 Junlei Hou1,2†
Junlei Hou1,2† Bo Zhu
Bo Zhu Haoran Zha
Haoran Zha
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Immunol. , 21 March 2024
Sec. Cancer Immunity and Immunotherapy
Volume 15 - 2024 | https://doi.org/10.3389/fimmu.2024.1371559
This article is part of the Research Topic Community Series in Biomarkers in the Era of Cancer Immunotherapy: Zooming in from Periphery to Tumor Microenvironment, Volume II View all 14 articles
Anti-PD-(L)1 therapy has shown great efficacy in some patients with cancer. However, a significant proportion of patients with cancer do not respond to it. Another unmet clinical need for anti-PD-(L)1 therapy is the dynamic monitoring of treatment effects. Therefore, identifying biomarkers that can stratify potential responders before PD-(L)1 treatment and timely monitoring of the efficacy of PD-(L)1 treatment are crucial in the clinical setting. The identification of biomarkers by liquid biopsy has attracted considerable attention. Among the identified biomarkers, circulating T cells are one of the most promising because of their indispensable contribution to anti-PD-(L)1 therapy. The present review aimed to thoroughly explore the potential of circulating T cells as biomarkers of anti-PD-(L)1 therapy and its advantages and limitations.
Over the past decade, anti-PD-(L)1 therapy has revolutionized the clinical treatment of cancer. However, only some patients with cancer benefit from anti-PD-(L)1 therapy, whereas most patients eventually experience disease progression. Furthermore, a few patients receiving anti-PD-(L)1 therapy succumb to hyperprogressive diseases. Therefore, identifying biomarkers that can stratify patients who will benefit from anti-PD-(L)1 therapy is crucial in clinical practice.
The US Food and Drug Administration (FDA) has approved some biomarkers that can predict the efficacy of anti-PD-(L)1 therapy in several cancer types, including PD-L1 expression (assessed using immunohistochemistry), mismatch repair deficient/microsatellite instability-high (dMMR/MSI-high), and tumor mutation burden (TMB). These biomarkers are mostly based on tumor lesions sampled through invasive surgery or biopsy. Unfortunately, Previous studies have revealed that tumors usually exhibit high temporal and spatial heterogeneity (1), making them unrepresentative of the entire immune landscape based on a biopsy sample from a single site. For example, approximately 55% of patients with advanced non-small cell lung cancer (NSCLC) who exhibit PD-L1 expression in at least 50% of tumor cells do not benefit from the therapy (2). Therefore, there is an urgent need to develop new approaches using robust biomarkers associated with anti-PD-(L)1 therapy.
Liquid biopsy has emerged as an appealing method for identifying new biomarkers that reflect the general immune landscape (3, 4). Among the identified biomarkers, circulating T cells are one of the most promising for three reasons: 1) It has been historically accepted that effective anti-PD-(L)1 therapy relies on the reactivation of pre-existing T cells of the tumor, which exhibit high programmed cell death protein 1 (PD-1) expression levels; however, recent data suggest that effective anti-PD-(L)1 therapy relies on the continuous recruitment of new T cells from circulation (5). Accordingly, recent studies demonstrated the existence of anti-PD-(L)1 therapy-responding T cell subsets in circulation (6, 7); 2) A previous theory suggested that activated T cells infiltrate the tumor, become dysfunctional, and finally die within tumor microenvironment (TME); however, recent research indicates that some tumor-infiltrating T cells can escape from the tumor and re-enter circulation (8). indicating that some circulating T cell subsets may reflect the status of tumor-infiltrating T cells; 3) Given the gut microbiota is involved in determining the efficacy of anti-PD-(L)1 therapy (9), circulating T cells against specific ectopic bacteria may serve as a biomarker of anti-PD-(L)1 therapy. Consistently, recent research has indicated that bacteria-specific T follicular helper (Tfh) cells exist in the circulation and are correlated with PD-(L)1 efficacy (10).
This review aimed to thoroughly explore circulating T cell subsets in the context of PD-(L)1 efficacy. The advantages and limitations of the different subsets and functional states of circulating T cells were also discussed.
The number of peripheral CD8+ T cell subsets is closely associated with immunotherapy efficacy (11–13). Herein, we reviewed the predictive impact of peripheral blood CD8+ T cell subsets on efficacy before and after anti-PD-(L)1 therapy (Table 1, Figure 1).
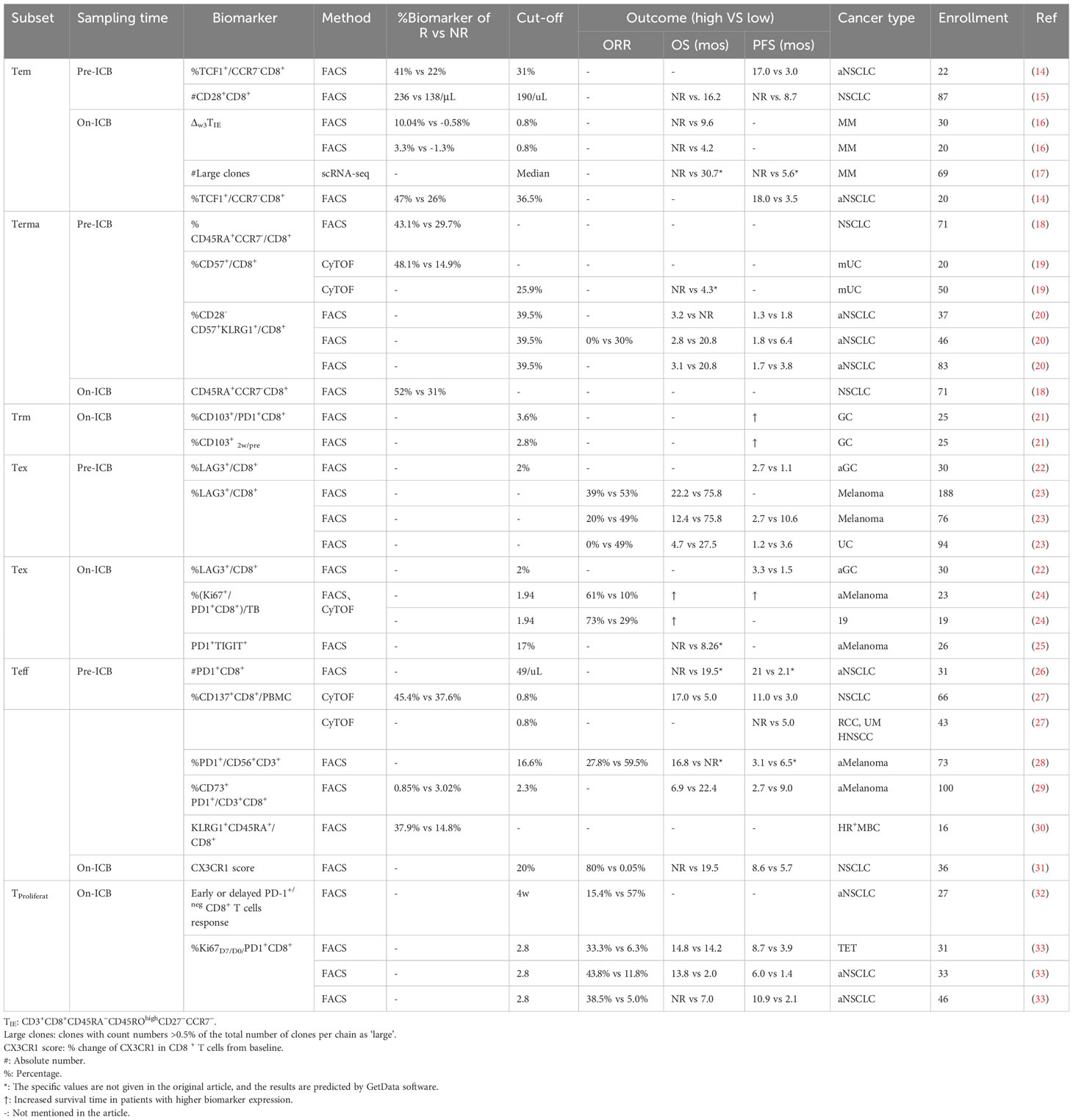
Table 1 Predictive outcome of peripheral conventional CD8+ T cell subsets in PD-(L)1 therapy.
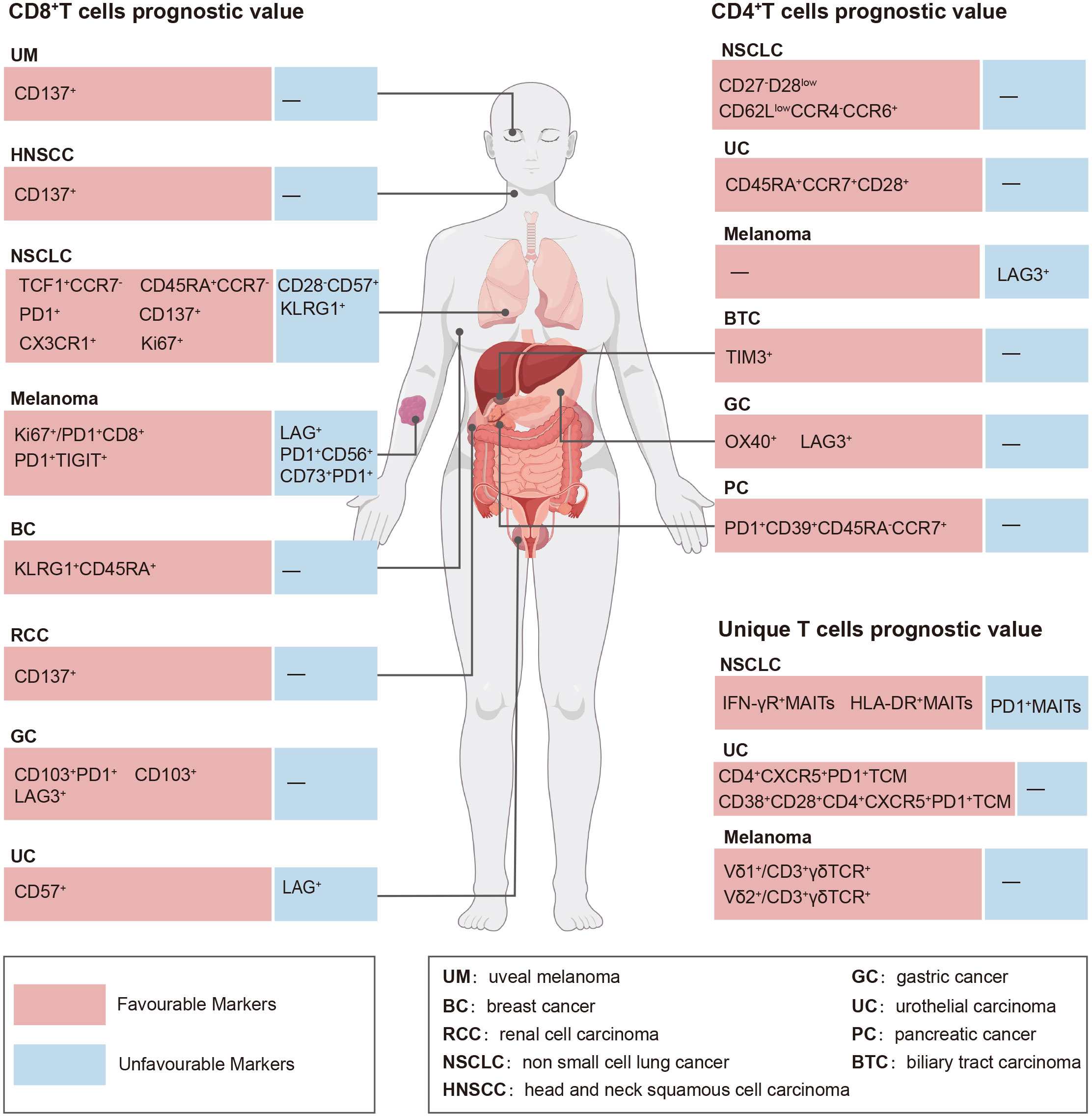
Figure 1 Prognostic value of circulating T cell subsets in the context of PD-(L)1 efficacy.
Upon antigen stimulation, naïve T cells differentiate into T effector (Teffs) cells. After antigen elimination, most Teff cells undergo apoptosis, whereas a small proportion of Teff cells differentiate into long-lived memory T cells (34). Memory CD8+ T cells recirculating throughout the bloodstream can induce a rapid and robust response upon antigen reengagement, thereby playing a crucial role in sustaining long-lasting protective immunity. Circulating memory CD8+ T cells can be categorized into three distinct subtypes: stem central memory (Tscm), central memory (Tcm), and effector memory (Tem). Furthermore, memory CD8+ T cells that reside within affected tissues and exhibit limited recirculation capacity are called tissue-resident memory T (Trm) cells. Trm cells can also re-enter circulation under specific contexts (35). Current evidence indicates that memory CD8+ T cells in the peripheral blood correlate with responsiveness to immunotherapy, indicating their predictive value as a biomarker.
Tem cells are usually characterized by a CD45RO+C-C motif chemokine receptor 7 (CCR7)-killer cell lectin-like receptor G1 (KLRG1)high phenotype in patients with cancer. Tem cells emerge among early responders to immunotherapy, and their early expansion in the circulation is correlated with a durable response and improved objective response rates (16). In addition, Tem cells persist in the peripheral blood of patients with melanoma who experience durable benefits from immunotherapy (36).
Tem cells have been extensively studied as predictive markers of the baseline response to anti-PD-(L)1 therapy in patients with NSCLC (14, 15). Recent studies have demonstrated that anti-PD-(L)1 therapy does not reverse the phenotype of terminally exhausted T cells but promotes the differentiation of self-renewing progenitor T cells into newly formed effector-like T cells, including memory T cells (37). T-cell factor 1 (TCF1) is an established marker of self-renewing T cells. In patients with NSCLC receiving anti-PD-(L)1 therapy, a higher proportion of TCF1-expressing T cells was observed in the circulating CD8+ Tem cells of patients who achieved durable clinical benefit (DCB) at baseline than in those who were resistant to treatment. Higher TCF-1 expression was associated with longer progression-free survival (PFS) (14). CD28 is a surface marker of TCF1-expressing CD8+ cells. Analysis of pre-treatment peripheral blood lymphocytes from 87 patients with different tumors undergoing first-line anti-PD-(L)1 therapy revealed significantly elevated circulating CD8+CD28+ T cell counts in patients who responded to treatment (median [range] counts: 236 (30–536) vs. 138 [36-460]/μL). Using 190/μL as the cut-off, patients with higher CD8+CD28+ T cell counts exhibited significantly prolonged median PFS than patients with lower counts (not reached vs. 8.7 months, p < 0.001). In addition, they had a significantly extended overall survival (OS) (not reached vs. 16.2 months, p < 0.001) (15). These findings suggest that Tem cells are strongly associated with response to PD(L)1 therapy.
In a clinical study investigating the impact of single-cycle anti-PD-(L)1 therapy on peripheral T cell dynamics in treatment-naive patients with metastatic melanoma, a significant expansion of CD27-CCR7-CD45ROhigh CD8+ T cells was observed during the third week of treatment with pembrolizumab or nivolumab. An amplification ratio > 0.8% in these cells indicated a favorable prognosis with a predictive accuracy of 90% (16). These findings highlight the significant role of Tem cells in response to PD-(L)1 immunotherapy.
Prolonged stimulation with antigens induces the progressive differentiation of Tem cells into terminally differentiated effector memory (Terma) cells, distinguished primarily by the re-emergence of CD45RA (CD45RA+CCR7-CD28-CD27-) and robust effector functionality, limited proliferative potential, and a profusion of senescence-related phenotypic traits (38). Terma cells are present in the peripheral blood of patients with different kinds of tumor (18, 19), and their presence is strongly correlated with the outcomes of PD-(L)1 immunotherapy (17, 18).
A study on patients with NSCLC receiving nivolumab reported increased numbers of CD45RA+CCR7-CD8+ T cells in patients who achieved a partial response (PR) at baseline and after the second or third dose (18). CD57 is a crucial marker of Terma cells (38). A study on patients with metastatic uroepithelial cancer (mUC) receiving PD-(L)1 (atezolizumab) therapy reported a remarkable prevalence of CD57 within circulating neoantigen-specific CD8+ T cells, particularly in individuals who responded to atezolizumab treatment, and responders to atezolizumab exhibited an increased proportion of Terma cells in the peripheral blood (19).
In contrast, some studies have suggested that circulating Terma cells are associated with an unfavorable prognosis in patients receiving PD-(L)1 immunotherapy (17, 20). A study examining the influence of immune senescence on anti-PD-(L)1 therapy in patients with advanced NSCLC found that CD28-CD57+KLRG1+CD8+ T cells (Terma-like CD8+ T cells) were associated with a lack of benefit from PD-(L)1 immunotherapy (20).
The ambiguity surrounding the predictive value of circulating Terma cells in immunotherapy may be attributed to variable tumor types and their clinical stages. Therefore, broadening the scope of the tumor species and increasing the sample size for further investigation on Terma cells are necessary. Terma cells are potential circulating markers for predicting the effectiveness of immunotherapy; however, further investigation is needed.
Trm cells persist within tissues and offer rapid and effective protective immunity against pathogens and metastatic cancer cells (39). Trm cells represent a distinct subpopulation of memory cells characterized by CD103+CD69+CD49a+CD8+ expression and unique transcriptome features, including RUNX family transcription factor 3, neurogenic locus notch homolog protein, B lymphocyte-induced maturation protein-1, and basic leucine zipper ATF-like transcription factor (40). Trm cells are mainly localized within tissues; however, they can also be detected in peripheral blood (35) and play a crucial role in the early response to anti-PD-(L)1 therapy[6. In patients with oral cancer receiving neoadjuvant treatment with either PD-1 monotherapy or PD-1 therapy combined with cytotoxic T-lymphocyte associated protein 4 (CTLA-4) inhibitor, single-cell and T-cell receptor (TCR) sequencing revealed that both peripheral blood and tumor-infiltrating CD8+ T cells exhibited activation and amplification, featuring highly overlapping clone types (6). Notably, the proportion of peripheral blood KLRG1-PD-1+CD8+ T cells was positively associated with pathological responses in both pre-and-on-treatment groups. The findings of the study highlight the pivotal role of neoadjuvant immunotherapy in the treatment of early-stage tumors involving Trm cells. In another study involving patients with gastric cancer who underwent surgery, individuals with a higher proportion of CD103 in peripheral blood PD-1+CD8+ T cells at week 2 of immunotherapy exhibited significantly improved PFS (21).
Tem, Terma, and Trm cells aid in distinguishing patients who are more likely to benefit from PD-(L)1 immunotherapy and predict their clinical prognosis. However, owing to contradictory reported data, the significance of Tem and Terma cells as predictors needs to be interpreted with caution. Additional studies are necessary to elucidate the specific markers of circulating memory cells associated with the prognosis of patients receiving PD-(L)1 immunotherapy.
T cell exhaustion is the specific differentiation state of T cells induced by persistent antigens and inflammatory signals, resulting in reduced effector function, diminished proliferative capacity, altered expression of multiple inhibitory receptors, and dysregulation of transcriptional mechanisms (41). Exhausted CD8+ T (Tex) cells exhibit high heterogeneity, which makes it challenging to define Tex cells using limited markers and molecular patterns. In the present review, Tex cells were defined using any of the following criteria: (1) expression of at least one additional co-inhibitory receptor other than PD-1, (2) high PD-1 expression levels, and (3) expression of at least one exhaustion-associated transcriptional factor, such as thymocyte selection-associated high mobility group-box (TOX), Eomesodermin (EOMES), nuclear receptor subfamily 4A, and nuclear factor of activated T cells 1. Using this category standard, we summarized the presence of heterogeneous Tex cells in the peripheral blood of patients with cancer and their predictive value in guiding the stratification of patients who may benefit from anti-PD-(L)1 therapy (22–25, 42).
Numerous studies have reported the baseline levels of Tex cell populations to predict clinical outcomes before treatment (22, 23, 42). Lymphocyte-activation gene 3 (LAG-3) is a surface inhibitory molecule highly expressed in Tex cells. In patients with gastric cancer receiving immunotherapy, a significant positive correlation was observed between LAG3+CD8+ T cells and PFS at baseline and after the initial dose (22). Another study identified a subset of LAG3+ T cells as an exhausted tumor-specific subpopulation that could be rejuvenated by the PD-1/PD-L1 blockade and was associated with an improved prognosis (22). Conversely, another study suggested that circulating LAG-3+ CD8+ T cells are predictive markers for identifying patients who are unlikely to benefit from PD-1 therapy. Analysis of pre-treatment blood samples from 188 patients with melanoma undergoing PD-1 therapy revealed a median survival discrepancy of > 4 years between patients with a LAG+ immunophenotype and those without (22.2 months vs. 75.8 months). Furthermore, in a validation cohort of 94 patients with bladder cancer treated with PD-1 therapy, those with the LAG+ immunophenotype demonstrated a response rate of 0. The LAG+CD8+ T cell immunotype is an independent prognostic marker (23). The noticeable discrepancy between the results of these two studies might be attributed to the patient population or methodological differences.
Patients with melanoma resistant to anti-PD-(L)1 therapy exhibited a distinct subpopulation of CD8+ T cells characterized by high levels of oxidative phosphorylation (OXPHOS), CD38 and CD39 expressions, and markers of exhaustion such as TOX, PD-1, and C-X-C motif chemokine ligand 13. Single-cell transcriptome analysis revealed an overlap between CD8+ and intratumoral CD8+ T cells. The study indicated that OXPHOS+CD8+ T cells among pre-treatment peripheral blood CD8+ T cells correlate with immune checkpoint inhibitor resistance in patients with malignant melanoma (42).
Changes in Tex cells after treatment can predict clinical outcomes because the systemic response to anti-PD-(L)1 therapy is dynamic and complex (43). Notably, several studies have investigated the role of post-treatment peripheral blood Tex cells in predicting the clinical outcome of anti-PD-(L)1 therapy (24, 25, 42).
In patients with stage IV melanoma receiving PD-1 (pembrolizumab) therapy, the magnitude of reinvigoration of circulating Tex cells, determined in relation to the pre-treatment tumor burden, correlated with the clinical response (24). The reinvigoration of circulating Tex cells is characterized by the expression of CD45AloCD27hiEomeshiT-betlo and co-expressed suppressor molecules (PD1, CTLA-4, and CD244), which correspond to the Tex cells’ characteristics (24). Immunoglobulin, immunoreceptor tyrosine-based inhibitory motif domains of T cell immunoreceptors, and PD1 double-positive T cells (DPOS) are present in the peripheral blood of patients with cancer and can serve as markers for predicting response to anti-PD-(L)1 therapy. In three cohorts of patients with cancer undergoing PD-1 therapy, a higher proportion of DPOS T cells in the peripheral blood after 1 month of treatment correlated with improved clinical response and extended OS (25).
In summary, peripheral circulating Tex cells play a crucial role in the response to PD-(L)1 treatment and can potentially serve as biomarkers for predicting the efficacy of anti-PD-(L)1 therapy.
CD8+ Teff cells express chemokine receptors, including CCR5, C-X-C motif chemokine receptor 3, and C-X3-C motif chemokine receptor 1 (44, 45), enabling their migration and infiltration from peripheral blood into the tumor microenvironment, where they exert their cytotoxic effects (45). Previous studies have shown that tumor-reactive T cells, including neoantigen-specific T cells, are present in the peripheral blood of patients with cancer and are enriched within the population of PD1+CD8+ T cells (46–48). Peripheral effector PD1lowCD8+ T cells exhibit an “effector-like” phenotype (32). Current evidence has revealed the response of peripheral circulating CD8+ Teff cells in patients with tumors to anti-PD-(L)1 therapy and their value in predicting the prognosis of patients with cancer (15, 26, 28, 29, 31, 49).
The predictive ability of circulating Teff cells before anti-PD-(L)1 therapy has been investigated in various tumor types, including malignant melanoma, NSCLC, and gastric cancer (26, 28, 29). One study reported that among patients with advanced NSCLC receiving nivolumab treatment, treatment responders exhibited approximately two-fold higher baseline levels of PD1+CD8+ T cells in the peripheral blood than non-responders. Increased levels of circulating PD1+CD8+ T cells are associated with prolonged OS and PFS (26). In another clinical cohort study involving metastatic tumors of various origins, a higher abundance of baseline CD137+CD8+ T cells was observed in the peripheral blood of patients who responded to anti-PD-(L)1 therapy. Elevated levels of CD137+CD8+ T cells in peripheral blood are associated with improved PFS and OS in patients (27). The CD137 receptor (4-1BB, tumor necrosis factor receptor [TNFR] superfamily) belongs to the costimulatory TNFR family and is expressed on activated CD8+ T cells (50). Similar results have been observed in patients with hormone receptor-positive metastatic breast cancer treated with the cyclin-dependent kinase 4/6 inhibitor (palbociclib) and PD-1 therapy (pembrolizumab) (30).
In contrast, certain observations have been made for specific subpopulations of PD1+ Teff cells. In patients with melanoma, the proportion of PD-1+CD56+ T cells in the peripheral blood is inversely correlated with clinical benefit (28). Another study reported a correlation between an elevated number of CD73+PD1+CD8+ T cells in the peripheral blood and an unfavorable response to anti-PD-(L)1 therapy (29). Among patients with advanced melanoma receiving nivolumab treatment, those who experienced clinical benefits exhibited considerably lower baseline proportions of circulating CD73+PD1+CD8+ T cells than non-responding patients (29).
Studies have shown that Tex cells within the tumor are replaced by CD8+ Teff cells recruited from the peripheral blood. This suggests that peripheral circulating T cells respond to anti-PD-(L)1 therapy (5). Circulating Teff cells are early responders to anti-PD-(L)1 therapy, and changes in circulating Teff cells caused by PD-(L)1 treatment strongly correlate with the prognosis of patients with tumor (31, 49).
A clinical trial investigating the impact of anti-PD-(L)1 therapy on CD8+ T cell function utilized single-cell sequencing of peripheral blood samples obtained from patients with melanoma undergoing immunotherapy. The results revealed the upregulation of natural killer cell granule protein-7 in CD8+ Teff cells in responders, whereas its downregulation was observed in non-responders (49). CX3CR1 belongs to a class of chemokine receptors that are highly expressed on the surface of Teff cells (45). Studies have shown that an elevated proportion of CX3CR1+ subpopulations within circulating CD8+ T cells at an early stage following anti-PD-1 therapy correlates with a favorable response and improved survival in patients with NSCLC (31). Furthermore, another study revealed an increase in neoantigen-reactive T cells among responders to PD-L1 therapy (51). However, this potential was constrained by the limited number of pre-and post-treatment sample pairs. Furthermore, the identification of tumor antigen-specific CD8+ T cells remains limited, and their universality necessitates further investigation.
Reinvigoration of pre-existing tumor-infiltrating T cells by anti-PD-(L)1 therapy is insufficient to inhibit tumor growth. Maximizing anti-PD-(L)1 therapy-mediated tumor control requires newly recruited T cells from the circulation (5). In patients with chronic infection and cancer, the TCF1-expressing subset of CD8+ T cells is responsible for the anti-PD-(L)1 therapy-driven T cell proliferation burst, which depends on CD28 signaling (52–54). Notably, most TCF1+CD8+ T cells in individuals with cancer reside in tumor-draining lymph nodes (55, 56). Under anti-PD-(L)1 therapy, TCF1+CD8+ T cells tend to proliferate (56). Subsequently, proliferating CD8+ T cells are released into the bloodstream, as evidenced by the increased proportion of Ki67+ CD8+ T cells in the blood after PD-1 therapy (53). The expansion of T cells in the peripheral blood of patients with cancer consistently predicts a better clinical response to anti-PD-L1 therapy (57). Numerous studies have consistently reported that Ki-67 expression in peripheral blood CD8+ T cell subsets exhibits a transient increase solely during the initial cycle after immunotherapy (32, 33). Therefore, the independent predictive and prognostic value of Ki-67-expressing CD8+ T cells as biomarkers of anti-PD-(L)1 therapy remains debatable.
A longitudinal analysis of blood samples from patients with advanced NSCLC undergoing PD-1 therapy revealed that approximately 70% of the patients exhibited an increased proportion of Ki67+PD-1+CD8+ T cells after the initial or second treatment cycle (32). This suggests that PD-1 therapy stimulates the proliferation of peripheral circulating PD1+CD8+ T cells. Among patients who experienced clinical benefits, 80% demonstrated a PD-1+CD8+ T-cell response within 4 weeks of treatment. PD-1 therapy-driven T cell proliferation is not uniform across different T cell subsets, as researchers found that Epstein–Barr virus-specific CD8+ T cells exhibit a diminished response to PD-1 therapy, indicating that the responsive cells might be specific to the tumor (32). Proliferating CD8+ T cells exhibit the characteristics of an effector-like phenotype (Human leukocyte antigen-DR+, CD38+, and B-cell lymphoma 2lo) (32). Comparable outcomes have been observed in patients with mUC treated with PD-L1 therapy. One study reported that within the identified neoantigen-reactive CD8+ T cells (NART), patients with disease control exhibited a Ki67+PD1+ effector phenotype in peripheral NART, indicating that the early amplification and activation of effector NART in patients with mUC is associated with a positive response to anti-PD-(L)1 therapy (58). Another study reported that an early proliferative response of PD1+CD8+ T cells was correlated with improved clinical outcomes. Among patients with thymic epithelial tumors and NSCLC undergoing PD-1 therapy, peripheral blood PD-1+CD8+ T cells (Ki-67D7/D0) exhibited a proliferative response within the initial week of treatment, enabling the differentiation between response to therapy and disease progression. This finding was confirmed in patients with NSCLC receiving PD-1 therapy. Tumor-specific CD8+ T cells exhibited a significant increase in Ki-67 expression on day 7, whereas virus-specific CD8+ T cells did not, indicating the specificity of PD-1 therapy for expanding tumor-reactive CD8+ T cells (33).
In summary, anti-PD-(L)1 therapy effectively restored the expansion of peripheral circulating tumor-reactive CD8+ T cells. The proliferation of peripheral circulating tumor-reactive CD8+ T cells has been associated with improved clinical outcomes and prolonged survival. However, additional studies are required to establish the optimal sampling time due to the highly dynamic expression of Ki-67 in circulating CD8+ T cells.
Similar to CD8+ T cells, CD4+ T cells, including naïve, Tcm, Tem, and Terma CD4+ T cell subsets and a unique group of regulatory CD4+ T cells (Tregs), characterized by CD25 and Forkhead box protein P3 expressions, are abundantly present in the peripheral blood. It is generally accepted that CD8+ T cells play a critical role in anti-PD-(L)1 therapy, whereas the importance of CD4+ T cells is underappreciated. However, few studies have revealed the significance of circulating CD4+ T cell subsets in predicting the efficacy of anti-PD-(L)1 therapy. Herein, we summarized the predictive value of CD4+ T cell subsets in patients with cancer treated with PD-(L)1 (Table 2, Figure 1).
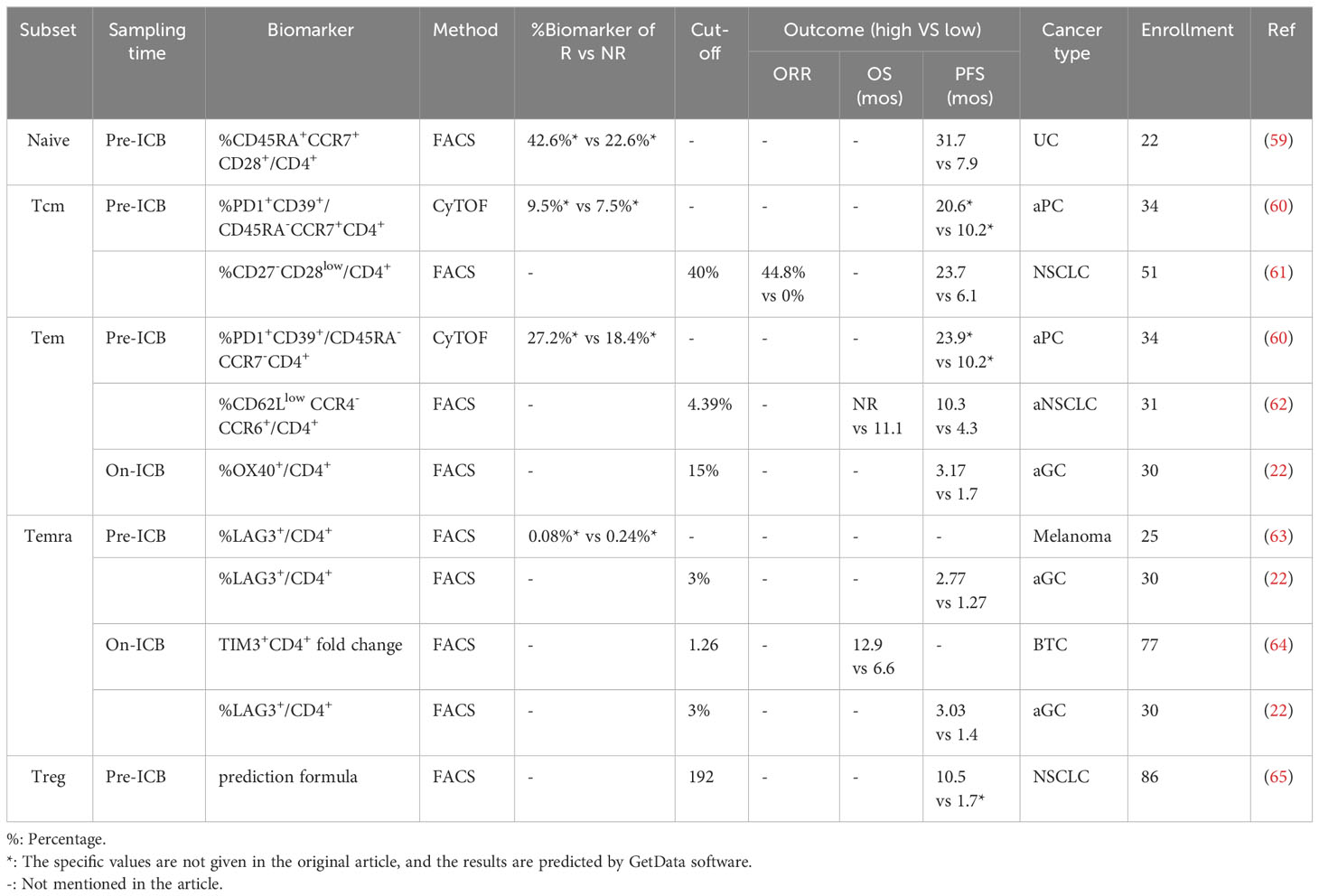
Table 2 Predictive outcome of peripheral CD4+ T cell subsets in PD-(L)1 therapy.
Naïve CD4+ T cells are characterized by CD45RA+ and CCR7+ expressions, whereas CD4+ Tcm cell subsets are characterized by CD45RA- and CCR7+ expressions. CD4+ Tcm cell subsets are T cells with long-term persistence. In a study involving 26 patients with bladder cancer who received PD-(L)1 (durvalumab) therapy, a group of naïve (CD45RAhi/int CCR7+CD28+) CD4+ T cells from pre-treatment (week 1) time points were enriched in the peripheral blood of responders but not in that of progressors (59). In addition, the pre-treatment proportions of circulating CD4+ Tcm cells were associated with prolonged survival after treatment with nivolumab combined with chemotherapy in patients with metastatic pancreatic cancer (60).
OX40 is a vital co-stimulator molecule (66). The proportion of baseline OX40-expressing circulating CD4+ T cells has been studied as a predictive marker of response to PD-1 therapy in patients with advanced solid tumors. CD4+PD1+OX40+ and CD4+α4β7+ cells among total CD4+ T cells have been reported to be present in higher proportions in patients with DCB, PR, or SD ≥ 6 months than in those without DCB (67). In addition to baseline level, circulating OX40+ CD4+ T cell/CD4+ T cell ≥ 15% was associated with better PFS in patients with advanced gastric cancer after treatment with nivolumab (22). In two independent phase I clinical trials, 19 patients with hormone receptor-positive metastatic breast cancer were treated with pembrolizumab. A study reported that the baseline proportion of circulating KLRG1+inducible T cell costimulator +CD4+ T cells was significantly higher in responders than in non-responders (22.6% vs. 8.7%) (30). Another study reported that the proportion of pre-treatment PD-1+CD39+CD4+ Tem cells was associated with > 1-year survival (60). Horimoto et al. recently suggested that the peripheral blood of patients with NSCLC receiving PD-1 therapy contains abundant CCR4-CCR6+ CD4+ T cells (Th7R cells). Patients with a higher proportion of baseline Th7R cells among CD4+ T cells (> 4.39%) showed significantly prolonged OS and PFS (62). In addition, a subgroup of CD27-CD28low cells, known as highly differentiated T cells, appears to be associated with the prognosis of anti-PD-(L)1 therapy (68).
CD4+ Terma cells (characterized by CD45RA+ and CCR7- expression) represent a group of terminally differentiated T cells with downregulation of costimulatory molecules and upregulation of inhibitory molecules. The proportions of CD38+CD39+CD127-GARP- CD4+ Terma cells were higher in the peripheral blood of patients with recurrence than in those without recurrence after treatment with nivolumab plus ipilimumab (69). In addition, an increased population of CD4+T cell immunoglobulin and mucin domain-containing protein 3 (TIM3+)T cells after dual mitogen-activated protein kinase kinase/PD-L1 inhibition correlated with worse OS (64). In contrast to TIM3, which predicts a poor prognosis, the relationship between LAG3+CD4+ T cells and prognosis is more variable. At baseline, a high proportion of LAG3-expressing CD4+ T cells indicates resistance to ipilimumab plus nivolumab treatment (63). In contrast, another study reported that patients with higher proportions of LAG3+CD4+ T cells at baseline (> 3%) and after the first administration(>3%) of nivolumab had longer PFS (22).
Current evidence indicates that regulatory T cells (Tregs) are essential for predicting the efficacy of anti-PD-(L)1 therapy. Notably, most studies have shown that Tregs are negatively correlated with prognosis, which is consistent with general beliefs. At baseline, the proportion of Tregs among CD4+ T cells was higher in patients with progressive disease (70). Furthermore, the expression of ki67 on Tregs is higher in patients with progressive disease (PD) (71). In addition, the percentage of circulating Tregs is significantly higher in patients resistant to nivolumab (65).
The predictive capability of immunotherapy has traditionally focused on classical CD8+ and CD4+ T cells; however, there is emerging evidence that some unique T cell subsets, including natural killer T cells (NKT), mucosa-associated invariant T cells (MAIT) (72), γδ T cells (73), CD8-CD4-T cells (double-negative T [DNT]) (74), and Tfh cells (10) can effectively identify individuals who will benefit from immunotherapy. Herein, we reviewed the current research on the role of these unique T cells in predicting immunotherapy outcomes and explored the need for further investigation into their significance in determining the predictive value of immunotherapy (Table 3, Figure 1).
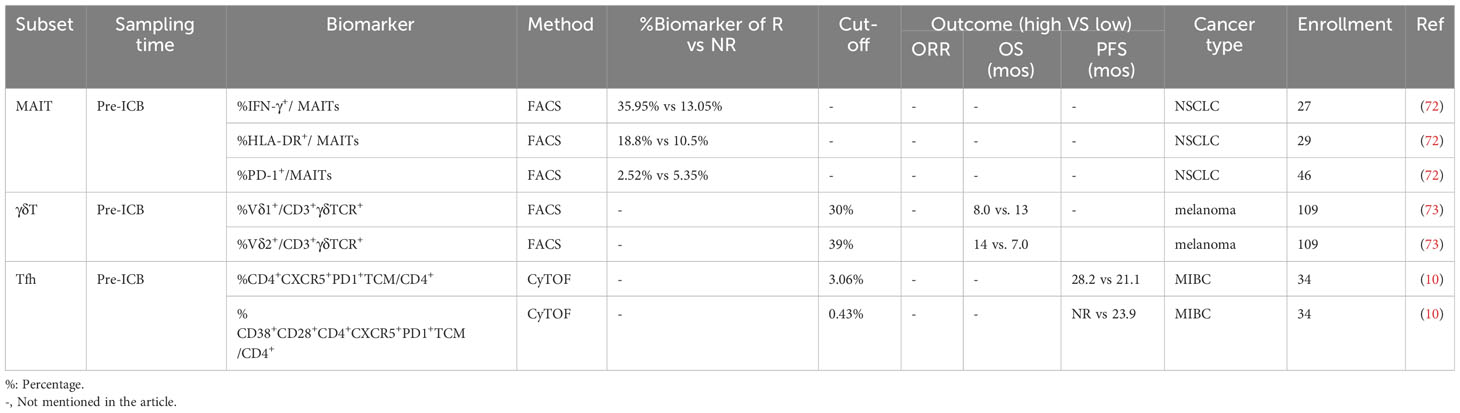
Table 3 Predictive outcome of peripheral unique T cell subsets in PD-(L)1 therapy.
MAIT cells, an atypical subset of T lymphocytes, are widely distributed throughout the human body and have been implicated in the pathogenesis of various human malignancies (75, 76). While predominantly located in mucosal tissues, MAIT cells also exist in the peripheral blood, lymphoid tissue, and organs including the liver (77). These cells express a conserved alpha chain (Vα7.2-Jα33) and a restricted range of beta chains (Vβ2 or Vβ13 in humans), facilitating the recognition of vitamin B metabolite antigens presented by the major histocompatibility complex (MHC)-I-related molecule (78). Among patients with NSCLC, those who responded to PD-1 therapy exhibited a higher proportion of circulating MAIT cells expressing the IFN-γ receptor (IFN-γR) in CD3+ T cells. Conversely, a lower proportion of MAIT cells expressed PD-1 (72). While these findings suggest that specific subsets of circulating MAIT cells expressing interferon (IFN)-γ receptor (MAIT-IFNGR cells) may serve as promising predictive markers for therapeutic response, MAIT cells expressing interleukin-17A (MAIT-IL17 cells) might indicate potential resistance to PD-1 therapy.
γδ T cells, a subset of unconventional T cells that operate independently of the conventional MHC restriction, constitute 1%–5% of the circulating T cell population and exhibit natural and adaptive immunity features (79). In patients with melanoma undergoing ipilimumab treatment, investigators observed that a decreased frequency of γδ T cells and an increased frequency of γδ T cells in the peripheral blood before treatment correlated with extended OS (73).
DNT are a unique subpopulation of CD3+ cells that express αβ or γδ T cell receptors but lack mature surface T cell markers, including CD4, CD8, and CD56 (80). These cells are thought to play a vital role in maintaining immune system homeostasis by eliminating self-reactive immune cells and modulating the allogeneic responses (81–83). In a prior study, it was observed that patients who responded to treatment exhibited reduced DNT and elevated NKT cell counts in their peripheral blood after undergoing treatment (74).
Tfh constitute a specialized subset of CD4+ Teff cells that play a vital role in germinal center responses, B-cell affinity maturation, and the orchestration of lymph node structure development (84, 85). In the context of Muscle-Invasive Bladder Cancer (MIBC), patients with a high proportion of circulating Tfh cells at baseline exhibited improved prognostic survival. Mechanistically, neoadjuvant therapy proves particularly beneficial for patients with MIBC who possess pre-existing circulating Tfh cells, as these cells play a pivotal role in orchestrating the development and maturation of tertiary lymphoid structures (TLS) like cells and in promoting the infiltration of CD8+ T cells into the tumor microenvironment (10).
Driven by chronic stimulation of tumor antigens, inflammation and hypoxic TME, tumor infiltrating T cells undergoes exhaustion, characterized by hierarchical loss of cytokine secretion and proliferation potential, while increase expression of various inhibitory receptors (86). Among the inhibitory receptors, PD-1 is the most attractive targets, due to its blockade refers to a significant success in cancer treatment (87). PD-1 was first cloned by Honjo et al. in 1992 (88). T cells are the major source of PD-1 in TME, while other types of cell also express it (89). PD-1 has two major ligands: PD-L1 and PD-L2 (87). PD-L1 is broadly expressed, while PD-L2 expression is restricted to hematopoietic cells, including dendritic cells, macrophages and B cells. Given the therapeutic efficacy of anti-PD-1 and anti-PD-L1 therapies are comparable in human, PD-L2 is probably of little importance in this context (87). Therefore, our review mainly focus on PD-1 and PD-L1. However, some recent studies suggest that PD-L2 are also promising therapeutic target (90, 91).
The advent of anti-PD-(L)1 therapy has greatly transformed the approach and prognosis of patients with cancer. Despite its notable impact, the response rate of anti-PD-(L)1 therapy remains low when applied in unselected patients with cancer. So far, the US FDA has approved three biomarkers that can predict the efficacy of anti-PD-1 therapy in several cancer types, including PD-L1 expression, dMMR/MSI-high, and TMB. These biomarkers are mostly based on tumor lesions sampled through invasive surgery or biopsy. Taking PD-L1 as an example, despite widespread use in clinic, PD-L1 expression as a biomarker presents limits due to technical and biological reasons, and its difficulty of assessment for pathologists (92, 93). In addition, the intra- and inter-tumor heterogeneity is another major issue for using PD-L1 as a biomarker (1, 94). Thus, the PD-L1 expression can be underestimated in small biopsies (such as bronchial and transthoracic biopsies), which are not representative of the entire tumor.
Circulating T cells has emerged as a promising biomarker that reflect the general immune landscape. Circulating T cells contains several T cell population, including memory (like) T cells, exhausted (like) T cells, effector T cells and proliferating T cells. Rencent studies suggest that anti-PD-(L)1 relies on the activation of memory (like) T cells or exhausted precursors T cells (52, 56), while activation of terminally exhausted T cells may promotes tumor progression by modulating cancer stem cells (95). Accordingly, existence of abundant number of memory (like) T cells in circulation was reported to be a favorable biomarker in patients receiving anti-PD-(L)1 therapy (14). Although numerous studies have explored the potential of circulating T cell subsets in predicting the efficacy of anti-PD-(L)1 therapy, a consensus is yet to be reached regarding the specific population and subsets of circulating T cell subsets. Consequently, prospective studies of large cohorts are urgently needed to validate the value of circulating T cell subsets in therapeutic decision-making. Lastly, it is important to note that circulating T cells are modifiable, which means a patient could been possiblely transformed toward an PD-(L)1-sensitive state before the initiation of therapy. For example, circulating bacteria-specific Tfh cells are correlated with PD-(L)1 efficacy (10). Accordingly, a recent study suggest that oral administration of bacteria or faecal microbiota transplantation were able to enhance efficacy of anti-PD-(L)1 threarpy in mice (61). Circulating T cells is a promising biomarker of PD-(L)1 therapy, as medicine becomes increasingly personalized, they may play a vital role in informing treatment decisions in the future.
JH: Writing – original draft. XY: Writing – original draft. SX: Writing – original draft. ZB: Conceptualization, Writing – review & editing. HZ: Conceptualization, Writing – original draft, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by the National Nature Science Foundation of China (No. 82272808 to HZ, No. 82230095 to BZ).
Thanks to Figdraw for the picture.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Jia Q, Wu W, Wang Y, Alexander PB, Sun C, Gong Z, et al. Local mutational diversity drives intratumoral immune heterogeneity in non-small cell lung cancer. Nat Commun. (2018) 9:5361. doi: 10.1038/s41467-018-07767-w
2. Reck M, Rodriguez-Abreu D, Robinson AG, Hui R, Csoszi T, Fulop A, et al. Pembrolizumab versus Chemotherapy for PD-L1-Positive Non-Small-Cell Lung Cancer. New Engl J Med. (2016) 375:1823–33. doi: 10.1056/NEJMoa1606774
3. Song R, Liu F, Ping Y, Zhang Y, Wang L. Potential non-invasive biomarkers in tumor immune checkpoint inhibitor therapy: response and prognosis prediction. biomark Res. (2023) 11:57. doi: 10.1186/s40364-023-00498-1
4. Hofman P, Heeke S, Alix-Panabières C, Pantel K. Liquid biopsy in the era of immuno-oncology: is it ready for prime-time use for cancer patients? Ann Oncol Off J Eur Soc Med Oncol. (2019) 30:1448–59. doi: 10.1093/annonc/mdz196
5. Yost KE, Satpathy AT, Wells DK, Qi Y, Wang C, Kageyama R, et al. Clonal replacement of tumor-specific T cells following PD-1 blockade. Nat Med. (2019) 25:1251–9. doi: 10.1038/s41591-019-0522-3
6. Luoma AM, Suo S, Wang Y, Gunasti L, Porter CBM, Nabilsi N, et al. Tissue-resident memory and circulating T cells are early responders to pre-surgical cancer immunotherapy. Cell. (2022) 185:2918–2935.e29. doi: 10.1016/j.cell.2022.06.018
7. Wang Z, Ahmed S, Labib M, Wang H, Wu L, Bavaghar-Zaeimi F, et al. Isolation of tumour-reactive lymphocytes from peripheral blood via microfluidic immunomagnetic cell sorting. Nat BioMed Eng. (2023) 7(9):1188–203. doi: 10.1038/s41551-023-01023-3
8. Li Z, Tuong ZK, Dean I, Willis C, Gaspal F, Fiancette R, et al. In vivo labeling reveals continuous trafficking of TCF-1+ T cells between tumor and lymphoid tissue. J Exp Med. (2022) 219:e20210749. doi: 10.1084/jem.20210749
9. Roulleaux Dugage M, Albarrán-Artahona V, Laguna JC, Chaput N, Vignot S, Besse B, et al. Biomarkers of response to immunotherapy in early stage non-small cell lung cancer. Eur J Cancer (Oxford Engl 1990). (2023) 184:179–96. doi: 10.1016/j.ejca.2023.01.029
10. Goubet AG, Lordello L, Alves Costa Silva C, Peguillet I, Gazzano M, Mbogning-Fonkou MD, et al. Escherichia coli-Specific CXCL13-Producing TFH Are Associated with Clinical Efficacy of Neoadjuvant PD-1 Blockade against Muscle-Invasive Bladder Cancer. Cancer Discovery. (2022) 12:2280–307. doi: 10.1158/2159-8290.CD-22-0201
11. Duchemann B, Naigeon M, Auclin E, Ferrara R, Cassard L, Jouniaux JM, et al. CD8(+)PD-1(+) to CD4(+)PD-1(+) ratio (PERLS) is associated with prognosis of patients with advanced NSCLC treated with PD-(L)1 blockers. J Immunother Cancer. (2022) 10:e004012. doi: 10.1136/jitc-2021-004012
12. Chalfin HJ, Pramparo T, Mortazavi A, Niglio SA, Schonhoft JD, Jendrisak A, et al. Circulating Tumor Cell Subtypes and T-cell Populations as Prognostic Biomarkers to Combination Immunotherapy in Patients with Metastatic Genitourinary Cancer. Clin Cancer Res. (2021) 27:1391–8. doi: 10.1158/1078-0432.CCR-20-2891
13. Martens A, Wistuba-Hamprecht K, Yuan J, Postow MA, Wong P, Capone M, et al. Increases in Absolute Lymphocytes and Circulating CD4+ and CD8+ T Cells Are Associated with Positive Clinical Outcome of Melanoma Patients Treated with Ipilimumab. Clin Cancer Res. (2016) 22:4848–58. doi: 10.1158/1078-0432.CCR-16-0249
14. Maniar R, Wang PH, Washburn RS, Kratchmarov R, Coley SM, Saqi A, et al. Self-Renewing CD8+ T-cell Abundance in Blood Associates with Response to Immunotherapy. Cancer Immunol Res. (2023) 11:164–70. doi: 10.1158/2326-6066.CIR-22-0524
15. Geng R, Tang H, You T, Xu X, Li S, Li Z, et al. Peripheral CD8+CD28+ T lymphocytes predict the efficacy and safety of PD-1/PD-L1 inhibitors in cancer patients. Front Immunol. (2023) 14:1125876. doi: 10.3389/fimmu.2023.1125876
16. Valpione S, Galvani E, Tweedy J, Mundra PA, Banyard A, Middlehurst P, et al. Immune-awakening revealed by peripheral T cell dynamics after one cycle of immunotherapy. Nat Cancer. (2020) 1:210–21. doi: 10.1038/s43018-019-0022-x
17. Fairfax BP, Taylor CA, Watson RA, Nassiri I, Danielli S, Fang H, et al. Peripheral CD8+ T cell characteristics associated with durable responses to immune checkpoint blockade in patients with metastatic melanoma. Nat Med. (2020) 26:193–9. doi: 10.1038/s41591-019-0734-6
18. Kunert A, Basak EA, Hurkmans DP, Balcioglu HE, Klaver Y, van Brakel M, et al. CD45RA+CCR7– CD8 T cells lacking co-stimulatory receptors demonstrate enhanced frequency in peripheral blood of NSCLC patients responding to nivolumab. J ImmunoTherapy Cancer. (2019) 7:149. doi: 10.1186/s40425-019-0608-y
19. Fehlings M, Kim L, Guan X, Yuen K, Tafazzol A, Sanjabi S, et al. Single-cell analysis reveals clonally expanded tumor-associated CD57(+) CD8 T cells are enriched in the periphery of patients with metastatic urothelial cancer responding to PD-L1 blockade. J immunotherapy Cancer. (2022) 10:e004759. doi: 10.1136/jitc-2022-004759
20. Ferrara R, Naigeon M, Auclin E, Duchemann B, Cassard L, Jouniaux JM, et al. Circulating T-cell Immunosenescence in Patients with Advanced Non-small Cell Lung Cancer Treated with Single-agent PD-1/PD-L1 Inhibitors or Platinum-based Chemotherapy. Clin Cancer Res an Off J Am Assoc Cancer Res. (2021) 27:492–503. doi: 10.1158/1078-0432.CCR-20-1420
21. Nose Y, Saito T, Yamamoto K, Yamashita K, Tanaka K, Yamamoto K, et al. The tissue-resident marker CD103 on peripheral blood T cells predicts responses to anti-PD-1 therapy in gastric cancer. Cancer Immunol Immunother. (2023) 72:169–81. doi: 10.1007/s00262-022-03240-2
22. Ohmura H, Yamaguchi K, Hanamura F, Ito M, Makiyama A, Uchino K, et al. OX40 and LAG3 are associated with better prognosis in advanced gastric cancer patients treated with anti-programmed death-1 antibody. Br J Cancer. (2020) 122:1507–17. doi: 10.1038/s41416-020-0810-1
23. Shen R, Postow MA, Adamow M, Arora A, Hannum M, Maher C, et al. LAG-3 expression on peripheral blood cells identifies patients with poorer outcomes after immune checkpoint blockade. Sci Trans Med. (2021) 13:eabf5107. doi: 10.1126/scitranslmed.abf5107
24. Huang AC, Postow MA, Orlowski RJ, Mick R, Bengsch B, Manne S, et al. T-cell invigoration to tumour burden ratio associated with anti-PD-1 response. Nature. (2017) 545:60–5. doi: 10.1038/nature22079
25. Simon S, Voillet V, Vignard V, Wu Z, Dabrowski C, Jouand N, et al. PD-1 and TIGIT coexpression identifies a circulating CD8 T cell subset predictive of response to anti-PD-1 therapy. J immunotherapy Cancer. (2020) 8:e001631. doi: 10.1136/jitc-2020-001631
26. Mazzaschi G, Facchinetti F, Missale G, Canetti D, Madeddu D, Zecca A, et al. The circulating pool of functionally competent NK and CD8+ cells predicts the outcome of anti-PD1 treatment in advanced NSCLC. Lung Cancer. (2019) 127:153–63. doi: 10.1016/j.lungcan.2018.11.038
27. Zizzari IG, Di Filippo A, Botticelli A, Strigari L, Pernazza A, Rullo E, et al. Circulating CD137+ T Cells Correlate with Improved Response to Anti-PD1 Immunotherapy in Patients with Cancer. Clin Cancer Res. (2022) 28:1027–37. doi: 10.1158/1078-0432.CCR-21-2918
28. Bochem J, Zelba H, Amaral T, Spreuer J, Soffel D, Eigentler T, et al. Peripheral PD-1+CD56+ T-cell frequencies correlate with outcome in stage IV melanoma under PD-1 blockade. PloS One. (2019) 14:e0221301. doi: 10.1371/journal.pone.0221301
29. Capone M, Fratangelo F, Giannarelli D, Sorrentino C, Turiello R, Zanotta S, et al. Frequency of circulating CD8+CD73+T cells is associated with survival in nivolumab-treated melanoma patients. J Transl Med. (2020) 18:121. doi: 10.1186/s12967-020-02285-0
30. Egelston C, Guo W, Yost S, Lee JS, Rose D, Avalos C, et al. Pre-existing effector T-cell levels and augmented myeloid cell composition denote response to CDK4/6 inhibitor palbociclib and pembrolizumab in hormone receptor-positive metastatic breast cancer. J Immunother Cancer. (2021) 9:e002084. doi: 10.1136/jitc-2020-002084
31. Yamauchi T, Hoki T, Oba T, Jain V, Chen H, Attwood K, et al. T-cell CX3CR1 expression as a dynamic blood-based biomarker of response to immune checkpoint inhibitors. Nat Commun. (2021) 12:1402. doi: 10.1038/s41467-021-21619-0
32. Kamphorst AO, Pillai RN, Yang S, Nasti TH, Akondy RS, Wieland A, et al. Proliferation of PD-1+ CD8 T cells in peripheral blood after PD-1-targeted therapy in lung cancer patients. Proc Natl Acad Sci U.S.A. (2017) 114:4993–8. doi: 10.1073/pnas.1705327114
33. Kim KH, Cho J, Ku BM, Koh J, Sun JM, Lee SH, et al. The First-week Proliferative Response of Peripheral Blood PD-1(+)CD8(+) T Cells Predicts the Response to Anti-PD-1 Therapy in Solid Tumors. Clin Cancer Res. (2019) 25:2144–54. doi: 10.1158/1078-0432.CCR-18-1449
34. Reading JL, Gálvez-Cancino F, Swanton C, Lladser A, Peggs KS, Quezada SA. The function and dysfunction of memory CD8(+) T cells in tumor immunit. Immunol Rev. (2018) 283:194–212. doi: 10.1111/imr.12657
35. Sievers C, Craveiro M, Friedman J, Robbins Y, Yang X, Bai K, et al. Phenotypic plasticity and reduced tissue retention of exhausted tumor-infiltrating T cells following neoadjuvant immunotherapy in head and neck cancer. Cancer Cell. (2023) 41:887–902.e5. doi: 10.1016/j.ccell.2023.03.014
36. Han J, Zhao Y, Shirai K, Molodtsov A, Kolling FW, Fisher JL, et al. Resident and circulating memory T cells persist for years in melanoma patients with durable responses to immunotherapy. Nat Cancer. (2021) 2:300–11. doi: 10.1038/s43018-021-00180-1
37. Siddiqui I, Schaeuble K, Chennupati V, Fuertes Marraco SA, Calderon-Copete S, Pais Ferreira D, et al. Intratumoral Tcf1(+)PD-1(+)CD8(+) T Cells with Stem-like Properties Promote Tumor Control in Response to Vaccination and Checkpoint Blockade Immunotherapy. Immunity. (2019) 50:195–211.e10. doi: 10.1016/j.immuni.2018.12.021
38. Henson SM, Riddell NE, Akbar AN. Properties of end-stage human T cells defined by CD45RA re-expression. Curr Opin Immunol. (2012) 24:476–81. doi: 10.1016/j.coi.2012.04.001
39. Molodtsov AK, Khatwani N, Vella JL, Lewis KA, Zhao Y, Han J, et al. Resident memory CD8(+) T cells in regional lymph nodes mediate immunity to metastatic melanoma. Immunity. (2021) 54:2117–2132.e7. doi: 10.1016/j.immuni.2021.08.019
40. Mami-Chouaib F, Blanc C, Corgnac S, Hans S, Malenica I, Granier C, et al. Resident memory T cells, critical components in tumor immunology. J Immunother Cancer. (2018) 6:87. doi: 10.1186/s40425-018-0399-6
41. Wherry EJ, Kurachi M. Molecular and cellular insights into T cell exhaustion. Nat Rev Immunol. (2015) 15:486–99. doi: 10.1038/nri3862
42. Li C, Phoon YP, Karlinsey K, Tian YF, Thapaliya S, Thongkum A, et al. A high OXPHOS CD8 T cell subset is predictive of immunotherapy resistance in melanoma patients. J Exp Med. (2022) 219:e20202084. doi: 10.1084/jem.20202084
43. Chen DS, Mellman I. Oncology Meets Immunology: The Cancer-Immunity Cycle. Immunity. (2013) 39:1–10. doi: 10.1016/j.immuni.2013.07.012
44. Franciszkiewicz K, Boissonnas A, Boutet M, Combadiere C, Mami-Chouaib F. Role of chemokines and chemokine receptors in shaping the effector phase of the antitumor immune response. Cancer Res. (2012) 72:6325–32. doi: 10.1158/0008-5472.CAN-12-2027
45. Ohta M, Tanaka F, Yamaguchi H, Sadanaga N, Inoue H, Mori M. The high expression of Fractalkine results in a better prognosis for colorectal cancer patients. Int J Oncol. (2005) 26:41–7. doi: 10.3892/ijo
46. Pasetto A, Gros A, Robbins PF, Deniger DC, Prickett TD, Matus-Nicodemos R, et al. Tumor- and Neoantigen-Reactive T-cell Receptors Can Be Identified Based on Their Frequency in Fresh Tumor. Cancer Immunol Res. (2016) 4:734–43. doi: 10.1158/2326-6066.CIR-16-0001
47. Gros A, Parkhurst MR, Tran E, Pasetto A, Robbins PF, Ilyas S, et al. Prospective identification of neoantigen-specific lymphocytes in the peripheral blood of melanoma patients. Nat Med. (2016) 22:433–8. doi: 10.1038/nm.4051
48. Gros A, Tran E, Parkhurst MR, Ilyas S, Pasetto A, Groh EM, et al. Recognition of human gastrointestinal cancer neoantigens by circulating PD-1+ lymphocytes. J Clin Invest. (2019) 129:4992–5004. doi: 10.1172/JCI127967
49. Wen T, Barham W, Li Y, Zhang H, Gicobi JK, Hirdler JB, et al. NKG7 Is a T-cell-Intrinsic Therapeutic Target for Improving Antitumor Cytotoxicity and Cancer Immunotherapy. Cancer Immunol Res. (2022) 10:162–81. doi: 10.1158/2326-6066.CIR-21-0539
50. Ye Q, Song DG, Poussin M, Yamamoto T, Best A, Li C, et al. CD137 accurately identifies and enriches for naturally occurring tumor-reactive T cells in tumor. Clin Cancer Res. (2014) 20:44–55. doi: 10.1158/1078-0432.CCR-13-0945
51. Fehlings M, Jhunjhunwala S, Kowanetz M, O'Gorman WE, Hegde PS, Sumatoh H, et al. Late-differentiated effector neoantigen-specific CD8+ T cells are enriched in peripheral blood of non-small cell lung carcinoma patients responding to atezolizumab treatment. J immunotherapy Cancer. (2019) 7:249. doi: 10.1186/s40425-019-0695-9
52. Im SJ, Hashimoto M, Gerner MY, Lee J, Kissick HT, Burger MC, et al. Defining CD8+ T cells that provide the proliferative burst after PD-1 therapy. Nature. (2016) 537:417–21. doi: 10.1038/nature19330
53. Kamphorst AO, Wieland A, Nasti T, Yang S, Zhang R, Barber DL, et al. Rescue of exhausted CD8 T cells by PD-1-targeted therapies is CD28-dependent. Sci (New York N.Y.). (2017) 355:1423–7. doi: 10.1126/science.aaf0683
54. Miller BC, Sen DR, Al Abosy R, Bi K, Virkud YV, LaFleur MW, et al. Subsets of exhausted CD8(+) T cells differentially mediate tumor control and respond to checkpoint blockade. Nat Immunol. (2019) 20:326–36. doi: 10.1038/s41590-019-0312-6
55. Schenkel JM, Herbst RH, Canner D, Li A, Hillman M, Shanahan SL, et al. Conventional type I dendric cells maintain a reservoir of proliferative tumor-antigen specific TCF-1(+) CD8(+) T cells in tumor-draining lymph nodes. Immunity. (2021) 54(10):2338–53.e6. doi: 10.1016/j.immuni.2021.08.026
56. Huang Q, Wu X, Wang Z, Chen X, Wang L, Lu Y, et al. The primordial differentiation of tumor-specific memory CD8(+) T cells as bona fide responders to PD-1/PD-L1 blockade in draining lymph nodes. Cell. (2022) 185(22):4049–66.e25. doi: 10.1016/j.cell.2022.09.020
57. Wu TD, Madireddi S, de Almeida PE, Banchereau R, Chen YJ, Chitre AS, et al. Peripheral T cell expansion predicts tumour infiltration and clinical response. Nature. (2020) 579:274–8. doi: 10.1038/s41586-020-2056-8
58. Holm JS, Funt SA, Borch A, Munk KK, Bjerregaard AM, Reading JL, et al. Neoantigen-specific CD8 T cell responses in the peripheral blood following PD-L1 blockade might predict therapy outcome in metastatic urothelial carcinoma. Nat Commun. (2022) 13:1935. doi: 10.1038/s41467-022-29342-0
59. Joshi M, Tuanquin L, Zhu J, Walter V, Schell T, Kaag M, et al. Concurrent durvalumab and radiation therapy (DUART) followed by adjuvant durvalumab in patients with localized urothelial cancer of bladder: results from phase II study, BTCRC-GU15-023. J Immunother Cancer. (2023) 11:e006551. doi: 10.1136/jitc-2022-006551
60. Padron LJ, Maurer DM, O'Hara MH, O'Reilly EM, Wolff RA, Wainberg ZA, et al. Sotigalimab and/or nivolumab with chemotherapy in first-line metastatic pancreatic cancer: clinical and immunologic analyses from the randomized phase 2 PRINCE trial. Nat Med. (2022) 28:1167–77. doi: 10.1038/s41591-022-01829-9
61. Gopalakrishnan V, Spencer CN, Nezi L, Reuben A, Andrews MC, Karpinets TV, et al. Gut microbiome modulates response to anti-PD-1 immunotherapy in melanoma patients. Sci (New York N.Y.). (2018) 359:97–103. doi: 10.1126/science.aan4236
62. Kagamu H, Yamasaki S, Kitano S, Yamaguchi O, Mouri A, Shiono A, et al. Single-Cell Analysis Reveals a CD4+ T-cell Cluster That Correlates with PD-1 Blockade Efficacy. Cancer Res. (2022) 82:4641–53. doi: 10.1158/0008-5472.CAN-22-0112
63. Machiraju D, Wiecken M, Lang N, Hulsmeyer I, Roth J, Schank TE, et al. Soluble immune checkpoints and T-cell subsets in blood as biomarkers for resistance to immunotherapy in melanoma patients. Oncoimmunology. (2021) 10:1926762. doi: 10.1080/2162402X.2021.1926762
64. Ruggieri AN, Yarchoan M, Goyal S, Liu Y, Sharon E, Chen HX, et al. Combined MEK/PD-L1 Inhibition Alters Peripheral Cytokines and Lymphocyte Populations Correlating with Improved Clinical Outcomes in Advanced Biliary Tract Cancer. Clin Cancer Res. (2022) 28:4336–45. doi: 10.1158/1078-0432.CCR-22-1123
65. Kagamu H, Kitano S, Yamaguchi O, Yoshimura K, Horimoto K, Kitazawa M, et al. CD4(+) T-cell Immunity in the Peripheral Blood Correlates with Response to Anti-PD-1 Therapy. Cancer Immunol Res. (2020) 8:334–44. doi: 10.1158/2326-6066.CIR-19-0574
66. Gao J, Bernatchez C, Sharma P, Radvanyi LG, Hwu P. Advances in the development of cancer immunotherapies. Trends Immunol. (2013) 34:90–8. doi: 10.1016/j.it.2012.08.004
67. Baldini C, Danlos FX, Varga A, Texier M, Halse H, Mouraud S, et al. Safety, recommended dose, efficacy and immune correlates for nintedanib in combination with pembrolizumab in patients with advanced cancers. J Exp Clin Cancer Res. (2022) 41:217. doi: 10.1186/s13046-022-02423-0
68. Zuazo M, Arasanz H, Fernández-Hinojal G, García-Granda MJ, Gato M, Bocanegra A, et al. Functional systemic CD4 immunity is required for clinical responses to PD-L1/PD-1 blockade therapy. EMBO Mol Med. (2019) 11:e10293. doi: 10.15252/emmm.201910293
69. Khushalani NI, Vassallo M, Goldberg JD, Eroglu Z, Kim Y, Cao B, et al. Nivolumab plus chemoradiotherapy in locally-advanced cervical cancer: the NICOL phase 1 trial. J Immunother Cancer. (2022) 10:e005684. doi: 10.1136/jitc-2022-005684
70. Gibney GT, KudChadkar RR, DeConti RC, Thebeau MS, Czupryn MP, Tetteh L, et al. Safety, correlative markers, and clinical results of adjuvant nivolumab in combination with vaccine in resected high-risk metastatic melanoma. Clin Cancer Res. (2015) 21:712–20. doi: 10.1158/1078-0432.CCR-14-2468
71. Rodrigues M, Vanoni G, Loap P, Dubot C, Timperi E, Minsat M, et al. Nivolumab plus chemoradiotherapy in locally-advanced cervical cancer: the NICOL phase 1 trial. Nat Commun. (2023) 14:3698. doi: 10.1038/s41467-023-39383-8
72. Shi L, Lu J, Zhong D, Song M, Liu J, You W, et al. Clinicopathological and predictive value of MAIT cells in non-small cell lung cancer for immunotherapy. J immunotherapy Cancer. (2023) 11:e005902. doi: 10.1136/jitc-2022-005902
73. Wistuba-Hamprecht K, Martens A, Haehnel K, Geukes Foppen M, Yuan J, Postow MA, et al. Proportions of blood-borne Vdelta1+ and Vdelta2+ T-cells are associated with overall survival of melanoma patients treated with ipilimumab. Eur J Cancer. (2016) 64:116–26. doi: 10.1016/j.ejca.2016.06.001
74. Strippoli S, Fanizzi A, Negri A, Quaresmini D, Nardone A, Armenio A, et al. Examining the Relationship between Circulating CD4- CD8- Double-Negative T Cells and Outcomes of Immuno-Checkpoint Inhibitor Therapy-Looking for Biomarkers and Therapeutic Targets in Metastatic Melanoma. Cells. (2021) 10:406. doi: 10.3390/cells10020406
75. Yan J, Allen S, McDonald E, Das I, Mak JYW, Liu L, et al. MAIT Cells Promote Tumor Initiation, Growth, and Metastases via Tumor MR1. Cancer Discovery. (2020) 10:124–41. doi: 10.1158/2159-8290.CD-19-0569
76. Loh L, Wang Z, Sant S, Koutsakos M, Jegaskanda S, Corbett AJ, et al. Human mucosal-associated invariant T cells contribute to antiviral influenza immunity via IL-18-dependent activation. Proc Natl Acad Sci U.S.A. (2016) 113:10133–8. doi: 10.1073/pnas.1610750113
77. Godfrey DI, Koay HF, McCluskey J, Gherardin NA. The biology and functional importance of MAIT cells. Nat Immunol. (2019) 20:1110–28. doi: 10.1038/s41590-019-0444-8
78. Gold MC, Cerri S, Smyk-Pearson S, Cansler ME, Vogt TM, Delepine J, et al. Human mucosal associated invariant T cells detect bacterially infected cells. PloS Biol. (2010) 8:e1000407. doi: 10.1371/journal.pbio.1000407
79. Godfrey DI, Le Nours J, Andrews DM, Uldrich AP, Rossjohn J. Unconventional T Cell Targets for Cancer Immunotherapy. Immunity. (2018) 48:453–73. doi: 10.1016/j.immuni.2018.03.009
80. Wu Z, Zheng Y, Sheng J, Han Y, Yang Y, Pan H, et al. CD3(+)CD4(-)CD8(-) (Double-Negative) T Cells in Inflammation, Immune Disorders and Cancer. Front Immunol. (2022) 13:816005. doi: 10.3389/fimmu.2022.816005
81. Fischer K, Voelkl S, Heymann J, Przybylski GK, Mondal K, Laumer M, et al. Isolation and characterization of human antigen-specific TCR alpha beta+ CD4(-)CD8- double-negative regulatory T cells. Blood. (2005) 105:2828–35. doi: 10.1182/blood-2004-07-2583
82. Priatel JJ, Utting O, Teh HS. TCR/self-antigen interactions drive double-negative T cell peripheral expansion and differentiation into suppressor cells. J Immunol. (2001) 167:6188–94. doi: 10.4049/jimmunol.167.11.6188
83. Zhang D, Yang W, Degauque N, Tian Y, Mikita A, Zheng XX. New differentiation pathway for double-negative regulatory T cells that regulates the magnitude of immune responses. Blood. (2007) 109:4071–9. doi: 10.1182/blood-2006-10-050625
84. Crotty S. Follicular helper CD4 T cells (TFH). Annu Rev Immunol. (2011) 29:621–63. doi: 10.1146/annurev-immunol-031210-101400
85. Chaurio RA, Anadon CM, Lee Costich T, Payne KK, Biswas S, Harro CM, et al. TGF-beta-mediated silencing of genomic organizer SATB1 promotes Tfh cell differentiation and formation of intra-tumoral tertiary lymphoid structures. Immunity. (2022) 55:115–128.e9. doi: 10.1016/j.immuni.2021.12.007
86. Zebley CC, Youngblood B. Mechanisms of T cell exhaustion guiding next-generation immunotherapy. Trends Cancer. (2022) 8:726–34. doi: 10.1016/j.trecan.2022.04.004
87. Chamoto K, Yaguchi T, Tajima M, Honjo T. Insights from a 30-year journey: function, regulation and therapeutic modulation of PD1. Nat Rev Immunol. (2023) 23:682–95. doi: 10.1038/s41577-023-00867-9
88. Ishida Y, Agata Y, Shibahara K, Honjo T. Induced expression of PD-1, a novel member of the immunoglobulin gene superfamily, upon programmed cell death. EMBO J. (1992) 11:3887–95. doi: 10.1002/embj.1992.11.issue-11
89. Zha H, Jiang Y, Wang X, Shang J, Wang N, Yu L, et al. Non-canonical PD-1 signaling in cancer and its potential implications in clinic. J immunotherapy Cancer. (2021) 9:e001230. doi: 10.1136/jitc-2020-001230
90. Park JS, Gazzaniga FS, Wu M, Luthens AK, Gillis J, Zheng W, et al. Targeting PD-L2-RGMb overcomes microbiome-related immunotherapy resistance. Nature. (2023) 617:377–85. doi: 10.1038/s41586-023-06026-3
91. Miao YR, Thakkar KN, Qian J, Kariolis MS, Huang W, Nandagopal S, et al. Neutralization of PD-L2 is Essential for Overcoming Immune Checkpoint Blockade Resistance in Ovarian Cancer. Clin Cancer Res an Off J Am Assoc Cancer Res. (2021) 27:4435–48. doi: 10.1158/1078-0432.CCR-20-0482
92. Hofman P. PD-L1 immunohistochemistry for non-small cell lung carcinoma: which strategy should be adopted? Expert Rev Mol Diagn. (2017) 17:1097–108. doi: 10.1080/14737159.2017.1398083
93. Tsao MS, Kerr KM, Kockx M, Beasley MB, Borczuk AC, Botling J, et al. PD-L1 Immunohistochemistry Comparability Study in Real-Life Clinical Samples: Results of Blueprint Phase 2 Project. J Thorac Oncol Off Publ Int Assoc Study Lung Cancer. (2018) 13:1302–11. doi: 10.1016/j.jtho.2018.05.013
94. Ilie M, Long-Mira E, Bence C, Butori C, Lassalle S, Bouhlel L, et al. Comparative study of the PD-L1 status between surgically resected specimens and matched biopsies of NSCLC patients reveal major discordances: a potential issue for anti-PD-L1 therapeutic strategies. Ann Oncol Off J Eur Soc Med Oncol. (2016) 27:147–53. doi: 10.1093/annonc/mdv489
Keywords: circulating T cells, anti-PD-(L)1 therapy, biomarker, CD8+ T lymphocyte subsets, CD4+ T cell
Citation: Hou J, Yang X, Xie S, Zhu B and Zha H (2024) Circulating T cells: a promising biomarker of anti-PD-(L)1 therapy. Front. Immunol. 15:1371559. doi: 10.3389/fimmu.2024.1371559
Received: 16 January 2024; Accepted: 07 March 2024;
Published: 21 March 2024.
Edited by:
Jehad Charo, Roche, SwitzerlandReviewed by:
Saptak Banerjee, Chittaranjan National Cancer Institute (CNCI), IndiaCopyright © 2024 Hou, Yang, Xie, Zhu and Zha. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Haoran Zha, emhhaGFvcmFuQGhvdG1haWwuY29t; Bo Zhu, Ym8uemh1QHRtbXUuZWR1LmNu
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.