
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Immunol., 31 January 2023
Sec. Nutritional Immunology
Volume 14 - 2023 | https://doi.org/10.3389/fimmu.2023.1072787
This article is part of the Research TopicThe Nutritional Immunological Effects and Mechanisms of Chemical Constituents from the Homology of Medicine and FoodView all 11 articles
 Nahed A. El-Shall1
Nahed A. El-Shall1 Shouqun Jiang2*
Shouqun Jiang2* Mayada R. Farag3
Mayada R. Farag3 Mahmoud Azzam4,5Abdulaziz A. Al-Abdullatif4Rashed Alhotan4Kuldeep Dhama6Faiz-ul Hassan7
Mahmoud Azzam4,5Abdulaziz A. Al-Abdullatif4Rashed Alhotan4Kuldeep Dhama6Faiz-ul Hassan7 Mahmoud Alagawany8*
Mahmoud Alagawany8*Increase in drug resistance as well as ineffective immunization efforts against various pathogens (viruses, bacteria and fungi) pose a significant threat to the poultry industry. Spirulina is one of the most widely used natural ingredients which is becoming popular as a nutritional supplement in humans, animals, poultry and aquaculture. It contains protein, vitamins, minerals, fatty acids, pigments, and essential amino acids. Moreover, it also has considerable quantities of unique natural antioxidants including polyphenols, carotenoids, and phycocyanin. Dietary supplementation of Spirulina can beneficially affect gut microbial population, serum biochemical parameters, and growth performance of chicken. Additionally, it contains polyphenolic contents having antibacterial effects. Spirulina extracts might inhibit bacterial motility, invasion, biofilm formation, and quorum sensing in addition to acting directly on the bacterium by weakening and making the bacterial cell walls more porous, subsequently resulting in cytoplasmic content leakage. Additionally, Spirulina has shown antiviral activities against certain common human or animal viruses and this capability can be considered to exhibit potential benefits against avian viruses also. Spirulan, a calcium-rich internal polysaccharide of Spirulina, is potentially responsible for its antiviral effect through inhibiting the entry of several viruses into the host cells, boosting the production of nitric oxide in macrophages, and stimulating the generation of cytokines. Comparatively a greater emphasis has been given to the immune modulatory effects of Spirulina as a feed additive in chicken which might boost disease resistance and improve survival and growth rates, particularly under stress conditions. This manuscript reviews biological activities and immune-stimulating properties of Spirulina and its potential use as a dietary supplement in poultry to enhance growth, gut health and disease resistance.
Numerous diseases continue to occur in the poultry industry despite the widespread use of vaccines and medications, resulting in financial losses. For instance, avian viral infections such as avian influenza (AI), infectious bronchitis (IB), infectious bursal disease (IBD), and Newcastle disease (ND) lead to significant economic losses, particularly in broilers, due to respiratory distress, increased mortality, reduced growth, and immunological suppression (1). The same is true for coccidiosis and bacterial infections like Salmonella, E. coli, etc. Until the emergence of resistant strains, chemical antimicrobials and antiprotozoals successfully inhibited and suppressed bacteria and protozoa. So there is a persistent need to use natural substances to address this issue. Algae, for instance, is a source of vital biological useful components, making the use of natural habitats as a source of these chemicals a viable strategy for creating innovative cuisines (2, 3). One of the greatest sources for organic nutrients among edible algae is the microscopic blue-green alga Spirulina (Arthrospira), which is used as a nutritional supplement for both human and animal feed globally (4). Spirulina platensis is a filamentous commercial cyanobacterium that is utilized as dietary and feed supplement in humans, aquaculture, livestock and poultry industry. Spirulina can grow in both saline and fresh water, and it is semi- and mass-cultivated in several countries. Dried spirulina is a rich nutritional source, with a high protein content (260-770 g/kg) representing 70% dry weight and a large fat content (10-140 g/kg) (5–7). Additionally, it has been observed that these microalgae have a high nutrient digestibility that was superior or equivalent to that of other vegetable diets and feeds (6, 8). Therefore, Spirulina has the potential to partially replace the traditional protein sources, particularly soybean meal (9). Oleic acid, linoleic acid, gamma-linolenic acid, docosahexaenoic acid (DHA), sulfolipids, and glycolipids are among the many polyunsaturated fatty acids found in spirulina, in addition to Omega-3 and -6 polyunsaturated fatty acids that are abundant in spirulina (25% and 60% of the total fatty acids) (5, 10). Spirulina also contains pigments, such as carotenoids (4000 mg/kg), which include β-carotene and zeaxanthin (10, 11), and chlorophyll pigments (12–14). Phycobiliproteins (15), vitamins (16), and macro- and micromineral components like calcium, iron, magnesium, manganese, potassium, zinc, and selenium are also found in spirulina (10, 17). Moreover, polysaccharides, pro-vitamin A, vitamin E, vitamin K and various B vitamins (10) as well as antioxidants are also important constituents of spirulina (18).
Spirulina is used as a dietary additive in a wide variety of food products due to its exceptional and impressive nutrient composition. This helps to improve the nutritional qualities of the products, as well as their potential to improve reproductive and productive performance, general health, and the symptoms of various animal diseases like arthritis, diabetes, anaemia, hypertension, and cardiovascular disorders. Spirulina are strong contenders as an alternative to antibiotics in chicken feed. These substances exhibited potential medicinal properties like antimicrobial, antioxidant, anti-cancer, anti-inflammatory, immune-enhancing, and colourants (18–20) in addition to their metalloprotective, radioprotective, and hypocholesterolemic effects (4, 10). Additionally, Spirulina (SPA) with antiviral properties has shown to strengthen the immune system, and its rich nutritional profile promoted growth performance by improving the intestinal villi length and number of the epithelial cells particlulary goblet cells (21). For a sustainable and feasible future of food security, spirulina is becoming a more affordable method of increasing poultry output (9). Here, therapeutic and immune-stimulating benefits of Spirulina were reviewed from another angle in case they could actually be used as nutritional supplement with antibiotic or vaccine to fight off various diseases of chicken (Figure 1).
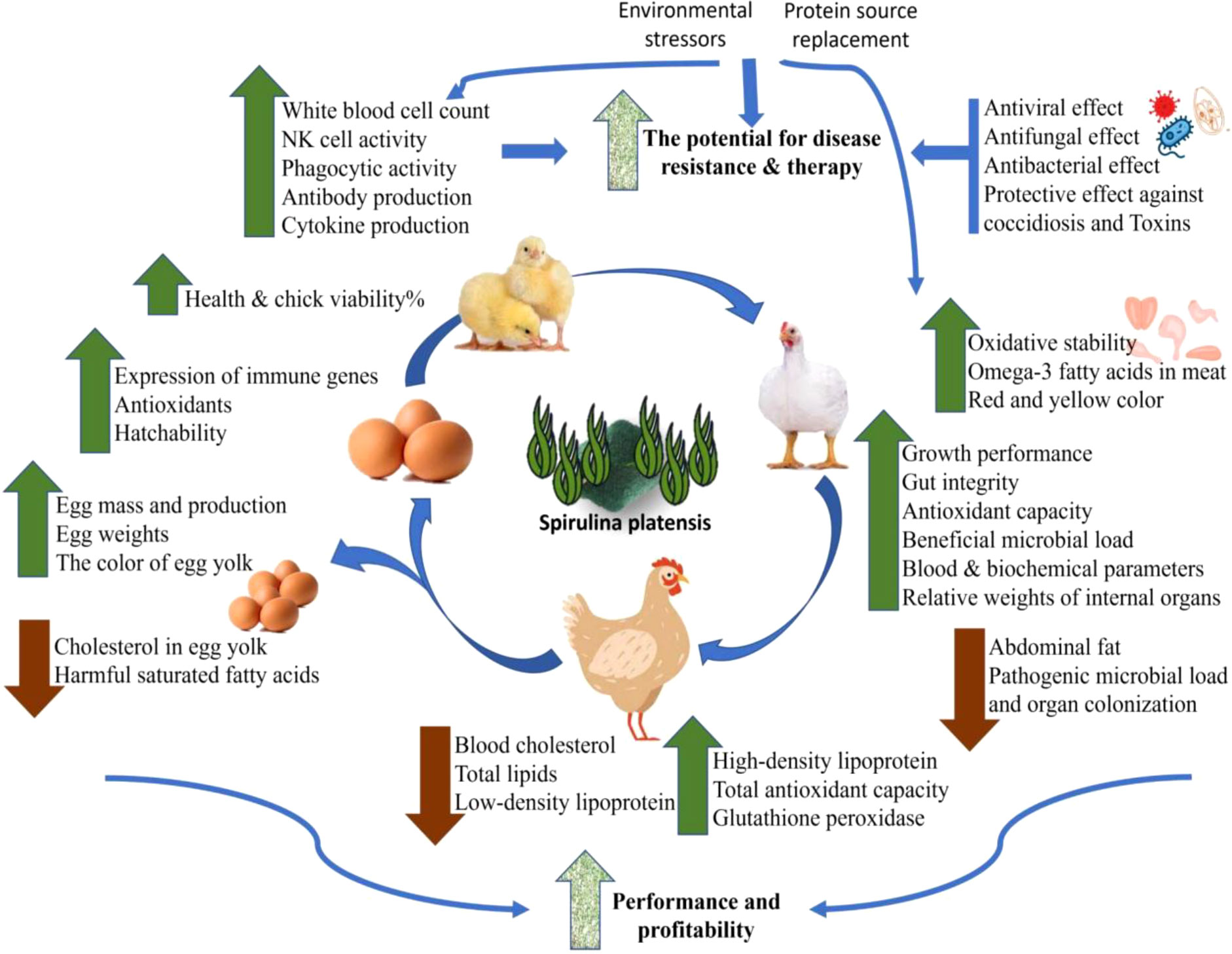
Figure 1 Overall applications and effects of Spirulina on poultry performance and health.
Numerous studies have supported spirulina’s ability to promote growth. From the embryonic stage until the egg is laid, spirulina can be supplemented in poultry diets. Spirulina platensis in ovo injection enhanced the expression of genes associated to immunity, antioxidants, and hatchability in quail chicks (22). Ibrahim et al. (23) reported that spirulina in drinking water for 4 weeks at levels of 0.5, 1 and 2 g/Liter significantly increased the average body weight gain, health, with the highest chick viability percent, best significant feed conversion ratio (FCR), feed efficiency, European Production Efficiency values, increase in the relative weights of carcass and internal organs with the significantly lowesr abdominal fat. Spirulina supplemented (1 or 2 g/kg diet) in diets containing vegetable and animal protein in Japanese quails improved the growth performance without affecting meat quality or gut flora in quails fed with vegetable protein source but no effect was observed in animal protein based diet (24).
K-strain White Cornell Leghorns and broiler chicks grown to ages of seven and three weeks, respectively, on diets containing varied concentrations (10, 100, 1,000, and 10,000 ppm) of Spirulina platensis exhibited higher body weights than control birds (25). With 0.7 and 0.9 g of Spirulina platensis per kg of feed, Cobb broiler chickens’ growth performance, blood parameters, biochemical changes in serum, and microbial load could all be improved (26).
Dietary supplementation of Spirulina at 10g/kg of diet showed significantly higher body weight gain and, consequently, linear improvements in FCR was observed in Cobb broilers during 35 days experimental period. Additionally, dietary Spirulina levels resulted in a rise in intestine Lactobacillus sp. while decreasing Escherichia coli populations (27).
Hajati and Zaghari (28) advocate utilizing Spirulina at dosages of 5 and 3 g/kg food during the Japanese quail’s growth and laying periods, respectively, using a range of varied doses. During the first 35 days of life, a diet of 5 g/kg induced a significant increase in body weight gain, breast percentage, and European production efficiency factor. When added to the diet of layers, Spirulina at a level of 3 g/kg considerably reduced the amount of cholesterol per gram of yolk while also improving the color of the egg yolks.
However, daily feed intake, FCR, the percentage of broken eggs, eggshell thickness, albumen height, Haugh unit, and egg weight were unaffected by dietary Spirulina up to 0.9%. When compared to non-supplemented birds, 0.6% algae significantly increased egg mass and production as well as egg yolk colour in laying chickens between 26 and 37 weeks of age (29).
For improved reproductive and productive performance, Mobarez et al. (30) suggested adding Spirulina to Golden Montazah layer diets. When given the basic diet with 2 or 3 g Spirulina/kg diet during the laying period from 29 to 40 weeks of age, hens and cocks had significantly improved FCR compared to the control group. Additionally, chickens fed a diet containing 3 g of Spirulina/kg had the highest levels of high-density lipoprotein (HDL), total antioxidant capacity (TAOC), glutathione peroxidase (GPx), egg quantity, egg weight, and hatchability percentage. Blood cholesterol, total lipids, and low-density lipoprotein (LDL) were all significantly reduced with Spirulina administration at both dosages.
Microalgae in quail diets (5%, 10%, and 15%) increased egg quality and provided benefits to consumer health by functioning as an immune-stimulant and antioxidant (31). However, they had no effect on egg production. This was because it boosted amounts of monounsaturated fatty acids, which are good for consumer health, and decreased levels of harmful saturated fatty acids. Additionally, egg yolk antioxidant levels increased, which reduced lipid peroxidation. On the other hand, 32 reported a lower performance of broiler chickens supplemented with Spirulina by 15%, for 2 weeks period (21st -35th day old), compared to non-supplemented birds. They attributed this negative effect to the high digesta viscosity induced by the gelation of indigestible proteins of Spirulina. Even addition of exogenous enzymes like lysozyme or Rovabio Excel AP resulted in the same worse findings, although lysozyme succeeded in breaking Spirulina’ cell wall. Pestana and hos co-authors suggested that this microalga’s proteins may be more easily digested and prevented from harmfully gelling if lysozyme and an exogenous specialised peptidase were combined.
Supplementing with spirulina boosts many immune processes. Spirulina has shown a specific action on monocytes and natural killer (NK) cells, which are essential components of the innate immune system. Administration of Spirulina exhibited to enhance macrophage phagocytic response and activity of NK cells in chicken and humans (25, 33, 34). The phagocytic activity of macrophages isolated from cats was also found to be increased in response to antigen exposure in the presence of Spirulina (25). A polysaccharide extract of Spirulina platensis has hsown to increase white blood cells in a haematopoietic system damaged by irradiation (35). Oral administration of Spirulina platensis in healthy male volunteers increased IFN-γ production and phagocytic activity of isolated NK cells stimulated with IL-12/18. Beside this, Spirulina also enhanced Toll like receptor (TLR)-2 and 4 mediated production of IL-12 from peripheral blood mononuclear cells, thus indicating Spirulina first activates monocytes and macrophages to produce cytokines that stimulate NK cells (34). An action through TLR-2 or -4, leading to NF-κB activation, has been suggested in studies in human monocytes (36, 37).
Spirulina’s immune-modulatory action on mice through increased IL-1 antibody production was observed in 1994 for the first time (38). In this regard, Kaoud (39) found that chicken groups fed diets containing spirulina had higher relative and absolute thymus and bursa weights than the control group. Similarly, in comparison to the untreated control, the addition of S. platensis at levels of 0.7 and 0.9 g/kg broiler diet resulted in a considerable rise in the weights of the bursa, thymus, and spleen (26). However, broilers and K-strain chicks given Spirulina (0, 10, 100, 1,000, and 10,000 ppm) did not change in bursal or splenic weight, but the K-strain chicks had significant larger thymuses than the controls (25).
A considerable rise in white blood cell count and increased macrophage phagocytic activity in broilers treated with S. platensis algae suggested that the immune system of the animals was strengthened (40).
According to Al-Batshan et al. (33) feeding Spirulina platensis increases macrophage phagocytic activity in terms of the average number of sheep red blood cells (SRBC) per phagocytic macrophage (range = 2.2 to 3.6 versus 1.8 to 2.5 in the basal group) and the overall phagocytic percentage (range = 28 to 39% versus 24 to 25% in the basal group). Over the course of the three developmental ages, Escherichia coli lipopolysaccharides-induced nitrite levels in macrophages (increased nitric oxide synthase activity) ranged from 60 to 278 microM in the basal diet group, but they were significantly higher in all Spirulina dietary groups (0.5% group range = 198 to 457 microM; 1.0% group range = 161 to 359 microM; and 2.0% group range = 204 to 420 microM).
Spirulina supplementation at 10000 ppm of diet also doubled the activity of NK cells and showed a greater PHA-P-mediated lymphoproliferative response compared to controls. All Spirulina groups (10, 100, 1000, 10000 ppm) demonstrated greater macrophages phagocytic capacity than the 0 ppm group in K-strain and broilers (25).
When compared to 1 g Spirulina/kg, laying Japanese quails given 3 or 5 g Spirulina/kg had significantly enhanced cutaneous basophil hypersensitivity after 12 or 24 hours of phytohemagglutinin injection (28). A substantially less heterophil and more lymphocytes than the control treatment were obtained by 1, 1.5, and 2 g spirulina/kg fed to broilers for 42 days (21). It has been demonstrated to increase the potential for disease resistance in chicken by activating their mononuclear phagocytic system (41). Improvement in cellular immunity observed in response to dietary supplementation of Sprirulina might be attributed to higher Zn concentration in spirulina like this (42, 43).
In Cornell K-strain White Leghorns and broiler chicks fed to 7 and 3 weeks of age, respectively, with meals containing varying amounts of Spirulina platensis (0, 10, 100, 1,000, and 10,000 ppm), anti-sheep red blood cell antibodies were not different throughout the initial reaction. However, all Spirulina-dietary groups with K-strain chicks exhibited greater total anti-SRBC titers during the secondary response, with the 10,000-ppm group having the highest (6.8 Log2) compared to the 0 ppm (5.5 Log2) group (25). In laying Japanese quails, different concentrations of Spirulina (1, 3 or 5 g/kg food) resulted in significantly greater levels of total antibody against SRBC and IgG titers (28).
Khan et al. (21) showed that the use of spirulina considerably enhanced growth performance, gut integrity, and immunity in broiler production while also providing better economics and supplementing with spirulina significantly affected the antibody titer against the ND vaccination.
In growing Japanese quail chicks, adding Spirulina at concentrations of 0.5, 1 and 2 g/Liter significantly raised the amount of serum antibodies against the Newcastle virus (NDV) and the plasma total protein profile (23).
Additionally, Golden Montazah laying hens and cocks supplemented with 3 g Spirulina/kg diet during the laying period recorded significantly higher antibody titers against NDV, Avian Influenza (AI), antibody against SRBC, and Interferon proteins (IFN- γ) concentration (30). Similarly Nia et al. (29) revealed that Spirulina by levels 0.3%, 0.6% or 0.9% in Lohmann Selected Leghorn (LSL) laying hens between the ages of 26 and 37 weeks had a substantial impact on the antibody titer in birds that had received the Newcastle vaccine. However, the ratio of heterophiles to lymphocytes, humoral immunity against SRBC, cell-mediated immunity response to PHA injection, and the relative weight of the bursa and spleen were not significantly affected by this dietary Spirulina.
Additionally, the immunosuppressive effects of diclofenac sodium, which were manifested by decreased phagocytic activity, phagocytic index, and a significant decrease in the titer of antibodies formed against NDV were significantly reversed by oral supplementation of broiler chicks with Spirulina at a dose of 10 gm/kg in their diet, either on therapeutic or preventive regimes (44).
Because microalgae are a significant source of C-phycocyanin, an antioxidant pigment with hypolipidemic activity, birds fed Spirulina showed improved antioxidant activity, which is another benefit of Spirulina feed (45, 46). Phycocyanin, carotene, and xanthophyll phytopigments, tocopherols, linolenic acid, and phenolic compounds are some examples of naturally occurring components in Spirulina that have been demonstrated to have strong antioxidant properties and potent scavenging activities against Reactive Oxygen Species like superoxide and hydrogen peroxide radicals (4).
Abdelkhalek et al. (47) reported that spirulina dramatically stimulates the activity of antioxidant enzymes preventing lipid peroxidation, DNA damage, and free radical scavenging. The total antioxidant capacity and thyroxin (T4) content were dramatically enhanced in growing Japanese quail chicks when Spirulina levels of 0.5, 1 and 2 g/Liter were used. Plasma cholesterol, total lipids, ALP, ALT, and AST activity, however, had significantly lower levels (23).
Additionally, in ovo injections of Arthrospira (Spirulina) platensis at doses of 0.75–3.5 mg/egg in Japanese quails and 25–35 mg/egg in broiler breeders enhanced chick hatchability and IFN-gamma expression. In quails, 2.5 or 3.5 mg Spirulina dramatically reduced expression of HSP70 and considerably boosted Catalase activity and GPX gene expression in hatchlings. The lowest HSP70 in chicks was induced by in ovo injection of 25 or 35 mg Spirulina in broiler breeders (22).
Moreover, spirulina inclusion in the poultry feed affects meat quality parameters like color, flavor, polyunsaturated fatty acids composition, and oxidative stability. Therefore, in terms of oxidative stability and the enhancement of omega-3 fatty acids like those of linolenic and docosahexaenoic acid by Spirulina feeding, 1.5% fed to Cobb 500 broiler chicks for 5 weeks could be potential functional ingredients to generate value-added broiler meat (breast and thigh meat) (48). Similarly, Abbas et al. (49) demonstrated that 3% and 4% of dietary Spirulina significantly increased the content of oleic acid, palmitic acid, docosahexaenoic acid, and linoleic acid in broiler carcass. Additionally, throughout the 30-day storage period, 4% Spirulina considerably decreased the value of peroxide. For the 60-day storage period, both Spirulina treatments significantly reduced the oxidation indices of total volatile nitrogen (TVN) and thiobarbituric acid (TBA), while the sensory evaluation ratings for the chest and thigh slices did not change. 50 reported an unchange of the amount of gamma-linolenic acid or omega-3 in the intramuscular fat by 75% and 50% Soy bean replacement by Spirulina in starter and grower feed, respectively, although the richness of Spirulina with poly unsaturated fatty acids. Moreover, when Spirulina-based meat samples were wrapped in highly oxygenated modified environment packaging, they showed higher rates of lipid oxidation than soybean meal-based meat samples (50). On another side, color of meat was improved by inclusion of spirulina at high doses in the feed that may be due to high level of carotenoids. Altmann et al. (51) found that when Spirulina replaces 50% of the soy protein in broiler diets, meat color could be increased to be dark reddish-yellowish flesh. Similarly, Altmann et al. (50) observed a more intensive color (higher red (a*) and yellow (b*)) for breast and thigh meat as well as an increased umami and chicken flavor were reported for broiler meat. 51 reported a positive effect of Spirulina on flavour breast filets through decreasing the score of metallic flavor (off-flovor) of breast meat, besides increasing pH value after 24 hours after death that was associated with softness and tenderness of breast filets. Feeding broiler chickens with Spirulina 15% for two, the breast and thighs exhibited higher values of yellowness (b*) compared to the control, as well as total carotenoids and saturated fatty acids were increased, but levels of n-3 polyunsaturated fatty acids and α-tocopherol were reduced (32). Neverthless, Park et al. (52) found that the quality of the breast meat of broilers fed Spirulina diets (0.25, 0.5, 0.75, or 1.0%) for 35 days was not substantially altered while seven-day drip loss was linearly reduced.
Dietary spirulina have shown good response in several infections in vivo in poultry (Table 1). A wide variety of influenza viruses, including oseltamivir-resistant strains, were blocked by the cold-water Spirulina extract from forming viral plaques. Inhibition of influenza hemagglutination was revealed to be one of the ways by which spirulina extract acts early in the course of infection to lower viral production in cells and increase survival in influenza-infected mice (58). The ethanol extract of Spirulina platensis had an antiviral effect in vitro, decreasing the infectious units of adenovirus types 7, Coxsackievirus B4, astrovirus types 1, rotavirus Wa strain, and adenovirus types 40 by respective amounts of 53.3%, 66.7%, 76.7%, 56.7%, and 50% (59).
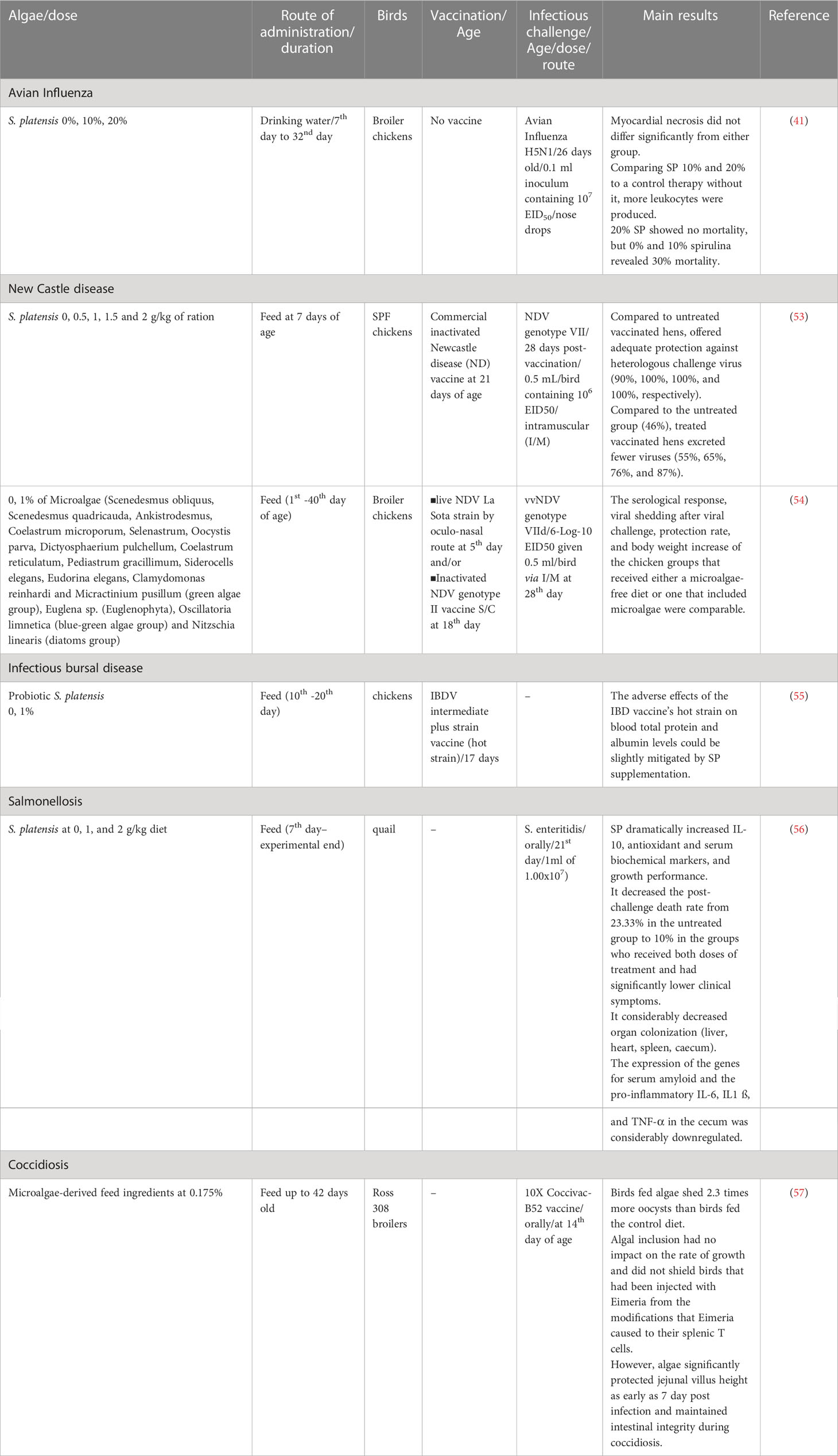
Table 1 Efficacy of Spirulina platensis (SP) supplementation in cases of various infectious diseases in poultry.
It was reported that a calcium-rich intracellular polysaccharide called spirulan found in Spirulina platensis prevents multiple viruses from replicating in vitro by preventing the virus’ entry into the various host cells that are being utilized (60, 61), increases macrophage nitric oxide synthesis and stimulates cytokine production (62). Broiler hens infected with the H5N1 Avian Influenza virus had cardiac necrosis, although 20% of spirulina had no discernible effect on this. However, it boosted leukocytes associated with an immune function, which prevented mortality vs 30% death in the non-supplemented group (41).
Regarding Newcastle disease, most investigations such as (53, 54) found that Spirulina had an impact on viral challenge in birds that had received live, attenuated, and/or inactivated vaccinations. As a result, the effect of Spirulina supplementation was seen as an immune-stimulating effect that dramatically improved clinical protection against heterologous strains and the capacity to decrease NDV shedding. In 2019, Kumari et al. discussed an indirect benefit of spirulina on infectious bursal disease. The decreasing effect of intermediate plus vaccine (hot strain given at the age of 17 days) on serum total protein concentration was greatly reduced by spirulina supplementation at 1.0% in feed from 10 to 20 days of age.
Cyanobacteria might be considered a suitable source for the manufacturing of antimicrobial agents as purified antimicrobial compound produced by S. platensis were more effective against Gram positive, Gram negative, and C. albicans, a unicellular fungus. Organic and aqueous extracts of S. platensis were tested in vitro and demonstrated broad-spectrum antibacterial and antifungal action. 63 showed that the highest biological activity of them was against Escherichia coli, Pseudomonas aeruginosa, Bacillus subtilis and Aspergillus niger. Compared to ethanol or aqueous extracts, methanol extract exhibited superior antibacterial activity against all tested bacterial strains (Gram positive bacteria, Gram negative bacteria, and Candida sp.), particularly against Gram positive bacteria (Staphylococcus aureus, Streptococcus pneumoniae, Bacillus cereus, and Enterococcus faecalis). For the various strains examined, the lowest inhibitory concentration value of ethanol and methanol extract ranged between 5-100 mg/mL (64). Similarly, compared to other extracts, the methanolic extract of Spirulina showed a larger total phenolic content and more antibacterial activity (65, 66).
Kaushik and Chauhan (67) reported that the minimum inhibitory concentrations (MIC) of the methanol extract against S. aureus and E. coli were 128 g/ml and 256 g/ml, respectively while it had no effect was against Klebsiella pneumoniae. Nevertheless, Spirulina acetone extract shown strong biological activity against Klebsiella pneumoniae and modest activity against Salmonella typhi and Pseudomonas aeruginosa (68). By using an ethanol extract of Spirulina platensis, inhibitory zones against Enterococcus faecalis and Candida albicans were seen using the disc diffusion technique (59). The effectiveness of Spirulina’s antibacterial actions in vivo was assessed by Abd El-Dayem et al. (56) as adding 1 and 2 g of spirulina per kg of diet significantly increased growth performance, antioxidant levels, and the production of the anti-inflammatory cytokine (IL-10) while reducing organ colonization and gene expressions of IL-6, IL-1ß, and TNF-α as compared to positive and negative control groups.
According to 69, pathogens use the same adhesion and invasion mechanisms to invade the guts of both people and animals, and the antibacterial action of spirulina might be attributed to its ability to prevent pathogen motility, invasion, biofilm formation, and quorum sensing. In addition, Spirulina’s bioactive ingredients have shown to weaken bacterial cell walls making it more permeable, that subsequently led to leakage of cytoplasmic contents (65).
Furthermore, when broilers were given the 10X Coccivac-B52 vaccine orally while being fed microalgae-derived feed components, the intestinal integrity of the birds during coccidiosis was conserved, and the jejunal villus height was protected as early as 7 days after the challenge (57). However, the algal element changed the immune response (splenic T cells) in a way that decreased recruitment from secondary lymphoid organs (57).
Spirulina was also a powerful binder for aflatoxins in broiler breeders (70). The negative effect of 300 ppb aflatoxin on broiler chicken growth rate and lymphoid organ weights might be partially mitigated by the addition of spirulina at a level of (0.05%) although there was no positive impact on feed consumption, the serum protein concentration, liver weights, or cholesterol levels (71). Likewise, Spirulina inclusion (0.02%) in feed had positive effects on growth, ready to cook yields, bursa weight, and cellular immune response in chicks fed aflatoxin (300 ppb) although there was no effect of Spirulina on feed intake, feed conversion efficiency, leg abnormality scores, SRBC response, and weights of the liver, giblets, spleen, and abdominal fat (72). Dietary inclusion of Spirulina is also recommended to protect against other toxins as shown in Table 1.
A crucial component of broiler productivity, especially with the use of antibiotics being reduced, is immune system health. Natural antibiotic alternatives and nutritional factors like crude protein % are being studied to understand how they affect immunity. The positive benefits of Spirulina in reversing the management and nutrient deficits in poultry were displayed in Table 2. A promising development for reducing feed costs without compromising the health of the bird is the ability of natural feed additives to counteract the negative effects that low crude protein has on immunity in birds (74). Spirulina is an alternative protein-containing component that manufacturers are considering because of both its great nutritional value and its capacity to strengthen the immune system. For instance, the addition of spirulina to low protein chicken diets resulted in reduction of systemic inflammation and bacterial translocation indicating its suitability as a alternative protein source (73). Additionally, the low crude protein (LCP) diet-Spirulina supplemented reversed the effects of the LCP diet, causing the monocyte proportions and concentrations in Ross 708 old male broilers to be similar statistically to those of the control base diet (74). However, compared to 20% crude protein without Spirulina, using a diet of 17% crude protein with 10% S. platensis for Ross 708 broiler females exhibited no detrimental effects on the health of broilers but substantial reduction in body weight in supplemented group led to economoic losses (75).
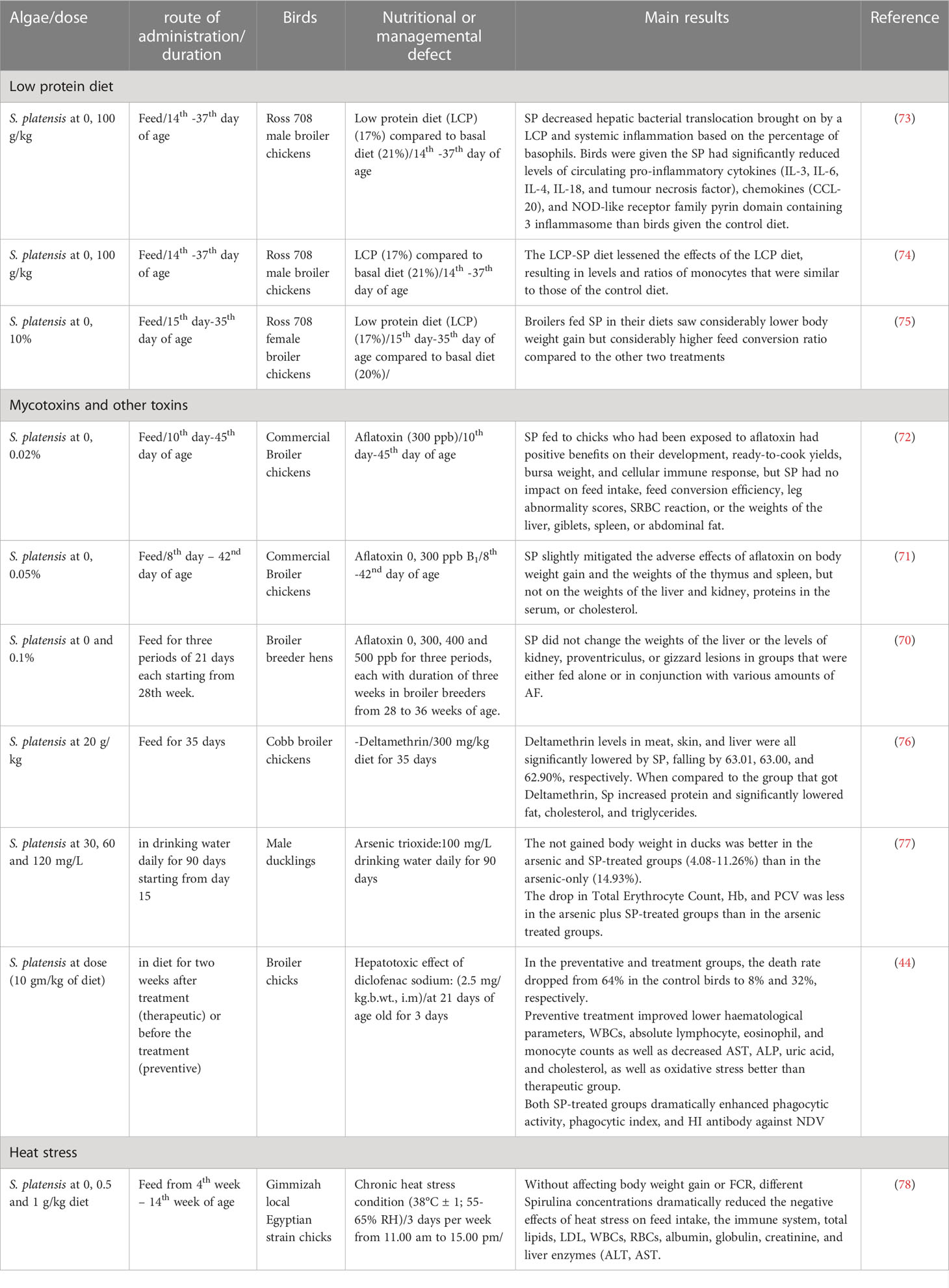
Table 2 Efficacy of Spirulina platensis (SP) in cases of nutritional deficiencies and managemental defects in poultry.
Dietary supplementation of S. platensis in broiler hens has shown to alleviate the adverse effects of high ambient temperature, including impaired enzymatic antioxidant system, raised stress hormone, and altered lipid profile (84). There was a dose-related modification of productivity, physiological, and immunological parameters when chickens under heat stress were given Spirulina in drinking water or feed (78, 82). Additionally, in ovo injection of S. platensis improved the broiler embryo’s ability to tolerate heat in the last days of incubation (22).
Spirulina as a feed additive can have implications for productivity and end product quality, depending on the system of animal production. Both swine growth performance and product quality were not negatively affected in response to dietary supplementation with spirulina which might be attributed to lower protein requirements in finishing diets. However, Spirulina negatively affected chicken and fish production performance besides altering product quality, particularly meat color, according to the consumer’s opinions (85). One of major challenges regarding use of Spirulina as a feed addtitve at high level is that the gelation of its indigestible proteins causing birds to perform worse due to the increased digesta viscosity (32). Therefore the golden standard level of Spirulina inclusion into the feed should be highlighted and interpreted to be applied in the field. In addition, trials to increase digistability of Spirulina should be rescearced. Another major challenge regarding use of Spirulina as a feed addtitve is its quite higher cost compared to other protein ingredients such as soybean meal (58, 86). However, improving production efficiency and using waste streams as culture media, spirulina could replace fishmeal by becoming competitive for fish feed due to higher cost of fishmeal (86). Second major challenge with spirulina is limitation of its large scale production as presently it is produced at a smaller scale primarily for the nutritional supplement sector with few exceptions (58). Third challenge is with sustainable production of spirulina as compared to the other protein ingredients like soybean which is mainly attributed to the sensitivity of spirulina to the production system and regional climate (87). Therefore serious research and development efforts are required to improve yield of spriulina and make its production more sustainable. For example some research initiative targeted to improve sustainability of production by using biogas effluent (88) or wastewater (89) as production media. Additionally, waste heat sources (e.g. heat produced during biogas production) can be integrated as spirulina requires warm temperature (35–37°C) for cultivation (5, 90). Consequently, above mentioned challenges could be overcomed by upscaling and optimizing production of spirulina. Moreover, advanced techniques can also fascilitate improvement in yield and protein quality of spirulina through breeding and Finally, although spirulina has a high proportion of crude protein, improvements to protein quality could be possible through breeding and nutrition/production research (91). Therefore, future research focused on sustainable production and product processing and acceptance should investigate the trade-offs of incorporating spirulina into poultry diets.
Bioactive metabolites are abundant in natural products and have been used for their medicinal properties. Spirulina that was regarded as a blue-green filamentous algae with a spiral shape, it has been identified as a genus of photosynthetic bacteria (Arthrospira) more recently. It is a highly nutritious and antioxidant natural product and having the ability to improve production performance either growth, hatchability, or egg production. The cell-mediated and humoral immune response as well as antimicrobial activities of spirulina promoted disease resistance and improved survival and growth rates in chicken. However, further studies on optimum dose of Spirulina for different poultry species, age groups, and production systems as well as the type of used Spirulina extract, organic or aqueous are required.
All the authors contributed significantly to this manuscript. NAE conceptualized the manuscript. NAE wrote the first draft with input from SJ, MRF, MA, AAA, RA, KD, FH, and MAl. Authors NAE, SJ, MRF, MA, AAA, RA, KD, FH, and MAl. reviewed and updated the manuscript. All authors contributed to revisions and approved the final manuscript.
China Agriculture Research System of MOF and MARA (CARS-41), the Key Realm R&D Program of Guangdong Province (2020B0202090004), National Key R&D Project (2021YFD1300404), Natural Science Foundation from Guangdong Province (2022A1515012069, 2021A1515012412, 2021A1515010830), the Science and Technology Plan Project of Guangzhou (202206010168), the Science and Technology Program of Guangdong Academy of Agricultural Sciences (202106TD, R2019PY-QF008), Introduction of Talents Program from Guangdong Academy of Agricultural Sciences (R2021YJ-YB3012), Guiding Agreement of Young Scholar from Guangdong Academy of Agricultural Sciences (R2021QD-024) P. R. China.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. El-Shall NA, Shewita RS, Abd El-Hack ME, AlKahtane A, Alarifi S, Alkahtani S, et al. Effect of essential oils on the immune response to some viral vaccines in broiler chickens, with special reference to Newcastle disease virus. Poult Sci (2020) 99(6):2944–54. doi: 10.1016/j.psj.2020.03.008
2. Lordan S, Paul Ross R, Stanton C. Marine bioactives as functional food ingredients: Potential to reduce the incidence of chronic diseases. Mar Drugs (2011) 9:1056–100. doi: 10.3390/md9061056
3. Draaisma RB, Wijffels RH, Slegers PE, Brentner LB, Roy A, Barbosa MJ. Food commodities from microalgae. Curr Opin Biotechnol (2013) 24:169–77. doi: 10.1016/j.copbio.2012.09.012
4. Farag MR, Mahmoud A, Mohamed Ezzat Abd E-H, Kuldeep D. Nutritional and healthical aspects of spirulina (Arthrospira) for poultry. Anim Human. Int J Pharmacol (2016) 12:36–51. doi: 10.3923/ijp.2016.36.51
5. Habib MAB, Parvin M, Huntington TC, Hasan MR.. A review on culture, production and use of spirulina as food for humans and feeds for domestic animals and fish. In: FAO Fisheries and Aquaculture Circular Rome: Food and agrculture organization of the United States Nations (2008), 1034.
6. Alvarenga RR, Rodrigues PB, Cantarelli V, Zangeronimo MG, Da Silva Junior JW, Da Silva LR, et al. Energy values and chemical composition of spirulina (Spirulina platensis) evaluated with broilers. Braz J Anim. Sci (2011) 40:992–6. doi: 10.1590/S1516-35982011000500008
7. Zahroojian N, Moravej H, Shivazad M. Effects of dietary marine algae (Spirulina platensis) on egg quality and production performance of laying hens. J Agric Sci Technol (2013) 15:1353–60.
8. Plaza M, Herrero M, Cifuentes A, Ibanez E. Innovative natural functional ingredients from microalgae. J Agric Food Chem (2009) 57:7159–70. doi: 10.1021/jf901070g
9. Deepika DS, Sowmya KL, Kiran KR. Efficacy of microalgal biomass in poultry nutrition journal of poultry fisheries & wildlife sciences. Poult Fish Wildl Sci (2021) 9(5):1000216.
10. Ghaeni M, Roomiani L. Review for application and medicine effects of spirulina, Spirulina platensis microalgae. J Advanced Agric Technol (2016) 3 (2). doi: 10.18178/joaat.3.2.114-117
12. Panyakampol J, Cheevadhanarak S, Sutheeworapong S, Chaijaruwanich J, Senachak J, Siangdung W, et al. Physiological and transcriptional responses to high temperature in arthrospira (Spirulina) platensis C1. Plant Cell Physiol (2014) 56(3):481–96. doi: 10.1093/pcp/pcu192
13. Ciferri O, Tiboni O. The biochemistry and industrial potential of spirulina. Ann Rev Microbiol (1985) 39:503–26. doi: 10.1146/annurev.mi.39.100185.002443
14. Ronda RR, Lele SS. Culture conditions stimulating high ylinolenic acid accumulation by Spirulina platensis. Braz J Microbiol (2008) 39:693–7. doi: 10.1590/S1517-83822008000400018
15. Eriksen NT. Production of phycocyanin - a pigment with applications in biology, biotechnology, foods and medicine. Appl Microbiol Biotechnol (2008) 80:1–14. doi: 10.1007/s00253-008-1542-y
16. Becker EW. Nutrition. In: Microalgae: Biotechnology and microbiology. Cambridge: Cambridge University Press (1994). p. 196–249.
17. Spolaore P, Joannis-Cassan C, Duran E, Isambert A. Commercial applications of microalgae. J Biosci Bioeng (2006) 101:87–96. doi: 10.1263/jbb.101.87
18. Christaki E, Bonos E, Giannenas I, Florou-Paneri P. Functional properties of carotenoids originating from algae. J Sci Food Agric (2013) 93:5–11. doi: 10.1002/jsfa.5902
19. Freitas AC, Rodrigues D, Rocha-Santos TAP, Gomes AMP, Duarte AC. Marine biotechnology advances towards applications in new functional foods. Biotechnol Adv (2012) 30:1506–15. doi: 10.1016/j.biotechadv.2012.03.006
20. Batista AP, Gouveia L, Bandarra NM, Franco JM, Raymundo A. Comparison of microalgal biomass profiles as novel functional ingredient for food products. Algal Res (2013) 2:164–73. doi: 10.1016/j.algal.2013.01.004
21. Khan S, Mobashar M, Mahsood FK, Javaid S, Abdel-Wareth AA, Ammanullah H, et al. Spirulina inclusion levels in a broiler ration: evaluation of growth performance, gut integrity, and immunity. Trop Anim Health Production (2020) 52:3233–40. doi: 10.1007/2Fs11250-020-02349-9
22. Hajati H, Zaghari M, Noori O, Negarandeh R, Cedraz de Oliveira H. Effects of in ovo injection of microalgae on hatchability, antioxidant and immunity-related genes expression, and post-hatch performance in broilers and Japanese quails. Ital J Anim Sci (2021) 20(1):985–94. doi: 10.1080/1828051X.2021.1910582
23. Ibrahim NS, Wakwak MM, Al-Gama MA. Productive performance and immune response in growing japanese quail supplemented with spirulina algae extract (arthrospira platensis) in drinking water. Egypt.Poult.Sci (2018) 38)(II):409–26.
24. Yusuf MS, Hassan MA, Abdel-Daim MM, Nabtiti AS, Ahmed AM, Moawed SA, et al. Value added by Spirulina platensis in two different diets on growth performance, gut microbiota, and meat quality of Japanese quails. Vet World. (2016) 9(11):1287–93. doi: 10.14202/vetworld.2016.1287-1293
25. Qureshi MA, Garlich JD, Kidd MT. Dietary Spirulina platensis enhances humoral and cell-mediated immune functions in chickens. Immunopharmacol Immunotoxicology (1996) 18:3:465–76. doi: 10.3109/08923979609052748
26. Fathi MA, Namra MMM, Ragab MS, Aly MMM. Effect of dietary supplementation of algae meal (Spirulina platensis) as growth promoter on performance of broiler chickens. Egypt. Poult. Sci J (2018) 38:375–89.
27. Alwaleed EA, El-Sheekh M, Abdel-Daim MM, Saber H. Effects of Spirulina platensis and amphora coffeaeformis as dietary supplements on blood biochemical parameters, intestinal microbial population, and productive performance in broiler chickens. Environ Sci pollut Res Int (2021) 28(2):1801–11. doi: 10.1007/s11356-020-10597-3
28. Hajati H, Zaghari M. Effects of Spirulina platensis on growth performance, carcass characteristics, egg traits and immunity response of Japanese quails. Iranian J Appl Anim Sci (2019) 9(2):347–57.
29. Nia EA, Mehri M, Shirmohammad F. Effects of dietary Spirulina platensis algae powder on performance, egg quality, ovarian follicles and immune system in lohmann LSL laying hens. Iran. J Anim. Sci (2021) 51(4):349–59. doi: 10.22059/ijas.2021.314088.653810
30. Mobarez SM, Rizk AM, Abdel latif AM, Osama AE. Effect of supplementing diet with Spirulina platensis algae or turmeric on productive and reproductive performance of golden montazah layers. Egypt. Poult. Sci J (2018) 38:109–25.
31. Boiago MM, Dilkin JD, Kolm MA, Barreta M, Souza CF, Baldissera MD, et al. Spirulina platensis in Japanese quail feeding alters fatty acid profiles and improves egg quality: Benefits to consumers. J Food Biochem (2019) 43(7):e12860. doi: 10.1111/jfbc.12860
32. Pestana JM, Puerta B, Santos H, Madeira MS, Alfaia CM, Lopes PA, et al. Impact of dietary incorporation of spirulina (Arthrospira platensis) and exogenous enzymes on broiler performance, carcass traits, and meat quality. Poult Sci (2020) 99(5):2519–32. doi: 10.1016/j.psj.2019.11.069
33. Al-Batshan HA, Al-Mufarrej SI, Homaidan AA, Qureshi MA. Enhancement of chicken macrophage phagocytic function and nitrite production by dietary Spirulina platensis. Immunopharmacol. Immunotoxicol. (2001) 23:281–9. doi: 10.1081/IPH-100103866
34. Hirahashi T, Matsumoto M, Hazeki K, Saeki Y, Ui M, Seya T. Activation of the human innate immune system by spirulina: augmentation of interferon production and NK cytotoxicity by oral administration of hot water extract of Spirulina platensis. Int Immunopharmacol. (2002) 2:423–34. doi: 10.1016/S1567-5769(01)00166-7
35. Zhang HQ, Lin AP, Sun Y, Deng YM. Chemo- and radio-protective effects of polysaccharide of Spirulina platensis on hemopoietic system of mice and dogs. Acta Pharmacol Sin (2001) 22:1121–4.
36. Balachandran P, Pugh ND, Ma G, Pasco DS. Toll-like receptor 2-dependent activation of monocytes by spirulina polysaccharide and its immune enhancing action in mice. Int Immunopharmacol. (2006) 6:1808–14. doi: 10.1016/j.intimp.2006.08.001
37. Kawanishi Y, Tominaga A, Okuyama H, Fukuoka S, Taguchi T, Kusumoto Y, et al. Regulatory effects of spirulina complex polysaccharides on growth of murine RSV-m glioma cells through toll-like receptor 4. Microbiol Immunol (2013) 57:63–73. doi: 10.1111/1348-0421.12001
38. Hayashi O, Hirahashi T, Katoh T, Miyajima H, Hirano T, Okuwaki Y. Class specific influence of dietary Spirulina platensis on antibody production in mice. J Nutr Sci Vitaminol (Tokyo). (1998) 44:841–51. doi: 10.3177/jnsv.44.841
39. Kaoud HA. Effect of Spirulina platensis as a dietary supplement on broiler performance in comparison with prebiotics. J Biol Sci (2015) 1:1–6.
40. Mariey Y, Samak H, Abou-Khashba H, Sayed M, Abou-Zeid A. Effect of using Spirulina platensis algae as a feed additives for poultry diets: 2 productive performance of broiler. Egypt. Poult. Sci (2014) 34:245–58.
41. Lokapirnasari WL, Yulianto AB, Legowo D, Agustono. The effect of spirulina as feed additive to myocardial necrosis and leukocyte of chicken with avian influenza (H5N1) virus infection. Proc Chem (2016) 18:213–217. doi: 10.1016/j.proche.2016.01.033
42. Abdel-Daim MM, Abuzead SM, Halawa SM. Protective role of Spirulina platensis against acute deltamethrininduced toxicity in rats. PloS One (2013) 8:e72991. doi: 10.1371/journal.pone.0072991
43. Abou-Gabal A, Aboul-Ela HM, Ali E, Ahemd E, Shalaby OK. Hepatoprotective, DNA damage prevention and antioxidant potential of Spirulina platensis on CCl4-induced hepatotoxicity in mice. Am J Biomedi. Res (2015) 3(2):29–34. doi: 10.12691/ajbr-3-2-3
44. Mokhbatly AA, Ghazy EW, Dabdelhady DH, Walid A, Abdelwahab M. Ameliorative effect of Spirulina platensis against diclofenac sodium-induced hepathotoxicity in broilers: hematological, biochemical, biological, antioxidant, immunological and histopathological study. Assiut Vet Med J (2018) 64 No. 156:164–77.
45. Khan Z, Bhadouria P, Bisen PS. Nutritional and therapeutic potential of spirulina. Curr Pharm Biotechnol (2005) 6:373–9. doi: 10.2174/138920105774370607
46. Wu LC, Ho JAA, Shieh and I.W. Lu MC. Antioxidant and antiproliferative activities of spirulina and chlorella water extracts. J Agric Food Chem (2005) 53:4207–12. doi: 10.1021/jf0479517
47. Abdelkhalek NK, Ghazy EW, Abdel-Daim MM. Pharmacodynamic interaction of Spirulina platensis and deltamethrin in freshwater fish Nile tilapia, oreochromis niloticus: impact on lipid peroxidation and oxidative stress. Environ Sci pollut Res (2015) 22(4):3023–31. doi: 10.1007/s11356-014-3578-0
48. Sharmin F, Sarker NR, Sarker MSK. Effect of using moringaoleifera and spirulinaplatensis as feed additives on performance, meat composition and oxidative stability and fatty acid profiles in broiler chicken. J Nutr Food Sci (2020) 10:772. doi: 10.35248/2155-9600.20.10.772
49. Abbas MS, Bandar LK, Alkhilani FMH. (2021). Effect of using different levels of spirulina algae (Spirulina platensis) in the diet on concentration, types of fatty acids, oxidation indicators, and sensory characteristics of broiler carcasses. IOP Conf Ser.: Earth Environ Sci 910:012044. doi: 10.1088/1755-1315/910/1/012044
50. Altmann BA, Wigger R, Ciulu M, Mörlein D. The effect of insect or microalga alternative protein feeds on broiler meat quality. J Sci Food Agric (2020) 100:4292–302. doi: 10.1002/jsfa.10473
51. Altmann BA, Neumann C, Velten S, Liebert F, Mörlein D. Meat quality derived from high inclusion of a micro-alga or insect meal as an alternative protein source in poultry diets: A pilot study. Foods (2018) 7:34. doi: 10.3390/foods7030034
52. Park JH, Lee SI, Kim IH. Effect of dietary spirulina (Arthrospira) platensis on the growth performance, antioxidant enzyme activity, nutrient digestibility, cecal microflora, excreta noxious gas emission, and breast meat quality of broiler chickens. Poult Sci (2018) 97(7):2451–9. doi: 10.3382/ps/pey093
53. Abotaleb MM, Mourad A, Abousenna MS, Helal AM, Nassif SA, Elsafty MM. The effect of spirulina algae on the immune response of SPF chickens to commercial inactivated Newcastle vaccine in poultry. VacciMonitor (2020) 29(2):51–7.
54. Abdo SM, Sameh Abdel-Moez A, Hagar Magdy A, Rehab Hamdy M, Abeer Abdallah S, Mohamed Abdel-Aziz K. Microalgae biomass application in commercial broilers nutrition and their efficacy against challenge with epidemic Newcastle disease virus in Egypt. J World Poult. Res (2019) 9(2):98–108. doi: 10.36380/jwpr.2019.12
55. Kumari P, Kundu P, Kajal S, Narang G. Effect of spirulina feeding on serum protein level in infectious bursal disease vaccinated chickens. Pharma Innovation J (2019) 8(2):543–6.
56. Abd El-Dayem GA, Saleh GK, Abd EL-Elwahab RA. Impact of dietary spirulina (arthrospira) platensis on growth performance, gene expression and antioxidant status of quail challenged with salmonella enteritidis. Mansoura Veterinary Med J (2021) 22:2:38–47. doi: 10.35943/mvmj.2021.66416.1040
57. Fries-Craft K, Meyer MM, Bobeck EA. Algae-based feed ingredient protects intestinal health during eimeria challenge and alters systemic immune responses with differential outcomes observed during acute feed restriction. Poult Sci (2021) 100(9):101369. doi: 10.1016/j.psj.2021.101369
58. Chen YH, Chang GK, Kuo SM, Huang SY, Hu IC, Lo YL, et al. Well-tolerated spirulina extract inhibits influenza virus replication and reduces virus-induced mortality. Sci Rep (2016) 6:24253. doi: 10.1038/srep24253
59. El-Baz FK, El-Senousy WM, El-Sayed AB, Kamel MM. In vitro antiviral and antimicrobial activities of Spirulina platensis extract. J Appl Pharm Sci (2013) 3:52–6. doi: 10.7324/JAPS.2013.31209
60. Hayashi T, Hayashi K, Maeda M, Kojima I. Calcium spirulan, an inhibitor of enveloped virus replication, from a blue-green alga Spirulina platensis. J Nat Prod (1996) 59:83–7. doi: 10.1016/j.intimp.2005.09.009
61. Hayashi K, Hayashi T, Kojima I. A natural sulphated polysaccharide, calcium spirulan, isolated from Spirulina platensis: In vitro and ex vivo evaluation of anti-herpes simplex virus and anti-human immunodeficiency virus. AIDS Res Hum Retrovir (1996) 12:1463–71. doi: 10.1089/aid.1996.12.1463
62. Bae SY, Yim JH, Lee HK, Pyo S. Activation of murine peritoneal macrophages by sulphated exopolysaccharide from marine microalga gyrodinium impudicum (strain KG03): Involvement of the NF-kappa b and JNK pathway. Int Immunopharmacol. (2006) 6:473–484. doi: 10.1016/j.intimp.2005.09.009
63. El-Sheekh MM, Daboor SM, Swelim MA, Mohamed S. Production and characterization of antimicrobial active substance from Spirulina platensis. Iran. J Microbiol (2014) 6:112–119.
64. Al-ghanayem A. Antimicrobial activity of Spirulina platensis extracts against certain pathogenic bacteria and fungi. Adv Biores. (2017) 8(6):96–101. doi: 10.15515/abr.0976-4585.8.6.96101
65. Abdel-Moneim EA-M, Mohamed TE-S, Abdelrazeq MS, Ahmed MS, Aldhumri SA, Ouda SM, et al. Antioxidant and antimicrobial activities of Spirulina platensis extracts and biogenic selenium nanoparticles against selected pathogenic bacteria and fungi. Saudi J Biol Sci (2021) 29(2):1319–562X. doi: 10.1016/j.sjbs.2021.09.046
66. Gheda SF, Ismail GA. Natural products from some soil cyanobacterial extracts with potent antimicrobial, antioxidant and cytotoxic activities. Anais da Academia Bras Ciências (2020) 92. doi: 10.1590/0001-3765202020190934
67. Kaushik P, Chauhan A. In vitro antibacterial activity of laboratory grown culture of Spirulina platensis. Indian J Microbiol (2008) 48:348–52. doi: 10.1007/s12088-008-0043-0
68. Mala R, Sarojini M, Saravanababu S, Umadevi G. Screening for antimicrobial activity of crude extracts of Spirulina platensis. J Cell Tissue Res (2009) 9:1951–5.
69. Saleh AA, Shukry M, Farrag F, Soliman MM, Abdel-Moneim A-ME. Effect of feeding wet feed or wet feed fermented by bacillus licheniformis on growth performance, histopathology and growth and lipid metabolism marker genes in broiler chickens. Animals (2021) 11:83. doi: 10.3390/ani11010083
70. Manafi M. Evaluation of different mycotoxin binders on broiler breeders induced with aflatoxin B1: Effects on visceral organ weight and organ lesions parameters. Adv Environ Biol (2011) 5(13):3795–9.
71. Raju MVLN, Rao SV, Radhika K, Chawak MM. Dietary supplementation of spirulina and its effects on broiler chicken exposed to aflatoxicosis. Indian J Poult. Sci (2005) 40:36–40. doi: 10.3389/fvets.2021.640968
72. Raju MVLN, Rama Rao SV, Radhika K, Chawak MM. Effects of Spirulina platensis or furazolidone on the performance and immune response of broiler chickens fed with aflatoxin contaminated diet. Indian J Anim Nutr (2004) 21(1):40–4.
73. Mullenix GJ, Greene ES, Emami NK, Tellez-Isaias G, Bottje WG, Erf GF, et al. Spirulina platensis inclusion reverses circulating pro-inflammatory (Chemo)cytokine profiles in broilers fed low-protein diets. Front Vet Sci (2021) 8:640968. doi: 10.3389/fvets.2021.640968
74. Glenn H, Mullenix GJ, Erf GF. Effects of a low crude protein diet with and without Spirulina platensis inclusion on white blood cell profiles in broilers. Discovery Student J Dale Bumpers Coll Agricultural Food Life Sci (2021) 22(1):38–44. doi: 10.1007/s11356-021-14617-8
75. Tarkington V. Effect of dietary Spirulina platensis on stress levels and growth of female broiler chickens. animal science undergraduate honors theses (2020). Available at: https://scholarworks.uark.edu/anscuht/42.
76. Ibrahim SS, Elsabagh R, Allam A, Youssef G, Fadl SE, Abdelhiee EY, et al. Bioremediation role of Spirulina platensis against deltamethrin-mediated toxicity and its chemical residues in chicken meat. Environ Sci pollut Res Int (2021) 28(40):56188–98. doi: 10.1007/s11356-021-14617-8
77. Islam MS, Awal MA, Mostofa M, Begum F, Khair A, Myenuddin M. Effect of spirulina on toxic signs, body weight and hematological parameters in arsenic induced toxicities in ducks. Int J Poultry Sci (2009) 8(1):75–9. doi: 10.5713/ajas.17.0483
78. Zeweil H, Abaza IM, Zahran SM, Ahmed MH, Aboul-Ela HM, Asmaa AS. Effect of Spirulina platensis as dietary supplement on some biological traits for chickens under heat stress condition. Asian J Biomed Pharm Sci (2016) 6(56):08–12. doi: 10.1590/1806-9061-2018-0977
79. Mirzaie S, Zirak-Khattab F, Hosseini SA, Donyaei-Darian H. Effects of dietary spirulina on antioxidant status, lipid profile, immune response and performance characteristics of broiler chickens reared under high ambient temperature. Asian-Australas J Anim Sci (2018) 31, No. 4:556–63. doi: 10.5713/ajas.17.0483
80. Hajati H, Zaghari M, Oliveira HC. Arthrospira (Spirulina) platensis can be considered as a probiotic alternative to reduce heat stress in laying Japanese quails. Braz J Poultry Sci (2020) 22:001–8. doi: 10.1016/j.jtherbio.2021.103100
81. Moustafa ES, Alsanie WF, Gaber A, Kamel NN, Alaqil AA, Abbas AO, et al. Blue-green algae (Spirulina platensis) alleviates the negative impact of heat stress on broiler production performance and redox status. Animals (2021) 11:1243. doi: 10.3390/ani11051243
82. Kolluri G, Marappan G, Yadav AS, Kumar A, Kumar MA, Tyagi JS, et al. Effects of spirulina (Arthrospira platensis) as a drinking water supplement during cyclical chronic heat stress on broiler chickens: Assessing algal composition, production, stress, health and immune-biochemical indices. J Thermal Biol (2022) 103:103100. doi: 10.1016/j.jtherbio.2021.103100
83. Abdel-Moneim AE, Shehata AM, Mohamed NG, Elbaz AM, Ibrahim NS. Synergistic effect of Spirulina platensis and selenium nanoparticles on growth performance, serum metabolites, immune responses, and antioxidant capacity of heat-stressed broiler chickens. Biol Trace Elem Res (2021). doi: 10.1007/s12011-021-02662-w
84. Mirzaie S, Zirak-Khattab F, Hosseini SA, Donyaei-Darian H. ffects of dietary spirulina on antioxidant status, lipid profile, immune response and performance characteristics of broiler chickens reared under high ambient temperature. Asian Australas J Anim Sci (2018) 31(4):556–63. doi: 10.5713/ajas.17.0483
85. Altmann BA, Rosenau S. Spirulina as animal feed: Opportunities and challenges. Foods. (2022) 11(7):965. doi: 10.3390/foods11070965
86. Ragaza JA, Hossain MS, Meiler KA, Velasquez SF, Kumar V. A review on spirulina: Alternative media for cultivation and nutritive value as an aquafeed. Rev Aquac (2020) 12:2371–95. doi: 10.1007/s00449-016-1726-2
87. Smetana S, Sandmann M, Rohn S, Pleissner D, Heinz V. Autotrophic and heterotrophic microalgae and cyanobacteria cultivation for food and feed: Life cycle assessment. Bioresour. Technol (2017) 245:162–70. doi: 10.1023/A:1023856702544
88. Hultberg M, Lind O, Birgersson GG, Asp HH. Use of the effluent from biogas production for cultivation of spirulina. Bioprocess Biosyst Eng (2017) 40:625–31. doi: 10.1016/j.resconrec.2015.05.013
89. Olguín EJ, Galicia S, Mercado G, Pérez T. Annual productivity of spirulina (Arthrospira) and nutrient removal in a pig wastewater recycling process under tropical conditions. J Appl Phycol (2003) 15:249–57. doi: 10.1016/j.anifeedsci.2016.05.003
90. Taelman SE, De Meester S, Van Dijk W, da Silva V, Dewulf J. Environmental sustainability analysis of a protein-rich livestock feed ingredient in the Netherlands: Microalgae production versus soybean import. Resour Conserv Recycl (2015) 101:61–72. doi: 10.1016/j.biotechadv.2006.11.002
Keywords: Spirulina platensis, poultry, production, immunity, antimicrobials, antiviral, anticoccidial, nutrition
Citation: El-Shall NA, Jiang S, Farag MR, Azzam M, Al-Abdullatif AA, Alhotan R, Dhama K, Hassan F-u and Alagawany M (2023) Potential of Spirulina platensis as a feed supplement for poultry to enhance growth performance and immune modulation. Front. Immunol. 14:1072787. doi: 10.3389/fimmu.2023.1072787
Received: 17 October 2022; Accepted: 06 January 2023;
Published: 31 January 2023.
Edited by:
Guiyan Yang, University of California, Davis, United StatesCopyright © 2023 El-Shall, Jiang, Farag, Azzam, Al-Abdullatif, Alhotan, Dhama, Hassan and Alagawany. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Shouqun Jiang, amlhbmdzaG91cXVuQGdkYWFzLmNu; Mahmoud Alagawany, ZHIubWFobW91ZC5hbGFnd2FueUBnbWFpbC5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.