 Vanessa Fernandes Rodrigues
Vanessa Fernandes Rodrigues Jefferson Elias-Oliveira
Jefferson Elias-Oliveira Ítalo Sousa Pereira
Ítalo Sousa Pereira Jéssica Assis Pereira
Jéssica Assis Pereira Sara Cândida Barbosa
Sara Cândida Barbosa Melissa Santana Gonsalez Machado
Melissa Santana Gonsalez Machado Daniela Carlos*
Daniela Carlos*- Department of Biochemistry and Immunology, Ribeirão Preto Medical School, University of São Paulo, Ribeirão Preto, Brazil
Akkermansia muciniphila is a Gram-negative anaerobic mucus-layer-degrading bacterium that colonizes the intestinal mucosa of humans and rodents. Metagenomic data have shown an inverse correlation between the abundance of A. muciniphila and diseases such as inflammatory bowel disease (IBD), obesity, and diabetes. Thus, in recent decades, the potential of this bacterium as an immunomodulatory probiotic for autoimmune and chronic inflammatory diseases has been explored in experimental models. Corroborating these human correlation data, it has been reported that A. muciniphila slows down the development and progression of diabetes, obesity, and IBD in mice. Consequently, clinical studies with obese and diabetic patients are being performed, and the preliminary results are very promising. Therefore, this mini review highlights the main findings regarding the beneficial roles of A. muciniphila and its action mechanisms in autoimmune and chronic inflammatory diseases.
Introduction
The intestine is mainly colonized by four phyla of bacteria: Firmicutes, Bacteroidetes, Proteobacteria, and Actinobacteria (1). Several factors, such as the use of antibiotics, diet, and pH can interfere with the gut microbiota. It is known that alterations in the gut microbiota (dysbiosis) are capable of inducing abnormal immune responses in the gut-associated lymphatic tissue and that these alterations can compromise the systemic immune response (2). The gut microbiota regulates the host immune response through two main mechanisms: activating the innate immune response via the Toll-like receptor (TLR) (3) and/or activating free fatty acid receptors (FFAR) via microbial metabolites such as short-chain fatty acids (SCFAs), including acetate, propionate, and butyrate. In addition, these metabolites can induce the differentiation of naive T cells into regulatory T cells (Tregs) or their migration into the intestine (4). Intestinal dysbiosis can lead to excessive activation of TLRs and a low production of SCFAs, contributing to the development of a number of gastrointestinal diseases, obesity, and diabetes (5–7). Because some probiotic bacteria in the gut can suppress chronic inflammatory and autoimmune diseases, the use of probiotics, like Bifidobacteria, Lactobacilli, Lactococci, and Streptococci, as prophylactics and/or therapeutic tools for these diseases has been investigated (8). More recently, Akkermansia muciniphila has been shown to be a promising probiotic (9).
A. muciniphila is a Gram-negative, anaerobic, oval-shaped bacterium that degrades the mucus layer. The analysis of the 16S rRNA gene sequence showed that this species belongs to the Verrucomicrobia phylum (10). A. muciniphila colonizes the intestinal tract early in life and comprises approximately 3% of the total microbiota in healthy adults (11). Upon degrading mucin, A. muciniphila produces acetate and propionate, which serve as substrates for other bacteria and the host (10, 12). SCFAs have also been linked to the regulation of body weight gain through their anorexic, anti-inflammatory, and metabolic effects (13, 14). As A. muciniphila is lower in the gut of humans and mice with autoimmune and metabolic diseases (15–17), this review highlights the immunomodulatory potential of A. muciniphila in these diseases.
Role of A. muciniphila in Protecting Against Inflammatory Bowel Disease
It is known that the impairment of homeostasis and the integrity of the intestinal barrier result in the development of metabolic and gastrointestinal disorders (18, 19). The intestinal mucosal barrier has evolved to maintain a balance between the absorption of essential nutrients and the prevention of pathogen translocation (20). The integrity of the intestinal epithelium is maintained by tight junctions (TJs), adherens junctions (AJs), and desmosome complexes of the epithelium, whose expression can be increased by probiotics or compounds produced by them, such as extracellular vesicles (EVs) or outer membrane microvesicles (OMVs) in the case of Gram-negative bacteria (21–23). The disruption of the integrity of the intestinal mucosa drives the development of inflammatory bowel disease (IBD), such as ulcerative colitis (UC) and Crohn’s disease (CD), which are chronic idiopathic inflammatory diseases characterized by an exaggerated immune response to gut microbiota, resulting in tissue damage (24, 25). It has been reported that antibiotic use during childhood alters the gut microbiota and increases susceptibility to IBD, suggesting an important role of gut microbiota in the maintenance of intestinal homeostasis (26). Several studies have found differences in the composition of the gut microbiota between healthy people and patients with IBD, with a remarkable reduction in A. muciniphila in patients with UC (16, 27). Furthermore, it has been reported that A. muciniphila or Amuc_1100 (an outer membrane protein of A. muciniphila) can attenuate DSS-induced colitis in mice. The modulating effect of A. muciniphila-derived Amuc_1100 in colitis was associated with a reduction in infiltrating macrophages, CD8+ cytotoxic T lymphocytes, and pro-inflammatory cytokines, such as tumor necrosis factor-α (TNF-α), interleukin (IL)-1α, IL-6, IL-12, macrophage inflammatory protein-1 (MIP-1) α, granulocyte colony-stimulating factor, and chemokine (C-X-C motif) ligand 1 (CXCL1) in the colon. In addition, A. muciniphila administration reduced CD16/32+ macrophages in the spleen and mesenteric lymph nodes (MLNs) of mice with colitis (28, 29).
A recent study has shown that Amuc_2109, an enzyme secreted by A. muciniphila, also attenuated DSS-induced colitis in mice, increasing the expression of TJs and reducing the expression of the NLRP3 inflammasome (30). However, the protective effect of viable A. muciniphila against DSS-induced colitis was shown to be dependent of NLRP3 activation (27). Indeed, the role of NLRP3 in the regulation of intestinal homeostasis was previously elucidated, since NLRP3-/- mice are more susceptible to the development of experimentally induced colitis (31).Additionally, it was demonstrated that the administration of A. muciniphila induced the proliferation of intestinal stem cells and boosted the differentiation of Paneth and goblet cells in the small intestine and colon of healthy mice or mice with gut damage caused by radiation and methotrexate. In the same study, the beneficial effect of A. muciniphila in the intestinal tract was associated with a greater amount of acetic and propionic acids in the cecal content of mice treated with A. muciniphila (32), thus demonstrating that this bacterium contributes to the tissue repair of the intestinal mucosa and that the production of SCFAs is involved in this process. Although A. muciniphila is a common component of the human and murine gastrointestinal tracts and has a beneficial role for the integrity of the intestinal mucosa, when intestinal dysbiosis occurs, the colonization by A. muciniphila can exacerbate the inflammation (11, 33). A previous study reported that treatment with A. muciniphila led to the worsening of intestinal inflammation caused by Salmonella enterica Typhimurium infection in gnotobiotic mice, which was related to a decrease in goblet cells and an increase in the expression of pro-inflammatory cytokines in the cecum (34). However, recently, Ring et al. (2019) demonstrated that in IL-10-deficient (IL-10-/-) mice, which spontaneously develop colitis, colonization by A. muciniphila had no effect on intestinal inflammation (35).
Interestingly, several studies have indicated that the outer membrane compounds of A. muciniphila, or pasteurized bacteria, have greater therapeutic potential for metabolic, inflammatory, and autoimmune diseases than live A. muciniphila (36–38). Notably, Kang et al. (2013) reported a change in the composition of EVs in the feces of mice with DSS-induced UC, such as a decrease in the EVs of A. muciniphila and Bacteroides acidifaciens. In the same study, OMVs from A. muciniphila (AmOMV) suppressed the production of IL-6 in colonic epithelial cells (CT26 cell line) stimulated with OMVs from Escherichia coli in vitro, and oral administration of AmOMV, but not viable bacteria, attenuated DSS-induced colitis in vivo (36). Additionally, in a murine model of high-fat diet (HFD)–induced intestinal dysbiosis, AmOMVs improved the intestinal mucosal barrier function, increased the expression of TJs and IL-10, and inhibited inflammatory markers in the colon. AmOMVs are also able to reduce intestinal permeability, increase the expression of TJs via AMP-activated protein kinase (AMPK), inhibit TLR-4 and interferon-alpha (IFN-α) expression, and increase TLR-2 expression and IL-4 production in Caco-2 cell lines in vitro (39, 40). These data indicate that A. muciniphila components and their OMVs may be potential therapeutic targets for IBD.
Role of A. muciniphila in Protecting Against Obesity and Metabolic Syndrome
The obesity is related to gut dysbiosis as an imbalance between energy consumption and expenditure favors the prevalence of pathobiont bacteria (41–43). Metagenomic studies have shown that the abundance of A. muciniphila is negatively correlated with body weight in humans (15, 44, 45). The analysis of gut microbiota in feces showed that obese and overweight children had reduced A. muciniphila concentrations compared with lean children (44). Significant multivariate linear associations using microbiome multivariable association with linear models revealed that the abundance of A. muciniphila was negatively correlated with fasting blood glucose levels and the body mass index (BMI), suggesting that these bacteria can act in the control of obesity and diabetes (46). Similarly, the experimental obesity model like mice fed an HFD or high-sucrose diet (HSD) and genetically obese mice (ob/ob) showed a negative correlation with the abundance of the Akkermansia genera and adiposity, body weight, liver and adipose tissue inflammation, blood glucose serum, insulin, and triglyceride levels (47–49).
Since there is an inverse correlation between the abundance of A. muciniphila and metabolic diseases, several experimental studies have used it as a potential probiotic for therapeutic tools for these diseases (15, 45–47). Similarly, some findings indicate that A. muciniphila adheres to the intestinal epithelium and strengthens the integrity of the enterocyte monolayer in vitro, suggesting its ability to contribute to the integrity of the intestinal barrier in obese individuals (50). In a murine model of obesity induced by HFD, it was demonstrated that treatment with A. muciniphila 108–109 Colony-forming unit (CFU)/mL was able to reduce body weight gain, the accumulation of white adipose tissue (WAT), and energy efficiency and improve liver function impaired by HFD in mice (Figure 1) (51, 52). Furthermore, it was reported in the HFD-induced obesity model that the pasteurized form or AmOMV has been successful in reducing or preventing hepatic steatosis and adipogenesis/lipogenesis (38, 53). In addition, the increase in Akkermansia ssp. abundance contributed to the antidiabetic effects of metformin in mice fed with HFD, improving glucose tolerance and increasing the population of goblet cells and Treg cells in adipose tissue (54). Kong et al. (2019) reported that intestinal dysbiosis triggered by high-calorie diets in mice can be alleviated by the administration of probiotics that restore proportions and increase the abundance of beneficial bacteria, including Akkermansia ssp (55).
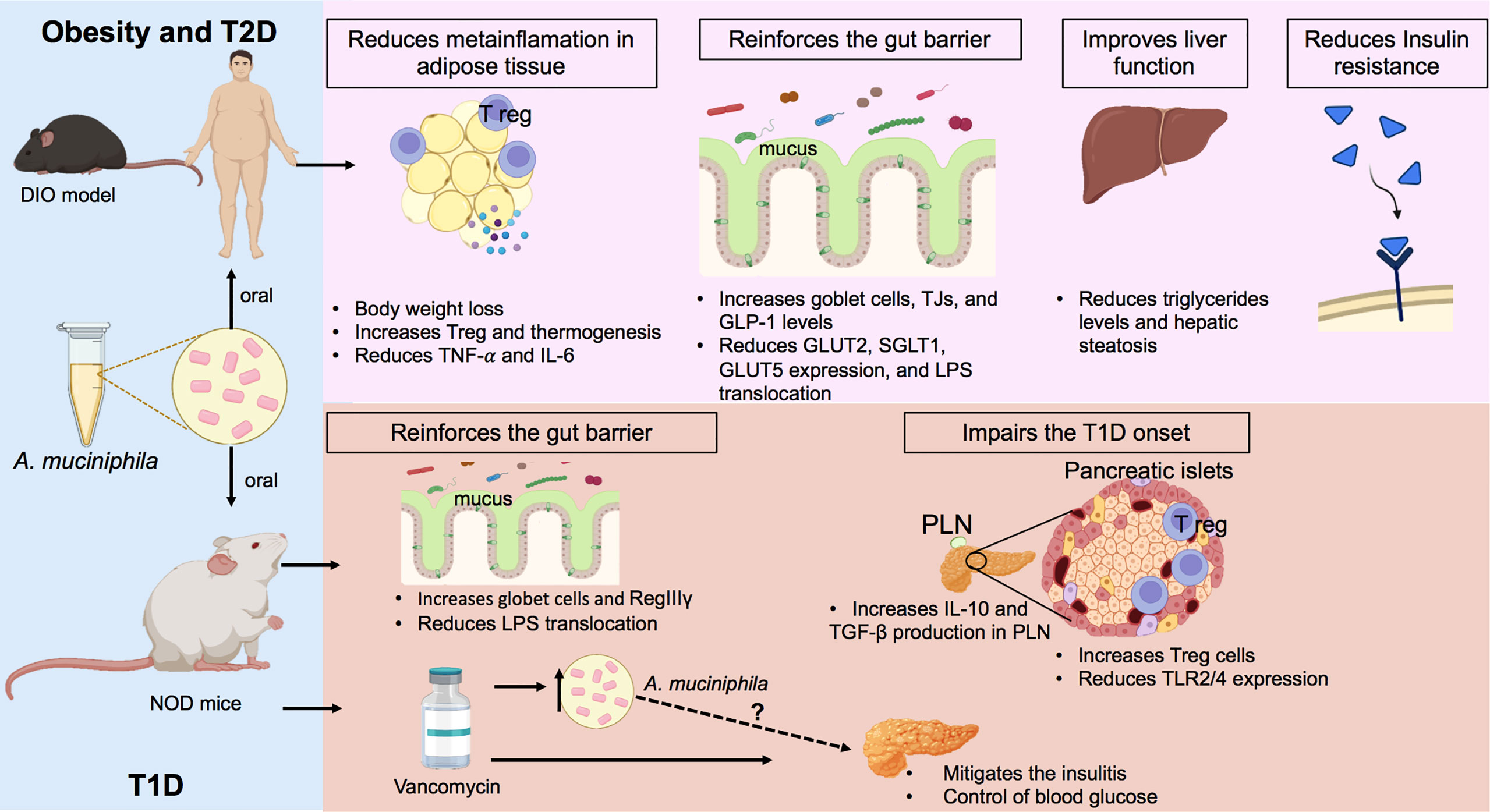
Figure 1 Regulatory effects of A. muciniphila on obesity, T2D, and T1D. In the models of diet-induced obesity (DIO), oral treatment with A. muciniphila reduces weight gain, controls fat accumulation, increases regulatory T cells (T regs), and decreases the production of pro-inflammatory cytokines in the adipose tissue of mice fed with a high-fat diet (HFD). In the intestine, this probiotic increases goblet cells and mucus production, in addition to inducing a greater production of glucagon-like peptide-1 (GLP-1), which controls glucose absorption. The administration of pasteurized A. muciniphila to obese subjects decreased body weight and hip circumference, improved insulin sensitivity, and reduced the markers of liver damage. In NOD mice, the model of T1D (autoimmune), A. muciniphila improved mucus production and increased the antimicrobial peptide RegIIIγ, which contributes to improved intestinal barrier function and the lower translocation of LPS into the circulation. In parallel, the probiotic increases the production of anti-inflammatory cytokines in pancreatic lymph nodes (PLNs) and potentiates the recruitment of T regs in the pancreas, culminating in a delay in the development of T1D. Vancomycin-treated NOD mice showed an enrichment of A. muciniphila in the gut, which is correlated with a lower degree of insulitis and glycemic control. Figure created with BioRender.com. TNF-α: tumor necrosis factor-α; IL-6: interleukin-6, GLP-1: glucagon-like peptide-1; GLUT2, glucose transporter 2; SGLT1, sodium-glucose cotransporter 1; GLUT5, glucose transporter 5; RegIIIγ: regeneration islet-derived III, IL-10: inyerleukin-10; TGF-β, transforming growth factor beta; TLR2/4, Toll-like receptor 2/4; TJs, tight junctions.
The mechanisms by which A. muciniphila regulates obesity and glucose levels have not yet been completely elucidated. A previous study showed that A. muciniphila was able to increase thermogenesis and the secretion of glucagon-like peptide-1 (GLP-1) and reduce the expression of proteins involved in adipose cell differentiation, and the gene expression of glucose and fructose transporters in the jejunum, suggesting that A. muciniphila reduces carbohydrate absorption (Figure 1) (52, 56, 57) Similarly, a clinical trial observed that patients who underwent Roux-en-Y gastric bypass surgery showed an increase in the Akkermansia population together with an improvement in anthropometric and clinical aspects, such as a reduction in BMI and glycated hemoglobin (HbA1c), as well as an increase in GLP-1 levels (58).
Interestingly, it has been reported that the pasteurization of A. muciniphila attenuates metabolic syndrome by reducing body weight, glucose intolerance, insulin resistance, the adipocyte diameter, and the serum levels of leptin and triglycerides (51, 59). In mice fed with HFD, it was also seen that AmOMV showed more suppressive effects, when compared with viable A. muciniphila, on lipid metabolism and the expression of inflammatory markers such as TNF-α and IL-6 in adipose tissues (40). Treatment with this probiotic or AmOMV increased the number of goblet cells and TJs in the colon and restored the gut bacterial diversity that has been affected by HFD (40, 51, 59), indicating that A. muciniphila can preserve gut homeostasis, which can impact the development of obesity and diabetes. Furthermore, A. muciniphila–derived proteins, Amuc_1100 (membrane protein) and P9 (secreted protein), have been reported to reduce obesity-related metabolic syndrome induced by HFD in mice (51, 56). In the case of P9, glucose homeostasis and obesity reduction are related to the interaction of P9 with intercellular adhesion molecule 2 (ICAM-2) and an increase in type 2 macrophages (M2) in an IL-6 dependent pathway (56).
Considering the promising effect of A. muciniphila as a therapeutic tool for metabolic syndrome in mice, some clinical studies have been conducted (60–63). In these clinical studies, obese and/or type 2 diabetes (T2D) patients supplemented with A. muciniphila, alone or in conjunction with other probiotics, showed improvement in the clinical and metabolic status (Table 1). Indeed, a positive effect was also observed in humans, since obese patients who underwent 3 months of treatment with 1 × 1010 CFU of pasteurized A. muciniphila showed a reduction in weight, fat mass, hip circumference, insulin resistance, plasma cholesterol levels, the markers of liver dysfunction, and systemic inflammation; these patients did not experience any side effects (60). It is important to mention that in addition to the two clinical trials already completed and published (Table 1), there are currently two other clinical trials in the recruitment phase, which aim to evaluate the effects of A. muciniphila in obesity and T2D (NCT: NCT04797442) and insulin resistance in healthy individuals with dysglycemia (NCT: NCT05114018).
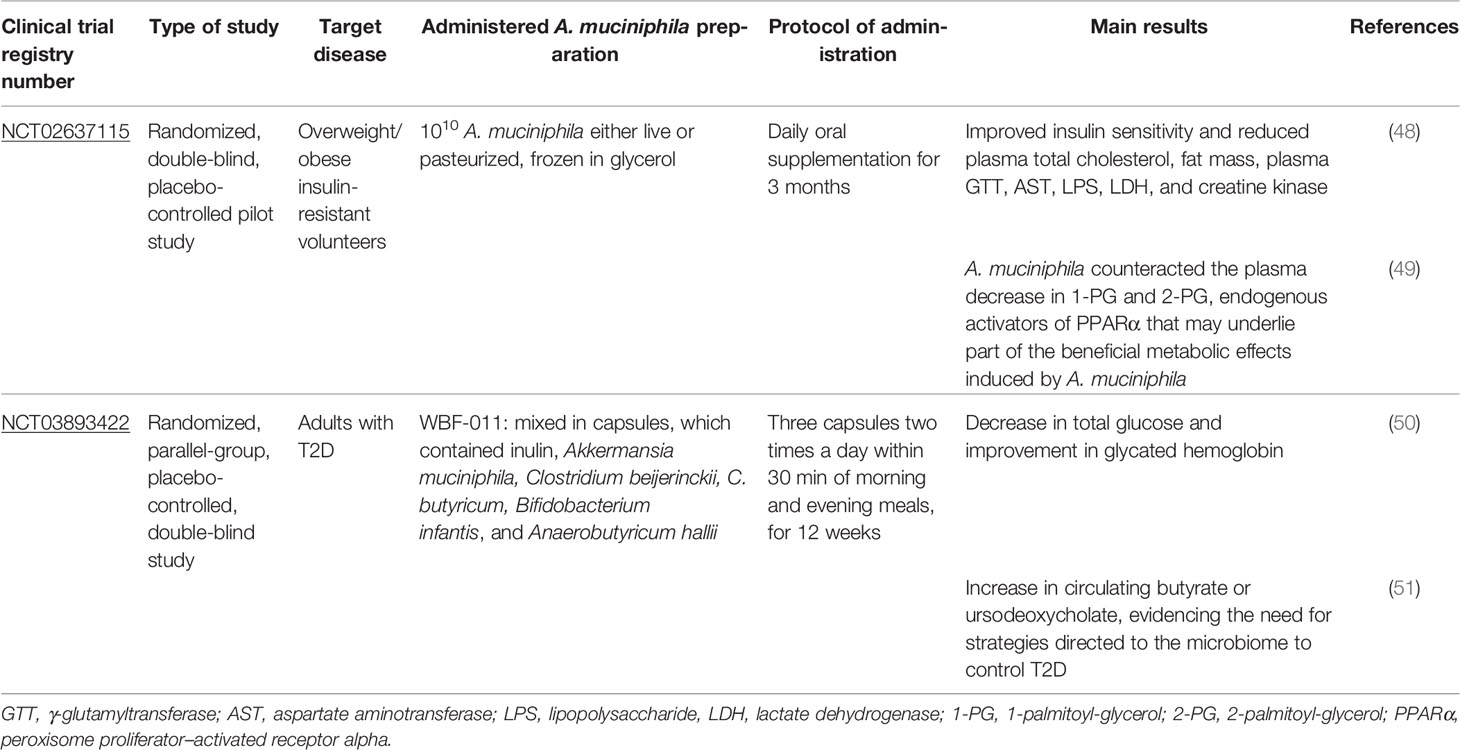
Table 1 Recorded and complete clinical trials using A. muciniphila administration.
Furthermore, obese patients with a higher abundance of A. muciniphila showed an improvement in metabolic profiles, such as total cholesterol levels and insulin sensitivity after caloric restriction, compared with patients with a low abundance of this bacterium (64). Another clinical trial with overweight and obese diabetic patients showed that inulin and butyrate administration decreased the diastolic blood pressure and the expression of TNF-α levels at the same time exerted bifidogenic effects and increased A. muciniphila abundance in these patients (65). Rodriguez et al. (2020) demonstrated that the transplantation of feces from different obese patients to mice with microbiota depleted by antibiotics and fed with HFD differentially responded to inulin supplementation, which was related to the initial gut microbiota composition. In the same study, a positive relationship was observed between an increase in the Akkermansia population and weight reduction, increased insulin production, and reduced hepatic and muscle fat in mice supplemented with inulin (66).
The accumulation of visceral fat that occurs in obesity is a risk factor for the development of T2D as its lipolytic activity and greater recruitment of macrophages with a pro-inflammatory phenotype favor insulin resistance (67, 68). In this regard, A. muciniphila has been shown to control fat accumulation and adipose tissue metabolism, as well as improve glucose homeostasis by reducing adiposity, fasting glucose, and insulin resistance caused by HFD (15). Studies in diet-induced obese mice have shown that a greater abundance of A. muciniphila promoted by the consumption of polyphenols is associated with the prevention of weight gain, reduction of visceral adiposity, prevention of intestinal inflammation, and reduction in circulating liposaccharide (LPS) levels. The results also showed that the increase in the population of Akkermansia ssp. improves insulin sensitivity through the reduction of LPS translocation as this bacterium reduces intestinal permeability (69, 70).
Role of A. muciniphila in Resistance Against Diabetes
The ample evidence for the effect of A. muciniphila in diabetes is controversial; a case–control two-stage metagenome broad association study with individuals with T2D in China indicated the enrichment of A. muciniphila DNA in feces at both evaluation stages (71). However, a variety of metagenomic studies have associated the inverse abundance of A. muciniphila in the gut microbiota of obese, prediabetic, and diabetic humans or mice (48, 72–74).Mice fed an HFD for 16 weeks exhibited an increased mRNA expression of inflammation markers in WAT, including TNF and CCL-2, in addition to developing hyperinsulinemia, hyperglycemia, and higher serum leptin concentrations. All of these metabolic and inflammatory changes were accompanied by a lower abundance of A. muciniphila after 3 weeks of diet and precede peripheral insulin resistance and T2D development (48). However, the influence of the relative abundance of A. muciniphila does not seem to be only involved in the T2D onset. Refractory T2D is characterized by an individual’s inability to achieve optimal glycemic control, marked by the serum levels of HbA1c less than or equal to 8%. A metagenomic study of stool samples indicated that patients with refractory T2D exhibited a lower relative abundance of A. muciniphila than diabetic subjects who achieved optimal glycemic control using metformin or other hypoglycemic agents (75). In an experimental model of T2D induced by streptozotocin (STZ) in rats, viable or pasteurized A. muciniphila attenuated T2D, which was associated with improved liver function and reduced plasma pro-inflammatory factors, gluco/lipotoxicity, and oxidative stress (76).
T2D is characterized by changes in glucose metabolism through the resistance of peripheral tissues to insulin, while T1D is an autoimmune disease in which it is possible to observe the progressive destruction of insulin-producing pancreatic β-cells by an inflammatory cell infiltrate and the production of autoreactive antibodies. Despite the differences in the nature of the development of these pathologies, both diseases are accompanied by gut dysbiosis (77–79). In case of T1D, the cytokine IFN-γ has an important role in the diabetes onset (80). Interestingly, IFN-γ-/- mice show better glucose tolerance and increased gut A. muciniphila relative abundance compared with wild-type mice. In the same study, IFN-γ-/- mice without A. muciniphila showed no improvement in glucose tolerance (81), indicating that the diabetogenic role of IFN-γ may be related to its ability to induce changes in the gut microbiota, especially in the reduction of A. muciniphila.
Interestingly, one study indicated that NOD mice that received oral treatment with the antibiotic vancomycin from birth to day 28 of life predominantly have A. muciniphila in their gut microbiota, which was accompanied by a lower incidence of T1D. Additionally, mice that received vancomycin treatment from the eighth week of life developed less insulitis associated with reduced destruction of pancreatic β-cells (Figure 1) (82). Similarly, diabetes-resistant antibiotic-treated STZ injected mice exhibited an increase in Akkermansia abundance (83). These results suggest a protective effect of A. muciniphila against T1D development. Furthermore, by observing two colonies of NOD mice with different incidence rates of diabetes, it was possible to notice that the colony of NOD mice with a low incidence of T1D presented a greater abundance of A. muciniphila in the gut microbiota. It is worth mentioning that administration of A muciniphila to the colony of NOD mice with high-incidence T1D delayed the T1D outcome (Figure 1) (84). It was seen that regulation of T1D/T2D by A. muciniphila was associated by a local modulation with goblet cell hyperplasia, increased M2 macrophage number and antimicrobial peptide expression in the colon, and the reduction of bacterial translocation. The Akkemansia-induced gut homeostasis was accompanied by increased insulin secretion and Treg lymphocyte numbers in the pancreas from diabetic mice models (74, 76, 84). Together, these studies elucidate the role of gut microbiota in the regulation of metabolic and autoimmune diseases, suggesting that probiotic bacteria, especially A. muciniphila, can be used as a therapeutic tool against diabetes.
Conclusion Remarks
Gut dysbiosis has been proposed to be a risk factor for the development of inflammatory and metabolic diseases. Restoring a balanced microbiota and modulating the gut–immune system axis using probiotics has been increasingly studied as a therapeutic strategy for these diseases. It has already been verified that A. muciniphila is capable of attenuating metabolic syndrome and damage to the intestinal mucosa by inducing an anti-inflammatory response and controlling intestinal homeostasis. Interestingly, the derived proteins A. muciniphila, AmOMV, and pasteurized A. muciniphila proved to be more efficient than viable bacteria for alleviating these diseases. Thus, A. muciniphila bacterial products or secreted proteins, named as postbiotics, have been proven to be promising targets as new therapeutic tools against chronic inflammatory and metabolic diseases.
Author Contributions
VR, JE-O, IP, JP, SB, and MM equally contributed to the manuscript writing. DC coordinated and reviewed the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This study was supported by grants from the São Paulo Research Foundation (FAPESP) (Process numbers: 2018/14815-0, 2020/05514-6).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We would like to thank Editage (www.editage.com) for English language editing.
References
1. Bibbò S, Dore MP, Pes GM, Delitala G, Delitala AP. Is There a Role for Gut Microbiota in Type 1 Diabetes Pathogenesis? Ann Med (2017) 49:11–22. doi: 10.1080/07853890.2016.1222449
2. Pabst O, Mowat AM. Oral Tolerance to Food Protein. Mucosal Immunol (2012) 5:232–9. doi: 10.1038/mi.2012.4
3. Xie Z, Huang G, Wang Z, Luo S, Zheng P, Zhou Z. Epigenetic Regulation of Toll-Like Receptors and its Roles in Type 1 Diabetes. J Mol Med (2018) 96:741–51. doi: 10.1007/s00109-018-1660-7
4. Scott NA, Andrusaite A, Andersen P, Lawson M, Alcon-Giner C, Leclaire C, et al. Antibiotics Induce Sustained Dysregulation of Intestinal T Cell Immunity by Perturbing Macrophage Homeostasis. Sci Transl Med (2018) 10:eaao4755. doi: 10.1126/scitranslmed.aao4755
5. Fava F. Intestinal Microbiota in Inflammatory Bowel Disease: Friend of Foe? World J Gastroenterol (2011) 17:557. doi: 10.3748/wjg.v17.i5.557
6. de Groot PF, Belzer C, Aydin Ö, Levin E, Levels JH, Aalvink S, et al. Distinct Fecal and Oral Microbiota Composition in Human Type 1 Diabetes, an Observational Study. PLoS One (2017) 12:e0188475. doi: 10.1371/journal.pone.0188475
7. Morrison DJ, Preston T. Formation of Short Chain Fatty Acids by the Gut Microbiota and Their Impact on Human Metabolism. Gut Microbes (2016) 7:189–200. doi: 10.1080/19490976.2015.1134082
8. He M, Shi B. Gut Microbiota as a Potential Target of Metabolic Syndrome: The Role of Probiotics and Prebiotics. Cell Biosci (2017) 7:54. doi: 10.1186/s13578-017-0183-1
9. Zhai Q, Feng S, Arjan N, Chen W. A Next Generation Probiotic, Akkermansia Muciniphila. Crit Rev Food Sci Nutr (2019) 59:3227–36. doi: 10.1080/10408398.2018.1517725
10. Derrien M, Vaughan EE, Plugge CM, de Vos WM. Akkermansia Muciniphila Gen. Nov., Sp. Nov., a Human Intestinal Mucin-Degrading Bacterium. Int J Syst Evol Microbiol (2004) 54:1469–76. doi: 10.1099/ijs.0.02873-0
11. Derrien M, Collado MC, Ben-Amor K, Salminen S, de Vos WM. The Mucin Degrader Akkermansia Muciniphila Is an Abundant Resident of the Human Intestinal Tract. Appl Environ Microbiol (2008) 74:1646–8. doi: 10.1128/AEM.01226-07
12. Lukovac S, Belzer C, Pellis L, Keijser BJ, de Vos WM, Montijn RC, et al. Differential Modulation by Akkermansia Muciniphila and Faecalibacterium Prausnitzii of Host Peripheral Lipid Metabolism and Histone Acetylation in Mouse Gut Organoids. MBio (2014) 5(4):e01438-14. doi: 10.1128/mBio.01438-14
13. Frost G, Sleeth ML, Sahuri-Arisoylu M, Lizarbe B, Cerdan S, Brody L, et al. The Short-Chain Fatty Acid Acetate Reduces Appetite via a Central Homeostatic Mechanism. Nat Commun (2014) 5:3611. doi: 10.1038/ncomms4611
14. Russell WR, Hoyles L, Flint HJ, Dumas M-E. Colonic Bacterial Metabolites and Human Health. Curr Opin Microbiol (2013) 16:246–54. doi: 10.1016/j.mib.2013.07.002
15. Everard A, Belzer C, Geurts L, Ouwerkerk JP, Druart C, Bindels LB, et al. Cross-Talk Between Akkermansia Muciniphila and Intestinal Epithelium Controls Diet-Induced Obesity. Proc Natl Acad Sci (2013) 110:9066–71. doi: 10.1073/pnas.1219451110
16. Pittayanon R, Lau JT, Leontiadis GI, Tse F, Yuan Y, Surette M, et al. Differences in Gut Microbiota in Patients With vs Without Inflammatory Bowel Diseases: A Systematic Review. Gastroenterology (2020) 158:930–46.e1. doi: 10.1053/j.gastro.2019.11.294
17. Zheng P, Li Z, Zhou Z. Gut Microbiome in Type 1 Diabetes: A Comprehensive Review. Diabetes Metab Res Rev (2018) 34:e3043. doi: 10.1002/dmrr.3043
18. Fasano A, Shea-Donohue T. Mechanisms of Disease: The Role of Intestinal Barrier Function in the Pathogenesis of Gastrointestinal Autoimmune Diseases. Nat Clin Pract Gastroenterol Hepatol (2005) 2:416–22. doi: 10.1038/ncpgasthep0259
19. Karl JP, Margolis LM, Madslien EH, Murphy NE, Castellani JW, Gundersen Y, et al. Changes in Intestinal Microbiota Composition and Metabolism Coincide With Increased Intestinal Permeability in Young Adults Under Prolonged Physiological Stress. Am J Physiol Liver Physiol (2017) 312:G559–71. doi: 10.1152/ajpgi.00066.2017
20. Salim SY, Söderholm JD. Importance of Disrupted Intestinal Barrier in Inflammatory Bowel Diseases. Inflammation Bowel Dis (2011) 17:362–81. doi: 10.1002/ibd.21403
21. Lutgendorff F, Nijmeijer RM, Sandström PA, Trulsson LM, Magnusson K-E, Timmerman HM, et al. Probiotics Prevent Intestinal Barrier Dysfunction in Acute Pancreatitis in Rats via Induction of Ileal Mucosal Glutathione Biosynthesis. PLoS One (2009) 4:e4512. doi: 10.1371/journal.pone.0004512
22. Ohland CL, MacNaughton WK. Probiotic Bacteria and Intestinal Epithelial Barrier Function. Am J Physiol Liver Physiol (2010) 298:G807–19. doi: 10.1152/ajpgi.00243.2009
23. Molina-Tijeras JA, Gálvez J, Rodríguez-Cabezas ME. The Immunomodulatory Properties of Extracellular Vesicles Derived From Probiotics: A Novel Approach for the Management of Gastrointestinal Diseases. Nutrients (2019) 11:1038. doi: 10.3390/nu11051038
24. Sairenji T, Collins KL, Evans DV. An Update on Inflammatory Bowel Disease. Prim Care Clin Off Pract (2017) 44:673–92. doi: 10.1016/j.pop.2017.07.010
25. Zhang Y-Z. Inflammatory Bowel Disease: Pathogenesis. World J Gastroenterol (2014) 20:91. doi: 10.3748/wjg.v20.i1.91
26. Shaw SY, Blanchard JF, Bernstein CN. Association Between the Use of Antibiotics in the First Year of Life and Pediatric Inflammatory Bowel Disease. Am J Gastroenterol (2010) 105:2687–92. doi: 10.1038/ajg.2010.398
27. Qu S, Fan L, Qi Y, Xu C, Hu Y, Chen S, et al. Akkermansia Muciniphila Alleviates Dextran Sulfate Sodium (DSS)-Induced Acute Colitis by NLRP3 Activation. Microbiol Spectr (2021) 9(2):e0073021. doi: 10.1128/Spectrum.00730-21
28. Bian X, Wu W, Yang L, Lv L, Wang Q, Li Y, et al. Administration of Akkermansia Muciniphila Ameliorates Dextran Sulfate Sodium-Induced Ulcerative Colitis in Mice. Front Microbiol (2019) 10:2259. doi: 10.3389/fmicb.2019.02259
29. Wang L, Tang L, Feng Y, Zhao S, Han M, Zhang C, et al. A Purified Membrane Protein From Akkermansia Muciniphila or the Pasteurised Bacterium Blunts Colitis Associated Tumourigenesis by Modulation of CD8 + T Cells in Mice. Gut (2020) 69(11):1988–97. doi: 10.1136/gutjnl-2019-320105. gutjnl-2019-320105.
30. Qian K, Chen S, Wang J, Sheng K, Wang Y, Zhang M. A β- N -Acetylhexosaminidase Amuc_2109 From Akkermansia Muciniphila Protects Against Dextran Sulfate Sodium-Induced Colitis in Mice by Enhancing Intestinal Barrier and Modulating Gut Microbiota. Food Funct (2022) 13:2216–27. doi: 10.1039/D1FO04094D
31. Hirota SA, Ng J, Lueng A, Khajah M, Parhar K, Li Y, et al. NLRP3 Inflammasome Plays a Key Role in the Regulation of Intestinal Homeostasis. Inflammation Bowel Dis (2011) 17:1359–72. doi: 10.1002/ibd.21478
32. Kim S, Shin Y-C, Kim T-Y, Kim Y, Lee Y-S, Lee S-H, et al. Mucin Degrader Akkermansia Muciniphila Accelerates Intestinal Stem Cell-Mediated Epithelial Development. Gut Microbes (2021) 13(1):1–20. doi: 10.1080/19490976.2021.1892441
33. Ahn IS, Lang JM, Olson CA, Diamante G, Zhang G, Ying Z, et al. Host Genetic Background and Gut Microbiota Contribute to Differential Metabolic Responses to Fructose Consumption in Mice. J Nutr (2020) 150:2716–28. doi: 10.1093/jn/nxaa239
34. Ganesh BP, Klopfleisch R, Loh G, Blaut M. Commensal Akkermansia Muciniphila Exacerbates Gut Inflammation in Salmonella Typhimurium-Infected Gnotobiotic Mice. PLoS One (2013) 8:e74963. doi: 10.1371/journal.pone.0074963
35. Ring C, Klopfleisch R, Dahlke K, Basic M, Bleich A, Blaut M. Akkermansia Muciniphila Strain ATCC BAA-835 Does Not Promote Short-Term Intestinal Inflammation in Gnotobiotic Interleukin-10-Deficient Mice. Gut Microbes (2019) 10:188–203. doi: 10.1080/19490976.2018.1511663
36. Sung KC, Ban M, Choi EJ, Moon HG, Jeon JS, Kim DK, et al. Extracellular Vesicles Derived From Gut Microbiota, Especially Akkermansia Muciniphila, Protect the Progression of Dextran Sulfate Sodium-Induced Colitis. PLoS One (2013) 8(10):e76520. doi: 10.1371/journal.pone.0076520
37. Ashrafian F, Keshavarz Azizi Raftar S, Shahryari A, Behrouzi A, Yaghoubfar R, Lari A, et al. Comparative Effects of Alive and Pasteurized Akkermansia Muciniphila on Normal Diet-Fed Mice. Sci Rep (2021) 11:17898. doi: 10.1038/s41598-021-95738-5
38. Choi Y, Bose S, Seo J, Shin J-H, Lee D, Kim Y, et al. Effects of Live and Pasteurized Forms of Akkermansia From the Human Gut on Obesity and Metabolic Dysregulation. Microorganisms (2021) 9:2039. doi: 10.3390/microorganisms9102039
39. Chelakkot C, Choi Y, Kim D-K, Park HT, Ghim J, Kwon Y, et al. Akkermansia Muciniphila-Derived Extracellular Vesicles Influence Gut Permeability Through the Regulation of Tight Junctions. Exp Mol Med (2018) 50:e450–0. doi: 10.1038/emm.2017.282
40. Ashrafian F, Shahriary A, Behrouzi A, Moradi HR, Keshavarz Azizi Raftar S, Lari A, et al. Akkermansia Muciniphila-Derived Extracellular Vesicles as a Mucosal Delivery Vector for Amelioration of Obesity in Mice. Front Microbiol (2019) 10:2155. doi: 10.3389/fmicb.2019.02155
41. Carlos D, Pérez MM, Leite JA, Rocha FA, Martins LMS, Pereira CA, et al. NOD2 Deficiency Promotes Intestinal CD4+ T Lymphocyte Imbalance, Metainflammation, and Aggravates Type 2 Diabetes in Murine Model. Front Immunol (2020) 11:1265. doi: 10.3389/fimmu.2020.01265
42. Cani PD, Bibiloni R, Knauf C, Neyrinck AM, Delzenne NM. Changes in Gut Microbiota Control Metabolic Diet–Induced Obesity and Diabetes in Mice. Diabetes (2008) 57:1470–81. doi: 10.2337/db07-1403.Additional
43. Cani PD, Amar J, Iglesias MA, Poggi M, Knauf C, Bastelica D, et al. Metabolic Endotoxemia Initiates Obesity and Insulin Resistance. Diabetes (2007) 56:1761–72. doi: 10.2337/DB06-1491
44. Karlsson CLJ, Önnerfält J, Xu J, Molin G, Ahrné S, Thorngren-Jerneck K. The Microbiota of the Gut in Preschool Children With Normal and Excessive Body Weight. Obesity (2012) 20:2257–61. doi: 10.1038/oby.2012.110
45. Santacruz A, Collado MC, García-Valdés L, Segura MT, Martín-Lagos JA, Anjos T, et al. Gut Microbiota Composition is Associated With Body Weight, Weight Gain and Biochemical Parameters in Pregnant Women. Br J Nutr (2010) 104:83–92. doi: 10.1017/S0007114510000176
46. Yassour M, Lim MY, Yun HS, Tickle TL, Sung J, Song Y-M, et al. Sub-Clinical Detection of Gut Microbial Biomarkers of Obesity and Type 2 Diabetes. Genome Med (2016) 8:17. doi: 10.1186/s13073-016-0271-6
47. Parks BW, Nam E, Org E, Kostem E, Norheim F, Hui ST, et al. Genetic Control of Obesity and Gut Microbiota Composition in Response to High-Fat, High-Sucrose Diet in Mice. Cell Metab (2013) 17:141–52. doi: 10.1016/J.CMET.2012.12.007
48. Schneeberger M, Everard A, Gómez-Valadés AG, Matamoros S, Ramírez S, Delzenne NM, et al. Akkermansia Muciniphila Inversely Correlates With the Onset of Inflammation, Altered Adipose Tissue Metabolism and Metabolic Disorders During Obesity in Mice. Sci Rep (2015) 5:16643. doi: 10.1038/srep16643
49. Suriano F, Vieira-Silva S, Falony G, Roumain M, Paquot A, Pelicaen R, et al. Novel Insights Into the Genetically Obese (Ob/Ob) and Diabetic (Db/Db) Mice: Two Sides of the Same Coin. Microbiome (2021) 9:147. doi: 10.1186/s40168-021-01097-8
50. Reunanen J, Kainulainen V, Huuskonen L, Ottman N, Belzer C, Huhtinen H, et al. Akkermansia Muciniphila Adheres to Enterocytes and Strengthens the Integrity of the Epithelial Cell Layer. Appl Environ Microbiol (2015) 81:3655–62. doi: 10.1128/AEM.04050-14/SUPPL_FILE/ZAM999116271SO1.PDF
51. Plovier H, Everard A, Druart C, Depommier C, Van Hul M, Geurts L, et al. A Purified Membrane Protein From Akkermansia Muciniphila or the Pasteurized Bacterium Improves Metabolism in Obese and Diabetic Mice. Nat Med (2017) 23:107–13. doi: 10.1038/nm.4236
52. Depommier C, Van Hul M, Everard A, Delzenne NM, De Vos WM, Cani PD. Pasteurized Akkermansia Muciniphila Increases Whole-Body Energy Expenditure and Fecal Energy Excretion in Diet-Induced Obese Mice. Gut Microbes (2020) 11:1231–45. doi: 10.1080/19490976.2020.1737307
53. Ashrafian F, Keshavarz Azizi Raftar S, Lari A, Shahryari A, Abdollahiyan S, Moradi HR, et al. Extracellular Vesicles and Pasteurized Cells Derived From Akkermansia Muciniphila Protect Against High-Fat Induced Obesity in Mice. Microb Cell Fact (2021) 20:219. doi: 10.1186/s12934-021-01709-w
54. Shin NR, Lee JC, Lee HY, Kim MS, Whon TW, Lee MS, et al. An Increase in the Akkermansia Spp. Population Induced by Metformin Treatment Improves Glucose Homeostasis in Diet-Induced Obese Mice. Gut (2014) 63:727–35. doi: 10.1136/GUTJNL-2012-303839
55. Kong C, Gao R, Yan X, Huang L, Qin H. Probiotics Improve Gut Microbiota Dysbiosis in Obese Mice Fed a High-Fat or High-Sucrose Diet. Nutrition (2019) 60:175–84. doi: 10.1016/J.NUT.2018.10.002
56. Yoon HS, Cho CH, Yun MS, Jang SJ, You HJ, Kim J, et al. Akkermansia Muciniphila Secretes a Glucagon-Like Peptide-1-Inducing Protein That Improves Glucose Homeostasis and Ameliorates Metabolic Disease in Mice. Nat Microbiol (2021) 6:563–73. doi: 10.1038/s41564-021-00880-5
57. Lee J, Song W, Lim JW, Choi T, Jo S, Jeon H, et al. An Integrative Multiomics Approach to Characterize Anti-Adipogenic and Anti-Lipogenic Effects of Akkermansia Muciniphila in Adipocytes. Biotechnol J (2022) 17:2100397. doi: 10.1002/biot.202100397
58. Palleja A, Kashani A, Allin KH, Nielsen T, Zhang C, Li Y, et al. Roux-En-Y Gastric Bypass Surgery of Morbidly Obese Patients Induces Swift and Persistent Changes of the Individual Gut Microbiota. Genome Med (2016) 8(1):67. doi: 10.1186/s13073-016-0312-1
59. Kim S, Lee Y, Kim Y, Seo Y, Lee H, Ha J, et al. Akkermansia Muciniphila Prevents Fatty Liver Disease, Decreases Serum Triglycerides, and Maintains Gut Homeostasis. Appl Environ Microbiol (2020) 86(7):e03004-19. doi: 10.1128/AEM.03004-19
60. Depommier C, Everard A, Druart C, Plovier H, Van Hul M, Vieira-Silva S, et al. Supplementation With Akkermansia Muciniphila in Overweight and Obese Human Volunteers: A Proof-of-Concept Exploratory Study. Nat Med (2019) 25:1096–103. doi: 10.1038/s41591-019-0495-2
61. Depommier C, Vitale RM, Iannotti FA, Silvestri C, Flamand N, Druart C, et al. Beneficial Effects of Akkermansia Muciniphila Are Not Associated With Major Changes in the Circulating Endocannabinoidome But Linked to Higher Mono-Palmitoyl-Glycerol Levels as New Pparα Agonists. Cells (2021) 10:185. doi: 10.3390/cells10010185
62. Perraudeau F, McMurdie P, Bullard J, Cheng A, Cutcliffe C, Deo A, et al. Improvements to Postprandial Glucose Control in Subjects With Type 2 Diabetes: A Multicenter, Double Blind, Randomized Placebo-Controlled Trial of a Novel Probiotic Formulation. BMJ Open Diabetes Res Care (2020) 8:e001319. doi: 10.1136/bmjdrc-2020-001319
63. McMurdie PJ, Stoeva MK, Justice N, Nemchek M, Sieber CMK, Tyagi S, et al. Increased Circulating Butyrate and Ursodeoxycholate During Probiotic Intervention in Humans With Type 2 Diabetes. BMC Microbiol (2022) 22:19. doi: 10.1186/s12866-021-02415-8
64. Dao MC, Everard A, Aron-Wisnewsky J, Sokolovska N, Prifti E, Verger EO, et al. Akkermansia Muciniphila and Improved Metabolic Health During a Dietary Intervention in Obesity: Relationship With Gut Microbiome Richness and Ecology. Gut (2016) 65:426–36. doi: 10.1136/gutjnl-2014-308778
65. Roshanravan N, Mahdavi R, Alizadeh E, Ghavami A, Saadat YR, Alamdari NM, et al. The Effects of Sodium Butyrate and Inulin Supplementation on Angiotensin Signaling Pathway via Promotion of Akkermansia Muciniphila Abundance in Type 2 Diabetes; A Randomized, Double-Blind, Placebo-Controlled Trial. J Cardiovasc Thorac Res (2017) 9:183. doi: 10.15171/JCVTR.2017.32
66. Rodriguez J, Hiel S, Neyrinck AM, Le Roy T, Pötgens SA, Leyrolle Q, et al. Discovery of the Gut Microbial Signature Driving the Efficacy of Prebiotic Intervention in Obese Patients. Gut (2020) 69:1975–87. doi: 10.1136/GUTJNL-2019-319726
67. Kosteli A, Sugaru E, Haemmerle G, Martin JF, Lei J, Zechner R, et al. Weight Loss and Lipolysis Promote a Dynamic Immune Response in Murine Adipose Tissue. J Clin Invest (2010) 120(10):3466–79. doi: 10.1172/JCI42845
68. Hirosumi J, Rol Tuncman G, Chang L, Gö Rgü CZ, Uysal KT, Maeda K, et al. A Central Role for JNK in Obesity and Insulin Resistance. (2002) 420(6913):333–6. doi: 10.1038/nature01137
69. Roopchand DE, Carmody RN, Kuhn P, Moskal K, Rojas-Silva P, Turnbaugh PJ, et al. Dietary Polyphenols Promote Growth of the Gut Bacterium Akkermansia Muciniphila and Attenuate High-Fat Diet-Induced Metabolic Syndrome. Diabetes (2015) 64:2847–58. doi: 10.2337/db14-1916
70. Anhê FF, Roy D, Pilon G, Dudonné S, Matamoros S, Varin TV, et al. A Polyphenol-Rich Cranberry Extract Protects From Diet-Induced Obesity, Insulin Resistance and Intestinal Inflammation in Association With Increased Akkermansia Spp. Population in the Gut Microbiota of Mice. Gut (2015) 64:872–83. doi: 10.1136/GUTJNL-2014-307142/-/DC1
71. Qin J, Li Y, Cai Z, Li S, Zhu J, Zhang F, et al. A Metagenome-Wide Association Study of Gut Microbiota in Type 2 Diabetes. Nature (2012) 490:55–60. doi: 10.1038/nature11450
72. Tilg H, Moschen AR. Microbiota and Diabetes: An Evolving Relationship. Gut (2014) 63:1513–21. doi: 10.1136/gutjnl-2014-306928
73. Zhang X, Shen D, Fang Z, Jie Z, Qiu X, Zhang C, et al. Human Gut Microbiota Changes Reveal the Progression of Glucose Intolerance. PLoS One (2013) 8:e71108. doi: 10.1371/journal.pone.0071108
74. Zhang J, Ni Y, Qian L, Fang Q, Zheng T, Zhang M, et al. Decreased Abundance of Akkermansia Muciniphila Leads to the Impairment of Insulin Secretion and Glucose Homeostasis in Lean Type 2 Diabetes. Adv Sci (2021) 8:2100536. doi: 10.1002/advs.202100536
75. Shih C-T, Yeh Y-T, Lin C-C, Yang L-Y, Chiang C-P. Akkermansia Muciniphila is Negatively Correlated With Hemoglobin A1c in Refractory Diabetes. Microorganisms (2020) 8:1360. doi: 10.3390/microorganisms8091360
76. Zhang L, Qin Q, Liu M, Zhang X, He F, Wang G. Akkermansia Muciniphila can Reduce the Damage of Gluco/Lipotoxicity, Oxidative Stress and Inflammation, and Normalize Intestine Microbiota in Streptozotocin-Induced Diabetic Rats. Pathog Dis (2018) 76(4):1–15. doi: 10.1093/femspd/fty028
77. Harbison JE, Roth-Schulze AJ, Giles LC, Tran CD, Ngui KM, Penno MA, et al. Gut Microbiome Dysbiosis and Increased Intestinal Permeability in Children With Islet Autoimmunity and Type 1 Diabetes: A Prospective Cohort Study. Pediatr Diabetes (2019) 20(5):574–83. doi: 10.1111/pedi.12865. pedi.12865.
78. Jamshidi P, Hasanzadeh S, Tahvildari A, Farsi Y, Arbabi M, Mota JF, et al. Is There Any Association Between Gut Microbiota and Type 1 Diabetes? A Systematic Review. Gut Pathog (2019) 11:49. doi: 10.1186/s13099-019-0332-7
79. Sato J, Kanazawa A, Ikeda F, Yoshihara T, Goto H, Abe H, et al. Gut Dysbiosis and Detection of “Live Gut Bacteria” in Blood of Japanese Patients With Type 2 Diabetes. Diabetes Care (2014) 37:2343–50. doi: 10.2337/dc13-2817
80. Debray-Sachs M, Carnaud C, Boitard C, Cohen H, Gresser I, Bedossa P, et al. Prevention of Diabetes in NOD Mice Treated With Antibody to Murine Ifnγ. J Autoimmun (1991) 4:237–48. doi: 10.1016/0896-8411(91)90021-4
81. Greer RL, Dong X, Moraes ACF, Zielke RA, Fernandes GR, Peremyslova E, et al. Akkermansia Muciniphila Mediates Negative Effects of Ifnγ on Glucose Metabolism. Nat Commun (2016) 7:13329. doi: 10.1038/ncomms13329
82. Hansen CHF, Krych L, Nielsen DS, Vogensen FK, Hansen LH, Sørensen SJ, et al. Early Life Treatment With Vancomycin Propagates Akkermansia Muciniphila and Reduces Diabetes Incidence in the NOD Mouse. Diabetologia (2012) 55:2285–94. doi: 10.1007/s00125-012-2564-7
83. Costa FRC, Françozo MCS, de Oliveira GG, Ignacio A, Castoldi A, Zamboni DS, et al. Gut Microbiota Translocation to the Pancreatic Lymph Nodes Triggers NOD2 Activation and Contributes to T1D Onset. J Exp Med (2016) 213:1223–39. doi: 10.1084/jem.20150744
Keywords: Akkermansia muciniphila, gut dysbiosis, obesity, diabetes, inflammatory bowel diseases
Citation: Rodrigues VF, Elias-Oliveira J, Pereira ÍS, Pereira JA, Barbosa SC, Machado MSG and Carlos D (2022) Akkermansia muciniphila and Gut Immune System: A Good Friendship That Attenuates Inflammatory Bowel Disease, Obesity, and Diabetes. Front. Immunol. 13:934695. doi: 10.3389/fimmu.2022.934695
Received: 02 May 2022; Accepted: 13 June 2022;
Published: 07 July 2022.
Edited by:
Jian Tan, The University of Sydney, AustraliaReviewed by:
Fatemeh Ashrafian, Pasteur Institute of Iran (PII), IranCopyright © 2022 Rodrigues, Elias-Oliveira, Pereira, Pereira, Barbosa, Machado and Carlos. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Daniela Carlos, ZGFuaWNhckB1c3AuYnI=