
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Immunol. , 16 February 2023
Sec. Alloimmunity and Transplantation
Volume 14 - 2023 | https://doi.org/10.3389/fimmu.2023.1137629
 Juliette Rousse1*
Juliette Rousse1* Pierre-Joseph Royer1
Pierre-Joseph Royer1 Gwénaëlle Evanno1Elsa Lheriteau1
Gwénaëlle Evanno1Elsa Lheriteau1 Carine Ciron1
Carine Ciron1 Apolline Salama2Françoise Shneiker1
Apolline Salama2Françoise Shneiker1 Roberto Duchi3
Roberto Duchi3 Andrea Perota3
Andrea Perota3 Cesare Galli3
Cesare Galli3 Emmanuele Cozzi4
Emmanuele Cozzi4 Gilles Blancho2Odile Duvaux1
Gilles Blancho2Odile Duvaux1 Sophie Brouard2
Sophie Brouard2 Jean-Paul Soulillou2
Jean-Paul Soulillou2 Jean-Marie Bach5Bernard Vanhove1
Jean-Marie Bach5Bernard Vanhove1Anti-thymocyte or anti-lymphocyte globulins (ATGs/ALGs) are immunosuppressive drugs used in induction therapies to prevent acute rejection in solid organ transplantation. Because animal-derived, ATGs/ALGs contain highly immunogenic carbohydrate xenoantigens eliciting antibodies that are associated with subclinical inflammatory events, possibly impacting long-term graft survival. Their strong and long-lasting lymphodepleting activity also increases the risk for infections. We investigated here the in vitro and in vivo activity of LIS1, a glyco-humanized ALG (GH-ALG) produced in pigs knocked out for the two major xeno-antigens αGal and Neu5Gc. It differs from other ATGs/ALGs by its mechanism of action excluding antibody-dependent cell-mediated cytotoxicity and being restricted to complement-mediated cytotoxicity, phagocyte-mediated cytotoxicity, apoptosis and antigen masking, resulting in profound inhibition of T-cell alloreactivity in mixed leucocyte reactions. Preclinical evaluation in non-human primates showed that GH-ALG dramatically reduced CD4+ (p=0.0005,***), CD8+ effector T cells (p=0.0002,***) or myeloid cells (p=0.0007,***) but not T-reg (p=0.65, ns) or B cells (p=0.65, ns). Compared with rabbit ATG, GH-ALG induced transient depletion (less than one week) of target T cells in the peripheral blood (<100 lymphocytes/L) but was equivalent in preventing allograft rejection in a skin allograft model. The novel therapeutic modality of GH-ALG might present advantages in induction treatment during organ transplantation by shortening the T-cell depletion period while maintaining adequate immunosuppression and reducing immunogenicity.
“Induction therapies” refer to early lymphodepletion or blockade to avoid acute graft rejection in solid organ transplantation. They also reduce nephrotoxicity by delaying the introduction of calcineurin inhibitors which are maintenance agents (1, 2). They comprise CD25 antagonists (3), anti-CD52 antibody and anti-lymphocyte/thymocyte globulins (ALGs/ATGs) (4, 5). ALGs/ATGs are polyclonal antibodies directed against T- and B-lymphocytes (6), obtained from rabbits or horses immunized with primary human thymocytes or human T-cell lines (7, 8). Their activity rely on apoptosis induction (9, 10), complement-dependent cytotoxicity (CDC), antibody-dependent cell cytotoxicity (ADCC), phagocytosis and other nondestructive mechanisms (6).
ALGs/ATGs have been used for 50 years (4, 6) and have proven efficacy in preventing acute graft rejection. However, they are endowed with a strong immunogenicity associated with side effects ranging from mild fever or skin rashes to more serious serum sickness disease (SSD) (11, 12) or anaphylactic shock (13–15). SSD is a hypersensitivity reaction toward glycoproteins from animal sources (16) caused by antibodies directed against glycans bearing the common mammalian sialic acid N-glycolylneuraminic acid (Neu5Gc) (17–21). Humans present a biased sialylation of glycoproteins and glycolipids. The lack of Neu5Gc (22) on proteins or lipids is due to a human lineage-specific genetic mutation in the enzyme cytidine monophosphate-N-acetylneuraminic acid hydroxylase (CMAH) (22). A direct consequence is that Neu5Gc epitopes are excluded from «self-tolerance» and elicit anti-Neu5Gc antibodies in humans (17, 23–26). In the same way, humans also lack the α1,3-galactosyl-transferase enzyme (GGTA1) (27) and are not tolerant to α1,3 galactose (Gal) epitopes; they present various levels of natural anti-a1,3Gal antibodies and increase their level of anti-a1,3Gal antibodies after infusion with animal-derived products (28–30).
Rabbit ATG given to diabetic patients resulted in highly significant increases in anti-Gal and -Neu5Gc IgG/IgM, still detectable one year post-infusion and responsible for the induction of SSD and immune complex disease in almost all patients (25). Even in the context of concomitant administration of immunosuppressants and steroids, however, such as in organ transplantation, SSD still occurs in 10% of recipients who received rabbit ALG/ATG (11). SSD, however, is a contributing factor to late graft loss following ALG/ATG induction (11) and high anti-Neu5Gc antibody level after kidney transplantation has also been associated with late graft loss (11). This effect may be due to the well-known passive incorporation of Neu5Gc residues from food origin into endothelial and epithelial cells (31, 32), possibly resulting in a chronic inflammation called “xenosialitis” in the presence of anti-Neu5Gc antibodies, although the detrimental role of these “natural antibodies” in human still needs to be confirmed (24, 33) (34),. In parallel, anti-Gal antibody responses can be involved in anaphylaxis-type reactions targeting glycoconjugates present on therapeutic products of animal origin (23, 25).
Rabbit ATG is known for its rapid, intense and persistent depletion. Indeed, it is characterized by a depletion of 83-92% of almost all lymphocyte subpopulations, observable on the day following treatment. It also results in leukopenia and thrombocytopenia (6). This depletion usually lasts for up to 3 months and may even extend to 6 months, which increases the risk of infections (35, 36). Anti-CD25 monoclonal antibodies (α chain of the IL-2 receptor) do not cause lymphocyte depletion. Its mechanism is based on blocking the binding of IL-2 to its receptor, which is only expressed by activated cells (37). Clinically, basiliximab has comparable efficacy to ATG in terms of survival at one year and reduces infections, but at the cost of a slightly higher rate of acute rejection (2, 38). Another anti-CD52 monoclonal antibody is also used: alemtuzumab. This causes massive depletion of peripheral blood T cells (CD52 is found on T and B lymphocytes, monocytes, macrophages and granulocytes) (39). However, alemtuzumab is not widely used in the clinic, as it has not yet been the subject of a prospective, randomized study (38).
To address these issues, a “glyco-humanized” polyclonal serum, has been developed in pigs knocked out for the two genes CMAH and GGTA1 so that the two main xenogenic glycoantigens have been changed into human-type glycoantigens: the a1,3Gal epitopes are replaced by Gal β1,4-GlucNac, and Neu5Gc is replaced by Neu5Ac. The mechanisms of action of GH-ALG were investigated in vitro on human PBMC and in vivo in nonhuman primates. They differ from human rabbit immunoglobulins because they have the unique property of not interacting with human FcγR, which alters the mechanism of action by altering ADCC (40).
Double KO defined high health status pigs were immunized with a human CD4+ CD8+ T-cell line. GH-ALG, the IgG fraction, was purified from serum in compliance with good manufacturing practice (GMP) and ICH guidelines. Details have been published elsewhere (40).
Human peripheral blood mononuclear cells (PBMCs) were plated at 1.105 cells per well in 96-well plates and then were labeled with GH-ALG (250 µg/mL) and/or with the following monoclonal antibodies used at a 1/100 dilution: anti-human TCRa/b-FITC (Clone T10B9.1A.31); anti-human CD2-PE (Clone RPA-2.10); anti-human CD3-PE (Clone HIT3a); anti-human CD4-PE (Clone RPA-T4); anti-human CD8-PE (RPA-T8); and anti-human CD28-PE (CD28.2) (all from BD Biosciences, Franklin Lakes, United States). Cell fluorescence was then measured by flow cytometry. A mean fluorescence intensity drop indicated competition between GH-ALG and the test antibody.
The active fraction of GH-ALG was determined by serial depletion against the cells used for immunization. Briefly, 30x106 target cells fixed in 4% paraformaldehyde were resuspended in 1 mL of GH-ALG (1.6 mg/ml) in phosphate buffered saline (PBS). After 20 min of incubation at room temperature, the cells were pelleted and the supernatant was transferred to a second tube containing a new fresh cell pellet to repeat the incubation. Five successive incubations were performed. The remaining specific antibodies were monitored after each incubation using a binding assay. The total IgG concentration was evaluated by spectrophotometry. The GH-ALG active fraction was defined as the fraction of IgG cleared using this repeated depletion procedure.
The binding of GH-ALG to PBMC, red blood cells and platelets from healthy was measured by flow cytometry. Briefly, 105 blood cells were incubated with increasing concentrations of pig or rabbit ALGs/ATGs for 30 min at 4°C. After 3 washes, bound IgG were detected using FITC-conjugated anti-pig or anti-rabbit antibody (both from Bio-Rad) or AF-488 conjugated protein G (Thermo Fisher, Waltham, MA), and analyzed on a BD CELESTA Flow cytometer.
GH-ALG and rabbit-ATG are coated on a maxisorp plate at 50ug/mL. After saturation and washing, revelation is performed with Alexa Fluor 488 conjugated protein G diluted at 1/200 or 1/400 and the reading performed in fluorescence with TECAN.
To perform the kinetic binding analysis of pig and rabbit IgG to protein G, we use the BLItz technique (VWR, Radnor, Pennsylvanie, United States) using bio-layer interferometry technology. Briefly, a first step of hydration of the G-protein biosensors for 10min is followed by loading the biosensors with GH-ALG or rabbit-ATG at 10ug/mL. Reading with BLItz for 2-5min. Results are interpreted using the affinity constant between IgGs and protein G.
PBMCs were mixed with increasing doses of GH-ALG or rabbit ATG in RPMI medium with 10% FCS. Nonimmune IgG was used as a negative control. After 3 h of culture (37°C, 5% CO2), the cells were labeled with AF488-conjugated Annexin V and DAPI (Thermo Fisher) before analysis by flow cytometry. The percentages of cells in early apoptosis (Annexin V+/DAPI- cells) and late apoptosis (Annexin V+/DAPI+) were combined to determine the overall % of apoptosis.
Human platelets purified from citrated blood were mixed with increasing doses of GH-ALG or rabbit ATG and aggregation was monitored for 60 minutes on a TA-8V STAGO aggregometer. To induce platelet aggregation, ristocetin (1.5 mg/ml), Thrombin Receptor Activator Peptide (TRAP,1.5 µM), arachidonic acid (0.3 mg/ml), ADP (adenosine diphosphate, 5 µM), epinephrine (5 µM) and collagen (1.25 µg/ml) were evaluated.
Monocytes were purified from PBMCs by CD14 positive magnetic selection (Miltenyi Biotec, Bergisch Gladbach, Germany) and plated at 2.106 cells/ml in RPMI 1640 medium containing 10% fetal calf serum (FCS). Monocyte differentiation into macrophages was performed for 5-7 days in the presence of human M-CSF (100 ng/ml) (R&D system). On the day of the assay, CFSE-labeled CD3 lymphocytes were incubated with GH-ALG in RPMI medium with 10% heat-inactivated FCS for 30 min at 4°C. Lymphocytes were washed twice with medium and cultured with human macrophages (ratio 1:1). After 3 hours of culture, the cells were washed twice, and macrophages were labeled with CD14-BV421 for 30 min at 4°C, and analysis by flow cytometry. Phagocytosis was assessed as the percentage of double-positive (CFSE+/CD14+) cells over CD14+ cells.
Proliferation was assessed using a CFSE dilution assay. Briefly, stimulator PBMCs were irradiated at 35 Gy and cultured (ratio 4:1) with CFSE-labeled responder PBMCs (Invitrogen, Waltham, Massachusetts, United States of America). The cells were cultured for 3 days in RPMI 10% FCS in the presence or absence of GH-ALG or rabbit ATG and analyzed by flow cytometry. Groups were compared using ANOVA and the Tukey−Kramer multiple comparison test. The results were expressed as %FITC low × number of lymphocytes to analyze cell proliferation of the residual cells.
Eighteen cynomolgus monkeys were used after authorization of the French Research Ministry (APAFIS 10717)/Citoxlab France Ethics Committee (CEC)/Animal Welfare Body of Cynbiose/Ethics Committee of VetAgro-Sup. The characteristics of these in vivo studies are reported in Supplementary Table 1. Cynomolgus monkeys received GH-ALG by daily intravenous administration at doses ranging from 40 to 75 mg/kg/administration for 5 days. GH-ALG was administered as a solution in the vehicle (sterile NaCl 0.9%) under a constant infusion rate. In the rabbit ATG group, 2 macaques were injected 5 times at 5mg/kg intravenously.
CD3+ T lymphocyte numeration was performed using the BD Trucount system (BD Biosciences). Whole blood cells were labeled with the fluorochrome-labeled monoclonal antibody anti-CD3 (BD Biosciences). Other lymphocyte subpopulations were studied by flow cytometry. Briefly, fresh peripheral blood cells were stained with fluorochrome-labeled monoclonal antibodies against CD3 (SP34-2), CD4 (RPA-T4), CD8 (RPA-T8), CD20 (2H7), CD25 (M-A251), CD27 (T27.1), CD28 (CD28.2), CD31 (WM-59), CD45 (D058-1283), CD45-RA (5H9), CD95 (DX2), CD127 (hIL-7R-M21), GzmB, and FoxP3 (236A/E7) (Beckton Dickinson, San Jose, CA, USA). Polychromatic flow cytometric analyses were performed using a Canto II analyzer (BD Biosciences).
The GH-ALG concentration in monkey serum was determined using a pig IgG ELISA (Bethyl, Montgomery, Tx) according to the manufacturer’s instructions.
Skin grafts were performed on 5 anesthetized female cynomolgus monkeys (Supplemental Table 1). After back skin shearing and asepsis, the animals were grafted in pairs (back skin collection using a 20 mm diameter template, followed by grafting and skin graft suturing). In the back of each animal, three grafts were performed: one autograft and two pairwise allografts. In each group, one animal received skin transplants of his congener and vice versa. Three of them (Group 1) received GH-ALG at 75 mg/kg once a day for 5 consecutive days. The injection was performed intravenously (concentration: 10 mg/mL; flow rate: 4 mL/kg/h; infusion duration: approximately 1 h50). The two remaining monkeys (Group 2) were used as controls and did not receive treatment.
All statistical analyses were performed using GraphPad Software (GraphPad Software, San Diego, CA). p values of <0.05 were considered statistically significant.
The binding of GH-ALG to human PBMCs, platelets or red blood cells was investigated by flow cytometry and compared with that of rabbit ATG (Figure 1). A dose-dependent and saturable signal was obtained for GH-ALG and rabbit ATG and was higher for rabbit ATG with maximum MFI at 40,000 for GH-ALG and 50,000 for thymoglobulin. IgG from nonimmunized pigs was used as a negative control and displayed negligible binding (Mean fluorescence intensity, MFI < 82). GH-ALG binding to red blood cells was very low (MFI < 5000; Figure 1A) for GH-ALG and rabbit ATG. Binding to platelets was barely detectable for both preparations (MFI<5000; Figure 1A). However, the comparison between GH-ALG and rabbit ATG binding was questionable because two different detection systems (anti-pig and anti-rabbit antibodies) were used. To accurately compare the binding of GH-ALG and rabbit ATG to platelets, we used AF-488-conjugated protein G to similarly detect both pig- and rabbit-bound IgG. The ability of Protein-G to equivalently recognize pig and rabbit IgG was first confirmed by ELISA and surface plasmon resonance (Supplemental Figure 1). The binding of GH-ALG to human platelets was significantly lower, from 100ug/mL to 1000ug/mL, than that of rabbit ATG in the presence of Protein G. Next, we performed platelet assays to determine whether aggregation was altered or induced by GH-ALG or rabbit ATG. No inhibition of induced platelet aggregation was observed with GH-ALG (not shown). Both preparations induced platelet aggregation in this assay but more strongly and faster (visible after 3 or 60 minutes respectively) with rabbit ATG and at lower concentrations (from 250ug/mL) than with GH-ALG (500ug/mL) (Figure 1C).
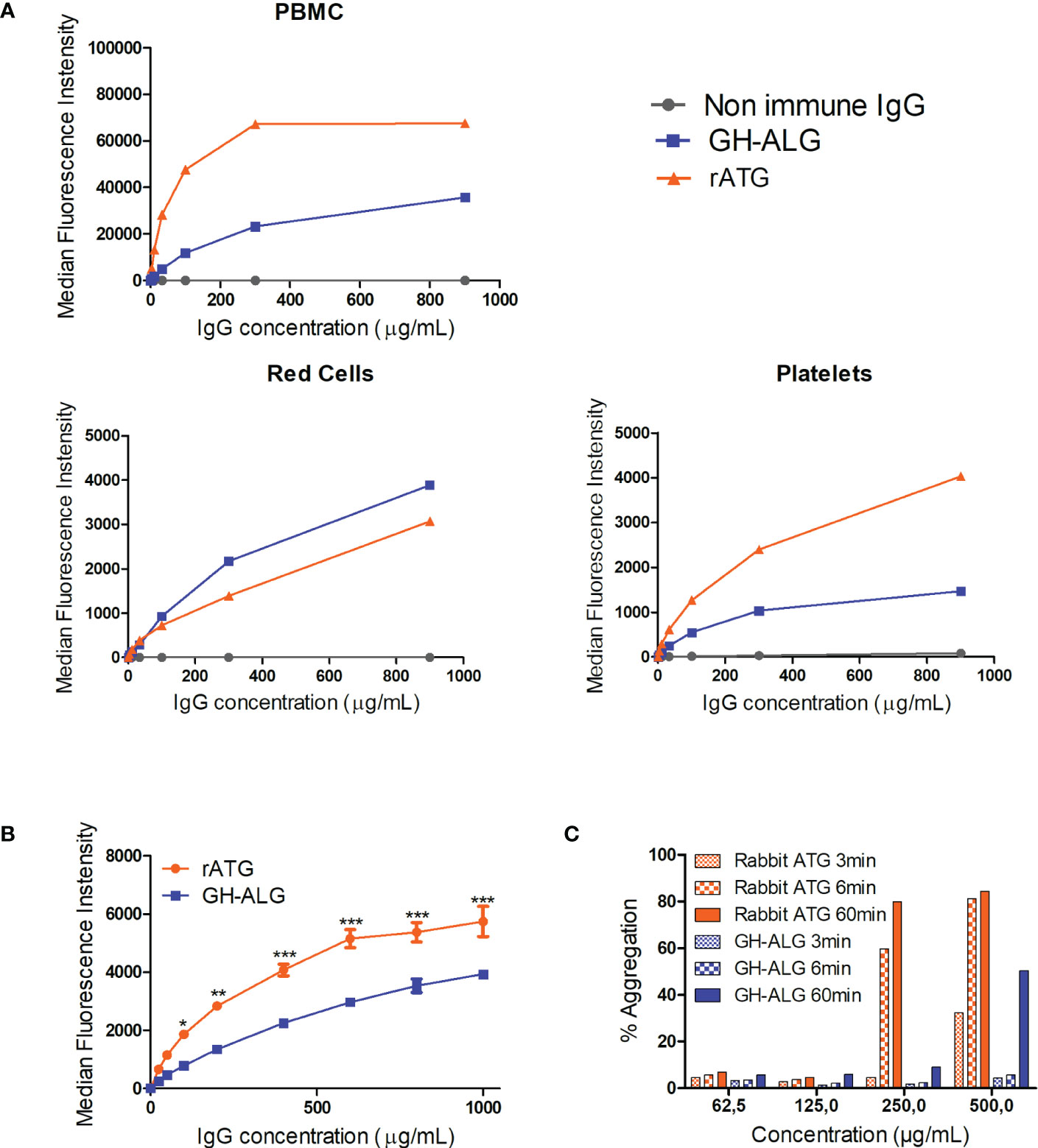
Figure 1 Interaction between GH-ALG and human blood cells. (A) Binding of GH-ALG or rabbit ATG to human PBMCs, red cells and platelets was investigated by flow cytometry. Cells were incubated with increasing concentrations of GH-ALG or rabbit ATG. Nonimmune IgG was used as a control. After washing, bound IgG was detected by flow cytometry using a FITC-conjugated anti-pig antibody or FITC-conjugated anti-rabbit antibody (N=3). (B) Direct comparison of GH-ALG and rabbit ATG binding to human platelets. Human platelets were incubated with increasing doses of GH-ALG or rabbit ATG. Attached antibodies were then detected using AF488-conjugated protein G and flow cytometry analysis. The data are expressed as means ± SEM (N=3). Two-way ANOVA (*, p<0.5; **, p<0.01; ***, p<0.005). (C) Platelet aggregation assays. Human platelets were purified from citrated blood, and aggregation was measured by turbidimetry using a TA-8 V platelet aggregometer (Stago). One representative experiment from two is shown. .
Antigenic targets of GH-ALG were determined using a competition assay against labeled-monoclonal antibodies targeting the classical T-cell markers—i.e., anti-TCR α/β, anti-CD2, anti-CD3, anti-CD4, anti-CD8 and anti-CD28 antibodies. In the presence of GH-ALG, we observed a strong decrease in the MFI for all antibodies evaluated (Table 1), indicating competition between mAbs and GH-ALG on these targets. The inhibition of the monoclonal signal ranges from 94% for CD8, 52% for CD4 to 23% inhibition for CD2.
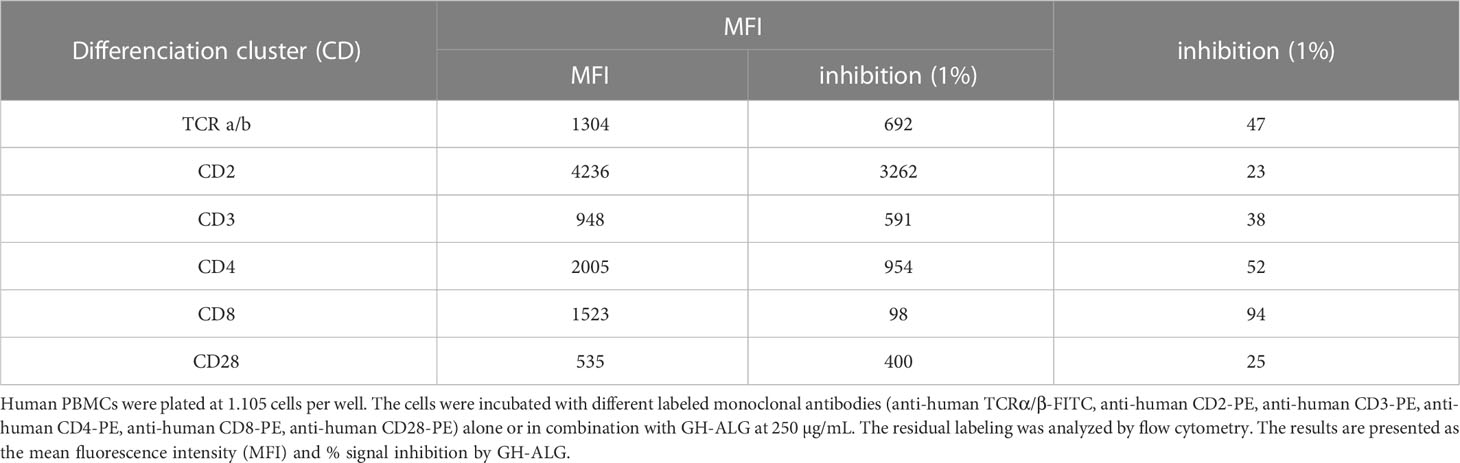
Table 1 Antigenic targets.
We then determined the fraction of GH-ALG able to bind target cells. Specific antibodies within GH-ALG polyclonal IgG were removed by successive adsorption on human PBMCs (Supplemental Figure 2). The amount of nonspecific GH-ALG antibodies remaining after 5 cycles of adsorption suggested that 40% of GH-ALG IgG was specific to human PBMCs.
ALG/ATG activity classically relies on lymphocyte depletion through complementary mechanisms such as CDC, ADCC, ADCP and apoptosis induction. We have previously shown that GH-ALG is particularly efficient in recruiting human C1q and mediating CDC. By contrast, GH-ALG could not interact with human Fcγ receptors and thus could not trigger ADCC in humans (40). Here, we investigated other mechanisms of lymphocyte depletion using rabbit ATG as a reference to assess GH-ALG activity.
Induction of apoptosis in human lymphocytes was investigated after exposure to increasing doses of GH-ALG. Nonimmune DKO pig IgG was used as a negative control. Induction of apoptosis, including early and late apoptosis, was monitored by flow cytometry using Annexin V and DAPI staining (Figure 2A). Significant apoptosis induction was detectable using 10 µg/ml of GH-ALG (Figure 2B) and increased with GH-ALG concentration, reaching a plateau at 300 µg/ml.
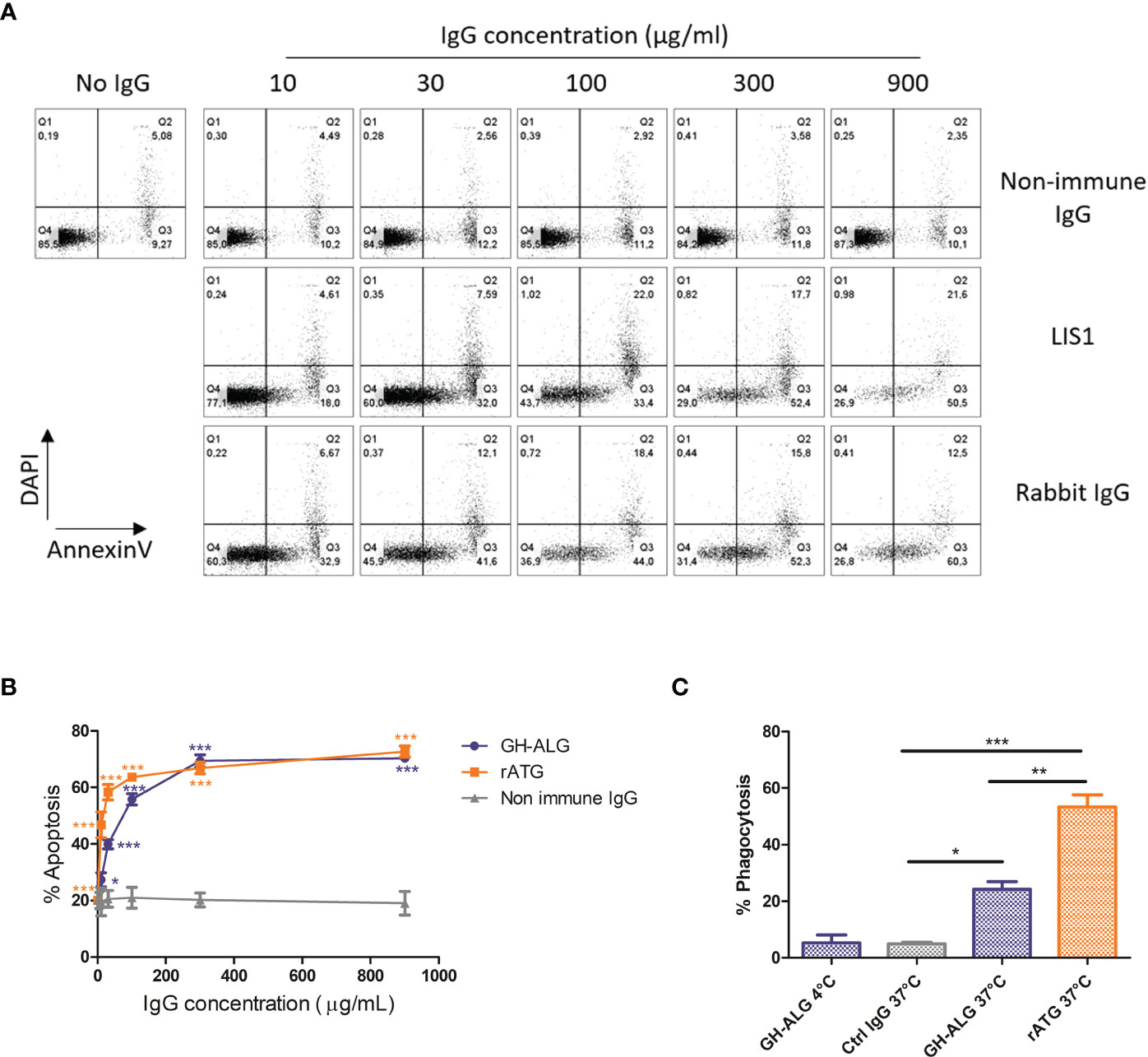
Figure 2 Mechanisms of action of GH-ALG. Induction of apoptosis by GH-ALG. (A, B) Human PBMCs were treated with increasing doses (10, 30, 100, 300 and 900 µg/ml) of GH-ALG. Rabbit ATG and nonimmune DKO pig IgG were used as positive and negative controls, respectively. After 3 h of culture (37°C, 5% CO2), apoptosis was monitored by flow cytometry by Annexin V and DAPI staining. (B) The mean percentage of apoptosis was then determined by adding the percentage of cells in early (Annexin V+/DAPI-) and late (Annexin V+/DAPI+) apoptosis. The data are expressed as means ± SEM (N=3). Two-way ANOVA (*, p<0.5; **, p<0.01; ***, p<0.005), comparison of GH-ALG or rabbit ATG vs. nonimmune IgG. Phagocytosis assay. (C) Phagocytosis of opsonized PBMCs by human monocyte-derived macrophages was assessed by flow cytometry. CFSE-labeled cells were incubated with IgG in RPMI 10% FCS medium for 30 min at 4°C. Lymphocytes were washed twice and cultured with human macrophages (ratio 1:1) in RPMI 10% FCS. After 3 hours of culture, the cells were washed twice, and macrophages were labeled with CD14-BV421 for 30 min at 4°C. Cells were washed twice before flow cytometry analysis. Phagocytosis was assessed as the percentage of double‐positive (CFSE+/CD14+) cells. The values were compared by ANOVA followed by Tukey’s post hoc test. (*, p<0.5; **, p<0.01; ***, p<0.001).
A significant uptake of T lymphocytes by monocyte-derived macrophages was observed after opsonization with GH-ALG or rabbit ATG (Figure 2C), with 23% or 53% of macrophages ingesting T cells, respectively. The absence of internalization when cultures were performed at 4°C demonstrated the active process involved. The active GH-ALG concentration was 10 µg/mL, a concentration with limited apoptosis. Therefore, the data presented reflected ADCP activity mediated by opsonization of targets and not phagocytosis of apoptotic cells.
We evaluated the ability of GH-ALG to block T-cell alloreactivity. One-way MLRs were performed in the presence of GH-ALG or rabbit ATG, and T-cell proliferation was monitored after 3 days. We observed a strong and dose-dependent inhibition of residual live T-cell proliferation when MLR was performed in the presence of GH-ALG (Figure 3). The inhibition reach 98% at a concentration of 500 µg/ml and remains significant up to 86ug/mL.
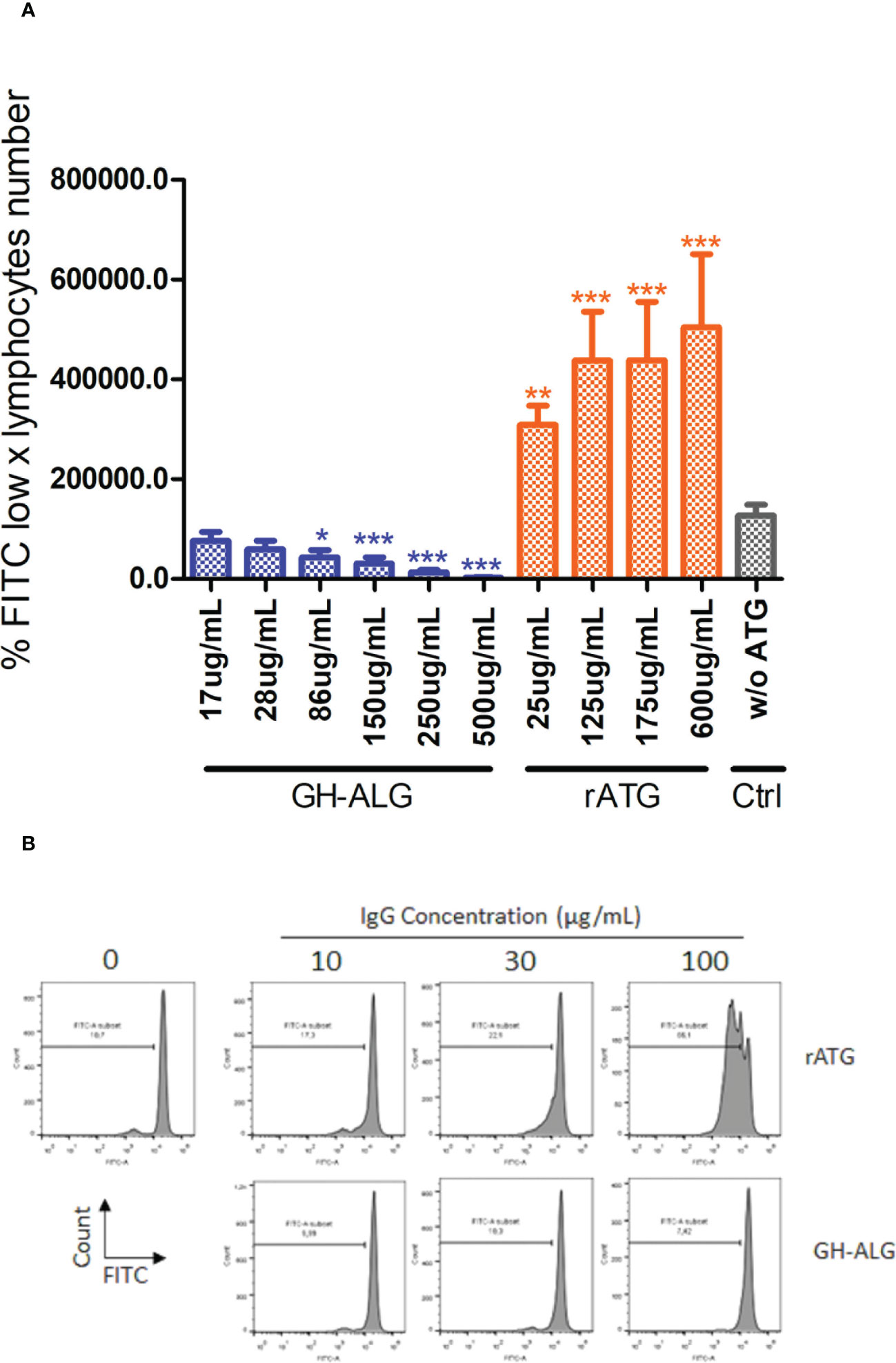
Figure 3 Mixed Lymphocyte Reaction assay. (A) Stimulator human PBMCs were irradiated with 35 Gy; responder PBMCs were labeled with CFSE. Stimulator cells were adjusted to 8.106 cells/mL, and responder cells were adjusted to 2 × 106 cells/ml of RPMI medium. Next, the PBMCs were cocultured in a total volume of 200 µl of RPMI medium at 37 °C in a 5% CO2 incubator in the dark for 3 days with or without 250 µg/mL of rabbit ATG or GH-ALG. Proliferation was evaluated by measuring CFSE incorporation using flow cytometry. Groups were compared using ANOVA and the Tukey‒Kramer multiple comparison test. (B) Representative flow cytometry histogram demonstrating serial dilution of FITC. * between 0.01 and 0.05. ** between 0.001 and 0.01. *** between 0 and 0.001.
To determine the ability of GH-ALG to blunt alloreactivity in vivo, allogeneic skin grafts were performed in monkeys (Figure 4A). Graft rejection was defined by the presence of a scabby aspect, brown coloring with loss of flexibility, or complete necrosis of the epidermis. Rejection of allogeneic skin grafts occurred from day 8 to 11 in the control group (untreated transplanted monkeys). Graft rejection was significantly delayed following treatment with GH-ALG (Figure 4B).
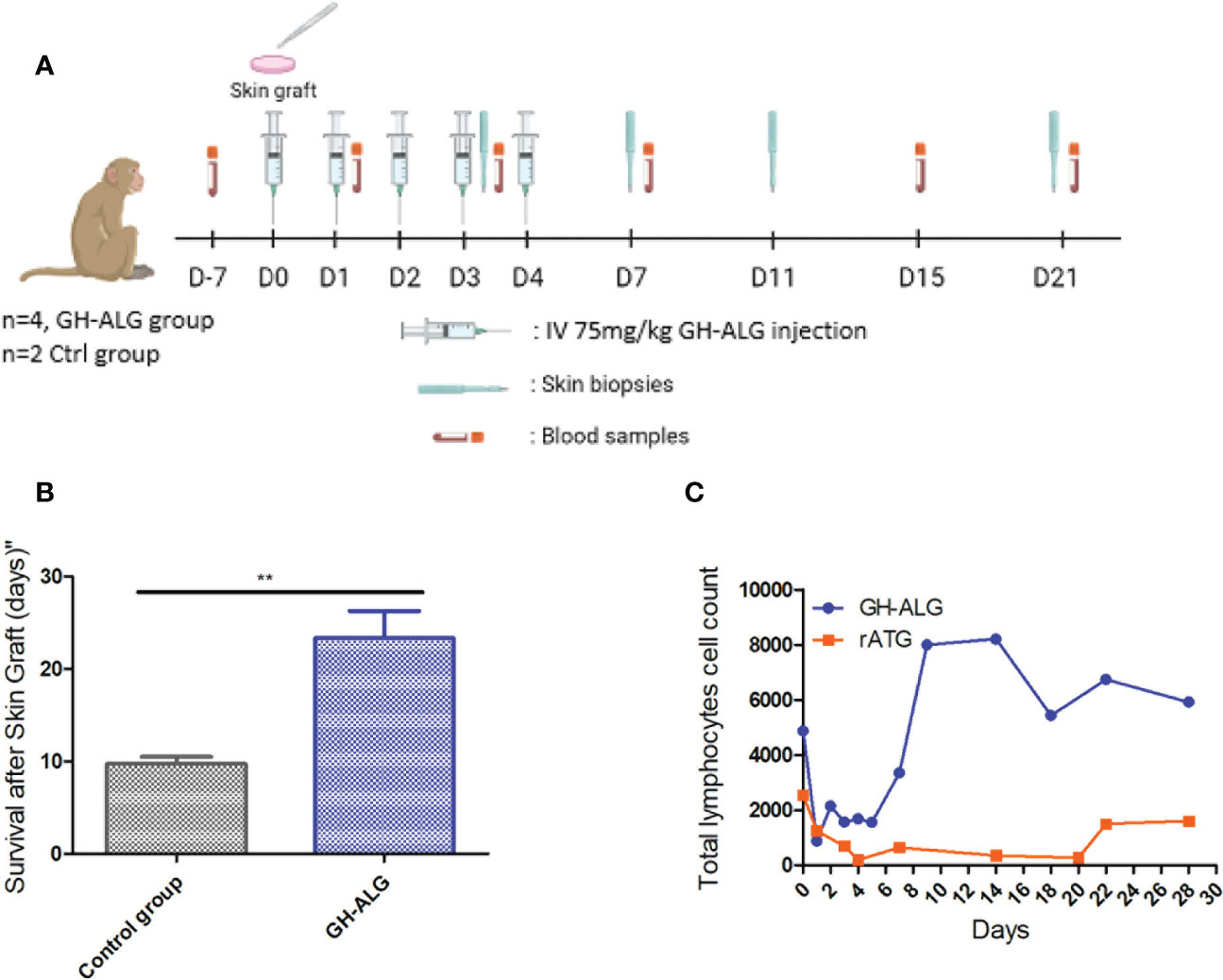
Figure 4 In vivo efficacy of anti-lymphocyte IgG in NHP. (A) Timeline of the NHP skin graft study. (B) Skin grafts were performed on anesthetized animals on Day 0. After back skin shearing and asepsis, the animals were grafted in pairs (back skin collection using a 20 mm diameter template, followed by grafting and skin graft suturing). In the back of each animal, three grafts were performed: one autograft (as a control) and two pairwise allografts (one animal received skin transplants of his congener). Four cynomolgus monkeys intravenously received GH-ALG at 75 mg/kg once a day for 5 consecutive days. Four cynomolgus monkeys were used as controls and did not receive treatment. Skin graft rejection was considered when the area of the necrotized part covered the full graft, and the whole graft site hardened. (C) GH-ALG and rabbit ATG-induced T-cell depletion in macaques. Cynomolgus monkeys (n=9) received I.V. daily doses of 50 mg/kg of GH-ALG for 5 days. Cynomolgus monkeys (n=2) received I.V. daily doses of 5 mg/kg of rabbit ATG for 5 days. ** between 0.001 and 0.01.
Depletion experiments were also performed in vivo in cynomolgus monkeys. A dramatic drop in the CD3 lymphocyte count was observed after the first injection of GH-ALG or rabbit ATG, from 4.8 giga lymphocytes/L to 0.8 for GH-ALG and from 2.5 to 0.2 for rabbit ATG. (Figure 4C). Lymphocyte depletion induced by GH-ALG was limited to one week, the rise above 1giga lymphocytes/L is observed from day 2 post injection.
We investigated the depletion of lymphocyte subpopulations after five I.V. infusions of GH-ALG (50 mg/kg/adm) in cynomolgus monkeys. GH-ALG induced selective depletion of peripheral effector naive T cells (CD4+ TN p=0.005; CD8+ TN, p=0.0002), memory effector T cells (CD4+ EM p=0.0134; CD8+ EM p=0.002) and recent thymic embryonic cells (CD4+ RTE, p=0.007; CD8+ RTE, p<0.007) between Day 0 and Day 3/4 (Figure 5). Depletion was stronger for CD8 cells (86% drop for naive CD8 cells and 95% for memory effectors) than CD4 cells (71% decrease for naive CD4 cells and 43% for memory effectors). By contrast, B cells and Tregs were unaffected by GH-ALG (Figure 5).
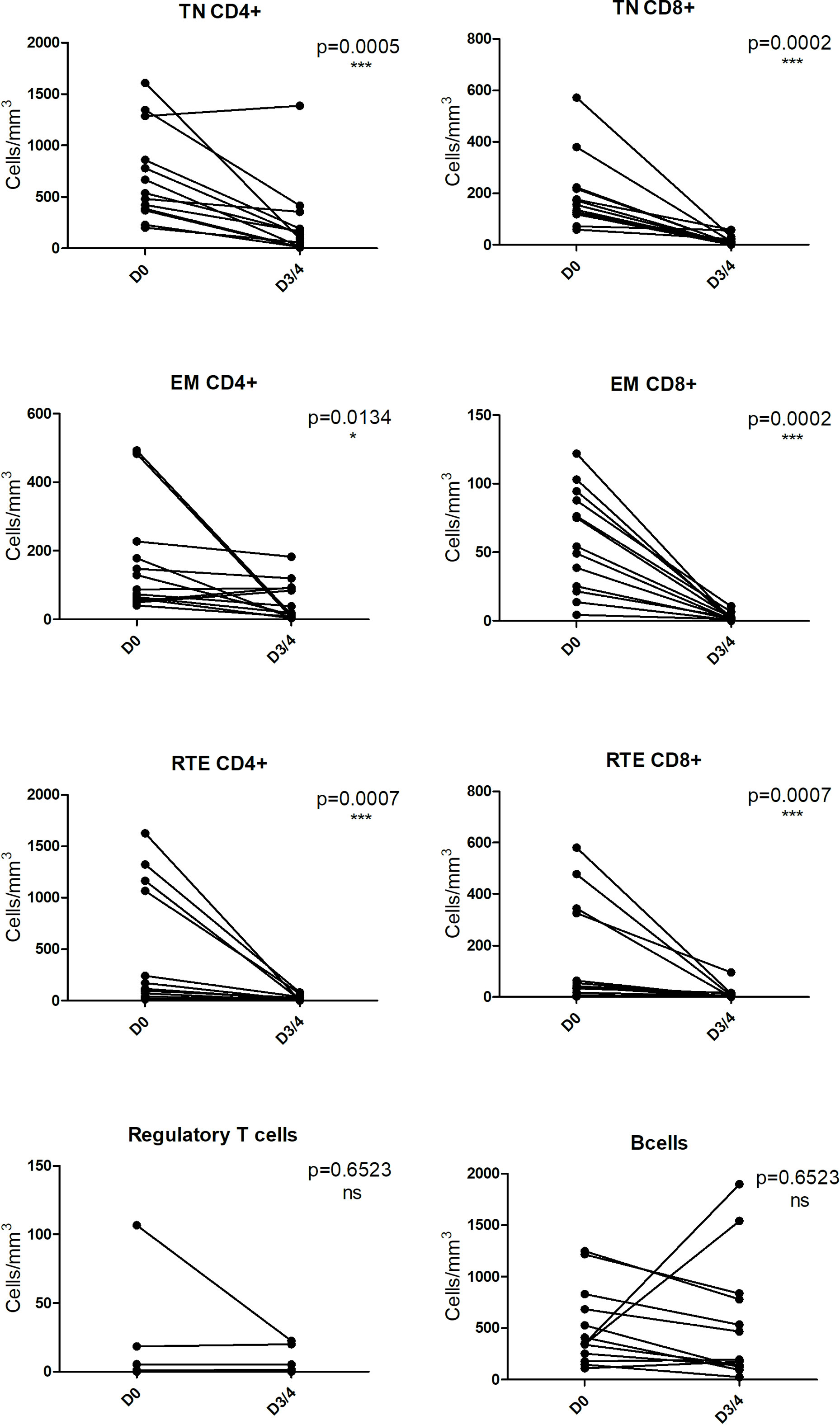
Figure 5 GH-ALG shows selective immunoreactivity to peripheral T cells. Cynomolgus monkeys (n=11) received I.V. daily doses of 50 mg/kg GH-ALG for 5 days. On Days 3-4, blood was sampled, and lymphocyte subpopulations were analyzed by flow cytometry. Whereas CD4+ and CD8+ T-cell subpopulations were reduced, Treg and B cells were unaffected. Monocytes were also unaffected (not shown). * between 0.01 and 0.05. *** between 0 and 0.001.
Serum samples were drawn and analyzed by ELISA. GH-ALG presented a Half-life of approx. 40 h, and a distribution profile compatible with the plasmatic compartment in primates (Figure 6). The mean systemic clearance was 0.044 and 0.048 mL/h/kg in females and males, respectively. Quantifiable serum concentrations were observed until the last collected time point (29 days after the last infusion on Day 5), reflecting a slow elimination rate of GH-ALG. No marked difference was observed between males and females, as the ratio of AUC0-t values was close to 1.
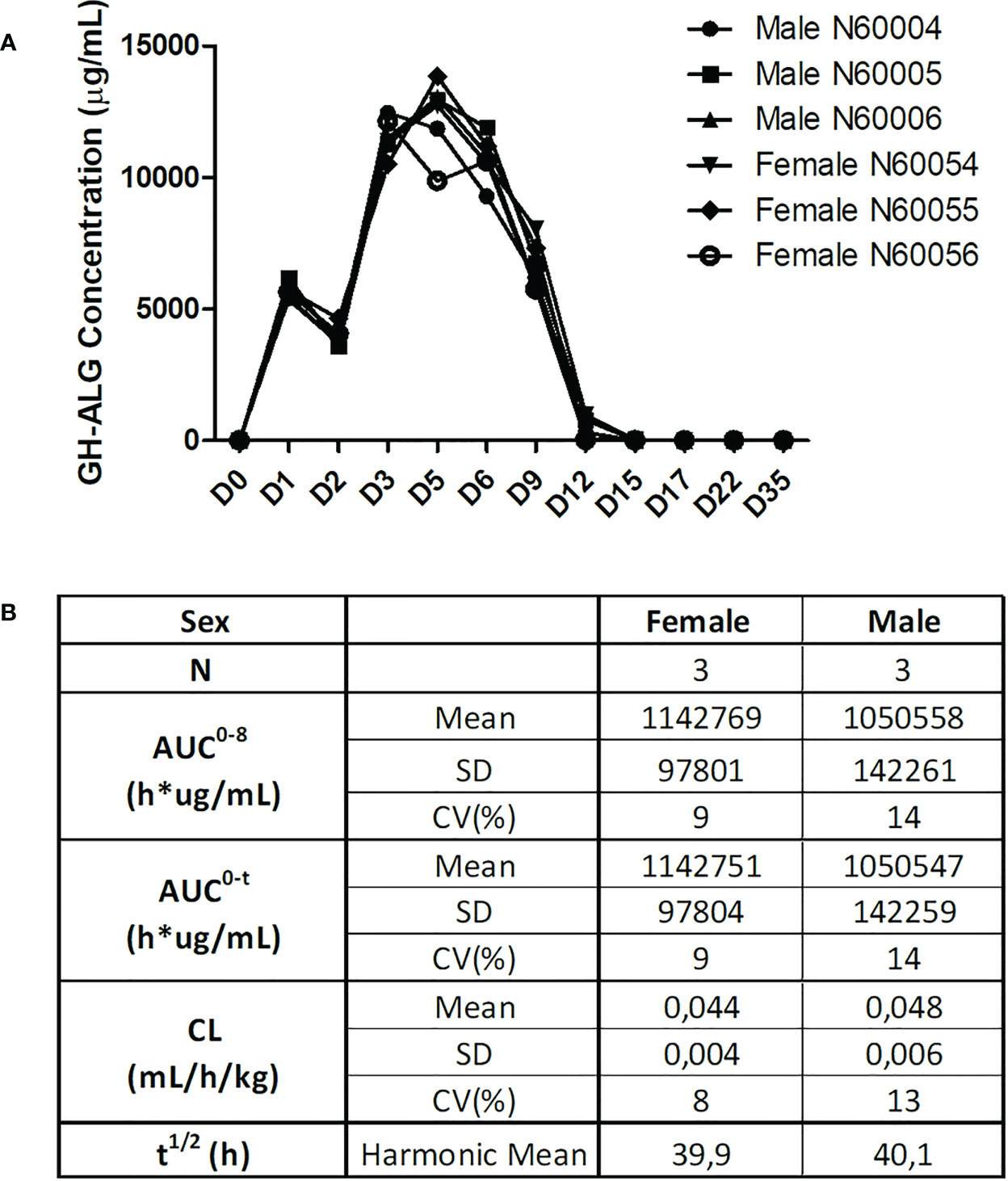
Figure 6 Pharmacokinetic evaluation. For the GH-ALG GLP toxicology study, one group of three male and three female cynomolgus monkeys received GH-ALG by daily intravenous administration (4-hour infusion) at 50 mg/kg/adm for 5 days. Serum samples were drawn on Days 1 (before infusion, 5 minutes, 24 and 96 hours after infusion) and 5 (before infusion and 5 minutes after infusion) and Days 6, 9, 12, 15, 17, and 22 and week 5. Pharmacokinetic parameters were analyzed by ELISA. (A) Pharmacokinetic profiles of GH-ALG-treated macaques. (B) Pharmacokinetic characteristics.
GH-ALG is a novel ALS obtained from GGTA1 and CMAH double KO pigs hyperimmunized with a double-positive human CD4+/CD8+ T-cell line. GH-ALG differs from other ALSs by its mechanism of action, which relies on apoptosis, CDC and inhibition of alloreactivity without ADCC.
The goal of induction therapy is to control early acute rejection following organ transplantation (41, 42). After infusion, rabbit ATG induces immediate immune cell depletion, particularly T lymphocyte depletion, through ADCC, CDC, opsonization resulting in phagocytosis of IgG-coated T cells apoptosis by activation of the Fas-Fas ligand pathway (35, 36, 43). Although these mechanisms allow marked depletion of target cells, they come at the cost of leukopenia and thrombocytopenia. Moreover, in some patients, CD4+ lymphopenia lasts for several months, increasing the risk of infection and other complications41,42.
One main challenge in organ transplantation is to define the best immunosuppressive strategy to ensure long-term graft and patient survival by preventing graft rejection and protecting against treatment side effects (2, 44, 45). Thrombocytopenia is a frequent complication, partly induced by immunosuppressive therapies. Thrombocytopenia is more common in patients receiving ATGs than in those receiving basiliximab in the first few days after transplantation (2, 45). Although thrombocytopenia is often mild and transient, for kidney transplantation, it may exacerbate the perioperative coagulation imbalance in patients with already compromised bleeding control. Because of differences in the source of cells used for immunization, GH-ALG and other ATGs have different antigenic profiles, and these differences may lead to clinical consequences (46), particularly for thrombocytopenia. We illustrate here that the binding and platelet aggregation of GH-ALG is very low, which might result in lower thrombocytopenia.
The mode of action of GH-ALG includes transient immunosuppression by inducing CDC, ADCP and apoptosis-mediated selective T-cell depletion. Using in vitro assays and in vivo pharmacodynamic and transplantation models in nonhuman primates (NHPs) or humanized RRGS rats (47), GH-ALG confirmed this mode of action and nevertheless demonstrated a strong impact on T-cell responses in a stringent skin graft rejection model.
Conventionally, ADCP is described as a mechanism by which antibody-opsonized target cells activate FcγRs on the surface of macrophages to induce phagocytosis (48). Thus, the mechanism underlying GH-ALG-generated ADCP is currently unknown because no interaction exists between porcine IgG and human FcγRs, except for FcRn (40). The active GH-ALG concentration was 10 µg/mL, a concentration with limited apoptosis. Therefore, the data presented reflected ADCP activity mediated by opsonization of targets and not phagocytosis of apoptotic cells. Studies have shown that FcRn expression on phagocytic cells (49, 50) increases the phagocytosis capacity of opsonized IgG particles (51, 52). This mechanism of phagocytosis (52, 53) would likely require external sensing and cellular activation via FcγR or pattern recognition receptors (C-type lectin receptors) (54). Phagocytosis by neutrophils and monocytes mediated by the interaction between C3b fragment from the cleavage of complement protein C3 and the CR1 receptor (CD35) is also a possible mechanism, although not explored here.
Additionally, in vitro, we showed that in mixed lymphocyte reaction (MLR) experiments, LIS1 demonstrated a potent immunosuppressive activity, a feature not found with a preparation of rabbit ATG assessed in parallel. It was well described for rabbit ATG (Thymoglobulin®) that at the concentration of 0.1 mg/ml onwards, it activated T cells in vitro (both CD4 and CD8 subpopulations) with the synthesis of IL-2, IFN-γ, and CD25 expression, inducing subsequent T cell proliferation (9). Presumably, LIS1 blocks T-cell receptors important for alloreactivity. This feature might be due to the incompatibility between pig IgG Fc domains with human Fc receptors, reducing FcR-mediated cross-linking and agonist events. This mechanism is reminiscent of the mechanism of another class of induction therapy, the anti-CD25 monoclonal antibodies (e.g., basiliximab), which completely and consistently block activated T cells and their proliferation via blocking the interleukin-2 receptor.
Rabbit ATG is obtained from rabbits immunized with a preparation of human thymocytes containing mature and immature T cells, Treg cells, progenitor cells, dendritic cells, epithelial cells and endothelial cells. It thus potentially targets a wide range of cells other than T lymphocytes. In a humanized mouse model, rabbit ATG has been shown to bind almost all human hematopoietic cell lineages, including hematopoietic stem cells, and thus likely depletes all subpopulations (55). The prolonged lymphodepletion observed in macaques treated with rabbit ATG (56), might originate from this broad impact of ATG on T cells and stem cells, also impacting T-cell reconstitution. Because GH-ALG is obtained by immunization with a selected effector-type human T-lymphocyte cell line, it shows selective immunoreactivity to peripheral effector T cells but not to B cells, Treg, recent thymic emigrant cells and myeloid-derived suppressor cells described in experimental models and humans to contribute to immunosuppression maintenance post kidney transplantation (57, 58). In vivo evaluation in nonhuman primates showed that GH-ALG robustly depleted target T cells from the peripheral blood. Interestingly, depletion was intense but short-term because T-cell counts recovered within two weeks. In a previous study in RRGS humanized rats reconstituted with human PBMCs (47), GH-ALG blocked human T-cell reconstitution by more than 98% and prevented graft versus host disease (GVHD).
Although potent T-cell depletion has been historically considered necessary to prevent acute rejection during the early posttransplant phase for patients with high immunological risk, the prolonged T-cell depletion (35) (59) is associated with an increased risk of infection, including bacterial infections (urinary tract and wound infections) and less commonly viral infections, such as cytomegalovirus (CMV), Epstein−Barr virus (EBV) and BK polyomavirus (BKV) infections (60). Studies have also reported that CD4+ T-cell lymphopenia is associated cardiovascular complications and mortality (61). An increased risk of infections is associated with the increased risk of posttransplant morbidity, rehospitalization and overall mortality (62, 63) and with decreased graft survival due to the infection itself and to the reduction in immunosuppressive therapy needed to manage infections (64). Our preclinical data show that after 5 consecutive days of GH-ALG infusion, T-cell depletion occurs acutely over 5 days and returns to baseline levels at the latest within 3 weeks (see Figure 4). If confirmed in human, these data suggest that GH-ALG may reduce the risk of infection associated with prolonged depletion observed with other ALS therapies. This regimen would for example benefit patients with a higher risk of infection, among which elderly kidney graft recipients (64). Avoiding prolonged T-cell depletion may also reduce the risk of neoplasm in patients, a possible additional benefit from induction with GH-ALG (65).
GH-ALG presented shorter and milder depleting activity than rabbit ATG and exhibited direct T-cell blockade activity in a mixed lymphocyte reaction. Nevertheless, in vivo efficacy data in a stringent skin graft model in cynomolgus monkeys showed an equivalent capacity of GH-ALG to prevent allograft rejection compared to published Thymoglobulin® data (56), 9 to 20-25 days with Thymoglobulin® and 9 to 20-29 days with GH-ALG. Whether the net effect of these differences allows similar protection against allograft rejection still remains to be proven in future clinical studies.
Breg cells prevent graft rejection and belong to the signature of tolerance (66–68). Our study shows that GH-ALG spares B cells, thus avoiding depletion of B-regulators, a major difference from rabbit ATG, which induces apoptosis of all B-cell subsets (6). Similarly, regulatory T cells (Tregs) play a crucial role in limiting renal transplant rejection and, potentially, in promoting transplant tolerance (69). Rabbit ATG induces and expands CD4+CD25+Foxp3+ cells with regulatory function ex vivo (70). We observed that Tregs were not affected by GH-ALG in nonhuman primates, at least in the short term. Whether these differences in the modulation of regulatory B and T cells may impact the outcomes of induction treatment is unknown.
GH-ALG is currently undergoing clinical evaluation in humans in de novo kidney transplant recipients (ClinicalTrials.gov Identifier: NCT04431219). This trial will investigate whether the short-term depletion observed in primates replicates in humans and allows safe induction treatment. The hypothesis of an improved posttransplant outcome due to faster T-cell reconstitution and avoidance of infections, while similarly controlling acute rejection needs to be assessed in larger cohorts.
Our paper suffers from limitations related to the complexity of the mode of action of polyclonal products and of the induction procedure in solid organ transplantation. Ideal duration of lymphodepletion post-induction is unclear, and it is unknown whether the short-term T cell depletion reported with GH-ALG plus the GH-ALG-induced blockade of alloreactivity will be sufficient to blunt acute rejection to the same extent as long-term depleting ATG. Similarly, the second role of induction would be to delay calcineurin inhibitors because of their nephrotoxicity, a shorter depletion may also not be suitable. Although absence of B cell depletion by GH-ALG might preserve Breg cells important in tolerance induction, it is unknown whether this presents advantage or drawback, owing to the possible lower control of anti-donor antibodies. The potentially different active titers of specific antibodies make elusive quantitative comparison of GH-ALG and rabbit ATG. However, our data suggest that GH-ALG may gather a specific pattern of properties that could increase the clinical advantage/disadvantage ratio of polyclonal anti human T cell preparations and widen their clinical usage. We have nevertheless tried to carry out as much as possible our in vitro experiments with an equivalent titer. Finally, as many data already exist in the literature for the rabbit-ATG skin grafting model, our grafting model proposes, for ethical reasons, a comparison with control animals and not with animals treated with rabbit-ATG. Our conclusions remain related to the differences in experimental conditions between the studies. As another limitation, we can add that the macaques used in our preclinical model are not CMAH deficient like humans (they are part of the old world monkey classification and have not had a mutation of their CMAH gene) (71), and therefore make Neu5Gc that can be found metabolically on the surface of their cells, but also develop a natural anti-Neu5Ac immunity. As GH-ALG is produced in a DKO CMAH-/GGTA1- pig model, the glycosylations carried by the IgGs are predominantly Neu5Ac. It has been shown (72, 73) that this difference in glycosylation could influence the recognition of PBMCs, which could be a bias to the analysis of our results on lymphocyte populations in the NHP model.
In summary, GH-ALG is a novel induction agent with potential interest for transplanted patients, overcoming problems related to xeno-antigenicity and prolonged target T-cell depletion. Its mechanism of action, combining mild T-cell depletion with inhibition of alloreactivity, warrants its clinical evaluation in kidney graft recipients.
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
The animal study was reviewed and approved by the French Research Ministry.
Conceived the study: J-MB, SB, OD, J-PS, and BV; designed and supervised some experiments: OD, J-PS, and BV; performed the experiments: CC, GE, JR, P-JR, and AS. All authors contributed to the article and approved the submitted version.
The authors thank Marc Fouassier (CHU Nantes) for his help in the platelet aggregation assay.
The authors of this manuscript have conflicts of interest to disclose: JR, P-JR, CC, GE, EL, FS, and BV are employees of Xenothera, a company developing glycol-humanized polyclonal antibodies as those described in this manuscript, and OD, J-PS, J-MB, and CG are cofounders of Xenothera.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2023.1137629/full#supplementary-material
| Supplemental Table 1 Overview of the in vivo analysis of cynomolgus monkeys. Cynomolgus monkeys (n=16) received I.V. daily doses ranging from 40 to 75 mg/kg GH-ALG for 5 days. The characteristics of these in vivo studies are described here.
| Supplemental Figure 1 Comparison of pig and rabbit IgG detection by G protein: (A) ELISA: GH-ALG and rabbit-ATG are coated on a maxisorp plate at 50ug/mL. After saturation and washing, revelation is performed with Alexa Fluor 488 conjugated protein G diluted at 1/200 or 1/400 and the reading performed in fluorescence with TECAN. (B): Analysis of binding kinetics by BLItz (bio-layer interferometry) of GH-ALG and rabbit-ATG on G protein.
| Supplemental Figure 2 Evaluation of the GH-ALG active fraction. One milliliter of GH-ALG diluted in PBS to a concentration of 1.6 mg/mL was incubated with 30.106 target lymphocyte cells. After 20 min of incubation at room temperature, the supernatant (SN1) was transferred to a second tube containing 30.106 target T lymphocytes. After 20 min of incubation at room temperature, the supernatant (SN2) was collected and stored until SN5. The supernatants obtained were serially diluted and then incubated (30 min, 4°C) with fresh XT1501 cells. After washing, a secondary anti-pig antibody was deposited, revealing the remaining XT1501-specific antibody fraction. GH-ALG was used as a positive specificity control. Nonimmune IgGs were used as a negative control.
1. Gaber AO, First MR, Tesi RJ, Gaston RS, Mendez R, Mulloy LL, et al. Results of the double-blind, randomized, multicenter, phase III clinical trial of thymoglobulin versus atgam in the treatment of acute graft rejection episodes after renal transplantation. Transplantation (1998) 66(1):29−37. doi: 10.1097/00007890-199807150-00005
2. Brennan DC, Daller JA, Lake KD, Cibrik D, Del Castillo D. Thymoglobulin induction study group. rabbit antithymocyte globulin versus basiliximab in renal transplantation. N Engl J Med (2006) 355(19):1967−77. doi: 10.1056/NEJMoa060068
3. Kapic E, Becic F, Kusturica J. Basiliximab, mechanism of action and pharmacological properties. Med Arh (2004) 58(6):373−6.
4. Woodruff MF. Purification of antilymphocytic antibody. Nature (1968) 217(5131):821−4. doi: 10.1038/217821a0
5. Gaber AO, Knight RJ, Patel S, Gaber LW. A review of the evidence for use of thymoglobulin induction in renal transplantation. Transplant Proc (2010) 42(5):1395−400. doi: 10.1016/j.transproceed.2010.04.019
6. Mohty M. Mechanisms of action of antithymocyte globulin: T-cell depletion and beyond. Leukemia (2007) 21(7):1387−94. doi: 10.1038/sj.leu.2404683
7. Thiyagarajan UM, Ponnuswamy A, Bagul A. Thymoglobulin and its use in renal transplantation: A review. Am J Nephrol (2013) 37(6):586−601. doi: 10.1159/000351643
8. Gharekhani A, Entezari-Maleki T, Dashti-Khavidaki S, Khalili H. A review on comparing two commonly used rabbit anti-thymocyte globulins as induction therapy in solid organ transplantation. Expert Opin Biol Ther (2013) 13(9):1299−313. doi: 10.1517/14712598.2013.822064
9. Bonnefoy-Bérard N, Vincent C, Revillard JP. Antibodies against functional leukocyte surface molecules in polyclonal antilymphocyte and antithymocyte globulins. Transplantation (1991) 51(3):669−73. doi: 10.1097/00007890-199103000-00024
10. Deeks ED, Keating GM. Rabbit antithymocyte globulin (thymoglobulin): A review of its use in the prevention and treatment of acute renal allograft rejection. Drugs (2009) 69(11):1483−512. doi: 10.2165/00003495-200969110-00007
11. Couvrat-Desvergnes G, Salama A, Le Berre L, Evanno G, Viklicky O, Hruba P, et al. Rabbit antithymocyte globulin–induced serum sickness disease and human kidney graft survival. J Clin Invest (2015) 125(12):4655−65. doi: 10.1172/JCI82267
12. Tanriover B, Chuang P, Fishbach B, Helderman JH, Kizilisik T, Nylander W, et al. Polyclonal antibody-induced serum sickness in renal transplant recipients: Treatment with therapeutic plasma exchange. Transplantation (2005) 80(2):279−81. doi: 10.1097/01.TP.0000165093.13046.B3
13. Brabant S, Facon A, Provôt F, Labalette M, Wallaert B, Chenivesse C. An avoidable cause of thymoglobulin anaphylaxis. Allergy Asthma Clin Immunol Off J Can Soc Allergy Clin Immunol (2017) 13:13. doi: 10.1186/s13223-017-0186-9
14. Sebeo J, Ezziddin O, Eisenkraft JB. Severe anaphylactoid reaction to thymoglobulin in a pediatric renal transplant recipient. J Clin Anesth (2012) 24(8):659−63. doi: 10.1016/j.jclinane.2012.04.014
15. Kandil E, Alabbas H, Distant D. Anaphylaxis to thymoglobulin: A case report and literature review. J La State Med Soc Off Organ La State Med Soc (2009) 161(5):279−81.
16. Watier H. De la sérothérapie aux anticorps recombinants « nus » - plus d’un siècle de succès en thérapie ciblée. médecine/sciences. (2009) 25(12):999−1009. doi: 10.1051/medsci/20092512999
17. Pham T, Gregg CJ, Karp F, Chow R, Padler-Karavani V, Cao H, et al. Evidence for a novel human-specific xeno-auto-antibody response against vascular endothelium. Blood (2009) 114(25):5225−35. doi: 10.1182/blood-2009-05-220400
18. Varki NM, Strobert E, Dick EJ, Benirschke K, Varki A. Biomedical differences between human and nonhuman hominids: Potential roles for uniquely human aspects of sialic acid biology. Annu Rev Pathol (2011) 6:365−93. doi: 10.1146/annurev-pathol-011110-130315
19. Pirofsky B, Ramirez-Mateos JC, August A. Foreign serum heterophile antibodies in patients receiving antithymocyte antisera. Blood (1973) 42(3):385−93. doi: 10.1182/blood.V42.3.385.385
20. Merrick JM, Zadarlik K, Milgrom F. Characterization of the hanganutziu-deicher (serum-sickness) antigen as gangliosides containing n-glycolylneuraminic acid. Int Arch Allergy Appl Immunol (1978) 57(5):477−80. doi: 10.1159/000232140
21. Higashi H, Naiki M, Matuo S, Ōkouchi K. Antigen of “serum sickness” type of heterophile antibodies in human sera: Identification as gangliosides with n-glycolylneuraminic acid. Biochem Biophys Res Commun (1977) 79(2):388−95. doi: 10.1016/0006-291x(77)90169-3
22. Irie A, Koyama S, Kozutsumi Y, Kawasaki T, Suzuki A. The molecular basis for the absence of n-glycolylneuraminic acid in humans. J Biol Chem (1998) 273(25):15866−71. doi: 10.1074/jbc.273.25.15866
23. Salama A, Evanno G, Harb J, Soulillou JP. Potential deleterious role of anti-Neu5Gc antibodies in xenotransplantation. Xenotransplantation (2015) 22(2):85−94. doi: 10.1111/xen.12142
24. Rousse J, Salama A, Leviatan Ben-Arye S, Hruba P, Slatinska J, Evanno G, et al. Quantitative and qualitative changes in anti-Neu5Gc antibody response following rabbit anti-thymocyte IgG induction in kidney allograft recipients. Eur J Clin Invest. avr (2019) 49(4):e13069. doi: 10.1111/eci.13069
25. Salama A, Evanno G, Lim N, Rousse J, Le Berre L, Nicot A, et al. Anti-gal and anti-Neu5Gc responses in nonimmunosuppressed patients after treatment with rabbit antithymocyte polyclonal IgGs. Transplantation (2017) 101(10):2501−7. doi: 10.1097/TP.0000000000001686
26. Taylor RE, Gregg CJ, Padler-Karavani V, Ghaderi D, Yu H, Huang S, et al. Novel mechanism for the generation of human xeno-autoantibodies against the nonhuman sialic acid n-glycolylneuraminic acid. J Exp Med (2010) 207(8):1637−46. doi: 10.1084/jem.20100575
27. Galili U. Evolution and pathophysiology of the human natural anti-alpha-galactosyl IgG (anti-gal) antibody. Springer Semin Immunopathol (1993) 15(2−3):155−71. doi: 10.1007/BF00201098
28. Berg EA, Platts-Mills TAE, Commins SP. Drug allergens and food–the cetuximab and galactose-α-1,3-galactose story. Ann Allergy Asthma Immunol Off Publ Am Coll Allergy Asthma Immunol (2014) 112(2):97−101. doi: 10.1016/j.anai.2013.11.014
29. Zhu A, Hurst R. Anti-n-glycolylneuraminic acid antibodies identified in healthy human serum. Xenotransplantation (2002) 9(6):376−81. doi: 10.1034/j.1399-3089.2002.02138.x
30. Padler-Karavani V, Yu H, Cao H, Chokhawala H, Karp F, Varki N, et al. Diversity in specificity, abundance, and composition of anti-Neu5Gc antibodies in normal humans: Potential implications for disease. Glycobiology (2008) 18(10):818−30. doi: 10.1093/glycob/cwn072
31. Bardor M, Nguyen DH, Diaz S, Varki A. Mechanism of uptake and incorporation of the non-human sialic acid n-glycolylneuraminic acid into human cells. J Biol Chem (2005) 280(6):4228−37. doi: 10.1074/jbc.M412040200
32. Tangvoranuntakul P, Gagneux P, Diaz S, Bardor M, Varki N, Varki A, et al. Human uptake and incorporation of an immunogenic nonhuman dietary sialic acid. Proc Natl Acad Sci U.S.A. (2003) 100(21):12045−50. doi: 10.1073/pnas.2131556100
33. Dhar C, Sasmal A, Varki A. From « serum sickness » to « xenosialitis »: Past, present, and future significance of the non-human sialic acid Neu5Gc. Front Immunol (2019) 10:807. doi: 10.3389/fimmu.2019.00807
34. Soulillou JP, Cozzi E, Bach JM. Challenging the role of diet-induced anti-Neu5Gc antibodies in human pathologies. Front Immunol (2020) 11:834. doi: 10.3389/fimmu.2020.00834
35. Mueller T. Mechanisms of action of thymoglobulin. Transplantation (2007) 84:S5−10. doi: 10.1097/01.tp.0000295420.49063.b1
36. Haudebourg T, Poirier N, Vanhove B. Depleting T-cell subpopulations in organ transplantation. Transpl Int Off J Eur Soc Organ Transplant (2009) 22(5):509−18. doi: 10.1111/j.1432-2277.2008.00788.x
37. Soulillou JP, Cantarovich D, Le Mauff B, Giral M, Robillard N, Hourmant M, et al. Randomized controlled trial of a monoclonal antibody against the interleukin-2 receptor (33B3.1) as compared with rabbit antithymocyte globulin for prophylaxis against rejection of renal allografts. N Engl J Med (1990) 322(17):1175−82. doi: 10.1056/NEJM199004263221702
38. Vanhove B. Anticorps monoclonaux en transplantation. médecine/sciences (2009) 25(12):1121−5. doi: 10.1051/medsci/200925121121
39. Ciancio G, Burke GW III. Alemtuzumab (Campath-1H) in kidney transplantation. Am J Transplant. (2008) 8(1):15−20. doi: 10.1111/j.1600-6143.2007.02053.x
40. Vanhove B, Duvaux O, Rousse J, Royer P, Evanno G, Ciron C, et al. High neutralizing potency of swine glyco-humanized polyclonal antibodies against SARS-CoV-2. Eur J Immunol (2021) 51(6):1412−1422. doi: 10.1002/eji.202049072
41. Hill P, Cross NB, Barnett ANR, Palmer SC, Webster AC. Polyclonal and monoclonal antibodies for induction therapy in kidney transplant recipients. Cochrane Database Syst Rev (2017) 1:CD004759. doi: 10.1002/14651858.CD004759.pub2
42. Bakr MA, Nagib AM, Donia AF. Induction immunosuppressive therapy in kidney transplantation. Exp Clin Transplant Off J Middle East Soc Organ Transplant. mars (2014) 12(Suppl 1):60−9. doi: 10.6002/ect.25liver.l58
43. Genestier L, Fournel S, Flacher M, Assossou O, Revillard JP, Bonnefoy-Berard N. Induction of fas (Apo-1, CD95)-mediated apoptosis of activated lymphocytes by polyclonal antithymocyte globulins. Blood (1998) 91(7):2360−8.
44. Castro MCR, Araujo LMP, Nahas WC, Arap S, David-Neto E, Ianhez LE. Induction versus noninduction therapy in kidney transplantation: Considering different PRA levels and different induction therapies. Transplant Proc (2004) 36(4):874−6. doi: 10.1016/j.transproceed.2004.03.084
45. Yang SL, Wang D, Wu W-Z, Lin W-H, Xu T-Z, Cai J-Q, et al. Comparison of single bolus ATG and basiliximab as induction therapy in presensitized renal allograft recipients receiving tacrolimus-based immunosuppressive regimen. Transpl Immunol (2008) 18(3):281−5. doi: 10.1016/j.trim.2007.08.002
46. Popow I, Leitner J, Grabmeier-Pfistershammer K, Majdic O, Zlabinger GJ, Kundi M, et al. A comprehensive and quantitative analysis of the major specificities in rabbit antithymocyte globulin preparations. Am J Transplant (2013) 13(12):3103−13. doi: 10.1111/ajt.12514
47. Ménoret S, Ouisse LH, Tesson L, Remy S, Usal C, Guiffes A, et al. In vivo analysis of human immune responses in immunodeficient rats. Transplantation (2020) 104(4):715−23. doi: 10.1097/TP.0000000000003047
48. Rosales C, Uribe-Querol E. Phagocytosis: A fundamental process in immunity. BioMed Res Int (2017) 2017:9042851. doi: 10.1155/2017/9042851
49. Zhu X, Meng G, Dickinson BL, Li X, Mizoguchi E, Miao L, et al. MHC class I-related neonatal fc receptor for IgG is functionally expressed in monocytes, intestinal macrophages, and dendritic cells. J Immunol Baltim Md 1950 (2001) 166(5):3266−76. doi: 10.4049/jimmunol.166.5.3266
50. Spiekermann GM, Finn PW, Ward ES, Dumont J, Dickinson BL, Blumberg RS, et al. Receptor-mediated immunoglobulin G transport across mucosal barriers in adult life. J Exp Med (2002) 196(3):303−10. doi: 10.1084/jem.20020400
51. Cervenak J, Bender B, Schneider Z, Magna M, Carstea BV, Liliom K, et al. Neonatal FcR overexpression boosts humoral immune response in transgenic mice. J Immunol Baltim Md 1950 (2011) 186(2):959−68. doi: 10.4049/jimmunol.1000353
52. Vidarsson G, Stemerding AM, Stapleton NM, Spliethoff SE, Janssen H, Rebers FE, et al. FcRn: an IgG receptor on phagocytes with a novel role in phagocytosis. Blood (2006) 108(10):3573−9. doi: 10.1182/blood-2006-05-024539
53. Baker K, Qiao SW, Kuo TT, Aveson VG, Platzer B, Andersen JT, et al. Neonatal fc receptor for IgG (FcRn) regulates cross-presentation of IgG immune complexes by CD8–CD11b+ dendritic cells. Proc Natl Acad Sci U.S.A. (2011) 108(24):9927−32. doi: 10.1073/pnas.1019037108
54. Kerrigan AM, Brown GD. C-type lectins and phagocytosis. Immunobiology (2009) 214(7):562−75. doi: 10.1016/j.imbio.2008.11.003
55. Jin F, He J, Jin C, Fan W, Shan Y, Zhang Z, et al. Antithymocyte globulin treatment at the time of transplantation impairs donor hematopoietic stem cell engraftment. Cell Mol Immunol (2017) 14(5):443−50. doi: 10.1038/cmi.2015.92
56. Préville X, Flacher M, LeMauff B, Beauchard S, Davelu P, Tiollier J, et al. Mechanisms involved in antithymocyte globulin immunosuppressive activity in a nonhuman primate model. Transplantation (2001) 71(3):460−8. doi: 10.1097/00007890-200102150-00021
57. Gurkan S, Luan Y, Dhillon N, Allam SR, Montague T, Bromberg JS, et al. Immune reconstitution following rabbit antithymocyte globulin. Am J Transplant Off J Am Soc Transplant Am Soc Transpl Surg (2010) 10(9):2132−41. doi: 10.1111/j.1600-6143.2010.03210.x
58. Esposito L, Kamar N, Tkaczuk J, Abbal M, Durand D, Rostaing L. Long-term evolution of lymphocytes subsets after induction therapy based on continuous versus discontinuous administration of anti-thymocyte globulins in renal-transplant patients. Transplant Proc (2005) 37(2):785−7. doi: 10.1016/j.transproceed.2004.12.200
59. Charpentier B, Rostaing L, Berthoux F, Lang P, Civati G, Touraine JL, et al. A three-arm study comparing immediate tacrolimus therapy with antithymocyte globulin induction therapy followed by tacrolimus or cyclosporine a in adult renal transplant recipients. Transplantation (2003) 75(6):844−51. doi: 10.1097/01.TP.0000056635.59888.EF
60. Issa NC, Fishman JA. Infectious complications of antilymphocyte therapies in solid organ transplantation. Clin Infect Dis Off Publ Infect Dis Soc Am (2009) 48(6):772−86. doi: 10.1086/597089
61. Ducloux D, Courivaud C, Bamoulid J, Crepin T, Chalopin JM, Tiberghien P, et al. Polyclonal antithymocyte globulin and cardiovascular disease in kidney transplant recipients. J Am Soc Nephrol JASN (2014) 25(6):1349−56. doi: 10.1681/ASN.2013060663
62. Fehr T, Cippà PE, Mueller NJ. Cytomegalovirus post kidney transplantation: Prophylaxis versus pre-emptive therapy? Transpl Int Off J Eur Soc Organ Transplant (2015) 28(12):1351−6. doi: 10.1111/tri.12629
63. Bicalho PR, Requião-Moura LR, Arruda ÉF, Chinen R, Mello L, Bertocchi APF, et al. Long-term outcomes among kidney transplant recipients and after graft failure: A single-center cohort study in Brazil. BioMed Res Int (2019) 2019:e7105084. doi: 10.1155/2019/7105084
64. Cippà PE, Schiesser M, Ekberg H, van GT, NJ M, CA C, et al. Risk stratification for rejection and infection after kidney transplantation. Clin J Am Soc Nephrol. (2015) 10(12):2213−20. doi: 10.2215/CJN.01790215
65. Wang K, Xu X, Fan M. Induction therapy of basiliximab versus antithymocyte globulin in renal allograft: A systematic review and meta-analysis. Clin Exp Nephrol (2018) 22(3):684−93. doi: 10.1007/s10157-017-1480-z
66. Cherukuri A, Mohib K, Rothstein DM. Regulatory b cells: TIM-1, transplant tolerance, and rejection. Immunol Rev (2021) 299(1):31−44. doi: 10.1111/imr.12933
67. Garcia SG, Sandoval-Hellín N, Franquesa M. Regulatory b cell therapy in kidney transplantation. Front Pharmacol (2021) 12:791450. doi: 10.3389/fphar.2021.791450
68. Chesneau M, Michel L, Degauque N, Brouard S. Regulatory b cells and tolerance in transplantation: From animal models to human. Front Immunol (2013) 4:497. doi: 10.3389/fimmu.2013.00497
69. Martin-Moreno PL, Tripathi S, Chandraker A. Regulatory T cells and kidney transplantation. Clin J Am Soc Nephrol CJASN (2018) 13(11):1760−4. doi: 10.2215/CJN.01750218
70. Lopez M, Clarkson MR, Albin M, Sayegh MH, Najafian N. A novel mechanism of action for anti-thymocyte globulin: Induction of CD4+CD25+Foxp3+ regulatory T cells. J Am Soc Nephrol JASN. (2006) 17(10):2844−53. doi: 10.1681/ASN.2006050422
71. Tector AJ, Mosser M, Tector M, Bach JM. The possible role of anti-Neu5Gc as an obstacle in xenotransplantation. Front Immunol (2020) 11:622. doi: 10.3389/fimmu.2020.00622
72. Estrada J, Martens G, Li P, Adams A, Newell K, Ford M, et al. Evaluation of human and nonhuman primate antibody binding to pig cells lacking GGTA1/CMAH/β4GalNT2 genes. Xenotransplantation (2015) 22(3):194−202. doi: 10.1111/xen.12161
Keywords: solid organ transplantation, induction, anti-lymphocyte globulin, anti-lymphocyte antibody, polyclonal antibodies (PAbs), kidney trans plantation, pig IgG
Citation: Rousse J, Royer P-J, Evanno G, Lheriteau E, Ciron C, Salama A, Shneiker F, Duchi R, Perota A, Galli C, Cozzi E, Blancho G, Duvaux O, Brouard S, Soulillou J-P, Bach J-M and Vanhove B (2023) LIS1, a glyco-humanized swine polyclonal anti-lymphocyte globulin, as a novel induction treatment in solid organ transplantation. Front. Immunol. 14:1137629. doi: 10.3389/fimmu.2023.1137629
Received: 04 January 2023; Accepted: 03 February 2023;
Published: 16 February 2023.
Edited by:
Jean Kwun, Duke University, United StatesReviewed by:
Joseph Ladowski, Duke University, United StatesCopyright © 2023 Rousse, Royer, Evanno, Lheriteau, Ciron, Salama, Shneiker, Duchi, Perota, Galli, Cozzi, Blancho, Duvaux, Brouard, Soulillou, Bach and Vanhove. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Juliette Rousse, anVsaWV0dGUucm91c3NlQHhlbm90aGVyYS5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.