 Yang Yang
Yang Yang Qing Hong
Qing Hong Xuehong Zhang
Xuehong Zhang Zhenmin Liu
Zhenmin Liu
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Immunol., 06 March 2024
Sec. Autoimmune and Autoinflammatory Disorders : Autoimmune Disorders
Volume 15 - 2024 | https://doi.org/10.3389/fimmu.2024.1331486
Rheumatoid arthritis (RA) is a systemic autoimmune disorder characterized by swollen joints, discomfort, stiffness, osteoporosis, and reduced functionality. Genetics, smoking, dust inhalation, high BMI, and hormonal and gut microbiota dysbiosis are all likely causes of the onset or development of RA, but the underlying mechanism remains unknown. Compared to healthy controls, patients with RA have a significantly different composition of gut microbiota. It is well known that the human gut microbiota plays a key role in the initiation, maintenance, and operation of the host immune system. Gut microbiota dysbiosis has local or systematic adverse effects on the host immune system, resulting in host susceptibility to various diseases, including RA. Studies on the intestinal microbiota modulation and immunomodulatory properties of probiotics have been reported, in order to identify their potential possibility in prevention and disease activity control of RA. This review summarized current studies on the role and potential mechanisms of gut microbiota in the development and progression of RA, as well as the preventative and therapeutic effects and potential mechanisms of probiotics on RA. Additionally, we proposed the challenges and difficulties in the application of probiotics in RA, providing the direction for the research and application of probiotics in the prevention of RA.
Rheumatoid arthritis (RA) is a chronic autoimmune disease that can lead to joint swelling, joint pain, morning stiffness, joint deformities, osteoporosis, and even disability (1). RA affects approximately 0.5-1.0% of adults aged 20-40 years globally and is more prevalent among individuals over the age of 75 (2, 3). Due to the unclear etiology and pathogenesis of RA, the main objective of treatment is to reduce joint inflammation and control the progression of lesions. The drugs used to treat RA include conventional synthetic disease-modifying antirheumatic drugs (csDMARDs), biologic DMARDs (bDMARDs), targeted synthetic DMARDs (tsDMARDs), and glucocorticoids (4). Early treatment is essential in RA management as it can effectively prevent irreversible structural damage and chronic functional impairment (5). Prior to the onset of clinical symptoms, RA is preceded by a preclinical phase that can last several years, during which RA-related autoimmunity is serologically detectable. This phase is characterized by the presence of autoantibodies, namely anti-citrullinated protein antibodies (ACPAs) and Fc domain-recognizing rheumatoid factors (RFs) (6). As RA progresses, synovial inflammation develops due to the infiltration of mononuclear cells, particularly CD4+ T cells, and macrophages, along with early activation of stromal cells. Inflammatory cells accumulate in the synovium, leading to the formation of a pannus that can destroy cartilage and further propagate the inflammatory process, eventually leading to clinical arthritis (7).
The symptoms of RA are associated with inflammation. Two key pathogenetic changes in the synovium in RA are an increase and activation of synoviocytes (8), as well as the infiltration of adaptive immune cells into the synovial sublining (9). These alterations are a prominent source of cytokines, proteases, and antibodies. The inflammation in the synovium leads to synovial hyperplasia and invasion into adjacent articular structures and eventually joint damage (10). The pathways involved in joint damage may include the release of cytokines and matrix metalloproteinases (MMPs) of macrophages, neutrophils (particularly in the synovial fluid) and mast cells (7). The maturation and activation of osteoclasts are the dominant factors of bone damage, while the interaction between ACPAs and citrullinated peptides expressed by osteoclasts and osteoclast precursors was the reason for osteoclast maturation and activation (11). However, studies suggest that the interaction above could precede the onset of synovial inflammation (12, 13).
A variety of risk factors are involved in the development of RA, including genetic and environmental factors. Studies on genetic predisposition to RA have implicated RA-related genes such as human leukocyte antigen (HLA) alleles as well as many other genes with low relative risk variants. Specific class II HLA loci can encode major histocompatibility complex (MHC) molecules that may contain the shared epitope (SE) (14). SE is a specific amino acid motif and is characterized by conserved amino acids within the peptide-binding groove, which are linked to an increased risk of developing RA (15). Moreover, environmental factors such as smoking, dust inhalation, high BMI, and hormonal factors have been demonstrated to contribute to the pathogenesis of RA (16). Heavy smoking, for example, heightens the likelihood of testing positive for ACPAs that are associated with joint symptoms (17, 18). Exposure to silica is also correlated with an elevated risk of future RA development (19).
In addition, various studies have suggested that mucosal environmental exposures and/or mucosal dysbiosis play a causal role in the period of susceptibility to RA (20–23). These researches support the ‘mucosal origins hypothesis’, which says that RA originates at one or more mucosal sites and then is transmitted to other organs. However, the hypothesis is not able to identify as to what stage of the development of RA this mucosal origination occurs. It is known that the gastrointestinal tract is the largest mucosal barrier, and is an essential getaway for the host to take in nutrition from food and drinks (24, 25). Up to 100 trillion symbiotic microbes live in the gut, and these microbes are involved in the digestion, metabolism, nutrition, disease control, and maintenance of general well-being (26, 27). The imbalance of gastrointestinal homeostasis, especially dysbiosis in gut microbiota composition and diversity, results in inflammatory responses and further triggers the occurrence of diseases such as RA (28, 29). Probiotics are a potential adjuvant to restore their original intestinal balance and behavior, and to modulate both innate and adaptive immunity in the host. The use of probiotics prevents or ameliorates the early stage of disease, and may be an effective therapy for chronic inflammatory diseases (30).
In this review, we summarize recent literature to help better understand the role of gut microbiome in RA, and to evaluate the therapeutic and preventative effects of probiotics in RA, providing a solid framework for future studies of optimizing the management of the health status of patients with RA.
The gut microbiota has gained more and more accentuated attention from specialists as a key contributor to the development and progression of RA (31, 32). In a study, fecal samples from RA patients were transplanted to germ-free mice, the result showed that mice inoculated microbiota from RA patients had an increased level of Th17 cells in the gut and an increased risk of developing severe arthritis (33). With continuous research, it has been found that perturbations of gut microbiota have been observed at different stages of RA, such as pre-clinical RA and established RA. In addition, these perturbations of gut microbiota are able to affect the host`s immune response (Figure 1).
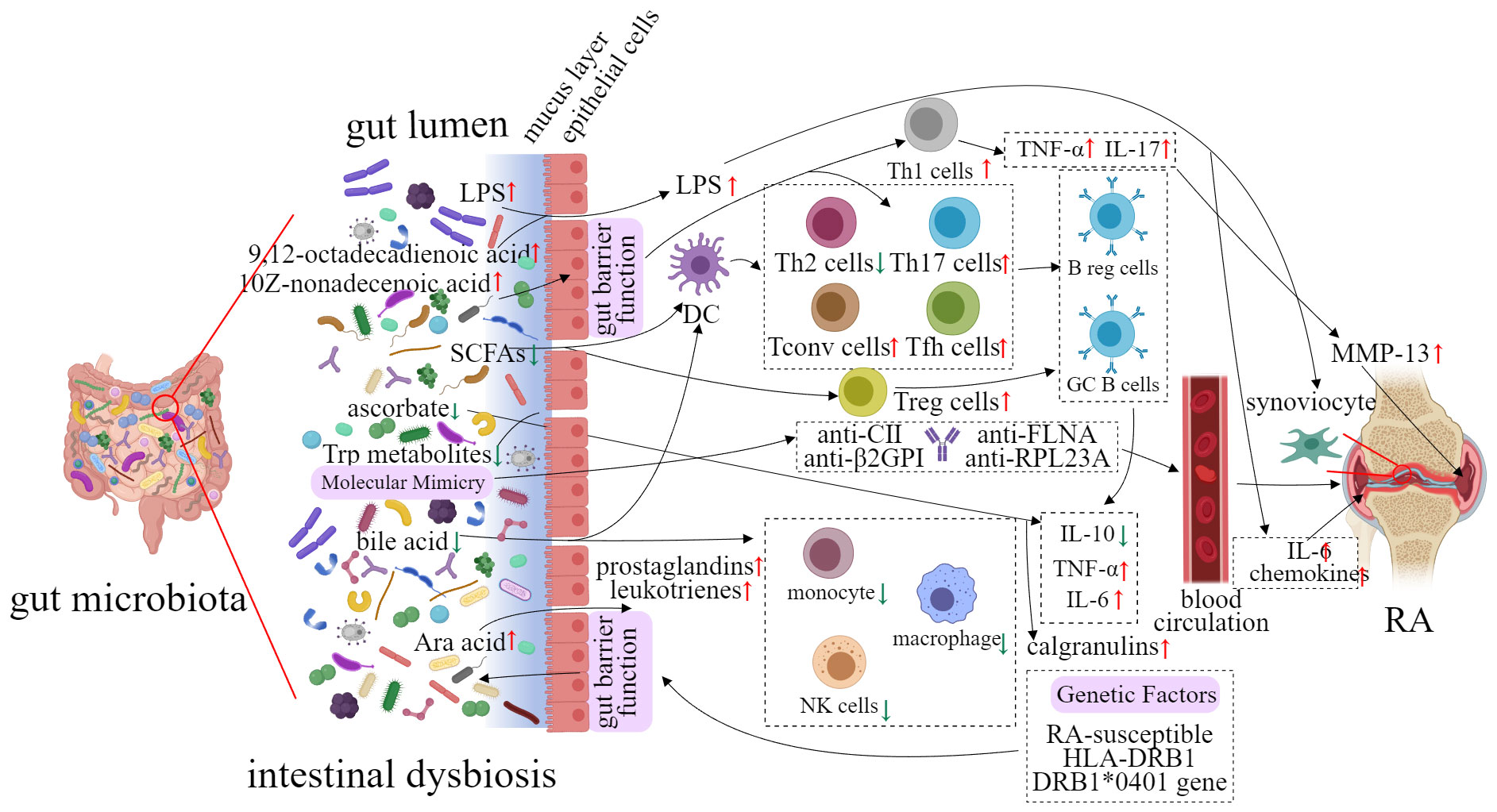
Figure 1 Potential implication of gut microbiota in the etiology of RA.
Rogier and his colleagues reported that the gut microbiota had a significant alteration in the pre-clinical stage of collagen-induced arthritis (CIA) mice (34). In pre-clinical RA mice, the abundance of the phylum Bacteroidete (e.g. S24-7 and Bacteroidaceae) was decreased, while Firmicutes and Proteobacteria (e.g. Ruminococcaceae, Lachnospiraceae, and Desulfovibrinocaceae) were increased (34). Interleukin (IL) 17-producing T helper 17 cell counts and the severity of RA was reduced when the gut microbiome was eliminated in established RA mice (34). In a study on anti-citrullinated protein (anti-CCP) positive individuals, researchers have sequenced the 16S rRNA gene of their gut microbiota (35). The abundance of Lachnospiraceae, Helicobacteraceae, Ruminococcaceae, Erysipelotrichaceae, and Bifidobacteriaceae were significantly increased in the intestine of anti-CCP-positive individuals (35). Furthermore, five out of twenty-five anti-CCP-positive individuals have progressed to RA during this trial (35).
A clinical trial involving 32 patients with RA and 30 healthy controls revealed significant differences in intestinal bacteria composition. Lactobacteria were found to be significantly decreased in RA patients (P < 0.05), whereas Enterococci and Clostridia were significantly increased (P < 0.05). In addition, the proportion of Bifidobacteria, Bacteroids, and Lactopositive colibacteria was observed to be reduced (P < 0.05), while the abundance of opportunistic Enterobacteria and Staphylococci showed increased (P < 0.05) (36). It is worth noting that opportunistic Enterobacteriaceae in urine and nasal mucosa were also detected (P < 0.05). This result implied the possible fact that opportunistic Enterobacteriaceae translocated from the intestines to other organs (36). In another cohort of individuals with RA and healthy subjects (at the age of 18-65), Zhang and his colleagues analyzed the change of microbiome in fecal, dental, and salivary samples respectively from RA patients through metagenomic sequencing and metagenome-wide association study (22). The results showed that Haemophilus spp. were depleted, while Lactobacillus salivarius was over-represented in the gut of RA patients (P < 0.05, Z-score≥1.6, 90% confidence). Similar findings were observed in their dental and salivary samples (22).
The gut microbiota plays an essential role in the maturation and development of the gut-associated lymphoid tissue (GALT) (37). GALT is composed of separate or aggregated lymphoid follicles, which form Peyer’s patches (PPs). PPs are considered to be an immune sensor in the gut, capable of controlling antigens and bacteria in the lumen area (38). Moreover, the interaction between follicle-associated epithelium and immune cells in PPs is associated with immune defense and immunologic tolerance (38). The lack of gut microbiota leads to a decreased level of mesenteric lymph nodes (MLNs) and PPs, as well as a reduced number of immune cells (39, 40). However, when gut microbiota is colonized in the intestines, the immune system can be rebuilt (41).
Faecalibacterium prausnitzii is one of the important commensal bacteria in the human gut microbiota, accounting for 3-5% of the total number of bacteria detected in stool samples of healthy people (42). A decrease in the abundance of F. prausnitzii in the intestine may be a sign of dysregulation such as the reduced capacity of self-defense against inflammatory reactions (43, 44). Furthermore, metabolites produced by the intestinal flora also influence intestinal immunity. Bacteroidetes fragilis, a symbiotic bacterium in humans, belongs to the phylum Bacteroidetes, which is one of the four major phyla of intestinal bacteria viz. Bacteroidetes, Firmicutes, Proteobacteria, and Actinobacteria. Polysaccharide A, secreted by Bacteroides fragilis, induces CD4+ T cells to transform into Foxp3+ regulatory T cells (Tregs) that produce IL-10 (45). Toll-like receptor 2 signaling is active by Tregs and IL-10 (45).
The above studies have demonstrated that the intestinal commensal microbiota is able to regulate T cell and Treg responses thereby protecting the host from pathogens. Intestinal commensal bacteria are crucial for the establishment of a regular innate immune system (46). As the human body’s first line of defense, innate immune responses rely on a family of receptors named pattern recognition receptors (PRRs). PRRs, especially Toll-like receptors (TLRs), are able to recognize pathogen-associated molecular patterns (PAMPs), and play a vital role in immune defense (47). TLRs are recognized by both gram-positive and gram-negative bacteria to regulate innate and adaptive immunity (48). Gram-positive bacteria, gram-negative bacteria, and their related PAMPs are identified by TLR-2 and TLR-4, respectively (49). TLR-2 pathway is activated by peptidoglycans and polysaccharides of gram-positive bacteria`s cytoderm. TLR-4 is bound to lipopolysaccharide of gram-negative bacteria (50). However, the imbalance of gut microbiota is likely to cause the alteration of the innate immune system. These alterations allow microbes and their metabolites to cross into the lamina propria and sub-epithelial spaces, which is related to the risk and severity of the disease such as RA (51).
Many studies suggest a potential link between RA development and intestinal dysbiosis (46, 52–56). Recent clinical studies have demonstrated that alterations in gut microbiota and their metabolite concentrations occurred before RA onset (57–61). These alterations activate autoreactive T cells and may induce systemic inflammation (33). The potential mechanism linking gut microbiota to RA will be elaborated below.
The mechanism of molecular mimicry has been known for a long time (62), but only in recent years have we begun to focus on the molecular mimicry of microbes that coexist with humans. Studies have revealed that human microorganisms express and produce some homologous proteins similar to host proteins, and these proteins may cause host immune imbalance under some circumstances, resulting in the development of autoimmune diseases (63–65).
Citrullination is a normal physiological process, involved in various physiological functions such as cell apoptosis, end-stage differentiation, gene regulation, and reproductive development. Peptidyl arginine deiminase (PAD) catalyzes the conversion of peptidyl arginine to citrulline in the involvement of Ca2+ (66). However, PAD secreted by Porphyromonas gingivalis (PPAD) enables catalyzed citrullination without Ca2+ (67). Porphyromonas gingivalis is the main cause of periodontal disease, but it is also highly correlated with RA (68). Patients with periodontal disease have a two-fold increased risk of RA (69). Severity of periodontitis is related to the severity of RA (70). PPAD can catalyze the citrullination of not only P. gingivalis`s proteins but also a variety of proteins such as vimentin, fibrinogen, and α-enolase, which are RA-specific autoantigens. Further studies also found that anti-CCP proteins were detectable in RA patients (71). The presence of citrullinated proteins was observed in the synovium of other inflammatory arthritis such as osteoarthritis, but there is no anti-CCP (72). These results suggested that citrullinated proteins may not be specific for RA, but the immune response against citrullinated proteins is unique to RA. This further illustrates that recognition of citrullinated proteins by autoreactive T cells and production of ACPAs by B cells may be even more important for RA. In addition to P. gingivalis, the others are also closely related to molecular mimicry and RA (Table 1).
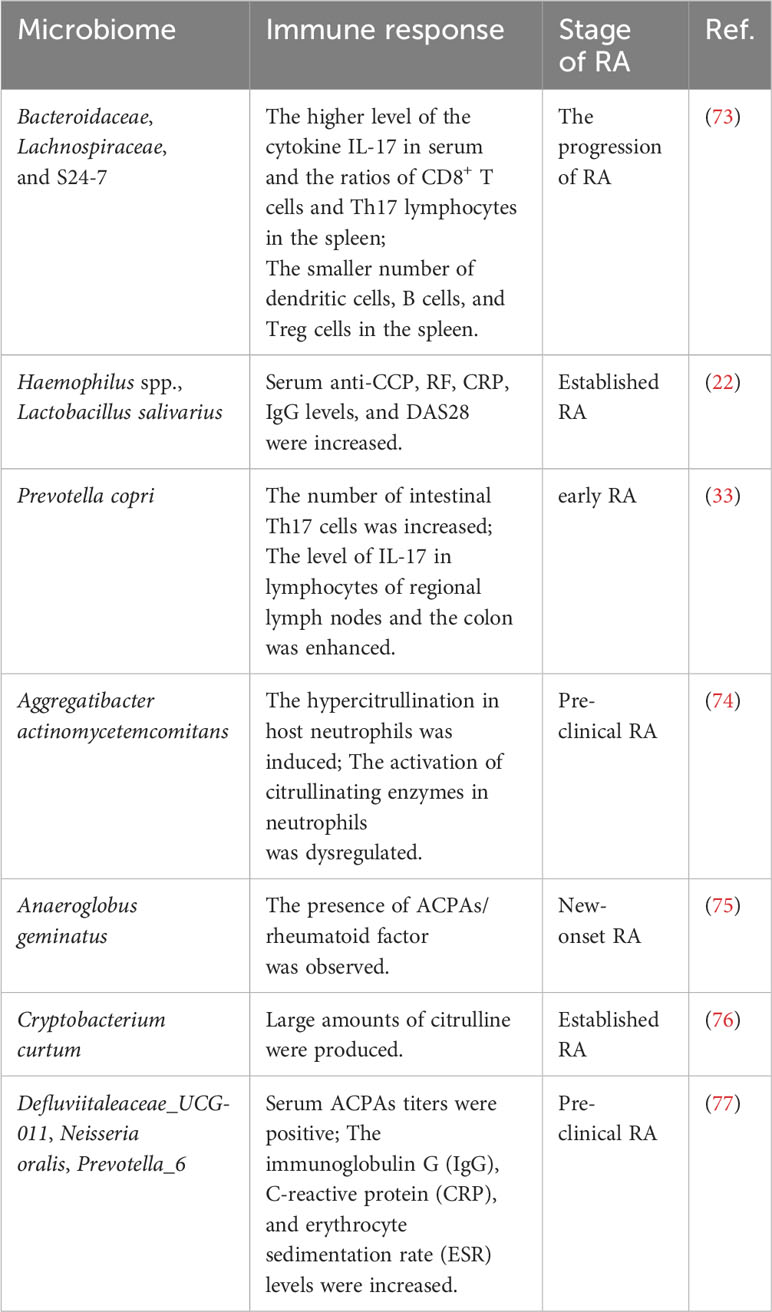
Table 1 Molecular mimicry of microbiome in RA.
Human leukocyte antigen (HLA) genes have been found to be associated with RA (78). It was found that the majority of RA patients share conserved amino acid sequences in the third hypervariable region (HV3) of the HLA DR molecule, which has been termed a ‘‘shared epitope’’ (SE) (14, 79). The SE-coding HLA-DRB1 alleles are the most significant genetic risk factor associated with RA (80, 81). It is worth noting that a cross-sectional study of 1650 TwinsUK participants showed that Prevotella spp was significantly associated with the RA polygenic risk score. And SCREEN-RA participants (n=133) carried established SE risk alleles. These results implied that Prevotella spp in the gut microbiota was associated with RA genotype before pre-clinical RA (58).
The mechanism underlying the role of the SE in RA remains unclear. There is a hypothesis that the SE may act as a signal transduction ligand that can trigger innate immune signaling (78). The hypothesis is based on the fact that the SE is located near the apex of α helical tri-dimensional structural motif of the major histocompatibility complex (MHC) gene family. The MHC, a group of genes, encodes major histocompatibility antigens in animals. The human MHC is named HLA. It is known that products coded by the MHC gene family have tri-dimensional homology (82). The top of α helical tri-dimensional structural motif seems to be enriched in signal transduction ligands. Therefore, it is presumed that SE is similar to class I MHC-coded molecules, resulting in stimulating innate immune responses (78). Studies show that SE is bound to the P-domain of calreticulin (CRT, a known innate immunity receptor) on the cell surface (83). The amino acid residues 217-224 of the CRT P-domain are a potential SE binding site (84). Further investigations have revealed that the SE-CRT pathway is involved in the activation of NO synthase and the production of reactive oxygen species (ROS), consequently leading to the inducing osteoclast-mediated bone destruction (85–87).
It is speculated that SE activates immune dysregulation. CRT is expressed on the surface of many cells including dendritic cells (DCs) (88). Apoptotic cells determine whether a pro- or anti-inflammatory reaction is activated in the host, whereas CRT plays a pivotal role in the clearance of apoptotic cells (89). This makes CRT play an important role in the junction between immunologic tolerance and autoimmunity (90). As professional antigen-presenting cells, DCs also induce tolerance mediated partly by indoleamine 2,3-dioxygenase (IDO) which is an enzyme that catalyzes the catabolism of tryptophan (91, 92). Thus, SE activates immune dysregulation by inhibiting the activity of IDO in DCs (93). In addition, a SE ligand stimulates the production of IL-6 and the differentiation of Th17 (94, 95). The above studies have gradually demonstrated that SE could affect RA pathogenesis.
The definition of probiotics is “Live microorganisms which when administered in adequate amounts confer a health benefit on the host.” (96). Probiotics exert various effects on the body, with different probiotics functioning in distinct ways. One possible way involves probiotics aiding the host in maintaining a healthy microbiome, as well as assisting in the restoration of intestinal microbial balance after dysbiosis. Additionally, probiotics have the ability to produce bioactive substances that elicit the desired effects and influence the immune responses of the host.
Probiotics have been studied in animal and human trials to assess their potential positive effects on the prevention and treatment of RA (Table 2). Probiotics such as Lactobacillus and Bifidobacterium have been widely investigated. In a study involving rats, different Lactobacillus species were orally administered two weeks before the induction of collagen-induced arthritis (CIA). The results demonstrated that different types of Lactobacillus exerted varying effects on RA. L. reuteri, L. casei, L. rhamnosus, and L. fermentum were found to alleviate RA by inhibiting species-specific pro-inflammatory cytokine and anti-CII antibody signaling pathways and regulating the balance of microflora and metabolites (e.g., short-chain fatty acids) through species-dependent immune regulation in the gut (106). In particular, L. reuteri and L. casei had a decreased effect on Th1 immune responses, whereas L. rhamnosus and L. fermentum had a decreased effect on Th17 immune responses. L. plantarum reduced both Th1 and Th17 immune responses, but it did not attenuate RA. Conversely, L. salivarius only postponed the onset time of RA without inducing the immune response (106). In a randomized and double-blind clinical trial, 46 RA patients were provided a daily capsule of L. casei 01 for 8 weeks. The serum level of pro-inflammatory cytokines (tumor necrosis factor-α, interleukin-6, interleukin-12) was significantly decreased. The serum regulatory cytokine (interleukin-10) was increased. The disease activity and inflammatory status of these participants were significantly decreased by L. casei 01 supplementation (107). Similarly, oral administration of L. rhamnosus GR-1 and L. reuteri RC-14 was found to improve the Health Assessment Questionnaire (HAQ) score of active RA patients and achieved an American College of Rheumatology (ACR) 20 response (108).
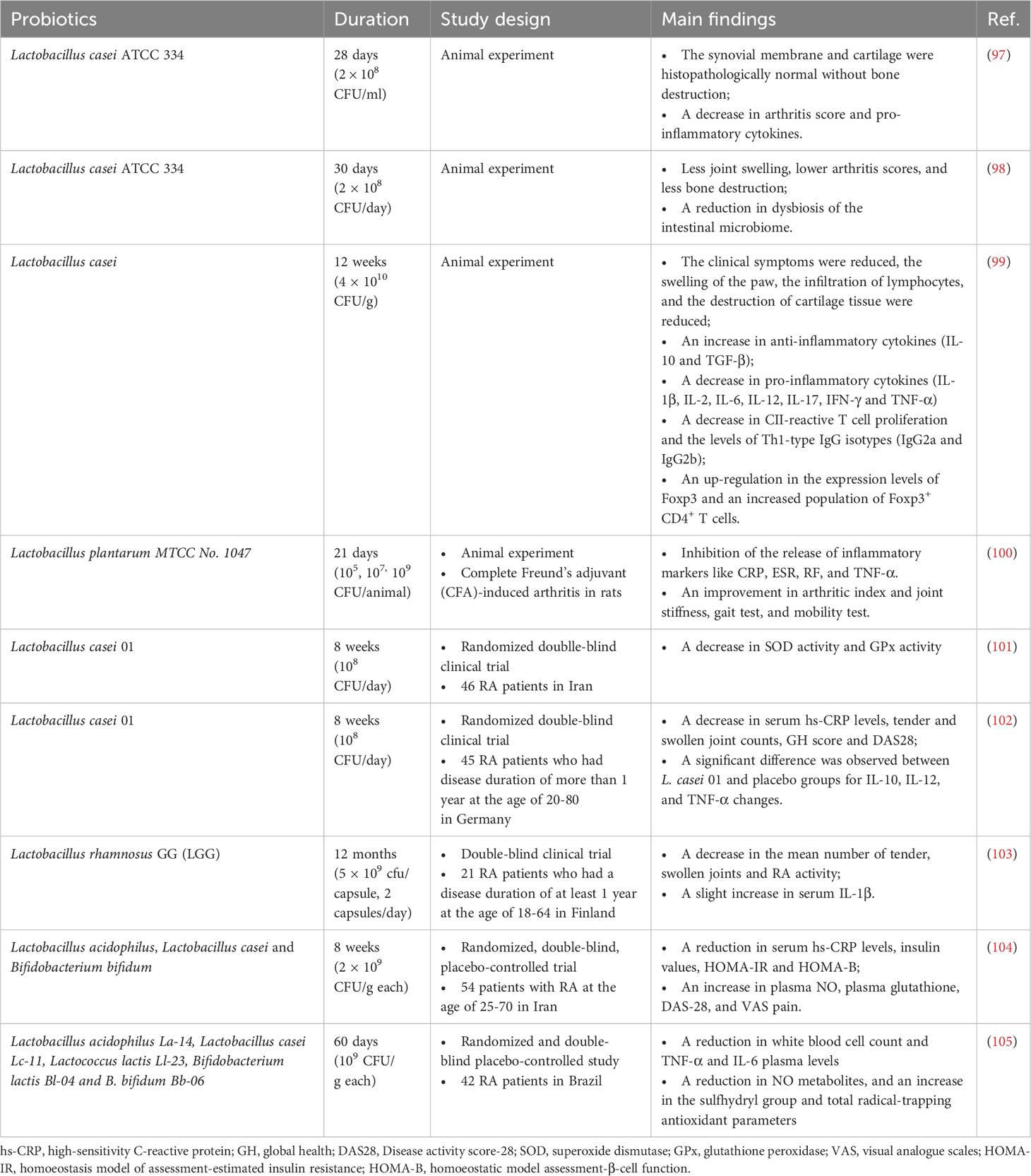
Table 2 List of human and animal studies on the therapeutic effect of probiotics.
Bifidobacterium species are also often studied for their effects on RA. In an early intervention experiment, five strains of Bifidobacterium adolescentis including HuNan2016-7-2, AHWH4-M1, FSDJN3Q1, M1DZ09M1, and FSDJN12W5 were given to CIA rats respectively by gavage. The clinical symptoms of CIA rats were alleviated, and the balance between pro- and anti-inflammatory responses was restored. Furthermore, the dysbiosis in the gut microbiota of the CIA rats was also rectified (109). In a pre-clinical model, intervention with B. longum RAPO in CIA mice, obese CIA, and humanized avatar model ameliorated the symptoms of RA such as reduced RA incidence, arthritis score, inflammation, bone damage, and cartilage damage. Additionally, B. longum RAPO may play a role in ameliorating RA via inhibiting the secretion of IL-17 and other pro-inflammatory mediators, suggesting its potential role in ameliorating RA (110). Furthermore, three strains containing L. acidophilus, L. casei, B. bifidum had beneficial effects on RA patients. The Disease Activity Score of 28 joints (DAS-28) of RA patients who were given probiotic capsules was improved. The homeostatic model assessment-B of cell function and serum high-sensitivity C-reactive protein concentrations had a significant decrease (111). In a patent, a newly isolated strain called Bifidobacterium bifidum ATT was found to be an effective composition for alleviating, preventing, or treating RA (112).
Prevotella is a newly discovered species that shares taxonomic similarities with Prevotella nigra and Prevotella villi. In a study by Marietta and colleagues (113), arthritis-susceptible HLA-DQ8 mice were treated with P. histicola, resulting in a significant decrease in the incidence and severity of arthritis in DQ8 mice. Moreover, the study observed a suppression of antigen-specific Th17 responses, an increase in the transcription of IL-10, and an upregulation of antimicrobial peptidase and tight junction proteins (113). On the other hand, it is possible that P. histicola colonization in the duodenum contributes to the restoration of Allobaculum population in the host’s gut. This colonization could potentially lead to an increase in butyrate production, and consequently higher levels of short-chain fatty acids (SCFAs). These elevated levels of SCFAs could potentially contribute to the maintenance of immune homeostasis (114).
Bacillus probiotics are a type of probiotic bacteria isolated from the human intestine, with Bacillus coagulans being one of the most common strains, known for its ability to produce lactic acid. In a double-blind trial, 45 patients with RA who were administered B. coagulans GBI-30, 6086 for 60 days showed a significant improvement in the Patient Pain Assessment score and Pain Scale compared to the placebo group. Moreover, some participants also demonstrated enhanced mobility, as evidenced by their ability to walk 2 miles and engage in daily activities (115). Additionally, B. coagulans also generates SCFAs including butyric acid, which is recognized for its role in promoting gut cell health and healing, as well as its involvement in regulating the mucosal immune system (116). In an in vivo experiment, arthritic rats were orally administered a combination of probiotic B. coagulans and prebiotic inulin to evaluate their possible influence on RA. The results indicated that pretreatment with B. coagulans and inulin significantly inhibits the production of serum amyloid A and fibrinogen, as well as the development of paw swelling in arthritic rats. Notably, the combination probiotic group displayed a significant reduction in pro-inflammatory cytokines, such as TNF-α (117).
The underlying mechanisms through which probiotics exert their anti-inflammatory and immunomodulatory effects are not yet fully understood. However, it has been observed that probiotics can regulate systemic inflammation through both direct and indirect pathways, ultimately leading to a reduction in RA disease activity (118) (Figure 2). In the direct pathway, specific probiotics have been shown to have an impact on various cells associated with innate and acquired immunity, including epithelial cells, macrophages, dendritic cells (DCs), natural killer cells (NKs), T cells, and B cells (119). The expression of PRRs on immune and non-immune cells (such as NK cells, DCs, macrophages, fibroblasts, and epithelial cells) is decreased by specific probiotics. Antigen-presenting cells (APCs) and Th cells are decreased. Th cells are also inhibited by the increase of IL-4 that is secreted by B cells (120). The pro-inflammatory cytokines such as IL-1β, TNF-α, IL-6, IL-17, and IL-23 are suppressed by anti-inflammatory cytokines IL-10 that is produced by APCs, resulting in the decrease of systemic inflammation (121). Furthermore, the increasing differentiation of B cells into IgA-secreting plasma cells leads to a restoration of the intestinal barrier function and a suppression in the bacterial adhesion to the epithelium. In the indirect pathway, probiotics are believed to promote the production of microbial metabolites such as SCFAs, tryptophan, adenosine, and histamine (122). This can lead to competition with pathogenic bacteria, reducing the likelihood of pathogenic microorganisms entering the system. Probiotics also stimulate mucosal secretion and promote the expression of tight junction proteins, further restoring the function of the intestinal barrier (123). Furthermore, microbial metabolites also play a role in regulating gut microbiota and changes in microbiota can impact the production of these microbial metabolites. Overall, these mechanisms contribute to the restoration of the balance of intestinal microbiomes and a reduction in systemic inflammation, ultimately leading to a decrease in RA disease activity (124).
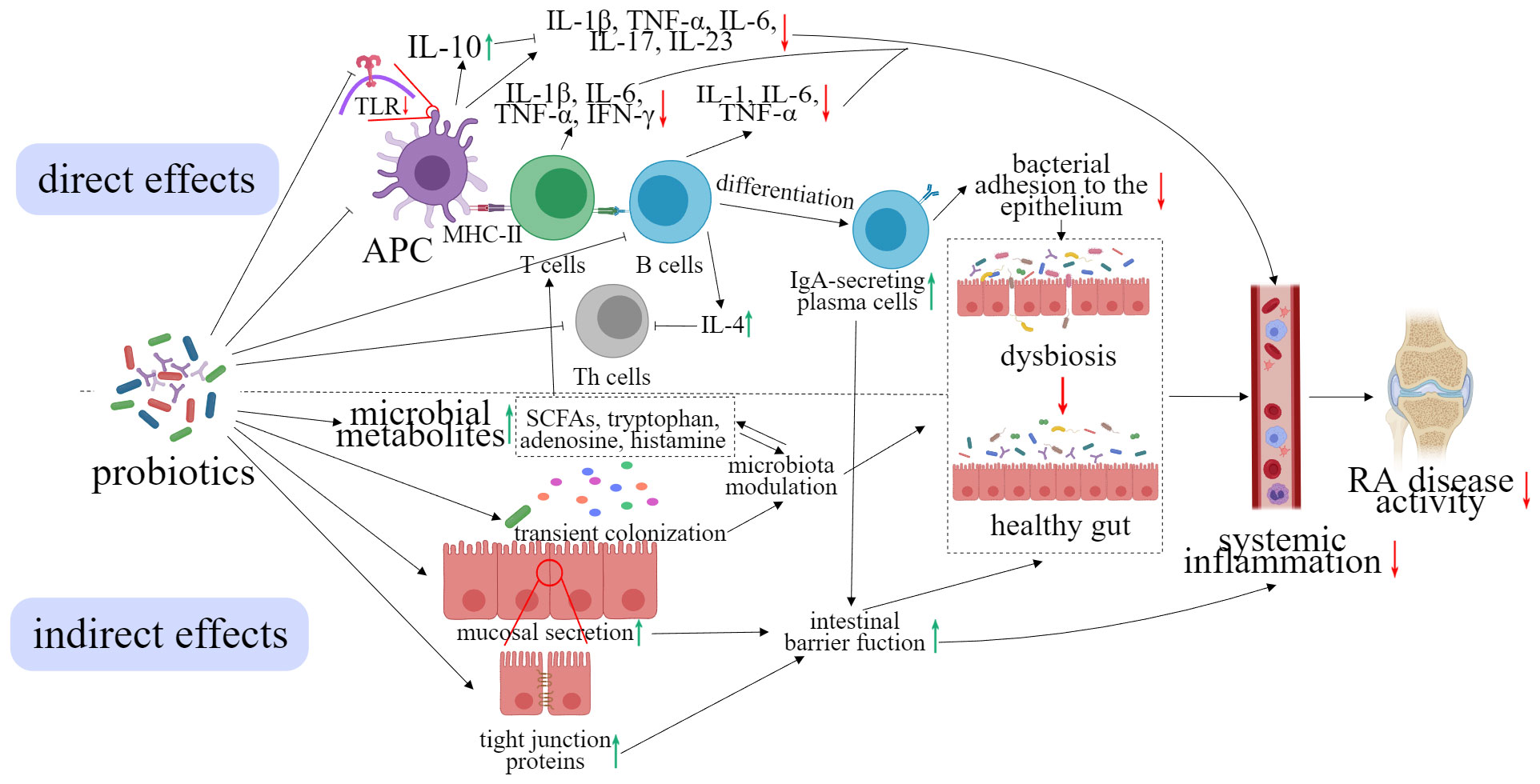
Figure 2 Effect of probiotics on the reduction of the symptoms of RA.
SCFAs are one of the most abundant metabolites of gut microbiota and play a role in modulating the immune system. Specifically, SCFAs have been shown to inhibit the differentiation of DCs (125) and alter the expression of cytokines by DCs (126). Additionally, SCFAs such as butyrate and propionate can induce apoptosis and affect the phagocytic capacity of neutrophils (127). Moreover, butyrate, acetate, and propionate have been found to down-regulation pro-inflammatory mediators in macrophages, that are induced by lipopolysaccharides (LPS) (128, 129). SCFAs also have a positive impact on intestinal barrier function by upregulating the expression of tight junction proteins (130). In addition, butyrate has been shown to suppress the secretion of inflammatory cytokines and inhibit RA through its regulation of Treg cells (131). This connection between SCFAs and their immunomodulatory effects has established their significance in the context of RA.
The etiology and pathogenesis of RA remain unresolved, making the search for highly effective treatment methods, preventive measures, and specific medicine an ongoing area of exploration (132). The primary objective in treating RA is to decrease joint inflammation, hinder the progression of lesions and irreversible bone damage, and safeguard the functionality of joints and muscles. The ultimate goal is to attain complete remission of the disease. Overall, RA exhibits a slow onset and chronic course, which can manifest varying degrees of difficulties and pain in daily life and work (133). However, with early diagnosis and prompt, comprehensive treatment, most patients can achieve varying degrees of effectiveness in alleviating pain and controlling disease progression (4).
Based on previous studies, probiotic treatment can alleviate symptoms of RA, but the underlying mechanism remains unknown. Several possible pathways have been proposed, including molecular mimicry (134), shared epitope (95), intestinal immune responses (6), metabolites produced by the intestinal flora (124), and compromised intestinal barrier function (135) associated with gut microbiota dysbiosis. To validate these pathways further research is needed. For instance, the ‘mucosal origins hypothesis’ says that inflammation is transmitted from one mucosal site to further ones, and it is necessary to investigate whether a single site is sufficient to trigger systemic spread through additional studies. The relationship between local mucosal findings and systemic immune changes should also be further examined (6). Additionally, it is important to evaluate T cell antigen-specific or innate immune reactivity systemically and at mucosal sites. Thus, ongoing research, such as APIPPRA (136) and TREAT EARLIER (137), has extended their observation period to up to 5 years to assess the long-term sustainability of the preventive effects of RA (138, 139). These long-term data will be crucial for a comprehensive understanding of the mechanisms underlying RA development and for reducing the disease burden in at-risk individuals.
A probiotic with optimal performance must undergo a series of processes including isolation, screening, evaluation, and final application, production, and promotion. These processes encompass various research aspects, such as basic research, functional research, clinical research, and industrialization, necessitating robust research and development capabilities. Advanced technologies like genomics, transcriptomics, and proteomics should be employed to analyze the molecular mechanisms behind strains’ environmental tolerance, intestinal adaptation, and probiotic function (140). Nevertheless, it is crucial to note that the benefits of probiotics are contingent upon the specific strain. Consequently, each strain must undergo a time-consuming development process, presenting challenges to the widespread implementation of probiotics. Fortunately, with the continuous emergence and advancement of new technologies, the comprehensive study of probiotics is expected to become less daunting.
A number of reports have demonstrated the various effects of probiotics on digestive tract health, immune health, and metabolic health (140). However, several issues arise. For instance, researchers have persistently faced the challenge of insufficient knowledge regarding probiotic strains and dosage. Due to the availability of probiotics without a prescription, many individuals begin or discontinue their usage without consulting medical professionals or seeking specialized advice (1). Enterprises often exaggerate the efficacy of probiotics when marketing them, leading to consumers experiencing different outcomes after consumption, thereby hindering the overall development of the probiotics industry. Furthermore, although research on probiotics is rapidly expanding, the availability of concrete evidence elucidating mechanisms, particularly the key substances and the reasons for variations between strains remains limited. Undoubtedly, this presents the most critical and difficult obstacle in probiotic research and development. We anticipate that this challenge will be overcome soon, paving the way for more possibilities in probiotic applications.
An important feature of probiotics is their ability to stabilize gut microbiota homeostasis and regulate immune responses, which has led to an increase in their application among patients worldwide. While traditional treatment for RA plays a fundamental role in its management and is typically a lifelong treatment, the long-term accumulation of multiple drugs often leads to adverse effects for patients. Consequently, researchers have been investigating a new therapy that targets intestinal bacteria through the use of probiotics in order to prevent or treat RA. As discussed in this review, the gut microbiota is closely associated with the development and progression of RA. Therefore, it is crucial to identify the causal relationship between the human microbiome and the pathogenesis of RA, as this knowledge is essential for improving treatment outcomes and restoring the health of patients. The manipulability of the gut microbiota makes it possible to locally or systematically manipulate the abundance of specific gut microbiota associated with diseases through interventions, potentially changing the treatment landscape for patients with RA. If proven feasible, this approach could lead to the development of entirely new prevention or treatment strategies in the clinical field. Furthermore, animal and clinical trials have demonstrated that probiotics supplementation ameliorates the symptoms of RA by regulating dysbiosis in the gut microbiota and modulating immune responses. Despite the various challenges that remain in probiotic research, such as determining targets, pathways, and mechanisms, the numerous benefits they offer to the human body continue to drive researchers in their quest for further exploration.
YY: Writing – review & editing. QH: Project administration, Writing – review & editing. XZ: Resources, Supervision, Writing – review & editing. ZL: Funding acquisition, Methodology, Supervision, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by the Shanghai Super post-doctoral Incentive Program (grant number 2021093); the Shanghai State-owned Assets Supervision and Administration Commission Enterprise Innovation Development and Capacity Enhancement Program (No. 2022013); the National Key R&D Program of China (2022YFD2100704).
The authors are thankful to the State Key Laboratory of Dairy Biotechnology, Shanghai Engineering Research Center of Dairy Biotechnology, Bright Dairy & Food Co., Ltd., Shanghai, China for providing the various resources for the completion of the current article.
Authors YY, QH, and ZL were employed by the company Bright Dairy & Food Co., Ltd.
The remaining author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Bungau SG, Behl T, Singh A, Sehgal A, Singh S, Chigurupati S, et al. Targeting probiotics in rheumatoid arthritis. Nutrients. (2021) 13(10):3376. doi: 10.3390/nu13103376
2. Xu B, Lin J. Characteristics and risk factors of rheumatoid arthritis in the United States: an NHANES analysis. PeerJ. (2017) 5:e4035. doi: 10.7717/peerj.4035
3. Sepriano A, Kerschbaumer A, Smolen JS, van der Heijde D, Landewé R. Safety of synthetic and biological DMARDs: a systematic literature review informing the 2019 update of the EULAR recommendations for the management of rheumatoid arthritis. Ann Rheum Dis. (2020) 79(6):760–70. doi: 10.1136/annrheumdis-2019-216653
4. Fraenkel L, et al. 2021 American College of Rheumatology Guideline for the Treatment of Rheumatoid Arthritis. (2021) 73 (3): 924–939
5. Finckh A, Liang MH, Herckenrode CMV, Pablo PD. Long-term impact of early treatment on radiographic progression in rheumatoid arthritis: A meta-analysis. Arthritis & Rheumatology. (2006) 55(6):864–72. doi: 10.1002/art.22353
6. Holers VM, Kristen DM, Kuhn KA, Buckner JH, Robinson WH, Yuko O, et al. Rheumatoid arthritis and the mucosal origins hypothesis: protection turns to destruction. Nat Rev Rheumatol. (2018) 14(9):542–57. doi: 10.1038/s41584-018-0070-0
7. Smolen JS, Aletaha D, Barton A, Burmester GR, Emery P, Firestein GS, et al. Rheumatoid arthritis. Nat Rev Dis Primers. (2018) 4(1):18001. doi: 10.1038/nrdp.2018.1
8. McInnes IB, Schett G. The pathogenesis of rheumatoid arthritis. New England Journal of Medicine. (2011) 365(23):2205–19. doi: 10.1056/NEJMra1004965
9. Ziff M. Relation of cellular infiltration of rheumatoid synovial membrane to its immune response. Arthritis Rheum. (1974) 17:313–9. doi: 10.1002/art.1780170317
10. Kiener HP, Niederreiter B, Lee DM, Jimenezboj E, Smolen JS, Brenner MB. Cadherin 11 promotes invasive behavior of fibroblast-like synoviocytes. Arthritis Rheum. (2009) 60(5):1305–10. doi: 10.1002/art.24453
11. Redlich K, Smolen JS. Inflammatory bone loss: pathogenesis and therapeutic intervention. Nat Rev Drug Discovery. (2012) 11:234–50. doi: 10.1038/nrd3669
12. Harre U, Georgess D, Bang H, Bozec A, Axmann R, Ossipova E, et al. Induction of osteoclastogenesis and bone loss by human autoantibodies against citrullinated vimentin. J Clin Invest. (2012) 122(5):1791–802. doi: 10.1172/JCI60975
13. Krishnamurthy A, Joshua V, Hensvold AH, Jin T, Sun M, Vivar N, et al. Identification of a novel chemokine-dependent molecular mechanism underlying rheumatoid arthritis-associated autoantibody-mediated bone loss. Ann Rheum Dis. (2016) 75(4):721–9. doi: 10.1136/annrheumdis-2015-208093
14. Gregersen PK, Silver J, Winchester RJ. The shared epitope hypothesis. an approach to understanding the molecular genetics of susceptibility to rheumatoid arthritis. Arthritis & Rheumatism. (1987) 30(11):1205–13. doi: 10.1002/art.1780301102
15. Nepom GT, Nepom BS. Prediction of susceptibility to rheumatoid arthritis by human leukocyte antigen genotyping. Rheumatic Dis Clinics North America. (1992) 18:785–92. doi: 10.1016/S0889-857X(21)00151-4
16. Karlson EW, Deane K. Environmental and gene-environment interactions and risk of rheumatoid arthritis. Rheumatic Disease Clinics of North America. (2012) 38(2):405–26. doi: 10.1016/j.rdc.2012.04.002
17. Hensvold AH, Magnusson PKE, Joshua V, Hansson M, Israelsson L, Ferreira RC, et al. Environmental and genetic factors in the development of anticitrullinated protein antibodies (ACPAs) and ACPA-positive rheumatoid arthritis: an epidemiological investigation in twins. Annals of the Rheumatic Diseases: A Journal of Clinical Rheumatology and Connective Tissue Research. (2013) 74(2):375–80. doi: 10.1136/annrheumdis-2013-203947
18. Van Zanten A, Arends S, Roozendaal C, Limburg PC, Maas F, Trouw LA, et al. Presence of anticitrullinated protein antibodies in a large population-based cohort from the Netherlands. (2017) 76(7):1184–90. doi: 10.1136/annrheumdis-2016-209991
19. Stolt P, Källberg H, Lundberg I, Sjögren B, Klareskog L, Alfredsson L, et al. Silica exposure is associated with increased risk of developing rheumatoid arthritis: results from the Swedish EIRA study. Annals of the Rheumatic Diseases. (2005) 64(4):582. doi: 10.1136/ard.2004.022053
20. Scher JU, Joshua V, Artacho A, Abdollahi-Roodsaz S, Ockinger J, Kullberg S, et al. The lung microbiota in early rheumatoid arthritis and autoimmunity. Microbiome. (2016) 4(1):60. doi: 10.1186/s40168-016-0206-x
21. Scher JU, Andrew S, Longman RS, Nicola S, Carles U, Craig B, et al. Expansion of intestinal Prevotella copri correlates with enhanced susceptibility to arthritis. Elife. (2013) 2(1629):e01202. doi: 10.7554/eLife.01202
22. Zhang X, Zhang D, Jia H, Feng Q, Wang D, Liang D, et al. The oral and gut microbiomes are perturbed in rheumatoid arthritis and partly normalized after treatment. Nat Med. (2015) 21(8):895–905. doi: 10.1038/nm.3914
23. Chen J, Wright K, Davis JM, Jeraldo P, Marietta EV, Murray J, et al. An expansion of rare lineage intestinal microbes characterizes rheumatoid arthritis. Genome Med. (2016) 8(1):43. doi: 10.1186/s13073-016-0299-7
24. Ramezani A, Raj DS. The gut microbiome, kidney disease, and targeted interventions. (2014) 25(4):657–70. doi: 10.1681/ASN.2013080905
25. Dias Bastos PA, Lara Santos L, Pinheiro Vitorino RM. How are the expression patterns of gut antimicrobial peptides modulated by human gastrointestinal diseases? A bridge between infectious, inflammatory, and Malignant diseases. Journal of Peptide Science. (2018) 24(3):e3071. doi: 10.1002/psc.3071
26. Boulangé CL, Neves AL, Chilloux J, Nicholson JK, Dumas ME. Impact of the gut microbiota on inflammation, obesity, and metabolic disease. Genome Med. (2016) 8(1):42. doi: 10.1186/s13073-016-0303-2
27. Kau AL, Ahern PP, Griffin NW, Goodman AL, Gordon JI. Human nutrition, the gut microbiome and the immune system. Nature. (2011) 474(7351):327–36. doi: 10.1038/nature10213
28. Bodkhe R, Balakrishnan B, Taneja V. The role of microbiome in rheumatoid arthritis treatment. Therapeutic Advances in Musculoskeletal Disease. (2019) 11:1759720X19844632. doi: 10.1177/1759720X19844632
29. Manasson J, Blank RB, Scher JU. The microbiome in rheumatology: Where are we and where should we go? Annals Of The Rheumatic Diseases. (2020) 79(6):727–33. doi: 10.1136/annrheumdis-2019-216631
30. Bedaiwi MK, Inman RD. Microbiome and probiotics: link to arthritis. Current Opinion in Rheumatology. (2014) 26(4):410–5. doi: 10.1097/BOR.0000000000000075
31. Xu L, Feng X, Tan W, Gu W, Guo D, Zhang M, et al. IL-29 enhances Toll-like receptor-mediated IL-6 and IL-8 production by the synovial fibroblasts from rheumatoid arthritis patients. Arthritis Res Ther. (2013) 15(5):R170. doi: 10.1186/ar4357
32. Thompson C, Davies R, Choy E. Anti cytokine therapy in chronic inflammatory arthritis. Cytokine. (2016) 86:92–9. doi: 10.1016/j.cyto.2016.07.015
33. Maeda Y, Kurakawa T, Umemoto E, Motooka D, Ito Y, Gotoh K, et al. Dysbiosis contributes to arthritis development via activation of autoreactive T cells in the intestine. Arthritis Research & Therapy. (2016) 68(11):2646–61. doi: 10.1002/art.39783
34. Rogier R, Evans-Marin H, Manasson J, Van dKPM, Walgreen B, Helsen MM, et al. Alteration of the intestinal microbiome characterizes preclinical inflammatory arthritis in mice and its modulation attenuates established arthritis. Sci Rep. (2017) 7(1):15613. doi: 10.1038/s41598-017-15802-x
35. Rooney CM, Mankia K, Mitra S, Moura IB, Wilcox MH. Perturbations of the gut microbiome in anti-CCP positive individuals at risk of developing rheumatoid arthritis. Rheumatology. (2020) 60(7):3380–7. doi: 10.1093/rheumatology/keaa792
36. Gul'neva M, Noskov SM. [Colonic microbial biocenosis in rheumatoid arthritis]. Klinicheskaia meditsina. (2011) 89(4):45–8.
37. Lamas B, Evariste L, Houdeau E. Dysregulation along the gut microbiota-immune system axis after oral exposure to titanium dioxide nanoparticles: A possible environmental factor promoting obesity-related metabolic disorders. Environ pollut. (2023) 330:121795. doi: 10.1016/j.envpol.2023.121795
38. Mohammadi-Motlagh H-R, Sadeghalvad M, Rezaei N. Organs and Tissues of the Immune System. In: Rezaei N, editor. Encyclopedia of Infection and Immunity. Elsevier, Oxford (2022). p. 14–23.
39. Smith K, McCoy KD, Macpherson AJ. Use of axenic animals in studying the adaptation of mammals to their commensal intestinal microbiota. Semin Immunol. (2007) 19:59–69. doi: 10.1016/j.smim.2006.10.002
40. Sommer F, Bäckhed F. The gut microbiota — masters of host development and physiology. Nat Rev Microbiol. (2013) 11(4):227–38. doi: 10.1038/nrmicro2974
41. Chung H, Pamp SJ, Hill JA, Surana NK, Edelman SM, Troy EB, et al. Gut immune maturation depends on colonization with a host-specific microbiota. Cell. (2012) 149(7):1578–93. doi: 10.1016/j.cell.2012.04.037
42. Breyner NM, Cristophe M, De SCS, Vilas BPB, Florian C, Azevedo VA, et al. Microbial Anti-Inflammatory Molecule (MAM) from Faecalibacterium prausnitzii Shows a Protective Effect on DNBS and DSS-Induced Colitis Model in Mice through Inhibition of NF-κB Pathway. Frontiers in Microbiology. (2017) 8:114. doi: 10.3389/fmicb.2017.00114
43. Zhang M, Qiu X, Zhang H, Yang X, Hong N, Yang Y, et al. Faecalibacterium prausnitzii inhibits interleukin-17 to ameliorate colorectal colitis in rats. PloS One. (2014) 9(10):e109146. doi: 10.1371/journal.pone.0109146
44. Quévrain E, Maubert MA, Michon C, Chain F, Marquant R, Tailhades J, et al. Identification of an anti-inflammatory protein from, Faecalibacterium prausnitzii, a commensal bacterium deficient in Crohn’s disease. Gut. (2016) 65(3):415. doi: 10.1136/gutjnl-2014-307649
45. Round JL, Mazmanian SK. Inducible Foxp3+ regulatory T-cell development by a commensal bacterium of the intestinal microbiota. Proceedings of the National Academy of Sciences. (2010) 107(27):12204–9. doi: 10.1073/pnas.0909122107
46. Jiao Y, Wu L, Huntington ND, Zhang X. Crosstalk between gut microbiota and innate immunity and its implication in autoimmune diseases. Frontiers in Immunology. (2020) 11:282. doi: 10.3389/fimmu.2020.00282
47. Bennike TB, Torkell E, Glerup H, Bonderup OK, Carlsen TG, Meyer MK, et al. A proteome analysis of rheumatoid arthritis gut mucosa. Journal of Proteome Research. (2016) 16(1):346–54. doi: 10.1021/acs.jproteome.6b00598
48. Walker WA. Initial intestinal colonization in the human infant and immune homeostasis. Ann Nutr Metab. (2013) 63(Suppl. 2):8–15. doi: 10.1159/000354907
49. Paul-Clark MJ, Sorrentino R, Bailey LK, Sriskandan S, Mitchell JA. Gram-positive and gram-negative bacteria synergize with oxidants to release CXCL8 from innate immune cells. Mol Med. (2008) 14(5):238–46. doi: 10.2119/2007-00098.Paul-Clark
50. Luckey D, Gomez A, Murray J, White B, Taneja V. Bugs & us: The role of the gut in autoimmunity. The Indian Journal of Medical Research. (2013) 138(5):732–43.
51. Firestein GS, McInnes IB. Immunopathogenesis of rheumatoid arthritis. Immunity. (2017) 46(2):183–96. doi: 10.1016/j.immuni.2017.02.006
52. Hannah J, Sonya AJR. The evidence for microbiome manipulation in inflammatory arthritis. Rheumatology. (2016) 56(9):1452–60. doi: 10.1093/rheumatology/kew374
53. Reyes-Castillo Z, Valdés-Miramontes E, Llamas-Covarrubias M, Muoz-Valle JF. Troublesome friends within us: the role of gut microbiota on rheumatoid arthritis etiopathogenesis and its clinical and therapeutic relevance. Clin Exp Med. (2021) 21(1):1–13. doi: 10.1007/s10238-020-00647-y
54. Gabriel HB, Romero-Figueroa MS, Montiel-Jarquín AJ, Pizano-Zárate ML, García-Mena J, Ramírez-Durán N, et al. Intestinal dysbiosis and rheumatoid arthritis: A link between gut microbiota and the pathogenesis of rheumatoid arthritis. Journal of Immunology Research. (2017) 2017:4835189. doi: 10.1155/2017/4835189
55. Zaiss MM, Wu HJJ, Mauro D, Schett G, Ciccia F. The gut–joint axis in rheumatoid arthritis. Nat Rev Rheumatol. (2021) 17(4):224–37. doi: 10.1038/s41584-021-00585-3
56. Brandl C, Bucci L, Schett G, Zaiss MM. Crossing the barriers: Revisiting the gut feeling in rheumatoid arthritis. European Journal of Immunology. (2021) 51(4):798–810. doi: 10.1002/eji.202048876
57. Alpizar-Rodriguez D, Lesker TR, Gronow A, Gilbert B, Raemy E, Lamacchia C, et al. Prevotella copri in individuals at risk for rheumatoid arthritis. Ann Rheumatic Dis. (2019) 78(5):590–593. doi: 10.1136/annrheumdis-2018-214514
58. Wells PM, Adebayo AS, Bowyer RCE, Freidin MB, Williams FMK. Associations between gut microbiota and genetic risk for rheumatoid arthritis in the absence of disease: a cross-sectional study. Lancet Rheumatol. (2020) 2(7):e418–27. doi: 10.1016/S2665-9913(20)30064-3
59. Mangalea MR, Paez-Espino D, Kieft K, Chatterjee A, Seifert JA, Feser ML. Individuals at risk for developing rheumatoid arthritis harbor differential intestinal bacteriophage communities with distinct metabolic potential. (2021) 29(5):726–39. doi: 10.1016/j.chom.2021.03.020
60. Allison A, Karine C, Harry S. Gut microbiota-derived metabolites as central regulators in metabolic disorders. Gut. (2021) 70(6):1174. doi: 10.1136/gutjnl-2020-323071
61. Chen Y, Ma C, Liu L, He J, Zhu C, Zheng F, et al. Analysis of gut microbiota and metabolites in patients with rheumatoid arthritis and identification of potential biomarkers. Aging. (2021) 13:23689–701. doi: 10.18632/aging.v13i20
62. Damian RT. Molecular mimicry: antigen sharing by parasite and host and its consequences. Am Nat. (1964) 98(900):129–49. doi: 10.1086/282313
63. Trost B, Lucchese G, Stufano A, Bickis M, Kanduc D. No human protein is exempt from bacterial motifs, not even one. Self/Nonself. (2010) 1:328–34. doi: 10.4161/self.1.4.13315
64. Kohm AP, Fuller KG, Miller SD. Mimicking the way to autoimmunity: an evolving theory of sequence and structural homology. Trends Microbiol. (2003) 11(3):101–5. doi: 10.1016/S0966-842X(03)00006-4
65. Lule S, Colpak AI, Balci-Peynircioglu B, Gursoy-Ozdemir Y, Peker S, Kalyoncu U, et al. Behçet Disease serum is immunoreactive to neurofilament medium which share common epitopes to bacterial HSP-65, a putative trigger. J Autoimmun. (2017) 84:87–96. doi: 10.1016/j.jaut.2017.08.002
66. Vossenaar ER, Zendman AJW, Venrooij WJV, Pruijn GJM. PAD, a growing family of citrullinating enzymes: genes, features and involvement in disease. Bioessays. (2003) 25(11):1106–18. doi: 10.1002/bies.10357
67. Mcgraw WT, Potempa J, Farley D, Travis J. Purification, characterization, and sequence analysis of a potential virulence factor from Porphyromonas gingivalis, peptidylarginine deiminase. Infection & Immunity. (1999) 67(7):3248. doi: 10.1128/IAI.67.7.3248-3256.1999
68. Li S, Yu Y, Yue Y, Zhang Z, Su K. Microbial infection and rheumatoid arthritis. J Clin Cell Immunol. (2013) 4(6):164. doi: 10.4172/2155-9899.1000174
69. de Pablo P, Dietrich T, McAlindon TE. Association of periodontal disease and tooth loss with rheumatoid arthritis in the US population. J Rheumatol. (2008) 35(1):70–6. doi: 10.1080/10582450802479693
70. Smit MD, Westra J, Vissink A, Meer DVD, Brouwer E, Winkelhoff AJV. Periodontitis in established rheumatoid arthritis patients: a cross-sectional clinical, microbiological and serological study. Arthritis Res Ther. (2012) 14(5):R222. doi: 10.1186/ar4061
71. Anne-Marie Q, Lugli EB, Wegner N, Hamilton BC, Charles P, Chowdhury M, et al. Heightened immune response to autocitrullinated Porphyromonas gingivalis peptidylarginine deiminase: a potential mechanism for breaching immunologic tolerance in rheumatoid arthritis. Ann Rheumatic Dis. (2014) 73(1):263–269. doi: 10.1136/annrheumdis-2012-202726
72. Vossenaar ER, Smeets TJM, Kraan MC, Raats JM, Venrooij WJV, Tak PP. The presence of citrullinated proteins is not specific for rheumatoid synovial tissue. Arthritis & Rheumatism. (2004) 50(11):3485–94. doi: 10.1002/art.20584
73. Liu X, Zeng B, Zhang J, Li W, Mou F, Wang H. Role of the gut microbiome in modulating arthritis progression in mice. Sci Rep. (2016) 6(1):30594. doi: 10.1038/srep30594
74. Konig MF, Abusleme L, Reinholdt J, Palmer RJ, Teles RP, Sampson K, et al. Aggregatibacter actinomycetemcomitans–induced hypercitrullination links periodontal infection to autoimmunity in rheumatoid arthritis. Sci Trans Med. (2016) 8(369):369ra176. doi: 10.1126/scitranslmed.aaj1921
75. Scher JU, Ubeda C, Equinda M, Khanin R, Buischi Y, Viale A, et al. Periodontal disease and the oral microbiota in new-onset rheumatoid arthritis. Arthritis & Rheumatism. (2012) 64(10):3083–94. doi: 10.1002/art.34539
76. Lopez-Oliva I, Paropkari AD, Saraswat S, Serban S, Yonel Z, Sharma P, et al. Dysbiotic subgingival microbial communities in periodontally healthy patients with rheumatoid arthritis. Arthritis & Rheumatology. (2018) 70(7):1008–13. doi: 10.1002/art.40485
77. Tong Y, Zheng L, Qing P, Zhao H, Li Y, Su L, et al. Oral microbiota perturbations are linked to high risk for rheumatoid arthritis. Front Cell Infection Microbiol. (2020) 9:00475. doi: 10.3389/fcimb.2019.00475
78. de Almeida DE, Ling S, Holoshitz J. New insights into the functional role of the rheumatoid arthritis shared epitope. FEBS Letters. (2011) 585(23):3619–26. doi: 10.1016/j.febslet.2011.03.035
79. Mattey DL, Dawes PT, Gonzalez-Gay MA, Garcia-Porrua C, Ollier WER. HLA-DRB1 alleles encoding an aspartic acid at position 70 protect against development of rheumatoid arthritis. Journal of Rheumatology. (2001) 28(2):232–9. doi: 10.1016/S1297-319X(00)00232-3
80. Sebastien V, Plant D, Bowes J, Lunt M, Eyre S, Barton A, et al. Genetic markers of rheumatoid arthritis susceptibility in anti-citrullinated peptide antibody negative patients. Ann Rheumatic Dis. (2012) 71(12):1984. doi: 10.1136/annrheumdis-2011-201225
81. Imboden JB. The immunopathogenesis of rheumatoid arthritis. (2009) 4(1):417–34. doi: 10.1146/annurev.pathol.4.110807.092254
82. Parham P. MHC class I molecules and KIRs in human history, health and survival. Immunology. (2005) 5(3):201–214. doi: 10.1038/nri1570
83. Ling S, Pi X, Holoshitz J. The rheumatoid arthritis shared epitope triggers innate immune signaling via cell surface calreticulin. Journal of Immunology. (2007) 179(9):6359–67. doi: 10.4049/jimmunol.179.9.6359
84. Ling S, Cheng A, Pumpens P, Michalak M, Holoshitz J. Identification of the rheumatoid arthritis shared epitope binding site on calreticulin. PloS One. (2010) 5:e11703. doi: 10.1371/journal.pone.0011703
85. Evans DM, Ralston SH. Nitric oxide and bone. Immunology. (1996) 11(3):300–5. doi: 10.1002/jbmr.5650110303
86. Lee NK, Choi YG, Baik JY, Han SY, Jeong DW, Bae YS, et al. A crucial role for reactive oxygen species in RANKL-induced osteoclast differentiation. Blood. (2005) 106(3):852–9. doi: 10.1182/blood-2004-09-3662
87. Lean JM, Davies JT, Fuller K, Jagger CJ, Kirstein B, Partington GA, et al. A crucial role for thiol antioxidants in estrogen-deficiency bone loss. J Clin Invest. (2003) 112(6):915–23. doi: 10.1172/JCI200318859
88. Zeng G, Aldridge ME, Xiaoli T, Seiler D. Dendritic cell surface calreticulin is a receptor for NY-ESO-1: direct interactions between tumor-associated antigen and the innate immune system. J Immunol. (2006) 177:3582–9. doi: 10.4049/jimmunol.177.6.3582
89. Henson PM, Bratton DL, Fadok VA. The phosphatidylserine receptor: a crucial molecular switch? Nat Rev Mol Cell Biol. (2001) 2(8):627. doi: 10.1038/35085094
90. Gardai SJ, Bratton DL, Ogden CA, Henson PM. Recognition ligands on apoptotic cells: a perspective. J Leukocyte Biol. (2006) 79:896–903. doi: 10.1189/jlb.1005550
91. Mellor A, Munn D, Mellor AL, Munn DH. IDO expression by dendritic cells: tolerance and tryptophan catabolism. Nat Rev Immunol. (2004) 4:762–74. doi: 10.1038/nri1457
92. Kolodziej LR, Paleolog EM, Williams RO. Kynurenine metabolism in health and disease. Amino Acids. (2011) 41(5):1173–83. doi: 10.1007/s00726-010-0787-9
93. Fallerino F, Vacca C, Orabona C, Belladonna ML, Bianchi R, Marshall B, et al. Functional expression of indoleamine 2,3-dioxygenase by murine CD8{alpha}+ dendritic cells. Int Immunol. (2002) 14(1):65–8. doi: 10.1093/intimm/14.1.65
94. Korn T, Bettelli E, Gao W, Awasthi A, Jger A, Strom TB, et al. IL-21 initiates an alternative pathway to induce proinflammatory T(H)17 cells. Nature. (2007) 448(7152):484–7. doi: 10.1038/nature05970
95. De Almeida DE, Ling S, Pi X, Hartmann-Scruggs AM, Pumpens P, Holoshitz J. Immune dysregulation by the rheumatoid arthritis shared epitope. J Immunol. (2010) 185(3):1927–34. doi: 10.4049/jimmunol.0904002
96. Who F. Health and nutritional properties of probiotics in food including powder milk with live lactic acid bacteria – joint FAO/WHO expert consultation. (2001), 1–34. doi: 10.1201/9781420009613.ch16
97. Amdekar S, Singh V, Singh R, Sharma P, Keshav P, Kumar A. Lactobacillus casei reduces the Inflammatory Joint Damage Associated with Collagen-Induced Arthritis (CIA) by Reducing the Pro-Inflammatory Cytokines. J Clin Immunol. (2011) 31:147–54. doi: 10.1007/s10875-010-9457-7
98. Pan H, Guo R, Ju Y, Wang Q, Liu L. A single bacterium restores the microbiome dysbiosis to protect bones from destruction in a rat model of rheumatoid arthritis. Microbiome. (2019) 7:107. doi: 10.1186/s40168-019-0719-1
99. So J-S, Lee CG, Kwon HK, Yi HJ, Chae CS, Park JA, et al. Lactobacillus casei potentiates induction of oral tolerance in experimental arthritis. Mol Immunol. (2008) 46:172–80. doi: 10.1016/j.molimm.2008.07.038
100. Gohil P, Patel V, Deshpande S, Chorawala M, Shah G. Anti-arthritic activity of cell wall content of Lactobacillus plantarum in freund’s adjuvant-induced arthritic rats: involvement of cellular inflammatory mediators and other biomarkers. Inflammopharmacology. (2018) 26:171–81. doi: 10.1007/s10787-017-0370-z
101. Vaghef-Mehrabany E, Homayouni-Rad A, Alipour B, Sharif SK, Vaghef-Mehrabany L, Alipour-Ajiry S. Effects of probiotic supplementation on oxidative stress indices in women with rheumatoid arthritis: A randomized double-blind clinical trial. J Am Coll Nutr. (2016) 35:291–9. doi: 10.1080/07315724.2014.959208
102. Alipour B, Homayouni-Rad A, Vaghef-Mehrabany E, Sharif SK, Vaghef-Mehrabany L, Asghari-Jafarabadi M, et al. Effects of Lactobacillus casei supplementation on disease activity and inflammatory cytokines in rheumatoid arthritis patients: a randomized double-blind clinical trial. Int J Rheumatic Dis. (2014) 17:519–27. doi: 10.1111/1756-185X.12333
103. Hatakka K, Martio J, Korpela M, Herranen M, Korpela R. Effects of probiotic therapy on the activity and activation of mild rheumatoid arthritis – a pilot study. Scandinavian J Rheumatol. (2003) 32:211–5. doi: 10.1080/03009740310003695
104. Zamani B, Farshbaf S, Golkar HR, Bahmani F. Synbiotic supplementation and the effects on clinical and metabolic responses in patients with rheumatoid arthritis: a randomised, double-blind, placebo-controlled trial. Br J Nutr. (2017) 117:1095–102. doi: 10.1017/S000711451700085X
105. Cannarella LAT, Mari NL, Alcântara CC, Iryioda TMV, Costa NT, Oliveira SR, et al. Mixture of probiotic reduces inflammatory biomarkers and improves the oxidative/nitrosative profile in patients with rheumatoid arthritis. Nutrition. (2021) 89:111282. doi: 10.1016/j.nut.2021.111282
106. Fan Z, Yang B, Ross RP, Stanton C, Chen W. The prophylactic effects of different Lactobacilli on collagen-induced arthritis in rats. Food Funct. (2020) 11(4):3681–94. doi: 10.1039/C9FO02556A
107. Vaghef-Mehrabany E, Alipour B, Homayouni-Rad A, Sharif SK, Asghari-Jafarabadi M, Zavvari S. Probiotic supplementation improves inflammatory status in patients with rheumatoid arthritis. Nutrition. (2014) 30(4):430–5. doi: 10.1016/j.nut.2013.09.007
108. Pineda Mde L, Thompson SF, Summers K, De Leon F, Pope J, Reid G. A randomized, double-blinded, placebo-controlled pilot study of probiotics in active rheumatoid arthritis. Med Sci Monit. (2011) 17(6):Cr347–354. doi: 10.12659/MSM.881808
109. Fan Z, Yang B, Ross RP, Stanton C, Shi G, Zhao J, et al. Protective effects of Bifidobacterium adolescentis on collagen-induced arthritis in rats depend on timing of administration. Food Funct. (2020) 11:4499–511. doi: 10.1039/D0FO00077A
110. Jeong Y, Jhun JY, Lee SY, Na HS, Choi JW, Cho KH, et al. Therapeutic potential of a novel bifidobacterium identified through microbiome profiling of RA patients with different RF levels. Front Immunol. (2021) 12:736196. doi: 10.3389/fimmu.2021.736196
111. Zamani B, Golkar HR, Farshbaf S, Emadi-Baygi M, Tajabadi-Ebrahimi M, Jafari P, et al. Clinical and metabolic response to probiotic supplementation in patients with rheumatoid arthritis: a randomized, double-blind, placebo-controlled trial. Clin Nutr. (2016) 19(9):869–79. doi: 10.1111/1756-185X.12888
112. Ji GE, Jeong Y, Fang H, Kwon B, Park SH, Cho ML, et al. Composition Containing Bifidobacterium for Alleviating, Preventing or Treating Rheumatoid Arthritis. Gangwom-do KR, Seoul KR, Republic of Korea. (2020).
113. Marietta EV, Murray JA, Luckey DH, Jeraldo PR, Taneja V. Suppression of inflammatory arthritis by human gut-derived prevotella histicola in humanized mice. Arthritis Rheumatol. (2016) 68(12):2878–88. doi: 10.1002/art.39785
114. Balakrishnan B, Luckey D, Bodhke R, Chen J, Marietta E, Jeraldo P, et al. Prevotella histicola protects from arthritis by expansion of allobaculum and augmenting butyrate production in humanized mice. Anaerobe. (2021) 12:102237. doi: 10.3389/fimmu.2021.609644
115. Mandel DR, Eichas K, Holmes J. Bacillus coagulans: a viable adjunct therapy for relieving symptoms of rheumatoid arthritis according to a randomized, controlled trial. BMC Complementary Altern Med. (2010) 10(1):1. doi: 10.1186/1472-6882-10-1
116. Kang Y, Cai Y, Zhang X, Kong X, Su J. Altered gut microbiota in RA: implications for treatment. Z für Rheumatologie. (2017) 76(5):451–7. doi: 10.1007/s00393-016-0237-5
117. Abhari K, Shekarforoush SS, Hosseinzadeh S, Nazifi S, Sajedianfard J, Eskandari MH. The effects of orally administered Bacillus coagulans and inulin on prevention and progression of rheumatoid arthritis in rats. Food Nutr Res. (2016) 60:30876. doi: 10.3402/fnr.v60.30876
118. Ferro M, Charneca S, Dourado E, Guerreiro CS, Fonseca JE. Probiotic supplementation for rheumatoid arthritis: A promising adjuvant therapy in the gut microbiome era. Front Pharmacol. (2021) 12:711788. doi: 10.3389/fphar.2021.711788
119. Cristofori F, Dargenio VN, Dargenio C, Miniello VL, Francavilla R. Anti-inflammatory and immunomodulatory effects of probiotics in gut inflammation: A door to the body. Front Immunol. (2021) 12:578386. doi: 10.3389/fimmu.2021.578386
120. Li M, Wang F. Role of intestinal microbiota on gut homeostasis and rheumatoid arthritis. J Immunol Res. (2021) 2021:8167283. doi: 10.1155/2021/8167283
121. Goh YJ, Klaenhammer TR. Functional roles of aggregation-promoting-like factor in stress tolerance and adherence of lactobacillus acidophilus NCFM. Appl Environ Microbiol. (2010) 76:5005–12. doi: 10.1128/AEM.00030-10
122. La Fata G, Weber P, Mohajeri MH. Probiotics and the gut immune system: indirect regulation. Probiotics Antimicrobial Proteins. (2018) 10:11–21. doi: 10.1007/s12602-017-9322-6
123. Cox AJ, West NP, Cripps AW. Obesity, inflammation, and the gut microbiota. Lancet Diabetes Endocrinol. (2015) 3:207–15. doi: 10.1016/S2213-8587(14)70134-2
124. Rasouli-Saravani A, Jahankhani K, Moradi S, Gorgani M, Shafaghat Z, Mirsanei Z, et al. Role of microbiota short-chain fatty acids in the pathogenesis of autoimmune diseases. Biomedicine Pharmacotherapy. (2023) 162:114620. doi: 10.1016/j.biopha.2023.114620
125. Nastasi C, Fredholm S, Willerslev-Olsen A, Hansen M, Woetmann A. Butyrate and propionate inhibit antigen-specific CD8+ T cell activation by suppressing IL-12 production by antigen-presenting cells. Sci Rep. (2017) 7:14516. doi: 10.1038/s41598-017-15099-w
126. Berndt BE, Zhang M, Owyang SY, Cole TS, Wang TW, Luther J, et al. Butyrate increases IL-23 production by stimulated dendritic cells. Am J Physiol. (2012) 303:G1384–92. doi: 10.1152/ajpgi.00540.2011
127. Aoyama M, Kotani J, Usami M. Butyrate and propionate induced activated or non-activated neutrophil apoptosis via HDAC inhibitor activity but without activating GPR-41/GPR-43 pathways. Nutrition. (2010) 26:653–61. doi: 10.1016/j.nut.2009.07.006
128. Bailón E, Cueto-Sola M, Utrilla P, Rodríguez-Cabezas ME, Comalada M. Butyrate in vitro immune-modulatory effects might be mediated through a proliferation-related induction of apoptosis. Immunobiology. (2010) 215:863–73. doi: 10.1016/j.imbio.2010.01.001
129. Iraporda C, Errea A, Romanin DE, Cayet D, Pereyra E, Pignataro O, et al. Lactate and short chain fatty acids produced by microbial fermentation downregulate proinflammatory responses in intestinal epithelial cells and myeloid cells. Immunobiology. (2015) 220(10):1161–1169. doi: 10.1016/j.imbio.2015.06.004
130. Fasano A. Zonulin and its regulation of intestinal barrier function: the biological door to inflammation, autoimmunity, and cancer. Physiol Rev. (2011) 91:151–75. doi: 10.1152/physrev.00003.2008
131. Takahashi D, Hoshina N, Kabumoto Y, Maeda Y, Suzuki A, Tanabe H, et al. Microbiota-derived butyrate limits the autoimmune response by promoting the differentiation of follicular regulatory T cells. EBioMedicine. (2020) 58:102913. doi: 10.1016/j.ebiom.2020.102913
132. Smolen JS, Aletaha D. Rheumatoid arthritis therapy reappraisal: strategies, opportunities and challenges. Nat Rev Rheumatol. (2015) 11:276–89. doi: 10.1038/nrrheum.2015.8
133. Smolen JS, Aletaha D, McInnes IB. Rheumatoid arthritis. Lancet. (2016) 388:2023–38. doi: 10.1016/S0140-6736(16)30173-8
134. Rojas M, Restrepo-Jiménez P, Monsalve DM, Pacheco Y, Acosta-Ampudia Y, Ramírez-Santana C, et al. Molecular mimicry and autoimmunity. J Autoimmun. (2018) 95:100–23. doi: 10.1016/j.jaut.2018.10.012
135. Di Tommaso N, Gasbarrini A, Ponziani FR. Intestinal barrier in human health and disease. Int J Environ Res Public Health. (2021) 18:12836. doi: 10.3390/ijerph182312836
136. van Steenbergen HW, Cope AP, van der Helm-van Mil AHM. Rheumatoid arthritis prevention in arthralgia: fantasy or reality? Nat Rev Rheumatol. (2023) 19:767–77. doi: 10.1038/s41584-023-01035-y
137. Krijbolder DI, Verstappen M, Dijk BTV, Dakkak Y, Burgers L, Boer A, et al. Intervention with methotrexate in patients with arthralgia at risk of rheumatoid arthritis to reduce the development of persistent arthritis and its disease burden (TREAT EARLIER): a randomised, double-blind, placebo-controlled, proof-of-concept trial. Lancet. (2022) 400:283–94. doi: 10.1016/S0140-6736(22)01193-X
138. Niemantsverdriet E, Dakkak YJ, Burgers LE, Bonte-Mineur F, van der Helm-van Mil AHM, et al. TREAT Early Arthralgia to Reverse or Limit Impending Exacerbation to Rheumatoid arthritis (TREAT EARLIER): a randomized, double-blind, placebo-controlled clinical trial protocol. Trials. (2020) 21(1):862. doi: 10.1186/s13063-020-04731-2
139. Al-Laith M, Jasenecova M, Abraham S, Bosworth A, Bruce IN, Buckley CD, et al. Arthritis prevention in the pre-clinical phase of RA with abatacept (the APIPPRA study): a multi-centre, randomised, double-blind, parallel-group, placebo-controlled clinical trial protocol. Trials. (2019) 20(1):429. doi: 10.1186/s13063-019-3403-7
Keywords: rheumatoid arthritis, gut microbiota, immune response, potential mechanism, probiotics therapy
Citation: Yang Y, Hong Q, Zhang X and Liu Z (2024) Rheumatoid arthritis and the intestinal microbiome: probiotics as a potential therapy. Front. Immunol. 15:1331486. doi: 10.3389/fimmu.2024.1331486
Received: 01 November 2023; Accepted: 19 February 2024;
Published: 06 March 2024.
Edited by:
Julio Villena, CONICET Reference Centre for Lactobacilli (CERELA), ArgentinaReviewed by:
Claudio Nicoletti, University of Siena, ItalyCopyright © 2024 Yang, Hong, Zhang and Liu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Zhenmin Liu, bGl1emhlbm1pbkBicmlnaHRkYWlyeS5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.