 Kerry L. Hilligan
Kerry L. Hilligan Sivaranjani Namasivayam
Sivaranjani Namasivayam Alan Sher1*
Alan Sher1*
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
PERSPECTIVE article
Front. Immunol. , 08 September 2023
Sec. Microbial Immunology
Volume 14 - 2023 | https://doi.org/10.3389/fimmu.2023.1232764
This article is part of the Research Topic The Covid-19 and TB Syndemic: Differences and Similarities View all 11 articles
The observation of reduced COVID-19 incidence and severity in populations receiving neonatal intradermal BCG vaccination vaccine raised the question of whether BCG can induce non-specific protection against the SARS-CoV-2 (SCV2) virus. Subsequent epidemiologic studies and clinical trials have largely failed to support this hypothesis. Furthermore, in small animal model studies all investigators have failed to observe resistance to viral challenge in response to BCG immunization by the conventional and clinically acceptable intradermal or subcutaneous routes. Nevertheless, BCG administered by the intravenous (IV) route has been shown to strongly protect both hamsters and mice against SCV2 infection and disease. In this Perspective, we review the current data on the effects of BCG vaccination on resistance to COVID-19 as well as summarize recent work in rodent models on the mechanisms by which IV administered BCG promotes resistance to the virus and discuss the translational implications of these findings.
The innate compartment of the immune system differs from the adaptive in its ability to provide non-specific defense against a wide variety of threats encountered by the body and its stimulation is an important strategy for enhancing host resistance to pathogens. Avirulent microbes and their products are themselves important triggers of innate immune function and recently have been described to do so with long term effects (1). BCG (Bacille Calmette Guérin) is a well-studied microbial stimulus for its effects on innate immunity. This attenuated Mycobacterium bovis strain is widely used to vaccinate against extrapulmonary tuberculosis (TB) in infants and children and was discovered in the mid-20th century to also promote non-specific resistance against tumors, a finding that led to its current employment as a treatment for some forms of bladder cancer (2). More recently BCG vaccination has been associated with lowering all-cause mortality in infants (3), reducing viremia after a yellow fever vaccine challenge in adults (4), and decreasing risk of respiratory infections in the elderly (5). Multiple mechanisms have been proposed to explain these effects, the most prominent of which involve the induction of “trained immunity” in which myelopoietic-derived innate effector cells become epigenetically modified so that they remain in a long-term primed state (up to 1 year in humans) (6–8).
BCG is typically administered to vaccinees by intradermal (ID) or subcutaneous (SC) injection although other routes (e.g. oral) have been employed in the past (9, 10). Although not clinically approved, the intravenous (IV) route of BCG administration has recently been employed in two important studies related to TB vaccination. In the first study, Kaufmann and colleagues showed that IV BCG preferentially induces trained immunity in mice because of its ability to access and infect long lived myelopoietic stem cells in the bone marrow (7). In the second report, Darrah and colleagues showed that IV in contrast to SC administered BCG induces sterile immunity against M. tuberculosis (Mtb) challenge in a rhesus monkey model (11), a dramatic finding that the authors attributed to the direct targeting of the lung and the induction of a strong local memory T cell response when the vaccine is given by this route (12). Recent studies indicate that in macaques such resistance can persist after the clearance of culturable BCG bacilli (13). Nevertheless, the contribution of BCG stimulated innate immune mechanisms to this striking protection is at present unclear.
Given its previously demonstrated ability to stimulate non-specific host resistance to certain other viral infections, BCG immunization was suggested in the early months of the COVID-19 pandemic as a possible prophylactic measure for the prevention of SCV2 infection and disease (14, 15). This concept was initially supported by a number of ecological/epidemiologic studies suggesting an association of prior BCG vaccination with a lower incidence of COVID-19 disease (16, 17) despite the relatively short period (up to 1 year) that “trained” responses have been reported to persist in vivo (8). This early work was followed up with a large number of more extensive investigations (summarized in Table 1) that in general have failed to confirm the protective effects of BCG vaccination on the incidence and severity of SCV2 infection (18–21, 26–29), including a recently published international multi-cohort randomized trial (BRACE) involving ID administration of BCG to adult health care workers (22). One study conducted with a small cohort of older adults in Greece did note some protection against the incidence of COVID-19 symptoms; however, the existence of SCV2 infection in these individuals was not confirmed by PCR or antibody testing (24). A significant reduction in the incidence and symptom severity of COVID-19 was also observed in a different study involving the follow-up of adult diabetes patients given 3 ID doses of intramural BCG over a 2-3 year period before the onset of the pandemic (25). The explanation for the unusual efficacy observed in the latter study is unclear but may relate to the multiple dosage, the use of a highly virulent BCG isolate (Tokyo strain) (30, 31), the spacing between BCG vaccination and SCV2 exposure, or possibly the diabetic state of the participants. Overall, there is currently no compelling evidence that a single-dose intradermal BCG inoculation provides protection against SCV2 infection and disease; however, there may be certain conditions that favor the protective outcomes observed with multiple BCG doses (25). Future studies examining prolonged or repeated mycobacterial exposures, either due to population level exposure to environmental mycobacteria and/or BCG re-vaccination strategies, may provide further insights into any potential protective effects (32).
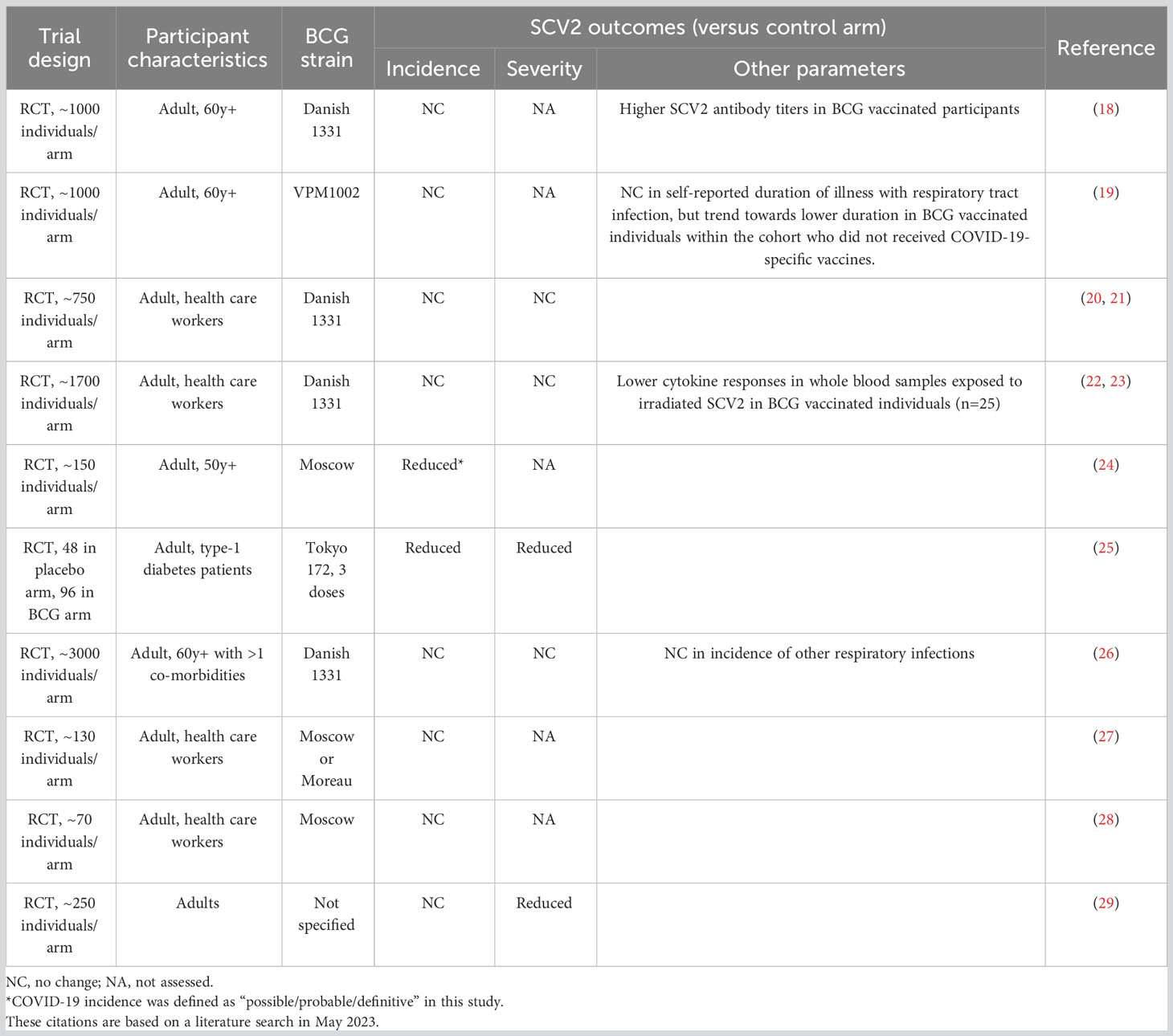
Table 1 Summary of human trials investigating BCG efficacy against COVID-19.
The hypothesis that prior BCG vaccination might offer protection against COVID-19 prompted a series of studies in different animal models to examine the effects of prior BCG administration on resistance to SCV2 challenge (Table 2). This work has generated a consensus that when inoculated by the conventional ID (or subcutaneous) route to mice (33, 35–37) or hamsters (35, 38) or by aerosol to monkeys (39), BCG fails to trigger significant protection against intranasal or intra-bronchial infection with the virus. Nevertheless, a number of independent studies have shown that when administered by the IV route to mice or hamsters, BCG can confer high levels of resistance to both SCV2 infection and disease (Table 2) (33, 34, 38, 40, 41). In the initial description of this effect, K18 transgenic mice which express the human ACE2 receptor (K18-hACE2) for the virus were IV inoculated with BCG (Pasteur strain) before intranasal SCV2 infection with a lethal dose of the WA/2020 strain (33). At 42 days following BCG administration, the virus challenged mice showed a striking protection from SCV2 induced weight loss and mortality along with pronounced reductions in pulmonary viral loads at 5 days post infection. This protection was still evident 112 days following BCG inoculation but at lower levels (33). To confirm that the COVID-19 resistance induced by IV BCG is not peculiar to hACE2 transgenic mice, the experiments were repeated using a second model in which wild type C57BL/6 mice were challenged with the more virulent B.1.1.7 SCV2 variant. In this situation unvaccinated mice support viral replication for 3-4 days before clearing the infection with minimal accompanying disease. Again, IV BCG induced striking protection against SCV2 with the majority of the BCG exposed mice showing no detectable virus in their lungs at 3 days following B.1.1.7 challenge (33). Consistent with the other studies cited above, no significant resistance against SCV2 was observed in mice inoculated with the same dose of BCG by the SC route in either of the two murine models. The ability of IV BCG to protect K18-hACE2 mice from early SCV2 infection was confirmed in a second study using the Tokyo strain of BCG and intranasal viral challenge with either an original “wild-type” strain or more virulent kappa or delta variants (40). In additional work, IV administered BCG (Tice strain) was shown to reduce viral loads and bronchopneumonia in Syrian hamsters challenged intranasally with the Wuhan-1 strain SCV2 (38). In contrast to the above findings, Kaufmann et al. reported that K18-hACE2 mice or hamsters given IV (or SC) BCG (Tice strain) showed no significant protection against intranasal (or in the case of mice either intranasal or intratracheal) challenge with a SCV2-B lineage variant. Nevertheless, the same BCG exposed mice displayed resistance to intranasally administered Influenza A virus (35). Ongoing follow up studies suggest that the negative results with SCV2 obtained in the latter study may relate to the BCG strain (42), its preparation and/or the dose employed for vaccination (Kaufmann and Hilligan, unpublished).
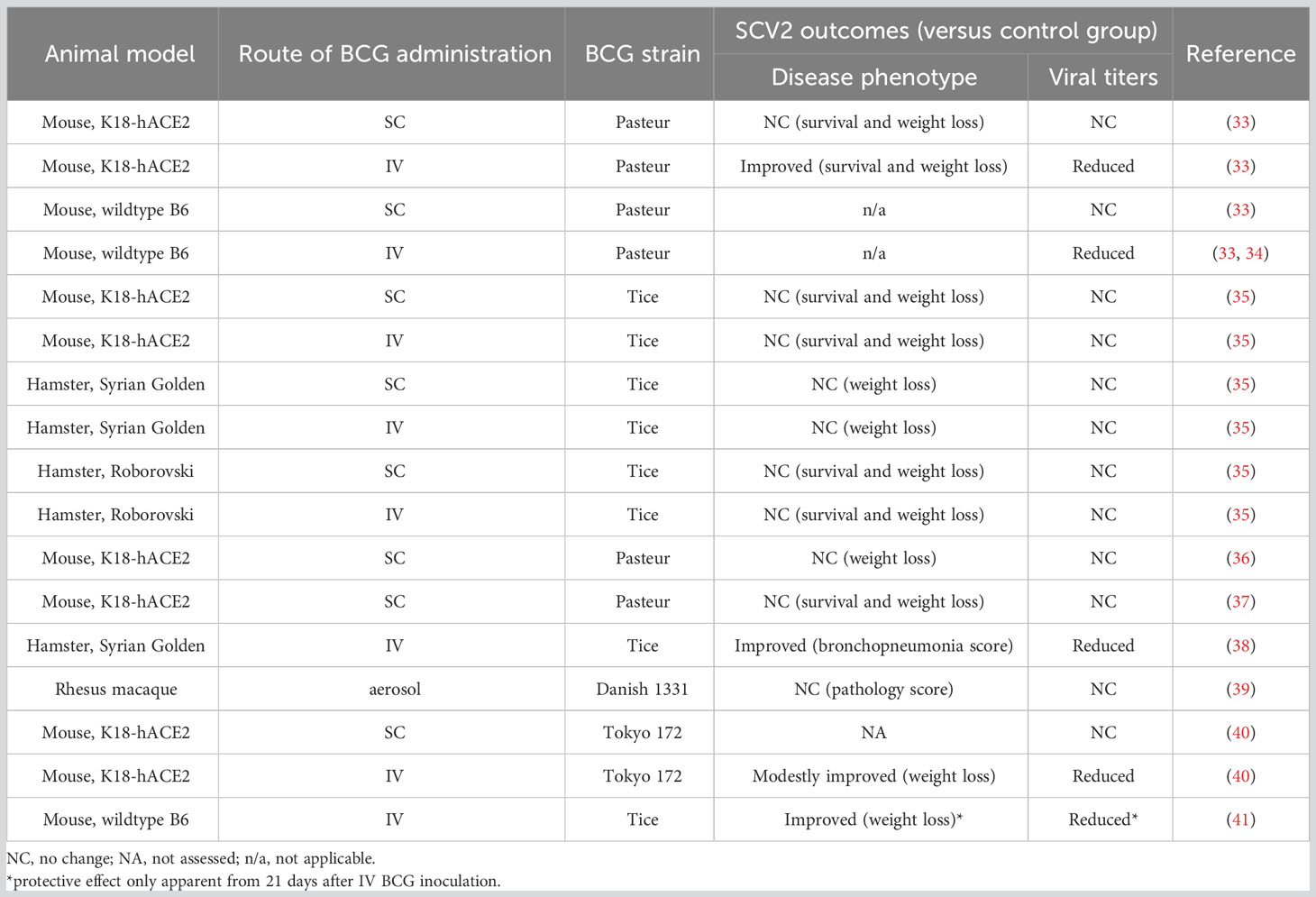
Table 2 Summary of animal studies assessing efficacy of BCG against SCV2 infection and disease.
The consistent failure of SC or ID inoculated BCG to provide protection against SCV2 infection suggests that the resistance conferred by IV BCG may relate to the long-term presence mycobacteria in the lungs and accompanying granulomatous inflammation occurring in animals inoculated by that route (33, 40). Consistent with this hypothesis, K18-hACE2 or non-transgenic mice infected by aerosol with virulent Mycobacterium tuberculosis and developing pulmonary TB, display high levels of resistance to SCV2 comparable to that reported in IV BCG exposed animals (37, 43, 44). Nevertheless, as noted above, in rhesus macaques BCG given by the aerosol route failed to induce protection against SCV2 challenge (39). Since pulmonary bacterial infection and local tissue responses were not evaluated in that study, it is difficult to ascertain whether this discrepancy with the rodent studies reflects the different host species employed or the local levels of BCG and/or immune responses occurring at that site. Indeed, a comparison between IV and aerosol inoculation of rhesus macaques by Darrah et al, showed that only IV BCG resulted in the formation of “microgranuloma” structures in the lung as well as increased numbers of CD4+ T cells and CD11c+ antigen-presenting cells (11).
It was originally proposed that ID (or SC) administered BCG might offer protection against COVID-19 because of its previously documented ability to enhance clinical resistance to other viral infections, effects that were attributed to the induction of trained immunity (14, 15). Since in nearly all studies humans vaccinated with BCG by this route fail to display significant resistance to COVID-19, it would appear that any response induced by a single-dose BCG inoculation is not sufficient to restrict SCV2. Nevertheless, it is still possible that boosting of the response by intradermal re-vaccination could induce more effective immunity and this could be the basis of the protection against COVID-19 observed by Faustman and colleagues in diabetes patients given multiple BCG inoculations (25).
Since with the latter exception BCG induced protection against SCV2 has not been documented in humans or non-human primates, nearly all the current information on anti-viral mechanisms derives from the studies on murine and hamster rodents involving IV administered bacteria. That route of inoculation has been previously shown in mice to preferentially stimulate myelopoiesis and the generation of monocyte/macrophages with a trained phenotype (7). Consistent with these earlier findings, Zhang and colleagues reported that IV BCG vaccinated mice challenged with SCV2 display enhanced bone marrow myelopoiesis, augmented pulmonary monocyte/macrophage infiltration and upregulated innate immune and metabolic gene signatures previously described as associated with training (40). Although not specifically addressing the issue of trained immunity, both the NIH murine model study of Hilligan et al. and hamster study of Singh and colleagues described enhanced pulmonary macrophage numbers in IV BCG inoculated animals that likely arise from bone marrow monocytes (33, 38). Given the long-term persistence of both mycobacteria and granulomatous inflammation in the lungs of IV BCG vaccinated mice (33, 40), it is unlikely that resistance to SCV2 challenge would require the type of trained myeloid cells previously described as arising in hosts exposed to a prior single intradermal bacterial inoculation.
In each of the three studies documenting protection against SCV2 induced by IV BCG, vaccination was shown to simultaneously reduce pulmonary viral load and virus induced bronchopneumonia, in some cases as early as 2 days post challenge. Consistent with the latter observation, in both mouse studies BCG inoculation resulted in lowered production of SCV2 induced IL-6 and MCP1 (CCL2) (33, 40). Although this decrease could reflect an effect of reduced viral load in the vaccinated animals, the results of a multivariate analysis performed in the NIH murine study revealed an inhibitory effect of prior IV BCG administration on the induction of these pathology associated cytokines independent of viral titer (33). These data align with results from the BRACE clinical trial that showed that while BCG vaccination did not protect against COVID-19 (22), BCG did limit SCV2-induced pro-inflammatory cytokine responses ex vivo, suggesting that BCG inoculation can modulate virus triggered immune responses independent of its protective effect (23).
In both mice and hamsters, IV BCG administration led to pronounced elevations in pulmonary T cells, while only a minor response was seen in mice given SC BCG. In mice, IV BCG enhanced lymphocytes were characterized as CD8+, FoxP3− CD4+, and FoxP3+ CD4+ T cells, as well as MAIT cells and their levels did not significantly increase following viral challenge (33, 38). Indeed, if anything, prior IV BCG administration appeared to suppress the CD8+ T cell expansion triggered by SCV2 infection. Somewhat in contrast, in the hamster model, prior IV BCG inoculation resulted in an expansion of cells with Th1, Th17, Treg, CTLs or Tmem transcriptional markers after viral challenge as well as the emergence of a new plasma cell population not present prior to SCV2 exposure and expressing genes associated with immunoglobulin production suggestive of accelerated antibody production. In the same hamster study, IV BCG vaccination also appeared to dampen the expression of T cell exhaustion markers triggered by SCV2 infection (38). Together these observations show that IV BCG triggers the recruitment of adaptive immune cells into the lung tissue that in addition to supplying a potential source of protective antibodies may be important in providing cytokines and other signals that shape the innate immune landscape. Another interesting possibility is that the response to the bacteria has hindered the ability of the host to respond to another inflammatory stimuli.
Type I IFNs are important for control of viral pathogens but in SCV2 and other virus infections these cytokines can also promote pathology (45, 46). Interestingly in the NIH mouse model study prior IV BCG inoculation appeared to suppress rather than augment the SCV2 triggered Type I IFN response consistent with the suppression of COVID-19-like pathology observed in these animals. BCG infection is classically associated with strong IFNγ production from CD4+ T, CD8+ T and NK cells and the cytokine was found to be heavily induced in the lungs of both mice and hamsters months after IV BCG inoculation (33, 38, 40). Importantly, this local Type II IFN response was minimal in mice vaccinated by the SC route consistent with the dearth of both BCG and its associated granulomatous tissue inflammation in lungs of these animals in contrast to IV inoculated mice. Recent functional studies in the murine models suggest that this IFNγ response deriving primarily from CD4+ T cells and acting on non-hematopoietic cells in the lung is required for the reduction in both SCV2 virus and its associated pathology and that the recombinant cytokine itself can trigger these effects (34, 41). Whether IV BCG induced protection against SCV2 is mediated entirely through this mechanism or also involves the myeloid, T or B lymphocytes changes reported to be associated with resistance in the studies discussed above is at present unclear. A summary of the different effector mechanisms currently proposed to explain the protection against SCV2 induced by IV BCG is presented in Figure 1.
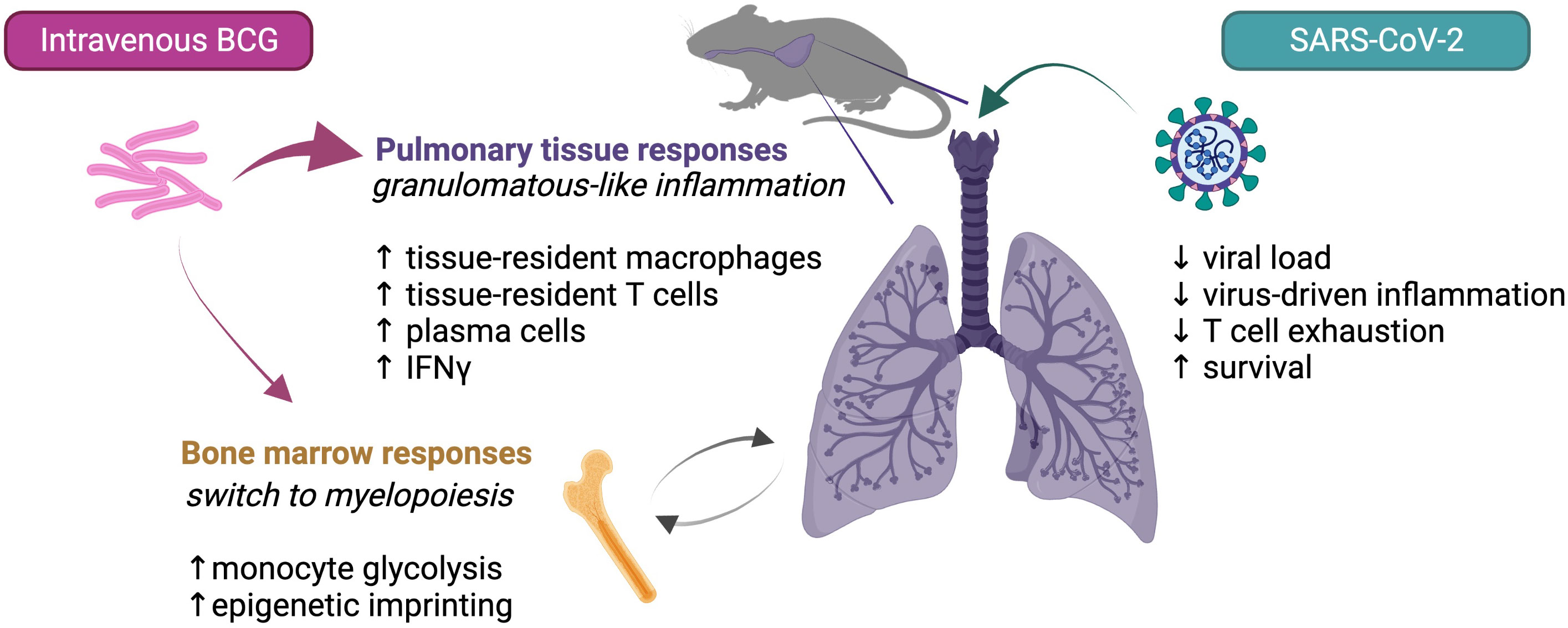
Figure 1 Possible mechanisms contributing to IV BCG conferred protection against SCV2 in mice.
The findings reviewed above establish a proof of principle in animal models that single dose BCG can stimulate protection against SCV2 but only when given IV, a mode of administration that is currently not clinically acceptable. The data do not rule out the possibility that through repeated boosting (25) or the use of a specially engineered bacterial strain (36) protection against COVID-19 could be generated through conventional ID or SC vaccination although it is likely that such resistance would involve a different mechanism. There is currently considerable interest in the possible use of IV administered BCG for vaccination against M. tuberculosis because of its ability to confer sterile immunity against this important pathogen in rhesus monkeys (11, 13). This has stimulated efforts to develop attenuated BCG mutants (e.g. auxotrophs) that would be safe for human intravenous use and such strains could be tested as candidates for protection against COVID-19 (47).
Regardless, the demonstration that bacterial stimulation of the lung can induce high levels of resistance against SCV2 could lead to the discovery of novel mechanisms of anti-viral protection with potential clinical applicability. For example, the recent evidence that BCG induced IFNγ can protect mice from SCV2 challenge (34, 41) raises the question of whether the cytokine could be used intranasally to protect subjects at high risk of infection possibly with less risk of toxicity than Type I IFN. It is also becoming clear that IV BCG is not a unique non-specific stimulus for host protection against experimental SCV2. In addition to prior M. tuberculosis infection (37, 43, 44), recent findings indicate that intranasally administered PRR ligands can also trigger host resistance in the same murine models (48–50) as can prior infection with a lung-transiting helminth (51). While seemingly distinct stimuli, it is possible that they all act by triggering the production of anti-viral effectors by pulmonary myeloid or epithelial cells.
As noted in the studies reviewed here, IV BCG infection can trigger long term changes in the cellular composition and adaptive immune responsiveness of lung tissue. While trained immunity may contribute [recently reviewed by Netea et al. (52)], other factors such as bacterial induced tissue remodeling and continuous immune stimulation by the bacteria surviving within granuloma-like structures in the lung are in this situation likely to play a more important role in promoting the long-lived property of the protection triggered by IV BCG at that tissue site (Figure 1).
Despite its limitations as a vaccine, studies on BCG continue to provide important insights on the interplay of innate and adaptive immunity in the host response to pathogens and in this case hopefully add to our understanding of how the lung can be stimulated to control both SCV2 and COVID-19 associated pathology.
Writing-draft: KH, AS. Writing-review and editing: KH, SN, AS. All authors contributed to the article and approved the submitted version.
KH, SN, AS were supported by the Intramural Research Program of the NIAID.
The authors thank Drs Katrin Mayer-Barber, Sonja Best, Stefan Kaufmann, Maziar Divangahi, Eva Kaufmann and Patricia Darrah for helpful discussion. Figure was created using BioRender.com.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Dominguez-Andres J, Dos Santos JC, Bekkering S, Mulder WJM, van der Meer JWM, Riksen NP, et al. Trained immunity: adaptation within innate immune mechanisms. Physiol Rev (2023) 103(1):313–46. doi: 10.1152/physrev.00031.2021
2. Pettenati C, Ingersoll MA. Mechanisms of BCG immunotherapy and its outlook for bladder cancer. Nat Rev Urol (2018) 15(10):615–25. doi: 10.1038/s41585-018-0055-4
3. Biering-Sorensen S, Aaby P, Lund N, Monteiro I, Jensen KJ, Eriksen HB, et al. Early BCG-Denmark and neonatal mortality among infants weighing <2500 g: A randomized controlled trial. Clin Infect Dis (2017) 65(7):1183–90. doi: 10.1093/cid/cix525
4. Arts RJW, Moorlag S, Novakovic B, Li Y, Wang SY, Oosting M, et al. BCG Vaccination Protects against Experimental Viral Infection in Humans through the Induction of Cytokines Associated with Trained Immunity. Cell Host Microbe (2018) 23(1):89–100.e5. doi: 10.1016/j.chom.2017.12.010
5. Giamarellos-Bourboulis EJ, Tsilika M, Moorlag S, Antonakos N, Kotsaki A, Dominguez-Andres J, et al. Activate: randomized clinical trial of BCG vaccination against infection in the elderly. Cell (2020) 183(2):315–23.e9. doi: 10.1016/j.cell.2020.08.051
6. Cirovic B, de Bree LCJ, Groh L, Blok BA, Chan J, van der Velden W, et al. BCG vaccination in humans elicits trained immunity via the hematopoietic progenitor compartment. Cell Host Microbe (2020) 28(2):322–34.e5. doi: 10.1016/j.chom.2020.05.014
7. Kaufmann E, Sanz J, Dunn JL, Khan N, Mendonca LE, Pacis A, et al. BCG educates hematopoietic stem cells to generate protective innate immunity against tuberculosis. Cell (2018) 172(1-2):176–90.e19. doi: 10.1016/j.cell.2017.12.031
8. Kleinnijenhuis J, Quintin J, Preijers F, Benn CS, Joosten LA, Jacobs C, et al. Long-lasting effects of BCG vaccination on both heterologous Th1/Th17 responses and innate trained immunity. J Innate Immun (2014) 6(2):152–8. doi: 10.1159/000355628
9. Cho T, KhatChadourian C, Nguyen H, Dara Y, Jung S, Venketaraman V. A review of the BCG vaccine and other approaches toward tuberculosis eradication. Hum Vaccin Immunother (2021) 17(8):2454–70. doi: 10.1080/21645515.2021.1885280
10. Calmette A, Guerin C BW-H. Essai d'immunisation contre l'infection tuberculeuse. Bull Acad Med (1924) 91:787–96.
11. Darrah PA, Zeppa JJ, Maiello P, Hackney JA, Wadsworth MH 2nd, Hughes TK, et al. Prevention of tuberculosis in macaques after intravenous BCG immunization. Nature (2020) 577(7788):95–102. doi: 10.1038/s41586-019-1817-8
12. Darrah PA, Zeppa JJ, Wang C, Irvine EB, Bucsan AN, Rodgers MA, et al. Airway T cells are a correlate of i.v. Bacille Calmette-Guerin-mediated protection against tuberculosis in rhesus macaques. Cell Host Microbe (2023) 31(6):962–77 e8. doi: 10.1016/j.chom.2023.05.006
13. Larson E, Ellis-Connell A, Rodgers M, Gubernat A, Gleim J, Moriarty R, et al. Vaccination with intravenous BCG protects macaques with pre-existing SIV infection from tuberculosis. Res Sq (2023). doi: 10.21203/rs.3.rs-2802306/v1
14. O'Neill LAJ, Netea MG. BCG-induced trained immunity: can it offer protection against COVID-19? Nat Rev Immunol (2020) 20(6):335–7. doi: 10.1038/s41577-020-0337-y
15. Curtis N, Sparrow A, Ghebreyesus TA, Netea MG. Considering BCG vaccination to reduce the impact of COVID-19. Lancet (2020) 395(10236):1545–6. doi: 10.1016/S0140-6736(20)31025-4
16. Escobar LE, Molina-Cruz A, Barillas-Mury C. BCG vaccine protection from severe coronavirus disease 2019 (COVID-19). Proc Natl Acad Sci USA (2020) 117(30):17720–6. doi: 10.1073/pnas.2008410117
17. Rivas MN, Ebinger JE, Wu M, Sun N, Braun J, Sobhani K, et al. BCG vaccination history associates with decreased SARS-CoV-2 seroprevalence across a diverse cohort of health care workers. J Clin Invest (2021) 131(2):e147157. doi: 10.1172/JCI145157
18. Moorlag S, Taks E, Ten Doesschate T, van der Vaart TW, Janssen AB, Muller L, et al. Efficacy of BCG vaccination against respiratory tract infections in older adults during the coronavirus disease 2019 pandemic. Clin Infect Dis (2022) 75(1):e938–e46. doi: 10.1093/cid/ciac182
19. Blossey AM, Bruckner S, May M, Parzmair GP, Sharma H, Shaligram U, et al. VPM1002 as prophylaxis against severe respiratory tract infections including coronavirus disease 2019 in the elderly: A phase 3 randomized, double-blind, placebo-controlled, multicenter clinical study. Clin Infect Dis (2023) 76(7):1304–10. doi: 10.1093/cid/ciac881
20. Ten Doesschate T, van der Vaart TW, Debisarun PA, Taks E, Moorlag S, Paternotte N, et al. Bacillus Calmette-Guerin vaccine to reduce healthcare worker absenteeism in COVID-19 pandemic, a randomized controlled trial. Clin Microbiol Infect (2022) 28(9):1278–85. doi: 10.1016/j.cmi.2022.04.009
21. Claus J, Ten Doesschate T, Gumbs C, van Werkhoven CH, van der Vaart TW, Janssen AB, et al. BCG vaccination of health care workers does not reduce SARS-coV-2 infections nor infection severity or duration: a randomized placebo-controlled trial. mBio (2023) 14(2):e0035623. doi: 10.1128/mbio.00356-23
22. Pittet LF, Messina NL, Orsini F, Moore CL, Abruzzo V, Barry S, et al. Randomized trial of BCG vaccine to protect against covid-19 in health care workers. N Engl J Med (2023) 388(17):1582–96. doi: 10.1056/NEJMoa2212616
23. Messina NL, Germano S, McElroy R, Rudraraju R, Bonnici R, Pittet LF, et al. Off-target effects of bacillus Calmette-Guerin vaccination on immune responses to SARS-CoV-2: implications for protection against severe COVID-19. Clin Transl Immunol (2022) 11(4):e1387. doi: 10.1002/cti2.1387
24. Tsilika M, Taks E, Dolianitis K, Kotsaki A, Leventogiannis K, Damoulari C, et al. ACTIVATE-2: A double-blind randomized trial of BCG vaccination against COVID-19 in individuals at risk. Front Immunol (2022) 13:873067. doi: 10.3389/fimmu.2022.873067
25. Faustman DL, Lee A, Hostetter ER, Aristarkhova A, Ng NC, Shpilsky GF, et al. Multiple BCG vaccinations for the prevention of COVID-19 and other infectious diseases in type 1 diabetes. Cell Rep Med (2022) 3(9):100728. doi: 10.1016/j.xcrm.2022.100728
26. Koekenbier EL, Fohse K, van de Maat JS, Oosterheert JJ, van Nieuwkoop C, Hoogerwerf JJ, et al. Bacillus Calmette-Guerin vaccine for prevention of COVID-19 and other respiratory tract infections in older adults with comorbidities: a randomized controlled trial. Clin Microbiol Infect (2023) 29:781-8. doi: 10.1016/j.cmi.2023.01.019
27. Santos AP, Werneck GL, Dalvi APR, Dos Santos CC, Tierno P, Condelo HS, et al. The effect of BCG vaccination on infection and antibody levels against SARS-CoV-2-The results of ProBCG: a multicenter randomized clinical trial in Brazil. Int J Infect Dis (2023) 130:8–16. doi: 10.1016/j.ijid.2023.02.014
28. Dos Anjos LRB, da Costa AC, Cardoso A, Guimaraes RA, Rodrigues RL, Ribeiro KM, et al. Efficacy and safety of BCG revaccination with M. bovis BCG moscow to prevent COVID-19 infection in health care workers: A randomized phase II clinical trial. Front Immunol (2022) 13:841868. doi: 10.3389/fimmu.2022.841868
29. Sinha S, Ajayababu A, Thukral H, Gupta S, Guha SK, Basu A, et al. Efficacy of bacillus calmette-guerin (BCG) vaccination in reducing the incidence and severity of COVID-19 in high-risk population (BRIC): a phase III, multi-centre, quadruple-blind randomised control trial. Infect Dis Ther (2022) 11(6):2205–17. doi: 10.1007/s40121-022-00703-y
30. Angelidou A, Conti MG, Diray-Arce J, Benn CS, Shann F, Netea MG, et al. Licensed Bacille Calmette-Guerin (BCG) formulations differ markedly in bacterial viability, RNA content and innate immune activation. Vaccine (2020) 38(9):2229–40. doi: 10.1016/j.vaccine.2019.11.060
31. Ritz N, Dutta B, Donath S, Casalaz D, Connell TG, Tebruegge M, et al. The influence of bacille Calmette-Guerin vaccine strain on the immune response against tuberculosis: a randomized trial. Am J Respir Crit Care Med (2012) 185(2):213–22. doi: 10.1164/rccm.201104-0714OC
32. Singh S, Maurya RP, Singh RK. "Trained immunity" from Mycobacterium spp. exposure or BCG vaccination and COVID-19 outcomes. PloS Pathog (2020) 16(10):e1008969. doi: 10.1371/journal.ppat.1008969
33. Hilligan KL, Namasivayam S, Clancy CS, O'Mard D, Oland SD, Robertson SJ, et al. Intravenous administration of BCG protects mice against lethal SARS-CoV-2 challenge. J Exp Med (2022) 219(2):e20211862. doi: 10.1084/jem.20211862
34. Hilligan KL, Namasivayam S, Clancy CS, Baker PJ, Old SI, Peluf V, et al. Pre-existing interferon gamma conditions the lung to mediate early control of SARS-CoV-2. bioRxiv (2023) 2023:07.15.549135. doi: 10.1101/2023.07.15.549135
35. Kaufmann E, Khan N, Tran KA, Ulndreaj A, Pernet E, Fontes G, et al. BCG vaccination provides protection against IAV but not SARS-CoV-2. Cell Rep (2022) 38(10):110502. doi: 10.1016/j.celrep.2022.110502
36. Counoupas C, Johansen MD, Stella AO, Nguyen DH, Ferguson AL, Aggarwal A, et al. A single dose, BCG-adjuvanted COVID-19 vaccine provides sterilising immunity against SARS-CoV-2 infection. NPJ Vaccines (2021) 6(1):143. doi: 10.1038/s41541-021-00406-4
37. Hildebrand RE, Chandrasekar SS, Riel M, Touray BJB, Aschenbroich SA, Talaat AM. Superinfection with SARS-CoV-2 Has Deleterious Effects on Mycobacterium bovis BCG Immunity and Promotes Dissemination of Mycobacterium tuberculosis. Microbiol Spectr (2022) 10(5):e0307522. doi: 10.1128/spectrum.03075-22
38. Singh AK, Wang R, Lombardo KA, Praharaj M, Bullen CK, Um P, et al. Dynamic single-cell RNA sequencing reveals BCG vaccination curtails SARS-CoV-2 induced disease severity and lung inflammation. bioRxiv (2022). doi: 10.1101/2022.03.15.484018
39. White AD, Sibley L, Sarfas C, Morrison AL, Bewley K, Churchward C, et al. Influence of aerosol delivered BCG vaccination on immunological and disease parameters following SARS-coV-2 challenge in rhesus macaques. Front Immunol (2021) 12:801799. doi: 10.3389/fimmu.2021.801799
40. Zhang BZ, Shuai H, Gong HR, Hu JC, Yan B, Yuen TT, et al. Bacillus Calmette-Guerin-induced trained immunity protects against SARS-CoV-2 challenge in K18-hACE2 mice. JCI Insight (2022) 7(11):e157393. doi: 10.1172/jci.insight.157393
41. Lee A, Floyd K, Wu S-Y, Fang Z, Tan TK, Li C, et al. Integrated Organ Immunity: Antigen-specific CD4-T cell-derived IFN-γ induced by BCG imprints prolonged lung innate resistance against respiratory viruses. bioRxiv (2023) 2023:07.31.551354. doi: 10.1101/2023.07.31.551354
42. Zhang L, Ru HW, Chen FZ, Jin CY, Sun RF, Fan XY, et al. Variable virulence and efficacy of BCG vaccine strains in mice and correlation with genome polymorphisms. Mol Ther (2016) 24(2):398–405. doi: 10.1038/mt.2015.216
43. Rosas Mejia O, Gloag ES, Li J, Ruane-Foster M, Claeys TA, Farkas D, et al. Mice infected with Mycobacterium tuberculosis are resistant to acute disease caused by secondary infection with SARS-CoV-2. PloS Pathog (2022) 18(3):e1010093. doi: 10.1371/journal.ppat.1010093
44. Baker PJ, Amaral EP, Castro E, Bohrer AC, Torres-Juarez F, Jordan CM, et al. Co-infection of mice with SARS-CoV-2 and Mycobacterium tuberculosis limits early viral replication but does not affect mycobacterial loads. Front Immunol (2023) 14. doi: 10.3389/fimmu.2023.1240419
45. Major J, Crotta S, Llorian M, McCabe TM, Gad HH, Priestnall SL, et al. Type I and III interferons disrupt lung epithelial repair during recovery from viral infection. Science (2020) 369(6504):712–7. doi: 10.1126/science.abc2061
46. Broggi A, Ghosh S, Sposito B, Spreafico R, Balzarini F, Lo Cascio A, et al. Type III interferons disrupt the lung epithelial barrier upon viral recognition. Science (2020) 369(6504):706–12. doi: 10.1126/science.abc3545
47. Dellagostin OA, Borsuk S, Oliveira TL, Seixas FK. Auxotrophic mycobacterium bovis BCG: updates and perspectives. Vaccines (Basel) (2022) 10(5):802. doi: 10.3390/vaccines10050802
48. Humphries F, Shmuel-Galia L, Jiang Z, Wilson R, Landis P, Ng SL, et al. A diamidobenzimidazole STING agonist protects against SARS-CoV-2 infection. Sci Immunol (2021) 6(59):eabi9002. doi: 10.1126/sciimmunol.abi9002
49. Li M, Ferretti M, Ying B, Descamps H, Lee E, Dittmar M, et al. Pharmacological activation of STING blocks SARS-CoV-2 infection. Sci Immunol (2021) 6(59):eabi9007. doi: 10.1126/sciimmunol.abi9007
50. Tamir H, Melamed S, Erez N, Politi B, Yahalom-Ronen Y, Achdout H, et al. Induction of innate immune response by TLR3 agonist protects mice against SARS-coV-2 infection. Viruses (2022) 14(2):189. doi: 10.3390/v14020189
51. Oyesola OO, Hilligan KL, Namasivayam S, Howard N, Clancy CS, Zhao M, et al. Exposure to lung-migrating helminth protects against murine SARS-CoV-2 infection through macrophage-dependent T cell activation. Sci Immunol (2023) 8(86):eadf8161. doi: 10.1126/sciimmunol.adf8161
Keywords: COVID-19, Bacille Calmette-Guérin (BCG), lung, interferon gamma (IFNγ), trained immunity
Citation: Hilligan KL, Namasivayam S and Sher A (2023) BCG mediated protection of the lung against experimental SARS-CoV-2 infection. Front. Immunol. 14:1232764. doi: 10.3389/fimmu.2023.1232764
Received: 01 June 2023; Accepted: 18 August 2023;
Published: 08 September 2023.
Edited by:
Rakesh K. Singh, Banaras Hindu University, IndiaReviewed by:
Max Bastian, Friedrich-Loeffler-Institute, GermanyCopyright © 2023 Hilligan, Namasivayam and Sher. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kerry L. Hilligan, a2hpbGxpZ2FuQG1hbGFnaGFuLm9yZy5ueg==; Alan Sher, YXNoZXJAbmlhaWQubmloLmdvdg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.