 Xin Chen
Xin Chen Lei Jiang
Lei Jiang
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Immunol., 27 July 2022
Sec. Cancer Immunity and Immunotherapy
Volume 13 - 2022 | https://doi.org/10.3389/fimmu.2022.954804
This article is part of the Research TopicNK cell modifications to advance their anti-tumor activitiesView all 13 articles
Immunotherapies focusing on rejuvenating T cell activities, like PD-1/PD-L1 and CTLA-4 blockade, have unprecedentedly revolutionized the landscape of cancer treatment. Yet a previously underexplored component of the immune system - natural killer (NK) cell, is coming to the forefront of immunotherapeutic attempts. In this review, we discuss the contributions of NK cells in the success of current immunotherapies, provide an overview of the current preclinical and clinical strategies at harnessing NK cells for cancer treatment, and highlight that NK cell-mediated therapies emerge as a major target in the next wave of cancer immunotherapy.
NK cells, a subset of lymphocytes that are principally innate immune cells, arise from common lymphoid progenitors and constitute the third lymphoid lineage in addition to T-cell and B-cell lineages (1). NK cells were initially discovered and named based on their ability to kill cancer cells in vitro (2). They express a broad repertoire of activating and inhibitory receptors, the “net weight” of which controls the final outputs. The biology of NK cells has been extensively reviewed elsewhere (3, 4). In this review, we mainly focus on the therapeutic potential of NK cells as the next wave in cancer immunity. We will discuss the prognostic roles of NK cells in cancers, summarize the contributions of NK cells in the success of immune checkpoint blockade (ICB) therapies and approaches including cell therapies to harness NK cells in the cancer treatment.
The immune surveillance role of NK cells in human cancers was first implicated in 1980s by reports revealing higher incidence of cancers in patients with NK cell defects (5, 6) and low NK cell activities in cancer patients or their families (7–11). Subsequently, a landmark 11-year following-up study reported a positive correlation between impaired NK cell functions and higher risk to develop numerous types of cancers (12). Meanwhile, the critical role of NK cells in control of tumor growth and metastasis was demonstrated in mice models in early studies (13, 14). However, due to the paucity of NK cells usually overserved in primary tumors in clinic, questions have been raised – as to whether NK cells play an important role in tumor control and prognosis, and whether NK cells contribute to therapies such as targeted antibody therapies, despite the role of NK cells in immune surveillance.
Subsequent to early findings, accumulating evidence have reported impaired functions of NK cells in chronic myelogenous leukemia (CML) (15) and acute myeloid leukemia (AML) (16, 17). Intriguingly, NK cells in AML patients have been reported to significantly down-regulate activating receptor NKp46 and up-regulate inhibitory receptor NKG2A compared to those in healthy age-matched controls (17). Furthermore, lower NKp46 expression on NK cells (18), phenotypic and functional defects of NK cells (17) or defective NK cell maturation (19) have been reported to be associated with adverse clinical outcomes in AML patients treated with allogeneic stem cell transplantation (allo-SCT) (18) or chemotherapy (17, 19).
Furthermore, the prognostic role of NK cells has not only been observed in chemotherapy-based studies in hematopoietic cancers, but also observed in targeted antibody therapy-based studies, in both liquid and solid tumors (Table 1). In diffuse large B-cell lymphoma (DLBCL) patients treated with Rituximab-CHOP (20), breast cancer patients treated with anti-HER2 monoclonal antibody (mAb) and chemotherapy (22), and in colorectal cancer patients treated with anti-EGFR mAb and chemotherapy (24), the tumor-infiltration of NK cells have been reported to positively correlate with clinical responses. Moreover, high baseline of antibody-dependent cellular cytotoxicity (ADCC) has been reported to correlate with a complete response (CR) and a long overall survival (OS) in head and neck cancer patients treated with anti-EGFR mAb and radiotherapy (23). Those evidence suggested a role of NK cells in targeted antibody therapy, probably mediated by ADCC, and support the development of tools harnessing ADCC activities of NK cells for enhanced anti-tumor efficacy. We will expand the discussion in later sessions of the review.

Table 1 Clinical correlations of NK cells with patient outcomes.
Another intriguing observation related to the prognostic and predictive role of NK cells comes from the studies on immune checkpoint blockades (ICBs) therapies. Higher NK cell infiltration has been found in responders to anti-PD-1 treatment compared to non-responders from independent studies (25, 26), and thus raise the question whether NK cells contribute to the success of ICBs.
Many inhibitory receptors including PD-1, LAG3, TIM3, TIGIT, NKG2A etc. are expressed and mediate inhibition on both NK cells and T cells (27) (Table 2). To date, anti-PD-1/PD-L1 therapies have achieved remarkable efficacy in a wide spectrum of cancers (28). Moreover, ICBs targeting LAG3 (29) and TIGIT (30) are displaying great potentials to further improve clinical outcomes in combination with anti-PD-1 therapy. Basically, the efficacy has been attributed to unleashing T cell responses, leaving the contributions of NK cells yet to be fully explored. Recently, growing evidence is suggesting a prognostic role of NK cell activation status and tumor infiltration in the success of ICB (25, 26, 31), thus raising considerable interests to fill the conceptual gap with respect to whether and how NK cells play a role in the ICB practice.
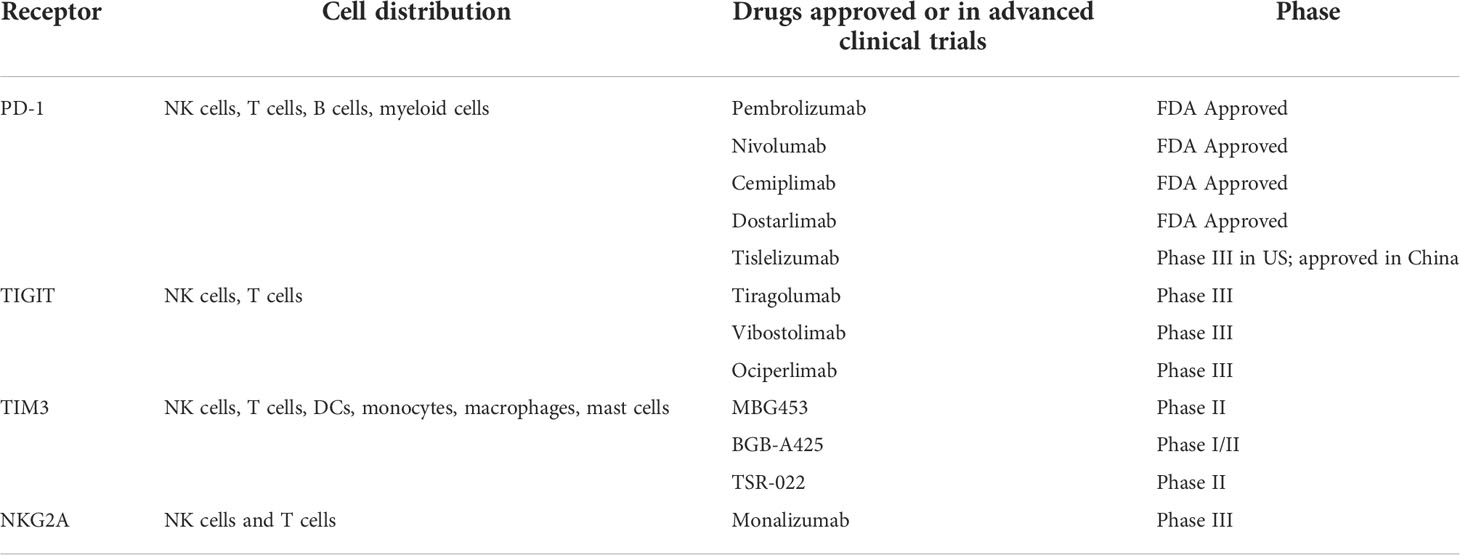
Table 2 Selected shared immune checkpoint receptors between NK cells and T cells.
First, NK cells may contribute to the ICB success by restraining the emergence of cancer cell clones that have escaped T cell attack through inactivation of antigen presentation. There is growing evidence that loss of genes associated with antigen presentation serves as an important mechanism of acquired resistance to ICB (32). In pre-clinical models, Nicolai et al. and Das et al. showed that NK cells mediate the rejection of CD8+ T cell resistant B2m-/- tumors (33, 34). It is in line with the longstanding observations that NK cells express inhibitory receptors binding to MHC-I, thereby maintaining “self-tolerance” to normal cells. When cancers down-regulate MHC-I on their surface to escape T cell attack, the “missing-self recognition” by NK cells is triggered, thus initiating NK cell mediated cytotoxicity against the “escapers” (35).
Second, the ICB may confer a direct modulation on NK cell activity. One study reported that PD-1 is upregulated on circulating and intra-tumoral NK cells in patients of Hodgkin lymphoma. PD-L1+ myeloid cells efficiently suppress the function of PD-1+ NK cells in vitro, while anti-PD-1 treatment can effectively reverse the suppression (36). Further evidence for the PD-1/PD-L1 signaling in NK cells comes from studies describing PD-1 upregulation in NK cells in non-small cell lung cancer (NSCLC) and head and neck cancer (HNC) patients. PD-L1 beads or PD-L1+ target cells impaired PD-1+ NK cell function, while anti-PD1 or anti-PD-L1 treatment significantly activated PD-1+ NK cells in vitro (37, 38). Moreover, there is evidence in in vivo mouse models that PD-1 is up-regulated on most activated tumor-infiltrating NK cells, and NK cells mediate full therapeutic efficacy of PD-1/PD-L1 blockade (39). Nevertheless, to what extent the anti-PD-1/PD-L1 therapies could directly activate NK cells in patients and thereby contribute to the efficacy remains an open question that needs to be further explored. Another shared checkpoint between T cells and NK cells, TIGIT, is constitutively expressed on PBMC-derived NK cells as well as in vitro activated human NK cells (40–43). In a recent publication, we have demonstrated the direct activation of NK cells by the therapeutic TIGIT blocking antibody ociperlimab (BGB-A1217) in an in vitro NK-cancer cell co-culture assay (44). Remarkably, the full Fc effector function of ociperlimab further elevated NK cell function in addition to checkpoint blockade (44), probably through the synergy between FcγRIIIa (CD16a) signaling and release of TIGIT mediated suppression on NK cells (45). Another immune checkpoint, TIM3, has been found to be up-regulated on NK cells from patients with melanoma (46), gastric cancer (47) and lung adenocarcinoma (48), and blockade of TIM3 has been reported to release the exhaustion of NK cells from advanced melanoma patients in vitro (46).
Recently, the NKG2A/CD94 blockade seems to carve a new path in the adoption of ICB in the cancer treatment via unleashing both T cells and NK cells. Pre-clinical data suggest a dual role of NKG2A blockade on NK cells and T cells (49–51). In clinic, monalizumab (49), a humanized IgG4 ICB targeting the NKG2A/CD94 receptor, blocking its interaction with HLA-E, is being investigated in the treatment of solid tumors. Encouraging results from a large, randomized Phase II trial showed monalizumab in combination with durvalumab, a PD-L1 blockade antibody, improved PFS and objective response rate (ORR) compared to durvalumab alone in patients with unresectable, stage III NSCLC. The 12-month PFS rate was 72.7% for durvalumab plus monalizumab, versus 33.9% with durvalumab alone (52).
From another perspective, it is noteworthy that the immune checkpoint blockade antibodies can activate NK cells through the Fc effector functions, as we reviewed previously (53). Direct evidence for this hypothesis comes from the Fc-competent TIGIT antibody Ociperlimab. TIGIT expression is highly expressed on Treg cells, relative to effector T cells, and is further elevated on Tregs in tumor microenvironment (44, 54). Our data have shown that the ligation of TIGIT on Tregs and Fcγ receptors on NK cells by Ociperlimab directly promoted NK cell activation and induced ADCC against cancer patient PBMC derived Tregs in vitro. In the CT26 mouse model, we also observed the decrease of intratumor Treg numbers (44). It is of great interest to further explore the potential mechanisms in clinical settings. Another T cell checkpoint, CTLA-4, is also expressed on cancer cells such as melanoma, leading to potential NK cell mediated ADCC against CTLA-4+ cancers induced by anti-CTLA-4 treatment (55). Nevertheless, CTLA-4 is also expressed on CTLs, thus rendering the overall mechanisms complicated.
Third, emerging evidence have suggested an essential role of NK cells in checkpoint therapy response through an NK-dendritic cell (DC) axis (25, 56). Conventional type 1 dendritic cells (cDC1) are a subtype of DC that stimulate robust T cell response to cancer. They adept at taking up dead cells and cross-present tumor antigen to CD8+ T cells (57, 58), attract T cells into tumor (59), and elicit tumor-specific T cell responses (60). Intriguingly, work from Bottcher et al. revealed a strong correlation between cDC1 signatures and NK cell signatures in cancers including skin cutaneous melanoma (SKCM), breast invasive carcinoma (BRCA), head and neck squamous cell carcinoma (HNSC) and lung adenocarcinoma (LUAD) (56). Moreover, NK cell signatures were found positively associated with patient survivals in all those cancers. Furthermore, they discovered in pre-clinical models that intratumor NK cells recruit cDC1 into tumors to promote tumor control (56). Similarly, Barry et al. observed that NK cell signatures positively correlate with stimulatory dendritic cells (SDC; intratumor cDC1) in melanoma, patient response to anti-PD-1 therapy and overall survival. In line with the data from Bottcher et al., they also uncovered a role of NK cells in the control of CD103+ SDC in a mouse tumor model (25).
Taken together, NK cells may contribute to ICBs success through multiple aspects. However, one caveat is that one should be cautious to interpret the data from pre-clinical models and translate from laboratory to clinic. Gaps exist between mouse models and human cancers, e.g., FcγRIII on mouse NK cells is actually not the homologue of FcγRIII on human NK cells. Human FcγRIIIa is functionally similar with a unique mouse FcγR – FcγRIV (61). Nonetheless mouse FcγRIV is not expressed on mouse NK cells, but abundantly on macrophages (53). In addition, syngeneic or xenograft tumor models may not truly mimic the NK cell infiltration status in human tumors, thereby suggesting translational gaps between pre-clinical tumor models and cancer patients.
NK cells express a broad range of activating and inhibitory receptors. Whether NK cells attack a target cell depends on the net equilibrium of the activating and inhibitory signals. Here we focus on emerging novel modalities for NK cell targeting, e.g., ADCC enhanced antibodies, bi- or tri-specifics, and iPSC-derived NK cells (iPSC-NK) therapies.
In humans, FcγRIIIa is the major type of FcγRs expressed on NK cells (62). Binding of Fc portion of human IgG to FcγRIIIa can trigger NK cell ADCC against mAb-opsonized target cells, as has been firmly established. Two alleles encode different FcγRIIIa variants that differ at the position 158, with either a valine (V) or phenylalanine (F). Between the two isoforms, FcγRIIIa-V158 exhibits higher affinity to IgG1, and mediated more efficient ADCC (63). In clinic, the FcγRIIIa dimorphism was strongly associated with the outcome of patients treated with anti-EGFR or anti-CD20 antibodies (64–69). Although it remains controversial about the relative contributions of different immune cells or effectors in the therapeutic efficacy of tumor-targeting mAbs (70–73), multiple studies have suggested a positive correlation of NK cell infiltration and activity with the response to tumor-targeting mAb treatment (22–24), and again, caution is warranted on the interpretation of mechanistic studies in mice given the discrepancies of FcγRs expression profiles between human and mice. Therefore, several strategies have been employed to develop ADCC-enhanced mAbs for harnessing NK cell functions.
Removal of core fucose from N-glycans attached to human IgG1 significantly enhances the binding affinity of IgG1 to FcγRIIIa and ADCC (74, 75), and has been the most widely adopted approach to harness the mAb mediated ADCC response in clinical practice (76). As of today, three afucosylated mAbs have been marketed for the treatment of human cancers: Obinutuzumab, a CD20-directed afucosylated antibody approved for the treatment of chronic lymphocytic leukemia (CLL); Poteligeo (mogamulizumab), a CCR4-targeting afucosylated mAb, approved for the treatment of Mycosis Fungoides (MF) and Sézary Syndrome (SS); and Fasenra (benralizumab), an afucosylated IL-5Rα targeting mAb for the treatment of patients with severe eosinophilic asthma. In addition, Rybrevant (amivantamab), an anti-EGFR and anti-cMet bispecific low fucose antibody with enhanced Fc function, have been approved for the treatment of NSCLC. Blenrep (belantamab mafodotin-blmf), consisting of an afucosylated humanized anti-BCMA IgG1 mAb conjugated to the tubulin inhibitor, monomethyl auristatin F (MMAF), for the treatment of adult patients with relapsed or refractory multiple myeloma, is the only FDA approved ADC with an afucosylated antibody (77). Nowadays, numerous afucosylated mAbs targeting a diverge range of receptors are actively in clinical development, with the outcomes yet to be revealed (78).
Fc engineering represents another approach to enhance ADCC (62, 79). Several Fc-enhanced mAbs through the genetic engineering approach are being investigated in clinical trials, with only one approved by FDA till now, Margenza (margetuximab), for the treatment of metastatic HER2-positive breast cancer. It is noteworthy that exploratory PFS analysis by FcγRIIIa genotype suggested that presence of a FcγRIIIa-F158 allele may predict margetuximab benefit over trastuzumab. Margetuximab provided no clinical benefit in FcγRIIIa-V158 homozygotes compared with trastuzumab (80). Since the Fc engineering of margetuximab-cmkb increases affinity for both FcγRIIIa allotypes, and FcγRIIIa-V158 per se has higher affinity to IgG1, the none-benefit in FcγRIIIa-V158 homozygotes might be attributed to the rapid cleavage and downregulation of FcγRIIIa due to stronger binding of the antibodies to FcγRIIIa-V158. From another perspective, strong binding to FcγRIIIa may induce enhanced antibody internalization by FcγRIIIa expressing cells, thus promoting the anti-drug antibody (ADA) production, to compromise the efficacy. The exact underlying mechanisms are yet to be elucidated.
There are a broad range of activating and inhibitory receptors on NK cells. The integration of signals for activation and inhibition determines the final outputs of NK cells. The loss of inhibitory signaling, like downregulation of MHC-I expression on tumor cells, renders tumor cells susceptible to NK cell cytotoxicity. Alternatively, NK cells can attack cancer cells that retain full expression of MHC-I if activating receptors on NK cells are engaged.
Recently, bi-specific or tri-specific antibodies targeting NK cell activating receptors are emerging as novel approaches to harness NK activity. Preclinical results provide the rationale for developing multi-specific NK cell engagers through ligation of tumor antigens and activating NK receptors. Examples include those targeting NKp46 (81), NKp30 (82), NKG2D (83, 84), and FcγRIIIa (CD16a) (85, 86). Encouraging data comes from a Phase I clinical study in which an anti-CD16/anti-CD30 bispecific NK-cell engager combined with pembrolizumab has shown an ORR of 83% and a CR rate of 46% in patients with relapsed or refractory Hodgkin Lymphoma (HL) (87). However, it should be noted that most of the activating receptors are not exclusively expressed on NK cells, instead are often shared with T cells or myeloid compartments. Targeting the activating receptors on NK cells may synergically augment both NK cells and other immune effectors.
A wide range of immune checkpoints are expressed on NK cells. As with the activating receptors, NK checkpoints are usually shared with other immune components (27). As discussed in the earlier part of the review, some blockades that can target checkpoints on both T cell and NK cells have obtained remarkable success or promising preliminary clinical responses (Table 2), albeit the contributions of NK cells therein are yet to be fully understood. In addition, blockers that target inhibitory killer Ig-like receptors (KIRs) have been investigated in clinical settings. KIRs are a group of receptors on NK cells that bind to HLA molecules to mediate inhibitory or activating signaling (88). Clinical evidence have suggested that adoptive NK cell transfer has the potential to improve outcomes of KIR ligand-mismatched recipients even further (89–91). Lirilumab, a humanized IgG4 mAb, binds to KIR2DL-1, KIR2DL-2 and KIR2DL-3 and thereby blocks their inhibitory signaling mediated by both HLA-C C1 and HLA-C C2 subtype molecules (92). It has shown good safety profiles in phase I trials, however, the phase II trial in patients with smoldering multiple myeloma failed to demonstrate clinical efficacy (93). The minimal efficacy may result from lack of the KIR matched HLA types from patients, and existing of other dominant inhibition signals (94). However, this does not rule out the possibility that inhibitory KIR blockers could synergistically work together with other ICBs or NK cell therapies to induce a combination efficacy.
Adoptive cell therapies, basically chimeric antigen receptor T (CAR-T) cell therapies, have exhibited remarkable clinical responses in treating hematologic malignancies, and thus spawned an explosion in the CAR-T field. As of today, six CAR-T cell therapies have been approved by FDA, wherein four targeting CD19, and two targeting BCMA. Although the efficacies have been notable (95–101), limitations are still obvious. First, as a highly personalized therapy, autologous CAR-T cells have to be individually prepared for each patient in a time- and material- consuming process that carries the risk for failure and demanding logistics. Patients who have already received multiple rounds of chemotherapy may not be able to mobilize enough T cells for the CAR-T cell preparation. Additionally, during the time waiting for CAR-T cells manufacturing, patients may experience disease progression. As such, the therapy results in low scalability and remains unacceptable and unaffordable to most of patients. Second, severe toxicity associated with CAR-T cells hampers the broad applicability of the treatment. In several patients, CAR-T cell treatments have been associated with substantial toxicity including cytokine release syndrome (CRS) and immune effector cell-associated neurotoxicity syndrome (ICANS), thus decreasing the feasibility due to demanding toxicity management, and inconvenient administration (102).
In contrast, NK cell therapies harbor high potential to overcome those hurdles. Firstly, the activating machineries of NK cells differ from the TCR system of T cells. NK cells do not require HLA matching to exert cytotoxicity against tumor cells. This allows for using NK cells in an allogeneic way. On the other hand, allogeneic NK cells do not result in graft versus host disease (GvHD), even in the setting of substantial HLA disparity between adoptive CAR-NK cells and the recipients (103), and thus can be provided as complete “off-the-shelf” products, significantly lowering the cost of manufacturing and logistics. Secondly, both autologous and allogeneic NK cells have exhibited excellent safety profile, without severe toxicity such as CRS or ICANS (103, 104). Compared to their T cell counterparts, NK cells present a safer cytokine profile, and differ in the crosstalk with myeloid cells (4). This property confers the feasibility of the NK cell therapy when specialized care units are unavailable. Thirdly, from the efficacy perspective, NK or CAR-NK cell therapies have shown inspiring clinical outcomes in early phase clinical trials when used alone (103) or in combination with other therapies (104), encouraging more endeavors in the field. Last but not least, repeated doses can be administrated given the short lifetime of NK cells, and NK cells from different donors can be sequentially dosed to circumvent rejection of donor NK cells by recipient memory T cells recognizing allo-antigens on the same donors.
NK cells for cell therapy can be generated from different sources and by a variety of methods (Table 3). Peripheral blood derived, and ex vivo expanded autologous NK cells have been well tolerated in clinical trials, whereas efficacy has been limited (105, 106). The low efficacy may be attributed to the suppression of autologous NK cells by self-HLA molecules. As such, allogeneic NK cells serve as a promising alternative approach to overcome the resistance. In a seminal study by Miller et al., a complete remission induced by haploidentical allogeneic NK-cell infusions in 5 of 19 poor-prognosis AML patients was observed (90). In a subsequent study in pediatric AML, all patients who received adoptive haploidentical NK cells remained in remission with a median follow-up time of 964 days (91). Later on, 53% complete remission was observed in AML patients treated with haploidentical NK cells combined with an IL-2 diphtheria toxin fusion protein, which was used to deplete host regulatory T cells (107); 32% complete remission was observed in AML patients treated with haploidentical NK cells combined with IL-15 (108), and 44% complete remission was observed in AML patients treated with allogeneic cytokine-induced memory-like NK cells, in separate studies (109).
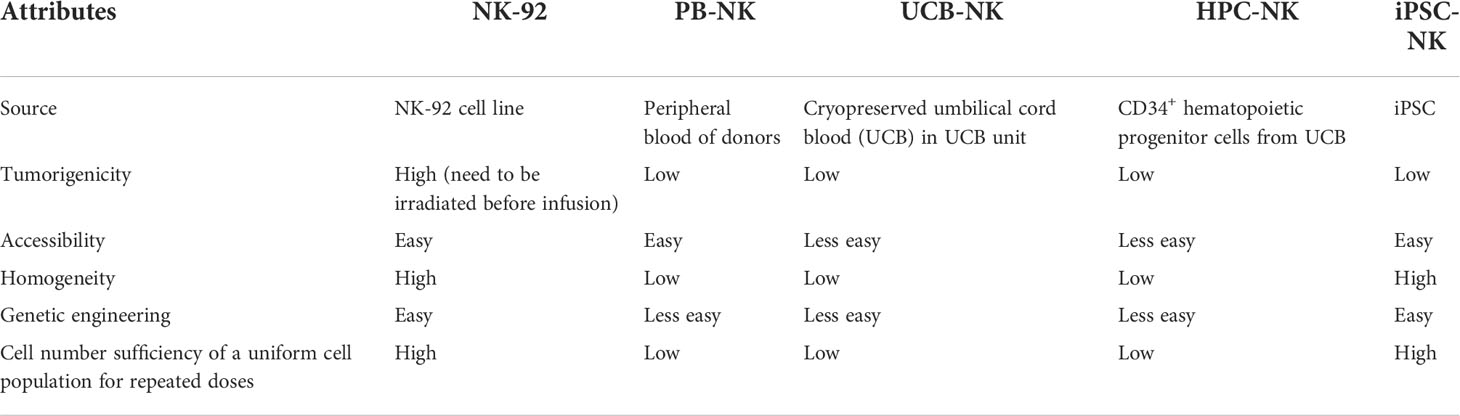
Table 3 Comparison of clinical-scale NK cells generated from distinct sources.
Besides peripheral blood-derived NK cells (PB-NK), an alternative approach to generate functional NK cells is to obtain NK cells from umbilical cord blood and expand ex vivo (110, 111). A recent clinical study by Liu et al. has presented inspiring results to show the remarkable efficacy and excellent safety profile of engineered umbilical cord blood derived NK cells (UCB-NK) in the treatment of CD19 positive relapsed or refractory lymphoid tumors. The HLA-mismatched UCB-NK cells were transduced with a retroviral vector encoding anti-CD19 CAR, IL-15, and inducible caspase 9 as a safety switch. Of the 11 patients who were treated, 8 (73%) had a response and 7 (64%) had a complete remission. Notably, no severe toxicity including CRS, neurotoxicity or GvHD were observed (103). Another method to derive NK cells from umbilical cord blood is to differentiate them from CD34+ hematopoietic progenitor cells (HPC) (112). In the first-in-human study, CD34+ HPC derived NK cells (HPC-NK) were administrated to 10 older AML patients after lymphodepleting chemotherapy without cytokine boosting. Preliminary data showed that HPC-NK cells were well tolerated, with neither GvHD nor toxicity observed. Notably, 2 of 4 patients with minimal residual disease (MRD) in bone marrow before HPC-NK cells infusion became MRD negative, which lasted for 6 months (113).
Albeit the encouraging efficacy achieved by those clinical trials using PB-NK or UCB-NK, limitations exist due to the requirement for collection from a donor by apheresis or from umbilical cord blood, the variability of NK cell yield influenced by donor variability, and the challenge in generic manipulation on differentiated cells with low proliferation capacity. To overcome those limitations, a number of studies used NK-92, a NK cell line originally established from a patient with non-Hodgkin’s lymphoma (114–116). There are several advantages of NK-92 cell line as a source for NK cell therapy – it provides a homogeneous master cell bank, can be expanded indefinitely and served as an uniform “off-the-shelf” product, is more amenable to genetic modification and allows sufficient cells for cell therapy (117). However, on the other hand, albeit NK-92 lack expression of most known KIRs and exhibit broad cytotoxicity against numerous cancers, it loses expression of typical activating receptors including NKp44, NKp46 and notably, FcγRIIIa, which mediated ADCC (117). Additionally, as a lymphoma cell line, NK-92 holds inherent draw backs such as potential tumorigenicity and latent infection by Epstein-Barr Virus (EBV). Thus, for safety considerations, NK-92 must be irradiated before administration to patients. The irradiation limits the proliferation and persistence of NK-92 in vivo, and eventually may impede the long-term anti-tumor efficacy. And this may account for the observed limited efficacy of NK-92 cells in clinic (118, 119).
In recent years, iPSC-NK technology emerges as a breakthrough innovation in the NK cell therapy field, offering the potential to overcome challenges often seen with other source-derived NK cells. Serial seminal studies from Kaufman et al. have significantly optimized the protocols to derive NK cells to a clinical-scale from embryonic stem cells (hESCs) or iPSC (120), and demonstrated for the first time that CAR-NK cells can be derived from iPSCs expressing CAR (121). Since pluripotent stem cells have the potential to grow indefinitely in an undifferenced state (122, 123), the iPSC can serve as a stable cell bank for uniform NK cell generation and allows for sufficient cell numbers for cell therapy. As such, the iPSC-NK can serve as a standardized “off-the-shelf” product. In addition, iPSC is amenable to genetic engineering. Once the genetically modified clones are selected, it can be expanded for a production of a uniform pool of iPSC-NK cells. Multiple genetical modifications, such as ectopic expression of IL-15/IL-15R fusion protein (124, 125), CAR (121), high-affinity non-cleavable variant of CD16a (125, 126), deletion of CISH (127) or CD38 (125) have been successfully introduced on iPSC-NK to achieve enhanced expansion, better in vivo persistence or greater cytotoxicity. The difference between iPSC-NK, PB-NK and UCB-NK are yet to be fully understood, yet some pre-clinical evidence have suggested that iPSC-NK may have comparable or superior activities relative to PB-NK or UCB-NK (128–130). To date, iPSC-NK cell therapies have entered phase I clinical trials, used alone or in combination with therapeutic monoclonal antibodies (mAbs) for the treatment of hematopoietic lymphomas or solid tumors (131–136). Remarkably, the first in human results are encouraging (137, 138). In a phase I trial, FT516, an iPSC-NK cell therapy using iPSC-NK cells engineered with a high-affinity, non-cleavable CD16a (hnCD16) that enables tumor targeting and enhanced ADCC in combination with a therapeutic mAb, was combined with rituximab to treat patients with relapsed or refractory B-cell lymphoma (BCL) (132). Eight of the 11 pts (73%) treated with ≥90 million FT516 cells achieved an objective response. Seven (64%) patients achieved CR, including 2 patients with prior CD19 CAR T-cell therapy (139). FT596 (140), is an iPSC-derived CAR-NK cell therapy armed with three modalities: a CD19-targeting CAR, a hnCD16, and IL15/IL-15 receptor fusion which promotes NK cell persistence by the autonomous cytokine. In a phase I trial, FT596 was administrated as monotherapy or in combination with rituximab or obinutuzumab for the treatment of relapsed or refractory BCLs and CLL (141). At single-dose levels of ≥90 million cells, 8 of 11 (73%) efficacy-evaluable patients achieved ORR, including 7 (64%) CR. Of 4 patients with prior CAR T-cell therapy treated at ≥90 million cells, 2 achieved CR (142). Notably, no dose-limiting toxicities, CRS, ICANS, or GvHD of any grade were observed for FT516 or FT596, and repeated doses were allowed (139, 142).
Nowadays, NK cell-based combination strategies have been investigated in the cancer immunotherapy and represent an important direction in the future.
As we have discussed in the previous sessions, activated or intra-tumor NK cells up-regulate checkpoint molecules (e.g., PD-1, TIGIT, TIM3, NKG2A) and blocked of those molecules unleash NK cell activity. In a pre-clinical model, iPSC-NK cells in combination with T cells and an anti-PD-1 antibody have been reported to eliminate tumors in a xenograft ovarian cancer mouse model (143). Combination of ICBs and adoptive NK cell therapy would be a promising approach to achieve optimized NK functions, and in concert with T cells.
Adoptive NK cell therapy in combination with tumor-targeting mAbs or other NK cell engagers represent an approach to fully augment the tumor-specific NK cell cytotoxicity. Impressive results have been obtained from a phase I clinical study that combine CAR-iPSC-NK cell therapy with anti-CD20 (139). Moreover, the combination of CB-NK and a bispecific CD30/CD16 antibody is being actively investigated in a phase I/II study (144, 145). As disclosed by Affimed on the 2022 AACR meeting, as of the cut-off date, the study had enrolled 22 patients with relapsed or refractory CD30+ Hodgkin and non-Hodgkin lymphoma having received a median of seven prior lines of therapy. Out of the 13 patients treated at the recommended phase 2 dose (RP2D), 13 patients (100%) achieved objective response, and 8 (62%) patients achieved CR after two cycles of treatment (146).
Sequential infusions of CAR-NK cells and CAR-T cells would be a good strategy to achieve better efficacy and safety. CAR-NK cells should rapidly decrease the tumor burden, particularly for patients with high tumor load. This may decrease the CRS and neurotoxicity risk imposed by CAR-T cells. Then subsequent CAR T cell infusion may eliminate residual tumor cells and provide a lasting anti-tumor effect through memory T cells that survive and persistence.
In addition to the approaches discussed above, strategies targeting cytokines, such as IL-12, IL-15 and IL18 (109, 124, 147), targeting intracellular checkpoints, such as CISH (127, 148), Cbl-b (149) GSK3 (150) and CDK8 (151, 152), or targeting tumor cells which can indirectly trigger NK cell surveillance through non–cell autonomous mechanisms (153) may also effectively augment NK cell functions and eventually result in novel therapeutic candidates.
Taken together, NK cell-based therapies have attracted intense interest and shown great potential in the treatment of cancers, emerging as the next wave in cancer immunotherapy. Multiple approaches, including checkpoint blockades, ADCC enhanced antibodies, agonist antibodies and multi-specific NK cell engagers, and adoptive NK cell therapies (particularly engineered iPSC NK cell therapies) have significantly widened the pool of potential clinical options. However, challenges exist with the opportunities. As a heterogenous population, NK cells are still not fully understood. It is crucial to continue to delineate the NK cell biology and characterize the differences of NK cells derived from distinct sources and methods. In addition, although NK cell-based therapies have demonstrated great potentials in the treatment of hematopoietic cancers, the advances in solid tumors remain limited. It is important to further understand NK homing capacities and the reasons underlying their poor infiltrations in solid tumors, which may eventually lead to the development of novel approaches to overcome the barriers. Furthermore, the questions about the persistence of NK cells, and the durability of the response, and the affordable cost for patients need to be considered. Along with the advancing of new technologies and methods, NK cell-based therapies will continue to evolve, and get closer to benefit patients with otherwise no treatment options. In summary, NK cell-mediated therapies have emerged as the next wave in cancer immunotherapy.
XC and LJ drafted the manuscript. XL reviewed the manuscript, and all authors were involved in revision of the manuscript. All authors contributed to the article and approved the submitted version.
This work was supported by internal company funding.
XC is a Senior Principal Investigator at the company BeiGene (Beijing) Co., Ltd. LJ is a Senior Scientist at the company BeiGene (Beijing) Co., Ltd. XL is the Vice President, Head of Biology at the company BeiGene (Beijing) Co., Ltd.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Chiossone L, Dumas PY, Vienne M, Vivier E. Natural killer cells and other innate lymphoid cells in cancer. Nat Rev Immunol (2018) 18:671–88. doi: 10.1038/s41577-018-0061-z
2. Trinchieri G. Biology of natural killer cells. Adv Immunol (1989) 47:187–376. doi: 10.1016/s0065-2776(08)60664-1
3. Huntington ND, Vosshenrich CA, Di Santo JP. Developmental pathways that generate natural-killer-cell diversity in mice and humans. Nat Rev Immunol (2007) 7:703–14. doi: 10.1038/nri2154
4. Long EO, Kim HS, Liu D, Peterson ME, Rajagopalan S. Controlling natural killer cell responses: integration of signals for activation and inhibition. Annu Rev Immunol (2013) 31:227–58. doi: 10.1146/annurev-immunol-020711-075005
5. Roder JC, Haliotis T, Klein M, Korec S, Jett JR, Ortaldo J, et al. A new immunodeficiency disorder in humans involving NK cells. Nature (1980) 284:553–5. doi: 10.1038/284553a0
6. Sullivan JL, Byron KS, Brewster FE, Purtilo DT. Deficient natural killer cell activity in x-linked lymphoproliferative syndrome. Science (1980) 210:543–5. doi: 10.1126/science.6158759
7. Hersey P, Edwards A, Honeyman M, McCarthy WH. Low natural-killer-cell activity in familial melanoma patients and their relatives. Br J Cancer (1979) 40:113–22. doi: 10.1038/bjc.1979.147
8. Nakajima T, Mizushima N, Nakamura J, Kanai K. Surface markers of NK cells in peripheral blood of patients with cirrhosis and hepatocellular carcinoma. Immunol Lett (1986) 13:7–10. doi: 10.1016/0165-2478(86)90117-3
9. Schantz SP, Shillitoe EJ, Brown B, Campbell B. Natural killer cell activity and head and neck cancer: a clinical assessment. J Natl Cancer Inst (1986) 77:869–75. doi: 10.1093/jnci/77.4.869
10. Strayer DR, Carter WA, Brodsky I. Familial occurrence of breast cancer is associated with reduced natural killer cytotoxicity. Breast Cancer Res Treat (1986) 7:187–92. doi: 10.1007/BF01806249
12. Imai K, Matsuyama S, Miyake S, Suga K, Nakachi K. Natural cytotoxic activity of peripheral-blood lymphocytes and cancer incidence: an 11-year follow-up study of a general population. Lancet (2000) 356:1795–9. doi: 10.1016/S0140-6736(00)03231-1
13. Talmadge JE, Meyers KM, Prieur DJ, Starkey JR. Role of natural killer cells in tumor growth and metastasis: C57BL/6 normal and beige mice. J Natl Cancer Inst (1980) 65:929–35. doi: 10.1093/jnci/65.5.929
14. Gorelik E, Wiltrout RH, Okumura K, Habu S, Herberman RB. Role of NK cells in the control of metastatic spread and growth of tumor cells in mice. Int J Cancer (1982) 30:107–12. doi: 10.1002/ijc.2910300118
15. Pierson BA, Miller JS. CD56+bright and CD56+dim natural killer cells in patients with chronic myelogenous leukemia progressively decrease in number, respond less to stimuli that recruit clonogenic natural killer cells, and exhibit decreased proliferation on a per cell basis. Blood (1996) 88:2279–87. doi: 10.1182/blood.V88.6.2279.bloodjournal8862279
16. Costello RT, Sivori S, Marcenaro E, Lafage-Pochitaloff M, Mozziconacci MJ, Reviron D, et al. Defective expression and function of natural killer cell-triggering receptors in patients with acute myeloid leukemia. Blood (2002) 99:3661–7. doi: 10.1182/blood.v99.10.3661
17. Stringaris K, Sekine T, Khoder A, Alsuliman A, Razzaghi B, Sargeant R, et al. Leukemia-induced phenotypic and functional defects in natural killer cells predict failure to achieve remission in acute myeloid leukemia. Haematologica (2014) 99:836–47. doi: 10.3324/haematol.2013.087536
18. Chretien AS, Devillier R, Fauriat C, Orlanducci F, Harbi S, Le Roy A, et al. NKp46 expression on NK cells as a prognostic and predictive biomarker for response to allo-SCT in patients with AML. Oncoimmunology (2017) 6:e1307491. doi: 10.1080/2162402X.2017.1307491
19. Chretien AS, Fauriat C, Orlanducci F, Galseran C, Rey J, Bouvier Borg G, et al. Natural killer defective maturation is associated with adverse clinical outcome in patients with acute myeloid leukemia. Front Immunol (2017) 8:573. doi: 10.3389/fimmu.2017.00573
20. Xu-Monette ZY, Xiao M, Au Q, Padmanabhan R, Xu B, Hoe N, et al. Immune profiling and quantitative analysis decipher the clinical role of immune-checkpoint expression in the tumor immune microenvironment of DLBCL. Cancer Immunol Res (2019) 7:644–57. doi: 10.1158/2326-6066.CIR-18-0439
21. Rea D, Henry G, Khaznadar Z, Etienne G, Guilhot F, Nicolini F, et al. Natural killer-cell counts are associated with molecular relapse-free survival after imatinib discontinuation in chronic myeloid leukemia: the IMMUNOSTIM study. Haematologica (2017) 102:1368–77. doi: 10.3324/haematol.2017.165001
22. Muntasell A, Rojo F, Servitja S, Rubio-Perez C, Cabo M, Tamborero D, et al. NK cell infiltrates and HLA class i expression in primary HER2(+) breast cancer predict and uncouple pathological response and disease-free survival. Clin Cancer Res (2019) 25:1535–45. doi: 10.1158/1078-0432.CCR-18-2365
23. Lattanzio L, Denaro N, Vivenza D, Varamo C, Strola G, Fortunato M, et al. Elevated basal antibody-dependent cell-mediated cytotoxicity (ADCC) and high epidermal growth factor receptor (EGFR) expression predict favourable outcome in patients with locally advanced head and neck cancer treated with cetuximab and radiotherapy. Cancer Immunol Immunother (2017) 66:573–9. doi: 10.1007/s00262-017-1960-8
24. Marechal R, De Schutter J, Nagy N, Demetter P, Lemmers A, Deviere J, et al. Putative contribution of CD56 positive cells in cetuximab treatment efficacy in first-line metastatic colorectal cancer patients. BMC Cancer (2010) 10:340. doi: 10.1186/1471-2407-10-340
25. Barry KC, Hsu J, Broz ML, Cueto FJ, Binnewies M, Combes AJ, et al. A natural killer-dendritic cell axis defines checkpoint therapy-responsive tumor microenvironments. Nat Med (2018) 24:1178–91. doi: 10.1038/s41591-018-0085-8
26. Lee H, Quek C, Silva I, Tasker A, Batten M, Rizos H, et al. Integrated molecular and immunophenotypic analysis of NK cells in anti-PD-1 treated metastatic melanoma patients. Oncoimmunology (2019) 8:e1537581. doi: 10.1080/2162402X.2018.1537581
27. Sivori S, Della Chiesa M, Carlomagno S, Quatrini L, Munari E, Vacca P, et al. Inhibitory receptors and checkpoints in human nk cells, implications for the immunotherapy of cancer. Front Immunol (2020) 11:2156. doi: 10.3389/fimmu.2020.02156
28. Chen S, Zhang Z, Zheng X, Tao H, Zhang S, Ma J, et al. Response efficacy of PD-1 and PD-L1 inhibitors in clinical trials: A systematic review and meta-analysis. Front Oncol (2021) 11:562315. doi: 10.3389/fonc.2021.562315
29. Tawbi HA, Schadendorf D, Lipson EJ, Ascierto PA, Matamala L, Castillo Gutierrez E, et al. Relatlimab and nivolumab versus nivolumab in untreated advanced melanoma. N Engl J Med (2022) 386:24–34. doi: 10.1056/NEJMoa2109970
30. Rodriguez-Abreu D, Johnson ML, Hussein MA, Cobo M, Patel AJ, Secen NM, et al. Primary analysis of a randomized, double-blind, phase II study of the anti-TIGIT antibody tiragolumab (tira) plus atezolizumab (atezo) versus placebo plus atezo as first-line (1L) treatment in patients with PD-L1-selected NSCLC (CITYSCAPE). J Clin Oncol (2020) 38:9503–3. doi: 10.1200/JCO.2020.38.15_suppl.9503
31. Zemek RM, De Jong E, Chin WL, Schuster IS, Fear VS, Casey TH, et al. Sensitization to immune checkpoint blockade through activation of a STAT1/NK axis in the tumor microenvironment. Sci Transl Med (2019) 11:eaav7816. doi: 10.1126/scitranslmed.aav7816
32. Schoenfeld AJ, Hellmann MD. Acquired resistance to immune checkpoint inhibitors. Cancer Cell (2020) 37:443–55. doi: 10.1016/j.ccell.2020.03.017
33. Nicolai CJ, Wolf N, Chang IC, Kirn G, Marcus A, Ndubaku CO, et al. NK cells mediate clearance of CD8(+) T cell-resistant tumors in response to STING agonists. Sci Immunol (2020) 5:eaaz2738. doi: 10.1126/sciimmunol.aaz2738
34. Das K, Eisel D, Lenkl C, Goyal A, Diederichs S, Dickes E, et al. Generation of murine tumor cell lines deficient in MHC molecule surface expression using the CRISPR/Cas9 system. PloS One (2017) 12:e0174077. doi: 10.1371/journal.pone.0174077
35. Ljunggren HG, Karre K. In search of the 'missing self': MHC molecules and NK cell recognition. Immunol Today (1990) 11:237–44. doi: 10.1016/0167-5699(90)90097-s
36. Vari F, Arpon D, Keane C, Hertzberg MS, Talaulikar D, Jain S, et al. Immune evasion via PD-1/PD-L1 on NK cells and monocyte/macrophages is more prominent in Hodgkin lymphoma than DLBCL. Blood (2018) 131:1809–19. doi: 10.1182/blood-2017-07-796342
37. Trefny MP, Kaiser M, Stanczak MA, Herzig P, Savic S, Wiese M, et al. PD-1(+) natural killer cells in human non-small cell lung cancer can be activated by PD-1/PD-L1 blockade. Cancer Immunol Immunother (2020) 69:1505–17. doi: 10.1007/s00262-020-02558-z
38. Concha-Benavente F, Kansy B, Moskovitz J, Moy J, Chandran U, Ferris RL. PD-L1 mediates dysfunction in activated PD-1(+) NK cells in head and neck cancer patients. Cancer Immunol Res (2018) 6:1548–60. doi: 10.1158/2326-6066.CIR-18-0062
39. Hsu J, Hodgins JJ, Marathe M, Nicolai CJ, Bourgeois-Daigneault MC, Trevino TN, et al. Contribution of NK cells to immunotherapy mediated by PD-1/PD-L1 blockade. J Clin Invest (2018) 128:4654–68. doi: 10.1172/JCI99317
40. Zhang Q, Bi J, Zheng X, Chen Y, Wang H, Wu W, et al. Blockade of the checkpoint receptor TIGIT prevents NK cell exhaustion and elicits potent anti-tumor immunity. Nat Immunol (2018) 19:723–32. doi: 10.1038/s41590-018-0132-0
41. Stanietsky N, Simic H, Arapovic J, Toporik A, Levy O, Novik A, et al. The interaction of TIGIT with PVR and PVRL2 inhibits human NK cell cytotoxicity. Proc Natl Acad Sci U S A (2009) 106:17858–63. doi: 10.1073/pnas.0903474106
42. Yu X, Harden K, Gonzalez LC, Francesco M, Chiang E, Irving B, et al. The surface protein TIGIT suppresses T cell activation by promoting the generation of mature immunoregulatory dendritic cells. Nat Immunol (2009) 10:48–57. doi: 10.1038/ni.1674
43. Wang F, Hou H, Wu S, Tang Q, Liu W, Huang M, et al. TIGIT expression levels on human NK cells correlate with functional heterogeneity among healthy individuals. Eur J Immunol (2015) 45:2886–97. doi: 10.1002/eji.201545480
44. Chen X, Xue L, Ding X, Zhang J, Jiang L, Liu S, et al. An fc-competent anti-human TIGIT blocking antibody ociperlimab (BGB-A1217) elicits strong immune responses and potent anti-tumor efficacy in pre-clinical models. Front Immunol (2022) 13:828319. doi: 10.3389/fimmu.2022.828319
45. Bryceson YT, March ME, Ljunggren HG, Long EO. Synergy among receptors on resting NK cells for the activation of natural cytotoxicity and cytokine secretion. Blood (2006) 107:159–66. doi: 10.1182/blood-2005-04-1351
46. da Silva IP, Gallois A, Jimenez-Baranda S, Khan S, Anderson AC, Kuchroo VK, et al. Reversal of NK-cell exhaustion in advanced melanoma by Tim-3 blockade. Cancer Immunol Res (2014) 2:410–22. doi: 10.1158/2326-6066.CIR-13-0171
47. Wang Z, Zhu J, Gu H, Yuan Y, Zhang B, Zhu D, et al. The clinical significance of abnormal tim-3 expression on NK cells from patients with gastric cancer. Immunol Invest (2015) 44:578–89. doi: 10.3109/08820139.2015.1052145
48. Xu L, Huang Y, Tan L, Yu W, Chen D, Lu C, et al. Increased Tim-3 expression in peripheral NK cells predicts a poorer prognosis and Tim-3 blockade improves NK cell-mediated cytotoxicity in human lung adenocarcinoma. Int Immunopharmacol (2015) 29:635–41. doi: 10.1016/j.intimp.2015.09.017
49. Andre P, Denis C, Soulas C, Bourbon-Caillet C, Lopez J, Arnoux T, et al. Anti-NKG2A mAb is a checkpoint inhibitor that promotes anti-tumor immunity by unleashing both T and NK cells. Cell (2018) 175:1731–43.e13. doi: 10.1016/j.cell.2018.10.014
50. Kamiya T, Seow SV, Wong D, Robinson M, Campana D. Blocking expression of inhibitory receptor NKG2A overcomes tumor resistance to NK cells. J Clin Invest (2019) 129:2094–106. doi: 10.1172/JCI123955
51. van Montfoort N, Borst L, Korrer MJ, Sluijter M, Marijt KA, Santegoets SJ, et al. NKG2A blockade potentiates CD8 T cell immunity induced by cancer vaccines. Cell (2018) 175:1744–55.e15. doi: 10.1016/j.cell.2018.10.028
52. Herbst RS, Majem M, Barlesi F, Carcereny E, Chu Q, Monnet I, et al. COAST: an open-label, phase II, multidrug platform study of durvalumab alone or in combination with oleclumab or monalizumab in patients with unresectable, stage III non-small-cell lung cancer J Clin Oncol (2022) JCO2200227. doi: 10.1200/JCO.22.00227
53. Chen X, Song X, Li K, Zhang T. FcgammaR-binding is an important functional attribute for immune checkpoint antibodies in cancer immunotherapy. Front Immunol (2019) 10:292. doi: 10.3389/fimmu.2019.00292
54. Preillon J, Cuende J, Rabolli V, Garnero L, Mercier M, Wald N, et al. Restoration of T-cell effector function, depletion of tregs, and direct killing of tumor cells: the multiple mechanisms of action of a-TIGIT antagonist antibodies. Mol Cancer Ther (2021) 20:121–31. doi: 10.1158/1535-7163.MCT-20-0464
55. Laurent S, Queirolo P, Boero S, Salvi S, Piccioli P, Boccardo S, et al. The engagement of CTLA-4 on primary melanoma cell lines induces antibody-dependent cellular cytotoxicity and TNF-alpha production. J Transl Med (2013) 11:108. doi: 10.1186/1479-5876-11-108
56. Bottcher JP, Bonavita E, Chakravarty P, Blees H, Cabeza-Cabrerizo M, Sammicheli S, et al. NK cells stimulate recruitment of cdc1 into the tumor microenvironment promoting cancer immune control. Cell (2018) 172:1022–37.e14. doi: 10.1016/j.cell.2018.01.004
57. Roberts EW, Broz ML, Binnewies M, Headley MB, Nelson AE, Wolf DM, et al. Critical role for CD103(+)/CD141(+) dendritic cells bearing CCR7 for tumor antigen trafficking and priming of T cell immunity in melanoma. Cancer Cell (2016) 30:324–36. doi: 10.1016/j.ccell.2016.06.003
58. Salmon H, Idoyaga J, Rahman A, Leboeuf M, Remark R, Jordan S, et al. Expansion and activation of CD103(+) dendritic cell progenitors at the tumor site enhances tumor responses to therapeutic PD-L1 and BRAF inhibition. Immunity (2016) 44:924–38. doi: 10.1016/j.immuni.2016.03.012
59. Spranger S, Dai D, Horton B, Gajewski TF. Tumor-residing batf3 dendritic cells are required for effector t cell trafficking and adoptive t cell therapy. Cancer Cell (2017) 31:711–23.e4. doi: 10.1016/j.ccell.2017.04.003
60. Broz ML, Binnewies M, Boldajipour B, Nelson AE, Pollack JL, Erle DJ, et al. Dissecting the tumor myeloid compartment reveals rare activating antigen-presenting cells critical for T cell immunity. Cancer Cell (2014) 26:638–52. doi: 10.1016/j.ccell.2014.09.007
61. Bruhns P. Properties of mouse and human IgG receptors and their contribution to disease models. Blood (2012) 119:5640–9. doi: 10.1182/blood-2012-01-380121
62. Bournazos S, Gupta A, Ravetch JV. The role of IgG fc receptors in antibody-dependent enhancement. Nat Rev Immunol (2020) 20:633–43. doi: 10.1038/s41577-020-00410-0
63. Koene HR, Kleijer M, Algra J, Roos D, von dem Borne AE, de Haas M. Fc gammaRIIIa-158V/F polymorphism influences the binding of IgG by natural killer cell fc gammaRIIIa, independently of the fc gammaRIIIa-48L/R/H phenotype. Blood (1997) 90:1109–14. doi: 10.1182/blood.V90.3.1109
64. Cartron G, Dacheux L, Salles G, Solal-Celigny P, Bardos P, Colombat P, et al. Therapeutic activity of humanized anti-CD20 monoclonal antibody and polymorphism in IgG fc receptor FcgammaRIIIa gene. Blood (2002) 99:754–8. doi: 10.1182/blood.v99.3.754
65. Veeramani S, Wang SY, Dahle C, Blackwell S, Jacobus L, Knutson T, et al. Rituximab infusion induces NK activation in lymphoma patients with the high-affinity CD16 polymorphism. Blood (2011) 118:3347–9. doi: 10.1182/blood-2011-05-351411
66. Treon SP, Hansen M, Branagan AR, Verselis S, Emmanouilides C, Kimby E, et al. Polymorphisms in FcgammaRIIIA (CD16) receptor expression are associated with clinical response to rituximab in waldenstrom's macroglobulinemia. J Clin Oncol (2005) 23:474–81. doi: 10.1200/JCO.2005.06.059
67. Calemma R, Ottaiano A, Trotta AM, Nasti G, Romano C, Napolitano M, et al. Fc gamma receptor IIIa polymorphisms in advanced colorectal cancer patients correlated with response to anti-EGFR antibodies and clinical outcome. J Transl Med (2012) 10:232. doi: 10.1186/1479-5876-10-232
68. Gavin PG, Song N, Kim SR, Lipchik C, Johnson NL, Bandos H, et al. Association of polymorphisms in FCGR2A and FCGR3A with degree of trastuzumab benefit in the adjuvant treatment of ERBB2/HER2-positive breast cancer: analysis of the NSABP b-31 trial. JAMA Oncol (2017) 3:335–41. doi: 10.1001/jamaoncol.2016.4884
69. Musolino A, Naldi N, Bortesi B, Pezzuolo D, Capelletti M, Missale G, et al. Immunoglobulin G fragment c receptor polymorphisms and clinical efficacy of trastuzumab-based therapy in patients with HER-2/neu-positive metastatic breast cancer. J Clin Oncol (2008) 26:1789–96. doi: 10.1200/JCO.2007.14.8957
70. Montalvao F, Garcia Z, Celli S, Breart B, Deguine J, Van Rooijen N, et al. The mechanism of anti-CD20-mediated b cell depletion revealed by intravital imaging. J Clin Invest (2013) 123:5098–103. doi: 10.1172/JCI70972
71. Stevenson GT. Three major uncertainties in the antibody therapy of cancer. Haematologica (2014) 99:1538–46. doi: 10.3324/haematol.2013.084640
72. Taylor RP, Lindorfer MA. The role of complement in mAb-based therapies of cancer. Methods (2014) 65:18–27. doi: 10.1016/j.ymeth.2013.07.027
73. Bakema JE, van Egmond M. Fc receptor-dependent mechanisms of monoclonal antibody therapy of cancer. Curr Top Microbiol Immunol (2014) 382:373–92. doi: 10.1007/978-3-319-07911-0_17
74. Shields RL, Lai J, Keck R, O'Connell LY, Hong K, Meng YG, et al. Lack of fucose on human IgG1 n-linked oligosaccharide improves binding to human fcgamma RIII and antibody-dependent cellular toxicity. J Biol Chem (2002) 277:26733–40. doi: 10.1074/jbc.M202069200
75. Shinkawa T, Nakamura K, Yamane N, Shoji-Hosaka E, Kanda Y, Sakurada M, et al. The absence of fucose but not the presence of galactose or bisecting n-acetylglucosamine of human IgG1 complex-type oligosaccharides shows the critical role of enhancing antibody-dependent cellular cytotoxicity. J Biol Chem (2003) 278:3466–73. doi: 10.1074/jbc.M210665200
76. Chenoweth AM, Wines BD, Anania JC, Mark Hogarth P. Harnessing the immune system via FcgammaR function in immune therapy: a pathway to next-gen mAbs. Immunol Cell Biol (2020) 98:287–304. doi: 10.1111/imcb.12326
77. Tong JTW, Harris PWR, Brimble MA, Kavianinia I. An insight into FDA approved antibody-drug conjugates for cancer therapy. Molecules (2021) 26:5847. doi: 10.3390/molecules26195847
78. Pereira NA, Chan KF, Lin PC, Song Z. The "less-is-more" in therapeutic antibodies: Afucosylated anti-cancer antibodies with enhanced antibody-dependent cellular cytotoxicity. MAbs (2018) 10:693–711. doi: 10.1080/19420862.2018.1466767
79. Wang X, Mathieu M, Brezski RJ. IgG fc engineering to modulate antibody effector functions. Protein Cell (2018) 9:63–73. doi: 10.1007/s13238-017-0473-8
80. Rugo HS, Im SA, Cardoso F, Cortes J, Curigliano G, Musolino A, et al. Efficacy of margetuximab vs trastuzumab in patients with pretreated ERBB2-positive advanced breast cancer: a phase 3 randomized clinical trial. JAMA Oncol (2021) 7:573–84. doi: 10.1001/jamaoncol.2020.7932
81. Gauthier L, Morel A, Anceriz N, Rossi B, Blanchard-Alvarez A, Grondin G, et al. Multifunctional natural killer cell engagers targeting NKp46 trigger protective tumor immunity. Cell (2019) 177:1701–13.e16. doi: 10.1016/j.cell.2019.04.041
82. Watkins-Yoon J, Guzman W, Oliphant A, Haserlat S, Leung A, Chottin C, et al. CTX-8573, an innate-cell engager targeting BCMA, is a highly potent multispecific antibody for the treatment of multiple myeloma. Blood (2019) 134:3182. doi: 10.1182/blood-2019-128749
83. Chang GP, Cheung AF, Haney W, LUNDE BM, Prinz B. United States Patent US20200231678A1. United States Patent and Trademark Office (2020)
84. US National library of medicine. ClinicalTrials.gov. Available at: https://www.clinicaltrials.gov/ct2/show/NCT04143711.
85. Wingert S, Reusch U, Knackmuss S, Kluge M, Damrat M, Pahl J, et al. Preclinical evaluation of AFM24, a novel CD16A-specific innate immune cell engager targeting EGFR-positive tumors. MAbs (2021) 13:1950264. doi: 10.1080/19420862.2021.1950264
86. Reusch U, Burkhardt C, Fucek I, Le Gall F, Le Gall M, Hoffmann K, et al. A novel tetravalent bispecific TandAb (CD30/CD16A) efficiently recruits NK cells for the lysis of CD30+ tumor cells. MAbs (2014) 6:728–39. doi: 10.4161/mabs.28591
87. Bartlett NL, Herrera AF, Domingo-Domenech E, Mehta A, Forero-Torres A, Garcia-Sanz R, et al. A phase 1b study of AFM13 in combination with pembrolizumab in patients with relapsed or refractory Hodgkin lymphoma. Blood (2020) 136:2401–9. doi: 10.1182/blood.2019004701
88. Pende D, Falco M, Vitale M, Cantoni C, Vitale C, Munari E, et al. Killer ig-like receptors (KIRs): Their role in NK cell modulation and developments leading to their clinical exploitation. Front Immunol (2019) 10:1179. doi: 10.3389/fimmu.2019.01179
89. Ruggeri L, Capanni M, Urbani E, Perruccio K, Shlomchik WD, Tosti A, et al. Effectiveness of donor natural killer cell alloreactivity in mismatched hematopoietic transplants. Science (2002) 295:2097–100. doi: 10.1126/science.1068440
90. Miller JS, Soignier Y, Panoskaltsis-Mortari A, McNearney SA, Yun GH, Fautsch SK, et al. Successful adoptive transfer and in vivo expansion of human haploidentical NK cells in patients with cancer. Blood (2005) 105:3051–7. doi: 10.1182/blood-2004-07-2974
91. Rubnitz JE, Inaba H, Ribeiro RC, Pounds S, Rooney B, Bell T, et al. NKAML: a pilot study to determine the safety and feasibility of haploidentical natural killer cell transplantation in childhood acute myeloid leukemia. J Clin Oncol (2010) 28:955–9. doi: 10.1200/JCO.2009.24.4590
92. Romagne F, Andre P, Spee P, Zahn S, Anfossi N, Gauthier L, et al. Preclinical characterization of 1-7F9, a novel human anti-KIR receptor therapeutic antibody that augments natural killer-mediated killing of tumor cells. Blood (2009) 114:2667–77. doi: 10.1182/blood-2009-02-206532
93. Korde N, Carlsten M, Lee MJ, Minter A, Tan E, Kwok M, et al. A phase II trial of pan-KIR2D blockade with IPH2101 in smoldering multiple myeloma. Haematologica (2014) 99:e81–3. doi: 10.3324/haematol.2013.103085
94. Carlsten M, Korde N, Kotecha R, Reger R, Bor S, Kazandjian D, et al. Checkpoint inhibition of KIR2D with the monoclonal antibody IPH2101 induces contraction and hyporesponsiveness of NK cells in patients with myeloma. Clin Cancer Res (2016) 22:5211–22. doi: 10.1158/1078-0432.CCR-16-1108
95. Munshi NC, Anderson LD Jr., Shah N, Madduri D, Berdeja J, Lonial S, et al. Idecabtagene vicleucel in relapsed and refractory multiple myeloma. N Engl J Med (2021) 384:705–16. doi: 10.1056/NEJMoa2024850
96. Schuster SJ, Bishop MR, Tam CS, Waller EK, Borchmann P, McGuirk JP, et al. Tisagenlecleucel in adult relapsed or refractory diffuse large b-cell lymphoma. N Engl J Med (2019) 380:45–56. doi: 10.1056/NEJMoa1804980
97. Neelapu SS, Locke FL, Bartlett NL, Lekakis LJ, Miklos DB, Jacobson CA, et al. Axicabtagene ciloleucel CAR T-cell therapy in refractory Large b-cell lymphoma. N Engl J Med (2017) 377:2531–44. doi: 10.1056/NEJMoa1707447
98. Locke FL, Miklos DB, Jacobson CA, Perales MA, Kersten MJ, Oluwole OO, et al. Axicabtagene ciloleucel as second-line therapy for Large b-cell lymphoma. N Engl J Med (2022) 386:640–54. doi: 10.1056/NEJMoa2116133
99. Anderson MK, Torosyan A, Halford Z. Brexucabtagene autoleucel: A novel chimeric antigen receptor t-cell therapy for the treatment of mantle cell lymphoma. Ann Pharmacother (2022) 56:609–19. doi: 10.1177/10600280211026338
100. Abramson JS, Palomba ML, Gordon LI, Lunning MA, Wang M, Arnason J, et al. Lisocabtagene maraleucel for patients with relapsed or refractory large b-cell lymphomas (TRANSCEND NHL 001): a multicentre seamless design study. Lancet (2020) 396:839–52. doi: 10.1016/S0140-6736(20)31366-0
101. Zhao WH, Liu J, Wang BY, Chen YX, Cao XM, Yang Y, et al. A phase 1, open-label study of LCAR-B38M, a chimeric antigen receptor T cell therapy directed against b cell maturation antigen, in patients with relapsed or refractory multiple myeloma. J Hematol Oncol (2018) 11:141. doi: 10.1186/s13045-018-0681-6
102. Neelapu SS, Tummala S, Kebriaei P, Wierda W, Gutierrez C, Locke FL, et al. Chimeric antigen receptor T-cell therapy - assessment and management of toxicities. Nat Rev Clin Oncol (2018) 15:47–62. doi: 10.1038/nrclinonc.2017.148
103. Liu E, Marin D, Banerjee P, Macapinlac HA, Thompson P, Basar R, et al. Use of CAR-transduced natural killer cells in CD19-positive lymphoid tumors. N Engl J Med (2020) 382:545–53. doi: 10.1056/NEJMoa1910607
104. Kim EJ, Cho YH, Kim DH, Ko DH, Do EJ, Kim SY, et al. A phase I/IIa randomized trial evaluating the safety and efficacy of SNK01 plus pembrolizumab in patients with stage IV non-small cell lung cancer. Cancer Res Treat (2021). doi: 10.4143/crt.2021.986
105. Parkhurst MR, Riley JP, Dudley ME, Rosenberg SA. Adoptive transfer of autologous natural killer cells leads to high levels of circulating natural killer cells but does not mediate tumor regression. Clin Cancer Res (2011) 17:6287–97. doi: 10.1158/1078-0432.CCR-11-1347
106. Sakamoto N, Ishikawa T, Kokura S, Okayama T, Oka K, Ideno M, et al. Phase I clinical trial of autologous NK cell therapy using novel expansion method in patients with advanced digestive cancer. J Transl Med (2015) 13:277. doi: 10.1186/s12967-015-0632-8
107. Bachanova V, Cooley S, Defor TE, Verneris MR, Zhang B, McKenna DH, et al. Clearance of acute myeloid leukemia by haploidentical natural killer cells is improved using IL-2 diphtheria toxin fusion protein. Blood (2014) 123:3855–63. doi: 10.1182/blood-2013-10-532531
108. Cooley S, He F, Bachanova V, Vercellotti GM, DeFor TE, Curtsinger JM, et al. First-in-human trial of rhIL-15 and haploidentical natural killer cell therapy for advanced acute myeloid leukemia. Blood Adv (2019) 3:1970–80. doi: 10.1182/bloodadvances.2018028332
109. Romee R, Rosario M, Berrien-Elliott MM, Wagner JA, Jewell BA, Schappe T, et al. Cytokine-induced memory-like natural killer cells exhibit enhanced responses against myeloid leukemia. Sci Transl Med (2016) 8:357ra123. doi: 10.1126/scitranslmed.aaf2341
110. Luevano M, Daryouzeh M, Alnabhan R, Querol S, Khakoo S, Madrigal A, et al. The unique profile of cord blood natural killer cells balances incomplete maturation and effective killing function upon activation. Hum Immunol (2012) 73:248–57. doi: 10.1016/j.humimm.2011.12.015
111. Shah N, Martin-Antonio B, Yang H, Ku S, Lee DA, Cooper LJ, et al. Antigen presenting cell-mediated expansion of human umbilical cord blood yields log-scale expansion of natural killer cells with anti-myeloma activity. PLoS One (2013) 8:e76781. doi: 10.1371/journal.pone.0076781
112. Spanholtz J, Preijers F, Tordoir M, Trilsbeek C, Paardekooper J, de Witte T, et al. Clinical-grade generation of active NK cells from cord blood hematopoietic progenitor cells for immunotherapy using a closed-system culture process. PLoS One (2011) 6:e20740. doi: 10.1371/journal.pone.0020740
113. Dolstra H, Roeven MWH, Spanholtz J, Hangalapura BN, Tordoir M, Maas F, et al. Successful transfer of umbilical cord blood CD34(+) hematopoietic stem and progenitor-derived NK cells in older acute myeloid leukemia patients. Clin Cancer Res (2017) 23:4107–18. doi: 10.1158/1078-0432.CCR-16-2981
114. Tonn T, Becker S, Esser R, Schwabe D, Seifried E. Cellular immunotherapy of malignancies using the clonal natural killer cell line NK-92. J Hematother Stem Cell Res (2001) 10:535–44. doi: 10.1089/15258160152509145
115. Gong JH, Maki G, Klingemann HG. Characterization of a human cell line (NK-92) with phenotypical and functional characteristics of activated natural killer cells. Leukemia (1994) 8:652–8.
116. Suck G, Odendahl M, Nowakowska P, Seidl C, Wels WS, Klingemann HG, et al. NK-92: an 'off-the-shelf therapeutic' for adoptive natural killer cell-based cancer immunotherapy. Cancer Immunol Immunother (2016) 65:485–92. doi: 10.1007/s00262-015-1761-x
117. Klingemann H, Boissel L, Toneguzzo F. Natural killer cells for immunotherapy - advantages of the NK-92 cell line over blood NK cells. Front Immunol (2016) 7:91. doi: 10.3389/fimmu.2016.00091
118. Boyiadzis M, Agha M, Redner RL, Sehgal A, Im A, Hou JZ, et al. Phase 1 clinical trial of adoptive immunotherapy using "off-the-shelf" activated natural killer cells in patients with refractory and relapsed acute myeloid leukemia. Cytotherapy (2017) 19:1225–32. doi: 10.1016/j.jcyt.2017.07.008
119. Williams BA, Law AD, Routy B, denHollander N, Gupta V, Wang XH, et al. A phase I trial of NK-92 cells for refractory hematological malignancies relapsing after autologous hematopoietic cell transplantation shows safety and evidence of efficacy. Oncotarget (2017) 8:89256–68. doi: 10.18632/oncotarget.19204
120. Knorr DA, Ni Z, Hermanson D, Hexum MK, Bendzick L, Cooper LJ, et al. Clinical-scale derivation of natural killer cells from human pluripotent stem cells for cancer therapy. Stem Cells Transl Med (2013) 2:274–83. doi: 10.5966/sctm.2012-0084
121. Li Y, Hermanson DL, Moriarity BS, Kaufman DS. Human iPSC-derived natural killer cells engineered with chimeric antigen receptors enhance anti-tumor activity. Cell Stem Cell (2018) 23:181–92.e5. doi: 10.1016/j.stem.2018.06.002
122. Thomson JA, Itskovitz-Eldor J, Shapiro SS, Waknitz MA, Swiergiel JJ, Marshall VS, et al. Embryonic stem cell lines derived from human blastocysts. Science (1998) 282:1145–7. doi: 10.1126/science.282.5391.1145
123. Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell (2006) 126:663–76. doi: 10.1016/j.cell.2006.07.024
124. Kim PS, Kwilas AR, Xu W, Alter S, Jeng EK, Wong HC, et al. IL-15 superagonist/IL-15RalphaSushi-Fc fusion complex (IL-15SA/IL-15RalphaSu-Fc; ALT-803) markedly enhances specific subpopulations of NK and memory CD8+ T cells, and mediates potent anti-tumor activity against murine breast and colon carcinomas. Oncotarget (2016) 7:16130–45. doi: 10.18632/oncotarget.7470
125. Woan KV, Kim H, Bjordahl R, Davis ZB, Gaidarova S, Goulding J, et al. Harnessing features of adaptive NK cells to generate iPSC-derived NK cells for enhanced immunotherapy. Cell Stem Cell (2021) 28:2062–75.e5. doi: 10.1016/j.stem.2021.08.013
126. Zhu H, Blum RH, Bjordahl R, Gaidarova S, Rogers P, Lee TT, et al. Pluripotent stem cell-derived NK cells with high-affinity noncleavable CD16a mediate improved antitumor activity. Blood (2020) 135:399–410. doi: 10.1182/blood.2019000621
127. Zhu H, Blum RH, Bernareggi D, Ask EH, Wu Z, Hoel HJ, et al. Metabolic reprograming via deletion of CISH in human iPSC-derived NK cells promotes In vivo persistence and enhances anti-tumor activity. Cell Stem Cell (2020) 27:224–37.e6. doi: 10.1016/j.stem.2020.05.008
128. Hermanson DL, Bendzick L, Pribyl L, McCullar V, Vogel RI, Miller JS, et al. Induced pluripotent stem cell-derived natural killer cells for treatment of ovarian cancer. Stem Cells (2016) 34:93–101. doi: 10.1002/stem.2230
129. Zeng J, Tang SY, Toh LL, Wang S. Generation of "off-the-shelf" natural killer cells from peripheral blood cell-derived induced pluripotent stem cells. Stem Cell Rep (2017) 9:1796–812. doi: 10.1016/j.stemcr.2017.10.020
130. Goldenson BH, Zhu H, Wang YM, Heragu N, Bernareggi D, Ruiz-Cisneros A, et al. Umbilical cord blood and ipsc-derived natural killer cells demonstrate key differences in cytotoxic activity and KIR profiles. Front Immunol (2020) 11:561553. doi: 10.3389/fimmu.2020.561553
131. US National library of medicine. ClinicalTrials.gov. Available at: https://www.clinicaltrials.gov/ct2/show/NCT04245722.
132. US National library of medicine. ClinicalTrials.gov. Available at: https://www.clinicaltrials.gov/ct2/show/NCT04023071.
133. US National library of medicine. ClinicalTrials.gov. Available at: https://www.clinicaltrials.gov/ct2/show/NCT04630769.
134. US National library of medicine. ClinicalTrials.gov. Available at: https://www.clinicaltrials.gov/ct2/show/NCT04614636.
135. US National library of medicine. ClinicalTrials.gov. Available at: https://www.clinicaltrials.gov/ct2/show/NCT05182073.
136. Goldenson BH, Hor P, Kaufman DS. iPSC-derived natural killer cell therapies - expansion and targeting. Front Immunol (2022) 13:841107. doi: 10.3389/fimmu.2022.841107
137. Bachanova V, Cayci Z, Lewis D, Maakaron JE, Janakiram M, Bartz A, et al. Initial clinical activity of FT596, a first-in-class, multi-antigen targeted, off-the-shelf, iPSC-derived CD19 CAR NK cell therapy in Relapsed/Refractory b-cell lymphoma. Blood (2020) 136:8. doi: 10.1182/blood-2020-141606
138. Strati P, Bachanova V, Goodman A, Pagel JM, Castro JE, Griffis K, et al. Preliminary results of a phase I trial of FT516, an off-the-shelf natural killer (NK) cell therapy derived from a clonal master induced pluripotent stem cell (iPSC) line expressing high-affinity, non-cleavable CD16 (hnCD16), in patients (pts) with relapsed/refractory (R/R) b-cell lymphoma (BCL). J Clin Oncol (2021) 39:7541. doi: 10.1200/JCO.2021.39.15_suppl.7541
139. Patel K, Bachanova V, Goodman AM, Pagel JM, Griffis K, Anderson M, et al. Phase I study of FT516, an off-the-Shelf iPSC-derived NK cell therapy, in combination with rituximab in patients with Relapsed/Refractory b-cell lymphoma. Blood (2021) 3873. doi: 10.1182/blood-2021-151520
140. Goodridge JP, Mahmood S, Zhu H, Gaidarova S, Blum R, Bjordahl R, et al. FT596: translation of first-of-kind multi-antigen targeted off-the-shelf CAR-NK cell with engineered persistence for the treatment of b cell malignancies. Blood (2019) 134:301. doi: 10.1182/blood-2019-129319
141. US National library of medicine. ClinicalTrials.gov. Available at: https://www.clinicaltrials.gov/ct2/show/NCT04245722?cond=NCT04245722.
142. Bachanova V, Ghobadi A, Patel K, Park JH, Flinn IW, Shah P, et al. Safety and efficacy of FT596, a first-in-class, multi-antigen targeted, off-the-shelf, iPSC-derived CD19 CAR NK cell therapy in relapsed/refractory b-cell lymphoma. Blood (2021) 138. doi: 10.1182/blood-2021-151185
143. Cichocki F, Bjordahl R, Gaidarova S, Mahmood S, Abujarour R, Wang H, et al. iPSC-derived NK cells maintain high cytotoxicity and enhance in vivo tumor control in concert with T cells and anti-PD-1 therapy. Sci Transl Med (2020) 12:eaaz5618. doi: 10.1126/scitranslmed.aaz5618
144. US National library of medicine. ClinicalTrials.gov. Available at: https://www.clinicaltrials.gov/ct2/show/NCT04074746.
145. Kerbauy LN, Marin ND, Kaplan M, Banerjee PP, Berrien-Elliott MM, Becker-Hapak M, et al. Combining AFM13, a bispecific CD30/CD16 antibody, with cytokine-activated blood and cord blood–derived NK cells facilitates CAR-like responses against CD30+ malignancies. Clin Cancer Res (2021) 27:3744–56. doi: 10.1158/1078-0432.CCR-21-0164
146. Affimed website. Available at: https://www.affimed.com/wp-content/uploads/04.
147. Souza-Fonseca-Guimaraes F, Cursons J, Huntington ND. The emergence of natural killer cells as a major target in cancer immunotherapy. Trends Immunol (2019) 40:142–58. doi: 10.1016/j.it.2018.12.003
148. Delconte RB, Kolesnik TB, Dagley LF, Rautela J, Shi W, Putz EM, et al. CIS is a potent checkpoint in NK cell-mediated tumor immunity. Nat Immunol (2016) 17:816–24. doi: 10.1038/ni.3470
149. Paolino M, Choidas A, Wallner S, Pranjic B, Uribesalgo I, Loeser S, et al. The E3 ligase cbl-b and TAM receptors regulate cancer metastasis via natural killer cells. Nature (2014) 507:508–12. doi: 10.1038/nature12998
150. Parameswaran R, Ramakrishnan P, Moreton SA, Xia Z, Hou Y, Lee DA, et al. Repression of GSK3 restores NK cell cytotoxicity in AML patients. Nat Commun (2016) 7:11154. doi: 10.1038/ncomms11154
151. Putz EM, Gotthardt D, Hoermann G, Csiszar A, Wirth S, Berger A, et al. CDK8-mediated STAT1-S727 phosphorylation restrains NK cell cytotoxicity and tumor surveillance. Cell Rep (2013) 4:437–44. doi: 10.1016/j.celrep.2013.07.012
152. Witalisz-Siepracka A, Gotthardt D, Prchal-Murphy M, Didara Z, Menzl I, Prinz D, et al. NK cell-specific CDK8 deletion enhances antitumor responses. Cancer Immunol Res (2018) 6:458–66. doi: 10.1158/2326-6066.CIR-17-0183
Keywords: natural killer (NK) cells, immune checkpoint, cancer, immunotherapy, NK cell therapy, iPSC-NK
Citation: Chen X, Jiang L and Liu X (2022) Natural killer cells: the next wave in cancer immunotherapy. Front. Immunol. 13:954804. doi: 10.3389/fimmu.2022.954804
Received: 27 May 2022; Accepted: 30 June 2022;
Published: 27 July 2022.
Edited by:
Hermann Einsele, Julius Maximilian University of Würzburg, GermanyReviewed by:
Alexander Steinle, Goethe University Frankfurt, GermanyCopyright © 2022 Chen, Jiang and Liu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xin Chen, WGluLkNoZW5AQmVpR2VuZS5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.