
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Immunol. , 27 July 2023
Sec. Parasite Immunology
Volume 14 - 2023 | https://doi.org/10.3389/fimmu.2023.1134020
This article is part of the Research Topic Women in Cytokines and Soluble Mediators in Immunity View all 63 articles
 Nasreddine Saidi1,2
Nasreddine Saidi1,2 Romain Blaizot1,3,4
Romain Blaizot1,3,4 Ghislaine Prévot1
Ghislaine Prévot1 Karim Aoun2,5
Karim Aoun2,5 Magalie Demar1,3,4Pierre André Cazenave1Aida Bouratbine2,5
Magalie Demar1,3,4Pierre André Cazenave1Aida Bouratbine2,5 Sylviane Pied1*
Sylviane Pied1*Cutaneous leishmaniasis (CL) caused by infection with the parasite Leishmania exhibits a large spectrum of clinical manifestations ranging from single healing to severe chronic lesions with the manifestation of resistance or not to treatment. Depending on the specie and multiple environmental parameters, the evolution of lesions is determined by a complex interaction between parasite factors and the early immune responses triggered, including innate and adaptive mechanisms. Moreover, lesion resolution requires parasite control as well as modulation of the pathologic local inflammation responses and the initiation of wound healing responses. Here, we have summarized recent advances in understanding the in situ immune response to cutaneous leishmaniasis: i) in North Africa caused by Leishmania (L.) major, L. tropica, and L. infantum, which caused in most cases localized autoresolutives forms, and ii) in French Guiana resulting from L. guyanensis and L. braziliensis, two of the most prevalent strains that may induce potentially mucosal forms of the disease. This review will allow a better understanding of local immune parameters, including cellular and cytokines release in the lesion, that controls infection and/or protect against the pathogenesis in new world compared to old world CL.
Leishmaniasis is a parasitic disease caused by a vector-borne protozoan parasite belonging to the Leishmania genus. It is transmitted as a flagellated promastigote via the bite of an infected sandfly (1, 2). Following its inoculation, promastigotes are ingested by innate cells mainly macrophages, neutrophils, and dendritic cells, where they evolve into amastigotes. Leishmania parasite are distributed in more than 98 countries, with 350 million people exposed and 12 million infected (3, 4). The cutaneous form is the most frequent, with an estimation of 600 000 to 1 million new cutaneous leishmaniasis (CL) cases per year. This disease is also responsible for significant psychosocial impacts due to the scars that persist after recovery and the associated social stigma (5). In the Old World, specifically in North Africa (NA), CL is caused by three species: L. major, L. tropica, and L. infantum. Cutaneous leishmaniasis is highly prevalent in Morocco, Algeria, and Tunisia. In the New World (NW), CL is widely distributed in South and Central America. It is mainly caused by L. amazonensis, L. guyanensis, L. panamensis, L. peruviana, L. mexicana, and L. braziliensis (6). These species can cause cutaneous and sometimes mucosal leishmaniasis (ML), with only a small potential for self-healing. French Guiana (FG) is a good example of an endemic South American country, where numerous works have been published on CL. The disease is basically divided into four clinical phenotypes (1): Localized CL (LCL) (2), mucocutaneous leishmaniasis, and (3) diffuse and (4) disseminated CL (7–9). Skin lesions result from a deregulated immune response which is unable to eliminate the intracellular parasites. This could explain the high number of parasites in the inflammatory infected zone for some forms of CL (10). Unbalanced T helper (Th)1/Th2 responses has been associated with an increased tissue destruction and a worsening of skin lesions. Moreover, in a susceptible murine model of L. major infection, improved resistance to infection was linked to the production of cytokines in lymph nodes. In most mouse strains, Th1 cells bring resistance through the secretion of IFN-γ. In contrast, susceptible mice generate a Th2 cell response characterized by the production of IL-4 and IL-13, which hampers the ability of IFN-γ to trigger toxic metabolites (11). The evolution of lesions is determined by a complex interaction between many factors triggered by the early immune responses, including innate and acquired immune mechanisms (10). However, there is a significant lack of information on the cutaneous immune response within CL lesions determining the evolution of the disease both in NA and FG. A better knowledge of the pattern of immune mechanisms and related factors involved in lesion progression or healing might provide helpful information for identifying new immunotherapeutic targets and new drugs.
In this review, we will sum up the current knowledge on clinical manifestations, lesion evolution and local immune response associated with L. infantum, L. major and L. tropica in NA and L. guyanensis and L. braziliensis CL in FG. We will also discuss similarities and divergences in immune mechanisms induced in NA and FG.
The clinical presentation and the outcome of CL depend on multiple factors, including species involved, lesion location, sandfly infectivity, comorbidities, treatment modalities, skin microbiota and host immune responses against Leishmania (Figure 1) (23–25). The clinical appearance of CL is initially characterized by an erythematous, non-specific papule or patch following an incubation period after the bite of the Leishmania infected sandfly. This sub-clinical or early CL has the potential for self-healing; it then evolves into ulcerated (85%) or non-ulcerated (nodules, papules, plaques 15%) specific lesions. Some lesions have a very small potential for self-healing, especially those in the New World, and almost always require a treatment; the median incubation of CL in French Guiana is 25 days. Hypopigmented, hyperpigmented or atrophic scars can persist after treatment, depending on the skin phototype (26–31). In some cases, the lesions can persist for more than one year and become a chronic non-healing form (32, 33). Therefore, some infected patients leading to disfigured (34–36). However, the disease outcome also depends on the parasite species (Table 1).
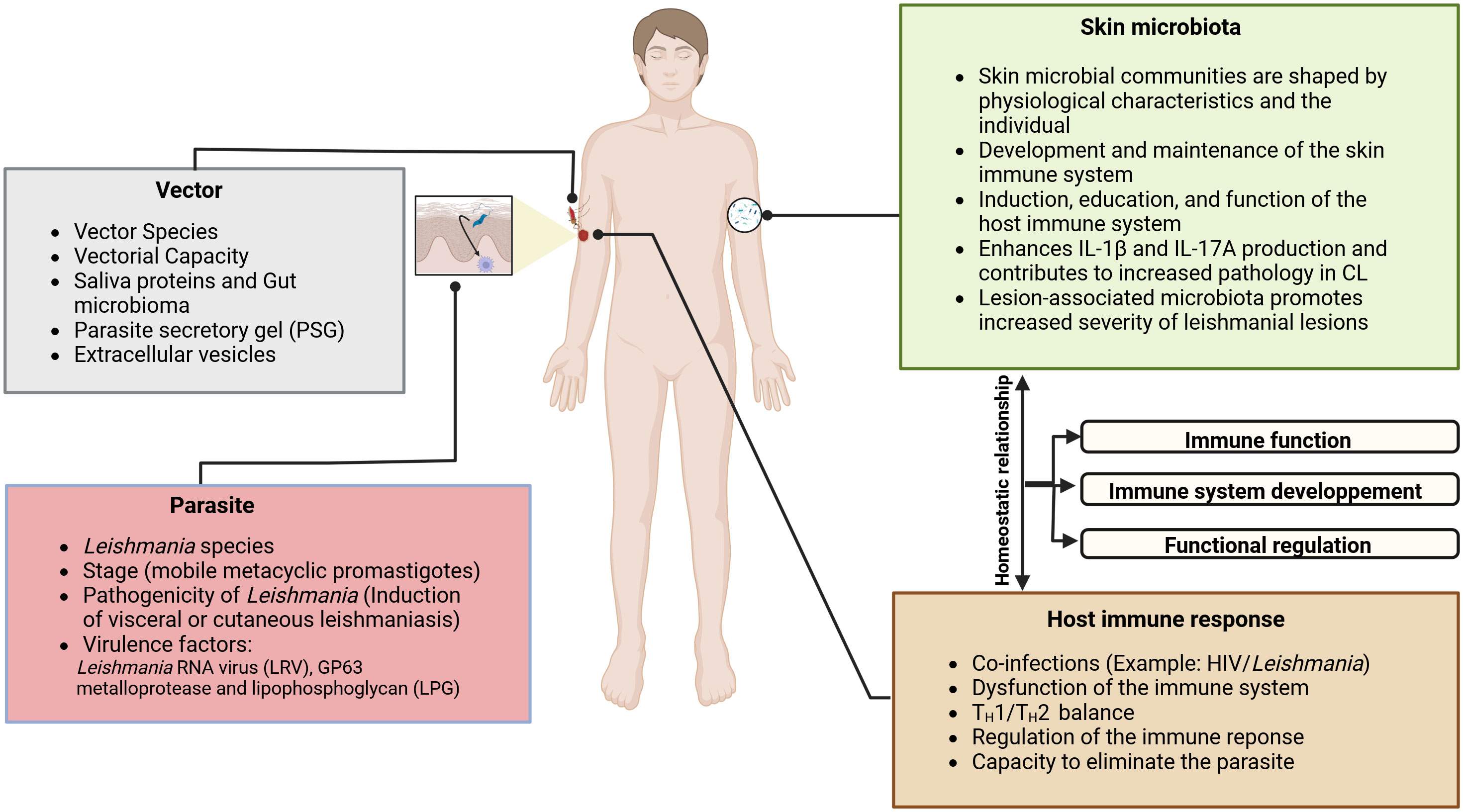
Figure 1 The pathogenesis of CL and the evolution of lesions are multifactorial; it depends on the complex interactions between the Leishmania parasite, the immune system, and the skin environment, including the vector-injected particles and the microbial skin communities. Sandfly saliva contains potent vasodilators, maxadilan, and adenosine, described respectively in Lutzomyia longipalpis and P. papatasi, that prevent clotting at the biting site (12), in addition of proteins that trigger a host immune response (13–15). As clearly demonstrated by several investigators, sandfly saliva contains immunomodulatory molecules that have been shown to enhance disease progression (16–18). Some of them, such as the parasite secretory gel (PSG), afford Leishmania protection from a hostile pro-inflammatory environment. They may directly regulate macrophages activation in dampening the early pro-inflammatory response and orchestrating wound repair and re-epithelialization (19). In fresh wounds, a robust pro-inflammatory response is required to sterilize the damaged tissue of potentially pathogenic bacteria. The skin microbiota plays a fundamental role in the host immune system’s induction, education, and function. In turn, the host immune system has evolved multiple means to maintain its homeostatic relationship with the microbiota. It has been shown that Staphylococcus spp., Streptococcus spp., Enterococcus spp, Pseudomonas spp, and other opportunistic bacteria are present in CL lesions (20–22). It was proven using a Leishmania-infected mouse with dysbiotic skin microbiota that naturally acquired dysbiosis can cause a change in inflammatory responses and disease progression. It has been demonstrated that the skin microbiome could modulate the skin’s immune response by enhancing IL17 production, which is essential in mediating inflammation in CL. While Th17 cells are a source of IL-17, it is possible that IL-17 produced by innate lymphoid cells present in the skin could contribute to disease progression. In effect, Scoot’s team has suggested that following infection by L. major, RORγt+ILCs produced IL-17 in the skin may contribute to disease progression.
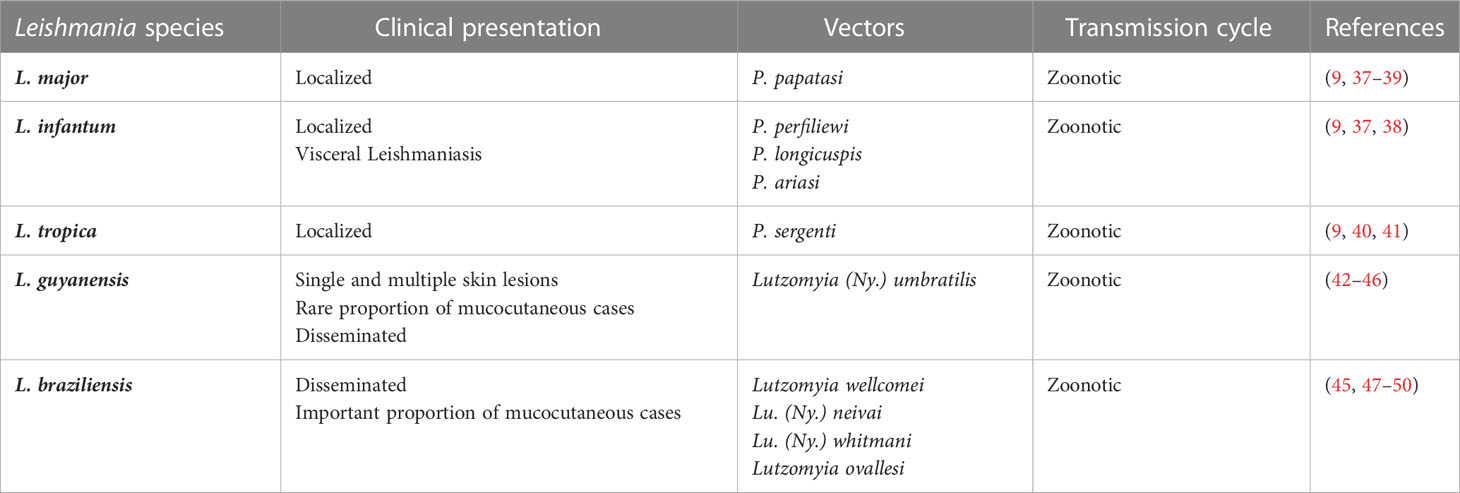
Table 1 Summary of the different Leishmania species, clinical presentations, vector and transmission cycles in North African countries (Old World) and French Guiana (New World).
Concerning L. major, this species is the most frequent in NA countries, where it remains a major public health problem, with more than 1000 to 8 000 cases yearly in Tunisia and 10,000 to 25,000 in Algeria (9, 40, 51, 52). Lesion can be single or multiple, usually with ulcerative nodules. These primary lesions can give birth to secondary peripheral lesions “satellite” (Figure 2) (9, 58). The most common type of ulcer is the ulcero-crusted form (59), also known as the “wet” or “rural” type. It is characterized by a painless ulcer with a clear raised border and a brownish scab covering it, commonly affecting the upper and lower limbs. CL caused by L. major has a wide range of clinical presentations, so it should be considered as a possibility in many skin diseases, including actinomycetoma, pyoderma gangrenosum, and various types of skin cancer. In Tunisia, about 11 uncommon clinical forms of CL due to L. major have been reported in zoonotic foci in Central and South regions of the country (60–62). This diversity may be due to the combination of the parasite’s genetics and the host’s immune response.
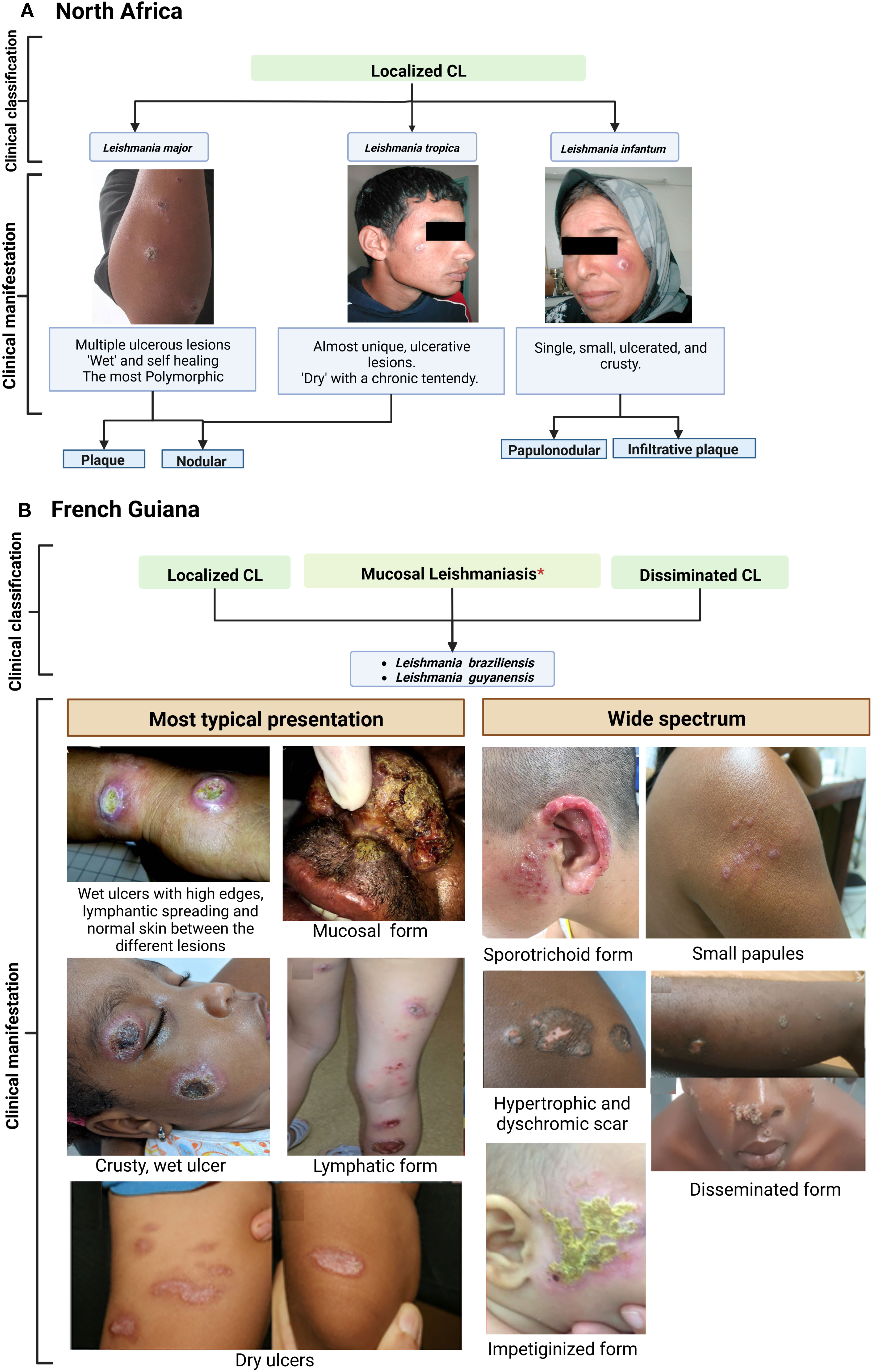
Figure 2 Clinical features of cutaneous leishmaniasis caused by L. major, L. tropica and L. infantum (Old world) (A) compared to L. braziliensis and L. guyanensis (New World) (B). Case photos were provided by the laboratory of Medical Parasitology and Mycology. Institute Pasteur of Tunis. and Dermatology Department, Centre Hospitalier de Cayenne, French Guiana. *Risk for mucosal involvement is about 6% for L. braziliensis and 1% for L. guyanensis. Reference: North Africa; French Guiana (9, 53–57).
L. tropica, is responsible for chronic CL (CCL). Among the different countries of NA, Morocco presents the widest endemic areas and the highest incidence for this species (9). Compared with L. major, cutaneous lesions are distinguished by their chronicity; their appearance is dry and slightly inflamed. Therefore, this makes L. tropica CL more difficult to diagnose, which causes a delay in patient care (9, 63–66). Lesions in CCL are drier than in L. major, with papules, nodules, or crusty ulcers. Also, they are different in location and size compared to other species, as it mainly appears on the face and are typically smaller, with a diameter of less than 2 cm (41, 51, 67, 68).
The third CL causative species in NA is L. infantum, the primary responsible agent of visceral leishmaniasis (VL) (69). Between 50 and 150 cases are reported annually in Tunisia; few data are available for other countries (9). Compared to the previous two, the clinical appearance of wounds is more significant (58, 70). The duration between the bite of the infective sandfly and the onset of symptoms may vary between several weeks to 1 year (average 3-6 months) (51, 71). Another characteristic is that lesions may last longer than those with L. major, persisting for several months or years; such lesions are often slow to heal and may leave large, disfiguring, or disabling scars. Clinically, L. infantum lesions may present as papules or nodules, typically on the extremities and bordered by a large zone of infiltration (9, 37). Plaque is the most frequent clinical manifestation of cutaneous L. infantum involvement, followed by ulcers, nodules, and papules (72). The clinical presentation is a single small plaque on the face (9). Cases of mucosal involvement have recently been reported in travelers returning to Europe but are seldom described by North African teams (73).
In French Guiana, CL is caused by certain species belonging to the Leishmania Viannia subgenus, which is endemic to Central and South America (74). L. guyanensis represents more than 80% of cases in FG, while L. braziliensis has emerged in the 2000s and represents about 10-15% of cases (75). Other species, such as L. lainsoni, L. naiffi, and L. amazonensis can also be found but remain rare and are not part of this review (76). Though a complex clinical classification has not been proposed in FG, a very wide spectrum of clinical lesions can be observed, including impetigo-like, lupoid-like, sporotrichoid, crusted ulcers, patches, plaques, nodules, papules, or even extensive sores (Figure 2). These atypical forms have also been described in other countries of South America, notably Brazil (77, 78). However, the typical form of CL in French Guiana and the rest of the Amazon basin consists of a wet, acute ulcer which is slightly painful and usually without secondary bacterial infection (44). Several lesions can be found, particularly when L. guyanensis is involved. The skin between the lesions is normal, without erythema. A lymphangitis with or without parasitic nodules can be observed. Compared to Old World (OW) leishmaniasis, New World (NW) forms are known for a greater tendency to disseminate systemically in the skin (6, 79).
In addition, a great number of lesions with lymphatic involvement are observed for L. guyanensis CL in FG (42, 43) (Figure 2). This species is known to occasionally cause mucosal issues, but it is less common than mucosal disease caused by L. braziliensis (44, 80–82). Indeed, less than 1% of L. guyanensis cases in FG display a mucosal involvement (81, 83). L. braziliensis CL can manifest in different ways, from a single ulcer on the skin, which is often associated with satellite adenomegaly, to the less frequently disseminated form (82, 84). Potentially severe mucosal involvement of the upper airways are typically described with this species (47, 82, 85–87). Single lesions are more frequent than with L. guyanensis (44, 83).
CL in NA and FG is characterized by various immunological features (Figure 3) (44, 47, 58, 74, 88–90). Once the Leishmania infected sandfly bites, the infection starts with an asymptomatic “silent phase” of variable duration, characterized by an inflammatory wave of poly morpho nuclear (PMN), dendritic cells (DC), and monocyte-derived macrophages which harbour a proliferation of amastigotes intracellular parasites (10, 91, 92). Then, a massive recruitment of CD4+ and CD8+ T lymphocytes, with enhanced pro-inflammatory responses participate in granuloma formation and parasite control. As a result, few parasites remain in the lesions and promote a delayed-type hypersensitivity (DTH) (Figure 4) (10, 93, 94). However, differences in intralesional immune profiles between the different CL forms are not well documented. Still, the infection outcome is clearly the consequence of a balance between pro and anti-inflammatory responses.

Figure 3 The T helper 1 (Th1)-type T helper 2 (Th2)-type balance across the cutaneous leishmaniasis severity. Mucosal and disseminated CL caused by L. guyanensis and L. braziliensis are the most severe form of the disease. They are on opposite sides of the Th1 and Th2 response, which enhances disease severity in an exaggerated response. An uncontrolled Th1 response in mucocutaneous leishmaniasis can cause an exaggerated cellular response, in which Leishmania amastigotes spread to the nasopharyngeal mucosa and cause tissue damage resulting in disfiguring lesions. For localized cutaneous leishmaniasis, mixed TH1 and TH2 responses have been observed during the active stage of infection; the Th1 profile is mainly associated with the healing of lesions.
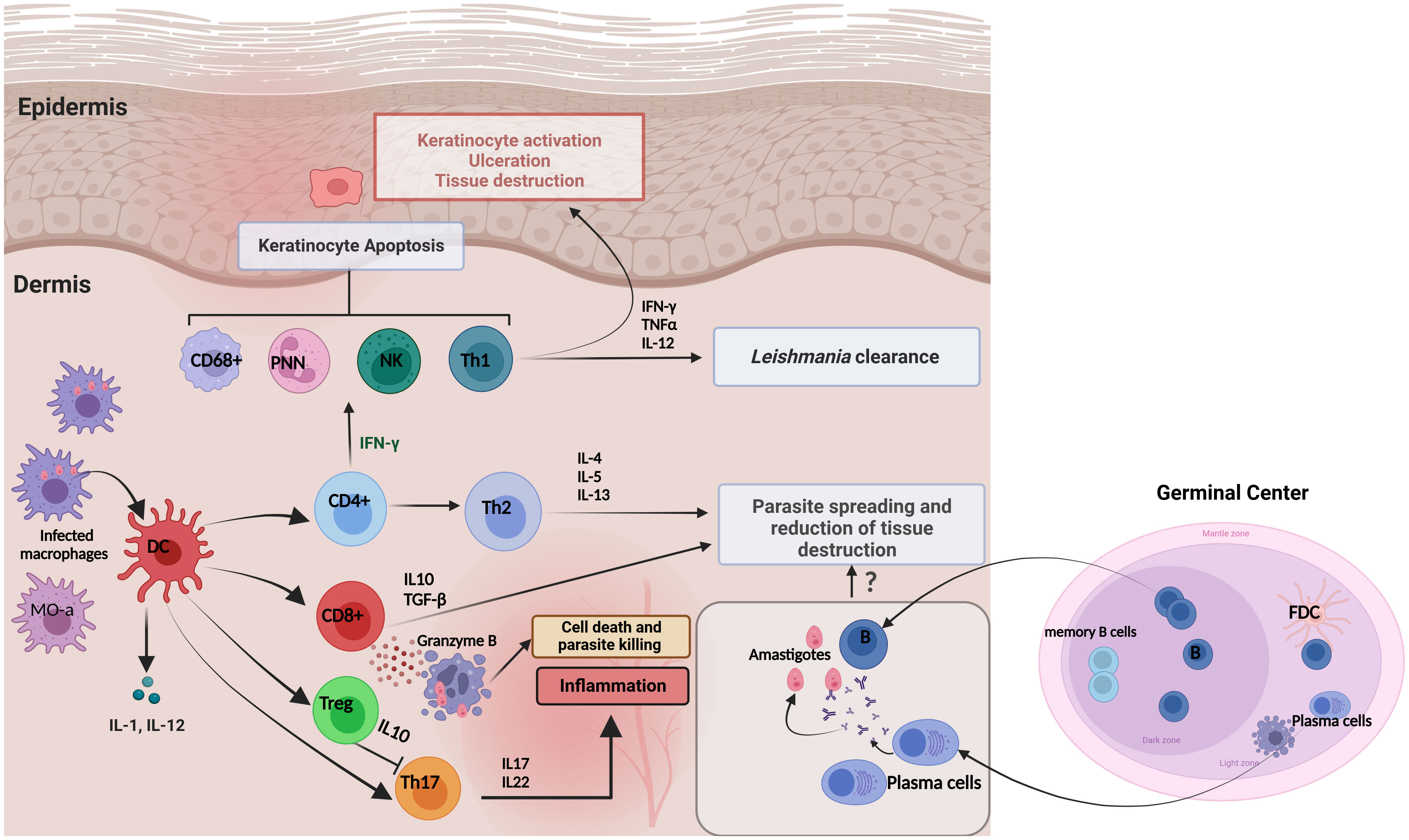
Figure 4 Immune responses and skin cytokine profile during cutaneous leishmaniasis infection. Pro-inflammatory cytokines are produced primarily to amplify the immune response to Leishmania infection. The major proinflammatory cytokines include TNF-α, IFN-γ, IL-1, IL-8, IL-12, IL-17 and Granzyme β. In contrast, anti-inflammatory cytokines are immunoregulatory molecules that counteract the effects of pro-inflammatory cytokines to limit the inflammation. These major anti-inflammatory cytokines include IL-5, IL-6, IL-4, IL-10, IL-13, and TGF-β. It has been proposed that the existence of Tregs in infected tissues could be an immune response from the host to maintain the balance to control Leishmania infection and reduce excessive inflammation that supports parasite survival. This is achieved by Tregs inhibiting Th17 cells through the production of IL-10. The role of the local humoral response to amastigote spreading and tissue destruction is still uncompletely understood.
During L. major infection, promastigotes deposition in the skin promotes neutrophils and dendritic cells (DCs) recruitment during the earliest stage of infection (95). In this stage, Chaves et al. demonstrated the anti-inflammatory functions of dermal tissue-resident macrophages, which was supported by the reduced parasite burdens observed in mice (96). The clearance and control of L. major multiplication involved mainly cytotoxic CD8+ T cells and CD4+ helper T cells and the production of IFN-γ (90, 97); in fact, non-healing lesions in mice are associated with a small number of TCD4+ (98, 99). However, PWK mice strain develops prolonged but self-healing lesions, with an immune response characterized by a mixed Th1-plus-Th2 pattern acquiring resistance to a secondary challenge (100). Formaglio et al. recently showed the implication of the resident memory CD4+ T cells in delayed-hypersensitivity response, without the involvement of circulating T cells but with the recruitment of activated inflammatory monocytes. These monocytes produce reactive oxygen species (ROS) and nitric oxide (NO) that play a role in the inhibition of L. major development in the mice (101). These findings underscore the central role of skin-resident CD4+ T cells’ in enhancing the protective immune response against L. major (102). Independently of the T resident memory cells, the acute availability of circulating CD4+ T helper 1 effector cells (Th1EFF) at the time of secondary infection is critical for the Th1 immune response. Th1EFF cell-phagocyte interactions proved crucial in preventing the establishment of a permissive L. major niche in lesions (103). However, if NK cells may play several roles in developing an effective T cell response against Leishmania, their contributions to the response to CL in humans are less clear. For instance, it has been shown that NK-cell-derived IFN-γ, in mice, is essential for activating the dendritic cells that mediate the T-cell-dependent protection against L. major infection (104). Another mechanism associated with Human skin lesions protection is granzyme B−dependent CD8 T-cell cytotoxicity, as it participates in controlling parasite multiplication through a cytotoxic process (88, 105). On the other hand, in mice, a negative regulatory role for IL-4 was identified in limiting the recruitment of Th1 cells to L. major infected tissues, which alters the Leishmania clearance (89, 106). Therefore, the study of the impact of IL-4 on lesion evolution could provide hints for a therapeutic trial (107–109). In addition, the cytokine balance IL-4/IFN-γ has also been demonstrated to participate in the commitment of local immune response. Indeed, IL-4 secretion at the site of L. major infection rather than low IFN-γ production may play a role in the prolongation of disease during acute and chronic CL lesions. A moderate increase of CD4 Tregs would be observed in chronic lesions. However, few studies have suggested the role of Tregs during human CL; Belkaid et al. demonstrated that Tregs are rapidly accumulated at sites of L. major infection, favoring the early parasite expansion, contributing to the maintenance of immunological self-tolerance, and coinciding with the expression of effective immune responses (98, 110). In a group of L.major infected patients, Hoseini et al. observed a significantly higher expression of Foxp3 in chronic lesions compared to acute lesions; The moderate increase of T reg in chronic lesions and their function in persistent infection is still not apparent (111).
L. infantum is mostly responsible for visceral leishmaniasis (LV) but can also cause CL; little is known about the pathogenesis of this form. Recently, the level of INF-γ was evaluated for sporadic CL and zoonotic CL and was found to be significantly higher in L. infantum lesions than due to L. major (90). As mentioned above, for CL caused by L. major, CD8+ T cells are an essential part of the defense mechanisms against the parasite; however, for L. infantum infection, IFN-γ has a long-range effect inducing skin tissue destruction and keratinocyte apoptosis (10, 112, 113). In addition, it has been shown that increased IL4 production in skin lesions of L. infantum- infected dogs is associated with severe clinical signs and a high parasite burden (114). Although the role of IL4 was little described for L. major infection, no data was found for L. infantum human cutaneous Leishmaniasis. Furthermore, there is still much to be learned regarding the role of T reg cells on skin lesions in human CL caused by L. infantum; authors have suggested that T reg cells and the regulatory cytokines, especially TGF-β, play an essential role in the immunopathogenesis of non-typical ulcerated leishmaniasis (NUCL), modulating the cellular immune response in the skin, avoiding tissue damage, and leading to low parasitic persistence in the skin (115, 116). These factors could play a role in regulating the cellular immune response balance, resulting in the maintenance of a low tissue parasitism that avoids lesion growth.
Although little is known about the in situ immune status of L. tropica-infected patients, cases develop a chronic CL with a strong delayed-type hypersensitivity (117). It has been recently, shown that L. tropica has an enhanced capacity to reduce NO production by macrophages in vitro, which could help to understand why L. tropica infection could induce chronic lesions (118). In a study by Ajdary et al., evaluating T-cell responses to Leishmania antigen in vitro, Th2 cell response was dominant in active CL cases, and Th1 characterized the group of healed patients (119). The level of specific cytokine was evaluated in vitro in the PBMC but not in the skin, and high levels of IFN-γ, IL-5, and IL-13 in non-healing patients were observed, suggesting a mixed Th1/Th2 response with chronic lesions. In contrast, patients with acute lesions respond to infection via a Th1-type response (119). However, such a result must be confirmed locally in skin lesions or animal models. It has been discovered that in the early stages of L. tropica infection, the amount of IL-4 is upregulated and linked to a higher parasite burden. This may play a role in the development of CL by inhibiting the protective immune response. These findings indicate that the parasites may rapidly multiply at the beginning of the infection, leading to a Th2-type immune response (120). Information on regulatory immune responses is lacking for L. tropica CL. However, L. tropica-infected lesions from Indian patients, the analysis of localized immune response reveals the presence of Th17 and T reg cells (121). Because of the difficulties in establishing infection in vivo, published data using animal models for leishmaniasis caused by L. tropica is limited (122). Nevertheless, it was suggested that the presence of Tregs in infected tissues may be a possible host immune response or homeostatic mechanism to control L. tropica infection and to reduce the excessive inflammation supporting parasite survival through IL-10. Still, additional investigations are needed to clarify this response.
The severity of lesions that develop in patients infected by L. braziliensis is mainly associated with a highly inflammatory cutaneous environment. In fact, patients with L. braziliensis infection exhibit a strong T-helper 1 (Th1) immune response, which leads to excessive inflammation and tissue damage (123). High levels of both IFN-γ and TNF-α are observed in CL caused by this species. Still, while IFN-γ may have a protective function (124), there is strong evidence to support the role of TNF-α in the pathology of cutaneous and mucosal lesions (125–130). Thiago Cardoso and colleagues (2014) studied the protective and pathological functions of CD8+ T Cells in L. braziliensis infection. The frequency of CD8+ T cells expressing granzyme in the lesions of severe CL patients is more significant than that in patients during early stage of CL (131). However, cytotoxic CD8 T cells are harmful to both L. braziliensis and infected host cells because cytotoxicity is higher in mucocutaneous leishmaniasis than in the localized form (132, 133). Recently, it has been observed a high proportion of senescent T cells (Tsen) with high inflammatory profiles in L. braziliensis lesions with mucosal involvement (134, 135), which is linked to the severity and tissue damage (136). Therefore, in vivo, senescent CD8+ T cells appear to be the most important cell populations mediating skin pathology (137). Other cells, particularly Treg cells, accumulate in lesions caused by L. braziliensis, and contribute to the local control of effector T-cell functions (138). Campanelli et al. research suggests that Treg accumulated at the sites of L. braziliensis infection may contribute to the local control of effector T cell functions. Still, a direct correlation with the pathogenesis is yet to be detected (138).
Little is known about the immune responses induced during human infection with L. guyanensis, which generally generates a mixed Th1/Th2/Th17 immune response. IL-13 is the main Th2 cytokine found in L. guyanensis LCL lesions, according to research by Bourreau and colleagues. As IL-13 has many similar effects with IL-4 (139), patients with L. guyanensis LCL are likely to have a Th2 response that includes the production of either IL-4 or IL-13. However, IL-13 plays a key role in maintaining the Th2 response in Human leishmaniasis by making certain cells resistant to IL-12 (140). In L. guyanensis infection, the levels of Foxp3 in the lesions were superior in chronic patients than in acute ones demonstrating the regulatory role of T reg. Furthermore, Tregs isolated from skin biopsy of L. guyanensis patients with acute CL had a suppressive effect on CD4+T effector (eff) cells. Also, Foxp3 expressions were higher in skin biopsies than in peripheral blood mononuclear cells (PBMC), confirming the recruitment of Tregs to the infection site (141, 142). L. guyanensis shows unique characteristics with a mixed immune response (Figures 5, 6), which warrants further investigation to uncover the mechanisms that regulate immune responses and control inflammation caused by this parasite. Recently, a proposed relationship between Leishmania RNA Virus (LRV) and the severity of leishmaniasis has been suggested, as various factors are linked to the pathogenicity of the disease (146). Zabala-Peñafiel and colleagues demonstrated that L. guyanensis metastatic strains had a higher rate of LRV1 positivity than non-metastatic strains (147). During macrophage infection, it caused over-expression of pro-inflammatory mediators such as TNF-α and IL-6. Ginouvès and colleagues found that 74% of the Leishmania (Viannia) subgenus clinical isolates in French Guiana contain LRV1, with the majority being L. braziliensis and L. guyanensis. Patients infected with LRV1-positive L. guyanensis have a high ratio of IL-17A and IFNγ in their skin lesions (146) (Figure 5). This is accompanied by higher levels of IL-17A in blood cell cultures after stimulation with live parasites, compared to those infected with LRV1-free L. guyanensis (148). This demonstrates that virus infection can exacerbate atypical tegumentary leishmaniasis caused by L. guyanensis. These findings suggest that the presence of LRV1 in the parasites not only increases the risk of developing ML but also causes complications in CL. Contradictorily, in a study conducted by Ginouvès et al., no correlation was found between the presence of LRV1 or its genotypes in L. guyanensis parasites and treatment failure, either after the first or second course of treatment of pentamidine (149). Additional research is required to assess a possible link between LRV1-infected parasites and their clinical symptoms.
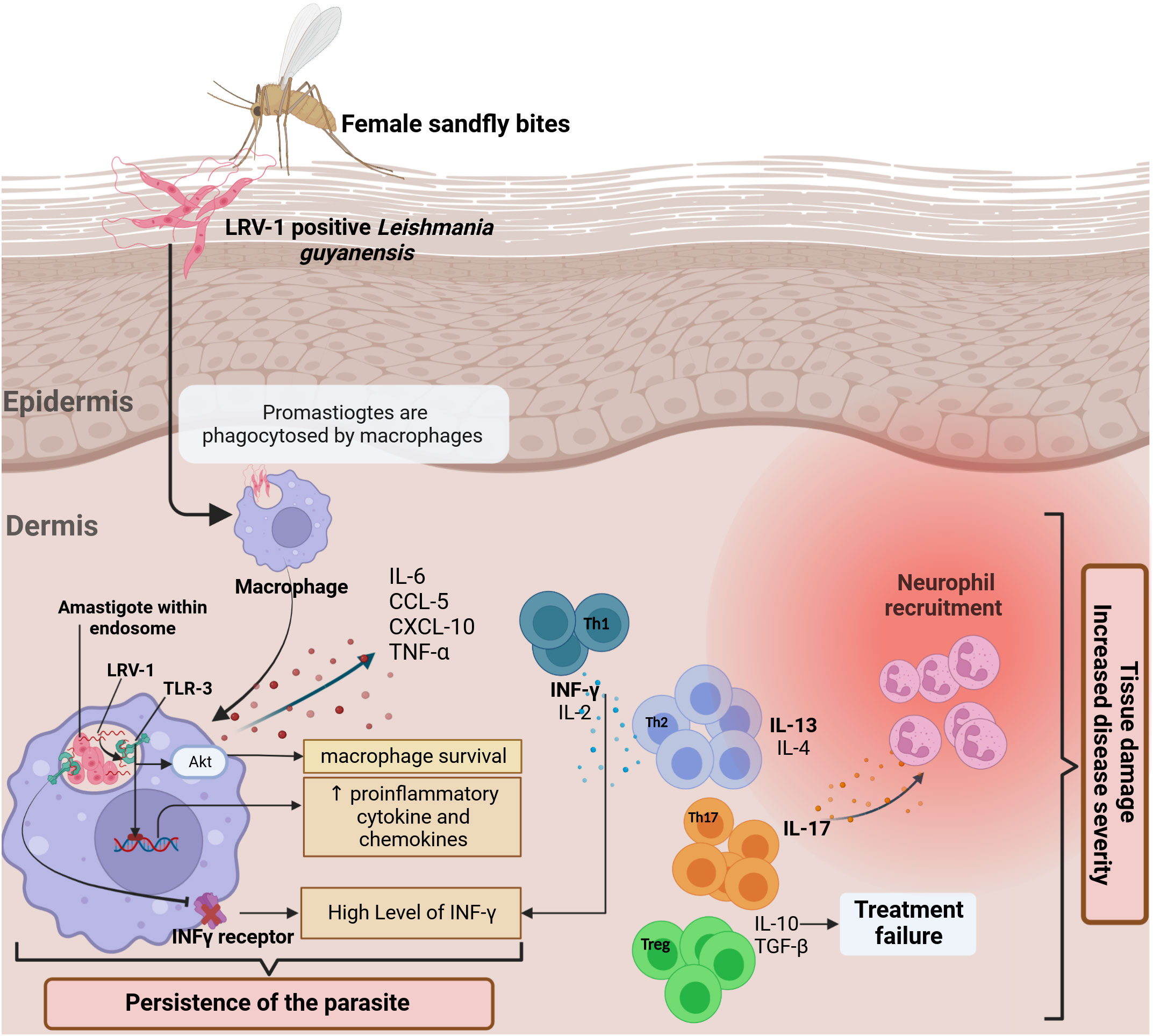
Figure 5 The in situ immune response against L. guyanensis contains a dsRNA virus called (LRV-1). The response to the virus is mediated by TLR 3 in the endosomal macrophage compartment, where the parasite lives and divides. A hypothesis is that after the infection, the viral RNA is released after parasite death and binds to TLR-3 (143), promoting pro-inflammatory cytokines and chemokines production such as IL-6, TNF-α, CXCL-10, and CCL-5 and controlling the severity of the disease (144). Generally, L. guyanensis infection induces a mixed Th1/Th2/Th17 immune response. Th-17 cells appear to predominate in lesions in the presence of Leishmania RNA with high production of IL17-A. TGF-β is essential for establishing infection and, together with IL-10, leads to therapeutic failures and increased disease severity. The stimulation of TLR3 results in the downregulating of IFN-γ receptor expression, reducing macrophage activation, which explains the high level of IFN-γ observed in lesions produced by Th-1. Indeed, via Akt (Protein kinase B) signaling, TLR3 activation by LRV1 promoted parasite persistence (145). Altogether, it enhances inflammation and thus exacerbates disease.

Figure 6 Comparison between the in situ immune profiles in lesions of patients infected with the five Leishmania species (L. major, L. infantum, L. tropica, L. guyanensis, L. braziliensis) and the implication for pathogenesis and the control of the diseases.
The outcome of Leishmania infection relies on the intricate equilibrium of pro- and anti-inflammatory immune responses generated by the host. Both innate and adaptive immunity are inextricably linked to each other as the cytokines produced by cells of the innate system determine the outcome and magnitude of the adaptive immune response. Therefore, the immune responses need to be tightly regulated to avoid immune-mediated pathology to host tissue. Here, we have highlighted recent progress made in understanding the immune response to cutaneous leishmaniasis caused by Leishmania major, L. tropica, and L. infantum in North Africa on the one hand, and by L. guyanensis and L. braziliensis in French Guiana on the other hand. If cellular and cytokines are released in the lesion during OW-CL, much remains to be known in NW-CL. Indeed, this review could significantly contribute to understanding local immune parameters that control and protect the pathogenesis during Leishmania infection. However, a better knowledge of the local immune response in Human CL may help to identify new mechanisms and targets to develop new and more adapted treatments. New local therapies for cutaneous leishmaniasis in NA, French Guiana, and all endemic countries are urgently needed. Drug development is considered a priority, as most current treatments are based on expensive drugs or potential side effects. The cost of anti-Leishmanial drugs is a crucial issue in low-income countries. Besides, therapeutic failures remain frequent, and cases of unresolved CL are often reported, possibly because all current drugs only target the parasite, not the host responses. This review demonstrates how these responses can shape the clinical lesions and lead to different forms. Research efforts should focus on testing immunotherapies that could reduce the severity of pathology seen in some cases of cutaneous leishmaniasis, which could consecutively lessen the duration of antiparasitic courses and their toxicity. In the last decade, several drugs have been developed for other skin inflammatory diseases or as immunotherapy for skin cancers. Some of these drugs inhibit IL-4 or cytotoxicity and might be helpful in combination with existing anti-parasitic drugs. Anti-IL 4 drugs could be useful for all species of Leishmania, while specific inhibitors of IL-13 and IL-17 could be used to dampen tissue damage in L. guyanensis infections (Figure 6). Dupilumab is currently used in atopic dermatitis both for its IL-4 and IL-13 effects and could be tested in association with antiparasitic drugs. Other candidates could include anti-IL17 used for inflammatory skin diseases, such as secukinumab, brodalimumab or ixekizumab. Furthermore, skin dysbiosis has been shown to drive the inflammatory response in L. major infected germ-free mice model (150). These observations emphasize the need to explore mechanisms by which skin microbiota is involved in the physiopathology of CL. In our opinion, it is essential that these advances in the role of microbiota should be included in the research for the development of new drugs and vaccines against CL.
SP designed, revised, and supervised the work. NS reviewed the literature, created the figures, and wrote the manuscript. RB, GP, KA, MD, PC, AB participated in the manuscript’s analysis, drafting, and revising. All authors revised and approved the manuscript’s final version.
This work was supported by LabEx PARAFRAP ANR-11-LABX-0024i and MALTOX FEDER CTG Guyane.
We wish to thank all the authors who have participated in this Research Topic and the reviewers for their insightful comments and suggestions. NS was funded by the Federation of African Immunological Societies (FAIS) Legacy Project (FLP), 2019, and the High-level scientific stays (SSHN) grant from the Minister of Europe and foreign affairs France, 2020.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Sunter J, Gull K. Shape, form, function and leishmania pathogenicity: From textbook descriptions to biological understanding. Open Biol (2017) 7(9). doi: 10.1098/rsob.170165
2. Steverding D. The history of leishmaniasis. Parasites Vectors (2017) 10(1):82. doi: 10.1186/s13071-017-2028-5
3. Ruiz-Postigo JA, Jain S, Maia-Elkhoury AMAN, Valadas S, Warusavithana S, Osman M, et al. Global leishmaniasis surveillance: 2019-2020, a baseline for the 2030 Roadmap/Surveillance mondiale de la leishmaniose: 2019-2020, une periode de reference pour la feuille de route a l’horizon 2030. Weekly Epidemiological Rec (2021) 96:401–19.
4. Torres-Guerrero E, Quintanilla-Cedillo MR, Ruiz-Esmenjaud J, Arenas R. Leishmaniasis: A review. F1000Research (2017) 6:750. doi: 10.12688/f1000research.11120.1
5. Bennis I, Belaid L, De Brouwere V, Filali H, Sahibi H, Boelaert M. “The mosquitoes that destroy your face”. social impact of cutaneous leishmaniasis in south-Eastern Morocco, a qualitative study. PloS One (2017) 12(12):e0189906. doi: 10.1371/journal.pone.0189906
6. Burza S, Croft SL, Boelaert M. Leishmaniasis. Lancet (London England) (2018) 392(10151):951–70. doi: 10.1016/s0140-6736(18)31204-2
7. Hashiguchi Y, Gomez EL, Kato H, Martini LR, Velez LN, Uezato H. Diffuse and disseminated cutaneous leishmaniasis: Clinical cases experienced in Ecuador and a brief review. Trop Med Health (2016) 44(1):2. doi: 10.1186/s41182-016-0002-0
8. Sinha S, Fernández G, Kapila R, Lambert WC, Schwartz RA. Diffuse cutaneous leishmaniasis associated with the immune reconstitution inflammatory syndrome. Int J Dermatol (2008) 47(12):1263–70. doi: 10.1111/j.1365-4632.2008.03804.x
9. Aoun K, Bouratbine A. Cutaneous leishmaniasis in north Africa: A review. Parasite (2014) 21:14. doi: 10.1051/parasite/2014014
10. Scott P, Novais FO. Cutaneous leishmaniasis: Immune responses in protection and pathogenesis. Nat Rev Immunol (2016) 16(9):581–92. doi: 10.1038/nri.2016.72
11. Reiner SL, Locksley RM. The regulation of immunity to leishmania major. Annu Rev Immunol (1995) 13(1):151–77. doi: 10.1146/annurev.iy.13.040195.001055
12. Maryam Ghafari S, Ebrahimi S, Nateghi Rostami M, Bordbar A, Parvizi P. Comparative evaluation of salivary glands proteomes from wild phlebotomus papatasi–proven vector of zoonotic cutaneous leishmaniasis in Iran. Veterinary Med Sci (2021) 7(2):362–9. doi: 10.1002/vms3.368
13. Mahamdallie SS, Ready PD. No recent adaptive selection on the apyrase of Mediterranean phlebotomus: Implications for using salivary peptides to vaccinate against canine leishmaniasis. Evolutionary Appl (2012) 5(3):293–305. doi: 10.1111/j.1752-4571.2011.00226.x
14. Ready PD. Biology of phlebotomine sand flies as vectors of disease agents. Annu Rev entomology (2013) 58:227–50. doi: 10.1146/annurev-ento-120811-153557
15. Zahirnia AH, Bordbar A, Ebrahimi S, Spotin A, Mohammadi S, Ghafari SM, et al. Predominance of leishmania major and rare occurrence of leishmania tropica with haplotype variability at the center of Iran. Braz J Infect Dis (2018) 22(4):278–87. doi: 10.1016/j.bjid.2018.07.005
16. Titus RG, Ribeiro JMC. Salivary gland lysates from the sand fly lutzomyia longipalpis enhance leishmania infectivity. Science (1988) 239(4845):1306–8. doi: 10.1126/science.3344436
17. Rogers KA, Titus RG. Immunomodulatory effects of maxadilan and phlebotomus papatasi sand fly salivary gland lysates on human primary in vitro immune responses. Parasite Immunol (2003) 25(3):127–34. doi: 10.1046/j.1365-3024.2003.00623.x
18. Adly E, Shehata MG, El-Demerdash E, Alfarraj S, Ali Alharbi S, Soliman DE. Impact of anti-sandfly saliva antibodies on biological aspects of phlebotomus papatasi (Diptera: Psychodidae), vector of cutaneous leishmaniasis. Saudi J Biol Sci (2021) 28(5):2695–700. doi: 10.1016/j.sjbs.2021.03.016
19. Leoni G, Neumann PA, Sumagin R, Denning TL, Nusrat A. Wound repair: Role of immune-epithelial interactions. Mucosal Immunol (2015) 8(5):959–68. doi: 10.1038/mi.2015.63
20. Shirazi MH, Ranjbar R, Asgari V, Mohebali M, Hamidian M. Study of bacterial infections among the patients with suspected cutaneous leishmaniasis. Pakistan J Biol sciences: PJBS (2007) 10(24):4555–8. doi: 10.3923/pjbs.2007.4555.4558
21. Vera LA, Macedo JL, Ciuffo IA, Santos CG, Santos JB. [Antimicrobial susceptibility of aerobic bacteria isolated from leishmaniotic ulcers in corte de pedra, ba]. Rev da Sociedade Bras Medicina Trop (2006) 39(1):47–50. doi: 10.1590/s0037-86822006000100009
22. Ziaei H, Sadeghian G, Hejazi SH. Distribution frequency of pathogenic bacteria isolated from cutaneus leishmaniasis lesions. Korean J Parasitol (2008) 46(3):191–3. doi: 10.3347/kjp.2008.46.3.191
23. Handler MZ, Patel PA, Kapila R, Al-Qubati Y, Schwartz RA. Cutaneous and mucocutaneous leishmaniasis: Clinical perspectives. J Am Acad Dermatol (2015) 73(6):897–908. doi: 10.1016/j.jaad.2014.08.051
24. Rojas R, Valderrama L, Valderrama M, Varona MX, Ouellette M, Saravia NG. Resistance to antimony and treatment failure in human leishmania (Viannia) infection. J Infect Dis (2006) 193(10):1375–83. doi: 10.1086/503371
25. Hartley MA, Drexler S, Ronet C, Beverley SM, Fasel N. The immunological, environmental, and phylogenetic perpetrators of metastatic leishmaniasis. Trends Parasitol (2014) 30(8):412–22. doi: 10.1016/j.pt.2014.05.006
26. Laboudi M, Sahibi H, Elabandouni M, Nhammi H, Ait Hamou S, Sadak A. A review of cutaneous leishmaniasis in Morocco: A vertical analysisto determine appropriate interventions for control and prevention. Acta tropica (2018) 187:275–83. doi: 10.1016/j.actatropica.2018.07.019
27. Beniklef R, Aoun K, Boudrissa K, Ben Abid M, Cherif K, Aissi W, et al. Cutaneous leishmaniasis in algeria; highlight on the focus of m’sila. Microorganisms (2021) 9(5). doi: 10.3390/microorganisms9050962
28. Bachi F, Icheboudene K, Benzitouni A, Taharboucht Z, Zemmouri M. [Epidemiology of cutaneous leishmaniasis in algeria through molecular characterization]. Bull la Societe pathologie exotique (1990) (2019) 112(3):147–52. doi: 10.3166/bspe-2019-0087
29. Pearson RD, Sousa A. Clinical spectrum of leishmaniasis. Clin Infect Dis (1996) 22(1):1–13. doi: 10.1093/clinids/22.1.1
30. Davidson RN. 123 - leishmaniasis. In: Cohen J, Powderly WG, Opal SM, editors. Infectious diseases (Fourth edition). Elsevier (2017). p. 1059–64.e1.
31. Yedidia Moser R, Ben-Shimol S, Sagi O, Horev A. Comparison between cutaneous leishmaniasis patients with facial and non-facial lesions. Int J Dermatol (2021) 60(9):1109–13. doi: 10.1111/ijd.15582
32. Akilov OE, Khachemoune A, Hasan T. Clinical manifestations and classification of old world cutaneous leishmaniasis. Int J Dermatol (2007) 46(2):132–42. doi: 10.1111/j.1365-4632.2007.03154.x
33. Mortazavi H, Sadeghipour P, Taslimi Y, Habibzadeh S, Zali F, Zahedifard F, et al. Comparing acute and chronic human cutaneous leishmaniasis caused by leishmania major and leishmania tropica focusing on arginase activity. J Eur Acad Dermatol Venereol: JEADV (2016) 30(12):2118–21. doi: 10.1111/jdv.13838
34. Bennis I, Thys S, Filali H, De Brouwere V, Sahibi H, Boelaert M. Psychosocial impact of scars due to cutaneous leishmaniasis on high school students in errachidia province, Morocco. Infect Dis poverty (2017) 6(1):46. doi: 10.1186/s40249-017-0267-5
35. Bennis I, De Brouwere V, Belrhiti Z, Sahibi H, Boelaert M. Psychosocial burden of localised cutaneous leishmaniasis: A scoping review. BMC Public Health (2018) 18(1):358. doi: 10.1186/s12889-018-5260-9
36. Chraiet-Rezgani K, Bouafif-Ben Alaya N, Habboul Z, Hajjej Y, Aoun K. [Epidemiological and clinical features of cutaneous leishmaniasis in kairouan-Tunisia and characteristics in children]. Bull la Societe pathologie exotique (1990) (2016) 109(2):80–3. doi: 10.1007/s13149-016-0475-4
37. Aoun K, Bouratbine A, Harrat Z, Guizani I, Mokni M, Bel Hadj Ali S, et al. [Epidemiologic and parasitologic data concerning sporadic cutaneous leishmaniasis in northern Tunisia]. Bull Soc Pathol Exot (2000) 93(2):101–3.
38. Aoun K, Kalboussi Y, Ben Sghaier I, Souissi O, Hammami H, Bellali H, et al. Assessment of incubation period of cutaneous leishmaniasis due to leishmania major in Tunisia. Am J Trop Med hygiene (2020) 103(5):1934–7. doi: 10.4269/ajtmh.20-0439
39. Bellali H, Chemak F, Nouiri I, Ben Mansour D, Ghrab J, Chahed MK. Zoonotic cutaneous leishmaniasis prevalence among farmers in central Tunisia, 2014. J agromedicine (2017) 22(3):244–50. doi: 10.1080/1059924x.2017.1318725
40. Tabbabi A. Review of leishmaniasis in the middle East and north Africa. Afr Health Sci (2019) 19(1):1329–37. doi: 10.4314/ahs.v19i1.4
41. Bousslimi N, Ben-Ayed S, Ben-Abda I, Aoun K, Bouratbine A. Natural infection of north African gundi (Ctenodactylus gundi) by leishmania tropica in the focus of cutaneous leishmaniasis, southeast Tunisia. Am J Trop Med hygiene (2012) 86(6):962–5. doi: 10.4269/ajtmh.2012.11-0572
42. Dedet J, Pradinaud R, Gay F. Epidemiological aspects of human cutaneous leishmaniasis in French Guiana. Trans R Soc Trop Med Hygiene (1989) 83(5):616–20. doi: 10.1016/0035-9203(89)90375-1
43. Dedet J-P. Cutaneous leishmaniasis in French Guiana: A review. (1990). doi: 10.4269/ajtmh.1990.43.25
44. Loiseau R, Nabet C, Simon S, Ginouves M, Brousse P, Blanchet D, et al. American Cutaneous leishmaniasis in French Guiana: An epidemiological update and study of environmental risk factors. Int J Dermatol (2019) 58(11):1323–8. doi: 10.1111/ijd.14625
45. Gramiccia M, Gradoni L. The current status of zoonotic leishmaniases and approaches to disease control. Int J Parasitol (2005) 35(11):1169–80. doi: 10.1016/j.ijpara.2005.07.001
46. Couppié P, Clyti E, Sainte-Marie D, Dedet JP, Carme B, Pradinaud R. Disseminated cutaneous leishmaniasis due to leishmania guyanensis: Case of a patient with 425 lesions. Am J Trop Med hygiene (2004) 71(5):558–60. doi: 10.4269/ajtmh.2004.71.558
47. Marsden P. Clinical presentations of leishmania braziliensis braziliensis. Parasitol Today (1985) 1(5):129–33. doi: 10.1016/0169-4758(85)90057-2
48. Simon S, Veron V, Carme B, Leishmania S. Identification by polymerase chain reaction-restriction fragment length polymorphism analysis and its applications in French Guiana. Diagn Microbiol Infect Dis (2010) 66(2):175–80. doi: 10.1016/j.diagmicrobio.2009.08.013
49. Rougeron V, Catzeflis F, Hide M, De Meeûs T, Bañuls AL. First clinical case of cutaneous leishmaniasis due to leishmania (Viannia) braziliensis in a domestic cat from French Guiana. Veterinary Parasitol (2011) 181(2-4):325–8. doi: 10.1016/j.vetpar.2011.04.028
50. Desjeux P, Dedet JP. Isoenzyme characterization of 112 leishmania isolates from French Guiana. Trans R Soc Trop Med Hygiene (1989) 83(5):610–2. doi: 10.1016/0035-9203(89)90373-8
51. Chaara D, Haouas N, Dedet JP, Babba H, Pratlong F. Leishmaniases in maghreb: An endemic neglected disease. Acta tropica (2014) 132:80–93. doi: 10.1016/j.actatropica.2013.12.018
52. Saïdi N, Galaï Y, Ben-Abid M, Boussoffara T, Ben-Sghaier I, Aoun K, et al. Imaging leishmania major antigens in experimentally infected macrophages and dermal scrapings from cutaneous leishmaniasis lesions in Tunisia. Microorganisms (2022) 10(6):1157.
53. Epelboin L, Abboud P, Abdelmoumen K, About F, Adenis A, Blaise T, et al. Panorama des pathologies infectieuses et non infectieuses de Guyane en 2022 [Overview of infectious and non-infectious diseases in French Guiana in 2022]. Med Trop Sante Int (2023) (1):mtsi.v3i1.2023.308. doi: 10.48327/mtsi.v3i1.2023.308
54. Malmontet T, Guarmit B, Gaillet M, Michaud C, Garceran N, Chanlin R, et al. Spectrum of skin diseases in Amerindian villages of the Upper Oyapock, French Guiana. Int J Dermatol (2020) 59(5):599–605. doi: 10.1111/ijd.14848
55. Henry K, Mayet A, Hernandez M, Frechard G, Blanc PA, Schmitt M, et al. Outbreak of Cutaneous Leishmaniasis among military personnel in French Guiana, 2020: Clinical, phylogenetic, individual and environmental aspects. PLOS Negl Trop Dis (2021) 15(11):e0009938. doi: 10.1371/journal.pntd.0009938
56. Valentin J, Niemetzky F, Gaillet M, Michaud C, Carbunar A, Demar M, et al. Spectrum of skin diseases in Maroon villages of the Maroni area, French Guiana. Int J Dermatol (2022) 61:1137–1144. doi: 10.1111/ijd.16324
57. Heleine M, Elenga N, Njuieyon F, Martin E, Piat C, Pansart C, et al. Using pentamidine to treat cutaneous leishmaniasis in children: a 10-year study in French Guiana. Clin Exp Dermatol (2023), llad146. doi: 10.1093/ced/llad146
58. Mokni M. [Cutaneous leishmaniasis]. Annales dermatologie venereologie (2019) 146(3):232–46. doi: 10.1016/j.annder.2019.02.002
59. Remadi L, Haouas N, Chaara D, Slama D, Chargui N, Dabghi R, et al. Clinical presentation of cutaneous leishmaniasis caused by leishmania major. Dermatol (Basel Switzerland) (2016) 232(6):752–9. doi: 10.1159/000456543
60. Masmoudi A, Hariz W, Marrekchi S, Amouri M, Turki H. Old world cutaneous leishmaniasis: Diagnosis and treatment. J Dermatol Case Rep (2013) 7(2):31. doi: 10.3315/jdcr.2013.1135
61. Masmoudi A, Ayadi N, Boudaya S, Meziou T, Mseddi M, Marrekchi S, et al. Clinical polymorphism of cutaneous leishmaniasis in centre and south of Tunisia. Bull la Societe pathologie exotique (1990) (2007) 100(1):36–40.
62. Masmoudi A, Kitar A, Rebai M, Bouassida S, Turki H, Zahaf A. [the cutaneous leishmaniasis of the face in gafsa area, Tunisia]. Bull la Societe pathologie exotique (1990) (2005) 98(5):374–9.
63. El Alem MMM, Hakkour M, Hmamouch A, Halhali M, Delouane B, Habbari K, et al. Risk factors and prediction analysis of cutaneous leishmaniasis due to leishmania tropica in southwestern Morocco. Infection Genet Evol (2018) 61:84–91. doi: 10.1016/j.meegid.2018.03.017
64. Baghad B, Razanapinaritra R, Maksouri H, El Bouri H, Outlioua A, Fellah H, et al. Possible introduction of leishmania tropica to urban areas determined by epidemiological and clinical profiles of patients with cutaneous leishmaniasis in Casablanca (Morocco). Parasite Epidemiol control (2020) 9:e00129. doi: 10.1016/j.parepi.2019.e00129
65. Chaara D, Ravel C, Bañuls A, Haouas N, Lami P, Talignani L, et al. Evolutionary history of leishmania killicki (Synonymous leishmania tropica) and taxonomic implications. Parasites Vectors (2015) 8:198. doi: 10.1186/s13071-015-0821-6
66. Dokhan MR, Jaouadi K, Salem S, Zenbil O, Gonzalez JP, Ben Salah A, et al. Natural infection of phlebotomus sergenti by leishmania tropica in Libya. Am J Trop Med hygiene (2018) 98(5):1339–42. doi: 10.4269/ajtmh.17-0814
67. Chiheb S, Guessous-Idrissi N, Hamdani A, Riyad M, Bichichi M, Hamdani S, et al. [Leishmania tropica cutaneous leishmaniasis in an emerging focus in north Morocco: New clinical forms]. Annales dermatologie venereologie (1999) 126(5):419–22.
68. Aoun K, Ben Abda I, Bousslimi N, Bettaieb J, Siala E, Ben Abdallah R, et al. Caractérisation comparative des trois formes de leishmaniose cutanée endémiques en tunisie. Annales dermatologie venereologie (2012) 139(6):452–8. doi: 10.1016/j.annder.2012.04.154
69. Hakkour M, El Alem MM, Hmamouch A, Rhalem A, Delouane B, Habbari K, et al. Leishmaniasis in northern Morocco: Predominance of leishmania infantum compared to leishmania tropica. BioMed Res Int (2019) 2019:5327287. doi: 10.1155/2019/5327287
70. del Giudice P, Marty P, Lacour JP, Perrin C, Pratlong F, Haas H, et al. Cutaneous leishmaniasis due to leishmania infantum. case reports and literature review. Arch Dermatol (1998) 134(2):193–8. doi: 10.1001/archderm.134.2.193
71. Meeting WECotCotL, Organization WH. Control of the leishmaniases: Report of a meeting of the who expert committee on the control of leishmaniases, Geneva, 22-26 march 2010. World Health Organization (2010).
72. Merino-Espinosa G, Corpas-López V, Díaz-Sáez V, Morillas-Márquez F, Tercedor-Sánchez J, Azaña-Defez JM, et al. Cutaneous leishmaniasis by leishmania infantum: Behind granulomatous lesions of unknown aetiology. J Eur Acad Dermatol Venereol (2018) 32(1):117–24. doi: 10.1111/jdv.14506
73. Guery R, Walker SL, Harms G, Neumayr A, Van Thiel P, Gangneux JP, et al. Clinical diversity and treatment results in tegumentary leishmaniasis: A European clinical report in 459 patients. PloS Negl Trop Dis (2021) 15(10):e0009863. doi: 10.1371/journal.pntd.0009863
74. Scorza BM, Carvalho EM, Wilson ME. Cutaneous manifestations of human and murine leishmaniasis. Int J Mol Sci (2017) 18(6):1296. doi: 10.3390/ijms18061296
75. Simon S, Nacher M, Carme B, Basurko C, Roger A, Adenis A, et al. Cutaneous leishmaniasis in French Guiana: Revising epidemiology with pcr-rflp. Trop Med Health (2017) 45:5. doi: 10.1186/s41182-017-0045-x
76. Ducharme O, Simon S, Ginouves M, Prévot G, Couppie P, Demar M, et al. Leishmania naiffi and lainsoni in French Guiana: Clinical features and phylogenetic variability. PloS Negl Trop Dis (2020) 14(8):e0008380. doi: 10.1371/journal.pntd.0008380
77. Meireles CB, Maia LC, Soares GC, Teodoro IPP, Gadelha M, da Silva CGL, et al. Atypical presentations of cutaneous leishmaniasis: A systematic review. Acta Trop (2017) 172:240–54. doi: 10.1016/j.actatropica.2017.05.022
78. Silveira FT, Lainson R, De Castro Gomes CM, Laurenti MD, Corbett CEP. Immunopathogenic competences of leishmania (V.) braziliensis and l. (L.) amazonensis in American cutaneous leishmaniasis. Parasite Immunol (2009) 31(8):423–31. doi: 10.1111/j.1365-3024.2009.01116.x
79. Rojas-Jaimes J, Rojas-Palomino N, Pence J, Lescano AG. Leishmania species in biopsies of patients with different clinical manifestations identified by high resolution melting and nested pcr in an endemic district in Peru. Parasite Epidemiol control (2019) 4:e00095–e. doi: 10.1016/j.parepi.2019.e00095
80. Santrich C, Segura I, Arias AL, Saravia NG. Mucosal disease caused by leishmania braziliensis guyanensis. Am J Trop Med hygiene (1990) 42(1):51–5. doi: 10.4269/ajtmh.1990.42.51
81. Naiff RD, Talhari S, Barrett TV. Isolation of leishmania guyanensis from lesions of the nasal mucosa. Memorias do Instituto Oswaldo Cruz (1988) 83(4):529–30. doi: 10.1590/s0074-02761988000400022
82. Marsden PD. Mucosal leishmaniasis (“Espundia” escomel, 1911). Trans R Soc Trop Med Hygiene (1986) 80(6):859–76. doi: 10.1016/0035-9203(86)90243-9
83. Borges AF, Gomes RS, Ribeiro-Dias F. Leishmania (Viannia) guyanensis in tegumentary leishmaniasis. Pathog Dis (2018) 76(4). doi: 10.1093/femspd/fty025
84. Costa J, Marsden P, Llanos-Cuentas E, Netto E, Carvalho E, Barral A, et al. Disseminated cutaneous leishmaniasis in a field clinic in bahia, Brazil: A report of eight cases. J Trop Med hygiene (1986) 89(6):319–23.
85. Llanos-Cuentas EA, Marsden PD, Lago EL, Barreto AC, Cuba CC, Johnson WD. Human mucocutaneous leishmaniasis in três braços, bahia-Brazil: An area of leishmania braziliensis braziliensis transmission. ii. cutaneous disease. presentation and evolution. Rev da Sociedade Bras Medicina Trop (1984) 17:169–77. doi: 10.1590/S0037-86821984000400003
86. Parise M, Pompeu M, Coehlo Filho J, Vasconcelos I, Lima J, Oliveira E, et al. Bubonic leishmaniasis: A common manifestation of leishmania (Viannia) braziliensis infection in ceara, Brazil. Am J Trop Med hygiene (1995) 53(4):380–5.
87. Romero GAS, de Farias Guerra MV, Paes MG, Macêdo V. Comparison of cutaneous leishmaniasis due to leishmania (Viannia) braziliensis and l. (V.) guyanensis in Brazil: Clinical findings and diagnostic approach. Clin Infect Dis (2001) 32(9):1304–12. doi: 10.1086/319990
88. Boussoffara T, Chelif S, Ben Ahmed M, Mokni M, Ben Salah A, Dellagi K, et al. Immunity against leishmania major infection: Parasite-specific granzyme b induction as a correlate of protection. Front Cell infection Microbiol (2018) 8:397. doi: 10.3389/fcimb.2018.00397
89. Louzir H, Melby PC, Ben Salah A, Marrakchi H, Aoun K, Ben Ismail R, et al. Immunologic determinants of disease evolution in localized cutaneous leishmaniasis due to leishmania major. J Infect Dis (1998) 177(6):1687–95. doi: 10.1086/515297
90. Boussoffara T, Boubaker MS, Ben Ahmed M, Mokni M, Guizani I, Ben Salah A, et al. Histological and immunological differences between zoonotic cutaneous leishmaniasis due to leishmania major and sporadic cutaneous leishmaniasis due to leishmania infantum. Parasite (Paris France) (2019) 26:9. doi: 10.1051/parasite/2019007
91. Misra P, Singh S. Site specific microbiome of leishmania parasite and its cross-talk with immune milieu. Immunol Lett (2019) 216:79–88. doi: 10.1016/j.imlet.2019.10.004
92. Dubie T, Mohammed Y. Review on the role of host immune response in protection and immunopathogenesis during cutaneous leishmaniasis infection. J Immunol Res (2020) 2020:2496713. doi: 10.1155/2020/2496713
93. Shamsi Meymandi S, Dabiri S, Eslammanesh T, Azadeh B, Nadji M, Shamsi Meymandi M, et al. Immunopathology of anthroponotic cutaneous leishmaniasis and incidental diagnostic tool of metastatic granuloma: A case-control study. Microbial pathogenesis (2021) 152:104654. doi: 10.1016/j.micpath.2020.104654
94. Wijesinghe H, Gunathilaka N, Semege S, Pathirana N, Manamperi N, de Silva C, et al. Histopathology of cutaneous leishmaniasis caused by leishmania donovani in Sri Lanka. BioMed Res Int (2020) 2020:4926819. doi: 10.1155/2020/4926819
95. Ribeiro-Gomes FL, Peters NC, Debrabant A, Sacks DL. Efficient capture of infected neutrophils by dendritic cells in the skin inhibits the early anti-leishmania response. PloS Pathog (2012) 8(2):e1002536. doi: 10.1371/journal.ppat.1002536
96. Chaves MM, Lee SH, Kamenyeva O, Ghosh K, Peters NC, Sacks D. The role of dermis resident macrophages and their interaction with neutrophils in the early establishment of leishmania major infection transmitted by sand fly bite. PloS Pathog (2020) 16(11):e1008674. doi: 10.1371/journal.ppat.1008674
97. Müller AJ, Filipe-Santos O, Eberl G, Aebischer T, Späth GF, Bousso P. Cd4+ T cells rely on a cytokine gradient to control intracellular pathogens beyond sites of antigen presentation. Immunity (2012) 37(1):147–57. doi: 10.1016/j.immuni.2012.05.015
98. Belkaid Y, Piccirillo CA, Mendez S, Shevach EM, Sacks DL. Cd4+Cd25+ regulatory T cells control leishmania major persistence and immunity. Nature (2002) 420(6915):502–7. doi: 10.1038/nature01152
99. Choi B-S, Kropf P. Evaluation of T cell responses in healing and nonhealing leishmaniasis reveals differences in T helper cell polarization ex vivo and in vitro. Parasite Immunol (2009) 31(4):199–209. doi: 10.1111/j.1365-3024.2009.01094.x
100. Babay BEC, Louzir H, Kebaïer C, Boubaker S, Dellagi K, Cazenave P-A. Inbred strains derived from feral mice reveal new pathogenic mechanisms of experimental leishmaniasis due to leishmania major. Infection Immun (2004) 72(8):4603–11. doi: 10.1128/IAI.72.8.4603-4611.2004
101. Formaglio P, Alabdullah M, Siokis A, Handschuh J, Sauerland I, Fu Y, et al. Nitric oxide controls proliferation of leishmania major by inhibiting the recruitment of permissive host cells. Immunity (2021) 54(12):2724–39.e10. doi: 10.1016/j.immuni.2021.09.021
102. Glennie ND, Volk SW, Scott P. Skin-resident Cd4+ T cells protect against leishmania major by recruiting and activating inflammatory monocytes. PloS Pathog (2017) 13(4):e1006349. doi: 10.1371/journal.ppat.1006349
103. Hohman LS, Mou Z, Carneiro MB, Ferland G, Kratofil RM, Kubes P, et al. Protective Cd4+ Th1 cell-mediated immunity is reliant upon execution of effector function prior to the establishment of the pathogen niche. PloS Pathog (2021) 17(9):e1009944. doi: 10.1371/journal.ppat.1009944
104. Remer KA, Roeger B, Hambrecht C, Moll H. Natural killer cells support the induction of protective immunity during dendritic cell-mediated vaccination against leishmania major. Immunology (2010) 131(4):570–82. doi: 10.1111/j.1365-2567.2010.03329.x
105. Trapani JA, Smyth MJ. Functional significance of the Perforin/Granzyme cell death pathway. Nat Rev Immunol (2002) 2(10):735–47. doi: 10.1038/nri911
106. Lazarski CA, Ford J, Katzman SD, Rosenberg AF, Fowell DJ. Il-4 attenuates Th1-associated chemokine expression and Th1 trafficking to inflamed tissues and limits pathogen clearance. PloS One (2013) 8(8):e71949. doi: 10.1371/journal.pone.0071949
107. McElrath MJ, Kaplan G, Nusrat A, Cohn ZA. Cutaneous leishmaniasis. the defect in T cell influx in Balb/C mice. J Exp Med (1987) 165(2):546–59. doi: 10.1084/jem.165.2.546
108. Nasseri M, Modabber FZ. Generalized infection and lack of delayed hypersensitivity in Balb/C mice infected with leishmania tropica major. Infection Immun (1979) 26(2):611–4. doi: 10.1128/iai.26.2.611-614.1979
109. Sadick MD, Heinzel FP, Holaday BJ, Pu RT, Dawkins RS, Locksley RM. Cure of murine leishmaniasis with anti-interleukin 4 monoclonal antibody. evidence for a T cell-dependent, interferon gamma-independent mechanism. J Exp Med (1990) 171(1):115–27. doi: 10.1084/jem.171.1.115
110. Suffia I, Reckling SK, Salay G, Belkaid Y. A role for Cd103 in the retention of Cd4+ Cd25+ TReg and control of leishmania major infection. J Immunol (2005) 174(9):5444–55. doi: 10.4049/jimmunol.174.9.5444
111. Hoseini SG, Javanmard SH, Zarkesh SH, Khamesipour A, Rafiei L, Karbalaie K, et al. Regulatory T-cell profile in early and late lesions of cutaneous leishmaniasis due to leishmania major. J Res Med Sci (2012) 17(6):513–8.
112. Antonelli LR, Dutra WO, Almeida RP, Bacellar O, Carvalho EM, Gollob KJ. Activated inflammatory T cells correlate with lesion size in human cutaneous leishmaniasis. Immunol Lett (2005) 101(2):226–30. doi: 10.1016/j.imlet.2005.06.004
113. Filipe-Santos O, Pescher P, Breart B, Lippuner C, Aebischer T, Glaichenhaus N, et al. A dynamic map of antigen recognition by Cd4 T cells at the site of leishmania major infection. Cell Host Microbe (2009) 6(1):23–33. doi: 10.1016/j.chom.2009.04.014
114. Brachelente C, Müller N, Doherr MG, Sattler U, Welle M. Cutaneous leishmaniasis in naturally infected dogs is associated with a T helper-2-Biased immune response. Veterinary Pathol (2005) 42(2):166–75. doi: 10.1354/vp.42-2-166
115. Araujo Flores GV, Sandoval Pacheco CM, Tomokane TY, Sosa Ochoa W, Zúniga Valeriano C, Castro Gomes CM, et al. Evaluation of regulatory immune response in skin lesions of patients affected by nonulcerated or atypical cutaneous leishmaniasis in Honduras, central America. Mediators Inflammation (2018) 2018:3487591. doi: 10.1155/2018/3487591
116. Abbehusen MMC, Cunha J, Suarez MS, Teixeira C, Almeida VDA, Pereira LDS, et al. Immunization of experimental dogs with salivary proteins from lutzomyia longipalpis, using DNA and recombinant canarypox virus induces immune responses consistent with protection against leishmania infantum. Front Immunol (2018) 9:2558. doi: 10.3389/fimmu.2018.02558
117. Sher A, Sacks DL, Scott PA. Host and parasite factors influencing the expression of cutaneous leishmaniasis. Ciba Found Symp (1983) 99:174–89. doi: 10.1002/9780470720806.ch10
118. Maksouri H, Dang PM-C, Rodrigues V, Estaquier J, Riyad M, Akarid K. Moroccan Strains of leishmania major and leishmania tropica differentially impact on nitric oxide production by macrophages. Parasites Vectors (2017) 10(1):506. doi: 10.1186/s13071-017-2401-4
119. Ajdary S, Riazi-Rad F, Alimohammadian MH, Pakzad SR. Immune response to leishmania antigen in anthroponotic cutaneous leishmaniasis. J infection (2009) 59(2):139–43. doi: 10.1016/j.jinf.2009.05.010
120. Kumar R, Bumb RA, Salotra P. Correlation of parasitic load with interleukin-4 response in patients with cutaneous leishmaniasis due to leishmania tropica. FEMS Immunol Med Microbiol (2009) 57(3):239–46. doi: 10.1111/j.1574-695X.2009.00607.x
121. Katara GK, Raj A, Kumar R, Avishek K, Kaushal H, Ansari NA, et al. Analysis of localized immune responses reveals presence of Th17 and treg cells in cutaneous leishmaniasis due to leishmania tropica. BMC Immunol (2013) 14:52. doi: 10.1186/1471-2172-14-52
122. Bastien P, Killick-Kendrick R. Leishmania tropica infection in hamsters and a review of the animal pathogenicity of this species. Exp Parasitol (1992) 75(4):433–41. doi: 10.1016/0014-4894(92)90256-a
123. Carvalho LP, Passos S, Schriefer A, Carvalho EM. Protective and pathologic immune responses in human tegumentary leishmaniasis. Front Immunol (2012) 3:301. doi: 10.3389/fimmu.2012.00301
124. Santos Cda S, Boaventura V, Ribeiro Cardoso C, Tavares N, Lordelo MJ, Noronha A, et al. Cd8(+) granzyme b(+)-mediated tissue injury vs. Cd4(+)Ifnγ(+)-mediated parasite killing in human cutaneous leishmaniasis. J Invest Dermatol (2013) 133(6):1533–40. doi: 10.1038/jid.2013.4
125. Faria DR, Gollob KJ, Barbosa J Jr., Schriefer A, Machado PR, Lessa H, et al. Decreased in situ expression of interleukin-10 receptor is correlated with the exacerbated inflammatory and cytotoxic responses observed in mucosal leishmaniasis. Infection Immun (2005) 73(12):7853–9. doi: 10.1128/iai.73.12.7853-7859.2005
126. Carvalho LP, Passos S, Bacellar O, Lessa M, Almeida RP, Magalhães A, et al. Differential immune regulation of activated T cells between cutaneous and mucosal leishmaniasis as a model for pathogenesis. Parasite Immunol (2007) 29(5):251–8. doi: 10.1111/j.1365-3024.2007.00940.x
127. Gollob KJ, Antonelli LR, Faria DR, Keesen TS, Dutra WO. Immunoregulatory mechanisms and Cd4-Cd8- (Double negative) T cell subpopulations in human cutaneous leishmaniasis: A balancing act between protection and pathology. Int Immunopharmacol (2008) 8(10):1338–43. doi: 10.1016/j.intimp.2008.03.016
128. Da-Cruz AM, de Oliveira MP, De Luca PM, Mendonça SC, Coutinho SG. Tumor necrosis factor-alpha in human American tegumentary leishmaniasis. Memorias do Instituto Oswaldo Cruz (1996) 91(2):225–9. doi: 10.1590/s0074-02761996000200019
129. Miranda Lessa M, Andrade Lessa H, Castro TWN, Oliveira A, Scherifer A, Machado P, et al. Mucosal leishmaniasis: Epidemiological and clinical aspects. Braz J otorhinolaryngol (2007) 73(6):843–7. doi: 10.1016/s1808-8694(15)31181-2
130. Lessa HA, Machado P, Lima F, Cruz AA, Bacellar O, Guerreiro J, et al. Successful treatment of refractory mucosal leishmaniasis with pentoxifylline plus antimony. Am J Trop Med hygiene (2001) 65(2):87–9. doi: 10.4269/ajtmh.2001.65.87
131. Cardoso TM, Machado Á, Costa DL, Carvalho LP, Queiroz A, Machado P, et al. Protective and pathological functions of Cd8+ T cells in leishmania braziliensis infection. Infection Immun (2015) 83(3):898–906. doi: 10.1128/IAI.02404-14
132. Melby PC, Andrade-Narvaez FJ, Darnell BJ, Valencia-Pacheco G, Tryon VV, Palomo-Cetina A. Increased expression of proinflammatory cytokines in chronic lesions of human cutaneous leishmaniasis. Infection Immun (1994) 62(3):837–42. doi: 10.1128/iai.62.3.837-842.1994
133. Bacellar O, Lessa H, Schriefer A, Machado P, Ribeiro de Jesus A, Dutra WO, et al. Up-regulation of Th1-type responses in mucosal leishmaniasis patients. Infection Immun (2002) 70(12):6734–40. doi: 10.1128/iai.70.12.6734-6740.2002
134. Covre LP, Martins RF, Devine OP, Chambers ES, Vukmanovic-Stejic M, Silva JA, et al. Circulating senescent T cells are linked to systemic inflammation and lesion size during human cutaneous leishmaniasis. Front Immunol (2019) 9:3001. doi: 10.3389/fimmu.2018.03001
135. Covre LP, Devine OP, Garcia de Moura R, Vukmanovic-Stejic M, Dietze R, Ribeiro-Rodrigues R, et al. Compartmentalized cytotoxic immune response leads to distinct pathogenic roles of natural killer and senescent Cd8(+) T cells in human cutaneous leishmaniasis. Immunology (2020) 159(4):429–40. doi: 10.1111/imm.13173
136. Fantecelle CH, Covre LP, Garcia de Moura R, Guedes HLM, Amorim CF, Scott P, et al. Transcriptomic landscape of skin lesions in cutaneous leishmaniasis reveals a strong Cd8(+) T cell immunosenescence signature linked to immunopathology. Immunology (2021) 164(4):754–65. doi: 10.1111/imm.13410
137. Milling S. Ageing dangerously; homing of senescent Cd8 T cells in cutaneous leishmaniasis. Immunology (2020) 159(4):355–6. doi: 10.1111/imm.13188
138. Campanelli AP, Roselino AM, Cavassani KA, Pereira MS, Mortara RA, Brodskyn CI, et al. Cd4+Cd25+ T cells in skin lesions of patients with cutaneous leishmaniasis exhibit phenotypic and functional characteristics of natural regulatory T cells. J Infect Dis (2006) 193(9):1313–22. doi: 10.1086/502980
139. Punnonen J, Aversa G, Cocks BG, McKenzie AN, Menon S, Zurawski G, et al. Interleukin 13 induces interleukin 4-independent Igg4 and ige synthesis and Cd23 expression by human b cells. Proc Natl Acad Sci (1993) 90(8):3730–4. doi: 10.1073/pnas.90.8.3730
140. Bourreau E, Prévot G, Pradinaud R, Launois P. Interleukin (Il)–13 is the predominant Th2 cytokine in localized cutaneous leishmaniasis lesions and renders specific Cd4+ T cells unresponsive to il-12. J Infect Dis (2001) 183(6):953–9. doi: 10.1086/319249
141. Bourreau E, Ronet C, Darcissac E, Lise MC, Marie DS, Clity E, et al. Intralesional regulatory T-cell suppressive function during human acute and chronic cutaneous leishmaniasis due to leishmania guyanensis. Infection Immun (2009) 77(4):1465–74. doi: 10.1128/IAI.01398-08
142. Bourreau E, Ronet C, Darsissac E, Lise M-C, Sainte Marie D, Clity E, et al. In leishmaniasis due to leishmania guyanensis infection, distinct intralesional interleukin-10 and Foxp3 mrna expression are associated with unresponsiveness to treatment. J Infect Dis (2009) 199(4):576–9. doi: 10.1086/596508
143. Ives A, Ronet C, Prevel F, Ruzzante G, Fuertes-Marraco S, Schutz F, et al. Leishmania rna virus controls the severity of mucocutaneous leishmaniasis. Science (2011) 331(6018):775–8. doi: 10.1126/science.1199326
144. Hartley M-A, Ronet C, Zangger H, Beverley S, Fasel N. Leishmania rna virus: When the host pays the toll. Front Cell Infection Microbiol (2012) 2:99. doi: 10.3389/fcimb.2012.00099
145. Eren RO, Reverte M, Rossi M, Hartley MA, Castiglioni P, Prevel F, et al. Mammalian innate immune response to a leishmania-resident rna virus increases macrophage survival to promote parasite persistence. Cell Host Microbe (2016) 20(3):318–28. doi: 10.1016/j.chom.2016.08.001
146. Ginouvès M, Simon S, Bourreau E, Lacoste V, Ronet C, Couppié P, et al. Prevalence and distribution of leishmania rna virus 1 in leishmania parasites from French Guiana. Am J Trop Med hygiene (2016) 94(1):102–6. doi: 10.4269/ajtmh.15-0419
147. Zabala-Peñafiel A, Fantinatti M, Dias-Lopes G, da Silva JL, Miranda LFC, Lyra MR, et al. First report of leishmania rna virus 1 in leishmania (Viannia) braziliensis clinical isolates from Rio de Janeiro state - Brazil. Memorias do Instituto Oswaldo Cruz (2022) 117:e210107. doi: 10.1590/0074-02760210107
148. Hartley MA, Bourreau E, Rossi M, Castiglioni P, Eren RO, Prevel F, et al. Leishmaniavirus-dependent metastatic leishmaniasis is prevented by blocking il-17a. PloS Pathog (2016) 12(9):e1005852. doi: 10.1371/journal.ppat.1005852
149. Ginouvès M, Couppié P, Simon S, Bourreau E, Rogier S, Brousse P, et al. Leishmaniavirus genetic diversity is not related to leishmaniasis treatment failure. Clin Microbiol infection (2021) 27(2):286.e1–.e5. doi: 10.1016/j.cmi.2020.04.037
Keywords: L. major, L. infantum, L. tropica, L. guyanensis, L. braziliensis, cutaneous leishmaniasis (CL), clinical manifestation, local immune response
Citation: Saidi N, Blaizot R, Prévot G, Aoun K, Demar M, Cazenave PA, Bouratbine A and Pied S (2023) Clinical and immunological spectra of human cutaneous leishmaniasis in North Africa and French Guiana. Front. Immunol. 14:1134020. doi: 10.3389/fimmu.2023.1134020
Received: 29 December 2022; Accepted: 10 March 2023;
Published: 27 July 2023.
Edited by:
Denis Sereno, Institut de Recherche Pour le Développement (IRD), FranceReviewed by:
Fernanda Nazaré Morgado, Oswaldo Cruz Institute, BrazilCopyright © 2023 Saidi, Blaizot, Prévot, Aoun, Demar, Cazenave, Bouratbine and Pied. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sylviane Pied, c3lsdmlhbmUucGllZEBwYXN0ZXVyLWxpbGxlLmZy
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.