
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
CORRECTION article
Front. Immunol., 27 September 2023
Sec. Molecular Innate Immunity
Volume 14 - 2023 | https://doi.org/10.3389/fimmu.2023.1283161
 Suado M. Abdillahi1*
Suado M. Abdillahi1* Ramesh Tati1Sara L. Nordin1Maria Baumgarten1Oskar Hallgren2
Ramesh Tati1Sara L. Nordin1Maria Baumgarten1Oskar Hallgren2 Leif Bjermer2Jonas Erjefält3
Leif Bjermer2Jonas Erjefält3 Gunilla Westergren-Thorsson4
Gunilla Westergren-Thorsson4 Birendra Singh5
Birendra Singh5 Kristian Riesbeck5*†Matthias Mörgelin1,6†
Kristian Riesbeck5*†Matthias Mörgelin1,6†by Abdillahi SM, Tati R, Nordin SL, Baumgarten M, Hallgren O, Bjermer L, Erjefä J, Westergren-Thorsson G, Singh B, Riesbeck K and Mörgelin M (2018) Front. Immunol. 9:1988. doi: 10.3389/fimmu.2018.01988
In the published article, there was an error in Figure 3 as published. Panel 3D does not show the correct bacterial specimen. The corrected Figure 3 and its caption “Targeting of NTHi surface adhesins PE and Hap by collagen VI VWA domains. (A) Titration of bacterial solutions with radiolabeled collagen VI microfibrils. Serial dilutions of bacteria were used: 1% (2 × 109 cfu/ml), 0.5% (1 × 109 cfu/ml), 0.1% (2 × 108 cfu/ml), 0.01% (2 × 107 cfu/ml), and 0.001% (2 × 106 cfu/ml). Wild type bacteria are compared to isogenic mutants as indicated. (B–F) negative staining and transmission electron microscopy of collagen VI networks bound to the bacterial surface. Wild type (B) and Δhia (C) bacteria interact with collagen VI (arrows) as opposed to Δhpe (D), Δhap (E), and ΔhpeΔhap (F). PE (G) and Hap (H) are frequently colocalized with collagen VI on the bacterial surface as visualized by antibodies conjugated with 5 nm (PE and Hap, arrowheads) and 10 nm (collagen VI, arrows) colloidal gold, respectively. The scale bars represent 200 nm (B–F) and 100 nm (G, H).” appear below.
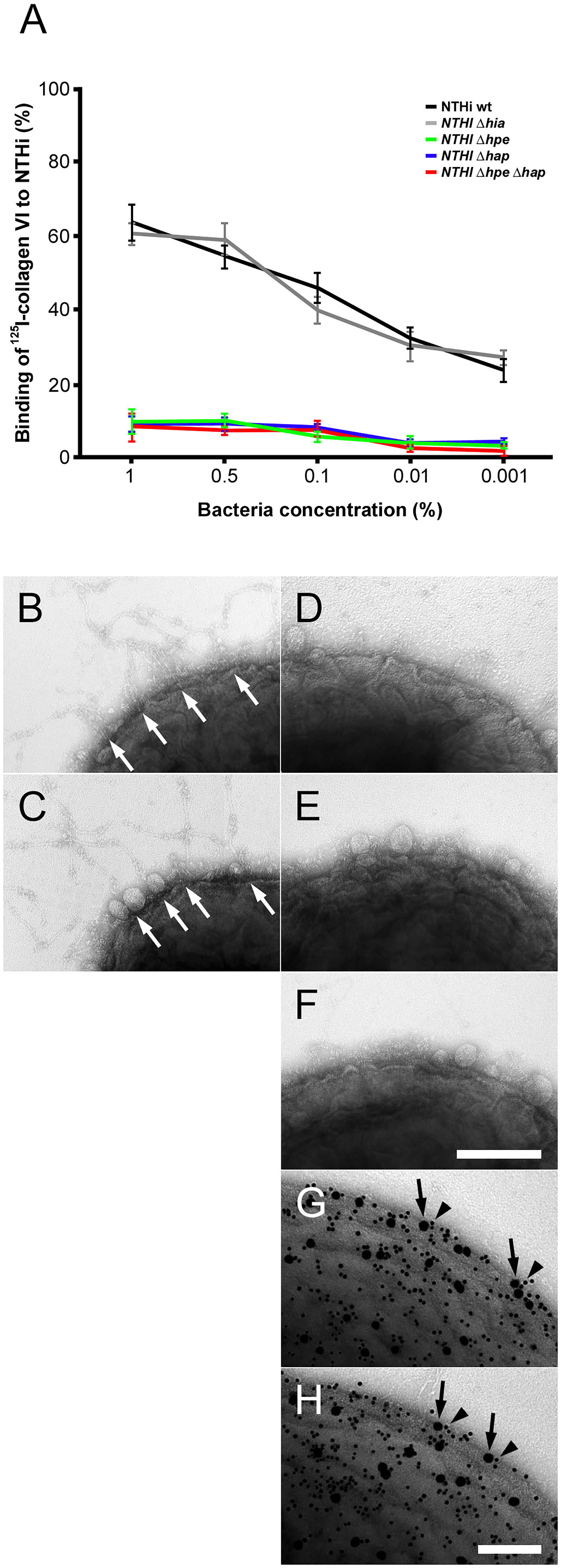
Figure 3 Targeting of NTHi surface adhesins PE and Hap by collagen VI VWA domains. (A) Titration of bacterial solutions with radiolabeled collagen VI microfibrils. Serial dilutions of bacteria were used: 1% (2 × 109 cfu/ml), 0.5% (1 × 109 cfu/ml), 0.1% (2 × 108 cfu/ml), 0.01% (2 × 107 cfu/ml), and 0.001% (2 × 106 cfu/ml). Wild type bacteria are compared to isogenic mutants as indicated. (B–F) negative staining and transmission electron microscopy of collagen VI networks bound to the bacterial surface. Wild type (B) and Δhia (C) bacteria interact with collagen VI (arrows) as opposed to Δhpe (D), Δhap (E), and ΔhpeΔhap (F). PE (G) and Hap (H) are frequently colocalized with collagen VI on the bacterial surface as visualized by antibodies conjugated with 5 nm (PE and Hap, arrowheads) and 10 nm (collagen VI, arrows) colloidal gold, respectively. The scale bars represent 200 nm (B–F) and 100 nm (G, H).
The authors apologize for this error and state that this does not change the scientific conclusions of the article in any way. The original article has been updated.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Keywords: antimicrobial activity, bronchopulmonary infection, collagen VI, COPD, extracellular matrix, Haemophilus influenzae, innate immunity, pulmonary fibrosis
Citation: Abdillahi SM, Tati R, Nordin SL, Baumgarten M, Hallgren O, Bjermer L, Erjefält J, Westergren-Thorsson G, Singh B, Riesbeck K and Mörgelin M (2023) Corrigendum: The pulmonary extracellular matrix is a bactericidal barrier against Haemophilus influenzae in chronic obstructive pulmonary disease (COPD): implications for an in vivo innate host defense function of collagen VI. Front. Immunol. 14:1283161. doi: 10.3389/fimmu.2023.1283161
Received: 25 August 2023; Accepted: 12 September 2023;
Published: 27 September 2023.
Edited and Reviewed by:
Uday Kishore, United Arab Emirates University, United Arab EmiratesCopyright © 2023 Abdillahi, Tati, Nordin, Baumgarten, Hallgren, Bjermer, Erjefält, Westergren-Thorsson, Singh, Riesbeck and Mörgelin. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Suado M. Abdillahi, c3VhZG8ubV9hYmRpbGxhaGlAbWVkLmx1LnNl; Kristian Riesbeck, a3Jpc3RpYW4ucmllc2JlY2tAbWVkLmx1LnNl
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.