
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Immunol., 29 March 2023
Sec. Inflammation
Volume 14 - 2023 | https://doi.org/10.3389/fimmu.2023.1101854
This article is part of the Research TopicImmune Imbalance in Obesity-associated DiseasesView all 12 articles
 Yisheng Cai1†
Yisheng Cai1† Xuemei Zuo1†Yuyang Zuo1Shuang Wu1Weiwei Pang1Keqiang Ma1Qiaorong Yi1Lijun Tan1
Xuemei Zuo1†Yuyang Zuo1Shuang Wu1Weiwei Pang1Keqiang Ma1Qiaorong Yi1Lijun Tan1 Hongwen Deng2
Hongwen Deng2 Xiaochao Qu1*
Xiaochao Qu1* Xiangding Chen1*
Xiangding Chen1*Background: Both obesity (OB) and periodontitis (PD) are chronic non-communicable diseases, and numerous epidemiological studies have demonstrated the association between these two diseases. However, the molecular mechanisms that could explain the association between OB and PD are largely unclear. This study aims to investigate the common gene signatures and biological pathways in OB and PD through bioinformatics analysis of publicly available transcriptome datasets.
Methods: The RNA expression profile datasets of OB (GSE104815) and PD (GSE106090) were used as training data, and GSE152991 and GSE16134 as validation data. After screening for differentially expressed genes (DEGs) shared by OB and PD, gene enrichment analysis, protein-protein interaction (PPI) network construction, GeneMANIA analysis, immune infiltration analysis and gene set enrichment analysis (GSEA) were performed. In addition, receiver operating characteristic (ROC) curves were used to assess the predictive accuracy of the hub gene. Finally, we constructed the hub gene-associated TF-miRNA-mRNA regulatory network.
Results: We identified a total of 147 DEGs shared by OB and PD (38 down-regulated and 109 up-regulated). Functional analysis showed that these genes were mainly enriched in immune-related pathways such as B cell receptor signalling, leukocyte migration and cellular defence responses. 14 hub genes (FGR, MNDA, NCF2, FYB1, EVI2B, LY86, IGSF6, CTSS, CXCR4, LCK, FCN1, CXCL2, P2RY13, MMP7) showed high sensitivity and specificity in the ROC curve analysis. The results of immune infiltration analysis showed that immune cells such as macrophages, activated CD4 T cells and immune B cells were present at high infiltration levels in both OB and PD samples.The results of GeneMANIA analysis and GSEA analysis suggested that five key genes (FGR, LCK, FYB1, LY86 and P2RY13) may be strongly associated with macrophages. Finally, we constructed a TF-miRNA-mRNA regulatory network consisting of 233 transcription factors (TFs), 8 miRNAs and 14 mRNAs based on the validated information obtained from the database.
Conclusions: Five key genes (FGR, LCK, FYB1, LY86, P2RY13) may be important biomarkers of OB and PD. These genes may play an important role in the pathogenesis of OB and PD by affecting macrophage activity and participating in immune regulation and inflammatory responses.
Obesity (OB) is a complex, multifactorial chronic inflammatory disease characterized by abnormal or excessive deposition of fat in adipose tissue (1). It is also a major risk factor for many diseases, including type 2 diabetes, cardiovascular disease, osteoarthritis and certain cancers (2). The prevalence of OB has tripled in the last few decades (3). The number of people with OB worldwide was reported to be as high as 671 million (12% of the world’s adult population) in 2016 (4). Periodontitis (PD) is one of the most common chronic multifactorial inflammatory diseases affecting the global population, leading to loss of connective tissue attachment, alveolar bone erosion, tooth loss and systemic inflammation (5, 6). There is evidence that OB increases susceptibility to PD (7). An earlier study reporting the association between OB and PD found changes in periodontal tissue in addition to greater alveolar bone resorption in obese rats compared to non-obese rats (8). Several recent studies have also suggested a comorbid effect between OB and PD (9, 10). OB increased the risk of PD by two to three times and was independent of traditional risk factors, including smoking, age, and gender (11). Animal studies have indicated that an increased alveolar bone loss in obese animals with PD and significantly greater alveolar bone loss in obese rats than in lean controls (12, 13). In addition, obese individuals who consume an excessively high-fat diet have an enhanced metabolic response to PD and show a metabolic susceptibility to increased periodontal destruction (14). These findings highlighted the existence of an association between OB and PD. However, the molecular mechanism of this association is still unknown. Therefore, exploring the common genetic features of OB and PD and their potential molecular mechanisms holds great promise for the diagnosis and treatment of OB and PD co-morbidities.
A growing body of clinical and experimental evidence suggests that immune cell infiltration and inflammatory factors play a critical role in the development of OB or PD (2, 14). On the one hand, mouse models of OB and diabetes were found to be characterized by impaired T and B lymphocyte-mediated immune responses (2). A recent study reported that natural killer T cells are regulators of adipose tissue inflammation in OB (15). Osborn O et al. (16) pointed that in the obesity-induced inflammatory response, immune cells are recruited and cause adipose tissue inflammation. Monocytes receive chemotactic signals and translocate into adipose tissue, polarizing it to a highly pro-inflammatory M1-like state. Once recruited, these M1-like macrophages secrete pro-inflammatory cytokines and act in a paracrine manner (16). In addition, OB induces a shift in the adipose tissue T-cell population, with decreased Treg content and increased CD4+ TH1 and CD8+ effector T cells that secrete pro-inflammatory cytokines (15). Recent studies have also indicated that increased B-cell numbers can promote T-cell activation and enhance M1-like macrophage polarization, inflammation, and insulin resistance (17). Notably, cytokines and chemokines from adipose tissue can also be released into the circulation and promote inflammation in other tissues in an endocrine manner (18). Meanwhile, T cells in adipose tissue are thought to play a role in obesity-induced inflammation by altering the number of adipose tissue macrophages and their activation status (19, 20). Nishimura et al. (21) showed that CD8+ T cells were increased in obese adipose tissue and promote the recruitment and activation of adipose tissue macrophages (21). On the other hand, Dutzan N et al. (22) revealed that IL-21, IL-1β, IL-17 and IL-23p19 were significantly overexpressed in periodontal disease tissues compared to healthy gingival tissues. In particular, IL-21 was overexpressed in chronic periodontitis gingival tissues and was associated with pro-inflammatory cytokines for periodontal destruction (22). IL-10 and TGF-β1 expression were down-regulated in periodontal lesions and may be regulators of inflammation and alveolar bone loss in periodontal disease (23). In addition, some researchers suggested that one of the mechanisms related to PD and OB is the increase in the production of inflammatory cytokines, i.e., OB leads to an increase in the inflammatory stimulation of adipose tissue, adipocytes secrete adipocytokines, which increases the release of inflammatory cytokines, thus leading to the imbalance between the reduction of anti-inflammatory mechanisms and persistent low-grade inflammation (24). For example, in humans, plasma levels of tumor necrosis factor α, interleukin-6 and C-reactive protein are strongly associated with OB (25). In a mouse model of OB/type 2 diabetes, resolvin E1 increased neutrophil phagocytosis in wild-type mice with Pseudomonas gingivalis, but had no effect on type 2 diabetic mice (26). Therefore, we speculate that immune factors and inflammatory responses may be one of the important reasons for the occurrence of OB and PD.
A number of previous studies have explored the potential impact of OB on the pathogenesis and progression of PD, highlighting the importance of common inflammatory processes and immune dysfunction, but the immune-related mechanisms involved in OB and PD remain to be elucidated. The aim of this study was to explore the molecular link between OB and PD using publicly available transcriptomic data. We used an integrated bioinformatics approach to study immune cell infiltration, reveal molecular regulatory networks, and explore the molecular mechanisms underlying the interaction of disease onset, hoping to provide new perspectives on the biological mechanisms of obesity-associated periodontitis.
We searched the Gene Expression Omnibus (GEO) database for gene expression profiles of obesity and periodontitis using the keywords “periodontitis” and “obesity”. Four data sets met the inclusion criteria: (1) the experimental data type was microarray or high-throughput sequencing; (2) the number of samples per cohort was greater than six; (3) the study samples were from humans. We used the GSE104815 and GSE106090 datasets as the discovery cohort for transcriptome analysis, and the GSE152991 and GSE16134 datasets as the validation cohort. For OB, the GSE104815 dataset contained 4 OB samples and 4 non-obese samples; the GSE152991 dataset contained 34 OB samples and 11 control samples. For PD, the GSE106090 dataset contained 18 samples, of which 6 PD samples and 6 healthy samples were selected, while 6 peri-implantitis samples were excluded; the GSE16134 dataset contained 241 PD samples and 69 control samples. For the samples in these datasets, we excluded the effects of medical history and medication, as these effects may introduce bias in our study. Finally, we used the limma/DEseq2 package to filter, log2 transform, and normalize all datasets. Among these, probes were annotated as gene symbols, and for genes matching multiple probes, the probe with the highest expression value was retained. Details of the platforms, experiment types and tissues of the four datasets are shown in Supplementary Table 1.
The empirical Bayesian approach in the limma package (27) in R was applied to screen the GSE106090 and GSE104815 datasets. Significant differentially expressed genes (DEGs) were identified based on the cutoff criteria of |log2FoldChange| ≥1 and adjusted p-value < 0.05, and the common DEGs between OB and PD were obtained by the intersection of the plotted Venn diagram using the ggVennDiagram (28) package in R.
To further reveal the functions of the common DEGs, Gene Ontology (GO) annotation and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis were performed. The biological properties of the DEGs were annotated as molecular function (MF), biological process (BP) and cellular component (CC) by GO enrichment analysis using the clusterProfiler (29) package in R, and the KEGG analysis was performed using the KOBAS (30) online tool. An adjusted p-value < 0.05 was set as the cut-off criterion.
Based on the common DEGs in PD and OB, the STRING (search tool for the retrieval of interacting genes) database (https://string-db.org/) was used to construct the PPI network, whose confidence score was set to the middle value (confidence score ≥ 0.4), and then Cytoscape software (31) was used to visualize the PPI network. The MCODE plugin (32) of Cytoscape was applied for module analysis to identify the key gene clusters, and the CytoHubba plugin (33) was used to identify the hub genes, through which four methods including maximal clique centrality (MCC), density of maximum neighborhood component (DMNC), degree and maximum neighborhood component (MNC) (34) were applied to identify the top 30 hub genes in the PPI network, respectively, and the genes obtained by taking the intersection of four gene lists were considered as hub genes.
The expression matrics of GSE16134 and GSE152991 were downloaded from the GEO database and the GREIN platform (http://www.ilincs.org/apps/grein/) (35), respectively, which were used to validate the expression levels of the hub genes. GSE16134 contains 241 PD and 69 control samples, and GSE152991 contains 34 OB and 11 control samples. Considering that our data do not conform to a normal distribution, the Wilcoxon test was used to compare the two groups in the two datasets defining a p-value < 0.05 as significant. Meanwhile, the pROC package (36) in R was used to plot receiver operating characteristic (ROC) curves to verify the validity and predictive accuracy of the hub genes. The hub genes with an area under the curve (AUC) > 0.7 were considered useful for disease diagnosis (37).
We used the GeneMANIA online website (38) to analyze the hub genes and their co-expressed gene network by entering the validated hub genes directly into the GeneMANIA website for query, and GeneMANIA will find functionally similar genes using a wealth of genomics and proteomics data, while also reporting the weights of the predicted values for each gene used for query, and the results will show in which biological terms and pathways the genes are enriched.
Immune infiltration analysis was performed using the ssGSEA (single sample gene set enrichment analysis) algorithm (39) for GSE15299 (OB) and GSE16134 (PD), respectively. The ssGSEA algorithm is an extension of the gene set enrichment analysis (GSEA) method (40), it is a ranking based method that defines a score indicating the absolute enrichment of a specific gene set in each sample (41, 42). ssGSEA scores can be used to quantify the relative abundance of immune cells in OB or PD tissues and determine the level of immune infiltration in each sample.
The immune infiltration gene set was downloaded from http://cis.hku.hk/TISIDB/ and the immune infiltration analysis was performed using the GSVA package in R. To reveal the relationship between the hub genes and the immune cells, we then performed the correlation analysis between the hub genes and the immune cells based on Spearman’s correlation coefficients.
To further explore the correlation between immune cell infiltration and hub genes, we performed GSEA analysis. Gene sets of pathways were obtained from the Molecular Signature Database (MSigDB) (https://www.gsea-msigdb.org/gsea/msigdb/). Based on the gene expression profile data of GSE152991 (OB) and GSE16134 (PD), the average expression value of each hub gene was calculated separately, and all samples in the dataset were divided into “high” and “low” groups according to whether the expression value of the gene was higher or lower than its average expression value, and the GSEA method was used to evaluate the relevant molecular mechanisms between the two groups. Terms with p-value < 0.05, |normalized enrichment score (NES)| > 1, and false positive rate (FDR) p-value < 0.25 were considered to be significant.
Transcription factors (TFs) are proteins that can bind to specific DNA sequences and regulate gene expression (43). MicroRNAs (miRNAs) are a class of endogenous short non-coding RNAs that mediate the degradation of target mRNAs or repress translation (44). TFs and miRNAs mostly act in a combinatorial manner, where many different TFs or miRNAs control the same gene, i.e., they act synergistically on the target mRNAs (45). Moreover, TFs and miRNAs can not only co-regulate the expression of target genes but also regulate each other (46). Therefore, it is helpful to learn about the dysregulation of gene expression in various physiological and disease conditions through the transcriptional regulatory network among TFs, miRNAs, and mRNAs. The Human microRNA Disease Database (HMDD) (47) is a database for manually collecting and organizing disease-associated miRNAs, and the association information is experimentally validated. We downloaded the miRNAs associated with OB and PD from the HMDD database and obtained the co-miRNAs of OB and PD by mapping. The multiMiR package (48) provides the target genes regulated by miRNAs, which are also experimentally validated. We used the multiMiR package to find genes with possible regulatory relationships with co-miRNAs of OB and PD. The TransmiR database (49) collected TF-miRNA regulatory relationships in publications, from which co-miRNA-related TFs of OB and PD were obtained. Finally, a TF-miRNA-mRNA network was constructed and visualized in Cytoscape.
A total of 875 DEGs were identified in the OB dataset using the limma package, of which 440 genes were up-regulated and 435 genes were down-regulated (Figure 1A), and 2399 DEGs were obtained in the PD dataset, of which 1336 genes were up-regulated and 1063 genes were down-regulated (Figure 1B). After taking the intersection of the DEGs in OB and PD datasets, a total of 147 overlapping DEGs were identified, including 109 commonly up-regulated genes and 38 commonly down-regulated genes (Figure 1C). Heatmaps of the overlapping DEGs in OB and PD are shown in Supplementary Figure 1.
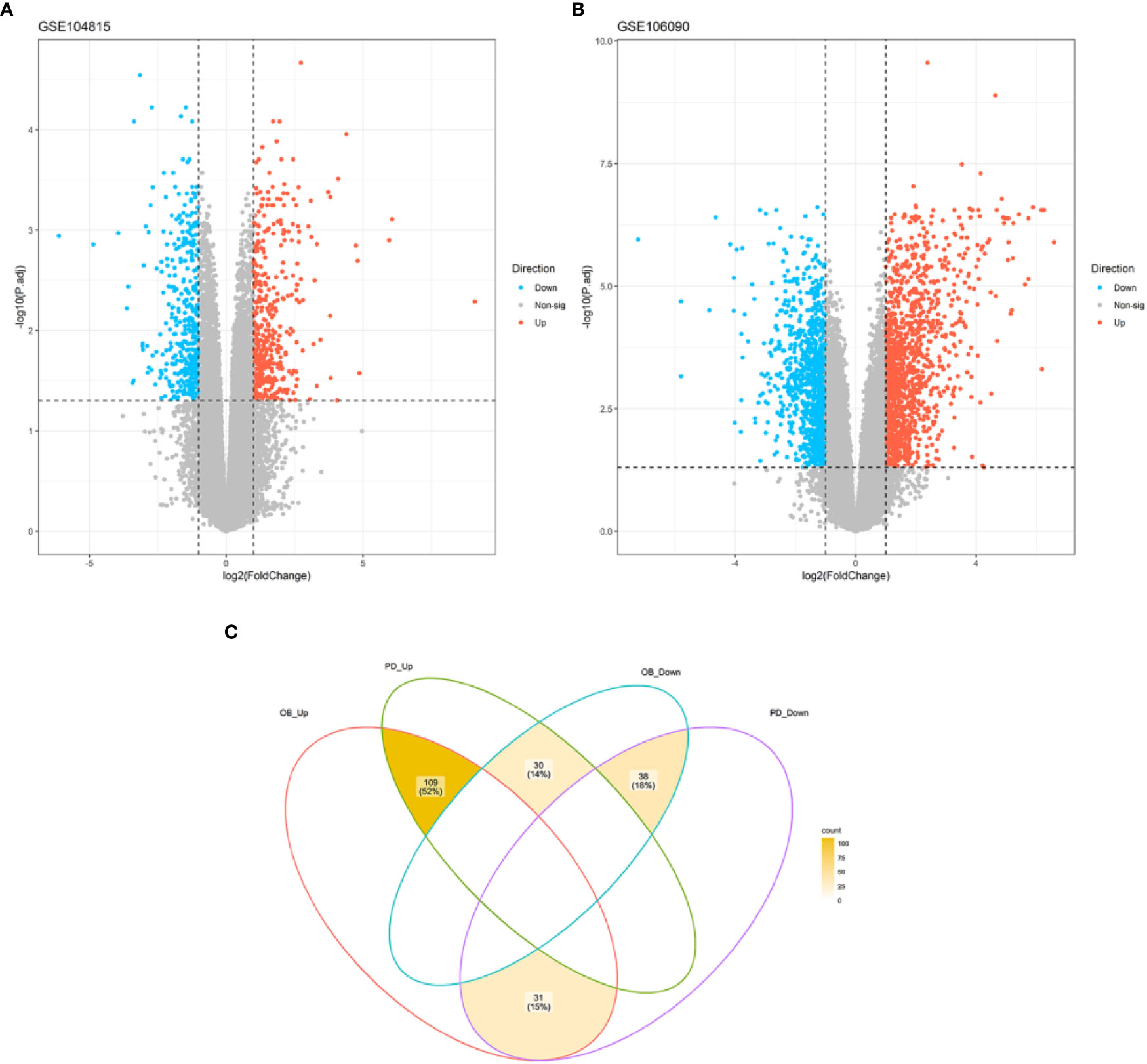
Figure 1 Volcano plot of DEGs and Venn diagram of common DEGs. (A) A total of 667 DEGs were identified between OB and healthy controls. (B) A total of 2191 DEGs were identified between PD and healthy controls. (C) A total of 109 common up-regulated genes and 38 common down-regulated genes were identified in OB and PD. DEGs, differentially expressed genes; OB, Obesity; PD, Periodontitis.
To investigate the potential biological processes and pathways of the DEGs, we separately performed GO and KEGG enrichment analysis using the clusterProfiler package and the KOBAS website. The results of KEGG enrichment analysis demonstrated that these genes were mainly enriched in “osteoclast differentiation”, “B cell receptor signaling pathway” and “viral protein interaction with cytokine and cytokine receptor” (Figure 2A). The results of GO analysis revealed that biological processes such as “leukocyte migration”, “cellular chemotaxis” and “cellular defense response” were significantly enriched (Figure 2B). In terms of cellular composition, terms such as “NADPH oxidase complex”, “secondary lysosome” and “high-density lipoprotein particle” were significantly enriched (Figure 2C). In terms of molecular function, terms such as “superoxide-generating NADPH oxidase activator activity”, “inhibitory MHC class I receptor activity” and “MHC class I receptor activity” were significantly enriched (Figure 2D). The detailed results of the GO and KEGG enrichment analysis are shown in Supplementary Tables 2, 3.
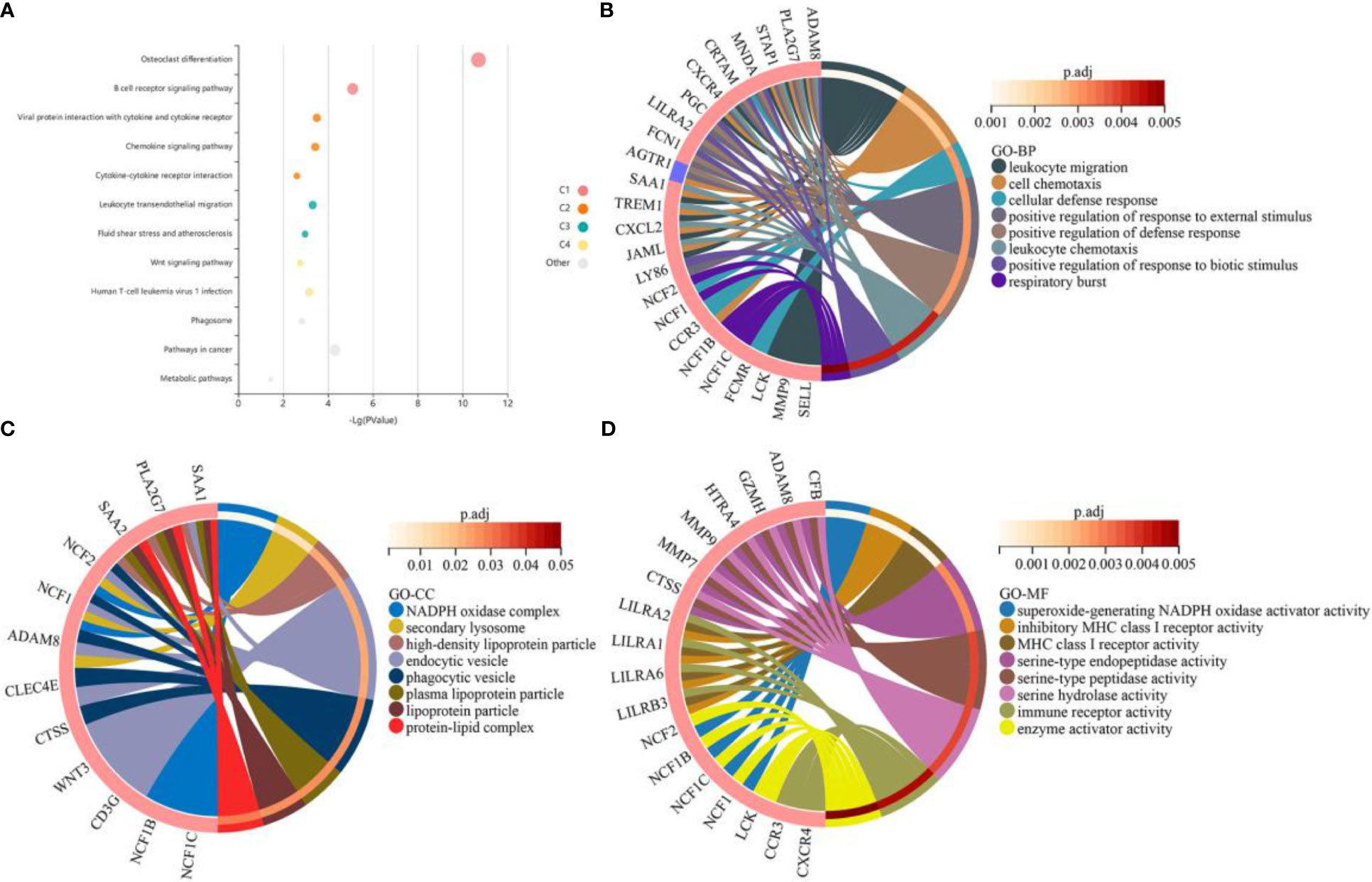
Figure 2 Functional enrichment analysis of the common differentially expressed genes (DEGs) between obesity and periodontitis. (A) KEGG pathway analysis of the DEGs. Where each bubble represents an enriched function, the size of the bubble represents 6 levels of enriched p-values, and the color of the bubble indicates the clustering of different pathways corresponding to different clusters (C1, C2, C3, etc.). (B–D) GO enrichment results of the DEGs for the categories of biological processes, cellular composition and molecular function. The red and blue colors of the gene represent up-regulation and down-regulation respectively. An adjusted p-value < 0.05 was considered statistically significant. DEGs, differentially expressed genes. KEGG, Kyoto Encyclopedia of Genes and Genomes. GO, Gene Ontology.
To further reveal the potential relationships among the common DEGs in OB and PD, a protein-protein interaction (PPI) network of these genes was constructed in the STRING database, which contained 72 nodes and 182 edges (Figure 3A). Module analysis was performed using the MCODE plugin in Cytoscape to detect key clustering modules, from which three modules were obtained, and module 1 contained 9 nodes and 17 edges with a cluster score of 4.25; module 2 contained 6 nodes and 10 edges with a score of 4, and module 3 contained 6 nodes and 8 edges with a score of 3.20 (Figure 3B).
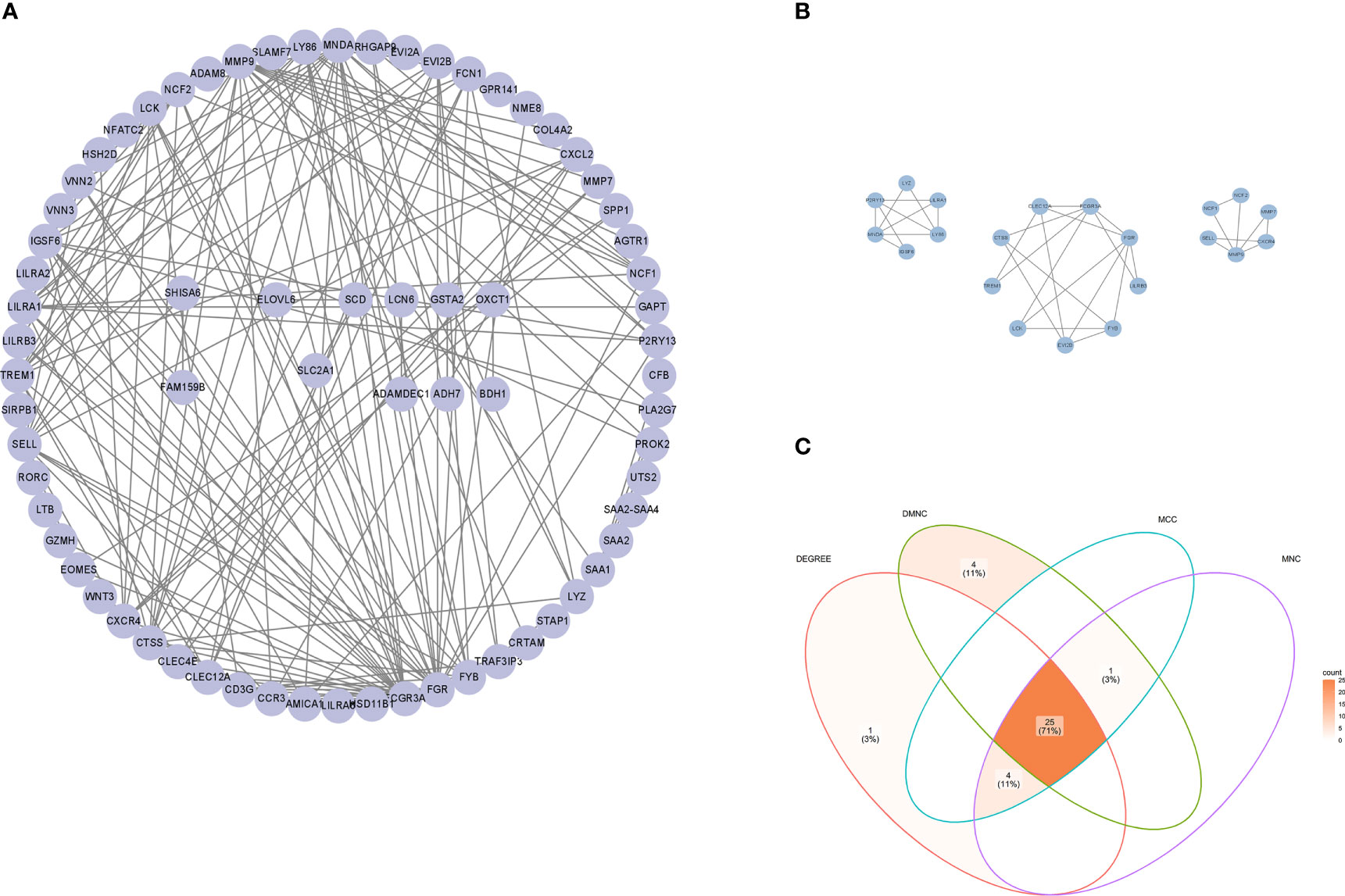
Figure 3 PPI network of hub genes. (A) PPI network of common DEGs constructed using the STRING database. (B) Three gene modules were identified by the MCODE plugin. (C) Venn diagram showing 25 common hub genes identified by MCC, MNC, Degree and DMNC algorithms using the CytoHubba plugin. PPI, protein-protein interaction; DEGs, differentially expressed genes; MCC, maximal clique centrality; DMNC, density of maximum neighborhood component; MNC, maximum neighborhood component.
To explore genes that may play an important role in the co-occurrence of OB and PD, the CytoHubba plugin was used to identify hub genes. Due to the heterogeneity of biological networks, several topological analysis algorithms including MCC, MNC, Degree and DMNC were applied in our research, and the top 30 most important hub genes in the PPI network were obtained. The intersection of four hub gene lists revealed 25 hub genes: FCGR3A, FGR, MNDA, SELL, NCF2, FYB1, EVI2B, LY86, TREM1, LILRA1, IGSF6, CTSS, CXCR4, LCK, CLEC12A, FCN1, CXCL2, VNN2 P2RY13, LYZ, CCR3, EOMES, MMP7, CD3G and CLEC4E (Figure 3C). Details of the hub genes are shown in Supplementary Table 4.
The 25 hub genes of OB and PD were validated using GSE152991 (for OB) and GSE16134 (for PD) datasets, respectively. The results demonstrated that 14 hub genes were significantly differentially expressed between the case and control groups in these two datasets (p-value < 0.05), all of which were found to be up-regulated in both OB and PD groups. The hub genes were FGR, MNDA, NCF2, FYB1, EVI2B, LY86, IGSF6, CTSS, CXCR4, LCK, FCN1, CXCL2, P2RY13 and MMP7 (Figure 4).
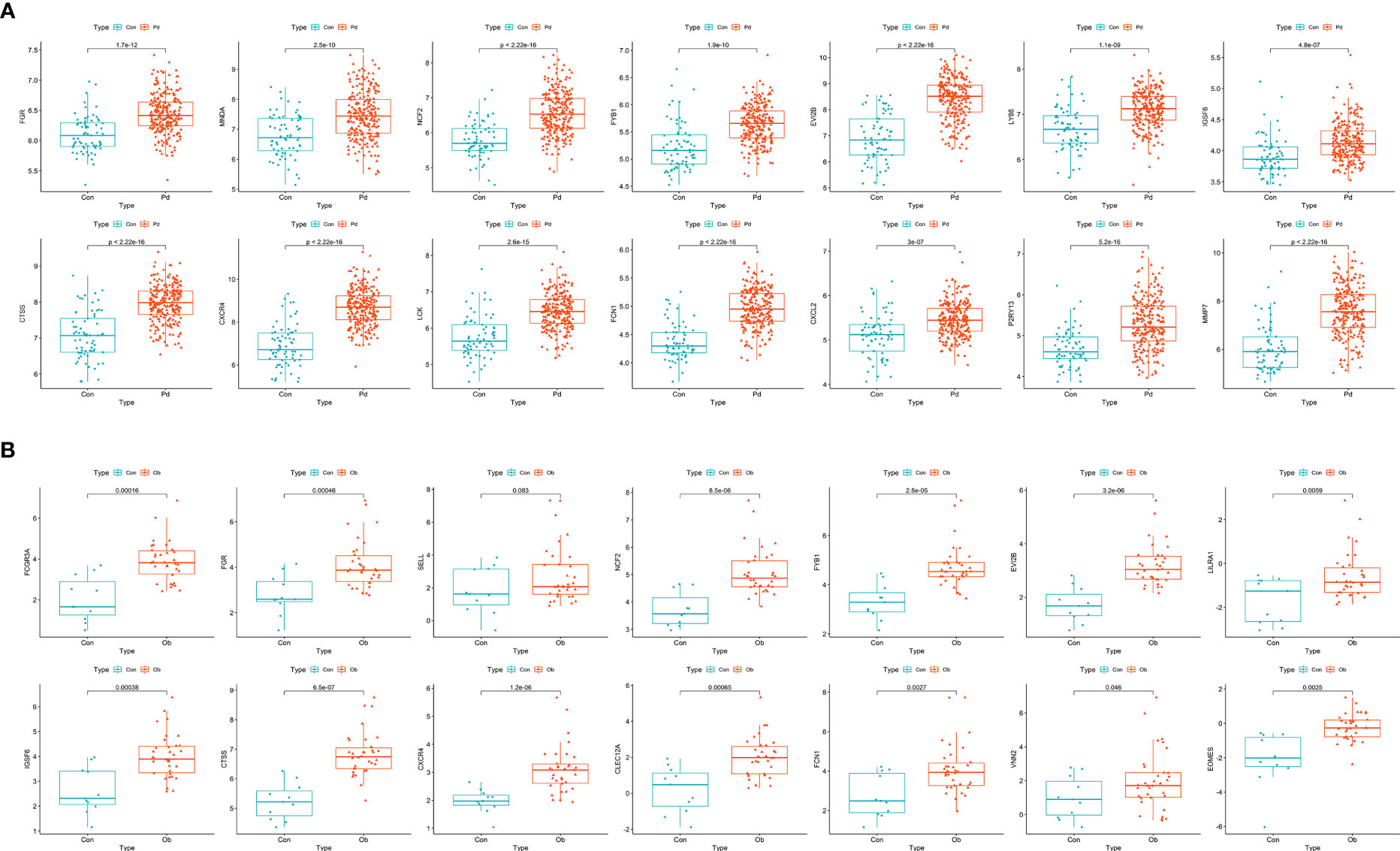
Figure 4 Identification and validation of hub genes. (A) Boxplots of the expression levels of hub genes in GSE152991. The expression levels of the 14 hub genes are significantly higher in the obesity group. (B) Boxplots of the expression levels of hub genes in GSE16134. The expression levels of the 14 hub genes are significantly higher in the periodontitis group. A p-value < 0.05 was considered statistically significant.
ROC analysis was performed on these two datasets to evaluate the accuracy of the diagnostic features of the hub genes. The AUC values of the 14 hub genes were all greater than 0.7 in the OB and PD datasets, indicating excellent predictive ability of these genes (Figure 5). The ROC curves of the hub genes in the four datasets are shown in Supplementary Figures 2, 3.
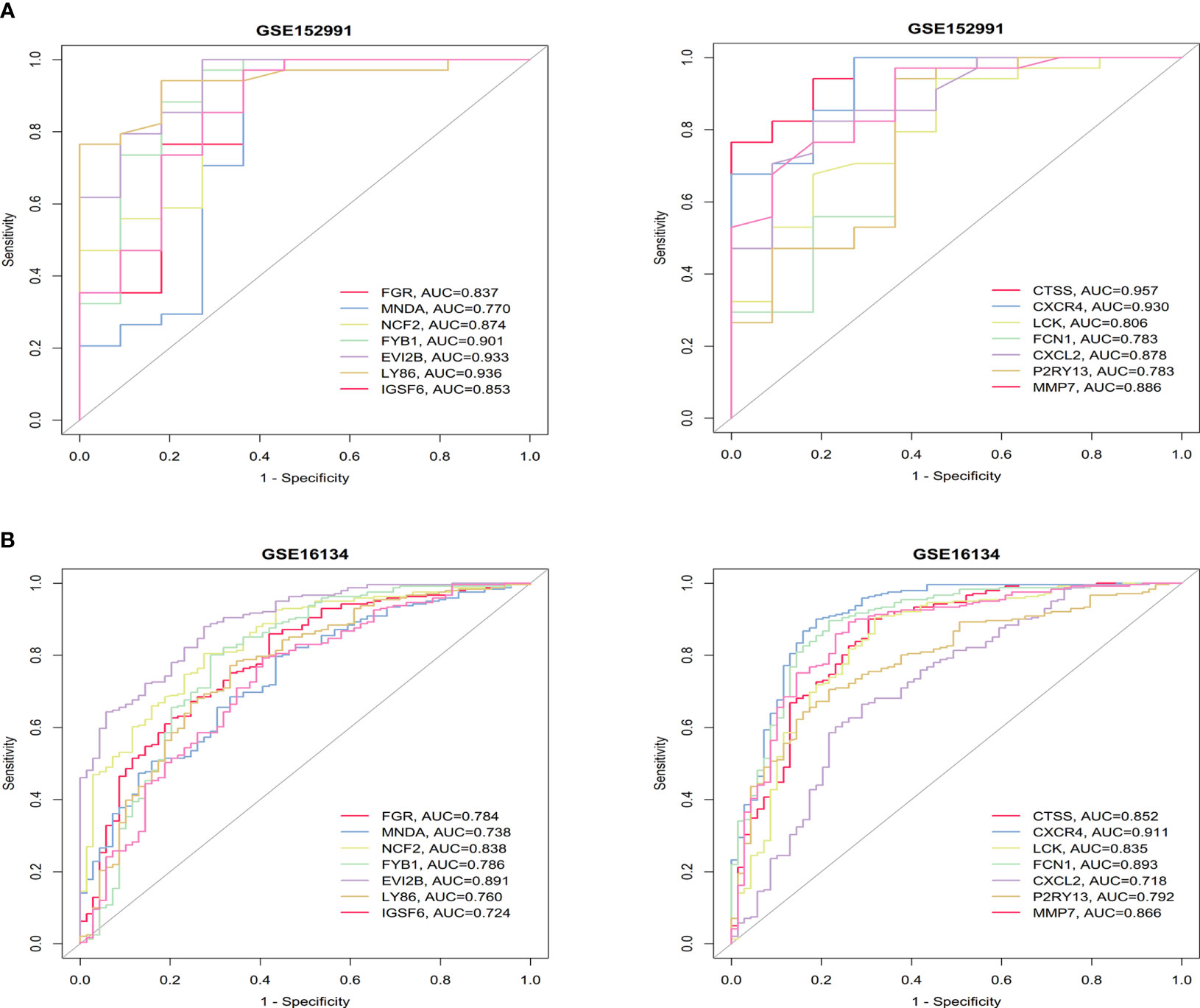
Figure 5 ROC curves of the 14 hub genes in obesity. (A) and periodontitis (B). The AUC values are listed in the lower right-hand corner. ROC, receiver operating characteristic; AUC, area under the curve.
The 14 hub genes were imported to GeneMANIA to find correlated genes based on physical interaction, co-expression, prediction, co-localization, genetic interaction and shared protein domains. The inner circle represents the hub genes, while the outer circle represents the related genes that were newly obtained from the database. The network revealed that these genes were significantly enriched in “macrophage activation”, “phagocytosis”, “leukocyte migration”, “regulation of mononuclear cell proliferation”, “Fc receptor signaling pathway”, and “antigen receptor-mediated signaling pathway” (Figure 6).
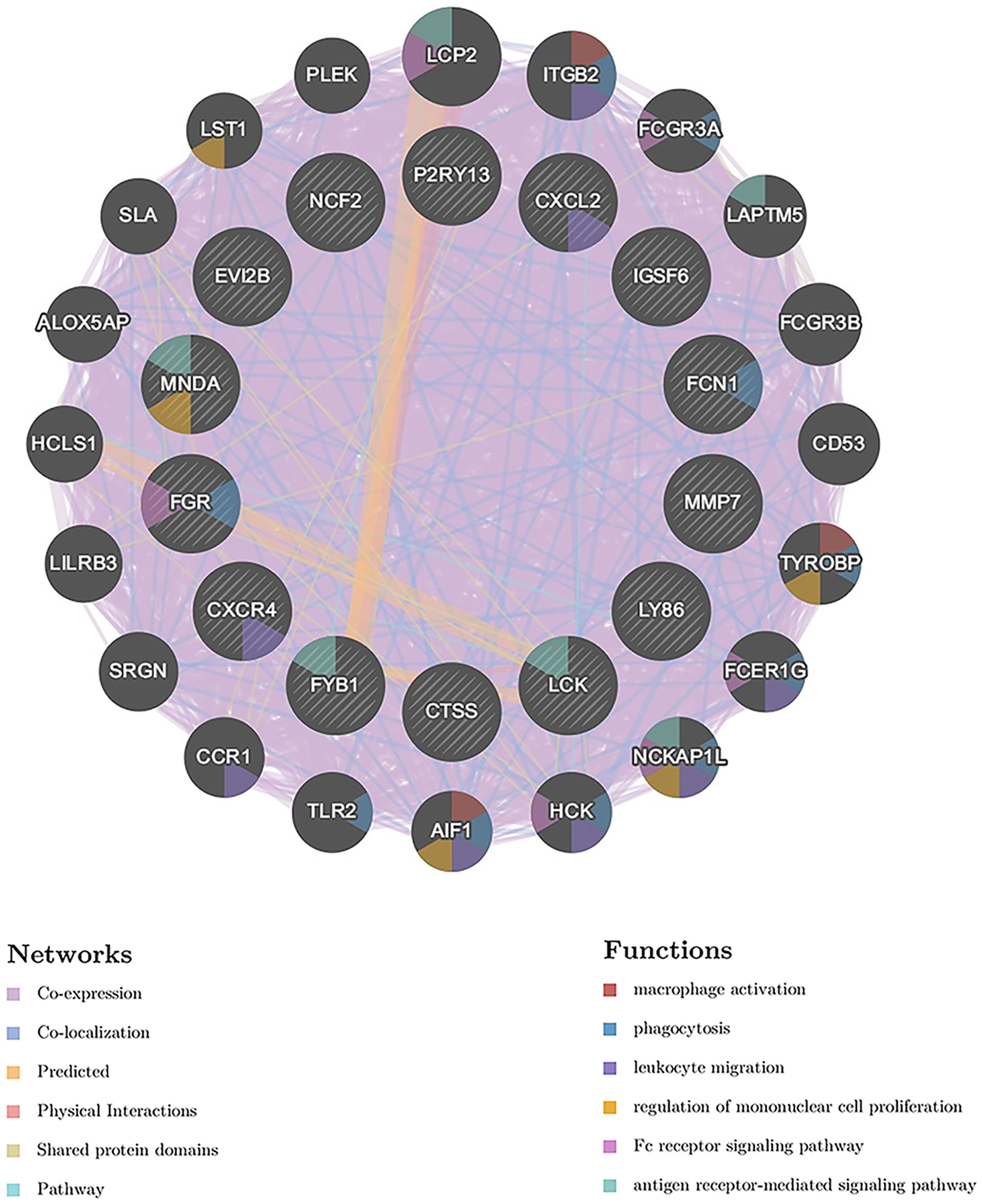
Figure 6 GeneMANIA analysis of the hub genes. The biological functions of the genes are shown. The hub genes are located in the inner circle and the genes correlated with the hub genes are located in the outer circle.
Immune infiltration analysis was performed to evaluate the infiltration level of 28 immune cell types, and the correlations between the 14 hub genes and 28 immune cells were analyzed using Spearman’s method. The infiltration level of immune cells including activated CD4 T cells, activated dendritic cells, central memory CD8 T cells, immune B cells, macrophages, MDSC, natural killer T cells, and plasmacytoid dendritic cells was significantly higher in the OB and PD samples compared with the control samples (Figure 7A; Supplementary Figure 4A), and the infiltration level of immune cells such as MDSC, regulatory T cells and macrophages was positively correlated with the 14 hub genes in both OB and PD. In addition, FGR, FYB1 and LCK were significantly associated with immature B cells, monocytes, and activated CD4 T cells. In PD samples from GSE16134, the 14 hub genes were positively associated with most cell types except effector memory CD4 T cells, type 2 T helper cells, and CD56 dim natural killer cells (Figure 7B; Supplementary Figure 5).
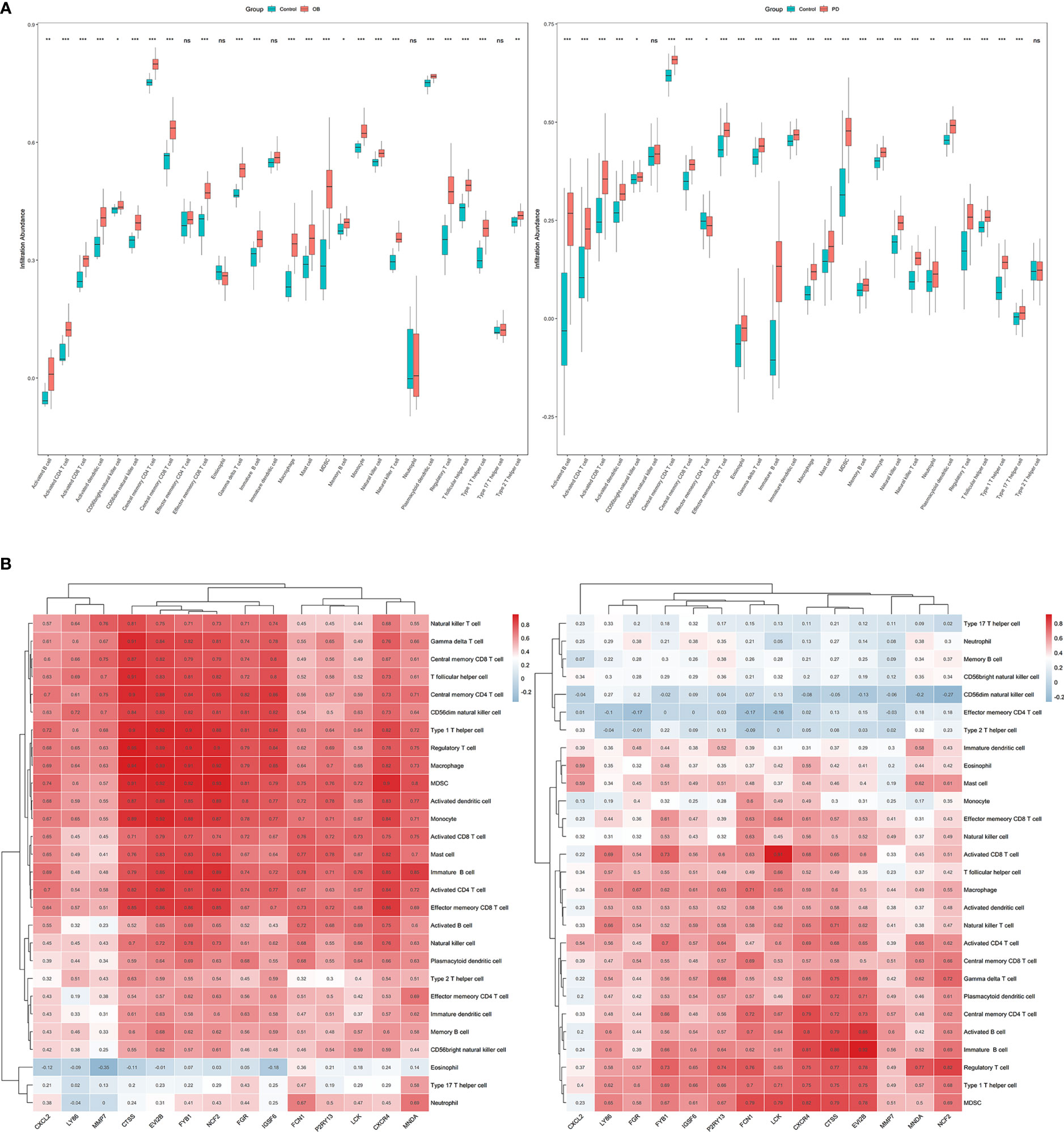
Figure 7 The results of immune infiltration analysis in obesity (OB) and periodontitis (PD) datasets. (A) Boxplots of the immune infiltration abundance in OB (left) and PD (right). (B) Heatmaps of the correlations between the 14 hub genes and the 28 immune cells in OB (left) and PD (right). * p < 0.05, ** p < 0.01, *** p < 0.001, ns, non-signicant.
Both the immune infiltration results and the GeneMANIA analysis suggested that the hub genes might be closely associated with macrophages. Therefore, we explored the enrichment of hub genes in macrophage-associated pathways based on their expression in the GSE152991 (OB) and GSE16134 (PD) datasets using GSEA analysis to determine whether these hub genes are also significantly associated with macrophage-associated pathways. Macrophage-related gene sets including “GOBP_MACROPHAGE_ACTIVATION”, “GOBP_MACROPHAGE_MIGRATION”, “GOBP_MACROPHAGE_CHEMOTAXIS”, “GOBP_MACROPHAGE_ACTIVATION_INVOLVED_IN_IMMUNE_RESPONSE”, “GOBP_MACROPHAGE_ACTIVATION_IN_IMMUNE_RESPONSE”, “GOBP_MACROPHAGE_CHEMOTAXIS”, “GOBP_MACROPHAGE_CYTOKINE_PRODUCTION”, “GOBP_MACROPHAGE_APOPTOTIC_PROCESS”, “GOBP_PHAGOCYTOSIS” and “GOBP_REGULATION_OF_MACROPHAGE_ACTIVATION” were downloaded from MSigDB database and subsequently used for GSEA analysis. The GSEA results indicated that high expression of FGR, FYB1, LY86, LCK and P2RY13 were significantly associated with several macrophage-related biological terms in both GSE152991 (Figure 8A) and GSE16134 (Figure 8B) datasets, such as “activation of macrophage”, “chemotaxis of macrophage”, “migration of macrophage”, “regulation of macrophage apoptotic process” and “macrophage activation involved in immune response”. The nominal p-values, NES and FDR p-values for the GSEA results of GSE152991 and GSE16134 are shown in Supplementary Tables 8, 9.
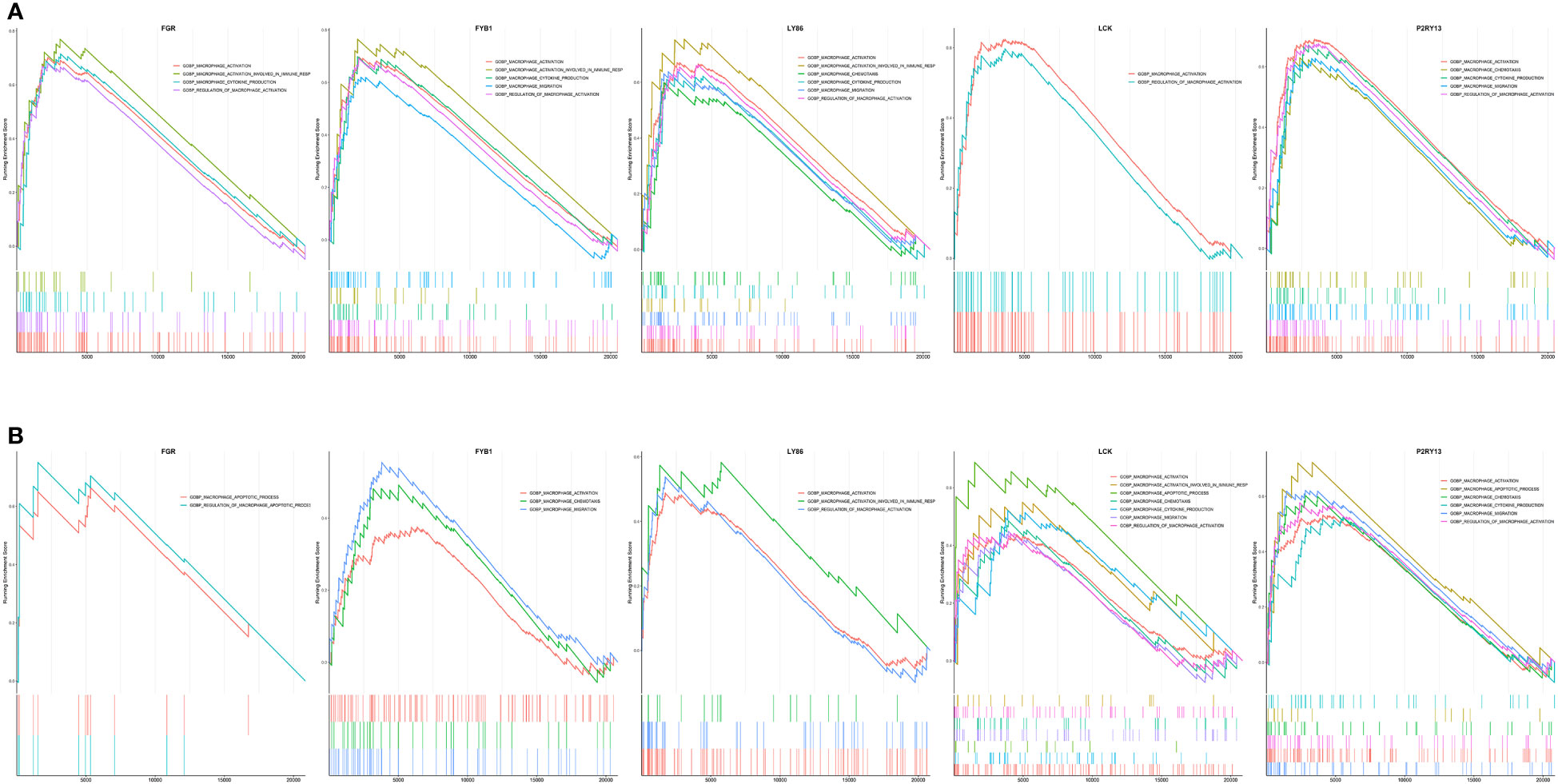
Figure 8 Merged enrichment plot of FGR, FYB1, LY86, LCK and P2RY13 from gene set enrichment analysis of GSE152991 (A) and GSE16134 (B) datasets. The threshold value of GSEA results was set as |NES| > 1.0, p-value < 0.05 & FDR p-value < 0.25. GSEA, gene set enrichment analysis; NES, normalized enrichment score; FDR, false positive rate.
The information of experimentally validated miRNAs and disease associations were downloaded from the HMDD database (Supplementary Table 5), from which we obtained a total of 80 miRNAs associated with PD and 33 miRNAs associated with OB. And 10 common miRNA (hsa-mir-17-5p, hsa-mir-130a-3p, hsa-mir-30a-5p, hsa-mir-126-3p, hsa-mir-146a-5p, hsa-mir-21-5p, hsa-mir-24-3p, hsa-mir-155-5p, hsa-mir-200b-3p and hsa-let-7b-5p) between OB and PD were obtained.
Functional enrichment analysis of the 10 miRNAs was then performed using the mirPath database, and the results indicated that several terms were significantly enriched, including “fatty acid biosynthesis”, “fatty acid metabolism”, “ErbB signaling pathway” and “Wnt signaling pathway and endocytosis” (Figure 9A). The “multiMiR” package was used to find validated target genes of the miRNAs (Supplementary Table 6), and the TransmiR database was used to find the regulatory information between TFs and the miRNAs, from which 233 TFs were obtained (Supplementary Table 7). Based on the regulatory relationships among TFs, mRNAs and miRNAs, a TF-miRNA-mRNA network was constructed (Figure 9B).
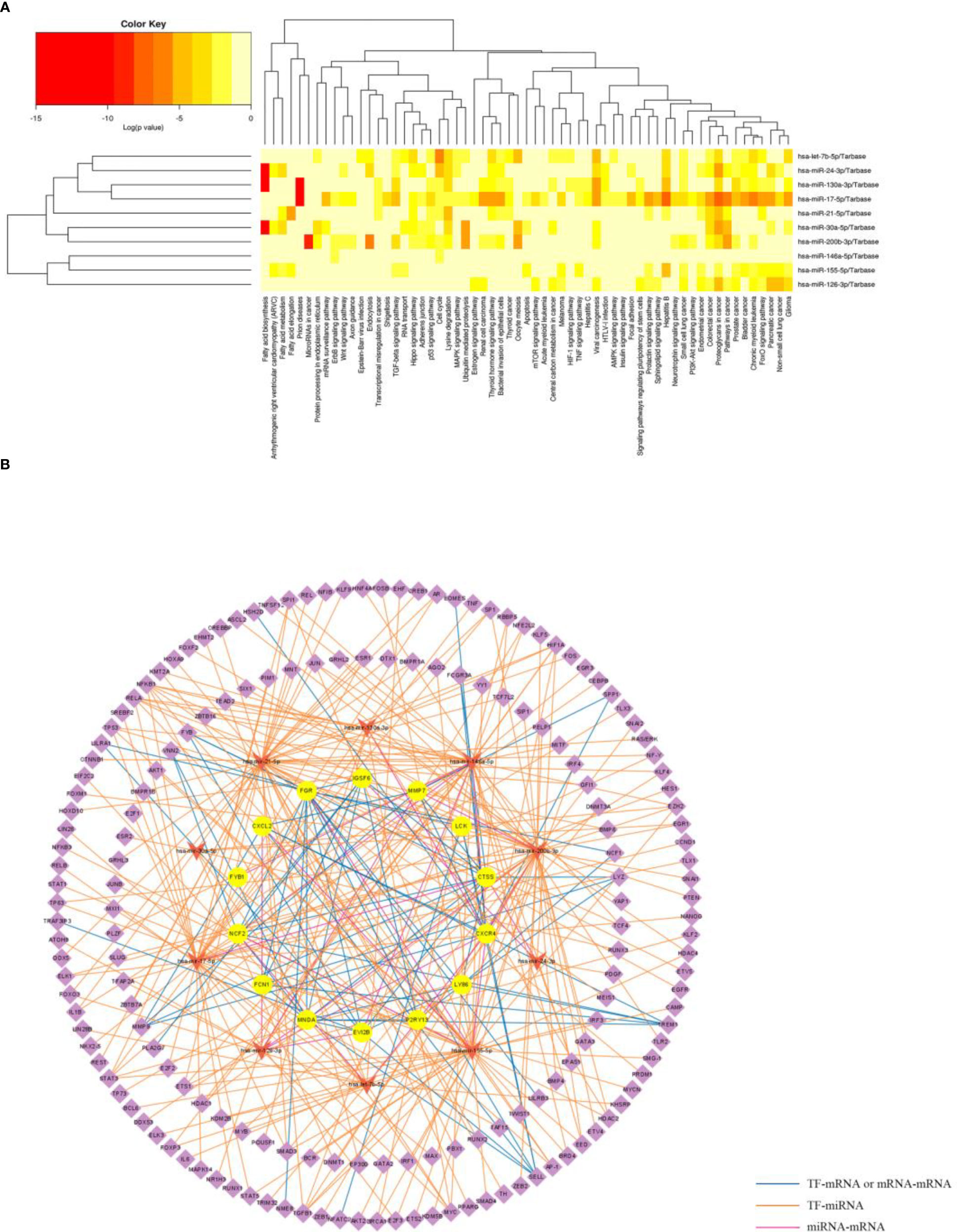
Figure 9 Functional enrichment analysis of the miRNAs and construction of TF-miRNA-mRNA network. (A) Significantly enriched biological pathways of the common miRNAs in OB and PD using the mirPath database. (B) The common TF-miRNA-mRNA regulatory network in OB and PD, where circles represent hub genes, inverted triangles represent miRNAs and diamonds represent transcription factors.
Both OB and PD are common health problems that now cause considerable economic damage and social burden worldwide (50). Previous studies have shown that the risk of periodontitis progression is 15% higher in obese than in healthy populations (51) and a systematic review of epidemiological studies has revealed a significant association between OB and PD (52). OB and PD may share overlapping pathogenic pathways, particularly immune cell infiltration and inflammation (6, 53). However, it is unclear how immune cell infiltration and inflammation accelerate the progression of OB-associated PD.
In our study, the characteristic genes shared by both OB and PD showed a close correlation with immune cell function. Based on integrated bioinformatics analysis, we screened the common DEGs between OB and PD, and functional enrichment analysis revealed that these genes were mainly involved in immune-related biological pathways such as “B cell receptor signaling pathway”, “chemokine signaling pathway”, “leukocyte migration”, “cellular defense response”, “phagocytic vesicle and immune receptor activity”. Through PPI network and hub gene analysis, we identified 25 common hub genes in OB and PD, among which 14 hub genes (FGR, MNDA, NCF2, FYB1, EVI2B, LY86, IGSF6, CTSS, CXCR4, LCK, FCN1, CXCL2, P2RY13, and MMP7) showed high sensitivity and specificity in the ROC curve analysis, indicating that these genes may be promising markers for diagnosis of OB and PD. GSEA analysis of the 14 genes demonstrated that 5 genes including FGR, LCK, FYB1, LY86 and P2RY13 were significantly involved in multiple immune-related GO terms, such as “activation of macrophage”, “chemotaxis of macrophage”, “migration of macrophage”, “regulation of macrophage apoptotic process” and “macrophage activation involved in immune response”. These results suggest that these co-DEGs may accelerate disease progression in OB and PD by affecting the activity of immune cells, especially macrophages.
Furthermore, macrophage infiltration plays a key role in inflammation in obese adipose tissue (54). In obese adipose tissue, macrophage infiltration leads to an increase in pro-inflammatory cytokines, which may lead to inflammation in other tissues via endocrine pathways, thus accelerating the progression of PD (55). In our study, the results of functional enrichment analysis of five common hub genes of OB and PD suggest a possible strong association with macrophages. Ortiz MA et al. (56) indicated that both FGR and LCK belong to the Src family of protein tyrosine kinases, a family of non-receptor tyrosine kinases consisting of nine members in humans: SRC, FGR, LCK, FYN, HCK, YES, LYN, YRK and BLK (56). Several Src family members are expressed in all cell types and are involved in a variety of cellular processes, of which FGR is mainly expressed in the hematopoietic system and LCK is mainly found in T lymphocytes and natural killer cells (57, 58). Like other members of the Src family, both FGR and LCK consist of a kinase structural domain, SH2, SH3 and unique N-terminal structural domains, and a regulatory C-terminal tail that phosphorylates tyrosine residues of a variety of proteins (59). FGR transduces signals from cell surface receptors lacking kinase activity and is involved in immunomodulatory responses, including macrophage, monocyte, neutrophil and mast cell function, cytoskeletal remodeling in response to extracellular stimuli, phagocytosis, cell adhesion and migration (60). LCK plays a key role in T cell antigen receptor (TCR)-related events: binding of the TCR to the peptide antigen-binding MHC complex facilitates the interaction of CD4 and CD8 with MHC class II and class I molecules, respectively, thereby recruiting the associated LCK protein to the vicinity of the TCR/CD3 complex. Subsequently, LCK phosphorylates tyrosine residues within the immunoreceptor tyrosine-based activation motif (ITAM) in the cytoplasmic tail of the TCR-γ chain and CD3 subunit, thereby initiating the TCR/CD3 signaling pathway (61). In the present study, the OB and PD enrichment results also showed that the co-DEGs were mainly enriched for molecular functions such as inhibitory MHC class I receptor activity and MHC class I receptor activity. Many studies have found that FGR expression is high in macrophages and monocytes and that high FGR expression can influence the inflammatory milieu through M1-type macrophage polarization, which in turn affects adipose tissue metabolism, leading to the development of inflammation and OB (62–64). He L et al. (65) also showed that high expression of LCK plays an important role in the imbalance of the immune system in periodontitis (65). In another study, FGR kinase was found to be a key regulator of pro-inflammatory adipose tissue macrophage activation, diet-induced OB, insulin resistance, and hepatic steatosis, knockdown of FGR reduced lipid accumulation and lipogenic gene expression, and low expression of FGR prevented macrophage polarization while preventing high-fat diet-induced OB in mice (66). This is consistent with our findings that FGR and LCK expression were significantly up-regulated in OB and PD, and functional enrichment analysis showed that they are involved in immune processes activated by macrophages. Therefore, we speculate that OB accelerates PD progression, at least in part, through immune cell infiltration and inflammatory response, and that FGR and LCK may play a key role in this process.
FYB1, known as adhesion and degranulation promoting linker protein (ADAP), is required for T cell activation as a bridging protein for the FYN and LCP2 signaling cascades in T cells (67). In addition, FYB1 can be expressed on primary natural killer cells and on lymphocyte-activated killer cells stimulated by interleukin-2, resulting in enhanced antitumor responses (68). Carmo AM et al. (69) indicated that post-translational modification of FYB1 could lead to increased tyrosine phosphorylation by affecting T-cell receptor attachment (69). The protein encoded by the LY86 gene is lymphocyte antigen 86, also known as MD-1 protein, a secreted glycoprotein associated with RP105 (Toll-like receptor family protein), which plays a key role in the B-cell surface expression of RP105, while the RP105/MD-1 complex is expressed in immune cells, including B cells, macrophages and dendritic cells (70). In a genome-wide methylation analysis study of OB, the methylation level of the LY86 gene was shown to be higher in obese cases compared to healthy controls (71). The product of the P2RY13 gene belongs to the G protein-coupled receptor family, a 354 amino acid-encoded gastrointestinal protein-coupled receptor that is involved in the pathogenesis of purine energy transfer pathways, cholesterol metabolism, inflammation and immune dysfunction mechanisms, mediating a variety of pathophysiological processes, such as apoptosis, autophagy, proliferation and metabolism (72, 73). Many studies have shown that P2RY13 promotes apoptosis and increases the release of pro-inflammatory factors (74, 75). In addition, P2RY13 is highly expressed in the inflamed intestinal tissue of ulcerative colitis patients (74, 76). The above series of studies have shown that FGR, LCK, FYB1, LY86, and P2RY13 may play an important role in the dysfunction of immune cells such as macrophages, T lymphocytes, and B lymphocytes during the pathology of OB or PD. Therefore, we speculate that macrophage infiltration and recruitment of other immune cells may be common mechanisms in OB and PD. A possible molecular mechanism is that in OB and PD, upregulation of LY86 gene expression affects B-cell function, and the increase in B-cell number promotes T-cell activation. Meanwhile, FYB1 is involved in T-cell activation and LCK phosphorylates the T-cell antigen receptor (TCR), but increased T cell activation may in turn promote M1-like macrophage polarization and inflammation. Among other things, FGR transduces signals from cell surface receptors lacking kinase activity and is involved in immunoregulatory responses such as macrophage function. Finally, P2RY13 promotes the release of inflammatory factors.
To further explore the potential molecular regulatory mechanisms between these immune-related hub genes and immune cells, especially macrophages, we constructed a TF-miRNA-mRNA regulatory network for OB and PD. The results showed that among the five hub genes, FGR was regulated by hsa-miR-155-5p and FYB1 was regulated by hsa-miR-146a-5p. In addition, TFs such as AKT1, BRCA1, and TP63 inhibited the regulatory effect of hsa-miR-155-5p, and TFs such as FOXP3, JUNB, NFKB1, and SMAD4 activated the regulation of hsa-miR-155-5p. On the other hand, the regulatory role of hsa-miR-146a-5p was inhibited by TFs such as HDAC1, TP53, and HDAC1, and could also be activated by TFs such as FOXP3, IL1B, and RELA. Notably, previous findings have shown that hsa-miR-155-5p expression is up-regulated under various inflammatory conditions (77). Langi G et al. (78) showed that the expression of hsa-miR-155-5p is down-regulated in OB patients undergoing bariatric surgery (78). In addition, miR-146a-5p expression was up-regulated in humans and mice obese adipose tissue and suppressed the inflammatory response in human adipocytes (79). Meanwhile, animal experiments revealed that deletion of the transcription factor AKT1 increased energy expenditure and prevented diet-induced obesity in mice (80), and AKT1 regulated macrophage polarization and alters periodontal inflammatory status (81). These findings are consistent with our study, where 14 hub genes were found to be up-regulated in both the OB and PD groups and may have a strong association with immune cells, especially macrophages. Therefore, we can reasonably speculate that the upregulation of FGR and FYB1 expression in OB and PD may be regulated by hsa-miR-155-5p and hsa-miR-146a-5p and TFs such as AKT1, FOXP3, TP53, and IL1B are involved in the regulatory process of hsa-miR-155-5p and hsa-miR-146a-5p.
In this study, we used transcriptome data to elucidate the common mechanisms of OB and PD. Our study is novel in that we analyzed the infiltration of 28 immune cell species in adipose tissue of OB patients and periodontal tissue of PD patients using the ssGSEA algorithm. It is comprehensive in that we elucidated the key genes, biological pathways, immune infiltration levels, and TF-miRNA-mRNA networks of OB and PD, which will be helpful in understanding the pathophysiological mechanisms shared between the two diseases and the treatment of PD patients with OB. However, there are still limitations to our study. First of all, there is limited clinical information available in public databases, and not all datasets have large sample sizes, which may lead to bias in the results. In addition, this study was mainly based on bioinformatics analysis. Although for hub genes with high diagnostic value, we used different datasets to validate this diagnostic value, and many previous studies were able to confirm our findings to some extent, further experimental validation of our findings is still needed.
Our study provides key co-diagnostic effector genes for OB and PD patients and reveals that the common key genes of both diseases are closely associated with immune cell infiltration. The possible molecular mechanism of accelerated PD progression in OB is that the secretion of pro-inflammatory cytokines increases with immune cell infiltration and inflammatory response in OB, causing inflammation in other tissues by endocrine means, thus accelerating PD progression. Five hub genes (FGR, LCK, FYB1, LY86, P2RY13) are promising biomarkers for OB and PD and may play an important role in the pathogenesis of OB and PD by influencing the activity of macrophages involved in immune regulation and inflammatory responses.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
XQ, YC, and XC designed the study and drafted the manuscript. YZ and SW contributed to the data collection. YC, XZ, WP, and KM contributed to data analysis. YC, XZ, YZ, and SW drafted the manuscript. XQ, LT and HD revised the manuscript. All authors contributed to the article and approved the submitted version.
This work was funded by Scientific Research Projects of Provincial Education Department of Hunan (22C0009), National Students’ Platform for Innovation and Entrepreneurship Training Program (202210542035), and the 2021 Hunan Provincial Excellent Graduate Demonstration Course, "Bioinformatics", Department of Education of Hunan Province, Xiang Jiao Tong [2022] No.116.
The authors thank the GEO database for providing open access to the data and the reviewers for their constructive comments.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2023.1101854/full#supplementary-material
1. Boesing F, Patiño J, Da Silva V, Moreira E. The interface between obesity and periodontitis with emphasis on oxidative stress and inflammatory response. Obes Rev(2009) 10(3):290–7. doi: 10.1111/j.1467-789X.2008.00555.x
2. Pamuk F, Kantarci A. Inflammation as a link between periodontal disease and obesity. Periodontol 2000. (2022) 90(1):186–96. doi: 10.1111/prd.12457
3. Zhao P, Xu A, Leung WKJB. Obesity, bone loss, and periodontitis: The interlink. Biomolecules (2022) 12(7):865. doi: 10.3390/biom12070865
4. Loos RJ, Yeo GSH. The genetics of obesity: From discovery to biology. Nat Rev Genet (2022) 23(2):120–33. doi: 10.1038/s41576-021-00414-z
5. Kinane DF, Stathopoulou PG, Papapanou PN. Periodontal diseases. Nat Rev Dis Primers. (2017) 3(1):17038. doi: 10.1038/nrdp.2017.38
6. Könönen E, Gursoy M, Gursoy UK. Periodontitis: A multifaceted disease of tooth-supporting tissues. J Clin Med (2019) 8(8):1135. doi: 10.3390/jcm8081135
7. Martinez-Herrera M, Silvestre-Rangil J, Silvestre F-J. Association between obesity and periodontal disease. A systematic review of epidemiological studies and controlled clinical trials. patol Oral y cirugia bucal (2017) 22(6):e708. doi: 10.4317/medoral.21786
8. Perlstein MI, Bissada NF. Influence of obesity and hypertension on the severity of periodontitis in rats. Oral surgery Oral med Oral pathol (1977) 43(5):707–19. doi: 10.1016/0030-4220(77)90055-X
9. Virto L, Cano P, Jiménez-Ortega V, Fernández-Mateos P, González J, Esquifino AI, et al. Obesity and periodontitis: An experimental study to evaluate periodontal and systemic effects of comorbidity. J Periodontol (2018) 89(2):176–85. doi: 10.1902/jop.2017.170355
10. Cecoro G, Annunziata M, Iuorio MT, Nastri L, Guida LJM. Periodontitis, low-grade inflammation and systemic health: a scoping review. Medicina (2020) 56(6):272. doi: 10.3390/medicina56060272
11. Al-Zahrani MS, Bissada NF, Borawskit EA. Obesity and periodontal disease in young, middle-aged, and older adults. J periodontol (2003) 74(5):610–5. doi: 10.1902/jop.2003.74.5.610
12. Martinez-Herrera M, Javier Silvestre F, Silvestre-Rangil J, Banuls C, Rocha M, Hernandez-Mijares A. Involvement of insulin resistance in normoglycaemic obese patients with periodontitis: A cross-sectional study. J Clin Periodontol (2017) 44(10):981–8. doi: 10.1111/jcpe.12773
13. Muluke M, Gold T, Kiefhaber K, Al-Sahli A, Celenti R, Jiang H, et al. Diet-induced obesity and its differential impact on periodontal bone loss. J Dental Res (2016) 95(2):223–9. doi: 10.1177/0022034515609882
14. Chen Z-y, Xu T-t, Liang Z-j, Zhao L, Xiong X-q, Xie K-k, et al. Untargeted and targeted gingival metabolome in rodents reveal metabolic links between high-fat diet-induced obesity and periodontitis. J Clin Periodontol (2021) 48(8):1137–48. doi: 10.1111/jcpe.13486
15. Huh JY, Park YJ, Ham M, Kim JB. Crosstalk between adipocytes and immune cells in adipose tissue inflammation and metabolic dysregulation in obesity. Molecules Cells (2014) 37(5):365–71. doi: 10.14348/molcells.2014.0074
16. Osborn O, Olefsky JM. The cellular and signaling networks linking the immune system and metabolism in disease. Nat Med (2012) 18(3):363–74. doi: 10.1038/nm.2627
17. Chatzigeorgiou A, Karalis K, Bornstein S, Chavakis TJD. Lymphocytes in obesity-related adipose tissue inflammation. Diabetologia (2012) 55(10):2583–92. doi: 10.1007/s00125-012-2607-0
18. Wu H, Ballantyne CM. Metabolic inflammation and insulin resistance in obesity. Circ Res (2020) 126(11):1549–64. doi: 10.1161/CIRCRESAHA.119.315896
19. Feuerer M, Herrero L, Cipolletta D, Naaz A, Wong J, Nayer A, et al. Lean, but not obese, fat is enriched for a unique population of regulatory T cells that affect metabolic parameters. Nat Med (2009) 15(8):930–9. doi: 10.1038/nm.2002
20. Winer S, Chan Y, Paltser G, Truong D, Tsui H, Bahrami J, et al. Normalization of obesity-associated insulin resistance through immunotherapy. Nat Med (2009) 15(8):921–9. doi: 10.1038/nm.2001
21. Nishimura S, Manabe I, Nagasaki M, Eto K, Yamashita H, Ohsugi M, et al. CD8+ effector T cells contribute to macrophage recruitment and adipose tissue inflammation in obesity. Nat Med (2009) 15(8):914–20. doi: 10.1038/nm.1964
22. Dutzan N, Vernal R, Vaque JP, García-Sesnich J, Hernandez M, Abusleme L, et al. Interleukin-21 expression and its association with proinflammatory cytokines in untreated chronic periodontitis patients. J Periodontol (2012) 83(7):948–54. doi: 10.1902/jop.2011.110482
23. Souza PPC, Palmqvist P, Lundberg P, Lundgren I, Hänström L, Souza JAC, et al. Interleukin-4 and interleukin-13 inhibit the expression of leukemia inhibitory factor and interleukin-11 in fibroblasts. Mol Immunol (2012) 49(4):601–10. doi: 10.1016/j.molimm.2011.10.009
24. Akman PT, Fentoğlu O, Yılmaz G, Arpak N. Serum plasminogen activator inhibitor-1 and tumor necrosis factor-α levels in obesity and periodontal disease. J Periodontol (2012) 83(8):1057–62. doi: 10.1902/jop.2011.110548
25. Kern PA, Ranganathan S, Li C, Wood L, Ranganathan G. Adipose tissue tumor necrosis factor and interleukin-6 expression in human obesity and insulin resistance. Am J Physiol Endocrinol Metab (2001) 280(5):E745–51. doi: 10.1152/ajpendo.2001.280.5.E745
26. Herrera BS, Hasturk H, Kantarci A, Freire MO, Nguyen O, Kansal S, et al. Impact of resolvin E1 on murine neutrophil phagocytosis in type 2 diabetes. Infection immunity. (2015) 83(2):792–801. doi: 10.1128/IAI.02444-14
27. Ritchie ME, Phipson B, Wu D, Hu Y, Law CW, Shi W, et al. Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res (2015) 43(7):e47–e. doi: 10.1093/nar/gkv007
28. Gao C-H, Yu G, Cai P. GgVennDiagram: An intuitive, easy-to-use, and highly customizable r package to generate venn diagram. Front Genet (2021) 2:706907. doi: 10.3389/fgene.2021.706907
29. Wu T, Hu E, Xu S, Chen M, Guo P, Dai Z, et al. clusterProfiler 4.0: A universal enrichment tool for interpreting omics data. Innovation (2021) 2(3):100141. doi: 10.1016/j.xinn.2021.100141
30. Bu D, Luo H, Huo P, Wang Z, Zhang S, He Z, et al. KOBAS-i: Intelligent prioritization and exploratory visualization of biological functions for gene enrichment analysis. Nucleic Acids Res (2021) 49(W1):W317–W25. doi: 10.1093/nar/gkab447
31. Shannon P, Markiel A, Ozier O, Baliga NS, Wang JT, Ramage D, et al. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res (2003) 13(11):2498–504. doi: 10.1101/gr.1239303
32. Saito R, Smoot ME, Ono K, Ruscheinski J, Wang P-L, Lotia S, et al. A travel guide to cytoscape plugins. Nat Methods (2012) 9(11):1069–76. doi: 10.1038/nmeth.2212
33. Chin C-H, Chen S-H, Wu H-H, Ho C-W, Ko M-T, Lin C-Y. cytoHubba: identifying hub objects and sub-networks from complex interactome. Bmc Syst Biol (2014) 8(4):1–7. doi: 10.1186/1752-0509-8-S4-S11
34. Wang M, Wang H, Zheng H. A mini review of node centrality metrics in biological networks. International Journal of Network Dynamics and Intelligence (2022) 1(1):99–110. doi: 10.53941/ijndi0101009
35. Mahi NA, Najafabadi MF, Pilarczyk M, Kouril M, Medvedovic M. GREIN: An interactive web platform for re-analyzing GEO RNA-seq data. Sci Rep (2019) 9(1):7580. doi: 10.1038/s41598-019-43935-8
36. Robin X, Turck N, Hainard A, Tiberti N, Lisacek F, Sanchez J-C, et al. pROC: An open-source package for r and s+ to analyze and compare ROC curves. BMC Bioinformatics (2011) 12(1):1–8. doi: 10.1186/1471-2105-12-77
37. Bantis LE, Young KJ, Tsimikas JV, Mosier BR, Gajewski B, Yeatts S, et al. Statistical assessment of the prognostic and the predictive value of biomarkers-a biomarker assessment framework with applications to traumatic brain injury biomarker studies. Res Methods in Med Health Sci (2023) 4(1):34–48. doi: 10.1177/26320843221141056
38. Franz M, Rodriguez H, Lopes C, Zuberi K, Montojo J, Bader GD, et al. GeneMANIA update 2018. Nucleic Acids Res (2018) 46(W1):W60–W4. doi: 10.1093/nar/gky311
39. Mootha VK, Lindgren CM, Eriksson K-F, Subramanian A, Sihag S, Lehar J, et al. PGC-1α-responsive genes involved in oxidative phosphorylation are coordinately downregulated in human diabetes. Nat Genet (2003) 34(3):267–73. doi: 10.1038/ng1180
40. Subramanian A, Tamayo P, Mootha VK, Mukherjee S, Ebert BL, Gillette MA, et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles.Proc Natl Acad Sci USA (2005) 102(43):15545–50. doi: 10.1073/pnas.0506580102
41. Finotello F, Trajanoski Z. Quantifying tumor-infiltrating immune cells from transcriptomics data. Cancer Immunol Immunother (2018) 67(7):1031–40. doi: 10.1007/s00262-018-2150-z
42. Bindea G, Mlecnik B, Tosolini M, Kirilovsky A, Waldner M, Obenauf AC, et al. Spatiotemporal dynamics of intratumoral immune cells reveal the immune landscape in human cancer. Immunity (2013) 39(4):782–95. doi: 10.1016/j.immuni.2013.10.003
43. Lv Y, Zhang T, Cai J, Huang C, Zhan S, Liu J. Bioinformatics and systems biology approach to identify the pathogenetic link of long COVID and myalgic Encephalomyelitis/Chronic fatigue syndrome. Front Immunol (2022) 13:5144. doi: 10.3389/fimmu.2022.952987
44. Bartel DP. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell (2004) 116(2):281–97. doi: 10.1016/S0092-8674(04)00045-5
45. Hobert O. Common logic of transcription factor and microRNA action. Trends Biochem Sci (2004) 29(9):462–8. doi: 10.1016/j.tibs.2004.07.001
46. Zhang H-M, Kuang S, Xiong X, Gao T, Liu C, Guo A-Y. Transcription factor and microRNA co-regulatory loops: Important regulatory motifs in biological processes and diseases. Brief Bioinform (2015) 16(1):45–58. doi: 10.1093/bib/bbt085
47. Huang Z, Shi J, Gao Y, Cui C, Zhang S, Li J, et al. HMDD v3. 0: A database for experimentally supported human microRNA–disease associations. Nucleic Acids Res (2019) 47(D1):D1013–D7. doi: 10.1093/nar/gky1010
48. Ru Y, Kechris KJ, Tabakoff B, Hoffman P, Radcliffe RA, Bowler R, et al. The multiMiR r package and database: Integration of microRNA–target interactions along with their disease and drug associations. Nucleic Acids Res (2014) 42(17):e133–e. doi: 10.1093/nar/gku631
49. Tong Z, Cui Q, Wang J, Zhou Y. TransmiR v2. 0: An updated transcription factor-microRNA regulation database. Nucleic Acids Res (2019) 47(D1):D253–D8. doi: 10.1093/nar/gky1023
50. Kim CM, Lee S, Hwang W, Son E, Kim TW, Kim K, et al. Obesity and periodontitis: A systematic review and updated meta-analysis. Front Endocrinol (Lausanne). (2022) 13:999455. doi: 10.3389/fendo.2022.999455
51. Charupinijkul A, Arunyanak S, Rattanasiri S, Vathesatogkit P, Thienpramuk L, Lertpimonchai A. The effect of obesity on periodontitis progression: The 10-year retrospective cohort study. Clin Oral Investig (2022) 26(1):535–42. doi: 10.1007/s00784-021-04031-2
52. Suvan J, D’Aiuto F, Moles DR, Petrie A, Donos N. Association between overweight/obesity and periodontitis in adults. A systematic review. Obes Rev (2011) 12(5):e381–404. doi: 10.1111/j.1467-789X.2010.00808.x
53. Ahmed B, Sultana R, Greene MW. Adipose tissue and insulin resistance in obese. Biomed Pharmacother (2021) 137:111315. doi: 10.1016/j.biopha.2021.111315
54. Lu J, Zhao J, Meng H, Zhang X. Adipose tissue-resident immune cells in obesity and type 2 diabetes. Front Immunol (2019) 10:1173. doi: 10.3389/fimmu.2019.01173
55. Hajishengallis G, Sahingur S. Novel inflammatory pathways in periodontitis. Adv Dent Res (2014) 26(1):23–9. doi: 10.1177/0022034514526240
56. Ortiz MA, Mikhailova T, Li X, Porter BA, Bah A, Kotula LJCC, et al. Src family kinases, adaptor proteins and the actin cytoskeleton in epithelial-to-mesenchymal transition. Cell Commun Signal (2021) 19(1):1–19. doi: 10.1186/s12964-021-00750-x
57. Zhai Y, Yang J, Zhang J, Yang J, Li Q, Zheng T. Src-family protein tyrosine kinases: A promising target for treating cardiovascular diseases. Int J Med Sci (2021) 18(5):1216. doi: 10.7150/ijms.49241
58. Singh PK, Kashyap A, Silakari O. Exploration of the therapeutic aspects of lck: A kinase target in inflammatory mediated pathological conditions. Biomed Pharmacother (2018) 108:1565–71. doi: 10.1016/j.biopha.2018.10.00259
59. Li H, Zhao C, Tian Y, Lu J, Zhang G, Liang S, et al. Src family kinases and pulmonary fibrosis: A review. Biomed Pharmacother (2020) 127:110183. doi: 10.1016/j.biopha.2020.110183
60. Roskoski R Jr. Src protein–tyrosine kinase structure and regulation. Biochem Biophys Res Commun (2004) 324(4):1155–64. doi: 10.1016/j.bbrc.2004.09.171
61. Bommhardt U, Schraven B, Simeoni L. Beyond TCR signaling: Emerging functions of lck in cancer and immunotherapy. Int J Mol Sci (2019) 20(14):3500. doi: 10.3390/ijms20143500
62. Covarrubias AJ, Aksoylar HI, Horng T. Control of macrophage metabolism and activation by mTOR and akt signaling. Semin Immunol (2015) 27(4):286–96. doi: 10.1016/j.smim.2015.08.001
63. Fitzer-Attas CJ, Lowry M, Crowley MT, Finn AJ, Meng F, DeFranco AL, et al. Fcγ receptor–mediated phagocytosis in macrophages lacking the src family tyrosine kinases hck, fgr, and Lyn. J Exp Med (2000) 191(4):669–82. doi: 10.1084/jem.191.4.669
64. Acín-Pérez R, Iborra S, Martí-Mateos Y, Cook EC, Conde-Garrosa R, Petcherski A, et al. Fgr kinase is required for proinflammatory macrophage activation during diet-induced obesity. Nat Metab (2020) 2(9):974–88. doi: 10.1038/s42255-020-00273-8
65. He L, Liu L, Li T, Zhuang D, Dai J, Wang B, et al. Exploring the imbalance of periodontitis immune system from the cellular to molecular level. Front Genet (2021) 12:653209. doi: 10.3389/fgene.2021.653209
66. Feng J, Zhang X, Shan C, Xia J, Zhang Z, Shi H, et al. Src family kinases involved in the differentiation of human preadipocytes. Mol Cell Endocrinol (2021) 533:111323. doi: 10.1016/j.mce.2021.111323
67. Parzmair GP, Gereke M, Haberkorn O, Annemann M, Podlasly L, Kliche S, et al. ADAP plays a pivotal role in CD4+ T cell activation but is only marginally involved in CD8+ T cell activation, differentiation, and immunity to pathogens. J Leukocyte Biol (2017) 101(2):407–19. doi: 10.1189/jlb.1A0216-090RR
68. Fostel LV, Dluzniewska J, Shimizu Y, Burbach BJ, Peterson EJ. ADAP is dispensable for NK cell development and function. Int Immunol (2006) 18(8):1305–14. doi: 10.1093/intimm/dxl063
69. Carmo AM, Henriques SN. Cell activation and signaling in lymphocytes. Tissue-Specific Cell Signaling (2020) 133–61. doi: 10.1007/978-3-030-44436-5
70. Nagai Y, Shimazu R, Ogata H, Akashi S, Sudo K, Yamasaki H, et al. Requirement for MD-1 in cell surface expression of RP105/CD180 and b-cell responsiveness to lipopolysaccharide. Blood (2002) 99(5):1699–705. doi: 10.1182/blood.V99.5.1699
71. Xu X, Su S, Barnes VA, De Miguel C, Pollock J, Ownby D, et al. A genome-wide methylation study on obesity: differential variability and differential methylation. Epigenetics (2013) 8(5):522–33. doi: 10.4161/epi.24506
72. Ishimaru M, Yusuke N, Tsukimoto M, Harada H, Takenouchi T, Kitani H, et al. Purinergic signaling via P2Y receptors up-mediates IL-6 production by liver macrophages/Kupffer cells. J Toxicol Sci (2014) 39(3):413–23. doi: 10.2131/jts.39.413
73. Wang N, Robaye B, Gossiel F, Boeynaems JM, Gartland A. The P2Y13 receptor regulates phosphate metabolism and FGF-23 secretion with effects on skeletal development. FASEB J (2014) 28(5):2249–59. doi: 10.1096/fj.13-243626
74. Wu X, Wei S, Chen M, Li J, Wei Y, Zhang J, et al. P2RY13 exacerbates intestinal inflammation by damaging the intestinal mucosal barrier via activating IL-6/STAT3 pathway. Int J Biol Sci (2022) 18(13):5056. doi: 10.7150/ijbs.74304
75. Tan C, Salehi A, Svensson S, Olde B, Erlinge DJC. ADP receptor P2Y 13 induce apoptosis in pancreatic β-cells. Cell Mol Life Sci (2010) 67:445–53. doi: 10.1007/s00018-009-0191-3
76. Voss U, Turesson MF, Robaye B, Boeynaems J-M, Olde B, Erlinge D, et al. The enteric nervous system of P2Y13 receptor null mice is resistant against high-fat-diet-and palmitic-acid-induced neuronal loss. Purinerg Signal (2014) 10(3):455–64. doi: 10.1007/s11302-014-9408-5
77. O’Connell RM, Rao DS, Baltimore D. microRNA regulation of inflammatory responses. Annu Rev Immunol (2012) 30:295–312. doi: 10.1146/annurev-immunol-020711-075013
78. Langi G, Szczerbinski L, Kretowski AJ. Meta-analysis of differential miRNA expression after bariatric surgery. J Clin Med (2019) 8(8):1220. doi: 10.3390/jcm8081220
79. Roos J, Enlund E, Funcke J-B, Tews D, Holzmann K, Debatin K-M, et al. miR-146a-mediated suppression of the inflammatory response in human adipocytes. Sci Rep (2016) 6(1):38339. doi: 10.1038/srep38339
80. Wan M, Easton RM, Gleason CE, Monks BR, Ueki K, Kahn CR, et al. Loss of Akt1 in mice increases energy expenditure and protects against diet-induced obesity. Mol Cell Biol (2012) 32(1):96–106. doi: 10.1128/MCB.05806-11
Keywords: obesity, periodontitis, bioinformatics, transcriptomic analysis, immune processes, macrophage infiltration, inflammation
Citation: Cai Y, Zuo X, Zuo Y, Wu S, Pang W, Ma K, Yi Q, Tan L, Deng H, Qu X and Chen X (2023) Transcriptomic analysis reveals shared gene signatures and molecular mechanisms between obesity and periodontitis. Front. Immunol. 14:1101854. doi: 10.3389/fimmu.2023.1101854
Received: 18 November 2022; Accepted: 24 February 2023;
Published: 29 March 2023.
Edited by:
Pingping Li, Chinese Academy of Medical Sciences and Peking Union Medical College, ChinaReviewed by:
Hong-Hui Wang, Hunan University, ChinaCopyright © 2023 Cai, Zuo, Zuo, Wu, Pang, Ma, Yi, Tan, Deng, Qu and Chen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xiaochao Qu, cXV4Y0BodW5udS5lZHUuY24=; Xiangding Chen, eGRjaGVuQGh1bm51LmVkdS5jbg==
†These authors have contributed equally to this work and share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.