K-Z. Identification of entomopathogenic bacteria associated with the invasive pest Drosophila suzukii in infested areas of Germany. J Invertebr Pathol. (2020) 173:107389. doi: 10.1016/j.jip.2020.107389
47. Hiebert N, Kessel T, Skaljac M, Spohn M, Vilcinskas A, Lee K-Z. The Gram-Positive Bacterium Leuconostoc pseudomesenteroides Shows Insecticidal Activity against Drosophilid and Aphid Pests. Insects. (2020) 11:471. doi: 10.3390/insects11080471
48. Marriott I, Huet-Hudson YM. Sexual dimorphism in innate immune responses to infectious organisms. Immunol Res. (2006) 34:177–92. doi: 10.1385/IR:34:3:177
49. Klein SL, Flanagan KL. Sex differences in immune responses. Nat Rev Immunol. (2016) 16:626–38. doi: 10.1038/nri.2016.90
50. Shahrestani P, Chambers M, Vandenberg J, Garcia K, Malaret G, Chowdhury P, et al. Sexual dimorphism in Drosophila melanogaster survival of Beauveria bassiana infection depends on core immune signaling. Sci Rep. (2018) 8:12501. doi: 10.1038/s41598–018-30527–1
51. Rera M, Clark RI, Walker DW. Intestinal barrier dysfunction links metabolic and inflammatory markers of aging to death in Drosophila. Proc Natl Acad Sci U.S.A. (2012) 109:21528–33. doi: 10.1073/pnas.1215849110
52. Landis GN, Abdueva D, Skvortsov D, Yang J, Rabin BE, Carrick J, et al. Similar gene expression patterns characterize aging and oxidative stress in Drosophila melanogaster. Proc Natl Acad Sci U.S.A. (2004) 101:7663–8. doi: 10.1073/pnas.0307605101
53. Pletcher SD, Macdonald SJ, Marguerie R, Certa U, Stearns SC, Goldstein DB, et al. Genome-wide transcript profiles in aging and calorically restricted Drosophila melanogaster. Curr Biol. (2002) 12:712–23. doi: 10.1016/s0960–9822(02)00808–4
54. Zerofsky M, Harel E, Silverman N, Tatar M. Aging of the innate immune response in Drosophila melanogaster. Aging Cell. (2005) 4:103–8. doi: 10.1111/j.1474-9728.2005.00147.x
Keywords: allyl-isothiocyanate, anti-microbial peptides, Drosophila melanogaster, infection, bioactive plant compound, survival, gene expression
Citation: Zimmermann C, Dähn S and Wagner AE (2024) Effect of allyl-isothiocyanate on survival and antimicrobial peptide expression following oral bacterial infections in Drosophila melanogaster. Front. Immunol. 15:1404086. doi: 10.3389/fimmu.2024.1404086
Received: 27 March 2024; Accepted: 24 April 2024;
Published: 13 May 2024.
Edited by:
Diego A. Moreno, Spanish National Research Council (CSIC), SpainReviewed by:
Ilias Kounatidis, The Open University, United KingdomCopyright © 2024 Zimmermann, Dähn and Wagner. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Christian Zimmermann, Y2hyaXN0aWFuLmEuemltbWVybWFubkBlcm5hZWhydW5nLnVuaS1naWVzc2VuLmRl
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.