
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Immunol., 20 February 2024
Sec. Inflammation
Volume 15 - 2024 | https://doi.org/10.3389/fimmu.2024.1355503
This article is part of the Research TopicNew insights into the role of cellular senescence in musculoskeletal system diseaseView all 4 articles
 Shuai Li1,2,3
Shuai Li1,2,3 Jinpeng Du1,3Yunfei Huang1,3Shenglong Gao1,2,3Zhigang Zhao1,3Zhen Chang1,3Xuefang Zhang1,3
Jinpeng Du1,3Yunfei Huang1,3Shenglong Gao1,2,3Zhigang Zhao1,3Zhen Chang1,3Xuefang Zhang1,3 BaoRong He1,3*
BaoRong He1,3*The incidence of lumbar disc herniation has gradually increased in recent years, and most patients have symptoms of low back pain and nerve compression, which brings a heavy burden to patients and society alike. Although the causes of disc herniation are complex, intervertebral disc degeneration (IDD) is considered to be the most common factor. The intervertebral disc (IVD) is composed of the upper and lower cartilage endplates, nucleus pulposus, and annulus fibrosus. Aging, abnormal mechanical stress load, and metabolic disorders can exacerbate the progression of IDD. Among them, high glucose and high-fat diets (HFD) can lead to fat accumulation, abnormal glucose metabolism, and inflammation, which are considered important factors affecting the homeostasis of IDD. Diabetes and advanced glycation end products (AGEs) accumulation- can lead to various adverse effects on the IVD, including cell senescence, apoptosis, pyroptosis, proliferation, and Extracellular matrix (ECM) degradation. While current research provides a fundamental basis for the treatment of high glucose-induced IDD patients. further exploration into the mechanisms of abnormal glucose metabolism affecting IDD and in the development of targeted drugs will provide the foundation for the effective treatment of these patients. We aimed to systematically review studies regarding the effects of hyperglycemia on the progress of IDD.
Low back pain (LBP) is a common public health concern worldwide (1, 2). Approximately 60-80% of patients experience chronic low back pain throughout their lifetime (3–5). Furthermore, LBP is the main cause of disability and productivity loss and seriously affects the quality of life of patients (6, 7). Although the cause of most cases of LBP is unclear, intervertebral disc degeneration (IDD) is considered to be the most common factor. The intervertebral disc (IVD) consists of the annulus fibrosus (AF), nucleus pulposus (NP) and cartilage endplate (CEP) (8–10). The NP is rich in proteoglycans and type II collagen and is highly hydrated. Therefore, physiological osmotic pressure can easily dissipate any mechanical force transmitted through the spine (11, 12). The AF is a layered structure mainly composed of type I collagen (13, 14), whereas the CEP is composed of transparent cartilage located between IVD soft tissue and the vertebral bone structures (15, 16). The CEP is crucial in the maintenance of mechanical integrity and nutrient exchange of the IVD. Furthermore, the IVD can increase the range of spinal movement, withstand pressure, cushion vibration and protect the spinal cord (17).
Degeneration of the IVD occurs naturally with age, weakening its elasticity and toughness (18, 19). Imbalance between anabolism and catabolism in the IVD can lead to changes in the composition of the extracellular matrix (ECM), cell loss, excessive oxidative stress and inflammation (20, 21). Additionally, abnormal spinal mechanical changes can easily cause a series of symptoms of intervertebral disc herniation. Age, repeated mechanical stress, occupational factors, metabolic disorders (such as obesity and diabetes), trauma, heredity, and even smoking may lead to the development of IDD (22–25). The possible factors affecting IDD are described in the Figure 1.
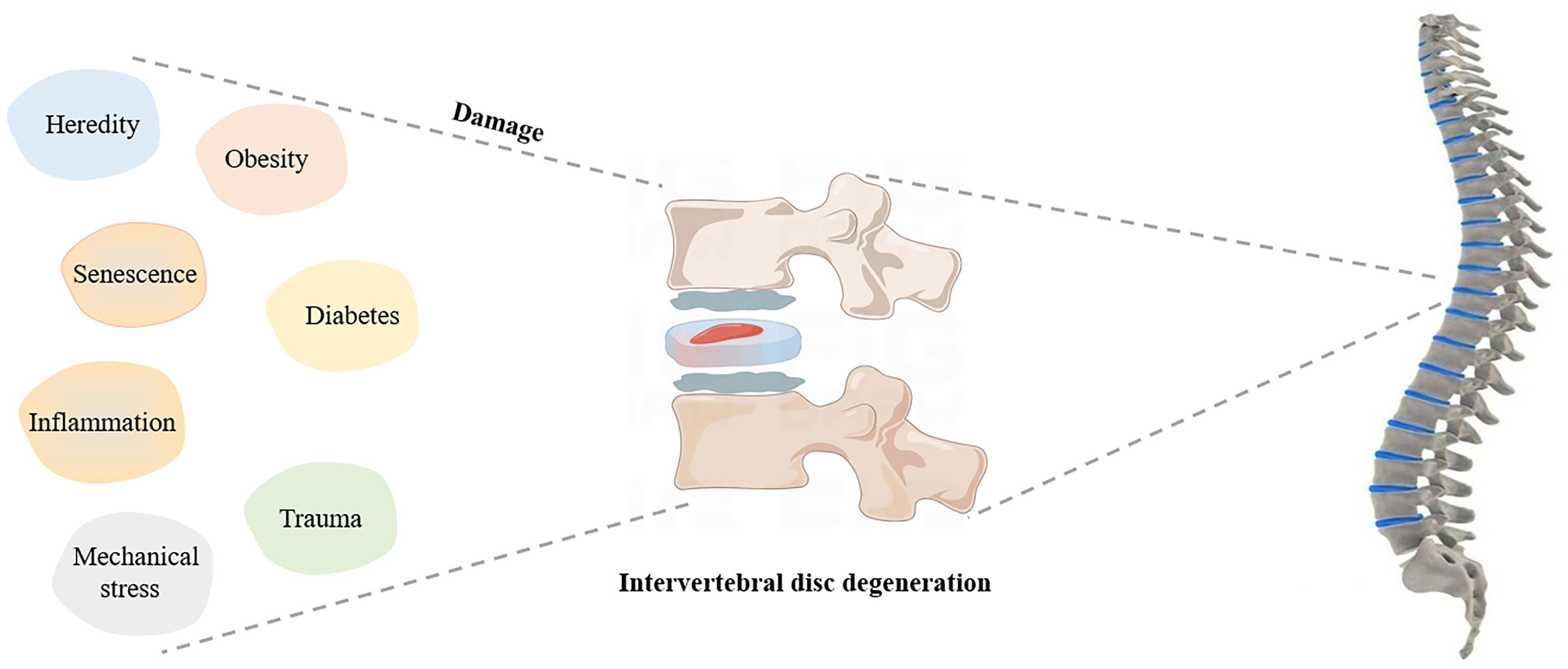
Figure 1 Possible factors affecting IDD.
The preferred treatment methods for lumbar and leg pain caused by IDD are lifestyle changes, encompassing weight management (reducing body weight as a means to alleviate spinal pressure), optimizing posture (acquiring and sustaining proper standing, sitting, and lifting postures), incorporating regular breaks (averting prolonged static positions and periodically altering postures), thermal therapy (alternating between cold and heat applications to mitigate pain and inflammation), refining sleep quality (utilizing appropriate mattresses and pillows, discovering sleep positions conducive to alleviating spinal pressure), spinal traction (alleviating pain and diminishing intervertebral disc pressure), acupuncture, and massage (inducing muscle relaxation and alleviating discomfort) (26, 27). Lifestyle changes have a significant effect. Physiotherapy also plays a vital role in relieving the symptoms of IDD. On the drug therapy aspect, opioids, steroids, and non-steroidal anti-inflammatory drugs play an important role in controlling pain and improving function and quality of life (28–30). While medications prove effective in managing pain and inflammation, prolonged use carries potential side effects, with opioids posing a risk of dependence. Emphasis must be placed on the crucial need to customize treatment plans based on individual patient needs and responses. Moreover, the sustained efficacy of these conservative measures varies within patient populations, underscoring the imperative for personalized therapeutic approaches that consider specific patient circumstances, comorbidities, and responses to initial treatments.
Surgical treatment includes discectomy, fusion, and total disc replacement, and can be considered as the gold standard of IDD treatment (31, 32). The decision for surgical intervention in cases of IDD is a complex and multifaceted process that requires a comprehensive evaluation, combining imaging studies such as CT, MRI, and bone density scans with an assessment of the patients overall physical condition. Typically, surgery becomes a pivotal consideration when conservative treatments, including medication and physical therapy, prove inadequate in providing substantial relief from symptoms. The presence of neurological symptoms such as numbness, muscle weakness, or difficulty in walking signifies compression of nerve roots or the spinal cord, serving as a critical determinant for opting for surgical decompression procedures. Additionally, the patients general health, encompassing coexisting conditions like diabetes, osteoporosis, or cardiovascular diseases, along with the patients willingness to undergo surgery, are pivotal factors influencing the decision for surgical intervention. Surgery can effectively relieve the symptoms of nerve compression and prevent the subsequent deterioration of muscle strength (33). With the development of three-dimensional (3D) printing technology and synthetic biomaterials, using artificial intervertebral discs to replace degenerative IVD has gradually become feasible (34–36). In summary, the decision to undergo surgical intervention should be guided by the substantial benefits observed in terms of pain alleviation, functional enhancement, and overall improvement in the quality of life during the evaluation of the patient. Surgical recommendation is warranted when there is a discernible and significant positive impact in these critical aspects.
IDD is a multi-factor process. Biomolecule therapy aims to target disease states characterized by decreased anabolism and increased catabolism (37–39). A variety of biotherapies have been evaluated in preclinical and clinical studies, including the use of growth factors and platelet-rich plasma (18, 40, 41). Biomolecule therapy focuses on repair due to the loss of proteoglycans and collagen, and its goal is to reduce the environment of pro-inflammatory catabolism and anti-anabolic metabolism in IDD.
Cell therapy is one of the best treatment strategies for patients with mid-term IDD, playing plays an active role by increasing the number of normal cells in the diseased IVD. Cell transplantation can secrete various cytokines, immune receptors, and anti-inflammatory molecules to regulate the microenvironment of host tissues (42, 43). Furthermore, extracellular vesicles (EVs), as acellular structures, can freely shuttle between cells and perform signaling and communication functions (44–46). EVs regulates apoptosis, senescence, proliferation and inflammation by delivering targeted non-coding RNA and various proteins to recipient cells (47, 48).
In addition to abnormal mechanical overload, abnormal fat accumulation, changes in glucose and lipid metabolism and inflammation are important factors affecting IVD homeostasis and pro-inflammatory pathophysiology (49–51). High concentrations of pro-inflammatory cytokines, adipokines, sugars, and lipids, aggravate systemic low-grade inflammation, which consequently impairs the metabolism of articular chondrocytes and IVD cells. Abnormal glucose metabolism is an important adverse factor in triggering and exacerbating IDD, and various biological processes and mechanisms of abnormal glucose metabolism affect cell senescence, apoptosis, inflammation, proliferation, and ECM degradation in IDD (Figure 2).
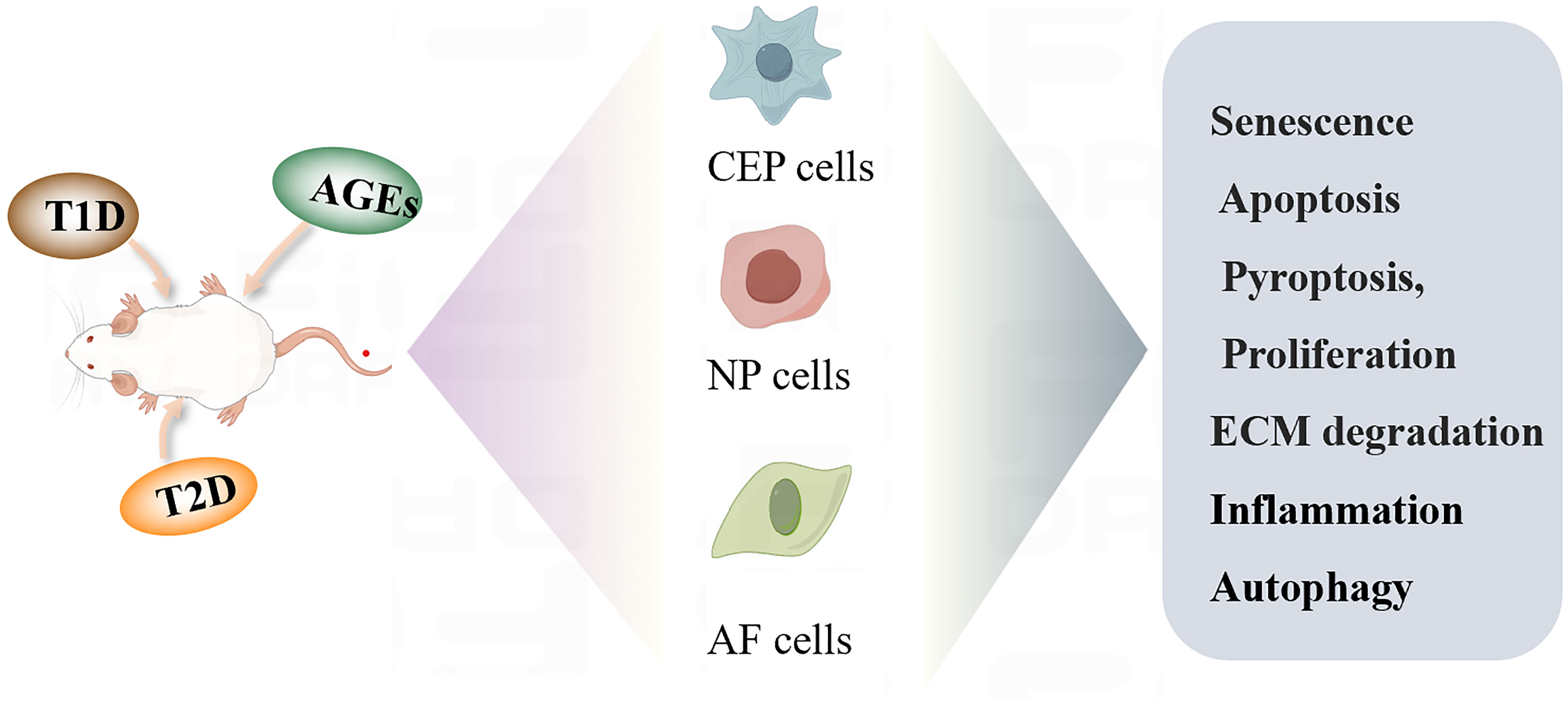
Figure 2 Diabetic IDD regulates the biological function of cells.
Type 1 diabetes (Non-obese diabetes, T1D) is an autoimmune disease characterized by destruction of pancreatic β cells, affecting more than 1.2 million children and adolescents worldwide. For these patients, daily insulin injections are necessary to maintain healthy blood sugar levels. Viral infection (enterovirus and coxsackievirus), streptozotocin, antibiotics and epigenetic modification are risk factors for the formation of T1D (52, 53). Ina clinical study, 118patients with T1D were retrospectively analyzed and evaluated by Pfirrmann score system and MRI. The results showed that IDD was serious in patients with poor blood glucose control. T1D can cause early degeneration of the IVD (54). At the cellular level, NP tissues from Streptozotocin (STZ) -induced T1D rats with IDD rats were collected for transcriptome sequencing and bioinformatics analysis. The results showed that BMP7 could be used as a core gene, and NLRP3 inflammasome and pyroptosis related markers were significantly increased in NP cells. Further experimental detection showed that BMP7 inhibits the activation of NLRP3 inflammasome and cell pyroptosis processes, thereby slowing down the IDD of T1D rats (55). In addition, in T1D mice, higher levels of ADAMTS were shown in the intervertebral disc matrix, which mediated glycan fragmentation and IVD apoptosis (56). These studies show that T1D is an important factor in promoting the progress of IDD, and that effective blood glucose control is the key strategy to delay IDD.
Type II diabetes (T2D) is a chronic metabolic disease characterized by long-term hyperglycemia and insulin resistance (IR), and the decrease of insulin absorption and glucose utilization leads to more compensatory insulin secretion, resulting in hyperinsulinemia (57, 58). Long-term hyperglycemia can lead to decreased organ function due to various health complications (59, 60). A clinical observation showed that the height of the IVD decreased significantly in T2D patients, which may lead to an increase in the risk of vertebral fracture in T2D patients (61). In addition, increased oxidative stress and AGE/RAGE-mediated interaction may be important factors leading to an increase in the incidence of T2D IVD disease (62). Contrastingly, a diabetic mouse model was established by feeding with a HFD, and it was found that HFD mice showed obvious IDD. A microarray analysis showed that the differential genes ADAMTS4 and ADAMTS5 were increased in IDD samples. Overexpression of ADAMTS4 and ADAMTS5 accelerated the degradation of the ECM and led to the occurrence of IDD. In mechanism, the complex formed by CBP-PGC-1 α-Runx2 binds to the initiation binding regions of ADAMTS4 and ADAMTS5 to promote gene expression (63).
Leptin receptor deficient knockout (db/db) mice are mature models for the study of T2D. Leptin receptor deficiency can lead to cortical and trabecular bone changes, decrease torsional destruction strength, and destruction of biomechanical behavior of IDD (64). Furthermore, knockdown of the leptin receptor can increase the level of MMP3 and induce apoptosis (65). Moreover, Vitamin K is also related to glucose metabolism and insulin sensitivity. In IDD induced by T2D, vitamin K2 diets may exert an anti-inflammatory function by regulating Socs3 and Hmox1 (66). In summary, the prevention and treatment of IDD induced by T2D cannot be ignored. The current research is still limited, and further exploration is needed to elucidate the mechanism of high glucose-induced IDD. Both Type 1 and Type 2 diabetes contribute to the exacerbation of IDD, albeit with slightly distinct mechanisms. Owing to an earlier onset and its autoimmune nature, T1D may precipitate an earlier onset of IDD with potentially accelerated progression. In contrast, IDD linked to T2D may manifest more covertly, exacerbated by the emergence of metabolic syndrome. For T1D, a concentrated effort on stringent blood glucose control and vigilant monitoring of early signs of IDD may prove beneficial. In the context of T2D, addressing the pervasive influence of metabolic syndrome and achieving optimal blood glucose control may be more efficacious in IDD management. In essence, although both diabetes types exert adverse effects on IDD, an in-depth comprehension of these mechanisms will facilitate the development of more targeted and efficacious treatment approaches for IDD within the diabetic milieu.
The continuous increase of blood glucose levels in patients with diabetes can lead to non-enzymatic macromolecular glycosylation and eventually to the formation of advanced glycation end products (AGEs) (67, 68). AGEs accumulation is the source of complications of diabetes. AGEs are mainly accumulated in proteins, including aggregating glycans and collagen (69, 70), causing the fibers to soften, rendering the dehydrated matrix unable to withstand the typical mechanical forces of the spine, resulting in biomechanical changes (71). Studies have determined the direct correlation between AGEs intake and the incidence of IDD. The NP samples of clinical IDD patients were detected, and it was found that compared with non-diabetic IDD patients, the level of AGEs in diabetic patients was significantly higher than that in non-diabetic controls (72). Transcriptome sequencing analysis showed that the expression of several MMP genes increased in IVD mice fed with HFD. AGEs treatment promotes the rise of the AP1-p300/CBP-PPRC pathway, and AGEs via PAK1/USP24/PPRC1-p300/CBP-AP1 signaling drives MMP to induce ECM degradation, which leads to IDD. Inhibition of this pathway by injection of inhibitors in vivo can effectively prevent the degeneration of the IVD (73). After ACG activation, Gal3 and RAGE can act as receptors of ACG to affect collagen, in which Gal3 has a protective effect on AGE attack, limiting collagen damage and biomechanical changes. RAGE is an essential receptor for collagen injury induced by AGEs (74). MRI image analysis showed that oral treatment with the AGE inhibitor PM could reduce NP dehydration after needle injury. PM also promoted the increase in the level of oligosaccharides in rats (75). In addition, AGEs-BSA can induce mitochondrial damage in NP cells through activation of NLRP3 inflammatory bodies, resulting in a secondary inflammatory response (76). High AGEs content diets increase vertebral cortical thickening and endplate calcification, decrease IVD height and GAG content, and increase COL-X expression, which promotes the hypertrophic differentiation of NP cells. This accelerates aging of the spinal structures (77). In AF cells, AGEs affects cell viability and proliferation by regulating mitochondrial apoptosis and ROS production in a dose-dependent manner (78). The formation of AGEs requires sustained hyperglycemia, so there is no change in AGEs during a short duration of hyperglycemia (79).
The CEP is composed of chondrocytes and matrix, located at the upper and lower edges of the vertebral body, and proteoglycan and type II collagen (80). The main function of the CEP is to distribute the mechanical load along the spine to protect the vertebrae from compressive vertebral atrophy under pressure and to provide nutrition for the IVD (81). After high glucose induction, the mitochondrial membrane potential of CEP cells decrease and ROS production and apoptosis related proteins increased. High glucose may cause damage to the CEP cells through oxidative stress and CEP apoptosis induced by high glucose is time-dependent (82). Moreover, lncRNA MALAT1 play an important role in promoting apoptosis of rat CEP cells by activating the p38/MAPK signal pathway. This suggests that MALAT1 can be used as a therapeutic target for diabetes related IDD (83).
The NP is located between the CEP and the AF, and is an elastic translucent gelatinous material composed of a crisscross fibrous reticular structure (84–86). The water content of the NP in infants is 80-90%. With an increase in age, the proteoglycan depolymerization in the NP increases, the water content decreases gradually and the collagen thickens and is gradually replaced by fibrocartilage (87, 88). Therefore, the incidence of IVD herniation in the elderly is significantly higher than that in young adults (89, 90).
The nutrition of the NP is mainly supplied by the vertebral body-cartilage endplate (84). During spinal movement, the NP rolls like a bearing to support the vertebral body and assists other parts of the spine to complete physiological activities (91). When the NP bears an external force, the force is uniformly transferred to the surrounding AF, which can balance the stress (86, 92). In a model of diabetes induced by streptozotocin injection, the protein levels of caspase-8, caspase-9, and caspase-3 apoptosis proteins in NP cells and the expression of aging marker p16lnk4A protein were significantly increased. In addition, the activation of the autophagy pathway in NP cells inhibits the expression of MMP-13 in diabetic IDD and triggers the protective mechanism of the NP cells (93, 94).
On the other hand, the addition of osteogenic protein-1 (OP-1) reverses NP matrix catabolism induced by high glucose (95). A high glucose concentration significantly decreases the proliferation, colony formation, migration, wound healing and dry maintenance of NP mesenchymal stem cells (NPMSCs) and accelerate the apoptosis and senescence of NPMSCs (96). In recent years, stem cell therapy has been widely used in cell-based regenerative medicine (97–99). Treatment of NPMSCs with hMSCs-CM inhibits the level of phosphorylated p38 MAPK (100). Autophagy has been shown to prevent premature aging under various conditions, and autophagy defects may lead to aging (101, 102). Through screening diabetes data -sets and tissue validation, it was found that HuR expression in diabetic NP tissue and high glucose treated NP cells was reduced. Knockdown of HuR promotes senescence of the NP cells. Further exploration of the mechanism shows that HuR can bind to the key autophagy gene Atg7 and regulate the stability of Atg7. In vivo injection of Atg7 overexpression improves IDD progression by promoting autophagy (103). It is of great significance to explore new methods to prevent IDD and cell senescence from the point of view of the NP and NPMSCs.
As the external structure of the IVD, the AF is used to protect the NP from being released from IVD during axial compression, tension and bending when the spine is under high load. The AF can maintain physiological intervertebral disc pressure. When AF cell degeneration also occurs, various pathological events may occur, including decreased IVD cells, upregulation of matrix degrading enzymes, and inflammation. Studies have shown that AF cells of young rats treated with high glucose for 1 and 3 days showed increased mitochondrial damage and increased cell senescence markers, excessive production of ROS and decreased average telomerase activity (104). Stress-induced accelerated premature senescence of young AF cells may be an emerging risk factor for premature disc degeneration in young DM patients. In addition, high glucose culture significantly increase the caspase-3 and caspase-9 activity of AF cells and promotes the activation of the apoptosis pathway. In mechanism, inhibition of JNK pathway and p38 MAPK signal pathways may attenuate the effect of high glucose on apoptosis of the AF cells (105). Furthermore, endoplasmic reticulum stress can lead to the disorder of steady-state behavior of many cells and induce IVD apoptosis to degenerate. After high glucose induction, the protein expression of ER stress markers CHOP, ATF-6, and ATF-6 significantly increased and induced apoptosis in the AF cells (106).
Surgery can directly and effectively solve the pain and neurological symptoms of patients, but it is not helpful for the prevention and progress of IDD. Therefore, effective drugs should be developed to intervene in the early stage of IDD or to prevent IDD in high-risk groups, to avoid trauma and risk of surgical treatment for patients. Research has found that adding resveratrol to high glucose induced NP cells can reduce apoptosis, ROS production, downregulate the expression of aging markers (p16 and p53), and increase telomerase activity and the expression of anti-apoptotic molecules (Bcl-2) in NP cells. On the contrary, inhibition of the PI3K/Akt pathway can counteract the beneficial effect of resveratrol in high glucose groups (107).
Glucagon-like peptide-1 (GLP-1) is a key intestinal insulin-stimulating hormone that regulates glucose and energy homeostasis (108, 109). Liraglutide (LIR), as a long-acting GLP-1 analog, is highly homologous to endogenous GLP-1 (110). LIR is considered to be a powerful therapeutic choice for T2D by binding to GLP-1R to regulate insulin and cell proliferation, differentiation and apoptosis (111, 112). In diabetic IDD, the addition of high concentration of LIR inhibited the apoptosis of NPs induced by high glucose, increasing cell activity and proliferation. The PI3K/Akt/caspase-3 pathway may be one of the mechanisms by which LIR plays a protective role in IDD (113, 114).
Marein is a flavonoid extracted from C. tinctoria, which can improve insulin resistance induced by high glucose levels. In IDD, Marein protects HNPC from HG-induced damage and ECM degradation by inhibiting the ROS/NF-κB pathway. Therefore, Marein is a promising therapeutic agent to delay IDD in patients with diabetes (115).
Cycloastragenol is the only telomerase activator that has anti-aging effects by increasing telomerase levels to delay telomere shortening (116, 117). Astragaloside IV (AG-IV) has a variety of drug properties, such as anti-inflammatory, anti-IR and neuroprotective effects (118–120). High glucose concentration is known to have adverse effects on telomerase reverse transcriptase (TERT) expression and telomere length in NPs. After treatment with CAG and AG-IV, the cell morphology and vitality significantly improve, the TERT expression of NPC and telomere length increased, and apoptosis and senescence an inhibited (121). At present, the exploration of novel pharmacotherapeutic strategies for treating IDD, particularly in the presence of concurrent diabetes, represents a highly promising avenue. Compounds such as resveratrol, liraglutide, and Marein exhibit diverse mechanisms of action, underscoring the potential of targeted drug interventions in mitigating IDD. It is crucial to comprehend how these drugs interact with conventional diabetes medications and their impact on blood glucose control. Longitudinal studies are imperative to assess the prolonged safety and efficacy of these drugs in IDD patients, especially considering the chronic nature of IDD and diabetes. The selection of pharmacological interventions should be personalized based on the individual patients circumstances, the stage of IDD, and the status of diabetes.
Studies have shown that both T1D and T2D can cause different degrees of IDD, and that high glucose and HFD are risk factors for diabetic IVD. High glucose can promote the aging of endplate cells, NP cells, and AF cells, increase cell apoptosis and inflammatory response, and hinder the formation of autophagy pathways and ECM degradation. The relevant mechanisms are described in Table 1.
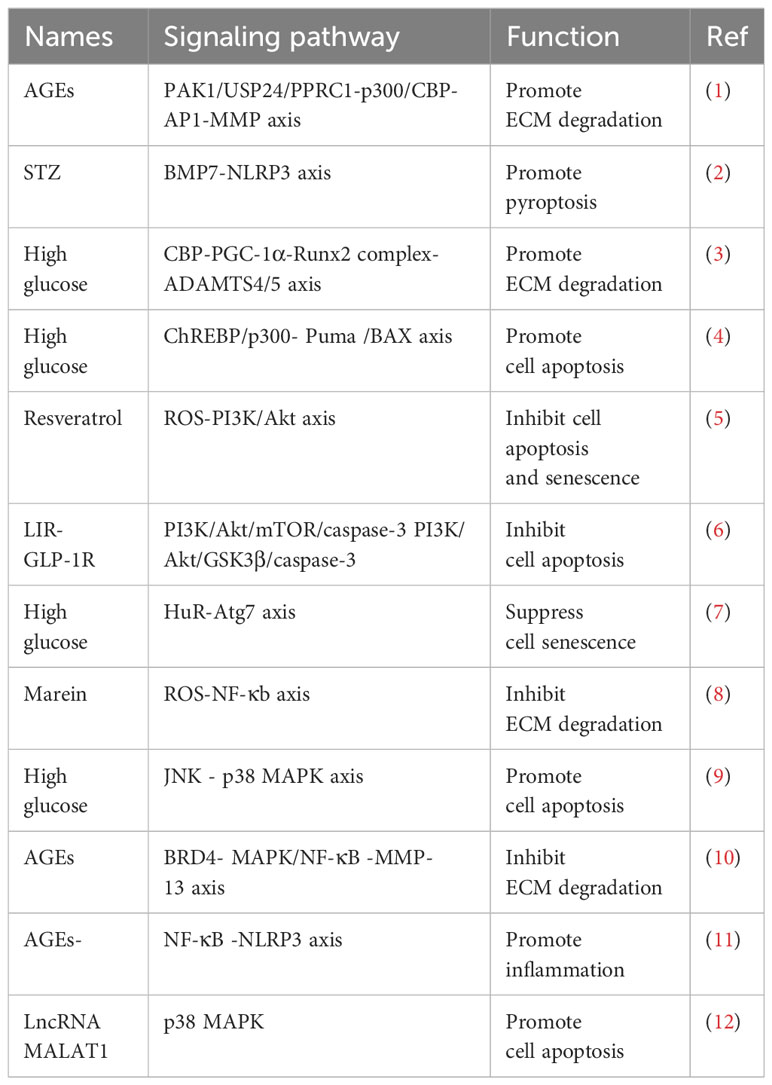
Table 1 Mechanisms involved in the progress of Diabetic IDD.
It is worth noting that some drugs have shown beneficial effects on alleviating high glucose induced IDD, but current drug therapy is still limited. We believe that health education for the population is the preferred strategy to preventing high glucose induced IDD in the future. Patients who adhere to a reasonable diet and regular living habits can effectively alleviate the progress of IDD. In addition, it is necessary to further reveal the mechanism of high glucose-induced IDD and develop specific targeting drugs. Drugs enter the body through blood circulation and are mostly metabolized by the liver and kidney. The effective dose to achieve the function of the IVD is usually greatly reduced. Therefore, the application of nanoparticles to encapsulate drug components and enable drugs to target the therapeutic area should be considered. For example, exosomes, EVs, and artificial vesicles can all serve as good carriers for drugs. In summary, current research provides a fundamental basis for the treatment of high glucose induced IDD patients. With the further exploration of more mechanisms of abnormal glucose metabolism affecting IDD and in the development of targeted drugs, it will bring dawn to the rehabilitation of patients.
SL: Writing – original draft. JD: Data curation, Writing – original draft. YH: Software, Writing – original draft. SG: Software, Writing – original draft. ZZ: Data curation, Writing – original draft. ZC: Investigation, Software, Writing – original draft. XZ: Formal Analysis, Funding acquisition, Software, Writing – original draft. BH: Supervision, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by Shaanxi Province key research and development plan project (2023JH−YXYB−0145) and Xian Science and Technology Bureau (23YXYJ0083).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Knezevic NN, Candido KD, Vlaeyen JWS, Van Zundert J, Cohen SP. Low back pain. Lancet (2021) 398(10294):78–92. doi: 10.1016/S0140-6736(21)00733-9
2. Vlaeyen JWS, Maher CG, Wiech K, Van Zundert J, Meloto CB, Diatchenko L, et al. Low back pain. Nat Rev Dis Primers (2018) 4(1):52. doi: 10.1038/s41572-018-0052-1
4. Meier ML, Vrana A, Schweinhardt P. Low back pain: the potential contribution of supraspinal motor control and proprioception. Neuroscientist (2019) 25(6):583–96. doi: 10.1177/1073858418809074
5. Kamali A, Ziadlou R, Lang G, Pfannkuche J, Cui S, Li Z, et al. Small molecule-based treatment approaches for intervertebral disc degeneration: Current options and future directions. Theranostics (2021) 11(1):27–47. doi: 10.7150/thno.48987
6. Hayden JA, Ellis J, Ogilvie R, Malmivaara A, van Tulder MW. Exercise therapy for chronic low back pain. Cochrane Database Syst Rev (2021) 9(9):Cd009790. doi: 10.1002/14651858.CD009790.pub2
7. Gianola S, Bargeri S, Del Castillo G, Corbetta D, Turolla A, Andreano A, et al. Effectiveness of treatments for acute and subacute mechanical non-specific low back pain: a systematic review with network meta-analysis. Br J Sports Med (2022) 56(1):41–50. doi: 10.1136/bjsports-2020-103596
8. Krut Z, Pelled G, Gazit D, Gazit Z. Stem cells and exosomes: new therapies for intervertebral disc degeneration. Cells (2021) 10(9). doi: 10.3390/cells10092241
9. Francisco V, Pino J, González-Gay M, Lago F, Karppinen J, Tervonen O, et al. A new immunometabolic perspective of intervertebral disc degeneration. Nat Rev Rheumatol (2022) 18(1):47–60. doi: 10.1038/s41584-021-00713-z
10. Mohd Isa IL, Mokhtar SA, Abbah SA, Fauzi MB, Devitt A, Pandit A. Intervertebral disc degeneration: biomaterials and tissue engineering strategies toward precision medicine. Adv Healthc Mater (2022) 11(13):e2102530. doi: 10.1002/adhm.202102530
11. Williams RJ, Tryfonidou MA, Snuggs JW, Le Maitre CL. Cell sources proposed for nucleus pulposus regeneration. JOR Spine (2021) 4(4):e1175. doi: 10.1002/jsp2.1175
12. Sun K, Jiang J, Wang Y, Sun X, Zhu J, Xu X, et al. The role of nerve fibers and their neurotransmitters in regulating intervertebral disc degeneration. Ageing Res Rev (2022) 81:101733. doi: 10.1016/j.arr.2022.101733
13. Torre OM, Mroz V, Bartelstein MK, Huang AH, Iatridis JC. Annulus fibrosus cell phenotypes in homeostasis and injury: implications for regenerative strategies. Ann N Y Acad Sci (2019) 1442(1):61–78. doi: 10.1111/nyas.13964
14. Zhang A, Cheng Z, Chen Y, Shi P, Gan W, Zhang Y. Emerging tissue engineering strategies for annulus fibrosus therapy. Acta Biomater (2023) 167:1–15. doi: 10.1016/j.actbio.2023.06.012
15. Zehra U, Tryfonidou M, Iatridis JC, Illien-Jünger S, Mwale F, Samartzis D. Mechanisms and clinical implications of intervertebral disc calcification. Nat Rev Rheumatol (2022) 18(6):352–62. doi: 10.1038/s41584-022-00783-7
16. Xiang Q, Zhao Y, Lin J, Jiang S, Li W. The Nrf2 antioxidant defense system in intervertebral disc degeneration: Molecular insights. Exp Mol Med (2022) 54(8):1067–75. doi: 10.1038/s12276-022-00829-6
17. Mohd Isa IL, Teoh SL, Mohd Nor NH, Mokhtar SA. Discogenic low back pain: anatomy, pathophysiology and treatments of intervertebral disc degeneration. Int J Mol Sci (2022) 24(1). doi: 10.3390/ijms24010208
18. Wang Y, Che M, Xin J, Zheng Z, Li J, Zhang S. The role of IL-1β and TNF-α in intervertebral disc degeneration. BioMed Pharmacother (2020) 131:110660. doi: 10.1016/j.biopha.2020.110660
19. Wang F, Cai F, Shi R, Wang XH, Wu XT. Aging and age related stresses: a senescence mechanism of intervertebral disc degeneration. Osteoarthritis Cartilage (2016) 24(3):398–408. doi: 10.1016/j.joca.2015.09.019
20. Zhang GZ, Liu MQ, Chen HW, Wu ZL, Gao YC, Ma ZJ, et al. NF-κB signalling pathways in nucleus pulposus cell function and intervertebral disc degeneration. Cell Prolif (2021) 54(7):e13057. doi: 10.1111/cpr.13057
21. Cao G, Yang S, Cao J, Tan Z, Wu L, Dong F, et al. The role of oxidative stress in intervertebral disc degeneration. Oxid Med Cell Longev (2022) 2022:2166817. doi: 10.1155/2022/2166817
22. Zhang HJ, Liao HY, Bai DY, Wang ZQ, Xie XW. MAPK /ERK signaling pathway: A potential target for the treatment of intervertebral disc degeneration. BioMed Pharmacother (2021) 143:112170. doi: 10.1016/j.biopha.2021.112170
23. Zhang Z, Lin J, Nisar M, Chen T, Xu T, Zheng G, et al. The sirt1/P53 axis in diabetic intervertebral disc degeneration pathogenesis and therapeutics. Oxid Med Cell Longev (2019) 2019:7959573. doi: 10.1155/2019/7959573
24. Wu J, Chen Y, Liao Z, Liu H, Zhang S, Zhong D, et al. Self-amplifying loop of NF-κB and periostin initiated by PIEZO1 accelerates mechano-induced senescence of nucleus pulposus cells and intervertebral disc degeneration. Mol Ther (2022) 30(10):3241–56. doi: 10.1016/j.ymthe.2022.05.021
25. Liang W, Han B, Hai Y, Sun D, Yin P. Mechanism of action of mesenchymal stem cell-derived exosomes in the intervertebral disc degeneration treatment and bone repair and regeneration. Front Cell Dev Biol (2021) 9:833840. doi: 10.3389/fcell.2021.833840
26. Zhang B, Xu H, Wang J, Liu B, Sun G. A narrative review of non-operative treatment, especially traditional Chinese medicine therapy, for lumbar intervertebral disc herniation. Biosci Trends (2017) 11(4):406–17. doi: 10.5582/bst.2017.01199
27. Lindback Y, Tropp H, Enthoven P, Abbott A, Öberg B. PREPARE: presurgery physiotherapy for patients with degenerative lumbar spine disorder: a randomized controlled trial. Spine J (2018) 18(8):1347–55. doi: 10.1016/j.spinee.2017.12.009
28. Liao Z, Su D, Liu H, Xu C, Wu J, Chen Y, et al. Dihydroartemisinin attenuated intervertebral disc degeneration via inhibiting PI3K/AKT and NF-κB signaling pathways. Oxid Med Cell Longev (2022) 2022:8672969. doi: 10.1155/2022/8672969
29. Lu R, Xu H, Deng X, Wang Y, He Z, Xu S, et al. Physalin A alleviates intervertebral disc degeneration via anti-inflammatory and anti-fibrotic effects. J Orthop Translat (2023) 39:74–87. doi: 10.1016/j.jot.2023.01.001
30. Wu PH, Kim HS, Jang IT. Intervertebral disc diseases PART 2: A review of the current diagnostic and treatment strategies for intervertebral disc disease. Int J Mol Sci (2020) 21(6). doi: 10.3390/ijms21062135
31. Chen BL, Guo JB, Zhang HW, Zhang YJ, Zhu Y, Zhang J, et al. Surgical versus non-operative treatment for lumbar disc herniation: a systematic review and meta-analysis. Clin Rehabil (2018) 32(2):146–60. doi: 10.1177/0269215517719952
32. Sandhu FA, Dowlati E, Garica R. Lumbar arthroplasty: past, present, and future. Neurosurgery (2020) 86(2):155–69. doi: 10.1093/neuros/nyz439
33. Eisenstein SM, Balain B, Roberts S. Current treatment options for intervertebral disc pathologies. Cartilage (2020) 11(2):143–51. doi: 10.1177/1947603520907665
34. De Pieri A, Byerley AM, Musumeci CR, Salemizadehparizi F, Vanderhorst MA, Wuertz-Kozak K. Electrospinning and 3D bioprinting for intervertebral disc tissue engineering. JOR Spine (2020) 3(4):e1117. doi: 10.1002/jsp2.1117
35. Bydon M, Goyal A, Yolcu YU. Novel intervertebral technologies. Neurosurg Clin N Am (2020) 31(1):49–56. doi: 10.1016/j.nec.2019.08.006
36. Liu Z, Wang H, Yuan Z, Wei Q, Han F, Chen S, et al. High-resolution 3D printing of angle-ply annulus fibrosus scaffolds for intervertebral disc regeneration. Biofabrication (2022) 15(1). doi: 10.1088/1758-5090/aca71f
37. Silwal P, Nguyen-Thai AM, Mohammad HA, Wang Y, Robbins PD, Lee JY, et al. Cellular senescence in intervertebral disc aging and degeneration: molecular mechanisms and potential therapeutic opportunities. Biomolecules (2023) 13(4). doi: 10.3390/biom13040686
38. Risbud MV, Shapiro IM. Role of cytokines in intervertebral disc degeneration: pain and disc content. Nat Rev Rheumatol (2014) 10(1):44–56. doi: 10.1038/nrrheum.2013.160
39. Vergroesen PP, Kingma I, Emanuel KS, Hoogendoorn RJ, Welting TJ, van Royen BJ, et al. Mechanics and biology in intervertebral disc degeneration: a vicious circle. Osteoarthritis Cartilage (2015) 23(7):1057–70. doi: 10.1016/j.joca.2015.03.028
40. Li M, Wu Y, Li H, Tan C, Ma S, Gong J, et al. Nanofiber reinforced alginate hydrogel for leak-proof delivery and higher stress loading in nucleus pulposus. Carbohydr Polym (2023) 299:120193. doi: 10.1016/j.carbpol.2022.120193
41. Chen S, Liu S, Ma K, Zhao L, Lin H, Shao Z. TGF-β signaling in intervertebral disc health and disease. Osteoarthritis Cartilage (2019) 27(8):1109–17. doi: 10.1016/j.joca.2019.05.005
42. Liao Z, Li S, Lu S, Liu H, Li G, Ma L, et al. Metformin facilitates mesenchymal stem cell-derived extracellular nanovesicles release and optimizes therapeutic efficacy in intervertebral disc degeneration. Biomaterials (2021) 274:120850. doi: 10.1016/j.biomaterials.2021.120850
43. Tong W, Lu Z, Qin L, Mauck RL, Smith HE, Smith LJ, et al. Cell therapy for the degenerating intervertebral disc. Transl Res (2017) 181:49–58. doi: 10.1016/j.trsl.2016.11.008
44. DiStefano TJ, Vaso K, Danias G, Chionuma HN, Weiser JR, Iatridis JC. Extracellular vesicles as an emerging treatment option for intervertebral disc degeneration: therapeutic potential, translational pathways, and regulatory considerations. Adv Healthc Mater (2022) 11(5):e2100596. doi: 10.1002/adhm.202100596
45. van Niel G, D'Angelo G, Raposo G. Shedding light on the cell biology of extracellular vesicles. Nat Rev Mol Cell Biol (2018) 19(4):213–28. doi: 10.1038/nrm.2017.125
46. Isaac R, Reis FCG, Ying W, Olefsky JM. Exosomes as mediators of intercellular crosstalk in metabolism. Cell Metab (2021) 33(9):1744–62. doi: 10.1016/j.cmet.2021.08.006
47. Fabbiano F, Corsi J, Gurrieri E, Trevisan C, Notarangelo M, D'Agostino VG. RNA packaging into extracellular vesicles: An orchestra of RNA-binding proteins? J Extracell Vesicles (2020) 10(2):e12043. doi: 10.1002/jev2.12043
48. Dooley K, McConnell RE, Xu K, Lewis ND, Haupt S, Youniss MR, et al. A versatile platform for generating engineered extracellular vesicles with defined therapeutic properties. Mol Ther (2021) 29(5):1729–43. doi: 10.1016/j.ymthe.2021.01.020
49. Jung M, Rospleszcz S, Löffler MT, Walter SS, Maurer E, Jungmann PM, et al. Association of lumbar vertebral bone marrow and paraspinal muscle fat composition with intervertebral disc degeneration: 3T quantitative MRI findings from the population-based KORA study. Eur Radiol (2023) 33(3):1501–12. doi: 10.1007/s00330-022-09140-4
50. Lintz M, Walk RE, Tang SY, Bonassar LJ. The degenerative impact of hyperglycemia on the structure and mechanics of developing murine intervertebral discs. JOR Spine (2022) 5(1):e1191. doi: 10.1002/jsp2.1191
51. Kang L, Zhang H, Jia C, Zhang R, Shen C. Epigenetic modifications of inflammation in intervertebral disc degeneration. Ageing Res Rev (2023) 87:101902. doi: 10.1016/j.arr.2023.101902
52. Atkinson MA, Eisenbarth GS, Michels AW. Type 1 diabetes. Lancet (2014) 383(9911):69–82. doi: 10.1016/S0140-6736(13)60591-7
53. Chellappan DK, Sivam NS, Teoh KX, Leong WP, Fui TZ, Chooi K, et al. Gene therapy and type 1 diabetes mellitus. BioMed Pharmacother (2018) 108:1188–200. doi: 10.1016/j.biopha.2018.09.138
54. Chen R, Liang X, Huang T, Zhong W, Luo X. Effects of type 1 diabetes mellitus on lumbar disc degeneration: a retrospective study of 118 patients. J Orthop Surg Res (2020) 15(1):280. doi: 10.1186/s13018-020-01784-6
55. Yu XJ, Wang YG, Lu R, Guo XZ, Qu YK, Wang SX, et al. BMP7 ameliorates intervertebral disc degeneration in type 1 diabetic rats by inhibiting pyroptosis of nucleus pulposus cells and NLRP3 inflammasome activity. Mol Med (2023) 29(1):30. doi: 10.1186/s10020-023-00623-8
56. Russo F, Ambrosio L, Ngo K, Vadalà G, Denaro V, Fan Y, et al. The role of type I diabetes in intervertebral disc degeneration. Spine (Phila Pa 1976) (2019) 44(17):1177–85. doi: 10.1097/BRS.0000000000003054
57. Laakso M. Biomarkers for type 2 diabetes. Mol Metab (2019) 27s:S139–s146. doi: 10.1016/j.molmet.2019.06.016
58. Demir S, Nawroth PP, Herzig S, Ekim Üstünel B. Emerging targets in type 2 diabetes and diabetic complications. Adv Sci (Weinh) (2021) 8(18):e2100275. doi: 10.1002/advs.202100275
59. Vekic J, Vujcic S, Bufan B, Bojanin D, Al-Hashmi K, Al-Rasadi K, et al. The role of advanced glycation end products on dyslipidemia. Metabolites (2023) 13(1). doi: 10.3390/metabo13010077
60. Bhatti JS, Sehrawat A, Mishra J, Sidhu IS, Navik U, Khullar N, et al. Oxidative stress in the pathophysiology of type 2 diabetes and related complications: Current therapeutics strategies and future perspectives. Free Radic Biol Med (2022) 184:114–34. doi: 10.1016/j.freeradbiomed.2022.03.019
61. Agius R, Galea R, Fava S. Bone mineral density and intervertebral disc height in type 2 diabetes. J Diabetes Complications (2016) 30(4):644–50. doi: 10.1016/j.jdiacomp.2016.01.021
62. Fields AJ, Berg-Johansen B, Metz LN, Miller S, La B, Liebenberg EC, et al. Alterations in intervertebral disc composition, matrix homeostasis and biomechanical behavior in the UCD-T2DM rat model of type 2 diabetes. J Orthop Res (2015) 33(5):738–46. doi: 10.1002/jor.22807
63. Tseng C, Han Y, Lv Z, Song Q, Wang K, Shen H, et al. Glucose-stimulated PGC-1α couples with CBP and Runx2 to mediate intervertebral disc degeneration through transactivation of ADAMTS4/5 in diet-induced obesity mice. Bone (2023) 167:116617. doi: 10.1016/j.bone.2022.116617
64. Natelson DM, Lai A, Krishnamoorthy D, Hoy RC, Iatridis JC, Illien-Jünger S. Leptin signaling and the intervertebral disc: Sex dependent effects of leptin receptor deficiency and Western diet on the spine in a type 2 diabetes mouse model. PloS One (2020) 15(5):e0227527. doi: 10.1371/journal.pone.0227527
65. Li X, Liu X, Wang Y, Cao F, Chen Z, Hu Z, et al. Intervertebral disc degeneration in mice with type II diabetes induced by leptin receptor deficiency. BMC Musculoskelet Disord (2020) 21(1):77. doi: 10.1186/s12891-020-3091-1
66. Mahmoud M, Kokozidou M, Gögele C, Werner C, Auffarth A, Kohl B, et al. Does vitamin K2 influence the interplay between diabetes mellitus and intervertebral disc degeneration in a rat model? Nutrients (2023) 15(13). doi: 10.3390/nu15132872
67. Khalid M, Petroianu G, Adem A. Advanced glycation end products and diabetes mellitus: mechanisms and perspectives. Biomolecules (2022) 12(4). doi: 10.3390/biom12040542
68. Lee J, Yun JS, Ko SH. Advanced glycation end products and their effect on vascular complications in type 2 diabetes mellitus. Nutrients (2022) 14(15). doi: 10.3390/nu14153086
69. Moldogazieva NT, Mokhosoev IM, Mel'nikova TI, Porozov YB, Terentiev AA. Oxidative stress and advanced lipoxidation and glycation end products (ALEs and AGEs) in aging and age-related diseases. Oxid Med Cell Longev (2019) 2019:3085756. doi: 10.1155/2019/3085756
70. Dong L, Li Y, Chen Q, Liu Y, Wu Z, Pan D, et al. Cereal polyphenols inhibition mechanisms on advanced glycation end products and regulation on type 2 diabetes. Crit Rev Food Sci Nutr (2023), 1–19. doi: 10.1080/10408398.2023.2213768
71. Krishnamoorthy D, Hoy RC, Natelson DM, Torre OM, Laudier DM, Iatridis JC, et al. Dietary advanced glycation end-product consumption leads to mechanical stiffening of murine intervertebral discs. Dis Model Mech (2018) 11(12). doi: 10.1101/342691
72. Xu YC, Gu Y, Yang JY, Xi K, Tang JC, Bian J, et al. RACK1 mediates the advanced glycation end product-induced degradation of HIF-1α in nucleus pulposus cells via competing with HSP90 for HIF-1α binding. Cell Biol Int (2021) 45(6):1316–26. doi: 10.1002/cbin.11574
73. Tseng C, Chen B, Han Y, Wang K, Song Q, Shen H, et al. Advanced glycation end products promote intervertebral disc degeneration by transactivation of matrix metallopeptidase genes. Osteoarthritis Cartilage (2023). doi: 10.1016/j.joca.2023.09.005
74. Gallate ZS, D'Erminio DN, Nasser P, Laudier DM, Iatridis JC. Galectin-3 and RAGE differentially control advanced glycation endproduct-induced collagen damage in murine intervertebral disc organ culture. JOR Spine (2023) 6(2):e1254. doi: 10.1002/jsp2.1254
75. Glaeser JD, Ju D, Tawackoli W, Yang JH, Salehi K, Stefanovic T, et al. Advanced glycation end product inhibitor pyridoxamine attenuates IVD degeneration in type 2 diabetic rats. Int J Mol Sci (2020) 21(24). doi: 10.3390/ijms21249709
76. Song Y, Wang Y, Zhang Y, Geng W, Liu W, Gao Y, et al. Advanced glycation end products regulate anabolic and catabolic activities via NLRP3-inflammasome activation in human nucleus pulposus cells. J Cell Mol Med (2017) 21(7):1373–87. doi: 10.1111/jcmm.13067
77. Illien-Junger S, Lu Y, Qureshi SA, Hecht AC, Cai W, Vlassara H, et al. Chronic ingestion of advanced glycation end products induces degenerative spinal changes and hypertrophy in aging pre-diabetic mice. PloS One (2015) 10(2):e0116625. doi: 10.1371/journal.pone.0116625
78. Hu Y, Shao Z, Cai X, Liu Y, Shen M, Yao Y, et al. Mitochondrial pathway is involved in advanced glycation end products-induced apoptosis of rabbit annulus fibrosus cells. Spine (Phila Pa 1976) (2019) 44(10):E585–e595. doi: 10.1097/BRS.0000000000002930
79. Tsai TT, Ho NY, Lin YT, Lai PL, Fu TS, Niu CC, et al. Advanced glycation end products in degenerative nucleus pulposus with diabetes. J Orthop Res (2014) 32(2):238–44. doi: 10.1002/jor.22508
80. Liu W, Ma Z, Wang Y, Yang J. Multiple nano-drug delivery systems for intervertebral disc degeneration: Current status and future perspectives. Bioact Mater (2023) 23:274–99. doi: 10.1016/j.bioactmat.2022.11.006
81. Chen J, Yang X, Feng Y, Li Q, Ma J, Wang L, et al. Targeting ferroptosis holds potential for intervertebral disc degeneration therapy. Cells (2022) 11(21). doi: 10.3390/cells11213508
82. Jiang Z, Lu W, Zeng Q, Li D, Ding L, Wu J. High glucose-induced excessive reactive oxygen species promote apoptosis through mitochondrial damage in rat cartilage endplate cells. J Orthop Res (2018) 36(9):2476–83. doi: 10.1002/jor.24016
83. Jiang Z, Zeng Q, Li D, Ding L, Lu W, Bian M, et al. Long non−coding RNA MALAT1 promotes high glucose−induced rat cartilage endplate cell apoptosis via the p38/MAPK signalling pathway. Mol Med Rep (2020) 21(5):2220–6. doi: 10.3892/mmr
84. Guerrero J, Häckel S, Croft AS, Hoppe S, Albers CE, Gantenbein B. The nucleus pulposus microenvironment in the intervertebral disc: the fountain of youth? Eur Cell Mater (2021) 41:707–38. doi: 10.22203/eCM
85. Lawson L, Harfe BD. Notochord to nucleus pulposus transition. Curr Osteoporos Rep (2015) 13(5):336–41. doi: 10.1007/s11914-015-0284-x
86. Molladavoodi S, McMorran J, Gregory D. Mechanobiology of annulus fibrosus and nucleus pulposus cells in intervertebral discs. Cell Tissue Res (2020) 379(3):429–44. doi: 10.1007/s00441-019-03136-1
87. Silagi ES, Shapiro IM, Risbud MV. Glycosaminoglycan synthesis in the nucleus pulposus: Dysregulation and the pathogenesis of disc degeneration. Matrix Biol (2018) 71-72:368–79.
88. Singh K, Masuda K, Thonar EJ, An HS, Cs-Szabo G. Age-related changes in the extracellular matrix of nucleus pulposus and anulus fibrosus of human intervertebral disc. Spine (Phila Pa 1976) (2009) 34(1):10–6. doi: 10.1097/BRS.0b013e31818e5ddd
89. Song C, Zhou Y, Cheng K, Liu F, Cai W, Zhou D, et al. Cellular senescence - Molecular mechanisms of intervertebral disc degeneration from an immune perspective. BioMed Pharmacother (2023) 162:114711. doi: 10.1016/j.biopha.2023.114711
90. Zhang Y, Yang B, Wang J, Cheng F, Shi K, Ying L, et al. Cell senescence: A nonnegligible cell state under survival stress in pathology of intervertebral disc degeneration. Oxid Med Cell Longev (2020) 2020:9503562. doi: 10.1155/2020/9503562
91. Mordechai HS, Aharonov A, Sharon SE, Bonshtein I, Simon C, Sivan SS, et al. Toward a mechanically biocompatible intervertebral disc: Engineering of combined biomimetic annulus fibrosus and nucleus pulposus analogs. J BioMed Mater Res A (2023) 111(5):618–33. doi: 10.1002/jbm.a.37519
92. Yang B, O'Connell GD. Intervertebral disc swelling maintains strain homeostasis throughout the annulus fibrosus: A finite element analysis of healthy and degenerated discs. Acta Biomater (2019) 100:61–74. doi: 10.1016/j.actbio.2019.09.035
93. Jiang L, Zhang X, Zheng X, Ru A, Ni X, Wu Y, et al. Apoptosis, senescence, and autophagy in rat nucleus pulposus cells: Implications for diabetic intervertebral disc degeneration. J Orthop Res (2013) 31(5):692–702. doi: 10.1002/jor.22289
94. Wang J, Hu J, Chen X, Huang C, Lin J, Shao Z, et al. BRD4 inhibition regulates MAPK, NF-κB signals, and autophagy to suppress MMP-13 expression in diabetic intervertebral disc degeneration. FASEB J (2019) 33(10):11555–66. doi: 10.1096/fj.201900703R
95. Liu Z, Zhang Z, Zhang A, Zhang F, Du W, Zhang Y, et al. Osteogenic protein-1 alleviates high glucose microenvironment-caused degenerative changes in nucleus pulposus cells. Biosci Rep (2019) 39(4). doi: 10.1042/BSR20190170
96. Liu Y, Li Y, Nan LP, Wang F, Zhou SF, Wang JC, et al. The effect of high glucose on the biological characteristics of nucleus pulposus-derived mesenchymal stem cells. Cell Biochem Funct (2020) 38(2):130–40. doi: 10.1002/cbf.3441
97. Qin Y, Ge G, Yang P, Wang L, Qiao Y, Pan G, et al. An update on adipose-derived stem cells for regenerative medicine: where challenge meets opportunity. Adv Sci (Weinh) (2023) 10(20):e2207334. doi: 10.1002/advs.202207334
98. Margiana R, Markov A, Zekiy AO, Hamza MU, Al-Dabbagh KA, Al-Zubaidi SH, et al. Clinical application of mesenchymal stem cell in regenerative medicine: a narrative review. Stem Cell Res Ther (2022) 13(1):366. doi: 10.1186/s13287-022-03054-0
99. Zhu Y, Ge J, Huang C, Liu H, Jiang H. Application of mesenchymal stem cell therapy for aging frailty: from mechanisms to therapeutics. Theranostics (2021) 11(12):5675–85. doi: 10.7150/thno.46436
100. Qi L, Wang R, Shi Q, Yuan M, Jin M, Li D. Umbilical cord mesenchymal stem cell conditioned medium restored the expression of collagen II and aggrecan in nucleus pulposus mesenchymal stem cells exposed to high glucose. J Bone Miner Metab (2019) 37(3):455–66. doi: 10.1007/s00774-018-0953-9
101. Kaushik S, Tasset I, Arias E, Pampliega O, Wong E, Martinez-Vicente M, et al. Autophagy and the hallmarks of aging. Ageing Res Rev (2021) 72:101468. doi: 10.1016/j.arr.2021.101468
102. Wong SQ, Kumar AV, Mills J, Lapierre LR. Autophagy in aging and longevity. Hum Genet (2020) 139(3):277–90. doi: 10.1007/s00439-019-02031-7
103. Shao Z, Ni L, Hu S, Xu T, Meftah Z, Yu Z, et al. RNA-binding protein HuR suppresses senescence through Atg7 mediated autophagy activation in diabetic intervertebral disc degeneration. Cell Prolif (2021) 54(2):e12975. doi: 10.1111/cpr.12975
104. Park JS, Park JB, Park IJ, Park EY. Accelerated premature stress-induced senescence of young annulus fibrosus cells of rats by high glucose-induced oxidative stress. Int Orthop (2014) 38(6):1311–20. doi: 10.1007/s00264-014-2296-z
105. Shan L, Yang D, Zhu D, Feng F, Li X. High glucose promotes annulus fibrosus cell apoptosis through activating the JNK and p38 MAPK pathways. Biosci Rep (2019) 39(7). doi: 10.1042/BSR20190853
106. Pang L, Yang K, Zhang Z. High-glucose environment accelerates annulus fibrosus cell apoptosis by regulating endoplasmic reticulum stress. Biosci Rep (2020) 40(7). doi: 10.1042/BSR20200262
107. Wang W, Li P, Xu J, Wu X, Guo Z, Fan L, et al. Resveratrol attenuates high glucose-induced nucleus pulposus cell apoptosis and senescence through activating the ROS-mediated PI3K/Akt pathway. Biosci Rep (2018) 38(2). doi: 10.1042/BSR20171454
108. Muller TD, Finan B, Bloom SR, D'Alessio D, Drucker DJ, Flatt PR, et al. Glucagon-like peptide 1 (GLP-1). Mol Metab (2019) 30:72–130.
109. Gribble FM, Reimann F. Metabolic Messengers: glucagon-like peptide 1. Nat Metab (2021) 3(2):142–8. doi: 10.1038/s42255-020-00327-x
110. Knudsen LB, Lau J. The discovery and development of liraglutide and semaglutide. Front Endocrinol (Lausanne) (2019) 10:155. doi: 10.3389/fendo.2019.00155
111. Yousef CC, Thomas A, Matar MA, Ghandoura L, Aldossary I, Almuhanna SM, et al. Liraglutide effects on glycemic control and weight in patients with type 2 diabetes Mellitus: A real-world, observational study and brief narrative review. Diabetes Res Clin Pract (2021) 177:108871. doi: 10.1016/j.diabres.2021.108871
112. Gentilella R, Pechtner V, Corcos A, Consoli A. Glucagon-like peptide-1 receptor agonists in type 2 diabetes treatment: are they all the same? Diabetes Metab Res Rev (2019) 35(1):e3070. doi: 10.1002/dmrr.3070
113. Ming-Yan Y, Jing Z, Shu-Qin G, Xiao-Liang B, Zhi-Hong L, Xue Z. Liraglutide inhibits the apoptosis of human nucleus pulposus cells induced by high glucose through PI3K/Akt/caspase-3 signaling pathway. Biosci Rep (2019) 39(8). doi: 10.1042/BSR20190109
114. Yao M, Zhang J, Li Z, Bai X, Ma J, Li Y. Liraglutide protects nucleus pulposus cells against high-glucose induced apoptosis by activating PI3K/akt/ mTOR/caspase-3 and PI3K/akt/GSK3β/caspase-3 signaling pathways. Front Med (Lausanne) (2021) 8:630962. doi: 10.3389/fmed.2021.630962
115. Yao M, Zhang J, Li Z, Guo S, Zhou X, Zhang W. Marein protects human nucleus pulposus cells against high glucose-induced injury and extracellular matrix degradation at least partly by inhibition of ROS/NF-κB pathway. Int Immunopharmacol (2020) 80:106126. doi: 10.1016/j.intimp.2019.106126
116. Yilmaz S, Bedir E, Ballar Kirmizibayrak P. The role of cycloastragenol at the intersection of NRF2/ARE, telomerase, and proteasome activity. Free Radic Biol Med (2022) 188:105–16. doi: 10.1016/j.freeradbiomed.2022.06.230
117. Ullah M, Sun Z. Klotho deficiency accelerates stem cells aging by impairing telomerase activity. J Gerontol A Biol Sci Med Sci (2019) 74(9):1396–407. doi: 10.1093/gerona/gly261
118. Xia ML, Xie XH, Ding JH, Du RH, Hu G. Astragaloside IV inhibits astrocyte senescence: implication in Parkinson's disease. J Neuroinflamm (2020) 17(1):105. doi: 10.1186/s12974-020-01791-8
119. Yao M, Zhang L, Wang L. Astragaloside IV: A promising natural neuroprotective agent for neurological disorders. BioMed Pharmacother (2023) 159:114229. doi: 10.1016/j.biopha.2023.114229
120. Yin Y, Qu H, Yang Q, Fang Z, Gao R. Astragaloside IV alleviates Schwann cell injury in diabetic peripheral neuropathy by regulating microRNA-155-mediated autophagy. Phytomedicine (2021) 92:153749. doi: 10.1016/j.phymed.2021.153749
Keywords: Intervertebral disc degeneration, high glucose, AGEs, cartilage endplates, nucleus pulposus, annulus fibrosus
Citation: Li S, Du J, Huang Y, Gao S, Zhao Z, Chang Z, Zhang X and He B (2024) From hyperglycemia to intervertebral disc damage: exploring diabetic-induced disc degeneration. Front. Immunol. 15:1355503. doi: 10.3389/fimmu.2024.1355503
Received: 14 December 2023; Accepted: 05 February 2024;
Published: 20 February 2024.
Edited by:
Xijie Yu, West China Hospital of Sichuan University, ChinaReviewed by:
Zhonghai Li, First Affiliated Hospital of Dalian Medical University, ChinaCopyright © 2024 Li, Du, Huang, Gao, Zhao, Chang, Zhang and He. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: BaoRong He, anpoYnJsc0AxMjYuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.