
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Immunol. , 29 November 2023
Sec. Cancer Immunity and Immunotherapy
Volume 14 - 2023 | https://doi.org/10.3389/fimmu.2023.1291292
This article is part of the Research Topic Glycans: Molecules at the Interface of Immunity and Disease View all 14 articles
 Michela Manni1,2
Michela Manni1,2 Natalia Rodrigues Mantuano1Andreas Zingg1Elisabeth A. Kappos3Anna-Janina Behrens2Jonathan Back2
Natalia Rodrigues Mantuano1Andreas Zingg1Elisabeth A. Kappos3Anna-Janina Behrens2Jonathan Back2 Rainer Follador2Amir Faridmoayer2
Rainer Follador2Amir Faridmoayer2 Heinz Läubli1,4*
Heinz Läubli1,4*Humans lack the enzyme that produces the sialic acid N-glycolyl neuraminic acid (Neu5Gc), but several lines of evidence have shown that Neu5Gc can be taken up by mammalian food sources and replace the common human sialic acid N-acetyl neuraminic acid (Neu5Ac) in glycans. Cancer tissue has been shown to have increased the presence of Neu5Gc and Neu5Gc-containing glycolipids such as the ganglioside GM3, which have been proposed as tumor-specific antigens for antibody treatment. Here, we show that a previously described antibody against Neu5Gc-GM3 is binding to Neu5GC-containing gangliosides and is strongly staining different cancer tissues. However, we also found a strong intracellular staining of keratinocytes of healthy skin. We confirmed this staining on freshly isolated keratinocytes by flow cytometry and detected Neu5Gc by mass spectrometry. This finding implicates that non-human Neu5Gc can be incorporated into gangliosides in human skin, and this should be taken into consideration when targeting Neu5Gc-containing gangliosides for cancer immunotherapy.
Cancer-associated glycan changes include the truncation of carbohydrate chains, increased N-glycan branching, and changes in sialic acid metabolism such as increased terminal sialic acid concentrations (1, 2). Such cancer-associated glycan changes have been targeted using tumor-specific antibodies to the cancer microenvironment (3, 4). One prominent example is the sialyl-Thomsen nouveau (sTn) antigen, a sialylated-truncated O-glycan (5). A trial targeting this glycan antigen in a phase I trial in solid cancer patients with an antibody–drug conjugate is ongoing (e.g., Seagen Inc, NCT04665921).
Sialic acids are monosaccharides that are commonly found on the outer terminal units of cell surfaces in vertebrates (1, 6). They play a crucial role in the interactions between cells and their microenvironment. The two main types of sialic acids found on mammalian cells are N-glycolyl-neuraminic acid (Neu5Gc) and N-acetyl-neuraminic acid (Neu5Ac) (1, 6, 7). Neu5Gc is particularly notable because it is deficient in humans due to a specific genetic deletion in the CMAH gene, which encodes the hydroxylase responsible for converting CMP-Neu5Ac to CMP-Neu5Gc (7). However, despite this deficiency, Neu5Gc can still be absorbed into human tissues through dietary sources, especially red meat, leading to higher concentrations of Neu5Gc in certain types of cancer (8–12). Breast carcinomas showed anti-Neu5GC reactivity in the majority of tumor cells and some related blood vessels (9). In addition, non-human Neu5Gc has also been detected in other healthy organs and biotherapeutic agents, although usually at rather lower levels in healthy tissue (13–15).
Previously, limited expression of N-glycolyl GM3 (NeuGcGM3) ganglioside in human normal tissues was reported, as well as its presence in melanoma and breast carcinoma using the 14F7 antibody (anti-NeuGcGM3)(16, 17). The expression of NeuGc-GM3 using the same antibody was found in a high proportion of neuroectodermal and Wilms tumors, common pediatric cancers (18, 19). The ganglioside Neu5Gc-GM3 has been previously defined as a potential tumor-specific target with little to no expression in healthy tissue (20–22). We further wanted to define the expression of NeuGc-GM3 ganglioside as a potential tumor target in different malignancies. We therefore performed staining with an 14F7 antibody clone.
Immunohistochemistry was performed by TriStar Technology Group using a Ventana BenchMark XT staining system. In brief, 4-μm FFPE TMAs (TriStar Technology Group) were deparaffinized and hydrated before antigen retrieval in Tris–EDTA buffer, pH 8.4, 100°C, for 8 min. Slides were then blocked in 3% hydrogen peroxide for 4 min at 37°C. Slides were then incubated with 14F7 (1:8,000 dilution, 1 μg/ml) or control mouse IgG antibodies at 37°C for 12 min. Primary antibodies were detected using the OptiView Universal DAB kit (Ventana, #760-700). TMAs (TA3162, TA2998, TA2472, TA2287, and TA2942) were scored by a trained pathologist at TriStar Technology Group. The score 0 to 3+ measures the intensity of the staining in the sample analyzed, assigning 0 for no staining, 1+ for weak staining, 2+ for intermediate staining, and 3+ for strong staining. The H-score was used to evaluate the degree immunoreactivity. The H-score was calculated using the following formula: three times the percentage of strongly stained cells plus two times the percentage of moderately stained cells, in addition to the percentage of weakly stained cells. This computation yields a numerical range spanning from 0 to 300.
Patients undergoing breast and abdominal reduction were consented and enrolled at the University Hospital of Basel. Fresh human skin was collected in PBS, subcutaneous fat was removed, and skin was extensively cut with a scalpel in a petri dish. Skin was then transferred into a 15-ml tube with 1 ml of the following mix per four punch biopsies: collagenase II (2.5 mg/ml) + collagenase IV (2.5 mg/ml) + DNAse (0.5 mg/ml) in PBS. Skin was then digested for 2 h at 37°C with shaking. Cells were next filtered on a 100-μm filter and washed with 5 ml DMEM+10% FBS. The cells were resuspended in FACS Buffer for staining.
A 14F7 sequence was obtained from published data (23, 24).
Heavy chain: MERHWIFLFLFSVTAGVHSQVQLQQSGNELAKPGASMKMSCRASGYSFTSYWIHWLKQRPDQGLEWIGYIDPATAYTESNQKFKDKAILTADRSSNTAFMYLNSLTSEDSAVYYCARESPRLRRGIYYYAMDYWGQGTTVTVSS.
Light chain: MVSHLKILGLLLFWISASRGDLVLTQSPATLSVTPGDSVSFSCRASQSISNNLHWYQQRTHESPRLLIKYASQSISGIPSRFSGSGSGTDFTLSIISVETEDFGMYFCQQSNRWPLTFGAGTKLELKRA
Expression and purification of the monoclonal antibody was performed at Evitria AG, Switzerland. In brief, the corresponding cDNA was cloned into the vector system. The vector plasmids were gene synthesized. Plasmid DNA was prepared under low-endotoxin conditions. The DNA concentration was determined by measuring the absorption at a wavelength of 260 nm. Correctness of the sequences was verified with Sanger sequencing.
CHO K1 cells were used for production. Cells were transfected in Evitria’s proprietary transfection reagent, and cells were grown after transfection in an animal-component free, serum-free medium. Supernatant was harvested by centrifugation and subsequent filtration (0.2-μm filter). The antibody was purified using MabSelect™ SuRe™.
The cell surface of human keratinocytes was treated with endoglycoceramidase I (EGCase I; New England Biolabs) according to the manufacturer’s instructions. In brief, cell pellets were digested with EGCase I for 3 h at 37°C, and the supernatant was collected and dried in a vacuum dryer. Oligosaccharides were thus released via the hydrolysis of the β-glycosidic linkage between the oligosaccharide and the ceramide in GM3-type gangliosides. The released oligosaccharides were further fluorescently labeled and analyzed by HILIC-UPLC FLR/MS, as described previously (25). Neu5Ac-GM3 and Neu5Gc-GM3 chemically synthesized standards (from ChemBind LLC) were analyzed as controls alongside the keratinocytes.
Anti-Neu5Gc-ganglioside clone 14F7 was obtained from Evitria AG. Single-cell suspension cells were first stained with Fixable Viability Dye (eBioscience) for 20 min in PBS at room temperature. Cells were washed and then incubated with antibodies (5 μg/ml 14F7) in FACS buffer (PBS + 1% FBS) in the presence of an Fc receptor binding inhibitor (Invitrogen) for 30 min on ice. Cells were washed and primary antibodies detected by staining with a goat anti-mouse IgG conjugated to Alexa Fluor 488 (Jackson ImmunoResearch). Data were acquired using a FACS Fortessa (Beckton Dickinson) and analyzed using FlowJo (TriStar).
In order to test statistical relevant differences, we used non-parametric Mann–Whitney test. We used GraphPad Prism 9.0 for the calculation of statistical differences.
The 14F7 antibody clone has been previously used to study the expression of Neu5Gc-GM3 in human tissues (22, 26–28). Different cancer types were found to have a high expression of Neu5Gc-GM3, whereas most of the healthy tissue had minimal staining as determined by immunohistochemistry (IHC). We wanted to characterize the binding of the 14F7 antibody to different glycolipids printed on a glass slide. Binding was highly specific to Neu5Gc-containing glycolipids with the strongest binding to Neu5Gc-containing GD3. Binding to Neu5Gc-GM3 was also clearly detectable (Supplementary Figure S1A). We next tested the specificity of the 14F7 antibody in an ELISA assay using Neu5Gc-GM3 and Neu5Ac-GM3, which demonstrated specific binding of 14F7 to Neu5Gc-containing gangliosides (Supplementary Figure S1B). We used the 14F7 antibody to stain various cancer tissues and analyzed the expression by classifying the staining intensity 1+ to 3+. We found that a significant percentage of TMA cores of intestinal epithelia had a positive staining (Figure 1A; Supplementary Figures S1C, D; Table 1). Several cores of colorectal cancer showed a strong staining (Figures 1A, B). When we compared the staining of normal colon tissue with colorectal cancer, we observed a clear increased H-score in many colorectal samples and no or minimal staining in samples of healthy colon tissue (Figure 1C).
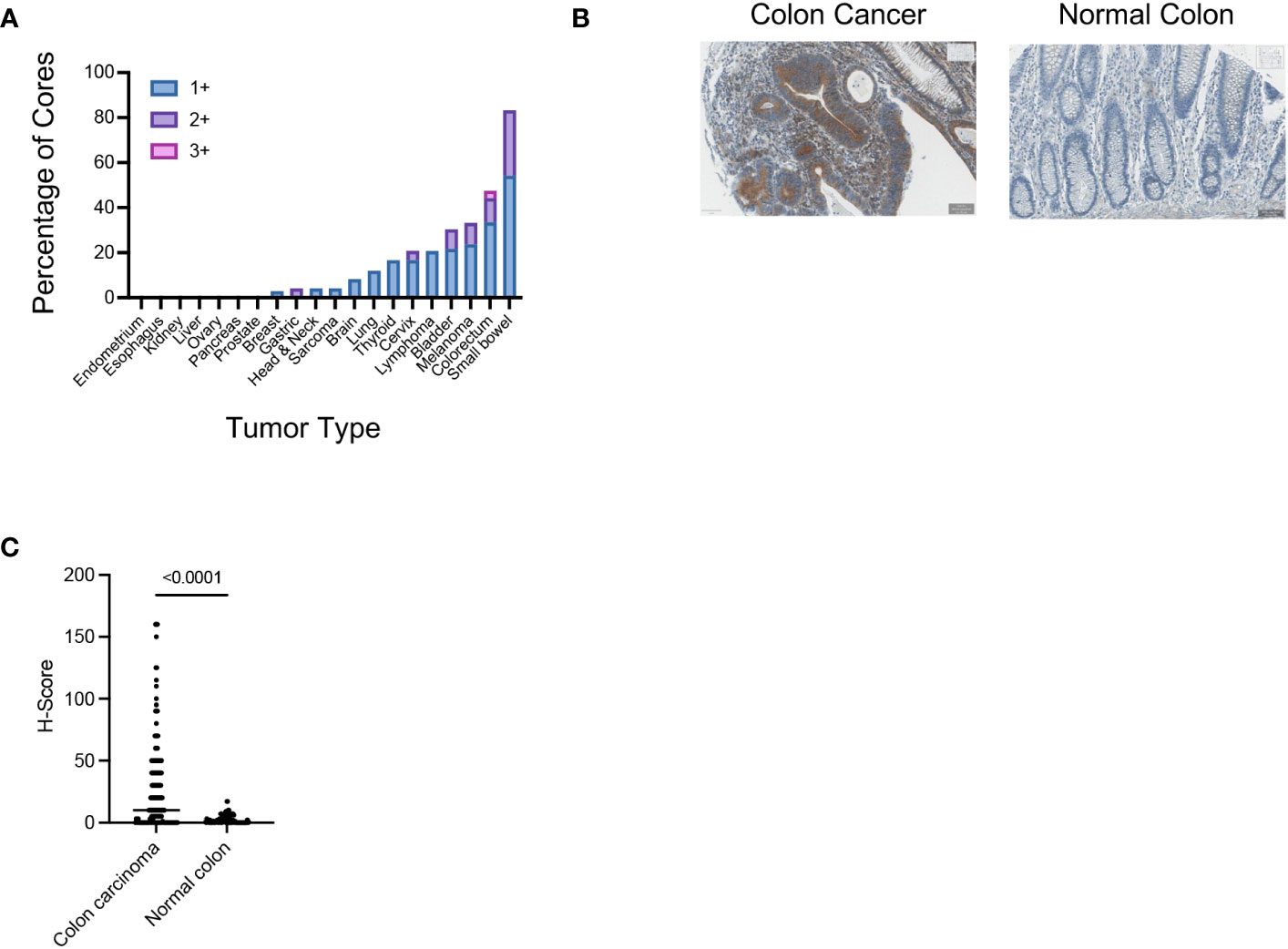
Figure 1 (A) Assessment of the Neu5Gc-GM3 expression in the indicated human tumor samples using the 14F7 antibody. Graph shows the percentage of TMA cores which show an IHC score of 1+, 2+, or 3+ in the indicated tumor samples. (B) Representative IHC image showing the expression of Neu5Gc-GM3 in colon cancer or normal colon as assessed by 14F7 staining. (C) Graph showing the Neu5Gc-GM3 level in colon cancer (140 cores) or normal colon (42 cores) expressed as H-Score. Each dot represents one TMA core.
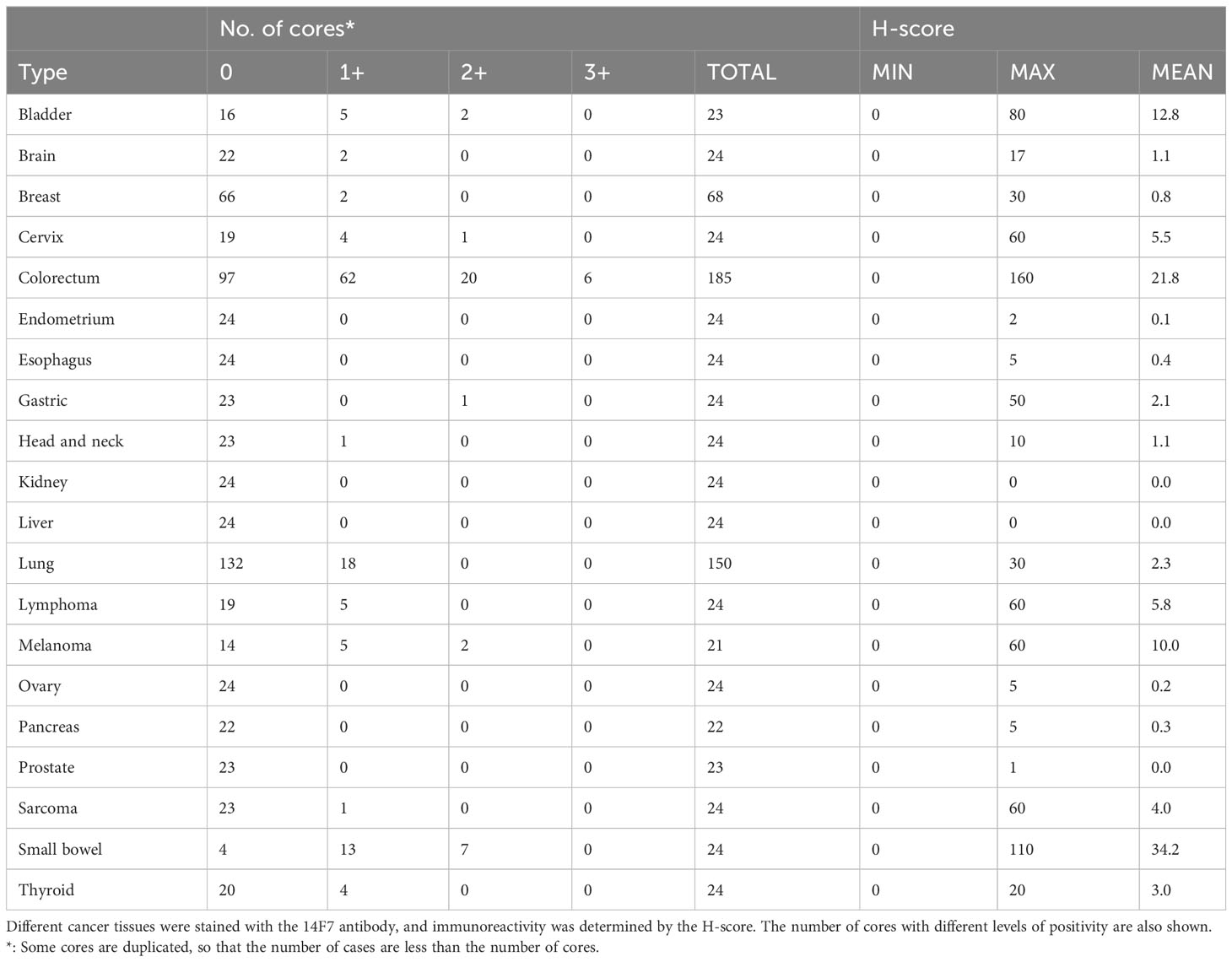
Table 1 Summary of 14F7 IHC results in various tumor tissues.
Taken together, these data reproduce previous observations showing an enriched staining with the 14F7 antibody in various cancer types and minimal staining in healthy tissue.
Our analysis also showed a significant staining of melanoma samples (Figures 2A, B). However, when we further compared the H-score from a TMA with additional healthy skin samples than used in the initial screening (Supplementary Figure S1C), we found a very strong staining of 14F7 which was mainly localized intracellularly in keratinocytes (Figures 2A, B). Indeed, the staining of healthy skin was even higher than in melanoma samples as measured by H-score determination (Figure 2B).
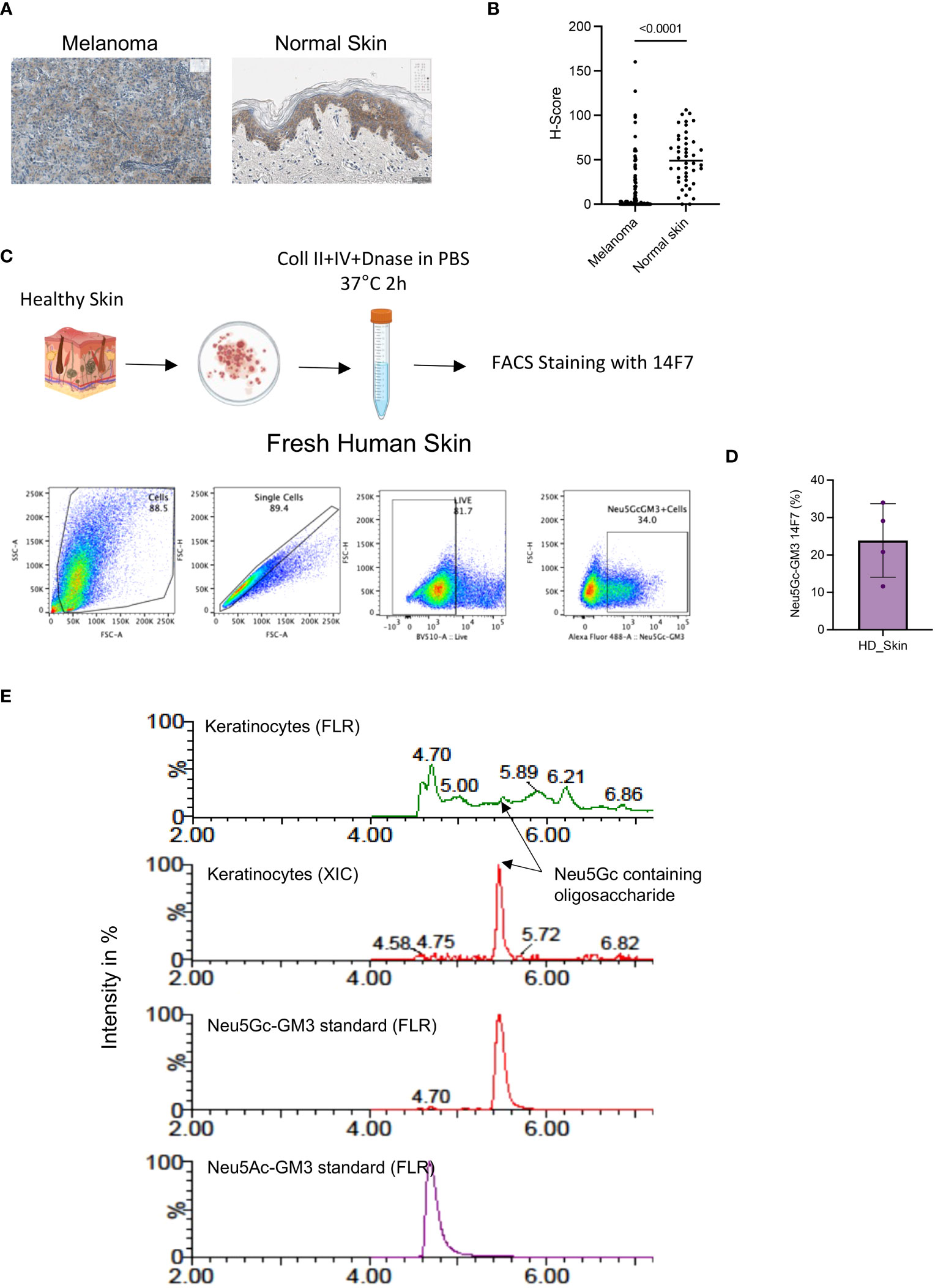
Figure 2 (A) Representative IHC image showing the expression of Neu5Gc-GM3 in melanoma or normal skin as assessed by 14F7 staining. (B) Quantification of Neu5Gc-GM3 expression as assessed by 14F7 staining. Graph shows the H-score in melanoma (119 cores) and normal skin (44 cores). Each dot represents one TMA core. (C) Gating strategy used to evaluate Neu5Gc-GM3 expression in fresh human skin by flow cytometry. (D) Quantification of (C). Graph showing the percentage of skin cells expressing Neu5Gc-GM3. Each dot represents a healthy skin donor. (E), HILIC-UPLC-FLR/MS analysis of GM3 in keratinocytes in comparison with a chemically synthesized Neu5Gc-GM3 standard. In the fluorescence trace (FLR), a distinctive peak at the retention time of the Neu5Gc-GM3 control is clearly visible that is confirmed by an extracted ion chromatogram (XIC).
These data show that the Neu5Gc-specific antibody 14F7 is strongly staining keratinocytes in healthy skin, although it is not excluded that other cell types in skin are also positive for Neu5Gc-containing gangliosides.
We further wanted to confirm the presence of Neu5Gc in human skins by additional methods. First, we wanted to test the binding of 14F7 to keratinocytes in fresh human tissue. Keratinocytes were collected from fresh healthy skin tissue obtained from patients undergoing breast reduction surgery. Dissociated tissue was processed into single-cell suspensions, which were analyzed by flow cytometry after staining with the 14F7 antibody (Figure 2C). Flow cytometry analysis of fresh unfixed skin showed a significant number of keratinocytes positive for 14F7 staining (Figure 2D and Supplementary Figure 2A).
The presence of Neu5Gc-GM3 ceramides in human keratinocytes was supported by mass spectrometry using HILIC-UPLC FLR/MS. EGCase I is a glycosphingolipid-specific enzyme that allows the specific analysis of ceramide-associated oligosaccharides. Figure 2E shows the analysis of a chemically synthesized Neu5Gc-GM3 standard in comparison with the surface ceramide-associated oligosaccharides of keratinocytes. The retention times in the fluorescence spectrum and masses observed in the analysis of human keratinocytes are identical to the ones generated from standards and support the presence of Neu5Gc-containing glycolipids in keratinocytes.
These results show that Neu5Gc is present in human healthy skin.
Changes in sialic acid uptake and sialylation of cancer-associated glycans have been described since many years (1, 8, 29, 30). In particular, upregulation of glycans containing sialic acid (hypersialylation) and incorporation of the non-human sialic acid Neu5Gc have been identified as the most common changes (4). Here, we describe the presence of the Neu5Gc-containing glycolipids in the skin of healthy humans, which is surprising since humans lack the hydroxylase that is responsible for the generation of the glycolyl group (15). However, it has been reported that Neu5Gc can be taken up by the consumption of different mammalian food sources and incorporated into human glycans (9, 10). This xeno-antigen also leads to antibodies that recognize this epitope (31).
Previous analyses have not discovered increased amounts of Neu5Gc in human skin (32), probably because general antibodies against Neu5Gc were used for the detection. Here, we used the specific antibody 14F7 for the detection of Neu5Gc-containing gangliosides by immunostaining. In addition, we confirmed the binding of the 14F7 antibody to formalin-fixed keratinocytes as well as to unfixed keratinocytes from fresh human skin. In addition, we corroborated our finding by an antibody-independent method using mass spectrometry from keratinocytes.
As other healthy tissues have not been found to have a significant positive staining for 14F7, Neu5Gc-GM3 could still be an interesting target, in particular as the main staining was found intracellular in keratinocytes. The 14F7 antibody was tested on cell lines in vitro (20, 30, 33), and humanized antibodies of 14F7 have been made and tested in preclinical in vitro and in vivo models (22). Gangliosides have been targeted for cancer therapy by antibodies and also CAR T cells. For example, GD2 is targeted with chimeric antigen receptor (CAR) T cells and antibody–drug candidates in neuroblastoma and glioma patients (34). However, further tests are needed to determine the potential use of the 14F7 antibody for therapeutic use.
Humans absorb and metabolically incorporate Neu5Gc, enriched in foods of mammalian origin, even while generating xenoreactive, and potentially autoreactive, antibodies against the same molecule (9). Although the detection of Neu5Gc-containing glycolipids in healthy human skin is interesting, it remains unclear how Neu5Gc is taken up by keratinocytes and further studies are needed to determine its exact role and metabolism.
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
The studies involving humans were approved by Ethical Committee of Central and Northwestern Switzerland (EKNZ). The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
MM: Conceptualization, Data curation, Formal Analysis, Investigation, Supervision, Writing – review & editing. NR: Formal Analysis, Project administration, Writing – original draft. AZ: Investigation, Writing – review & editing. EK: Resources, Writing – review & editing. A-JB: Data curation, Investigation, Writing – review & editing. JB: Supervision, Writing – review & editing. RF: Conceptualization. AF: Funding acquisition, Supervision, Writing – review & editing. HL: Conceptualization, Funding acquisition, Resources, Supervision, Visualization, Writing – original draft, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was further supported by an Innosuisse Grant (42408.1 IP-LS).
We thank Charlotte Ossenbrink for her support with some experiments.
HL received travel grants and consultant fees from Bristol-Myers Squibb BMS and Merck, Sharp and Dohme MSD. HL received research support from BMS, Novartis, GlycoEra, and Palleon Pharmaceuticals. MM, A-JB, JB, RF, and AF are employees of GlycoEra. HL and NR are co-founders of Glycocalyx Therapeutics.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2023.1291292/full#supplementary-material
ELISA, enzyme-linked immunosorbent assay; FLR, fluorescence trace; IHC, immunohistochemistry; MS, mass spectrometry; Neu5Ac, N-acetyl-neuraminic acid; Neu5Gc, N-glycolyl-neuraminic acid; TMA, tissue microarray.
1. Pearce OM, Laubli H. Sialic acids in cancer biology and immunity. Glycobiology (2016) 26:111–28. doi: 10.1093/glycob/cwv097
2. Bellis SL, Reis CA, Varki A, Kannagi R, Stanley P. Glycosylation changes in cancer. In: Varki A, Cummings RD, Esko JD, Stanley P, Hart GW, Aebi M, Mohnen D, Kinoshita T, Packer NH, Prestegard JH, Schnaar RL, Seeberger PH, editors. Essentials of glycobiology. NY: Cold Spring Harbor (2022).
3. Mereiter S, Balmana M, Campos D, Gomes J, Reis CA. Glycosylation in the era of cancer-targeted therapy: where are we heading? Cancer Cell (2019) 36:6–16. doi: 10.1016/j.ccell.2019.06.006
4. Rodrigues Mantuano N, Natoli M, Zippelius A, Laubli H. Tumor-associated carbohydrates and immunomodulatory lectins as targets for cancer immunotherapy. J Immunother Cancer (2020) 8. doi: 10.1136/jitc-2020-001222
5. Prendergast JM, Galvao da Silva AP, Eavarone DA, Ghaderi D, Zhang M, Brady D, et al. Novel anti-Sialyl-Tn monoclonal antibodies and antibody-drug conjugates demonstrate tumor specificity and anti-tumor activity. MAbs (2017) 9:615–27. doi: 10.1080/19420862.2017.1290752
6. Lewis AL, Chen X, Schnaar RL, Varki A. Sialic acids and other nonulosonic acids. In: Varki A, Cummings RD, Esko JD, Stanley P, Hart GW, Aebi M, Mohnen D, Kinoshita T, Packer NH, Prestegard JH, Schnaar RL, Seeberger PH, editors. Essentials of glycobiology. NY: Cold Spring Harbor (2022).
7. Varki A, Gagneux P. Multifarious roles of sialic acids in immunity. Ann N Y Acad Sci (2012) 1253:16–36. doi: 10.1111/j.1749-6632.2012.06517.x
8. Malykh YN, Schauer R, Shaw L. N-Glycolylneuraminic acid in human tumours. Biochimie (2001) 83:623–34. doi: 10.1016/S0300-9084(01)01303-7
9. Tangvoranuntakul P, Gagneux P, Diaz S, Bardor M, Varki N, Varki A, et al. Human uptake and incorporation of an immunogenic nonhuman dietary sialic acid. Proc Natl Acad Sci U.S.A. (2003) 100:12045–50. doi: 10.1073/pnas.2131556100
10. Banda K, Gregg CJ, Chow R, Varki NM, Varki A. Metabolism of vertebrate amino sugars with N-glycolyl groups: mechanisms underlying gastrointestinal incorporation of the non-human sialic acid xeno-autoantigen N-glycolylneuraminic acid. J Biol Chem (2012) 287:28852–64. doi: 10.1074/jbc.M112.364182
11. Samraj AN, Pearce OM, Laubli H, Crittenden AN, Bergfeld AK, Banda K, et al. A red meat-derived glycan promotes inflammation and cancer progression. Proc Natl Acad Sci U.S.A. (2015) 112:542–7. doi: 10.1073/pnas.1417508112
12. Alisson-Silva F, Kawanishi K, Varki A. Human risk of diseases associated with red meat intake: Analysis of current theories and proposed role for metabolic incorporation of a non-human sialic acid. Mol Aspects Med (2016) 51:16–30. doi: 10.1016/j.mam.2016.07.002
13. Diaz SL, Padler-Karavani V, Ghaderi D, Hurtado-Ziola N, Yu H, Chen X, et al. Sensitive and specific detection of the non-human sialic Acid N-glycolylneuraminic acid in human tissues and biotherapeutic products. PloS One (2009) 4:e4241. doi: 10.1371/journal.pone.0004241
14. Pham T, Gregg CJ, Karp F, Chow R, Padler-Karavani V, Cao H, et al. Evidence for a novel human-specific xeno-auto-antibody response against vascular endothelium. Blood (2009) 114:5225–35. doi: 10.1182/blood-2009-05-220400
15. Varki A. Multiple changes in sialic acid biology during human evolution. Glycoconj J (2009) 26:231–45. doi: 10.1007/s10719-008-9183-z
16. Blanco R, Rengifo E, Cedeno M, Rengifo CE, Alonso DF, Carr A. Immunoreactivity of the 14F7 mab raised against N-glycolyl GM3 ganglioside in epithelial Malignant tumors from digestive system. ISRN Gastroenterol (2011) 2011:645641. doi: 10.5402/2011/645641
17. Blanco R, Rengifo E, Rengifo CE, Cedeno M, Frometa M, Carr A. Immunohistochemical reactivity of the 14F7 monoclonal antibody raised against N-glycolyl GM3 ganglioside in some benign and Malignant skin neoplasms. ISRN Dermatol (2011) 2011:848909. doi: 10.5402/2011/848909
18. Scursoni AM, Galluzzo L, Camarero S, Pozzo N, Gabri MR, de Acosta CM, et al. Detection and characterization of N-glycolyated gangliosides in Wilms tumor by immunohistochemistry. Pediatr Dev Pathol (2010) 13:18–23. doi: 10.2350/08-10-0544.1
19. Scursoni AM, Galluzzo L, Camarero S, Lopez J, Lubieniecki F, Sampor C, et al. Detection of N-glycolyl GM3 ganglioside in neuroectodermal tumors by immunohistochemistry: an attractive vaccine target for aggressive pediatric cancer. Clin Dev Immunol (2011) 2011:245181. doi: 10.1155/2011/245181
20. Dorvignit D, Garcia-Martinez L, Rossin A, Sosa K, Viera J, Hernandez T, et al. Antitumor and cytotoxic properties of a humanized antibody specific for the GM3(Neu5Gc) ganglioside. Immunobiology (2015) 220:1343–50. doi: 10.1016/j.imbio.2015.07.008
21. Labrada M, Dorvignit D, Hevia G, Rodriguez-Zhurbenko N, Hernandez AM, Vazquez AM, et al. GM3(Neu5Gc) ganglioside: an evolution fixed neoantigen for cancer immunotherapy. Semin Oncol (2018) 45:41–51. doi: 10.1053/j.seminoncol.2018.04.003
22. Dorvignit D, Boligan KF, Relova-Hernandez E, Clavell M, Lopez A, Labrada M, et al. Antitumor effects of the GM3(Neu5Gc) ganglioside-specific humanized antibody 14F7hT against Cmah-transfected cancer cells. Sci Rep (2019) 9:9921. doi: 10.1038/s41598-019-46148-1
23. Carr A, Mullet A, Mazorra Z, Vazquez AM, Alfonso M, Mesa C, et al. A mouse IgG1 monoclonal antibody specific for N-glycolyl GM3 ganglioside recognized breast and melanoma tumors. Hybridoma (2000) 19:241–7. doi: 10.1089/02724570050109639
24. Rojas G, Talavera A, Munoz Y, Rengifo E, Krengel U, Angstrom J, et al. Light-chain shuffling results in successful phage display selection of functional prokaryotic-expressed antibody fragments to N-glycolyl GM3 ganglioside. J Immunol Methods (2004) 293:71–83. doi: 10.1016/j.jim.2004.07.002
25. Behrens AJ, Duke RM, Petralia LM, Harvey DJ, Lehoux S, Magnelli PE, et al. Glycosylation profiling of dog serum reveals differences compared to human serum. Glycobiology (2018) 28:825–31. doi: 10.1093/glycob/cwy070
26. Blanco R, Quintana Y, Blanco D, Cedeno M, Rengifo CE, Frometa M, et al. Tissue reactivity of the 14F7 mab raised against N-glycolyl GM3 ganglioside in tumors of neuroectodermal, mesodermal, and epithelial origin. J biomark (2013) 2013:602417. doi: 10.1155/2013/602417
27. Lahera T, Calvo A, Torres G, Rengifo CE, Quintero S, Arango Mdel C, et al. Prognostic role of 14F7 mab immunoreactivity against N-glycolyl GM3 ganglioside in colon cancer. J Oncol (2014) 2014:482301. doi: 10.1155/2014/482301
28. Blanco R, Dominguez E, Morales O, Blanco D, Martinez D, Rengifo CE, et al. Prognostic significance of N-glycolyl GM3 ganglioside expression in non-small cell lung carcinoma patients: new evidences. Patholog Res Int (2015) 2015:132326. doi: 10.1155/2015/132326
29. Cheresh DA, Reisfeld RA, Varki AP. O-acetylation of disialoganglioside GD3 by human melanoma cells creates a unique antigenic determinant. Science (1984) 225:844–6. doi: 10.1126/science.6206564
30. Cheresh DA, Varki AP, Varki NM, Stallcup WB, Levine J, Reisfeld RA. A monoclonal antibody recognizes an O-acylated sialic acid in a human melanoma-associated ganglioside. J Biol Chem (1984) 259:7453–9. doi: 10.1016/S0021-9258(17)42812-2
31. Taylor RE, Gregg CJ, Padler-Karavani V, Ghaderi D, Yu H, Huang S, et al. Novel mechanism for the generation of human xeno-autoantibodies against the nonhuman sialic acid N-glycolylneuraminic acid. J Exp Med (2010) 207:1637–46. doi: 10.1084/jem.20100575
32. Samraj AN, Laubli H, Varki N, Varki A. Involvement of a non-human sialic Acid in human cancer. Front Oncol (2014) 4:33. doi: 10.3389/fonc.2014.00033
33. Fernandez-Marrero Y, Roque-Navarro L, Hernandez T, Dorvignit D, Molina-Perez M, Gonzalez A, et al. A cytotoxic humanized anti-ganglioside antibody produced in a murine cell line defective of N-glycolylated-glycoconjugates. Immunobiology (2011) 216:1239–47. doi: 10.1016/j.imbio.2011.07.004
Keywords: sialic acid, glycolipids, N-acetyl-neuraminic acid, N-glycolyl-neuraminic acid, cancer
Citation: Manni M, Mantuano NR, Zingg A, Kappos EA, Behrens A-J, Back J, Follador R, Faridmoayer A and Läubli H (2023) Detection of N-glycolyl-neuraminic acid-containing glycolipids in human skin. Front. Immunol. 14:1291292. doi: 10.3389/fimmu.2023.1291292
Received: 08 September 2023; Accepted: 10 November 2023;
Published: 29 November 2023.
Edited by:
Stevan Springer, University of Prince Edward Island, CanadaReviewed by:
Jeremy Alastair O’Sullivan, Northwestern University, United StatesCopyright © 2023 Manni, Mantuano, Zingg, Kappos, Behrens, Back, Follador, Faridmoayer and Läubli. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Heinz Läubli, aGVpbnoubGFldWJsaUB1bmliYXMuY2g=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.