 Maoyu Wang
Maoyu Wang Zhensheng Zhang1†
Zhensheng Zhang1† Yasheng Zhu
Yasheng Zhu Chuanliang Xu
Chuanliang Xu
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Immunol. , 11 July 2023
Sec. Cancer Immunity and Immunotherapy
Volume 14 - 2023 | https://doi.org/10.3389/fimmu.2023.1226057
This article is part of the Research Topic Community Series in Post-Translational Modifications of Proteins in Cancer Immunity and Immunotherapy, volume II View all 12 articles
With the rapidly increasing incidence of bladder cancer in China and worldwide, great efforts have been made to understand the detailed mechanism of bladder cancer tumorigenesis. Recently, the introduction of immune checkpoint inhibitor-based immunotherapy has changed the treatment strategy for bladder cancer, especially for advanced bladder cancer, and has improved the survival of patients. The ubiquitin–proteasome system, which affects many biological processes, plays an important role in bladder cancer. Several E3 ubiquitin ligases and deubiquitinases target immune checkpoints, either directly or indirectly. In this review, we summarize the recent progress in E3 ubiquitin ligases and deubiquitinases in bladder cancer tumorigenesis and further highlight the implications for bladder cancer immunotherapies.
Bladder cancer (BCa) is one of the most common types of cancer, with 550,000 new cases and 200,000 deaths annually (1). While the 5-year survival rate of all bladder cancer patients is 77.1%, the rate drops dramatically to 36.3% for regional disease and 4.6% for metastatic disease (2). Therefore, adjunctive therapy is needed to improve the prognosis of invasive and metastatic diseases. Cisplatin and gemcitabine combination chemotherapy has been applied for advanced bladder cancer (3); however, no major improvements in survival rate have been achieved until recently. The 5-year survival rate for patients with metastasis is 15% (3).
Immunotherapy, especially immune checkpoint inhibitors, is widely used for the treatment of different cancers (4, 5). BCa has been reported to be relatively sensitive to immunotherapy (6, 7). In May 2016, atezolizumab was the first PD-L1 inhibitor approved by the Food and Drug Administration (FDA) for bladder cancer (8). Since then, another four immune checkpoint inhibitors targeting PD-1 or PD-L1 for locally advanced and metastatic bladder cancer, including Nivolumab, Pembrolizumab, Avelumab, and Durvalumab have been approved by FDA for bladder cancer (8–10). However, owing to a lack of response, only a small group of patients with BCa can benefit from these agents (11). Taking PD-L1 as example, many studies have verified that PD-L1 expression is correlated with anti-PD-1/PD-L1 treatment, where high PD-L1 expression is equal to a good response to anti-PD-1/PD-L1 treatment (12). Thus, exploring the mechanism and identifying other reagents that can improve the efficacy of immune checkpoint blockade (ICB) is urgently needed (13). A series of mechanisms of PD-L1 regulation by post-translational modifications have been revealed in different cancers among recent research, including bladder cancer (14–16).
Ubiquitination and deubiquitinating modifications are highly conserved posttranslational modifications (PTMs) in mammals that play important roles in many biological processes and diseases, including cancers. The ubiquitin-activating enzyme E1, ubiquitin-conjugating enzyme E2, and ubiquitin ligase E3 contribute to the step-by-step process of ubiquitination. Ubiquitination involves the transfer of the C-terminal glycine of ubiquitin to the -NH2 group of the substrate lysine residue. Monoubiquitination, multiubiquitination, and polyubiquitination, which lead to proteolysis and signal transduction, are the three main types of ubiquitination (17). On the other hand, deubiquitinases (DUBs) can reverse ubiquitination by removing ubiquitin chains, thereby preserving the expression of the substrate protein while preventing ubiquitination. Most elements of biological activity depend on the interplay between ubiquitination and deubiquitination (13).
Numerous studies have demonstrated that the ubiquitin proteasome system (UPS) is related to the occurrence and progression of bladder cancer and that E3 ubiquitin ligases may be promising therapeutic targets (18–21). Meanwhile, the interaction between ubiquitination modification and immune-related molecules is emerging as a crucial regulatory mechanism and has recently draws great research interest (16, 22–25).
In this review, we summarize recent findings on protein ubiquitination and deubiquitinating enzymes in bladder cancer tumorigenesis and progression, as well as recent advances in the regulation of cancer immunotherapy effects.
Over 600 types of E3 ubiquitin ligases involved in the degradation of proteins have been discovered in humans (26). E3 ligases are classified into three subtypes: the interesting new gene (RING)-type, the homologous to E6AP carboxyl terminus (HECT)-type, and the RING-between-RING (RBR)-type (27). RING E3 ligases contain multiple subtypes, including monomers (c-CBL,E4B), homodimers (cIAP, CHIP), heterodimers (MDM2-MDMX), cullin-RING ligases (CRLs), and other RING E3s (28). CRLs are comprised of multiple subunits, which consist of four components: a cullin (CUL1,2,3,4A,4B,5,7,9), an adaptor protein, a substrate-recognizing receptor, and one RING protein (29–32). Moreover, SCF is the largest complex, consisting of SKP1, Cullin1, RBX1, and F-box proteins (29, 33). HECT structures are divided into three subfamilies: NEDD4 subfamily, HERC subfamily, and other HECT E3 ligases (34). RBRs are grouped into the Ariadne family and other RBRs (35). In particular, E3 ubiquitin ligases determine substrate specificity in the ubiquitination process.
In addition to maintaining the balance of intracellular proteins, E3 ligases are involved in multiple non-degradable functions including intracellular transport, autophagy, DNA damage repair, and metabolism (36). Thus, E3 ubiquitin ligases are critical for cellular processes. Therefore, their dysregulation may have a potential effect on the pathogenesis of cancer. Disorders of E3 ligases result in aberrant activation or inactivation of signaling pathways and the accumulation of misfolded or dysfunctional proteins (37), which promotes the occurrence and progression of cancer.
Numerous E3 ligases have been reported to be involved in bladder cancer tumorigenesis. They are involved in the regulation of key molecules including PD-L1, PTEN, and p53 (Table 1). In this section, we provide a detailed description of each E3 ligase in bladder cancer.
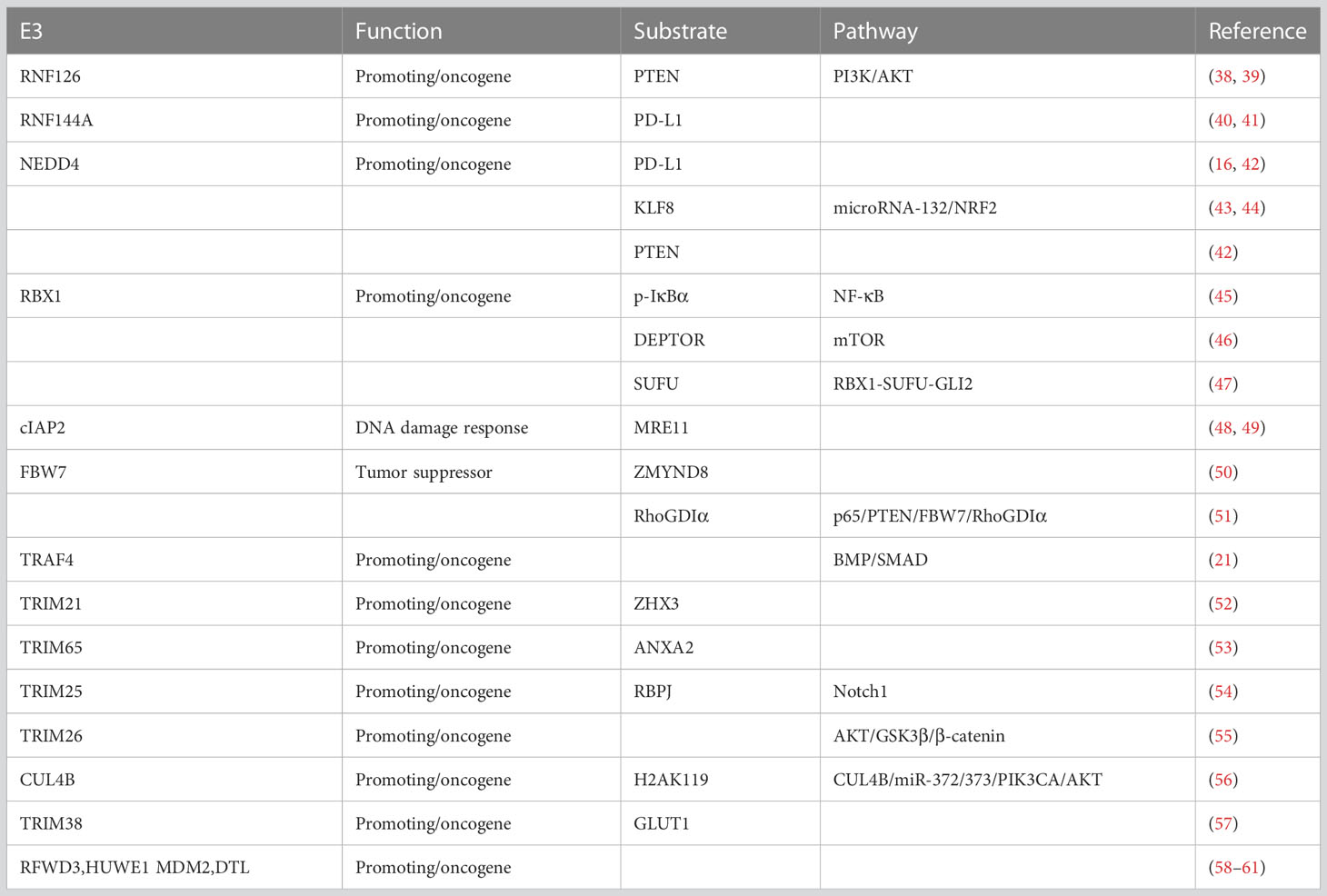
Table 1 E3 ligases in bladder cancer tumorigenesis.
RNF126 is a RING domain E3 ligase. A group of RNF126 substrates has been identified, including frataxin (62–64), epidermal growth factor receptor (64), pyruvate dehydrogenase kinases (65) and insulin-like growth factor II receptor (66). RNF126 is highly expressed in various cancers and strongly associated with tumorigenesis, including bladder cancer (38, 67–69). In BCa, RNF126 directly binds to PTEN via its C-terminal containing the RING domain and promotes the polyubiquitination and degradation of PTEN through the proteasome pathway (38). In vivo and in vitro studies have demonstrated that PTEN acts as an anti-oncogene, and PTEN silencing is closely related to the poor prognosis of patients with BCa (70). RNF126 silencing stabilizes PTEN, which antagonizes PI3K/AKT signaling pathway (38, 39), and promotes cell proliferation and metastasis when activated.
Moreover, previous studies revealed that RNF126 promotes the repair of DNA double-strand breaks via NHEJ and HR through different mechanisms (71, 72). The Ku70-Ku80 heterodimer recognizes DNA double-strand breaks (DSBs) and recruits proteins responsible for DNA repair by non-homologous end joining (NHEJ). While prolonged retention of Ku70/80 at DSBs prevents the completion of DNA repair, RNF126 ubiquitylates Ku80 at DSBs and promotes Ku70/80 dissociation from DSBs. In contrast, RNF126 can ubiquitinate and quench RNF168 function in the DNA damage response (71). Cisplatin has been widely used as first-line treatment for patients with advanced BCa (73). Furthermore, cisplatin induces cell apoptosis by accumulating DNA double-strand breaks. RNF126 depletion markedly increases the effect of cisplatin in inducing apoptosis in BCa cells (38). It has also been reported that RNF126 can directly bind and regulate PTEN stability through polyubiquitination, making RNF126 an attractive target for augmenting cisplatin-based chemotherapy and regulating bladder cancer tumorigenesis.
RNF144A belongs to the RBR E3 ubiquitin ligase family. Epigenetic depletion of RNF144A has been detected in numerous human cancers, including glioblastoma (74), breast cancer (75), and bladder cancer (40), indicating that RNF144A may act as a tumor suppressor. Previous studies have found that RNF144A is upregulated by various DNA-damaging agents (76) and further promotes cancer cell apoptosis of cancer cells by ubiquitinating and degrading DNA-PKcs and BMI1 (74, 77).
In a recent study, the basal-squamous subtype of bladder cancer has been found to express relatively low levels of RNF144A and high levels of immune checkpoint protein programmed cell death ligand-1(PD-L1) (41). The carboxyl-terminal region (aa 250–292) of RNF144A is responsible for its interaction with PD-L1, and RNF144A mainly targets glycosylated PD-L1 for degradation (40), further indicating a complex mechanism between protein ubiquitination and glycosylation.
NEDD4 is a HECT family E3 ubiquitin ligase (78). Mounting evidence has demonstrated that NEDD4 participates in the tumorigenesis of human cancers, such as cervical cancer (79), hepatocellular carcinoma (80), and breast cancer (81). NEDD4 is highly expressed in bladder cancer and promotes tumor cell migration and invasion (42, 43). KLF8 acts as a transcription factor in the Sp/KLF family and stimulates and promotes migration of bladder cancer cells. Moreover, miR-132 is downregulated by KLF8, which is overexpressed in bladder cancer. NEDD4 is conformed to interact with KLF8 (44). In bladder cancer, NEDD4 depletion significantly downregulated endogenous KLF8 ubiquitination, which affected the K63-linked polyubiquitination of KLF8, while K48-linked polyubiquitination remained unchanged. NEDD4 intensifies the stability and transcriptional activity of KLF8 through ubiquitination and affects the miR-132/NRF2 axis, thereby promoting tumor progression (44).
The ubiquitin ligase activity of NEDD4 can be promoted by FGFR1 and EGFR activation via tyrosine phosphorylation of NEDD4 (82). Previous studies have demonstrated that there is relatively decreased expression of PD-L1 in bladder cancer with FGFR3 mutations or high expression (41, 83, 84). Jing et al. (16)have indicated that the activation of FGFR3 promoted NEDD4 binding and phosphorylation and it had been reported that NEDD4 can be phosphorylated to greatly improve its ubiquitination capacity. NEDD4 depletion using CRISPR/Cas9-sgRNA remarkably upregulated PD-L1 expression in bladder cancer cells. NEDD4 targets and catalyzes the K48-linked polyubiquitination of PD-L1. These results reveal that NEDD4 is a critical regulator of PD-L1 expression in bladder cancer upon FGFR3 activation. This study provides powerful evidence for the combination of anti-PD-1 antibody therapy and erdafitinib, a tyrosine kinase inhibitor of FGFR1–4 (16).
As mentioned earlier, PTEN acts as an oncogene in bladder cancer. NEDD4 regulates PTEN levels in several types of human cancers (85). In bladder cancer, PTEN levels were increased by NEDD4 silencing (42). NEDD4 downregulation inhibits cell proliferation and apoptosis. However, the precise mechanism by which NEDD4 regulates PTEN expression has not been fully elucidated.
The cullin/RING ubiquitin ligase (CRL)family is the largest UPS E3 family (86). RBX1 forms the catalytic core of CRL complexes with different Cullin subunits (87). RBX1 is widely reported to be associated with poor clinical prognosis and is highly expressed in many cancers, including bladder cancer. In particular, RBX1 expression is significantly higher in muscle-invasive BCa and positively correlated with epithelial–mesenchymal transition (EMT) via inhibition of mTOR kinase activity by accumulation of the cullin-RING ligase (CRL) substrate mTOR-inhibitory protein DEPTOR (46).
Moreover, RBX1 has been confirmed to be positively correlated with activation of the NF-κB signaling pathway and nuclear p65 expression (45). p65 plays a key role in the canonical NF-κB pathway and is inactive in the cytoplasm upon binding to IκBα. Upon receiving the relevant signals, IκBα is phosphorylated, which is then ubiquitinated and degraded. Finally, p65 enters the nucleus and activates gene transcription (88). Therefore, IκBα-p65 is a key regulatory factor in the NF-κB signaling pathway. Activation of the NF-κB signaling pathway promotes tumor progression (89). By enhancing p-IκBα ubiquitination and degradation, RBX1 activates NF-κB signaling, which promotes p65 nuclear translocation and causes the transcription of several metastasis-related target genes including matrix metalloproteinase 9 (MMP9), vascular cell adhesion molecule 1 (VCAM1), and urokinase-type plasminogen activator receptor (uPAR) (45). Recently, Wang et al. demonstrated that RBX1 can activate the hedgehog pathway through the ubiquitinate suppressor of fused homolog (SUFU) for degradation, and dysregulation of the RBX1–SUFU–GLI2 axis play a pivotal role in bladder cancer progression (47).
IAP family members have been indicated to act as a key role in the regulation of NF-κB signaling and participate in intrinsic and extrinsic cell death pathways (90). cIAP2 is a RING-type E3 ligase in the IAP family and has been demonstrated to play a pivotal role in DNA repair (91, 92). Although the expression of cIAP1 examined by immunohistochemical testing is highly correlated to bladder cancer TNM stage, tumor grade, disease recurrence, and tumor-related death (93) and cIAP2 precise function and substrate specificity is unclear, previous studies have a common sense that there is redundancy between cIAP1 and cIAP2 in the regulation of cell death (94, 95). Recently, cIAP2 was reported to be involved in regulating radiosensitization in bladder cancer (48).
Histone deacetylase (HDAC) inhibitors exhibit low toxicity in normal cells, and panobinostat, an HDAC inhibitor, is a promising radiosensitizer (96). Panobinostat downregulates MRE11 (49), which is a key player in DNA repair, leading to a decreased ability to repair DNA, thereby enhancing radio sensitization. In T24 cells, transfecting cIAP2 into cells in increasing quantities, a growing decrease in MRE11 levels was observed. cIAP2 downregulates MRE11 via proteasomal pathways and increases the ubiquitination of MRE11. Furthermore, T24 cells became more radiosensitive after panobinostat treatment when cIAP2 was silenced.
F-box and WD repeat domain-containing 7(FBW7) is a member of the RING E3 ligase family, which is a subunit of the SKP1, cullin1, and F-box protein ubiquitin ligase complex (29). Low expression and mutation of FBW7 has been frequently detected in various human tumors such as breast cancer (97), colon cancer (98), and gastric cancer (99). Therefore, FBW7 is generally considered a tumor suppressor. According to the analysis of public datasets TCGA-BLCA and GSE13507, it has been verified that the mRNA expression levels of FBW7 are significantly downregulated in bladder tumors compared with normal samples (50). Kaplan–Meier analysis suggested that patients with BCa with high FBW7 expression levels exhibited longer survival times. Collectively, these results indicate that FBW7 may serve as a tumor suppressor in bladder cancer. ZMYND8 was acted as a common oncogene in numerous tumors, including bladder cancer (50). Bioinformatics predictive analysis from the UbiBrowser platform (http://ubibrowser.ncpsb.org/) and ubiquitination assays demonstrated that in T24 cells, ZMYND8 was a substrate target of FBW7. FBW7 is a tumor suppressor that is and downregulated in BCa. Low expression of FBW7 can increase the protein levels of ZMYND8 and promote BCa progression (50). This result was further confirmed in clinical samples.
Moreover, FBW7 was verified to be an NF-κBp65 downstream effector. Through promoting RHO guanosine diphosphate dissociation inhibitor (RhoGDIα) protein degradation, FBW7 significantly inhibited BCa migration (51). Mechanistically, p65 inhibited PTEN mRNA transcription, whereas PTEN accelerated FBW7 protein degradation. This revealed the function of the p65/PTEN/FBW7/RhoGDIα axis in mediating bladder cancer migration and expands the theoretical support for the regulation of the NF-κBp65 and PTEN pathways in BCa treatment.
MDM2 is reported to mainly target p53 protein in various types of cancer, including bladder cancer (100). The SNP309 polymorphisms of MDM2 is associated with an improved survival rate of bladder cancer (101). MDM2 is upregulated by the OCT3/4/TET1/NRF2 axis, which contributes to increased immune escape in bladder cancer (102). Amounts of inhibitors, such as MDM2 exerted an influence on immunity in the tumor microenvironment, such as APG-115 and AMG-232. APG-115 can enhance the efficacy of PD-L1 blockade (103) and AMG-232 (104) can increase the ability to kill T cells. Furthermore, gene amplification of MDM2 can act as a predictive marker for PD-L1 targeted therapy response (105).
Several other E3 ubiquitin ligases are also involved in bladder tumorigenesis. RFWD3 is highly expressed in bladder cancer tissue and correlates with a higher N stage and poorer prognosis (58). A bladder cancer genome-wide CRISPR/Cas9 KO screen showed that HUWE1 was correlated with cisplatin sensitivity in bladder cancer; however, the underlying mechanism has not been elucidated (59). MDM2 binds to PPARγ to ubiquitinate and downregulate its PPARγ expression (60). Denticleless E3 ubiquitin protein ligase homolog (DTL) is overexpressed in BCa, and increased DTL expression correlates with malignant biological behavior and promotes BCa progression through the AKT/mTOR pathway (61). A pan-cancer study also showed that DLT could be a potential immunotherapy biomarker (106).
TRAF4 can bind to and target another E3 ligase, SMURF1, for proteasomal degradation (21). As SMURF1 is a negative regulator of the BMP/SMAD signaling pathway, TRAF4 can promote BMP/SMAD signaling and inhibit bladder cancer progression (21). TRIM21 acts as a ubiquitin E3 ligase to degrade ZHX3, which is involved in bladder cancer progression and metastasis (52). The expression level of TRIM65 is frequently upregulated and ANXA2 is ubiquitinated and degraded by TRIM65. Bladder cancer patients with low ANXA2 expression and high TRIM65 expression showed the poorest outcome (53). RITA1 recruits TRIM25 to ubiquitinate RBPJ to accelerate its degradation via the proteasome, which leads to transcriptional inhibition of Notch1 downstream targets (54). TRIM26 plays an oncogenic role in bladder cancer by regulating cell proliferation, migration, and invasion via the AKT/GSK3β/β-catenin pathway (55). CUL4B is a scaffold protein in the CUL4B–RING ubiquitin ligase (CRL4B) complexes. CUL4B levels are overexpressed and positively associated with the malignancy of BCa, and CUL4B epigenetically represses the transcription of miR-372/373 by catalyzing the monoubiquitination of H2AK119 in the gene cluster encoding miR-372/373, which further leads to the upregulation of PIK3CA and activation of AKT (56).
Reprogramming cell metabolism is a hallmark of cancer (107, 108). Aerobic glycolysis has been extensively studied in several cancers, including bladder cancer (107). It is characterized by increased glucose uptake and lactate production under normal oxygen conditions. Elevated glycolytic flux in cancer cells is mediated by glycolysis-associated signature genes, including GLUT1 (109). GLUT1 driven glycolytic reprogramming is considered necessary for tumor cell growth (110).Wang et al. identified GLUT1 as the downstream substrate of TRIM38 and TRIM38 can constrain bladder tumor progression through ubiquitination and degradation of GLUT1 (57). TRIM38 has been verified to be a predictive biomarker related to prognosis, with low expression in BCa (57).
Deubiquitinases (DUBs) are proteases that remove ubiquitin from substrates or cleave ubiquitin chains to regulate ubiquitination (111). It is important to regulate the processes of deubiquitination and ubiquitination (112). DUBs consist of cysteine proteases and metalloproteinases that specifically cleave ubiquitin molecules on protein substrates (113). Approximately 100 different DUBs can be broadly classified into seven distinct superfamilies (114). Six of these families are cysyrine-based DUBs, including Ub C-terminal hydrolases (UCHs), Ub-specific proteases (USPs), Machado-Josephin domain proteases (MJDs), ovarian tumor proteases (OTUs), motifs interacting with the Ub-containing novel DUB family (MINDY), zinc-finger-containing Ub peptidase (ZUP1), and Jab1/Mov34/MPN+ protease (JAMM) family members, which are zinc-binding metalloproteases (115).
Numerous studies have demonstrated that the effect of protein deubiquitination is associated with the occurrence and development of cancers, such as prostate cancer, lung cancer, stomach cancer, and bladder cancer (116–120). A summary of the deubiquitinases involved in BCa is presented in Table 2.
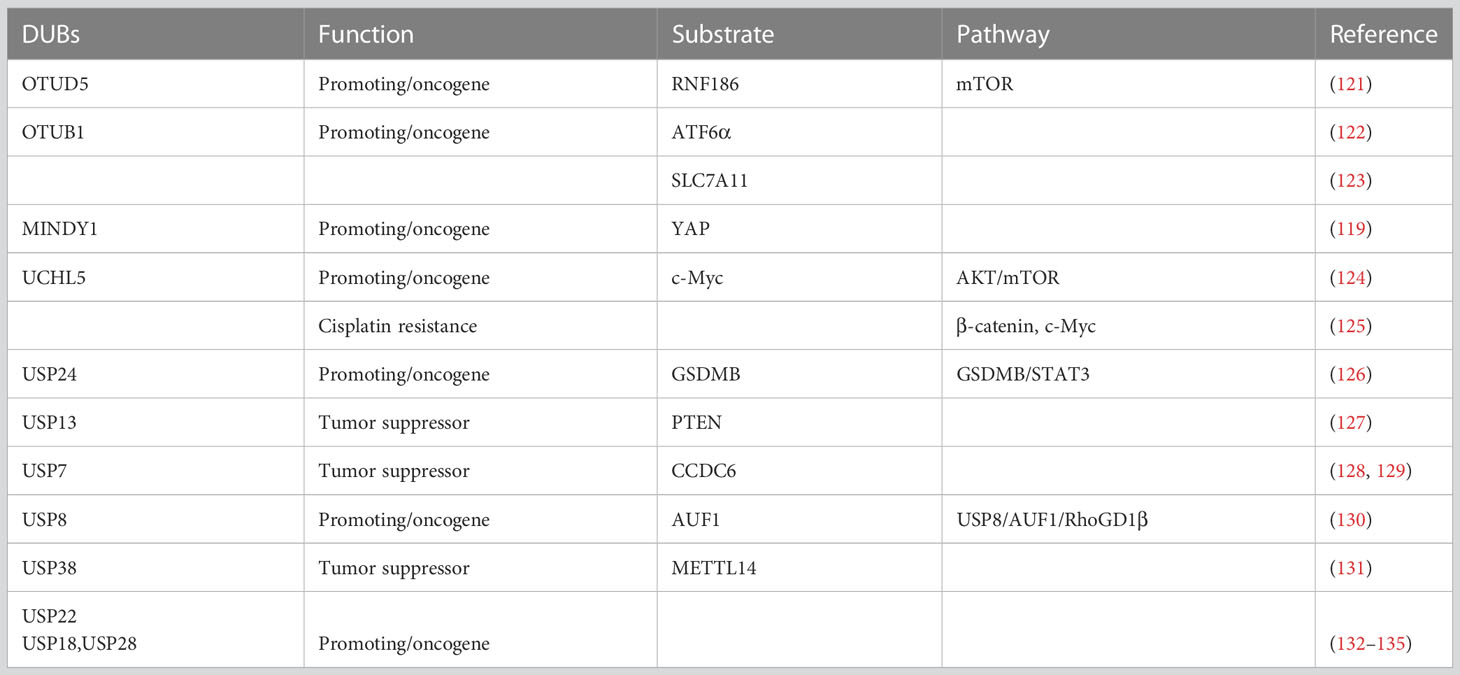
Table 2 Deubiquitinases in bladder cancer tumorigenesis.
There are 16 types of cysteine protease OTU family members, including OTUB, OTUD, A20-like, and OTULIN subfamily (113). The OTUD family is one of the subfamilies including OTUD1, OTUD2/YOD1, OTUD3, OTUD4, OTUD5/DUBA, OTUD6A, OTUD6B, and ALG13 (113, 136). OTUD5 has been the focus of numerous studies and plays pivotal roles in various cellular processes. The first report of function of OTUD5 is to negatively regulate IFN-1 expression by cleaving the polyubiquitin chains on TRAF3 (137). Furthermore, OTUD5 regulates DNA damage repair, transcription, and innate immunity (138, 139).
In bladder cancer, OTUD5 has been shown that is highly expressed in tumor tissues compared with normal urothelial cells (121). OTUD5 knockdown inhibited the cell proliferation, and OTUD5 positively regulated the mTOR signaling pathway to promote cell proliferation. Specifically, OTUD5 stabilizes RNF186 by deubiquitination, leading to sestrin2 degradation, which acts as a feedback inhibitor of the mTOR signaling pathway (140, 141). Everolimus treatment, an mTOR inhibitor, with simultaneous OTUD5 knockdown seems to be an ideal strategy for bladder cancer treatment (121).
The deubiquitinase OTUB1 is significantly more highly expressed in bladder cancer tumor tissues than in normal tissues (122). Kaplan–Meier survival analysis confirmed that bladder cancer patients with low OTUB1 expression had significantly superior overall survival compared to those with high OTUB1 expression. It has been found that OTUB1 can stabilize activating transcription factor 6α (ATF6α) in response to endoplasmic reticulum stress and promote bladder cancer progression (122). Numerous studies have indicated that ferroptosis is an important and independent mechanism of tumor suppression (142). Solute carrier family 7, membrane 11 (SLC7A11), a 12-pass transmembrane protein, acts as a potential biomarker for protecting cancer cells from oxidative stress and ferroptosis (143). Liu et al. discovered a distinct mechanism by which OTUB1 mediates ferroptosis in bladder cancer via the stabilization of SLC7A11 (123).
MINDY1 (also known as FAM63A) has been reported that contains MIU motifs with high selectivity for binding and cleaving K48-linked polyUb (144). The Hippo signaling pathway has emerged as a critical pathway in the regulation of bladder cancer tumorigenesis, and TAZ and YAP are important effectors of this pathway (145–147). MINDY1 removes the K48-linked ubiquitin chain from YAP, thus inhibiting proteasome-mediated YAP degradation, which will in turn promote the expression of YAP downstream genes, CTGF, ANKRD1, and CYR61 (119).
UCHL5 is abnormally upregulated in human cancer tissues and cell lines, such as pancreatic adenocarcinoma, gastric cancer, endometrial cancer, and bladder cancer (124, 148–150). Upregulation of the TGF signaling pathway is the main mechanism by which UCHL5 modulates malignant tumor progression (151–153). UCHL5 is overexpressed in patients with bladder cancer patients, and high expression is associated with poor prognosis and tumor progression. Mechanistically, UCHL5 activates the AKT/mTOR signaling pathway and increases c-Myc expression, which promotes tumor occurrence and progression (124). Meanwhile, it has been reported that the UCHL5 inhibitor b-AP15 suppresses bladder cancer stemness by inhibiting the β-catenin and c-Myc signaling pathways and overcomes cisplatin resistance (125). b-AP15 has been demonstrated to have synergistic effects in combination with cisplatin, gefitinib, gemcitabine, and vinorelbine in lung cancer cells (154). In bladder cancer cell lines and mouse xenograft models, b-AP15 combined with cisplatin showed superior therapeutic effects compared to cisplatin monotherapy (125). These studies indicate that UCHL5 may act as a potential therapeutic target, and that b-AP15 may be a new choice for patients with cisplatin resistance.
Ubiquitin-specific peptidase 24 (USP24), consisting of 2,620 amino acids, serves as a deubiquitinase (155). However, the biological function of USP24 in cancer is poorly understood. It has been reported that USP24 binds to GSDMB to deubiquitinate and stabilize GSDMB. GSDMB promotes cancer cell growth by activating STAT3, which increases the expression of HK2, LDNA, ENO2, and IGFBP3 to enhance glycolysis in bladder cancer cells (126). EOAI3402143, a USP24 inhibitor, can block this process, which provides a therapeutic strategy for inhibiting the GSDMB/STAT3 axis (126).
USP13 belongs to the Ub-specific protease subfamily of deubiquitinase family. USP13 has been indicated in suppressing tumor occurrence by deubiquitinating anti-oncogenes, including p53 (156), PTEN (157), and MITF (158), and subsequently stabilizing these proteins. As mentioned above, PTEN acts as a key tumor suppressor in bladder cancer via inhibition of the PI3K/AKT/mTOR signaling pathway. Otherwise, NF-κB activation has been reported to be essential for inhibition of PTEN expression (159, 160). PTEN is deubiquitinated by USP13 in bladder cancer, and its stabilized expression suppresses tumor progression (127). There is also a potential regulatory loop in which NF-κB induces miR-130b/301b overexpression, decreasing USP13 expression and subsequently leading to the downregulation of PTEN overexpression (127).
Several studies have demonstrated that other USPs serve as oncogenes in BCa tumorigenesis (128, 130, 132–134, 161). Jeong et al. detect the mRNA expression of USP2a in bladder cancer tissues and normal tissues. The results indicate that the expression of USP2a in bladder cancer is downregulated and that high stage muscle invasive bladder cancer (MIBC) has lower USP2a expression. USP2a can be specifically used as a potential marker to stratify the more invasive phenotype of MIBC (132).
USP7 has been reported to modulate CCDC6 levels in bladder cancer and lung neuroendocrine cancers (129). CCDC6 acts as a tumor suppressor, its deficiency determines the sensitivity of PARP-inhibitors (162, 163). In a recent study, P5091, an inhibitor of USP7, promoted CCDC6 degradation and sensitized bladder cancer cells to the cytotoxic effect of the PARP-inhibitor olaparib (128).
The non-canonical NF-κB subunit p52 upregulates USP8 expression at the transcriptional level, and USP8 modulates AUF1 protein degradation. USP8 plays a significant role in the p52/miR-145/Sp1/USP8/AUF1/RhoGD1β axis, which can act as a positive regulator of bladder cancer invasion (130).
USP22 is a positive regulator of tumor growth. Silencing USP22 by interfering with RNA inhibits proliferation and induces cell cycle arrest in BCa cells (133). USP18 and USP28 have been reported to serve as prognostic markers for bladder cancer (134, 135). A study also revealed a feedback loop of USP38 and METTL14 in bladder cancer to suppress BCa progression. METTL14 stabilizes USP38 mRNA expression through YTHDF2-dependant m6A modification and USP38 enhances the stability of METTL14 by deubiquitination of METTL14 (131).
The concept of immunotherapies for bladder cancer can be divided into cytokine-based treatment, genetically engineered immune cells (adoptive cell therapy), oncolytic viruses, bispecific antibodies, intravesical therapy with Bacillus Calmette–Guerin (BCG) vaccine, immune checkpoint inhibitors (ICIs), and antibody–drug conjugates (ADCs) (10, 164, 165).
BCG immunotherapy remains the gold standard treatment for patients with non-muscle-invasive bladder cancer (NMIBC) at a high risk of progression or recurrence (166). Although it has been used in clinical practice since 1976, the mechanism of the BCG vaccine in BCa is not completely understood. Upon attachment to the urothelium and internalization, it is thought to induce innate and adaptive immune responses. However, whether a combination of reagents targeting E3 ligases or DUBs can augment the response to BCG or conquer certain patients’ unresponsiveness to BCG warrants further exploration (167).
The adoption of ICIs in bladder cancer has dramatically changed its treatment landscape (168). ICIs are now approved for the treatment of BCa at all stages, depending on the specific tumor characteristics (10). Immune checkpoint inhibitors can enhance T-cell responses and provide promising clinical outcomes in bladder cancer. However, this treatment strategy has only a 13%–24% response rate among patients with bladder cancer. A deeper exploration of the mechanisms that regulate PD-1/PD-L1 expression and stability may help increase clinical effectiveness. During the last decade, intensive evidence has demonstrated that PD-1/PD-L1 protein expression is regulated by the ubiquitin-mediated proteasome degradation pathway (169–172).
RNF144A and NEDD4 have been reported to participate in the regulation of PD-L1 expression (Figure 1). The basal-squamous subtype of bladder cancer expresses relatively low levels of RNF144A and high levels of immune checkpoint protein programmed cell death ligand-1 (PD-L1) (41). The carboxyl-terminal region (aa 250–292) of RNF144A is responsible for its interaction with PD-L1 and RNF114A mainly targets glycosylated PD-L1 for degradation (40). PD-L1, primarily in the insoluble fraction, interacts with RNF144A, which contains the plasma membrane and intracellular vesicles (40). RNF114A knockout stabilizes PD-L1 and leads to a reduction in tumor-infiltrating CD8+ T-cell populations in BBN-induced bladder tumors (40). Thus, RNF144A E3 ligase may be a promising therapeutic target for immunotherapy or combined therapy.
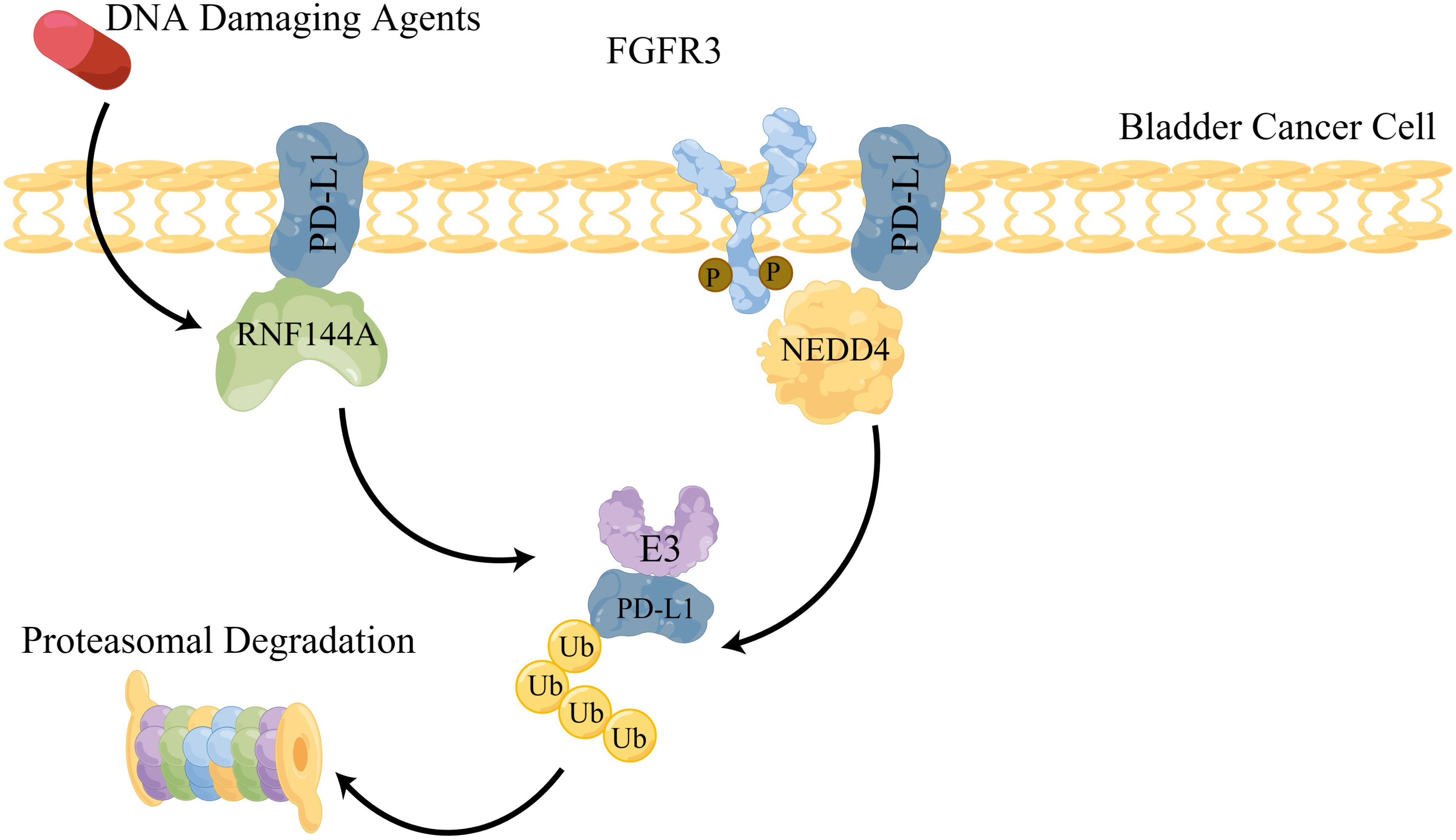
Figure 1 Graphic model of interaction between PD-L1 and E3 ligases in bladder cancer.
FGFR3 is an eligible target for the treatment of bladder cancer. p-FGFR3 and NEDD4 co-localized at the cell surface of bladder cancer cells. It has been demonstrated that NEDD4 can be phosphorylated to greatly improve its ubiquitination capacity by FGFR3 (16). NEDD4 depletion using CRISPR/Cas9-sgRNA remarkably upregulated PD-L1 expression in bladder cancer cells. NEDD4 targets and catalyzes the K48-linked polyubiquitination of PD-L1. These results revealed that NEDD4 is a critical regulator of PD-L1 expression in bladder cancer with FGFR3 activation (16). Thus, NEDD4 E3 ligase may be a promising therapeutic target in the bladder with immunotherapy or combined therapy.
USP7 has been shown to regulate anti-tumor immune responses. The activity of Treg cells is impeded by its inhibitor and the polarization of tumor-related macrophages is enhanced (173). One study reported that USP7 expression is positively related to PD-L1 expression and USP7 directly binds to PD-L1 which stabilized it in gastric cancer (117).However, the function of USP7 inhibitors in enhancing the immune response in bladder cancer remains unclear. Therefore, it is essential to investigate the role of USP7 in bladder cancer.
Although some other DUBs, including USP22 (174) and USP9X (175), have been shown to regulate PD-1/PD-L1 expression, no research has been conducted on bladder cancer. Because ubiquitination or deubiquitination of certain molecules can be cellular context-dependent, E3 ligases and DUBs targeting PD-1/PD-L1 in other tumors should be further verified in bladder cancer. Several E3 ligases and DUBs, especially DUBs, can be directly targeted by small molecular drugs; thus the combination of specific inhibitors and ICIs might be attractive and promising for enhancing ICI treatment effects (176). Notably, deubiquitinating enzymes are potential biomarkers for treatment selection and prognosis prediction (177).
In addition to PD-1 or PD-L1 based immunotherapy, antibody–drug conjugates (ADCs) have recently shown great progress. An ADC targeting nectin-4 (Enfortumab Vedotin) has shown significantly prolonged survival in patients with locally advanced or metastatic urothelial carcinoma who previously received platinum-containing chemotherapy and progressed after treatment with a PD-1 or PD-L1 inhibitor (178). For patients who are not eligible for cisplatin-containing chemotherapy, Enfortumab Vedotin Plus Pembrolizumab may be a safe and effective surrogate for previously untreated advanced bladder cancer patients (179, 180). Nectin-4 is a transmembrane protein overexpressed in bladder cancer and several other malignancies, making it an appropriate target antigen for ADCs. However, little is known about its role in tumor development, progression, and immunomodulatory functions. It might also be interesting to investigate the regulation of stabilization and degradation (180).
Casitas B lymphoma-b (Cbl-b) is an E3 ligase that can modulate PD-L1 ubiquitination and degradation after inhibition of PI3K/Akt, Jak/Stat, and MAPK-Erk signaling (181). Cbl-b can also target the ubiquitination of PI3K NEDD4, PLCγ, and the zeta-subunit of TCR. Interestingly, Cbl-b also serves as a downstream regulator of both CD28 and CTLA-4 signaling pathways. Thus, both innate and adaptive immune cells are regulated by E3 ubiquitin ligase, promoting an immunosuppressive tumor microenvironment. This implicated a complex regulatory loop between CTLA-4, E3 ligase Cbl-b, and PD-L1. Novel Cbl-b inhibitors offer antigen-specific immune stimulation and are promising therapeutic tools in the field of immune-oncology (182).
In summary, patients with advanced bladder cancer have poor survival rates, and immunotherapy may be a promising method for these patients. The use of single-agent immunotherapy or combined immunotherapy may be a further direction for treating advanced bladder cancer. A better understanding of bladder cancer progression and its regulation of immune-related molecules will help us to develop better therapeutic drugs and select appropriate patients. However, the overall efficacy is unsatisfactory, and a large number of patients cannot benefit from these agents due to a lack of response. PTMs have been indicted to play a significant role in the regulation of protein stabilization of the PD-1/PD-L1 axis. The ubiquitinase–protease system plays a pivotal role in bladder cancer, including in tumor progression, cisplatin resistance, tumor suppression, and predictive biomarkers. Notably, numerous E3 ligases and DUBs act as oncogenes, including RBX1, cIAP2, CUL4B, OTUD5, MINDY1, and USP24. FBW7, USP13, USP2a, USP8, and USP7 serve as tumor suppressors. Furthermore, emerging evidence has demonstrated that RNF114A and NEDD4 can modulate PD-L1 ubiquitination, which in turn leads to the subsequent modulation of immunosuppression and anticancer effects.
This review highlights the significant role of the UPS in bladder cancer carcinogenesis and in the regulation of certain immune therapy-related molecules, including PD-1/PD-L1. These findings indicate that E3 ligases and DUBs may act as potential targets for bladder cancer therapy or a promising therapeutic approach to promote immunotherapy effectiveness by regulating ubiquitination and deubiquitination.
MW, ZZ, and ZL wrote the manuscript and YZ and CX edited it. All authors contributed to the article and approved the submitted version.
This work was supported by the Shanghai Sailing Program (20YF1448100), grants from the Science and Technology Commission of Shanghai Municipality (22Y11905200), the “Voyaging Talents” Fund of the Naval Medical University (2021008149), and National Natural Science Foundation of China (82203450, 81772720,81972391).
Figure 1 is drawn using Figdraw. Export ID is YTIYU24832 (https://www.figdraw.com).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Richters A, Aben KKH, Kiemeney L. The global burden of urinary bladder cancer: an update. World J Urol (2020) 38(8):1895–904. doi: 10.1007/s00345-019-02984-4
2. Saginala K, Barsouk A, Aluru JS, Rawla P, Padala SA, Barsouk A. Epidemiology of bladder cancer. Med Sci (Basel) (2020) 8(1):15. doi: 10.3390/medsci8010015
3. von der Maase H, Hansen SW, Roberts JT. Gemcitabine and cisplatin versus methotrexate, vinblastine, doxorubicin, and cisplatin in advanced or metastatic bladder cancer: results of a large, randomized, multinational, multicenter, phase III study. J Clin Oncol (2000) 18(17):3068–77. doi: 10.1200/JCO.2000.18.17.3068
4. Bai R, Lv Z, Xu D, Cui J. Predictive biomarkers for cancer immunotherapy with immune checkpoint inhibitors. biomark Res (2020) 8(2050-7771(2050-7771 (Print):34. doi: 10.1186/s40364-020-00209-0
5. Darvin P, Toor SM, Sasidharan Nair V, Elkord E. Immune checkpoint inhibitors: recent progress and potential biomarkers. Exp Mol Med (2018) 50(12):1–11. doi: 10.1038/s12276-018-0191-1
6. Feld E, Harton J, Meropol NJ. Effectiveness of first-line immune checkpoint blockade versus carboplatin-based chemotherapy for metastatic urothelial cancer. Eur Urol (2019) 76(4):524–32. doi: 10.1016/j.eururo.2019.07.032
7. Lawrence MS, Stojanov P, Polak P. Mutational heterogeneity in cancer and the search for new cancer-associated genes. Nature (2013) 499(7457):214–8. doi: 10.1038/nature12213
8. Lopez-Beltran A, Cimadamore A, Blanca A. Immune checkpoint inhibitors for the treatment of bladder cancer. Cancers (Basel) (2021) 13(1):131. doi: 10.3390/cancers13010131
9. Twomey JD, Zhang B. Cancer immunotherapy update: FDA-approved checkpoint inhibitors and companion diagnostics. AAPS J (2021) 23(2):39. doi: 10.1208/s12248-021-00574-0
10. Abd El-Salam MA, Smith CEP, Pan CX. Insights on recent innovations in bladder cancer immunotherapy. Cancer Cytopathol (2022) 130(9):667–83. doi: 10.1002/cncy.22603
11. Zou W, Wolchok JD, Chen L. PD-L1 (B7-H1) and PD-1 pathway blockade for cancer therapy: mechanisms, response biomarkers, and combinations. Sci Transl Med (2016) 8(328):328rv4. doi: 10.1126/scitranslmed.aad7118
12. Patel VG, Oh WK, Galsky MD. Treatment of muscle-invasive and advanced bladder cancer in 2020. CA Cancer J Clin (2020) 70(5):404–23. doi: 10.3322/caac.21631
13. Liu J, Cheng Y, Zheng M. Targeting the ubiquitination/deubiquitination process to regulate immune checkpoint pathways. Signal Transduct Target Ther (2021) 6(1):28. doi: 10.1038/s41392-020-00418-x
14. Zhang J, Bu X, Wang H. Cyclin d-CDK4 kinase destabilizes PD-L1 via cullin 3-SPOP to control cancer immune surveillance. Nature (2018) 553(7686):91–5. doi: 10.1038/nature25015
15. Hsu JM, Li CW, Lai YJ, Hung MC. Posttranslational modifications of PD-L1 and their applications in cancer therapy. Cancer Res (2018) 78(22):6349–53. doi: 10.1158/0008-5472.Can-18-1892
16. Jing W, Wang G, Cui Z. FGFR3 destabilizes PD-L1 via NEDD4 to control T-cell-Mediated bladder cancer immune surveillance. Cancer Res (2022) 82(1):114–29. doi: 10.1158/0008-5472.CAN-21-2362
17. Zhao Y, Li J, Chen J, Ye M, Jin X. Functional roles of E3 ubiquitin ligases in prostate cancer. J Mol Med (Berl) (2022) 100(8):1125–44. doi: 10.1007/s00109-022-02229-9
18. Zhai F, Wang J, Yang W, Ye M, Jin X. The E3 ligases in cervical cancer and endometrial cancer. Cancers (Basel) (2022) 14(21):5354. doi: 10.3390/cancers14215354
19. Jayaprakash S, Hegde M, Bharathwaj Chetty B. Unraveling the potential role of NEDD4-like E3 ligases in cancer. Int J Mol Sci (2022) 23(20):12380. doi: 10.3390/ijms232012380
20. Liu TT, Yang H, Zhuo FF. Atypical E3 ligase ZFP91 promotes small-molecule-induced E2F2 transcription factor degradation for cancer therapy. Ebiomedicine (2022) 86(2352-3964(2352-3964 (Electronic):104353. doi: 10.1016/j.ebiom.2022.104353
21. Iyengar PV, Marvin DL, Lama D. TRAF4 inhibits bladder cancer progression by promoting BMP/SMAD signaling. Mol Cancer Res (2022) 20(10):1516–31. doi: 10.1158/1541-7786.MCR-20-1029
22. Zhu L, Kuang X, Zhang G. Albendazole induces immunotherapy response by facilitating ubiquitin-mediated PD-L1 degradation. J Immunother Cancer (2022) 10(5):e003819. doi: 10.1136/jitc-2021-003819
23. Xiong WJ, Gao XL, Zhang TT. USP8 inhibition reshapes an inflamed tumor microenvironment that potentiates the immunotherapy. Nat Commun (2022) 13(1):1700. doi: 10.1038/s41467-022-29401-6
24. Wang X, Tokheim C, Wang B. In vivo CRISPR screens identify E3 ligase Cop1 as a modulator of macrophage infiltration and cancer immunotherapy target. Cell (2021) 184(21):5357–74. doi: 10.1016/j.cell.2021.09.006
25. Han S, Wang R, Zhang Y. The role of ubiquitination and deubiquitination in tumor invasion and metastasis. Int J Biol Sci (2022) 18(6):2292–303. doi: 10.7150/ijbs.69411
26. Wang H, Lu Y, Wang M, Wu Y, Wang X, Li Y. Roles of E3 ubiquitin ligases in gastric cancer carcinogenesis and their effects on cisplatin resistance. J Mol Med (Berl) (2021) 99(2):193–212. doi: 10.1007/s00109-020-02015-5
27. Dale B, Cheng M, Park KS, Kaniskan HU, Xiong Y, Jin J. Advancing targeted protein degradation for cancer therapy. Nat Rev Cancer (2021) 21(10):638–54. doi: 10.1038/s41568-021-00365-x
28. Deshaies RJ, Joazeiro CA. RING domain E3 ubiquitin ligases. Annu Rev Biochem (2009) 78(1545-4509(1545-4509 (Electronic):399–434. doi: 10.1146/annurev.biochem.78.101807.093809
29. Cardozo T, Pagano M. The SCF ubiquitin ligase: insights into a molecular machine. Nat Rev Mol Cell Biol (2004) 5(9):739–51. doi: 10.1038/nrm1471
30. Genschik P, Sumara I, Lechner E. The emerging family of CULLIN3-RING ubiquitin ligases (CRL3s): cellular functions and disease implications. EMBO J (2013) 32(17):2307–20. doi: 10.1038/emboj.2013.173
31. Zhao YC, Sun Y. Cullin-RING ligases as attractive anti-cancer targets. Curr Pharm Design (2013) 19(18):3215–25. doi: 10.2174/13816128113199990300
32. Sun Y. Introduction. Adv Exp Med Biol (2020) 1217(0065-2598(0065-2598 (Print):1–8. doi: 10.1007/978-981-15-1025-0_1
33. Reed SI. Ratchets and clocks: the cell cycle, ubiquitylation and protein turnover. Nat Rev Mol Cell Biol (2003) 4(11):855–64. doi: 10.1038/nrm1246
34. Bernassola F, Chillemi G, Melino G. HECT-type E3 ubiquitin ligases in cancer. Trends Biochem Sci (2019) 44(12):1057–75. doi: 10.1016/j.tibs.2019.08.004
35. Wang XS, Cotton TR, Trevelyan SJ. The unifying catalytic mechanism of the RING-between-RING E3 ubiquitin ligase family. Nat Commun (2023) 14(1):168. doi: 10.1038/s41467-023-35871-z
36. Grabbe C, Husnjak K, Dikic I. The spatial and temporal organization of ubiquitin networks. Nat Rev Mol Cell Biol (2011) 12(5):295–307. doi: 10.1038/nrm3099
37. Hoeller D, Dikic I. Targeting the ubiquitin system in cancer therapy. Nature (2009) 458(7237):438–44. doi: 10.1038/nature07960
38. Xu HM, Ju LG, Xiong YY. E3 ubiquitin ligase RNF126 affects bladder cancer progression through regulation of PTEN stability. Cell Death Dis (2021) 12(3):239. doi: 10.1038/s41419-021-03521-1
39. Song MS, Salmena L, Pandolfi PP. The functions and regulation of the PTEN tumour suppressor. Nat Rev Mol Cell Biol (2012) 13(5):283–96. doi: 10.1038/nrm3330
40. Ho SR, Lee YC, Ittmann MM, Lin FT, Chan KS, Lin WC. RNF144A deficiency promotes PD-L1 protein stabilization and carcinogen-induced bladder tumorigenesis. Cancer Lett (2021) 520(1872-7980(1872-7980 (Electronic):344–60. doi: 10.1016/j.canlet.2021.08.008
41. Robertson AG, Kim J, Al-Ahmadie H. Comprehensive molecular characterization of muscle-invasive bladder cancer. Cell (2017) 171(3):540–56.e25. doi: 10.1016/j.cell.2017.09.007
42. Wen W, Li J, Wang L. Inhibition of NEDD4 inhibits cell growth and invasion and induces cell apoptosis in bladder cancer cells. Cell Cycle (2017) 16(16):1509–14. doi: 10.1080/15384101.2017.1338220
43. Mao M, Yang L, Hu J. Oncogenic E3 ubiquitin ligase NEDD4 binds to KLF8 and regulates the microRNA-132/NRF2 axis in bladder cancer. Exp Mol Med (2022) 54(1):47–60. doi: 10.1038/s12276-021-00663-2
44. Sun A, Hao J, Yu L. Regulation of Krüppel-like factor 8 by the NEDD4 E3 ubiquitin ligase. Am J Transl Res (2019) 11(3):1521–30.
45. Wu Q, Zhou X, Li P. ROC1 promotes the malignant progression of bladder cancer by regulating p-IkappaBalpha/NF-kappaB signaling. J Exp Clin Cancer Res (2021) 40(1):158. doi: 10.1186/s13046-021-01935-5
46. Wang W, Chen H, Liu Z. Regulator of cullins-1 expression knockdown suppresses the malignant progression of muscle-invasive transitional cell carcinoma by regulating mTOR/DEPTOR pathway. Br J Cancer (2016) 114(3):305–13. doi: 10.1038/bjc.2015.444
47. Wang W, Qiu J, Qu P. Regulator of cullins-1 (ROC1) negatively regulates the Gli2 regulator SUFU to activate the hedgehog pathway in bladder cancer. Cancer Cell Int (2021) 21(1):75. doi: 10.1186/s12935-021-01775-5
48. Nicholson J, Jevons SJ, Groselj B. E3 ligase cIAP2 mediates downregulation of MRE11 and radiosensitization in response to HDAC inhibition in bladder cancer. Cancer Res (2017) 77(11):3027–39. doi: 10.1158/0008-5472.CAN-16-3232
49. Groselj B, Kerr M, Kiltie AE. Radiosensitisation of bladder cancer cells by panobinostat is modulated by Ku80 expression. Radiotherapy Oncol (2013) 108(3):429–33. doi: 10.1016/j.radonc.2013.06.021
50. Qiu F, Jin Y, Pu J. Aberrant FBXW7-mediated ubiquitination and degradation of ZMYND8 enhances tumor progression and stemness in bladder cancer. Exp Cell Res (2021) 407(2):112807. doi: 10.1016/j.yexcr.2021.112807
51. Zhu J, Li Y, Chen C. NF-kappaB p65 overexpression promotes bladder cancer cell migration via FBW7-mediated degradation of RhoGDIalpha protein. Neoplasia (2017) 19(9):672–83. doi: 10.1016/j.neo.2017.06.002
52. Deng MH, Wei WS, Duan JL. ZHX3 promotes the progression of urothelial carcinoma of the bladder via repressing of RGS2 and is a novel substrate of TRIM21. Cancer Science (2021) 112(5):1758–71. doi: 10.1111/cas.14810
53. Wei WS, Chen X, Guo LY. TRIM65 supports bladder urothelial carcinoma cell aggressiveness by promoting ANXA2 ubiquitination and degradation. Cancer Lett (2018) 435(1872-7980(1872-7980 (Electronic):10–22. doi: 10.1016/j.canlet.2018.07.036
54. Tang H, Li X, Jiang L. RITA1 drives the growth of bladder cancer cells by recruiting TRIM25 to facilitate the proteasomal degradation of RBPJ. Cancer Sci (2022) 113(9):3071–84. doi: 10.1111/cas.15459
55. Xie X, Li H, Pan J, Han X. Knockdown of TRIM26 inhibits the proliferation, migration and invasion of bladder cancer cells through the Akt/GSK3beta/beta-catenin pathway. Chem Biol Interact (2021) 337(1872-7786(1872-7786 (Electronic):109366. doi: 10.1016/j.cbi.2021.109366
56. Liu X, Cui J, Gong L. The CUL4B-miR-372/373-PIK3CA-AKT axis regulates metastasis in bladder cancer. Oncogene (2020) 39(17):3588–603. doi: 10.1038/s41388-020-1236-1
57. Wang X, He H, Rui W, Zhang N, Zhu Y, Xie X. TRIM38 triggers the uniquitination and degradation of glucose transporter type 1 (GLUT1) to restrict tumor progression in bladder cancer. J Transl Med (2021) 19(1):508. doi: 10.1186/s12967-021-03173-x
58. Jiang P, Xu Z, Wu S, Sun J, Tian J, Chen X. RFWD3 acts as a tumor promotor in the development and progression of bladder cancer. Histol Histopathol (2022) 1699-5848(Electronic):18558. doi: 10.14670/HH-18-558
59. Kumar G, Ritch E, Oo HZ. Genome-wide CRISPR screen reveals SLFN11 as a potent mediator of cisplatin sensitivity in muscle-invasive bladder cancer. Urologic Oncology-Seminars Original Investigations (2020) 38(12):905. doi: 10.1016/j.urolonc.2020.10.050
60. Cai H, Xu H, Lu H. LncRNA SNHG1 facilitates tumor proliferation and represses apoptosis by regulating PPARgamma ubiquitination in bladder cancer. Cancers (Basel) (2022) 14(19):4740. doi: 10.3390/cancers14194740
61. Luo Y, He Z, Liu W, Zhou F, Liu T, Wang G. DTL is a prognostic biomarker and promotes bladder cancer progression through regulating the AKT/mTOR axis. Oxid Med Cell Longev (2022) 2022(1942-0994(1942-0994 (Electronic):3369858. doi: 10.1155/2022/3369858
62. Benini M, Fortuni S, Condo I. E3 ligase RNF126 directly ubiquitinates frataxin, promoting its degradation: identification of a potential therapeutic target for friedreich ataxia. Cell Rep (2017) 18(8):2007–17. doi: 10.1016/j.celrep.2017.01.079
63. Castro IH, Ferrari A, Herrera MG. Biophysical characterisation of the recombinant human frataxin precursor. FEBS Open Bio (2018) 8(3):390–405. doi: 10.1002/2211-5463.12376
64. Smith CJ, Berry DM, McGlade CJ. The E3 ubiquitin ligases RNF126 and Rabring7 regulate endosomal sorting of the epidermal growth factor receptor. J Cell Sci (2013) 126(Pt 6):1366–80. doi: 10.1242/jcs.116129
65. Yoshino S, Hara T, Nakaoka HJ. The ERK signaling target RNF126 regulates anoikis resistance in cancer cells by changing the mitochondrial metabolic flux. Cell Discovery (2016) 2(2056-5968(2056-5968 (Print):16019. doi: 10.1038/celldisc.2016.19
66. Huang CY, Lee FL, Peng SF. HSF1 phosphorylation by ERK/GSK3 suppresses RNF126 to sustain IGF-IIR expression for hypertension-induced cardiomyocyte hypertrophy. J Cell Physiol (2018) 233(2):979–89. doi: 10.1002/jcp.25945
67. Wang Y, Deng O, Feng Z. RNF126 promotes homologous recombination via regulation of E2F1-mediated BRCA1 expression. Oncogene (2016) 35(11):1363–72. doi: 10.1038/onc.2015.198
68. Zhi X, Zhao D, Wang Z. E3 ubiquitin ligase RNF126 promotes cancer cell proliferation by targeting the tumor suppressor p21 for ubiquitin-mediated degradation. Cancer Res (2013) 73(1):385–94. doi: 10.1158/0008-5472.CAN-12-0562
69. Migita K, Matsumoto S, Wakatsuki K. RNF126 as a marker of prognosis and proliferation of gastric cancer. Anticancer Res (2020) 40(3):1367–74. doi: 10.21873/anticanres.14078
70. Ashrafizadeh M, Zarrabi A, Samarghandian S, Najafi M. PTEN: what we know of the function and regulation of this onco-suppressor factor in bladder cancer? Eur J Pharmacol (2020) 881(1879-0712(1879-0712 (Electronic):173226. doi: 10.1016/j.ejphar.2020.173226
71. Zhang L, Wang Z, Shi R. RNF126 quenches RNF168 function in the DNA damage response. Genomics Proteomics Bioinf (2018) 16(6):428–38. doi: 10.1016/j.gpb.2018.07.004
72. Ishida N, Nakagawa T, Iemura SI. Ubiquitylation of Ku80 by RNF126 promotes completion of nonhomologous end joining-mediated DNA repair. Mol Cell Biol (2017) 37(4):e00347–16. doi: 10.1128/MCB.00347-16
73. Massari F, Santoni M, Ciccarese C. Emerging concepts on drug resistance in bladder cancer: implications for future strategies. Crit Rev Oncol Hematol (2015) 96(1):81–90. doi: 10.1016/j.critrevonc.2015.05.005
74. Jin X, Kim LJY, Wu Q. Targeting glioma stem cells through combined BMI1 and EZH2 inhibition. Nat Med (2017) 23(11):1352–61. doi: 10.1038/nm.4415
75. Zhang Y, Yang YL, Zhang FL, Liao XH, Shao ZM, Li DQ. Epigenetic silencing of RNF144A expression in breast cancer cells through promoter hypermethylation and MBD4. Cancer Med (2018) 7(4):1317–25. doi: 10.1002/cam4.1324
76. Ho SR, Mahanic CS, Lee YJ, Lin WC. RNF144A, an E3 ubiquitin ligase for DNA-PKcs, promotes apoptosis during DNA damage. Proc Natl Acad Sci USA (2014) 111(26):E2646–55. doi: 10.1073/pnas.1323107111
77. Wu YH, Hong CW, Wang YC. A novel histone deacetylase inhibitor TMU-35435 enhances etoposide cytotoxicity through the proteasomal degradation of DNA-PKcs in triple-negative breast cancer. Cancer Letters (2017) 400(1872-7980(1872-7980 (Electronic):79–88. doi: 10.1016/j.canlet.2017.04.023
78. Rotin D, Kumar S. Physiological functions of the HECT family of ubiquitin ligases. Nat Rev Mol Cell Biol (2009) 10(6):398–409. doi: 10.1038/nrm2690
79. Platta HW, Abrahamsen H, Thoresen SB, Stenmark H. Nedd4-dependent lysine-11-linked polyubiquitination of the tumour suppressor beclin 1. Biochem J (2012) 441(1):399–406. doi: 10.1042/BJ20111424
80. Zheng H, Ke X, Li D. NEDD4 promotes cell growth and motility in hepatocellular carcinoma. Cell Cycle (2018) 17(6):728–38. doi: 10.1080/15384101.2018.1440879
81. Wan LF, Liu T, Hong ZP. NEDD4 expression is associated with breast cancer progression and is predictive of a poor prognosis. Breast Cancer Res (2019) 21(1):148. doi: 10.1186/s13058-019-1236-7
82. Persaud A, Alberts P, Mari S. Tyrosine phosphorylation of NEDD4 activates its ubiquitin ligase activity. Sci Signal (2014) 7(346):ra95. doi: 10.1126/scisignal.2005290
83. Rosenberg JE, Hoffman-Censits J, Powles T. Atezolizumab in patients with locally advanced and metastatic urothelial carcinoma who have progressed following treatment with platinum-based chemotherapy: a single-arm, multicentre, phase 2 trial. Lancet (2016) 387(10031):1909–20. doi: 10.1016/S0140-6736(16)00561-4
84. Tully KH, Jutte H, Wirtz RM. Prognostic role of FGFR alterations and FGFR mRNA expression in metastatic urothelial cancer undergoing checkpoint inhibitor therapy. Urology (2021) 157(1527-9995(1527-9995 (Electronic):93–101. doi: 10.1016/j.urology.2021.05.055
85. Hang X, Zhu S, Di H. NEDD4 depletion inhibits hepatocellular carcinoma growth via targeting PTEN. Cell Physiol Biochem (2016) 39(2):768–79. doi: 10.1159/000445667
86. Petroski MD, Deshaies RJ. Function and regulation of cullin-RING ubiquitin ligases. Nat Rev Mol Cell Biol (2005) 6(1):9–20. doi: 10.1038/nrm1547
87. Hua Z, Vierstra RD. The cullin-RING ubiquitin-protein ligases. Annu Rev Plant Biol (2011) 62(1545-2123(1545-2123 (Electronic):299–334. doi: 10.1146/annurev-arplant-042809-112256
88. Taniguchi K, Karin M. NF-kappaB, inflammation, immunity and cancer: coming of age. Nat Rev Immunol (2018) 18(5):309–24. doi: 10.1038/nri.2017.142
89. Zhang Q, Lenardo MJ, Baltimore D. 30 years of NF-kappaB: a blossoming of relevance to human pathobiology. Cell (2017) 168(1-2):37–57. doi: 10.1016/j.cell.2016.12.012
90. Tenev T, Bianchi K, Darding M. The ripoptosome, a signaling platform that assembles in response to genotoxic stress and loss of IAPs. Mol Cell (2011) 43(3):432–48. doi: 10.1016/j.molcel.2011.06.006
91. Han X, Zhang L, Chung J. UbcH7 regulates 53BP1 stability and DSB repair. Proc Natl Acad Sci USA (2014) 111(49):17456–61. doi: 10.1073/pnas.1408538111
92. Hu S, Yang X. Cellular inhibitor of apoptosis 1 and 2 are ubiquitin ligases for the apoptosis inducer Smac/DIABLO. J Biol Chem (2003) 278(12):10055–60. doi: 10.1074/jbc.M207197200
93. Che XY, Yang DY, Zong HF. Nuclear cIAP1 overexpression is a tumor stage- and grade-independent predictor of poor prognosis in human bladder cancer patients. Urologic Oncology-Seminars Original Investigations (2012) 30(4):450–6. doi: 10.1016/j.urolonc.2010.12.016
94. Mahoney DJ, Cheung HH, Mrad RL. Both cIAP1 and cIAP2 regulate TNFalpha-mediated NF-kappaB activation. Proc Natl Acad Sci USA (2008) 105(33):11778–83. doi: 10.1073/pnas.0711122105
95. Zarnegar BJ, Wang Y, Mahoney DJ. Noncanonical NF-kappaB activation requires coordinated assembly of a regulatory complex of the adaptors cIAP1, cIAP2, TRAF2 and TRAF3 and the kinase NIK. Nat Immunol (2008) 9(12):1371–8. doi: 10.1038/ni.1676
96. Groselj B, Sharma NL, Hamdy FC, Kerr M, Kiltie AE. Histone deacetylase inhibitors as radiosensitisers: effects on DNA damage signalling and repair. Br J Cancer (2013) 108(4):748–54. doi: 10.1038/bjc.2013.21
97. Ibusuki M, Yamamoto Y, Shinriki S, Ando Y, Iwase H. Reduced expression of ubiquitin ligase FBXW7 mRNA is associated with poor prognosis in breast cancer patients. Cancer Sci (2011) 102(2):439–45. doi: 10.1111/j.1349-7006.2010.01801.x
98. Lee YS, Song SJ, Hong HK, Oh BY, Lee WY, Cho YB. The FBW7-MCL-1 axis is key in M1 and M2 macrophage-related colon cancer cell progression: validating the immunotherapeutic value of targeting PI3Kgamma. Exp Mol Med (2020) 52(5):815–31. doi: 10.1038/s12276-020-0436-7
99. Yokobori T, Mimori K, Iwatsuki M. p53-altered FBXW7 expression determines poor prognosis in gastric cancer cases. Cancer Res (2009) 69(9):3788–94. doi: 10.1158/0008-5472.CAN-08-2846
100. Klein AM, de Queiroz RM, Venkatesh D, Prives C. The roles and regulation of MDM2 and MDMX: it is not just about p53. Genes Dev (2021) 35(9-10):575–601. doi: 10.1101/gad.347872.120
101. Shinohara A, Sakano S, Hinoda Y. Association of TP53 and MDM2 polymorphisms with survival in bladder cancer patients treated with chemoradiotherapy. Cancer Sci (2009) 100(12):2376–82. doi: 10.1111/j.1349-7006.2009.01331.x
102. Mao M, Yang L, Hu J. OCT3/4 enhances tumor immune response by upregulating the TET1-dependent NRF2/MDM2 axis in bladder cancer. Genomics (2021) 113(4):2122–33. doi: 10.1016/j.ygeno.2021.04.033
103. Fang DD, Tang Q, Kong Y. MDM2 inhibitor APG-115 synergizes with PD-1 blockade through enhancing antitumor immunity in the tumor microenvironment. J Immunother Cancer (2019) 7(1):327. doi: 10.1186/s40425-019-0750-6
104. Sahin I, Zhang S, Navaraj A, Zhou L, Dizon D, Safran H, et al. AMG-232 sensitizes high MDM2-expressing tumor cells to T-cell-mediated killing. Cell Death Discov (2020) 6(2058-7716(2058-7716 (Print):57. doi: 10.1038/s41420-020-0292-1
105. Brunelli M, Tafuri A, Cima L. MDM2 gene amplification as selection tool for innovative targeted approaches in PD-L1 positive or negative muscle-invasive urothelial bladder carcinoma. J Clin Pathol (2022) 75(1):39–44. doi: 10.1136/jclinpath-2020-207089
106. Tang Y, Lei Y, Gao P. Pan-cancer analysis and experimental validation of DTL as a potential diagnosis, prognosis and immunotherapy biomarker. BMC Cancer (2023) 23(1):328. doi: 10.1186/s12885-023-10755-z
107. Burns JE, Hurst CD, Knowles MA, Phillips RM, Allison SJ. The warburg effect as a therapeutic target for bladder cancers and intratumoral heterogeneity in associated molecular targets. Cancer Sci (2021) 112(9):3822–34. doi: 10.1111/cas.15047
108. Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell (2011) 144(5):646–74. doi: 10.1016/j.cell.2011.02.013
109. Ancey PB, Contat C, Meylan E. Glucose transporters in cancer - from tumor cells to the tumor microenvironment. FEBS J (2018) 285(16):2926–43. doi: 10.1111/febs.14577
110. Freemerman AJ, Johnson AR, Sacks GN. Metabolic reprogramming of macrophages: glucose transporter 1 (GLUT1)-mediated glucose metabolism drives a proinflammatory phenotype. J Biol Chem (2014) 289(11):7884–96. doi: 10.1074/jbc.M113.522037
111. Lange SM, Armstrong LA, Kulathu Y. Deubiquitinases: from mechanisms to their inhibition by small molecules. Mol Cell (2022) 82(1):15–29. doi: 10.1016/j.molcel.2021.10.027
112. Li Q, Chao Q, Liu Y. Deubiquitinase ZRANB1 drives hepatocellular carcinoma progression through SP1-LOXL2 axis. Am J Cancer Res (2021) 11(10):4807.
113. Zhao J, Guo J, Wang Y. Research progress of DUB enzyme in hepatocellular carcinoma. Front Oncol (2022) 12(2234-943X:920287(2234-943X (Print). doi: 10.3389/fonc.2022.920287
114. Clague MJ, Urbe S, Komander D. Breaking the chains: deubiquitylating enzyme specificity begets function. Nat Rev Mol Cell Biol (2019) 20(6):338–52. doi: 10.1038/s41580-019-0099-1
115. Zhou XJ, Li R, Liu X, Qu YQ. Advances in deubiquitinating enzymes in lung adenocarcinoma. J Cancer (2021) 12(18):5573–82. doi: 10.7150/jca.56532
116. Dai X, Lu L, Deng S. USP7 targeting modulates anti-tumor immune response by reprogramming tumor-associated macrophages in lung cancer. Theranostics (2020) 10(20):9332–47. doi: 10.7150/thno.47137
117. Wang ZR, Kang WT, Li OW. Abrogation of USP7 is an alternative strategy to downregulate PD-L1 and sensitize gastric cancer cells to T cells killing. Acta Pharm Sin B (2021) 11(3):694–707. doi: 10.1016/j.apsb.2020.11.005
118. Peng Y, Liu J, Wang Z. Prostate-specific oncogene OTUD6A promotes prostatic tumorigenesis via deubiquitinating and stabilizing c-myc. Cell Death Differ (2022) 29(9):1730–43. doi: 10.1038/s41418-022-00960-x
119. Luo Y, Zhou J, Tang J. MINDY1 promotes bladder cancer progression by stabilizing YAP. Cancer Cell Int (2021) 21(1):395. doi: 10.1186/s12935-021-02095-4
120. Guo Y, Cui S, Chen Y, Guo S, Chen D. Ubiquitin specific peptidases and prostate cancer. PeerJ (2023) 11(2167-8359(2167-8359 (Electronic):e14799. doi: 10.7717/peerj.14799
121. Hou T, Dan W, Liu T. Deubiquitinase OTUD5 modulates mTORC1 signaling to promote bladder cancer progression. Cell Death Dis (2022) 13(9):778. doi: 10.1038/s41419-022-05128-6
122. Zhang HH, Li C, Ren JW. OTUB1 facilitates bladder cancer progression by stabilizing ATF6 in response to endoplasmic reticulum stress. Cancer Sci (2021) 112(6):2199–209. doi: 10.1111/cas.14876
123. Liu T, Jiang L, Tavana O, Gu W. The deubiquitylase OTUB1 mediates ferroptosis via stabilization of SLC7A11. Cancer Res (2019) 79(8):1913–24. doi: 10.1158/0008-5472.CAN-18-3037
124. Cao Y, Yan X, Bai X. UCHL5 promotes proliferation and migration of bladder cancer cells by activating c-myc via AKT/mTOR signaling. Cancers (Basel) (2022) 14(22):5538. doi: 10.3390/cancers14225538
125. Chow PM, Dong JR, Chang YW. The UCHL5 inhibitor b-AP15 overcomes cisplatin resistance via suppression of cancer stemness in urothelial carcinoma. Mol Ther Oncolytics (2022) 26(2372-7705(2372-7705 (Print):387–98. doi: 10.1016/j.omto.2022.08.004
126. He H, Yi L, Zhang B. USP24-GSDMB complex promotes bladder cancer proliferation via activation of the STAT3 pathway. Int J Biol Sci (2021) 17(10):2417–29. doi: 10.7150/ijbs.54442
127. Man X, Piao C, Lin X, Kong C, Cui X, Jiang Y. USP13 functions as a tumor suppressor by blocking the NF-kB-mediated PTEN downregulation in human bladder cancer. J Exp Clin Cancer Res (2019) 38(1):386. doi: 10.1186/s13046-019-1262-4
128. Morra F, Merolla F, Criscuolo D. CCDC6 and USP7 expression levels suggest novel treatment options in high-grade urothelial bladder cancer. J Exp Clin Cancer Res (2019) 38(1):90. doi: 10.1186/s13046-019-1087-1
129. Malapelle U, Morra F, Ilardi G. USP7 inhibitors, downregulating CCDC6, sensitize lung neuroendocrine cancer cells to PARP-inhibitor drugs. Lung Cancer (2017) 107(1872-8332(1872-8332 (Electronic):41–9. doi: 10.1016/j.lungcan.2016.06.015
130. Xu J, Hua X, Jin H. NFkappaB2 p52 stabilizes rhogdibeta mRNA by inhibiting AUF1 protein degradation via a miR-145/Sp1/USP8-dependent axis. Mol Carcinog (2019) 58(5):777–93. doi: 10.1002/mc.22970
131. Huang J, Zhou WM, Hao C, He QM, Tu XH. The feedback loop of METTL14 and USP38 regulates cell migration, invasion and EMT as well as metastasis in bladder cancer. PloS Genet (2022) 18(10):e1010366. doi: 10.1371/journal.pgen.1010366
132. Jeong P, Ha YS, Yun SJ. Assess the expression of ubiquitin specific protease USP2a for bladder cancer diagnosis. BMC Urol (2015) 15(1471-2490(1471-2490 (Electronic):80. doi: 10.1186/s12894-015-0074-x
133. Lv L, Xiao XY, Gu ZH, Zeng FQ, Huang LQ, Jiang GS. Silencing USP22 by asymmetric structure of interfering RNA inhibits proliferation and induces cell cycle arrest in bladder cancer cells. Mol Cell Biochem (2011) 346(1-2):11–21. doi: 10.1007/s11010-010-0585-4
134. Kim YH, Kim WT, Jeong P. Novel combination markers for predicting survival in patients with muscle invasive bladder cancer: USP18 and DGCR2. J Korean Med Sci (2014) 29(3):351–6. doi: 10.3346/jkms.2014.29.3.351
135. Guo GY, Xu YY, Gong MC, Cao Y, An RH. USP28 is a potential prognostic marker for bladder cancer. Tumor Biol (2014) 35(5):4017–22. doi: 10.1007/s13277-013-1525-1
136. Tyagi A, Haq S, Ramakrishna S. Redox regulation of DUBs and its therapeutic implications in cancer. Redox Biol (2021) 48(2213-2317(2213-2317 (Electronic):102194. doi: 10.1016/j.redox.2021.102194
137. Kayagaki N, Phung Q, Chan S. DUBA: a deubiquitinase that regulates type I interferon production. Science (2007) 318(5856):1628–32. doi: 10.1126/science.1145918
138. de Vivo A, Sanchez A, Yegres J, Kim J, Emly S, Kee Y. The OTUD5-UBR5 complex regulates FACT-mediated transcription at damaged chromatin. Nucleic Acids Res (2019) 47(2):729–46. doi: 10.1093/nar/gky1219
139. Rutz S, Kayagaki N, Phung QT. Deubiquitinase DUBA is a post-translational brake on interleukin-17 production in T cells. Nature (2015) 518(7539):417–21. doi: 10.1038/nature13979
140. Wolfson RL, Chantranupong L, Saxton RA. METABOLISM Sestrin2 is a leucine sensor for the mTORC1 pathway. Science (2016) 351(6268):43–8. doi: 10.1126/science.aab2674
141. Budanov AV, Karin M. p53 target genes sestrin1 and sestrin2 connect genotoxic stress and mTOR signaling. Cell (2008) 134(3):451–60. doi: 10.1016/j.cell.2008.06.028
142. Tang D, Chen X, Kang R, Kroemer G. Ferroptosis: molecular mechanisms and health implications. Cell Res (2021) 31(2):107–25. doi: 10.1038/s41422-020-00441-1
143. Koppula P, Zhuang L, Gan B. Cystine transporter SLC7A11/xCT in cancer: ferroptosis, nutrient dependency, and cancer therapy. Protein Cell (2021) 12(8):599–620. doi: 10.1007/s13238-020-00789-5
144. Abdul Rehman SA, Kristariyanto YA, Choi SY. MINDY-1 is a member of an evolutionarily conserved and structurally distinct new family of deubiquitinating enzymes. Mol Cell (2016) 63(1):146–55. doi: 10.1016/j.molcel.2016.05.009
145. Yu FX, Zhao B, Guan KL. Hippo pathway in organ size control, tissue homeostasis, and cancer. Cell (2015) 163(4):811–28. doi: 10.1016/j.cell.2015.10.044
146. Kedan A, Verma N, Saroha A, et al. PYK2 negatively regulates the hippo pathway in TNBC by stabilizing TAZ protein. Cell Death Dis (2018) 9(10):985. doi: 10.1038/s41419-018-1005-z
147. Baek SW, Mun JY, Jang IH. YAP1 activation is associated with the progression and response to immunotherapy of non-muscle invasive bladder cancer. Ebiomedicine (2022) 81:104092. doi: 10.1016/j.ebiom.2022.104092
148. Yang Y, Cao L, Guo Z, Gu H, Zhang K, Qiu Z. Deubiquitinase UCHL5 stabilizes ELK3 to potentiate cancer stemness and tumor progression in pancreatic adenocarcinoma (PAAD). Exp Cell Res (2022) 421(2):113402. doi: 10.1016/j.yexcr.2022.113402
149. Arpalahti L, Laitinen A, Hagstrom J. Positive cytoplasmic UCHL5 tumor expression in gastric cancer is linked to improved prognosis. PloS One (2018) 13(2):e0193125. doi: 10.1371/journal.pone.0193125
150. Liu D, Song Z, Wang X, Ouyang L. Ubiquitin c-terminal hydrolase L5 (UCHL5) accelerates the growth of endometrial cancer via activating the wnt/beta-catenin signaling pathway. Front Oncol (2020) 10(2234-943X:865(2234-943X (Print). doi: 10.3389/fonc.2020.00865
151. Cutts AJ, Soond SM, Powell S, Chantry A. Early phase TGFbeta receptor signalling dynamics stabilised by the deubiquitinase UCH37 promotes cell migratory responses. Int J Biochem Cell Biol (2011) 43(4):604–12. doi: 10.1016/j.biocel.2010.12.018
152. Fang Y, Shen X. Ubiquitin carboxyl-terminal hydrolases: involvement in cancer progression and clinical implications. Cancer Metastasis Rev (2017) 36(4):669–82. doi: 10.1007/s10555-017-9702-0
153. Wicks SJ, Haros K, Maillard M. The deubiquitinating enzyme UCH37 interacts with smads and regulates TGF-beta signalling. Oncogene (2005) 24(54):8080–4. doi: 10.1038/sj.onc.1208944
154. Sooman L, Gullbo J, Bergqvist M, Bergstrom S, Lennartsson J, Ekman S. Synergistic effects of combining proteasome inhibitors with chemotherapeutic drugs in lung cancer cells. BMC Res Notes (2017) 10(1):544. doi: 10.1186/s13104-017-2842-z
155. Wang YC, Wu YS, Hung CY. USP24 induces IL-6 in tumor-associated microenvironment by stabilizing p300 and beta-TrCP and promotes cancer malignancy. Nat Commun (2018) 9(1):3996. doi: 10.1038/s41467-018-06178-1
156. Liu J, Xia H, Kim M. Beclin1 controls the levels of p53 by regulating the deubiquitination activity of USP10 and USP13. Cell (2011) 147(1):223–34. doi: 10.1016/j.cell.2011.08.037
157. Zhang J, Zhang P, Wei Y. Deubiquitylation and stabilization of PTEN by USP13. Nat Cell Biol (2013) 15(12):1486–94. doi: 10.1038/ncb2874
158. Zhao X, Fiske B, Kawakami A, Li J, Fisher DE. Regulation of MITF stability by the USP13 deubiquitinase. Nat Commun (2011) 2(2041-1723(2041-1723 (Electronic):414. doi: 10.1038/ncomms1421
159. Vasudevan KM, Gurumurthy S, Rangnekar VM. Suppression of PTEN expression by NF-kappa b prevents apoptosis. Mol Cell Biol (2004) 24(3):1007–21. doi: 10.1128/MCB.24.3.1007-1021.2004
160. Kim S, Domon-Dell C, Kang J, Chung DH, Freund JN, Evers BM. Down-regulation of the tumor suppressor PTEN by the tumor necrosis factor-alpha/nuclear factor-kappaB (NF-kappaB)-inducing kinase/NF-kappaB pathway is linked to a default IkappaB-alpha autoregulatory loop. J Biol Chem (2004) 279(6):4285–91. doi: 10.1074/jbc.M308383200
161. Yuan F, Lou Z, Zhou Z, Yan X. Long non−coding RNA KCNQ1OT1 promotes nasopharyngeal carcinoma cell cisplatin resistance via the miR−454/USP47 axis. Int J Mol Med (2021) 47(4):54. doi: 10.3892/ijmm.2021.4887
162. Morra F, Merolla F, Damia G. The disruption of the CCDC6 - PP4 axis induces a BRCAness like phenotype and sensitivity to PARP inhibitors in high-grade serous ovarian carcinoma. J Exp Clin Cancer Res (2022) 41(1):245. doi: 10.1186/s13046-022-02459-2
163. Morra F, Merolla F, D'Abbiero D. Analysis of CCDC6 as a novel biomarker for the clinical use of PARP1 inhibitors in malignant pleural mesothelioma. Lung Cancer (2019) 135(1872-8332(1872-8332 (Electronic):56–65. doi: 10.1016/j.lungcan.2019.07.011
164. Wolacewicz M, Hrynkiewicz R, Grywalska E. Immunotherapy in bladder cancer: current methods and future perspectives. Cancers (Basel) (2020) 12(5):1181. doi: 10.3390/cancers12051181
165. Ward Grados DF, Ahmadi H, Griffith TS, Warlick CA. Immunotherapy for bladder cancer: latest advances and ongoing clinical trials. Immunol Invest (2022) 51(8):2226–51. doi: 10.1080/08820139.2022.2118606
166. Pettenati C, Ingersoll MA. Mechanisms of BCG immunotherapy and its outlook for bladder cancer. Nat Rev Urol (2018) 15(10):615–25. doi: 10.1038/s41585-018-0055-4
167. Rhea LP, Mendez-Marti S, Kim D, Aragon-Ching JB. Role of immunotherapy in bladder cancer. Cancer Treat Res Commun (2021) 26(2468-2942(2468-2942 (Electronic):100296. doi: 10.1016/j.ctarc.2020.100296
168. Rhea LP, Aragon-Ching JB. Advances and controversies with checkpoint inhibitors in bladder cancer. Clin Med Insights Oncol (2021) 15(1179-5549(1179-5549 (Print):11795549211044963. doi: 10.1177/11795549211044963
169. Lim SO, Li CW, Xia W. Deubiquitination and stabilization of PD-L1 by CSN5. Cancer Cell (2016) 30(6):925–39. doi: 10.1016/j.ccell.2016.10.010
170. Meng X, Liu X, Guo X. FBXO38 mediates PD-1 ubiquitination and regulates anti-tumour immunity of T cells. Nature (2018) 564(7734):130–5. doi: 10.1038/s41586-018-0756-0
171. Mezzadra R, Sun C, Jae LT. Identification of CMTM6 and CMTM4 as PD-L1 protein regulators. Nature (2017) 549(7670):106–10. doi: 10.1038/nature23669
172. Hou B, Chen T, Zhang H, Li J, Wang P, Shang G. The E3 ubiquitin ligases regulate PD-1/PD-L1 protein levels in tumor microenvironment to improve immunotherapy. Front Immunol (2023) 14(1664-3224:1123244(1664-3224 (Electronic). doi: 10.3389/fimmu.2023.1123244
173. Korenev G, Yakukhnov S, Druk A. USP7 inhibitors in cancer immunotherapy: current status and perspective. Cancers (Basel) (2022) 14(22):5539. doi: 10.3390/cancers14225539
174. Wang Y, Sun Q, Mu N. The deubiquitinase USP22 regulates PD-L1 degradation in human cancer cells. Cell Commun Signal (2020) 18(1):112. doi: 10.1186/s12964-020-00612-y
175. Jingjing W, Wenzheng G, Donghua W, Guangyu H, Aiping Z, Wenjuan W. Deubiquitination and stabilization of programmed cell death ligand 1 by ubiquitin-specific peptidase 9, X-linked in oral squamous cell carcinoma. Cancer Med (2018) 7(8):4004–11. doi: 10.1002/cam4.1675
176. Wang X, Zhang Y, Wu Y, Cheng H, Wang X. The role of E3 ubiquitin ligases and deubiquitinases in bladder cancer development and immunotherapy. Front Immunol (2023) 14:1202633. doi: 10.3389/fimmu.2023.1202633
177. Poondla N, Chandrasekaran AP, Kim KS, Ramakrishna S. Deubiquitinating enzymes as cancer biomarkers: new therapeutic opportunities? BMB Rep (2019) 52(3):181–9. doi: 10.5483/BMBRep.2019.52.3.048
178. Powles T, Rosenberg JE, Sonpavde GP. Enfortumab vedotin in previously treated advanced urothelial carcinoma. N Engl J Med (2021) 384(12):1125–35. doi: 10.1056/NEJMoa2035807
179. Hoimes CJ, Flaig TW, Milowsky MI. Enfortumab vedotin plus pembrolizumab in previously untreated advanced urothelial cancer. J Clin Oncol (2023) 41(1):22–31. doi: 10.1200/JCO.22.01643
180. Wong JL, Rosenberg JE. Targeting nectin-4 by antibody-drug conjugates for the treatment of urothelial carcinoma. Expert Opin Biol Ther (2021) 21(7):863–73. doi: 10.1080/14712598.2021.1929168
181. Wang S, Xu L, Che X. E3 ubiquitin ligases cbl-b and c-cbl downregulate PD-L1 in EGFR wild-type non-small cell lung cancer. FEBS Lett (2018) 592(4):621–30. doi: 10.1002/1873-3468.12985
Keywords: bladder cancer, E3 ubiquitin ligase, deubiquitinases, immunotherapy, tumorigenesis
Citation: Wang M, Zhang Z, Li Z, Zhu Y and Xu C (2023) E3 ubiquitin ligases and deubiquitinases in bladder cancer tumorigenesis and implications for immunotherapies. Front. Immunol. 14:1226057. doi: 10.3389/fimmu.2023.1226057
Received: 20 May 2023; Accepted: 23 June 2023;
Published: 11 July 2023.
Edited by:
Xiangpeng Dai, Jilin University, ChinaReviewed by:
Jianli Tao, Boston Children’s Hospital and Harvard Medical School, United StatesCopyright © 2023 Wang, Zhang, Li, Zhu and Xu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yasheng Zhu, enlzMDU2MkBmb3htYWlsLmNvbQ==; Chuanliang Xu, Y2h1YW5saWFuZ194dUAxMjYuY29t
†These authors have contributed equally to this work and share first authorship
‡These authors share last authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.